Спосіб покращення толерантності рослини до засухи та дії солі
Номер патенту: 106489
Опубліковано: 10.09.2014
Автори: Хуанг Ксіньюан, Чжу Мейчжен, Лін Хонгксуан, Гао Цзіпін, Ші Мін, Чао Даіїн


















Формула / Реферат
1. Спосіб покращення толерантності до засухи та дії солі у рослини, який включає:
інгібування функції активації транскрипції фактора транскрипції, де фактор транскрипції включає домен ″цинкові пальці″ типу С2H2, що містить послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2;
причому
інгібування здійснюють шляхом введення полінуклеотиду, що кодує неконсервативний мутантний фактор транскрипції, який являє собою білок, що має послідовність, яка включає Asn69Asp (N69D) мутацію та Ala162Thr (A162T) мутацію у послідовності SEQ ID NO: 2; або
інгібування здійснюють шляхом інгібування експресії фактора транскрипції введенням RNAi або антисмислового олігонуклеотиду у рослину.
2. Спосіб за п. 1, в якому фактор транскрипції вибраний з:
(a) поліпептиду, що містить амінокислотну послідовність SEQ ID NO: 2;
(b) поліпептиду, одержаного з (a), в якому 1-50 амінокислотних залишків заміщено, видалено або інсертовано, та який здатний до підвищення чутливості до засухи та дії солі у рослини.
3.Спосіб за п. 1, в якому фактор транскрипції має амінокислотну послідовність SEQ ID NO: 2.
4. Спосіб за п. 1, в якому інгібування здійснюють шляхом введення полінуклеотиду, що кодує неконсервативний мутантний фактор транскрипції, який не здатний до підвищення чутливості до засухи та дії солі у рослини.
5. Спосіб покращення за п. 1 або 4, в якому введення полінуклеотиду включає:
(1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить полінуклеотид;
(2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих полінуклеотидом; та
(3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2).
6. Спосіб за п. 1, в якому RNAi або антисмисловий олігонуклеотид направлений проти послідовності SEQ ID NO: 1.
7. Спосіб за п. 1 або 6, в якому інгібування експресії фактора транскрипції здійснюють:
(1) трансформуванням рослинної клітини, рослинної тканини або органу рослини конструктом, що містить RNAi або антисмисловий олігонуклеотид;
(2) відбором рослинної клітини, рослинної тканини або органу рослини, трансформованих олігонуклеотидом; та
(3) регенерацією рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2).
8. Спосіб за п. 1, в якому інгібування здійснюють шляхом інгібування зв′язування фактора транскрипції з цис-діючим елементом.
9. Спосіб за п. 8, в якому цис-діючий елемент містить послідовність SEQ ID NO: 3.
10. Рослина, одержана способом відповідно до будь-якого з пп. 1-4, 6, 8-9, що передбачає:
(1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить полінуклеотид;
(2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих полінуклеотидом; та
(3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2); або
(1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить RNAi або антисмисловий олігонуклеотид;
(2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих олігонуклеотидом; та
(3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2).
11. Рослина за п. 10, яка відрізняється тим, що рослина є культурою, вибраною з Gramineae, Malvaceae gossypium, Cruciferae brassica, Compositae, Solanaceae, Labiatae або Umbelliferae.
12. Рослина за п. 11, яка відрізняється тим, що рослина є культурою, вибраною з рису, кукурудзи, пшениці, ячменю, цукрової тростини, сорго, арабідопсису, бавовни або каноли.
13. Спосіб відбору рослини, толерантної до засухи та дії солі, що включає:
(i) визначення у кандидатної рослини рівня фактора транскрипції, що містить
домен ″цинкові пальці″ типу С2H2, що містить послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2;
рівня експресії полінуклеотиду, що кодує послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2; та/або рівня зв′язування між цис-діючим елементом, що містить послідовність SEQ ID NO: 3 та фактором транскрипції; та
(ii) порівняння рівня у кандидатної рослини, визначеного на стадії (i) з відповідним рівнем у контрольної рослини, якщо рівень у кандидатної рослини, нижчий за рівень у контрольної рослини, тоді кандидатна рослина є рослиною толерантною до засухи та дії солі.
Текст
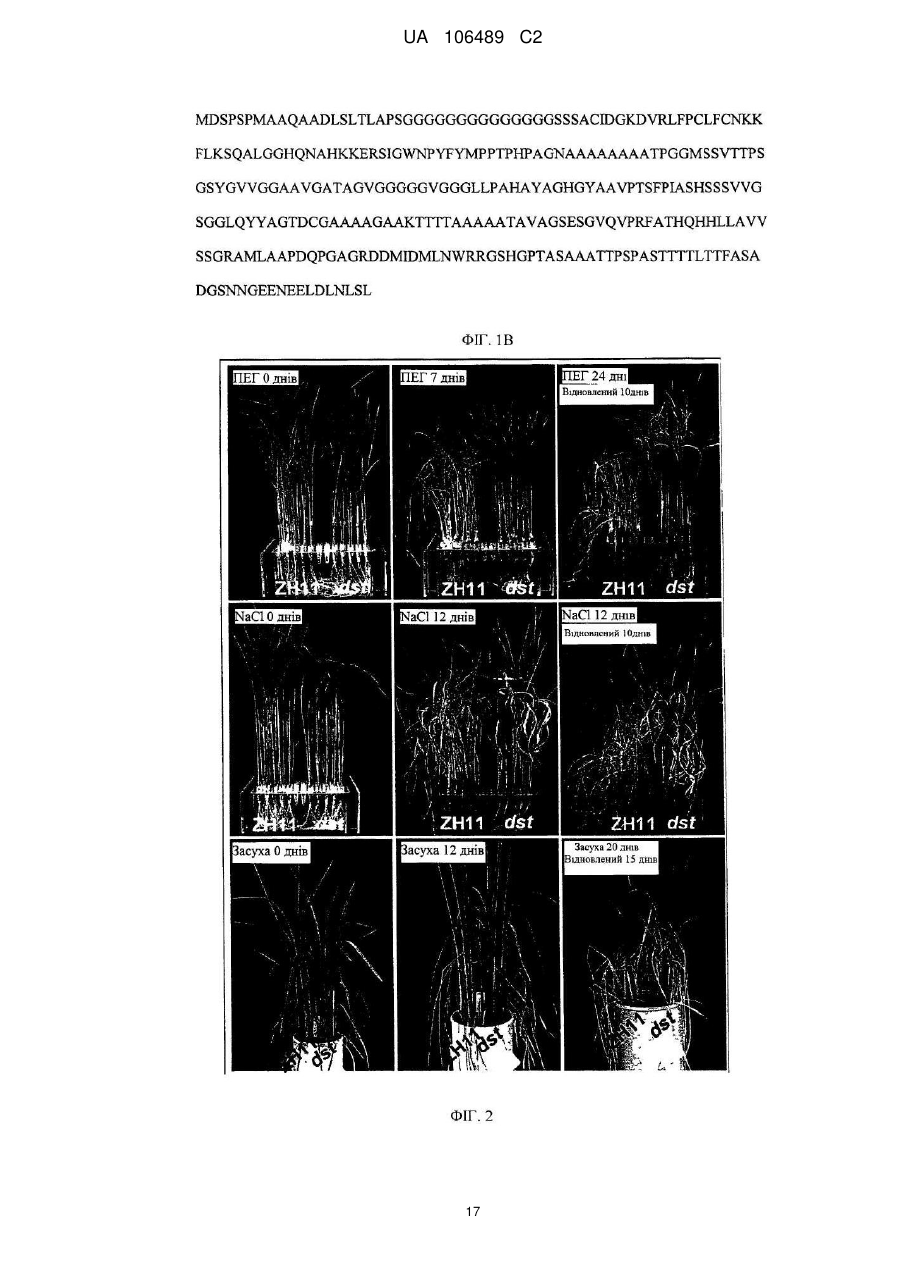
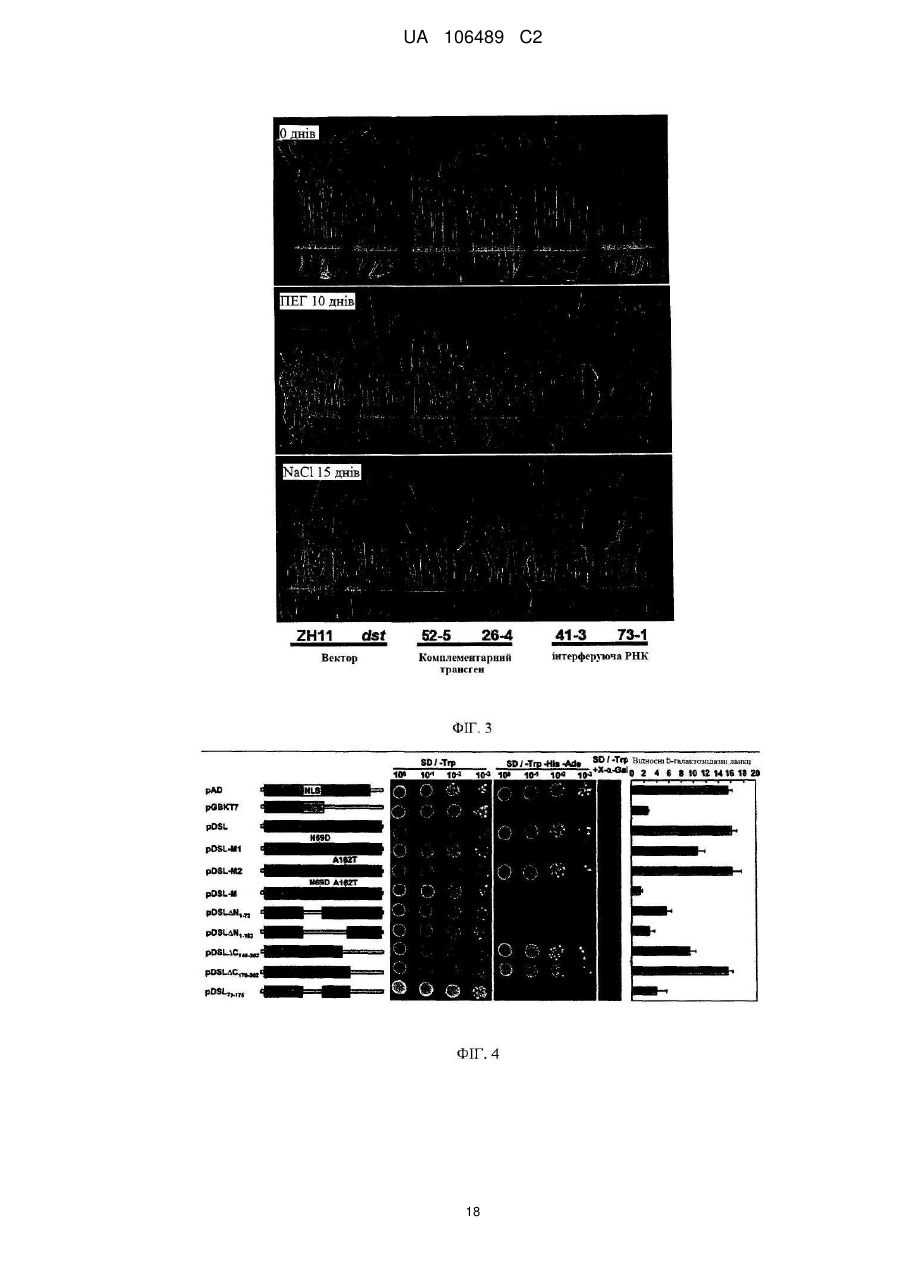
Реферат: Винахід належить до способу покращення толерантності до засухи та дії солі у рослини, який включає:інгібування функції активації транскрипції фактора транскрипції, де фактор транскрипції включає домен ″цинкові пальці″ типу С2H2, що містить послідовність амінокислот UA 106489 C2 (12) UA 106489 C2 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2; причому інгібування здійснюють шляхом введення полінуклеотиду, що кодує неконсервативний мутантний фактор транскрипції, який являє собою білок, що має послідовність, яка включає Asn69Asp (N69D) мутацію та Ala162Thr (A162T) мутацію у послідовності SEQ ID NO: 2; або інгібування здійснюють шляхом інгібування експресії фактора транскрипції введенням RNAi або антисмислового олігонуклеотиду у рослину. UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 ГАЛУЗЬ ВИНАХОДУ Даний винахід відноситься до галузі рослинної біоінженерії та генної інженерії для покращення рослин. Конкретно, даний винахід відноситься до застосування нових білків "цинкові пальці" генів фактору транскрипції рису та їх кодованих білків або поліпептидів для підвищення толерантності до засухи та дії солі у рослин, способам підвищення резистентності до солей та/або резистентності до засухи у рослин шляхом інгібування генів, описаних вище, або їх експресованих білків, та до трансгенних рослин. РІВЕНЬ ТЕХНІКИ Підвищена світова потреба у їжі та постійне скорочення сільськогосподарських земель створила постійне навантаження на національну продовольчу безпеку. При виробництві їжі, засуха та сіль є основними небіологічними (абіотичними) стресами, призводячи до значного скорочення кількості та якості зернових кожного року. Додатково, засуха часто супроводжує дію солі. Наприклад, засолення ґрунту є загальним явищем при погіршенні ґрунту на засушливих територіях. Доступні дані демонструють, що щорічні втрати риса через засуху у Китаї коштують щонайменше 2 мільярда доларів США, та засолення сільськогосподарських земель є також основною причиною для зменшення продукції та низьких врожаїв. Засуха та дія солі являли собою дві серйозні проблеми, що перешкоджали сільському господарству Китаю. Тому, велике значення має, як покращити толерантність до засухи та/або дії солі у культур для підвищення врожаїв, вирішення проблем з їжею у Китаї та у світі. Дослідження абіотичного стресу є одним з найбільш термінових та проблемних галузей при дослідженні рослин. На даний час, було клоновано деякі гени, пов'язані з резистентністю до засухи та дії солі, включаючи декілька факторів транскрипції, та було одержано деякі резистентні до стресу штами рослин або культур шляхом застосування методів генної інженерії. Метою генної інженерії для одержання резистентності до стресу є покращення резистентності до стресу рослин шляхом модулювання генної транскрипції та експресії, в яких фактори транскрипції відіграють основні ролі. На сьогодні, було клоновано деякі фактори транскрипції, що задіяні у генній експресії у відповідь на стрес. Проте, через те, що така велика кількість факторів транскрипції задіяна у цих складних процесах стресової відповіді рослин, такі фактори транскрипції являють собою тільки верхню частину айсберга. Набагато більшу кількість факторів транскрипції необхідно відкрити, дослідити та застосовувати при розведенні культур, які мають резистентність до стресу. Тому, існує термінова потребі у цій галузі у дослідженні факторів транскрипції, які задіяні у генні експресії у відповідь на стрес, для розробки нових способів розмноження резистентних до стресу культур та для розробки таких нових культур, таким чином збільшуючи кількість та покращуючи якість культур. СТИСЛИЙ ОПИС ВИНАХОДУ Однією з цілей даного винаходу є забезпечення нового гена, що має безпосереднє відношення до толерантності рослин (особливо зернових) до дії солі та засухи – фактору транскрипції білків "цинкових пальців" DST рису, та підтвердження того, що цей фактор транскрипції є негативним регулятором толерантності до дії солі та засухи у рослин. Іншою ціллю винаходу є забезпечення нового способу підвищення резистентності до дії солі та/або засухи у рослин. Іншою ціллю даного винаходу є забезпечення способів одержання трансгенних рослин з підвищеною толерантністю до дії солі та засухи та трансгенних рослин, одержаних такими способами. Іншою ціллю даного винаходу є забезпечення способів скринінгу рослин, що мають підвищену толерантність до дії солі та засухи, одержаних такими способами скринінгу. У першому аспекті, даний винахід забезпечує виділені фактори транскрипції білків "цинкових пальців", включаючи поліпептиди, що мають амінокислотну послідовність SEQ ID NO: 2, поліпептиди, що мають консервативні мутації у поліпептидах-попередниках, або гомологи поліпептидів-попередників. В одному переважному втіленні, вищевказані поліпептиди включають структурний домен типу "цинкових пальців" Cys-2/His-2. В іншому переважному втіленні, вказані поліпептиди задіяні у регуляції генів, пов'язаних з ферментами, що функціонують на основі пероксидів, контролюючи накопичення пероксидів та/або регулюючи продиховий отвір, таким чином, впливаючи на толерантність до засухи та дії солі у рису. В одному втіленні даного винаходу, вказаний поліпептид вибирають з такої групи: (a) поліпептиду, що має амінокислотну послідовність SEQ ID NO: 2; 1 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 (b) поліпептиду, що одержаний (a) шляхом однієї або більше заміщень, делецій або інсерцій амінокислотного залишку, та здатного підвищувати чутливість до засухи та дії солі у рослини; або (c) поліпептидного гомологу поліпептиду групи (a) або (b), що має Cys-2/His-2 структурний домен типу "цинкових пальців" та здатний до підвищення чутливості до засухи та дії солі у рослини. У переважних втіленнях, вказані рослини є дводольними рослинами або однодольними рослинами, переважно зерновими. В іншому переважному втіленні, вказані рослини вибирають з: зернових, бавовни сімейства мальвових, брюссельської капусти сімейства хрестоцвітних, складноцвітних, пасльонових, губоцвітних, або зонтичних, переважно зернових. В іншому переважному втіленні, вказані рослини вибирають з: рису, кукурудзи, пшениці, ячменю, цукрової тростини, сорго, Арабідопсису, бавовни або каноли, більш переважно, рису, кукурудзи, пшениці, ячменю, цукрової тростини або сорго. В іншому переважному втіленні, вказані дії солі відносься до: хлориду натрію, сульфату натрію, карбонату натрію або бікарбонату натрію. У іншому аспекті, даний винахід забезпечує виділені полінуклеотиди, що містять нуклеотидні послідовності, які кодують поліпептиди відповідно до даного винаходу. В одному переважному втіленні, вказаний полінуклеотид кодує амінокислотну послідовність SEQ ID NO: 2 або її поліпептидний гомолог. В одному втіленні даного винаходу, вказану полінуклеотидну послідовність вибирають з: (a) нуклеотидної послідовності, що містить послідовність SEQ ID NO: 1; (b) нуклеотидної послідовності, що містить нуклеотиди 1-435 у SEQ ID NO: 1; або (c) полінуклеотидної послідовності, комплементарної одній з нуклеотидних послідовностей (a)-(b). У третьому аспекті, даний винахід забезпечує вектор, що містить полінуклеотид відповідно до даного винаходу. У переважних втіленнях, вказаний вектор вибирають з: бактеріального плазміду, фага, дріжджового плазміду, рослинного вірусу, або вірусу ссавців; переважно, pCAMBIA1301, ПЕГFP1, pBI121, pCAMBIA1300, pCAMBIA2301 або pHB, та, більш переважно, pCAMBIA1301. У четвертому аспекті, даний винахід забезпечує створені методами генної інженерії клітинихазяіни, що містять вектор відповідно до даного винаходу, або що містять полінуклеотид відповідно до даного винаходу, інтегрований у геном. В переважних втіленнях, вказану клітину-хазяїн вибирають з прокаріотної клітини, нижчої еукаріотної клітини або вищої еукаріотної клітини, переважно бактеріальної клітини, дріжджової клітини або рослинної клітини, більш переважно, кишкової палички, стрептоміцетів, агробактерій, дріжджів, найбільш переважно агробактерій, де вказана агробактерія включає, не обмежуючись наведеним: EHA105, SOUP1301 або C58, переважно, EHA105. У п'ятому аспекті, даний винахід забезпечує цис-діючий елемент, що має послідовність SEQ ID NO: 3, здатну зв'язувати фактор транскрипції відповідно до даного винаходу. У переважних втіленнях, вказаний цис-діючий елемент має послідовність TGCTANN(A/T)TTG, в якій N вибирають з A, C, G або T. В іншому переважному втіленні, вказаний цис-діючий елемент зв'язує структурний домен типу "цинкових пальців" фактора транскрипції відповідно до даного винаходу. В іншому переважному втіленні, зв'язування вказаного цис-діючого елементу із фактором транскрипції відповідно до даного винаходу може підвищувати чутливість до засухи та дії солі у рослин. У шостому аспекті, даний винахід забезпечує антагоністи для білків або полінуклеотидів фактора транскрипції "цинкових пальців". У переважних втіленнях, антагоністами є низькомолекулярні інтерферуючі РНК, антитіла або антисмислові олігонуклеотиди. У сьомому аспекті, даний винахід забезпечує способи покращення толерантності до засухи та дії солі у рослин, де вказані способи включають інгібування факторів транскрипції "цинкових пальців" відповідно до даного винаходу, інгібування експресії полінуклеотидів відповідно до даного винаходу, або інгібування зв'язування цис-діючого елементу з факторами транскрипції "цинкових пальців" відповідно до даного винаходу. В одному переважному втіленні, вказане інгібування проводять способами делеції, мутації, РНК інтерференції, антисмислової або домнантної негативної регуляції. В іншому переважному втіленні, вказане інгібування включає введення однієї або більше амінокислот або нуклеотидного заміщення, делеції або інсерції у фактори транскрипції 2 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 відповідно до даного винаходу або полінуклеотиди відповідно до даного винаходу, що призводить до того, що вказані рослини мають покращену толерантність до засухи та дії солі. В іншому переважному втіленні, відповідно до амінокислотної послідовності SEQ ID NO: 2, вказане інгібування включає мутування аспарагіну на аспарагінову кислоту при амінокислоті 69, мутування аланіну на треонін при амінокислоті 162, що призводить до того, що рослини з такими мутантними послідовностями мають покращену толерантність до засухи та дії солі. В іншому переважному втіленні, де вказані способи включають застосування антагоністу відповідно до даного винаходу до рослини. В іншому переважному втіленні, вказане інгібування включає: трансформування рослин вектором що містить малу інтерферуючу РНК, направлену на фактор транскрипції білків "цинкові пальці" відповідно до даного винаходу, або трансформування рослин за допомогою клітини-хазяїна, що містить вказані вектори. В іншому переважному втіленні, вказані способи додатково включають кросбридинг рослин, що мають підвищену толерантність до засухи та дії солі, одержаних способами, описаними вище, з нетрансгенними рослинами або іншими трансгенними рослинами. В іншому переважному втіленні, вказані дії солі відносяться до: хлориду натрію, сульфату натрію, карбонату натрію або бікарбонату натрію. У восьмому аспекті, даний винахід забезпечує способи скринінгу рослин на предмет толерантності до засухи та дії солі, де вказані способи включають: (i) детекцію у кандидатної рослини рівня фактору транскрипції білків "цинкові пальці" відповідно до даного винаходу, рівню експресії полінуклеотиду відповідно до даного винаходу, та/або рівні зв'язування цис-діючого елементу відповідно до даного винаходу із фактором транскрипції білків "цинкові пальці" відповідно до даного винаходу; та (ii) порівняння рівня у кандидатній рослині, детектованої на стадії (i) з рівнем у контрольної рослини, якщо рівень у кандидатній рослині нижчий за рівень контрольної рослини, де вказана кандидатна рослина є рослиною, толерантною до засухи та дії солі. У переважних втіленнях, вказані дії солі відносяться до: хлориду натрію, сульфату натрію, карбонату натрію або бікарбонату натрію. У дев'ятому аспекті, даний винахід забезпечує способи одержання фактору транскрипції білків "цинкові пальці", що відрізняються тим, що вказані способи включають: (a) культивування клітини-хазяїна відповідно до даного винаходу за умов, придатних для експресії; та (b) виділення фактору транскрипції білків "цинкові пальці" з культурального середовища. В іншому аспекті, даний винахід забезпечує застосування інгібітору або неконсервативної мутантної послідовності фактору транскрипції білків "цинкові пальці" або нуклеотидної послідовності відповідно до даного винаходу для покращення толерантності до засухи та дії солі у рослини. В одному переважному втіленні, вказаний інгібітор є низькомолекулярною інтерферуючою РНК, антитілом, або антисмисловим олігонуктеотидом, що направляє вказаний фактор транскрипції або нуклеотидну послідовність. В іншому переважному втіленні, вказана неконсервативна мутантна послідовність інгібує трансляцію або експресію фактору транскрипції білків "цинкові пальці" або нуклеотидної послідовності відповідно до даного винаходу у рослини, що містить вказану неконсервативну мутантну послідовність, що призводить до кращої толерантності до засухи та дії солі, ніж толерантність рослин дикого типу, що не містять неконсервативну мутантну послідовність. В іншому переважному втіленні, вказана неконсервативна мутантна послідовність є полінуклеотидною послідовністю SEQ ID NO: 1 з двома мутаціями: A у положенні 205 мутувала до G та G у положенні 484 мутувала до A; або амінокислотну послідовність SEQ ID NO: 2 з двома мутаціями: аспарагін у положенні 69 мутував на аспарагінову кислоту та аланін у положенні 162 мутував на треонін. В одному втіленні даного винаходу, вказане покращення толерантності до засухи та дії солі у рослин включає: (i) безпосереднє нанесення інгібітору (антагоніста), описаного вище, на рослину; (ii) введення неконсервативної мутантної послідовності, описаної вище, у рослину; або (iii) розробку молекулярного маркера, специфічного до неконсервативної мутантної послідовності, та застосування вказаного молекулярного маркера для скринінгу потомства, одержаного від кросбридингу мутантної рослини, що містить неконсервативну мутантну послідовність, з іншим видом рису для відбору індивідуального нащадка, що містить неконсервативну мутантну послідовність. 3 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 В одному переважному втіленні відповідно до даного винаходу, вказаний молекулярний маркер містить пару праймерів, що мають послідовності, показані у SEQ ID NO: 10 та SEQ ID NO: 11, та/або пару праймерів, що мають послідовності, показані у SEQ ID NO: 12 та SEQ ID NO: 13. В іншому аспекті, даний винахід забезпечує способи покращення толерантності до засухи та дії солі у рослини, де вказаний спосіб включає: (A) забезпечення інгібітору або неконсервативної мутантної послідовності для фактор транскрипції білків "цинкові пальці" або полінуклеотидної послідовності відповідно до даного винаходу; (B) піддавання рослини одній або більше обробкам, що вибирають з: (i) нанесення вказаного інгібітору безпосередньо на рослину; (ii) введення неконсервативної мутантної послідовності у рослину; або (iii) розробки молекулярного маркера, специфічного до неконсервативної мутантної послідовності, та застосування вказаного молекулярного маркера для скринінгу потомства від кросбридингу мутантної рослини, що містить неконсервативну мутантну послідовність та іншого штаму рису для відбору індивідуального нащадка, що містить неконсервативну мутантну послідовність. В одному переважному втіленні відповідно до даного винаходу, вказаний молекулярний маркер є парою праймерів, що мають послідовності, показані у SEQ ID NO: 10 та SEQ ID NO: 11, та/або парою праймерів, що мають послідовності, показані у SEQ ID NO: 12 та SEQ ID NO: 13. В іншому аспекті, даний винахід забезпечує способи одержання трансгенної рослини, де вказані способи включають: (1) трансфектування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить неконсервативну мутантну послідовність фактору транскрипції білків "цинкові пальці" відповідно до даного винаходу або конструктом, що містить неконсервативну мутантну послідовність полінуклеотиду відповідно до даного винаходу; (2) відбір рослинної клітини, рослинної тканини або органу рослини, що містить неконсервативну мутантну послідовність; та (3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини, одержаних на стадії (2), Де одержана трансгенна рослина має більш високу толерантність до засухи та дії солі ніж нетрансгенна рослина. В іншому переважному втіленні, вказані способи також включають кросбридинг одержаної трансгенної рослини з нетрансгенної рослини або іншої трансгенної рослини, таким чином одержуючи гібридне потомство, що містить неконсервативну мутантну послідовність, де вказане гібридне потомство має більш високу толерантність до засухи та дії солі, ніж нетрансгенна рослина, переважно вказане гібридне потомство має стабільні генетичні ознаки. В іншому переважному втіленні, вказані способи також включають розробку молекулярного маркера, специфічного до неконсервативної мутантної послідовності, для скринінгу потомства, одержаного від кросбридингу трансгенної рослини з одержанням рослини, що має покращену толерантність до засухи та дії солі. Виходячи з даного опису, інші аспекти даного винаходу будуть очевидними фахівцю у цій галузі. ОПИС КРЕСЛЕНЬ ФІГ. 1: Генна послідовність DST рису (ФІГ. 1A) та її кодована амінокислотна послідовність (ФІГ. 1B). ФІГ. 2: Порівняння фенотипів генного мутанту dst рису DST та фенотипів рису дикого типу за умов засухи та дії солі. В кожній панелі, дикий тип (Zhonghua 11, ZH11) зображено зліва, а мутант dst - справа. ФІГ. 3: Порівняння фенотипів, за умов засухи та дії солі, мутанту dst дикого типу, одержаного DST генною комплементацією, та рослин зі зниженою DST функцією за допомогою РНК інтерференції. ФІГ. 4: Аналіз DST транскрипційної активації із застосуванням двогібридної системи TM дріжджів Matchmaker GAL4 3 (Clontech). ФІГ. 5: Результати аналізу вимірювання електрофоретичної рухливості (EMSA). ФІГ. 6: Аналіз вирівнювання послідовностей гомологічних доменів білків DST "цинкові пальці" серед культур зернових. ДЕТАЛЬНИЙ ОПИС ВИНАХОДУ Після довгого та інтенсивного дослідження, винахідники даного винаходу відкрили новий ген фактору транскрипції білків "цинкові пальці" DST (ген толерантності до засухи та дії солі) рису та підтвердили, що цей ген є негативним регуляторним фактором толерантності до засухи та дії солі, здатним регулювати толерантність до засухи та дії солі у рослин, та що інгібування 4 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 експресії даного гена може підвищувати резистентність до стресу від дії солі та засухи у рослин. Тому цей ген відіграє важливу роль у розведенні рослин, резистентних до засухи та дії солі. Виходячи з цього, винахідники втілили свій винахід на практиці. Конкретно, застосовуючи мутантну бібліотеку рису (EMS мутагенез) та методики клонування за допомогою картування, винахідники здійснили великомасштабний скринінг за умов стресу від дії солі з одержанням нового гену DST, що регулює толерантність до засухи та дії солі у рису. Довжина геномного DST гену становить 906bp, що не включає жодного інтрону. Тому, повнорозмірна ORF (відкрита рамка зчитування) становить 906bp. Цей ген кодує 301 амінокислот, білок, що має приблизно 29КДа, що містить консервативний домен "цинкових пальців". Цей білок являє собою фактор транскрипції. Результати фенотипної ідентифікації демонструють, що мутанти цього гену (наприклад, DST гену з нуклеотидними мутаціями, що призводить до 2 амінокислотних заміщено) проявляють як толерантність до засухи, так і до дії солі. За допомогою РНК інтерференції для пригнічення експресії цього гена також одержують підвищену толерантність до засухи та дії солі. Результати біохімічні досліджень демонструють, що DST є фактором транскрипції, що містить не лише домен транскрипційної активації, але також домен ДНК зв'язування. Аналіз генного чипу демонструє, що DST функціонує як фактор транскрипції, що регулює серію нижніх генів. Функціональні дослідження демонструють, що, порівняно з диким типом, мутанти мають більшу кількість пероксиду водню (H2O2), що накопичується біля продиху, мають менші продихові отвори, що дозволяє листю зберігати відносно більший рівень вологи за умов засухи. Тому, мутанти мають вищу толерантність до засухи. Додатково, через менші продихові отвори у мутантів, продихова провідність є більш низькою, та більш низькою є швидкість утворення + водяної пари. В результаті, транспорт Na іонів з коренів до частин над ґрунтом (листя, та інш.) + зменшено, та тому Na токсичність знижена, таким чином підвищуючи толерантність до дії солі. Це дослідження демонструє, що DST задіяний у регуляції генів, пов'язаних із перокидазою, регулює накопичення пероксиду водню (H2O2), регулює продиховий отвір, таким чином впливаючи на толерантність до засухи та дії солі у рису. Вищенаведені дослідження демонструють, що ген DST є фактором негативної регуляції для резистентності до засухи та резистентності до дії солі, інгібування його експресії може підвищити резистентність до стресу від дії солі та засухи у рослин. Така властивість може бути застосована для одержання трансгенних рослин зі значно вищою резистентністю до стресу від дії солі та засухи. Таким чином, DST ген має великий потенціал для покращення здатності зернових переносити несприятливі стреси, такі, як стреси від дії солі та від засухи. Додатково пошук у базах даних виявив: один DST гомологічний ген у геномі сорго (Сорго двокольоровий) з подібністю білків 54,3 %; три DST гомологічних гена у геномі кукурудзи (Zea mays) з подібністю білків 51,7 %, 36,1 %, та 33,5 %; один DST гомологічний ген у геномі ячменю (Hordeum vulgare) з подібністю білків 38,4 %; та три DST гомологічних гена у геномі цукрової тростини (Saccharum officinarum) з подібністю білків 38,2 %, 38,2 % та 34,5 %. Всі ці гомологічні гени мають консервативний C2H2-типу структурний домен "цинкових пальців", що у високому ступеню подібний при N кінці, передбачаючи, що DST гомологічні гени в інших рослинах (переважно, зернових) будуть мати функції, подібні до функцій DST гену рису. DST БІЛКИ або ПОЛІПЕПТИДИ ТА ЇХ КОДУЮЧИ ПОСЛІДОВНОСТІ У даному винаході, терміни "DST білки або поліпептиди, " "DST ген, кодований білками або поліпептидами, " або "фактор транскрипції білків "цинкові пальці»" відноситься до білків або поліпептидів, кодованих DST генами відповідно до даного винаходу. Ці визначення включають мутанти описаних вище білків або поліпептидів консервативними мутаціями, або їх гомологічні поліпептиди. Всі вони мають Cys-2/His-2 типу структурний домен типу "цинкових пальців", та якщо інгібується експресіявказаних білків або поліпептидів, то резистентність рослин до стресів від дії засухи або дії солі може бути підвищена. В одному втіленні даного винаходу, вказаний фактор транскрипції задіяний у регуляції генів, зв'язаних із пероксидазою, регулює накопичення пероксиду водню та/або регулює вустичний отвір, таким чином, впливаючи на толерантність до засухи та дії солі у рису. Вказані DST білкові або поліпептидні послідовності вибирають з: (a) поліпептидів, що мають амінокислотну послідовність SEQ ID NO: 2; (b) поліпептидів, одержаних від (a), що мають одну або більше заміщень амінокислотних залишків, делецій або інсерцій у амінокислотній послідовності SEQ ID NO: 2, та здатний до підвищення чутливості до засухи або дії солі у рослин; або (c) поліпептидних гомологів поліпептидів (a) або (b), що мають Cys-2/His-2 типу структурний домен типу "цинкових пальців" та здатні до підвищення чутливості до засухи або дії 5 UA 106489 C2 5 10 15 20 25 солі у рослин. Переважно, вказані білки або поліпептиди можуть зв'язуватися із TGCTANN(A/T)TTG, де N являє собою A, C, G або T. Білки та поліпептиди відповідно до даного винаходу можуть бути очищеними природними продуктами, або хімічно синтезованими продуктами, або одержаними, за допомогою рекомбінантної технології, з прокаріотних або еукаріотних клітин-хазяїв (наприклад, бактерій, дріжджів, вищих рослин, комах та клітин ссавців). DST білки або поліпептиди відповідно до даного винаходу, переважно є кодованими геном зернових (переважно, рису) DST або його гомологічними генами або сімейством генів. Типи мутацій у білках або поліпептидах відповідно до даного винаходу включають, не обмежуючись наведеним: делецію, інсерцію та/або заміщення однієї або більше (зазвичай 1-50, переважно 1-30, більш переважно, 1-20, найбільш переважно 1-10, наприклад, 1, 2, 3, 4, 5, 6, 7, 8, 9, або 10) амінокислот, та додавання на C кінці та/або N кінці однієї або декількох (зазвичай 20 або менше, переважно менш ніж 10, найбільш переважно менш ніж 5) амінокислот. Наприклад, з рівня техніки відомо, що, при заміщенні амінокислот, що мають зв'язані або подібні властивості, функції білків або поліпептидів зазвичай не змінюються. Як інший приклад, додавання однієї або більше амінокислот на C кінці та/або N кінці зазвичай не змінює функції білків або поліпептидів. Наприклад, DST білки або поліпептиди відповідно до даного винаходу можуть містити, а може не містити вихідний метіоніновий залишок та зберігають активність для підвищення резистентності до стресу, викликаного дією важких металів або солей у рослин. Фахівець у цій галузі, виходячи із загальних знань у цій галузі та/або рутинних експериментів, може легко ідентифікувати ці різні типи мутацій, що не впливатимуть на активність білків та поліпептидів. У даному винаході, термін "консервантивні мутантні поліпептиди" відноситься до поліпептидів, порівняно із амінокислотною послідовністю SEQ ID NO: 2, що має аж до 20, переважно аж до 10, більш переважно, аж до 5, найбільш переважно аж до 3 амінокислот, заміщених амінокислотами, що мають зв'язані або подібні властивості. Такі консервантивні мутантні поліпептиди можуть бути найкращим чином одержані відповідно до наступної таблиці амінокислотних заміщень: Таблиця Амінокислотні залишки Ala (A) Arg (R) Asn (N) Asp (D) Cys (C) Gln (Q) Glu (E) Gly (G) His (H) Ile (I) Leu (L) Lys (K) Met (M) Phe (F) Pro (P) Ser (S) Thr (T) Trp (W) Tyr (Y) Val (V) Репрезентативні заміщення Val; Leu; Ile Lys; Gln; Asn Gln; His; Lys; Arg Glu Ser Asn Asp Pro; Ala Asn; Gln; Lys; Arg Leu; Val; Met; Ala; Phe Ile; Val; Met; Ala; Phe Arg; Gln; Asn Leu; Phe; Ile Leu; Val; Ile; Ala; Tyr Ala Thr Ser Tyr; Phe Trp; Phe; Thr; Ser Ile; Leu; Met; Phe; Ala Переважні заміщення Val Lys Gln Glu Ser Asn Asp Ala Arg Leu Ile Arg Leu Leu Ala Thr Ser Tyr Phe Leu 30 35 Вказані білки та поліпептиди від (b) можуть бути одержані шляхом піддавання дії випромінювання або мутагенів з одержанням випадкового мутагенезу, або через сайтнаправлений мутагенез або за допомогою інших відомих методів молекулярної біології. Послідовності, що кодують білки або поліпептиди, можуть бути застосовані для конструювання трансгенних рослин для скринінгу та ідентифікації білків або поліпептидів, виходячи з того, або мають трансгенні рослини змінені характеристики. 6 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 Мутантні форми вказаних поліпептидів включають: гомологічні послідовності, консервантивні мутанти, алельні мутанти, природні мутанти, індуковані мутанти, білки, кодовані послідовностями, що можуть гібридизувати кодуючими послідовностями для DST білка за умов високої або низької жорсткості, та поліпептиди або білки, одержані за допомогою анті-DST білковою антисироваткою. Інші поліпептиди можуть бути також використані у даному винаході, такі, як гібридизаційні білки, що містять DST білок або його фрагмент. Додатково до майже повнорозмірних поліпептидів, даний винахід також включає розчинні фрагменти DST білків. Загалом, вказані розчинні фрагменти містять щонайменше приблизно 10 послідовних амінокислот у DST білковій послідовності, зазвичай щонайменше приблизно 30 послідовних амінокислот, переважно щонайменше приблизно 50 послідовних амінокислот, більш переважно, щонайменше приблизно 80 послідовних амінокислот, найбільш переважно щонайменше приблизно 100 послідовних амінокислот. В залежності від хазяїв, які застосовують для одержання рекомбінантів, білки або поліпептиди відповідно до даного винаходу можуть бути гліколізовані або можуть бути негліколізовані. Термін також включає активні фрагменти та активні похідні DST білків. У даному описі, терміни "DST ген, " "рослинний DST ген, " або "кодуючі послідовності факторів транскрипції відповідно до даного винаходу" є взаємозамінними. Всі вони відносяться до послідовностей, що кодують DST білки або поліпептиди відповідно до даного винаходу. Вони високогомологічні до генної послідовності DST рису (див. SEQ ID NO: 1); вони являють собою молекули, що можуть гібридизувати зі вказаною генною послідовністю за жорстких умов; або вони являють собою молекули сімейства генів, високогомологічні вказаним молекулами. Інгібування вказаної генної експресії призводить до визначного покращення резистентності до стресі, викликаного дією засухи або солі у рослин. В одному втіленні даного винаходу, вказаний полінуклеотид включає: (a) нуклеотидні послідовності SEQ ID NO: 1; (b) нуклеотидную послідовність, що містить нуклеотиди 1-435 SEQ ID NO: 1; або (c) полінуклеотиди. Комплементарні одній з нуклеотидних послідовностей у (a)-(b). У даному описі, термін "жорсткі умови" стосується: (1) гібридизації та промивання за низької іонної сили та високих температурах, наприклад 0,2× SSC, 0,1 % SDS (натрій додецилсульфату), 60°С; або (2) гібридизації у присутності денатуруючого агента, такого, як 50 % (об./об.) формамід, 0,1 % теляча сироватка/0,1 % Фіколл, 42°С, та інш.; або (3) гібридизації, що відбувається тільки якщо гомологія між двома послідовностями досягає щонайменше 50 %, переважно 55 % або більше, 60 % або більше, 65 % або більше, 70 % або більше, 75 % або більше, 80 % або більше, 85 % або більше, або 90 % або більше, більш переважно, 95 % або більше. Наприклад, вказані послідовності можуть бути послідовностями, комплементарними послідовностям, визначеним в (a). Повнорозмірні або фрагменти DST генних нуклеотидних послідовностей відповідно до даного винаходу можуть бути зазвичай одержані за допомогою ПЦР ампліфікації, рекомбінантних або синтетичних способів. Для ПЦР ампліфікації, родинні послідовності можуть бути одержані шляхом розробки праймерів, виходячи з родинних нуклеотидних послідовностей, описаних у даному винаході, конкретно відкритої рамки зчитування, та з використанням комерційно доступних кДНК бібліотек або кДНК бібліотек, генерованих традиційними способами, відомих фахівцю у цій галузі як темплати. При розгляді довгих послідовностей, зазвичай необхідні дві або більше ПЦР ампліфікації, та потім зборка фрагментів, одержаних з ампліфікації відповідно до правильних порядків. Необхідно розуміти, що ген DST відповідно до даного винаходу одержують переважно з рису. Інші гени, які одержують з інших рослин, що мають високу гомологічність до гену DST рису (наприклад, 50 % або більше, переважно 55 % або більше, 60 % або більше, 65 % або більше, 70 % або більше, 75 % або більше, 80 % або більше, більш переважно, 85 % або більше, наприклад, 85 %, 90 %, 95 % або навіть 98 % ідентичностей послідовностей) також розглядають як такий, що входить до обсягу даного винаходу. Способи та інструменти для порівняння ідентичності послідовностей також добре відомі з рівня техніки, наприклад, BLAST. РОСЛИНИ ТА ЇХ РЕЗИСТЕНТНІСТЬ ДО СТРЕСУ ПІД ДІЄЮ СОЛІ ТА/АБО ЗАСУХИ Як вживають у даному описі, вказані "рослини" включають (не обмежуючись наведеним): зернові, рослини бавовни сімейства мальвових, капусту сімейства хрестоцвітних, складноцвітні, пасльонові, губоцвітні рослини, або зонтичні, та інш. Переважно, вказані рослини є зерновими рослинами, більш переважно, культурами зернових. Наприклад, вказані рослини можуть бути вибрані з: рису, кукурудзи, пшениці, ячменю, цукрової тростини, сорго, Арабідопсису, бавовни або каноли, більш переважно, рису, кукурудзи, пшениці, ячменю, цукрової тростини або сорго. Як вживають у даному описі, термін "зернові" відноситься до рослин, що мають промислову цінність зерновому, бавовняному, олійному та інш. сільському господарстві та промисловості. 7 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 Промислова цінність може бути відображена насінням рослин, плодами, коренями, стеблинами, листям та іншими корисними частинами. Культури включають, не обмежуючись наведеним: дводольні або однодольні. Переважними однодольними рослинами є зернові рослини, більш переважно, рис, пшениця, ячмінь, кукурудза, сорго та інш. Переважні дводольні рослини включають, не обмежуючись наведеним: рослини бавовни сімейства мальвових, хрестоцвітні рослини, наприклад, капуста, більш переважно, бавовна та канола. Як вживають у даному описі, термін "стрес під дією солі" відноситься до: явища, при якому, при зростанні рослин у ґрунті або воді, що містить високі концентрації солей, їх зростання буде інгібовано або навіть вони помруть. Солі, що викликають сольовий стрес, включають (не обмежуючись наведеним): хлорид натрію, сульфат натрію, карбонат натрію або бікарбонат натрію. DST гени відповідно до даного винаходу або їх кодовані білки або поліпептиди можуть підвищувати резистентність до стресу під дією солі у рослин. Підвищена резистентність може спостерігатися шляхом порівняння з контрольними рослинами, які не обробляли вказаними генами, білками, або поліпептидами. На зростання та розвиток вказаних рослин не впливають або меншою мірою впливають високі концентрації солей, або вказані рослини можуть виживати при більш високих концентраціях солей. Як вживають у даному описі, термін "стрес від засухи" відноситься до: явища, при якому, коли рослини зростають у сухому ґрунті або іншому середовищі засухи, їх зростанні буде інгібовано або навіть вони помруть. DST гени відповідно до даного винаходу або їх кодовані білки або поліпептиди можуть підвищувати резистентність рослин до стресу від засухи, підвищена резистентність може спостерігатися шляхом порівняння з контрольними рослинами, які не обробляли вказаними генами, білками, або поліпептидами. На зростання та розвиток вказаних рослин не впливає або меншою мірою впливає відсутність води, або вказані рослини можуть виживати при більш жорстких умовах засухи. ВЕКТОРИ, ХАЗЯЄВА ТА ТРАНСГЕННІ РОСЛИНИ Даний винахід також відноситься до векторів, що містять DST гени, та до клітин-хазяїв, що містить вказані вектори, одержані методами генної інженерії, та до трансгенних рослин, одержаних шляхом генної трансфекції та експресії високих рівнів DST. За допомогою традиційної рекомбінантної ДНК технології (Science, 1984; 224:1431), кодуючі послідовності відповідно до даного винаходу можуть бути застосовані для експресії або продукування рекомбінантних DST білків. Загалом, це включає такі стадії: (1) трансфектування або трансформація придатних клітин-хазяїв полінуклеотидами (чи мутантами), що кодують DST білки відповідно до даного винаходу, або рекомбінантними векторами експресії, що містять вказані полінуклеотиди; (2) культивування клітин-хазяїв в придатних культуральних середовищах; та (3) виділення та очистку білків або поліпептидів з культурального середовища або з клітин. У даному винаході, терміни "вектори" та "вектори рекомбінантної експресії" можуть бути застосовані взаємозамінно, та вони стосуються бактеріальних плазмідів, бактеріофагів, дріжджових плазмідів, вірусів рослинних клітин, вірусів клітин ссавців або інших векторів, що добре відомі з рівня техніки. Стисло, можуть бути застосовані будь-які плазміни або вектори, оскільки вони можуть реплікувати та є стабільними зсередини клітин-хазяїв. Однією важливою ознакою векторів експресіїi є те, що вони зазвичай містять джерело реплікації, промотор, маркерний ген та елемент контролю трансляції. Для конструювання векторів експресії, що містять DST кодуючи послідовності та придатний контрольний сигнал транскрипції/трансляції, можуть бути використані способи, добре відомі фахівцю у цій галузі. Ці способи включають in vitro рекомбінантну ДНК технологію, технологію синтезу ДНК, та in vivo рекомбінантну технологію, та інш. Вказані ДНК послідовності можуть бути ефективно зв'язані з придатними промоторами в векторах експресії, направляючи синтез іРНК. Вектори експресії також включають сайти рибосомного зв'язування для сайтів початку трансляції та припинення транскрипції. У даному винаході, переважно застосовують ПЕГFP-1, pBI121, pCAMBIA1300, pCAMBIA1301, pCAMBIA2301 або pHB. Додатково, вектори експресії переважно включають один або більше генів маркерної селекції, забезпечуючи фенотипи для відбору трансфектованих клітин-хазяїв, таких, як дигідрофолат редуктаза, резистентність до неоміцину та зеленого флуоресцентного білку(GFP) для застосування в еукаріот ній клітинній культурі, або резистентність до тетрацикліну або ампіциліну для застосування у кишковій паличці. Вектори, що містять описані вище ДНК послідовності, та придатні промотори або контрольні елементи можуть бути застосовані для трансформації придатних клітин-хазяїв, дозволяючи ним експресувати білки або поліпептиди. Клітини-хазяїни можуть бути прокаріотними клітинами, такими, як бактеріальні клітини; або нижчими еукаріотними клітинами, такими, як дріжджові 8 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 клітини; або вищими еукаріотними клітинами, такими, як рослинні клітини. Репрезентативні приклади включають: кишкову паличку, стрептоміцети, агробактерії; клітини грибів, такі, як дріжджі; рослинні клітини, та інш. У даному винаході, клітини-хазяїни являють собою переважно агробактерії. Для експресії полінуклеотидів відповідно до даного винаходу у вищих еукаріотних клітинах, транскрипція може бути підвищена, якщо енхансерні послідовності вводять у вектори. Енхансерами є ДНК цис-діючі фактори, зазвичай приблизно від 10 до 300 bp, що діють на промотори для підсилення генної транскрипції. Фахівець у цій галузі буде знати як відбирати придатні вектори, промотори, енхансери та клітини-хазяїни. Одержані трансформанти можуть бути культивовані за допомогою традиційних способів, що експресують поліпептиди, кодовані генами відповідно до даного винаходу. В залежності від клітин-хазяїв, культуральні середовища, які застосовують для культивування, можуть бути вибрані з будь-яких традиційних середовищ. Потім може бути проведено культивування за умов, придатних для зростання клітин-хазяїв. Коли клітини-хазяїни зростають до придатної клітинної щільності, застосовують придатні способи (наприклад, температурний зсув або хімічну індукцію) для індукування відібраних промоторів, та продовжують культивування протягом ще одного періоду часу. Рекомбінантні поліпептиди, одержані за допомогою вказаних вище способів, можуть бути експресовані у клітинах, або клітинній мембрані, або декретовані поза клітинами. У разі необхідності, рекомбінантні білки можуть бути виділені та очищені за допомогою різних с способів виділення, виходячи з їх фізичних, хімічних та інших властивостей. Такі способи добре відомі фахівцю у цій галузі. Приклади цих способів включають, не обмежуючись наведеним: традиційну денатураційну обробку, обробку агентами осадження білків (спосіб висалювання), центрифугування, осмоліз бактерій, обробку ультразвуком, ультрацентрифугування, хроматографію молекулярних сит (гель-фільтрацію), адсорбційну хроматографію, іон-обмінну хроматографію, високоефективну рідинну хроматографію (ВЕРХ) та різні інші технології рідинної хроматографії та їх комбінації. Трансформування рослин може бути досягнуто за допомогою трансформації агробактерій або трансформаціє генної гармати, та інш., наприклад агробактерія-опосередковану трансформацію пластин листя. Трансформовані рослинні клітини, тканини або органи можуть бути регенеровані у рослинах за допомогою традиційних способів, що призводить до одержання рослин з покращеною резистентністю до захворювань. ЦИС-ДІЮЧІ ЕЛЕМЕНТИ Як застосовують у цьому описі, термін "цис-діючі елементи" відноситься до послідовностей, розташованих у фланкуючих областях генів та вони можуть впливати на генну експресію. Їх функціями є участь у регуляції генної експресії. Цис-діючий елемент сам по собі не кодує жодний білок; а тільки забезпечує діючий сайт. Він взаємодіє з транс-діючими факторами для досягнення їх функцій. При дослідження, винахідники даного винаходу відкрили: DST білки відповідно до даного винаходу мають здатність ДНК зв'язування Їх основні елементи зв'язування являють собою цисдіючі елементи, TGCTANN(A/T)TTG, де N являє собою A, C, G або T. Цис-діючі елементи відповідно до даного винаходу зв'язують DST фактори транскрипції, підвищуючи чутливість до засухи та дії солі у рослин, таким чином, знижуючи толерантність до засухи та дії солі у рослин. Вказані цис-діючі елементи переважно взаємодіють з доменом "цинкових пальців" DST. Навпаки, якщо взаємодії між цис-діючими елементами та DST факторами транскрипції відповідно до даного винаходу інгібовано, це призведе до підвищеної толерантності до засухи та дії солі у рослин. СПОСОБИ ПІДВИЩЕННЯ ТОЛЕРАНТНОСТІ ДО ЗАСУХИ ТА ДІЇ СОЛІ У РОСЛИН Як описано у даному описі, DST білки відповідно до даного винаходу, їх кодуючі послідовності або зв'язування DST білків з цис-діючими елементами мають тісний зв'язок із толерантністю до засухи та дії солі у рослин. Інгібування DST білків, їх кодуючи послідовностей або зв'язуванню DST білків із цис-діючими елементами призведе до підвищеної толерантності до засухи та дії солі у рослин. Тому, даний винахід також забезпечує способи підвищення толерантності до засухи та дії солі у рослин шляхом інгібування DST білків, їх кодуючи послідовностей, або зв'язування DST білків із цис-діючими елементами. В одному втіленні даного винаходу, антагоністи DST білків або їх кодуючі послідовності можуть бути застосовані для інгібування їх експресії. Вказані антагоністи включають, не обмежуючись наведеним: низькомолекулярні інтерферуючі РНК, домінантні негативні регулятори або антисмислові олігонуклеотиди. Фахівець у цій галузі, знаючи DST білки або їх 9 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 кодуючи послідовності, знатиме, як застосувати традиційні способи та тести для скринінгу та одержання вказаних антагоністів. Як вживають у даному винаході, термін "неконсервативна мутація" відноситься до одного або більше амінокислотного або нуклеотидного заміщення, делеції або інсерції (переважно, неконсервативної) у DST білку або його кодуючої послідовності, що призводить до підвищеної толерантності до засухи та дії солі у рослин. В іншому втіленні відповідно до даного винаходу, способи, відомі з рівня техніки, можуть бути застосовані для введення неконсервативних мутацій у DST білки відповідно до даного винаходу або їх кодуючи послідовності. Наприклад, нуклеотидні мутації у DST білкових кодуючи послідовностях можуть бути застосовані для введення неконсервативних мутацій у амінокислотних послідовностях DST білків, що містять SEQ ID NO: 2, забезпечуючи рослини, що містять мутантні послідовності, підвищеною толерантністю до засухи та дії солі. Наприклад, мутація при амінокислоті 69, з аспарагіну на аспарагінову кислоту, або мутація при амінокислоті 162, з аланіну на треонін. В іншому втіленні відповідно до даного винаходу, можуть бути одержані трансгенні рослини з інгібованою експресією DST генів або білків, та ці трансгенні рослини можуть бути необов'язково схрещені з нетрансгенними рослинами або іншими трансгенними рослинами. Наприклад, рослини можуть бути трансформовані векторами, що містять низькомолекулярні інтерферуючі РНК, анти смислові вектори, вектори домінантної негативної регуляції, специфічно направлені DST білки або їх кодовані білки або клітини-хазяїни, що містять вказані вектори. СПОСОБИ СКРИНІНГУ РОСЛИН, ТОЛЕРАНТНИХ ДО ЗАСУХИ ТА ДІЇ СОЛІ Відповідно до унікальних властивостей DST генів відповідно до даного винаходу та їх кодованих білків, даний винахід додатково включає способи скринінгу рослин, толерантних до засухи та дії солі. В одному втіленні, спосіб скринінгу відповідно до даного винаходу включає: (i) оцінку, у кандидатної рослини, рівня DST фактору транскрипції білків "цинкові пальці" відповідно до даного винаходу, рівню експресії його кодую чого полінуклеотиду, та/або рівню зв'язування цисдіючого елементу відповідно до даного винаходу із DST фактором транскрипції білків "цинкові пальці"; (ii) порівняння рівня, детектованого у кандидатної рослині на стадії (i) з відповідним рівнем у контрольної рослини, якщо рівень у кандидатній рослині нижчий за рівень у контрольної рослини, тоді кандидатна рослина є рослиною, толерантною до засухи та дії солі. В іншому втіленні, можуть бути застосовані методи відбору за допомогою молекулярних маркерів, відомі з рівня техніки, для введення толерантного до засухи та дії солі DST гену в інші варіанти для скринінгу та культивування нових варіантів, які толерантні до засухи та дії солі. Вказані способи можуть застосовувати традиційні способи кросбридингу. Його перевагою є те, що не потрібен генний перенос, уникаючи питань безпеки генного переносу. Вказані способи можуть включати: розробку молекулярних маркерів, специфічних для неконсервативних мутантних послідовностей, застосування вказаних молекулярних маркерів для скринінгу потомства від кросбридингу мутантів, що мають неконсервативні мутантні послідовності та інші варіанти рису, таким чином відбираючи окремі рослини, що містять вказані неконсервативні мутантні послідовності. ОСНОВНІ ПЕРЕВАГИ ДАНОГО ВИНАХОДУ Основні переваги даного винаходу включають: (1) ідентифікацію DST генів та їх кодованих білків або поліпептидів, та підтвердження їх зв'язку із толерантністю до засухи та дії солі у рослин, таким чином забезпечуючи нові способи дослідження толерантності до засухи та дії солі у рослин; (2) забезпечення трансгенних рослин, що мають підвищену резистентність до дії солі та засухи, таким чином забезпечуючи відмінну сировину та продукти для одержання та переробки зернових, бавовни та олії; та (3) забезпечення способів скринінгу для толерантного до дії солі та засухи потомства за допомогою молекулярних маркерів, що можуть бути реалізовані за допомогою традиційних способів кросбридингу, без потреби у переносі, таким чином, уникаючи проблем з безпекою генного переносу. Даний винахід забезпечує нові підходи для покращення резистентності до стресу від дії солі та засухи у рослин з великим потенціалом до застосування. ПРИКЛАДИ Наведений у цій заявці опис, у сполученні з конкретними прикладами, додатково ілюструє даний винахід. Необхідно розуміти, що ці приклади застосовують для пояснення даного винаходу та вони не повинні бути використані для обмеження обсягу даного винаходу. 10 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 У наступних прикладах, коли умови не вказані в експериментальних способах, вони зазвичай базуються на традиційних умовах (наприклад, див., Sambrook et al, "Molecular Cloning: A Laboratory Manual”, third edition, 2001, Cold Spring Harbor Laboratory Press) або умовах відповідно до пропозицій виробника. Якщо не вказано інше, процентні вмісти та співвідношення розраховують, виходячи з мас. Якщо не визначено інше, усі професійні та наукові терміни, вжиті у даному описі, мають ті ж значення, що добре відомі фахівцю у цій галузі. Додатково, будь-які способи та матеріали, подібні або еквівалентні описаним у даному описі, можуть бути застосовані у даному винаході. Переважні способи та речовин, описані у даному описі, застосовують тільки для ілюстрації. Різні середовища, що застосовують у прикладах (YEB рідке культуральне середовище, AB рідке культуральне середовище, AAM рідке культуральне середовище, N6D2 культуральне середовище, N6D2C культуральне середовище, спів-культуральне середовище, селекційне культуральне середовище N6D2S1, N6D2S2, пре-диференційне культуральне середовище, диференційне культуральне середовище, 1/2 MS0H культуральне середовище, культуральне середовище рису, SD культуральне середовище, та інш.) одержують відповідно до описів у зв'язаній літературі (Molecular Cloning: Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989; Hiei, Y., та інш., Plant J., 1994, 6, 271-282). ПРИКЛАД 1: ЕКСПЕРИМЕНТИ З DST ГЕННОГО ПЕРЕНОСУ РИСУ 1. Одержання DST мутантів, що мають високу толерантність до засухи та дії солі, їх характеристики та субклітинна локалізація. Насіння рису обробляли 0,6 % EMS (етил метансульфонат) для конструювання бібліотеки мутантів рису, що містить приблизно 9,000 мутантних ліній рису. Великомасштабний скринінг бібліотеки мутантів рису проводили за умов стресу, викликаного дією солі 140мМ хлориду натрію. Фенотип з толерантністю до дії солі та засухи перевіряли шляхом піддавання кандидат них мутанти повторному стресу від діє солі 140мМ хлориду натрію та 20 % ПЕГ4000 модельованому стресу від засухи. Одержували мутант з високою толерантністю до дії засухи та дії солі (dst). Із застосуванням молекулярних маркерів, DST ген попередньо розташований на хромосомі рису 3. Шляхом кросбридингу конструюють dst мутант із сіль-чутливими штамами, конструюють крупномасштабне F2 потомство. Із застосуванням молекулярних маркерів для скринінгу для кросбридингового потомства з групи, скомбінованої з генотипами та фенотипами росбридингового потомства, виконували клонування на основі картування. Це призвело до успішного клонування DST гену. Вказаний DST ген кодує білок "цинкові пальці" (фактор транскрипції) невідомої функції, що має консервативний C2H2 типу домен "цинкових пальців". Жодні інші DST гомологічні копії не знайдені у геномі рису, та жодний гомологічний ген на знайдений у геномі Арабідопсису. Довжина вказаного геномного гену становить 906 bp, без інтронів. Повнорозмірна ORF (відкрита рамка зчитування) має довжину 906 bp, кодуючи 301 амінокислот. Молекулярна маса білкового продукту, як оцінили, становила 29КДа (ФІГ. 1). Аналіз порівняння послідовностей демонструє, що DST ген у цьому мутанті містить 2 нуклеотидні мутації, що призводить до 2 амінокислотних заміщень (амінокислота 69 мутувала з аспарагіну на аспарагінову кислоту, та амінокислота 162 мутувала з аланіну на треонін) та призвело до утворення фенотипу, резистентного до засухи та дії солі. Це спостереження вказує на те, що DST є негативним регулятором резистентності до засухи та дії солі. Для визначення субклітинної локалізації DST, конструкт гібридизації DST та GFP (зеленого флуоресцентного білка) був одержаний та перенесений у епідермальні клітини цибулі за допомогою способу генної гармати для експресії, яку передають. Розташування флуоресценції всередині клітин досліджували за допомогою флуоресцентного конфокального мікроскопу. За допомогою дослідження субклітинної локалізації DST, як знайдено, розташований конкретно в ядрі. 2. Конструювання трансгенних плазмідів, що містять DST геномні фрагменти: Клони рису дикого типу BAC дигестували ApaLI рестриктазою, а потім T4 ДНК полімеразою з одержанням тупих кінців, які потім дигестували SalI рестриктазою. Потім відновлювали геномний фрагмент дикого типу 4.6-kb (що містить повнорозмірну ORF DST, промоторний регіон та стоп кодон з нижнім регіоном). Бінарний вектор рослинної експресії pCAMBIA1301 (придбаний від CAMBIA) дигестували EcoRI, а потім T4 DNA полімеразою з одержанням тупих кінців, які дигестували SalI а потім лігували відновлені фрагменти, вказані вище, для успішного конструювання p-DST плазміда, який застосовують для трансформації мутантів та проведення комплементарних експериментів. Усі ферменти придбають від New England Biolabs. 3. Конструювання плазмидів експресії DST-РНК інтерференції: 11 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 Застосування олігонуклеотидів на 5' та 3' кінцях як праймерів (SEQ ID NO: 4 та 5) для ампліфікації DST фрагмента, що має унікальний кодуючий регіон (535-bp) за допомогою ПЦР. Лігування цього фрагменту вектором інтерференції p1300РНК (одержаного шляхом модифікації pCAMBIA1300 через введення каталазного нітрону як лінкера, фланкованого полі-A та полі-T на обох кінцях) для конструювання DST-РНК інтерференційного плазміда. 5' олігонуклеотидна праймерна послідовність являє собою: 5'-AAGCTTTCCTTGCGAAGCCAAATAGC-3' (SEQ ID NO. 4) 3' праймерна послідовність являє собою: 5'-GGATCCCGAGGCTCAAGTTGAGGTCGA-3' (SEQ ID NO. 5) 4. DST трансгенного рису: Два рекомбінантні плазміни, одержані вище, переносили у штам агробактерій EHA105 за допомогою способу заморожування-відтаювання. Додавали 0,5-1 мкг (приблизно 10 мкл) плазмідної ДНК у кожні 200 мкл EHA105 компетентні клітини, змішували, та поміщали послідовно у крижу, у рідкий азот та у 37°С водяну баню на 5 хвилин кожна. Реакційну суміш розводили до 1 мл свіжим YEB рідким культуральним середовищем та потім інкубували при струшуванні при 28°С протягом 2-4 годин. Відбирали 200 мкл аліквоти та поміщали у YEB планшету, що містила канаміцинові (Kan) антибіотики (50 мкг/мл). Інкубували планшету при 28°С протягом 2-3 днів. Штрихували одержані колонії тричі на YEB планшетах, що містили Kan (50 мкг/мл) для відбору одинарних колоній. Збирали одинарні колонії агробактерій з YEB планшети та інокулювали в 3 мл YEB рідкого культурального середовища, що містило g 50 мкг/мл Kan антибіотиків та інкубували при струшуванні при 28°С протягом ночі. На день 2, переносили 1 % інокулюм до 50 мл AB рідкого середовища, що містив 50 мкг/мл Kan антибіотиків та продовжували інкубацію при струшуванні при 200 об./хв., доки OD600 досягне приблизно 0,6-0,8. Центрифугували свіжі агробактеріальні культури при 5000 об./хв. та 4°С протягом 5 хвилин. Збирали та повторно суспендували та гранулювали у 1/3 об'ємі AAM рідких культуральних середовищ. Ця суспензія можу бути застосована для трансформації різних рецепієнтних речовин рису. Цей експеримент застосовує традиційний агробактерії-опосредекований спосіб трансформації для трансформації зародкового каллюса рису Zhonghua 11 (або його мутантів). Завантажували недозріле насіння Zhonghua 11 (через 12-15 днів після запилення) у 70 % етанол протягом 1 хвилини, стерилізували них в NaClO розчині (змішували з водою при 1:3, додавали 2-3 краплини Tween 20) протягом 90 хвилин або більше, та промивали насіння стерильною водою 4-5 разів. Потім, зародки насіння збирали за допомогою скальпелю та пінцету та поміщали на N6D2 культуральне середовище для індукування утворення калюжної тканини, шляхом культивування при 26±1°С, у темряві. Через 4 дня, вони готові для трансформації. Зануряли одержані зародкові калюсні тканини у свіже AAM агробактеріальне рідке середовище при частому струшуванні. Видаляли речовину рису через 20 хвилин, застосовували стерильні фільтрувальні папери для видалення надлишкового бактеріального розчину, потім переносили на N6D2C культуральне середовище, покрите стерильним фільтрувальним папером, та продовжували при 26°С протягом 3 днів. Додавали ацетосирінгон до співкультурального середовища як генного активатора Vir, при концентрації 100 мкмоль/л. Через 3 днів, видаляли калюсні тканини зі співкультурального середовища, видаляли ембріони та переносили їх до N6D2S1 селекційного середовища (N6D2 середовища, що містило 25 мг/л Hyg) для відбору. Через 7-12 днів, переносили резистентні калюсні тканини до N6D2S2 (N6D2 середовища, що містило 50мг/л Hyg) селекційного середовища та продовжували відбір. Через 10-12 днів, переносили сильно зростаючі резистентні калюсні тканини до предиференційного культурального середовища та інкубували протягом приблизно 1 тижня. Потім, переносили їх до диференційного культурального середовища, дозволяючи їх диференціацію (12 годин світла/день). Після того, як регенероване насіння проростало у 1/2 MS0H культуральному середовищі, їх переносять у грунт у горщиках та зрощують у камерах з кліматконтролем. Після того, як регенеровані рослини вижили після трансплантації, ідентифікували позитивні трансгенні рослини за допомогою способів, відомих з рівня техніки, шляхом детекції βглюкозидази (бета-глюкоронідаза, GUS, див. Jefferson et al. EMBO J. 6, 3901-3907, 1987) або шляхом нанесення на листя з 0,1 % гербіцидом. Екстрагували весь ДНК з листя позитивних трансгенних рослин, та застосовували ПЦР для додаткової перевірки цих трансгенних рослин. У наступних експериментах, T2 покоління трансгенних рослин, одержаних вищезазначеними способами, застосовували для вивчення фенотипів, толерантних до засухи та дії солі, за умов стресу від дії засухи та солі, обробок (140 мМ NaCl) для підтвердження функцій гену DST. 12 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 ПРИКЛАД 2: ТРАНСГЕННА РОСЛИНА ТА КУЛЬТИВУВАННЯ ТА АНАЛІЗ НА СТРЕС ВІД ДІЇ СОЛІ ТА ВІД ЗАСУХИ Відбирали насіння трансгенного рису, одержаного у ПРИКЛАДІ 1, та інкубували його у печі при 45°С протягом тижня для порушення стану спокою. Потім просочували їх водопровідною водою при кімнатній температурі протягом 3 днів, та починали пророщування при 37°С протягом 2 днів. Після пророщування, висаджували паростки у планшети на 96 лунок. Потім, переносили їх у світові інкубатори, інкубували при 30°С, та піддавали дії світла протягом 13 годин на день. Через один день, поступово знижували температуру до 28°С та 26°С та інкубували їх протягом одного дня кожний та культивували їх при 20°С вночі. Після того, як паростки виросли, замінювали водопровідну воду а рисове культуральне середовище та продовжували культивування. Через приблизно 14 днів культивування, паростки пророщували до стану двох листків та одної серцевини. Піддавали їх сольовій обробці у рисовому середовищі, що містило 140 мМ NaCl протягом 12 днів, або ПЕГ обробці у рисовому середовищі, що містило 20 % (мас./об.) ПЕГ-4000 протягом 7 днів для стимулювання стресу від засухи. Щодо обробки засухою у ПВП трубках (полівінілпіролідонова трубка, 1,2 м заввишки, 20 см у діаметрі, два дренажні отвори на дні трубки), паростки трансгенної рослини проростали у водному інкубаторі протягом 25 днів у ПВП трубки, що містили грунт та культуру паростків у камері з клімат-контролем. Температура становила 24°С ~30°С, а вологість 50 %~60 %. Воду дренували протягом 30 днів після трансплантації, відкривали дренажні отвори на дні для дренажу води та виконували обробку засухою протягом 12 днів. ФІГ. 2 та ФІГ. 3 демонструють результати експериментів. Як показано на ФІГ. 2, рисовий мутант dst має значно вищу толерантність до засухи та дії солі, ніж дикий тип (Zhonghua 11, ZH11). Шляхом спостереження та порівняння, також знайдено: порівняно із диким типом, мутанти мають більше накопичення пероксиду водню (H2O2) навколо продиху, менші продихові отвори, відносно вищий вміст води у листя у стані стресу від засухи. Таким чином, мутанти мають вищу резистентність до засухи. Додатково, оскільки мутанти мають менші продихові отвори, меншу продихову провідність, та менші швидкості випаровування води, таким чином зменшуючи транспорт Na+ іонів з коренів до надземних частин (листя та інш.) та знижуючи токсичність Na+ та, тому, підвищуючи толерантність до дії солі. Як показано на ФІГ. 3: трансфектування DST геномного фрагмента від рису дикого типу (Zhonghua 11, ZH11) у dst мутант відновлює фенотип, чутливий до засухи та дії солі дикого типу у трансгенних комплементарних рослинах; тоді як рівень експресії DST зменшено РНК інтерференцією (трансформуванням Zhonghua 11), толерантності до засухи та дії солі у Zhonghua 11 значно підвищена. Ці результати демонструють: DST ген успішно клоновано та за допомогою методів генної інженерії за участю DST, резистентність до стресу у рису може бути значно підвищена. ПРИКЛАД 3: АНАЛІЗ DST ТРАНСКРИПЦІЙНОЇ АКТИВАЦІЙНОЇ АКТИВНОСТІ Маркер GAL4 для дріжджової двогібридної системи 3 (Clontech) застосовували для аналізу транскрипційної активності DST. Для конструювання позитивного регулюючого вектора pAD, NLSта GAL4 активуючі доменні (AD) послідовності ампліфікували за допомогою ПЦР та вводили у pGBKT7 (придбаний від Clontech) BamHI/SalI масковані сайти (праймерами є SEQ ID NO: 6 та 7) для гібридизації з GAL4 DNA доменом зв'язування (BD) in pGBKT7. Потім, ПЦР застосовували для ампліфікації DST повнорозмірної ORF (праймерами є SEQ ID NO: 8 та 9). Після підтвердження секвенуванням, ПЦР продукт конструювали у pGBKT7 вектор у BamHI та SalI сайти для гібридизації GAL4 DNA зв'язування домену з одержанням pGBKT7-DST вектору. Різні вектори потім трансформували у дріжджі AH109. Після зростання протягом ночі, культуру розводили та висаджували на планшети у SD культуральному середовищі без Trp або без трьох амінокислот (-Trp/-His/-Ade). Потім, спостерігали зростання дріжджів та визначали транскрипційну активаційну активність DST. Олігонуклеотидними праймерними послідовностями є: 5'-AAAGGATCCAAGCGGAATTAATTCCCGAG-3' (SEQ ID NO: 6); 5'-AAAGTCGACCCTCTTTTTTTGGGTTTGGTGG-3' (SEQ ID NO: 7); 5'-AAAGGATCCTGATGGACTCCCCGTCGCCT-3' (SEQ ID NO: 8); 5'-AAAGTCGACCGAGGCTCAAGTTGAGGTCGAG-3' (SEQ ID NO: 9). Результати наведено на ФІГ. 4. Як показано на фігурі, : рисовий DST білок має транскрипційну активаційну активність, тоді як мутантні DST білки та білки з N-кінцевою делецією втрачають транскрипційну активаційну активність. 13 UA 106489 C2 5 10 15 20 25 30 35 40 45 50 55 60 Результати вказують на те, що pGBKT7-DST має більш сильну транскрипційну активаційну активність та домен транскрипційної активаційної активності розташовано на N кінці, вказуючи на те, що DST є фактором транскрипції, що має транскрипційну активаційну активність. ПРИКЛАД 4: АНАЛІЗ DST БІЛКІВ ТА АНАЛІЗ ЗСУВУ ЕЛЕКТРОФОРЕТИЧНОЇ РУХЛИВОСТІ 1. Експресія прокаріот них білків: Дігестували вектор pGBKT7-DST EcoRI та SalI з одержанням DST повнорозмірної кДНК, яку потім реконструювали у pET32a(+). Трансфектували вектор прокаріотної експресії pET32a(+), що містить рекомбінантні DST, у BL21, застосовували IPTG для індукування прокаріотної експресії та потім очищували білки за допомогою His-tag колонок (основ). 2. Генерування антитіл: Імунізували кроликів очищеними білками, описаними вище, із застосуванням традиційних способів для генерування анти-DST антитіла. 3. Синтезували зонди, маркували них біотином, очищували на PAGE гелях та відновлювали марковані зонди електроелюцією. 4. Дозволяли маркованим пробам реагувати з очищеними прокаріотними експресованими DST білками. Потім, комплекси піддавали нативному PAGE електрофорезу та переносили на нейлонові мембрани за допомогою напівсухих способів переносу. Нейлонові мембрани потім піддавали дії рентгенівських плавок для проведення ауторадіографії для спостереження смуг зсуву. На ФІГ. 5 показані результати експериментів. Як показано на ФІГ. 5, DST має здатність до ДНК зв'язування. Основним елементом DST зв'язування є: цис-діючий елемент TGCTANN(A/T)TTG (SEQ ID NO: 3). Дане дослідження демонструє, що DST зв'язування із вказаним цис-діючим елементом може регулювати експресію нижніх генів, таким чином впливаючи на толерантність до засухи та дії солі у рослин. Тому, DST зв'язування із цис-діючим елементом відіграє важливу роль у негативній регуляції толерантності до засухи та дії солі. ПРИКЛАД 5: ІСНУВАННЯ DST ГЕННИХ ГОМОЛОГІВ У РІЗНИХ РОСЛИНАХ Пошук у базах даних (http://plantta.jcvi.org/index.shtml) виявив, один DST генний гомолог у геномі сорго (Сорго двокольоровий), з 54,3 % подібністю білків; три генних гомологи у геномі кукурудзи (Zea mays) з 51,7 %, 36,1 %, та 33,5 % подібністю білків; один DST генний гомолог у геномі ячменю (Hordeum vulgare), з 38,4 % подібністю білків; три генні гомологи у геномі цукрової тростини (Saccharum officinarum), з 38,2 %, 38,2 % та 34,5 % подібністю білків. Усі ці генні гомологи мали консервативні C2H2-типу домени типу "цинкових пальців". Вони мають однаковий домен типу "цинкових пальців". Одночасно, подібності є високими у Nкінцевих доменах (ФІГ. 6 демонструє аналогічну послідовність цих генних гомологів. Аналогічна послідовність являє собою DGKDVRLFPCLFCNKKFLKSQALGGHQNAHKKERSIGWNPYFYM, тобто, положення 42-85 у SEQ ID NO: 2). C2H2 типу білки "цинкові пальці" зв'язують цис-діючі елементи через домен типу "цинкових пальців". Тому, наявний відповідний зв'язок між цими генними гомологами та цис-діючими елементами, що вказує на те, що DST генні гомологи інших культур зернових можуть мати аналогічні функції, такі, як функції гену DST рису. ПРИКЛАД 6: ЗАСТОСУВАННЯ МЕТОДІВ ЗА ДОПОМОГОЮ ГЕННОГО DST МОЛЕКУЛЯРНОГО МАРКЕРУ РИСУ ТА ІНШИХ КУЛЬТУР ПРИ РОЗВЕДЕНІ КУЛЬТУР ДЛЯ ПОКРАЩЕННЯ РЕЗИСТЕНТНОСТІ У КУЛЬТУР Шляхом хімічного мутагенезу (EMS), були одержані дві нуклеотидні мутації у гені DST (A мутувала до G при нуклеотиді 205, та G мутувала до A при нуклеотиді 484), що спричинили два амінокислотні заміщення (аспарагін мутував на аспарагінову кислоту у положенні 69, та аланін мутував на треонін у положенні 162), що призводить до утворення фенотипу із резистентністю до засухи та дії солі. Розробили наступні дві пари праймерів, SNP5 та SNP3, у вказаному гені, таким чином, щоб ампліфіковані продукти містили першу точкову мутацію та другу точкову мутацію, відповідно. SNP-5S: ATGGACTCCCCGTCGCCT (SEQ ID NO: 10) SNP-5A: GTGCGCCGGGAGAAGCCC (SEQ ID NO: 11) SNP-3S: GCGGTGCCGACGTCGTTCCC (SEQ ID NO: 12) SNP-3A: GCCGCCGTCGTCGTCGTCTTC (SEQ ID NO: 13) Перша точкова мутація генерую ScrFI маскований сайт рестриктази, тоді як друга точкова мутація руйнує BstUI маскований сайт рестриктази. Дигестували ампліфіковані продукти, одержані за допомогою праймерів SNP5, а ScrFI утворював фрагменти 311 bp, 85 bp, та 31 bp у дикому типі, тоді як мутантний ампліфікований продукт утворював фрагменти 202 bp, 109 bp, 85 bp, та 31 bp, таким чином, одержуючи поліморфізм. Дигестували ампліфіковані продукти, одержані за допомогою праймерів SNP3 та BstUI утворював фрагменти 66 bp, 40 bp, та 25 bp у 14 UA 106489 C2 5 10 15 дикому типі, тоді як мутантний ампліфікований продукт утворював фрагменти 91 bp та 40 bp, таким чином також одержуючи поліморфізм. Тому, ці дві пари праймерів SNP5 та SNP3 можуть бути застосовані як молекулярні маркери для застосування у селективному розведенні за допомогою молекулярних маркерів. Застосування мутантів dst з резистентністю до засухи та дії солі для схрещування з варіантами рису, та скринінг потомства на один молекулярний маркер або два молекулярні маркера для відбору тих, що мають DST мутантний ген. Потім, культивували нові варіанти (лінії), що мали підвищену резистентність до засухи та дії солі. Вказаний спосіб застосовує традиційні способи кросбридингу, без генного переносу, таким чином уникаючи проблем з безпекою, пов'язаних із генним переносом. Тому, такі способи є переважними. Всі документи, процитовані у даному винаході, застосовують як посилання у даному описі, таким чином, ніби кожний з документів процитовано окремо. Додатково, має бути зрозуміло, що фахівець у цій галузі, який прочитав опис даного винаходу, описаного вище, зможе змінити або модифікувати різні аспекти відповідно до даного винаходу. Ці еквіваленти підпадають під обсяг формули винаходу, що додається до даної заявки. ФОРМУЛА ВИНАХОДУ 20 25 30 35 40 45 50 55 60 1. Спосіб покращення толерантності до засухи та дії солі у рослини, який включає: інгібування функції активації транскрипції фактора транскрипції, де фактор транскрипції включає домен "цинкові пальці" типу С2H2, що містить послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2; причому інгібування здійснюють шляхом введення полінуклеотиду, що кодує неконсервативний мутантний фактор транскрипції, який являє собою білок, що має послідовність, яка включає Asn69Asp (N69D) мутацію та Ala162Thr (A162T) мутацію у послідовності SEQ ID NO: 2; або інгібування здійснюють шляхом інгібування експресії фактора транскрипції введенням RNAi або антисмислового олігонуклеотиду у рослину. 2. Спосіб за п. 1, в якому фактор транскрипції вибраний з: поліпептиду, що містить амінокислотну послідовність SEQ ID NO: 2; поліпептиду, одержаного з (a), в якому 1-50 амінокислотних залишків заміщено, видалено або інсертовано, та який здатний до підвищення чутливості до засухи та дії солі у рослини. 3. Спосіб за п. 1, в якому фактор транскрипції має амінокислотну послідовність SEQ ID NO: 2. 4. Спосіб за п. 1, в якому інгібування здійснюють шляхом введення полінуклеотиду, що кодує неконсервативний мутантний фактор транскрипції, який не здатний до підвищення чутливості до засухи та дії солі у рослини. 5. Спосіб покращення за п. 1 або 4, в якому введення полінуклеотиду включає: (1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить полінуклеотид; (2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих полінуклеотидом; та (3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2). 6. Спосіб за п. 1, в якому RNAi або антисмисловий олігонуклеотид направлений проти послідовності SEQ ID NO: 1. 7. Спосіб за п. 1 або 6, в якому інгібування експресії фактора транскрипції здійснюють: (1) трансформуванням рослинної клітини, рослинної тканини або органу рослини конструктом, що містить RNAi або антисмисловий олігонуклеотид; (2) відбором рослинної клітини, рослинної тканини або органу рослини, трансформованих олігонуклеотидом; та (3) регенерацією рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2). 8. Спосіб за п. 1, в якому інгібування здійснюють шляхом інгібування зв′язування фактора транскрипції з цис-діючим елементом. 9. Спосіб за п. 8, в якому цис-діючий елемент містить послідовність SEQ ID NO: 3. 10. Рослина, одержана способом відповідно до будь-якого з пп. 1-4, 6, 8-9, що передбачає: (1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить полінуклеотид; (2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих полінуклеотидом; та 15 UA 106489 C2 5 10 15 20 (3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2); або (1) трансформування рослинної клітини, рослинної тканини або органу рослини конструктом, що містить RNAi або антисмисловий олігонуклеотид; (2) відбір рослинної клітини, рослинної тканини або органу рослини, трансформованих олігонуклеотидом; та (3) регенерацію рослини з рослинної клітини, рослинної тканини або органу рослини зі стадії (2). 11. Рослина за п. 10, яка відрізняється тим, що рослина є культурою, вибраною з Gramineae, Malvaceae gossypium, Cruciferae brassica, Compositae, Solanaceae, Labiatae або Umbelliferae. 12. Рослина за п. 11, яка відрізняється тим, що рослина є культурою, вибраною з рису, кукурудзи, пшениці, ячменю, цукрової тростини, сорго, арабідопсису, бавовни або каноли. 13. Спосіб відбору рослини, толерантної до засухи та дії солі, що включає: (i) визначення у кандидатної рослини рівня фактора транскрипції, що містить домен ″цинкові пальці″ типу С2H2, що містить послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2; рівня експресії полінуклеотиду, що кодує послідовність амінокислот 42-85 SEQ ID NO: 2 або консервативний варіант послідовності амінокислот 42-85 SEQ ID NO: 2; та/або рівня зв′язування між цис-діючим елементом, що містить послідовність SEQ ID NO: 3, та фактором транскрипції; та (ii) порівняння рівня у кандидатної рослини, визначеного на стадії (i) з відповідним рівнем у контрольної рослини, якщо рівень у кандидатної рослини, нижчий за рівень у контрольної рослини, тоді кандидатна рослина є рослиною толерантною до засухи та дії солі. 16 UA 106489 C2 17 UA 106489 C2 18 UA 106489 C2 Комп’ютерна верстка С. Чулій Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 19
ДивитисяДодаткова інформація
Назва патенту англійськоюRice zinc finger protein transcription factor dst and use thereof for regulating drought and salt tolerance
Автори англійськоюLin, Hongxuan, Huang, Xinyuan, Chao, Daiyin, Gao, Jiping, Zhu, Meizhen, Shi, Min
Автори російськоюЛин Хонгксуан, Хуанг Ксиньюан, Чао Даиин, Гао Цзипин, Чжу Мейчжен, Ши Мин
МПК / Мітки
МПК: C12N 15/29, A01H 1/00, C12N 15/63
Мітки: солі, засухи, покращення, дії, спосіб, толерантності, рослини
Код посилання
<a href="https://ua.patents.su/21-106489-sposib-pokrashhennya-tolerantnosti-roslini-do-zasukhi-ta-di-soli.html" target="_blank" rel="follow" title="База патентів України">Спосіб покращення толерантності рослини до засухи та дії солі</a>
Спосіб покращення врожайності і/або покращення сили рослини і/або збільшення стійкості або опірності рослини до кліматичних факторів стресу за допомогою амідної сполуки та її застосування

Номер патенту: 105897
Опубліковано: 10.07.2014
Автори: Праде Алешандер Гуттенкунст, Таварес-Родрігес Марко-Антоніо, Фьосте Дірк, Мерк Міхаель, Вільгельм Рональд, Гладуін Роберт Джон, Брам Лутц
МПК: A01P 21/00, A01N 43/56
Мітки: сполуки, факторів, застосування, збільшення, покращення, кліматичних, рослини, стійкості, врожайності, сили, спосіб, амідної, допомогою, стресу, опірності
Формула / Реферат:
1. Спосіб покращення врожайності і/або покращення сили, і/або покращення стійкості або опірності до кліматичних факторів стресу принаймні одного сорту рослини, який включає обробку рослини і/або місця, в якому рослина росте або повинна рости, амідом формули І, Iв якій замісники мають наступні значення:R4 являє собою дифторметил або...
Спосіб поліпшення резистентності або толерантності до нематод у рослини та її потомства, молекула днк, злитий протеїн та рослина, що його експресує

Номер патенту: 71901
Опубліковано: 17.01.2005
Автори: МакФерсон Майкл Джон, Ервін Пітер Едвард, Аткінсон Говард Джон
МПК: C12N 15/82, A01H 5/00, C07K 14/81, C12N 5/10, C12N 15/09, G01N 33/53, G01N 33/15, C07K 19/00, G01N 33/50, C12N 9/02, C12N 9/00
Мітки: рослина, рослини, резистентності, поліпшення, толерантності, протеїн, молекула, нематод, експресує, злитий, потомства, днк, спосіб
Формула / Реферат:
1. Спосіб поліпшення резистентності або толерантності до нематод у рослини та її потомства, який включає включення у геном вищезгаданої рослини гена, що кодує злитий протеїн, який включає: перший протеїн або домен протеїну з антипатогенною активністю, пептид-лінкер та другий протеїн або домен протеїну з антипатогенною активністю, у котрого щонайменше один з протеїнів або доменів протеїнів з антипатогенною активністю...
Композиція для покращення життєздатності рослини

Номер патенту: 102221
Опубліковано: 25.06.2013
Автори: Бегліоміні Едсон, Таварес-Родрігес Марко-Антоніо, Фьосте Дірк, Хаден Егон
МПК: A01N 47/02, A01P 7/04, A01N 43/60, A01N 43/24, A01P 3/00, A01N 51/00, A01N 43/56, A01N 43/653
Мітки: покращення, композиція, рослини, життєздатності
Формула / Реферат:
1. Спосіб обробки рослини комбінацією активних речовин, який включає нанесення на матеріали для розмноження рослини, з яких рослина росте, принаймні одного інсектицидного компонента (В) і нанесення на рослини один або кілька разів, коли вона знаходиться на стадії росту ВВСН 09-49 або під час її цвітіння або розквіту, принаймні однієї сполуки І компонента (А): (А) сполука формули І
Спосіб визначення критичного водного дефіциту у деревних рослин унаслідок повітряної засухи

Номер патенту: 16328
Опубліковано: 29.08.1997
Автори: Разнополов Олег Миколайович, Надеждіна Надія Євстафійовна
МПК: A01G 7/00
Мітки: повітряної, спосіб, деревних, водного, засухи, критичного, визначення, рослин, унаслідок, дефіциту
Формула / Реферат:
Способ определения критического водного дефицита в древесных растениях вследствие воздушной засухи, включающий измерение относительной скорости потока пасоки, отличающийся тем, что, с целью повышения точности и чувствительности определения потребности древесных пород в проведении освежительных или освежительно-увлажнительных поливов, относительную скорость потока определяют непрерывно или дискретно в течение нескольких часов до восхода...
Пестицидна комбінація, спосіб контролю або попередження пошкодження, спосіб захисту матеріалу для розмноження рослини, спосіб обробки рослини та матеріал для розмноження рослини
Номер патенту: 96316
Опубліковано: 25.10.2011
Автори: Брандл Франц, Вайсс Мартін
МПК: A01N 43/56, A01N 43/16, A01P 7/04, A01P 3/00, A01C 1/08, A01N 43/54, A01N 43/78, A01N 37/50
Мітки: розмноження, захисту, спосіб, пошкодження, обробки, рослини, пестицидна, матеріалу, матеріал, комбінація, попередження, контролю
Формула / Реферат:
1. Пестицидна комбінація, яка містить принаймні два компоненти, що являють собою діючі речовини, де компонент (І) являє собою принаймні одну сполуку або її сіль, або складний ефір, вибрану з групи, що включає флудіоксоніл, азоксистробін, дифеноконазол, флуоксастробін, тебуконазол, тіабендазол, трифлоксистробін, opтo-циклопропілкарбоксанілід формули II-IIV,, ІІ
Попередній патент: Пристрій для шліфування безперервно-литого виробу
Наступний патент: Застосування пропінебу як репеленту для птахів
Випадковий патент: Склад для обмеження припливу пластової води у свердловину