Спосіб визначення дат початку вегетації у деревних рослин у разі перенесення їх з одних фізико-географічних районів в інші




Формула / Реферат
Спосіб визначення дат початку вегетації у деревних рослин у разі перенесення їх з одних фізико-географічних районів в інші, який включає визначення сум ефективних температур понад біологічний нуль, який відрізняється тим, що додатково визначають тривалість дня, будують для конкретного виду, популяції, сорту рослин графік залежності між сумами ефективних температур понад біологічний нуль і тривалістю дня на початок вегетації у вихідному районі, будують графік середньорічного ходу наростання сум ефективних температур понад біологічний нуль у заданому районі, за точкою перетину ліній графіків знаходять тривалість дня, а звідси і дату початку вегетації виду, популяції, сорту рослин у заданому районі.
Текст
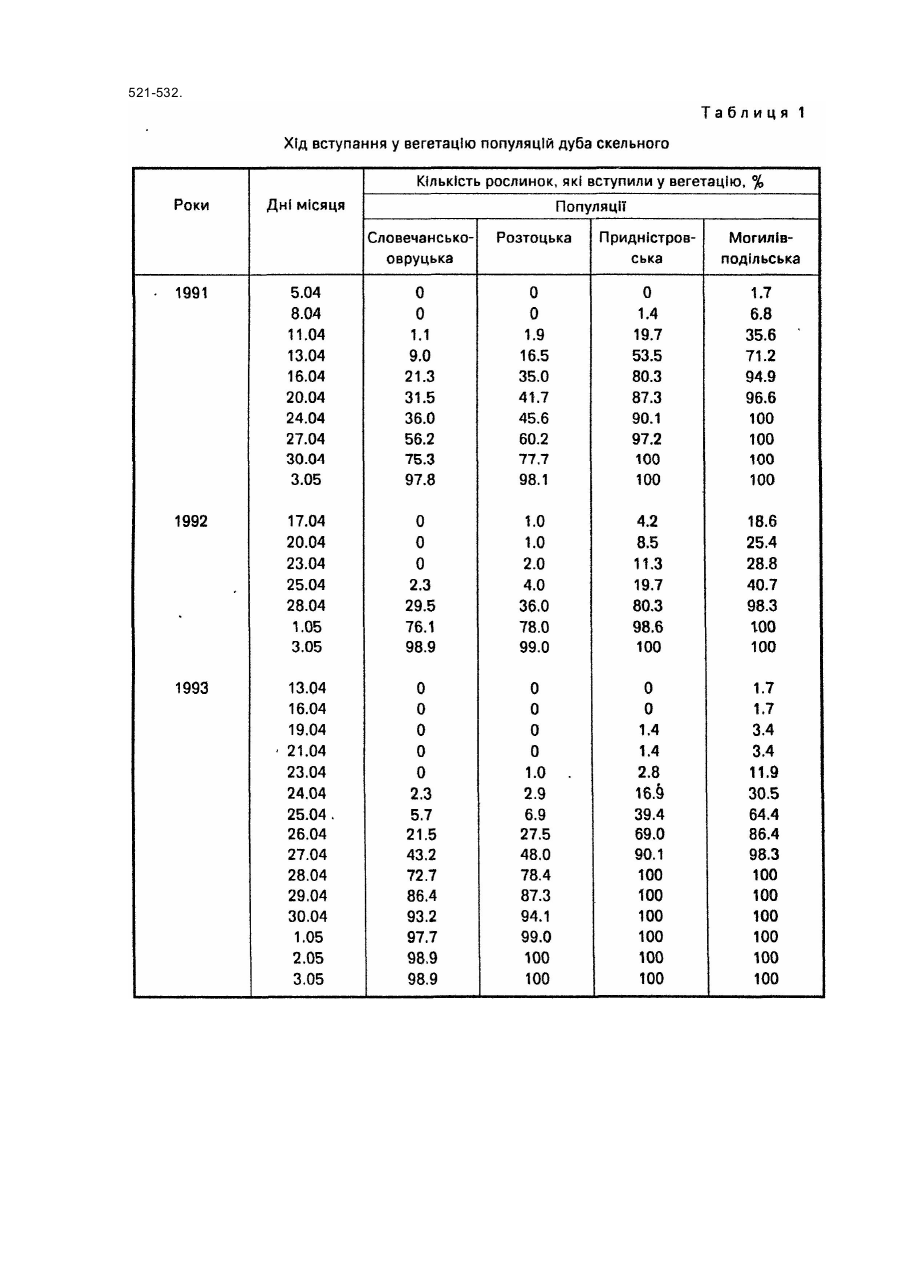
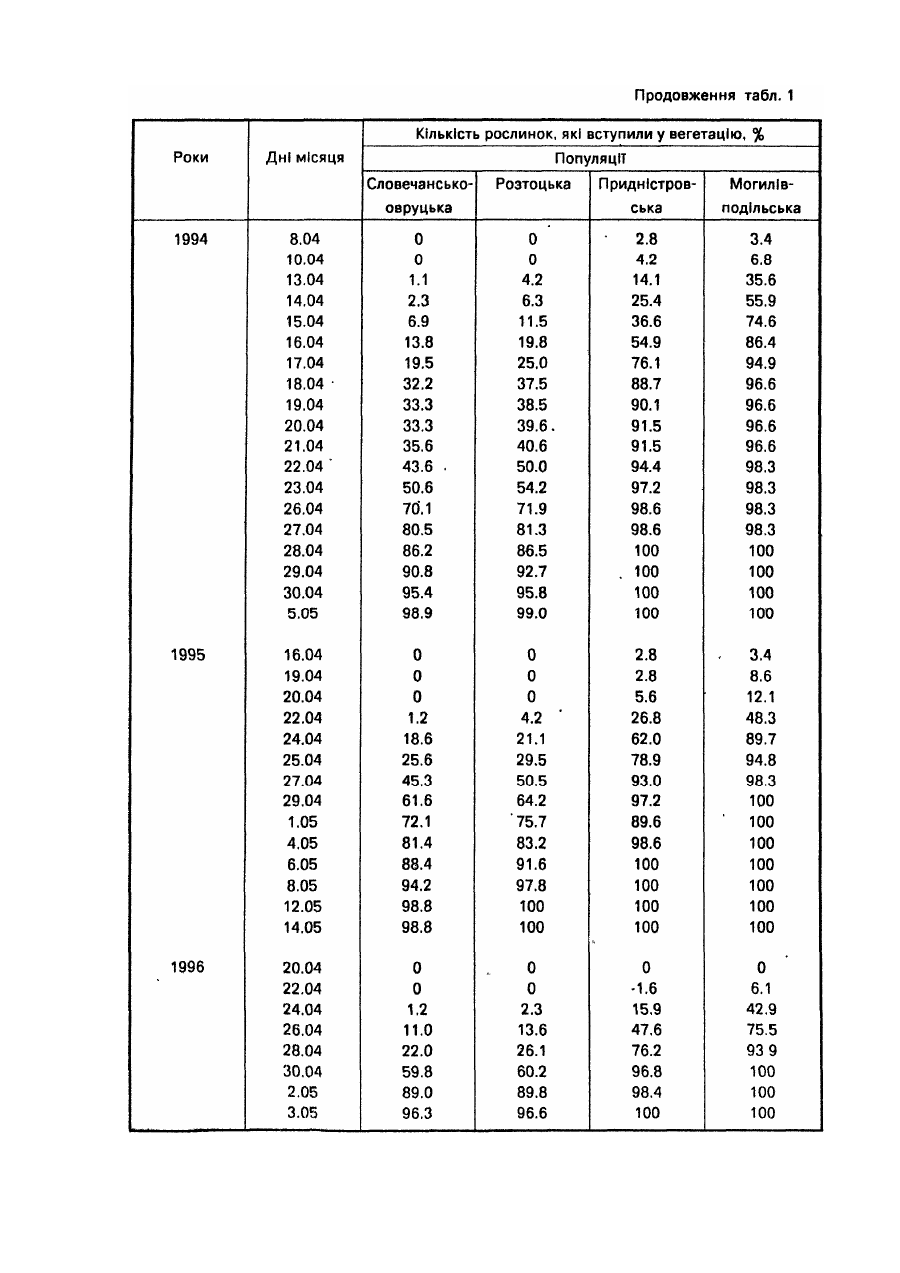
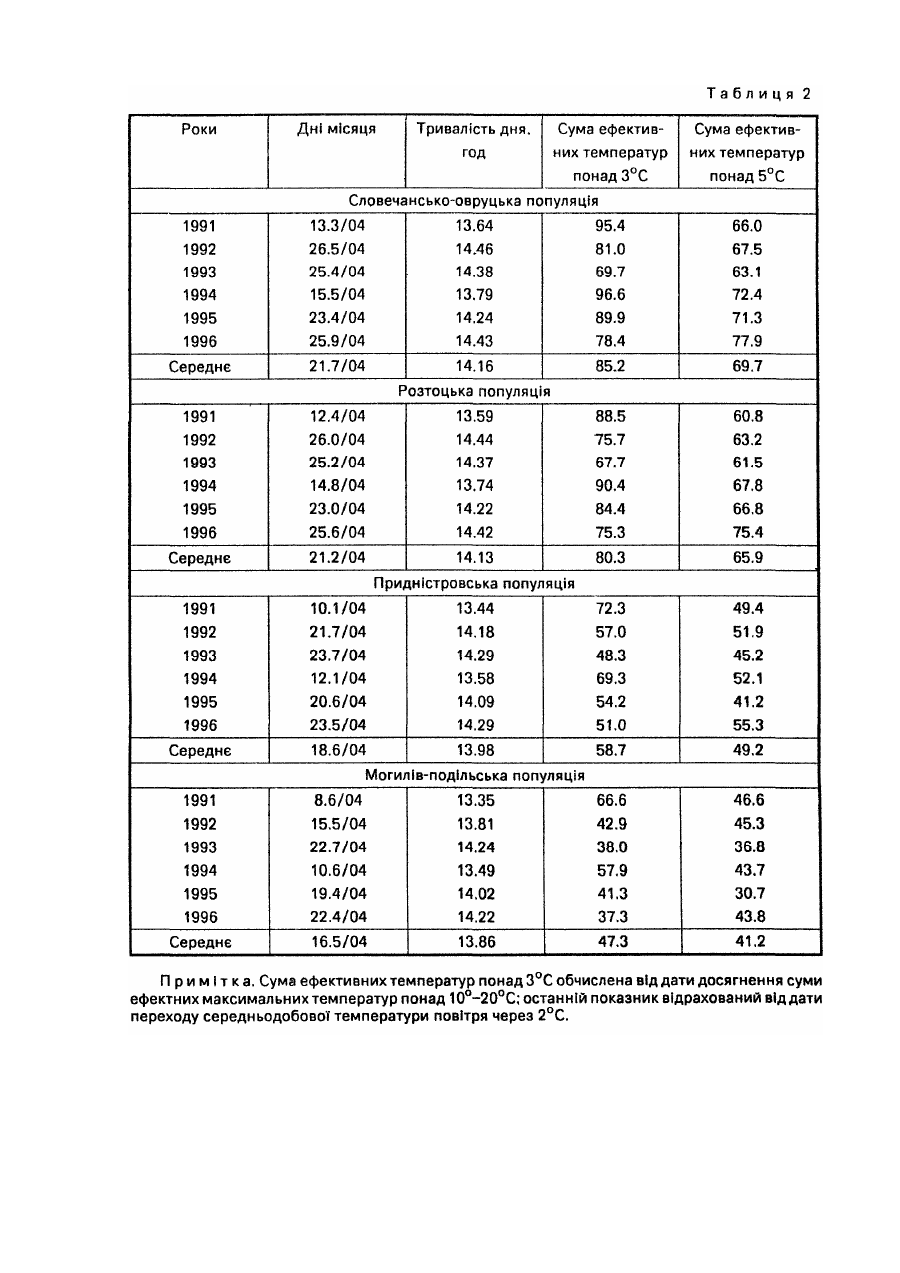
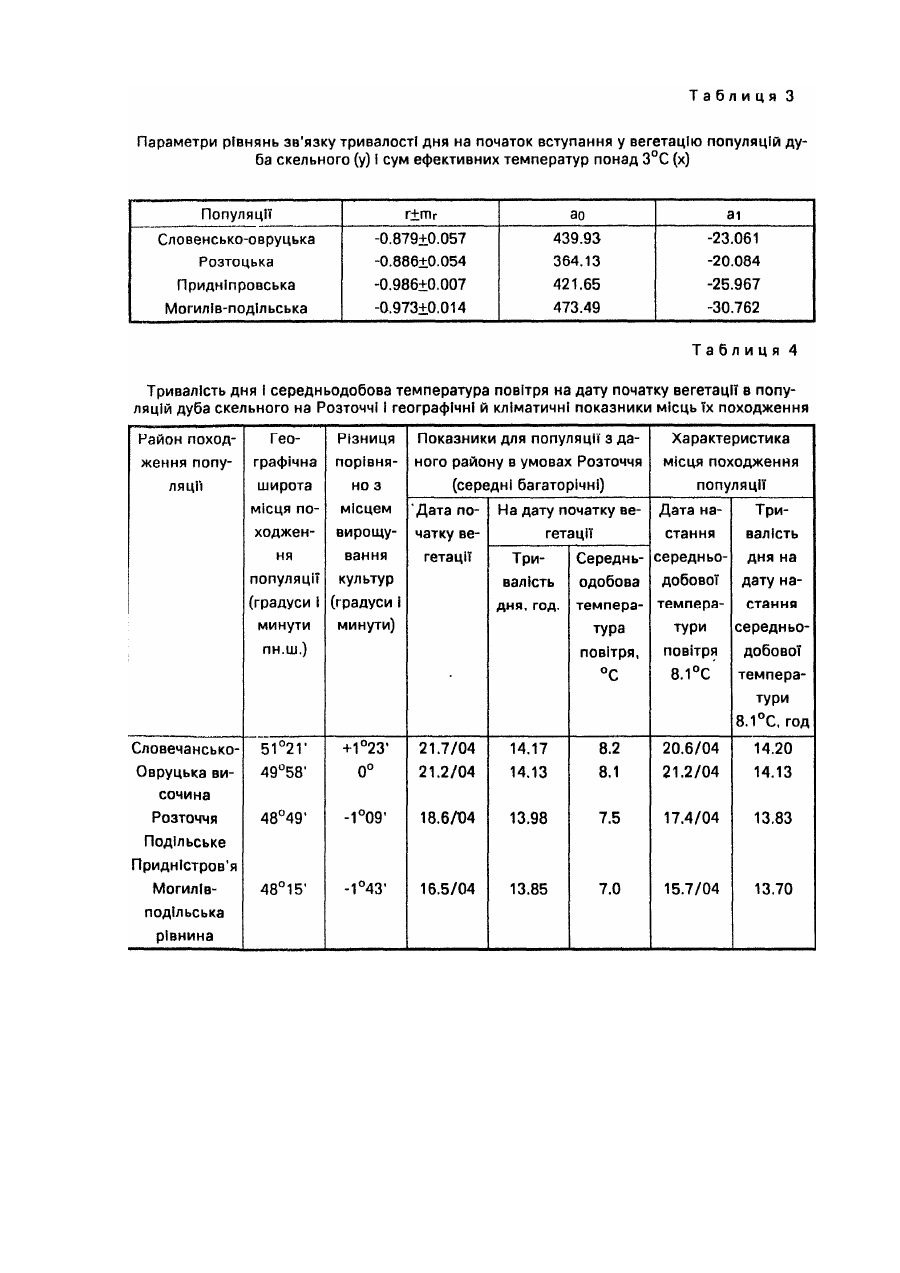
Спосіб належить до галузі лісового господарства і садівництва, може бути використаний при відборі найбільш відповідних за сезонним розвитком видів, популяцій, кліматипів, сортів тощо деревних рослин для вирощування у заданих районах. У разі Інтродукції ріст рослин, репродуктивна здатність, а загалом і життєвість значною мірою визначаються адекватністю ритму їх сезонного розвитку сезонній динаміці кліматичних умов районів, у які вони вводяться, Від даного показника залежить як їх стійкість до несприятливих кліматичних факторів, так і повнота реалізації кліматичних ресурсів. Експериментально встановлено, що однією з ознак перспективності деревних порід-інтродуцентів для середньої смуги помірного поясу є ранній у порівнянні з аборигенними деревними породами початок вегетації [5; 6; 9]. Відомий метод визначення дат настання вегетації у рослин-інтродуцентів, який грунтується на безпосередніх фенологічних спостереженнях за цими рослинами, висадженими у заданому районі [4 та ін.]. Метод вимагає попереднього висаджування рослин-інтродуцентів у заданому районі і проведення за ними кількарічних спостережень. Відомий метод фенологічного прогнозування - метод фенологічного лагу [15, с 168-174], названий ще фенолого-статистичним [7], за яким дати настання фенологічних явищ, що прогнозуються, визначають за датами появи більш ранніх сезонних явищ-індикаторів, корелятивно зв'язаними з датами настання явищ, які прогнозуються. Метод дає надійний прогноз (результат) лише у тому випадку, коли рослина-індикатор і досліджувана рослина належить до аналогічних внутрівидових рас за строками настання відповідних фенофаз і зростають у подібних мікрокеологічних умовах. Цей метод не може бути використаний для визначення дат настання певних фенологічних фаз у рослин у випадку перенесення їх з одних фізико-географічних районів в Інші, бо необхідною для нього умовою у даному разі була б наявність одних і тих же явищ-індикаторів або рослин-індикаторів, як у вихідному, так і в Інтродукційному районі, що далеко не завжди є можливим. До того ж фенологічні Індикатори у різних частинах їх ареалів мають неоднакове Індикаційне значення, а лаг-періоди між одними і тими ж явищами у різних географічних пунктах можуть суттєво відрізнятися один від одного. Усе це істотно ускладнює і знижує надійність визначення середніх багаторічних дат початку вегетації, як і інших фенологічних фаз, у деревних рослин поза районом їх зростання методом фенологічного лагу. Відомі метеоролого-фенологічні методи [15, с 174]. Вони полягають у математичному описі зв'язків між явищами, що прогнозуються і метеорологічними (кліматичними) факторами. Останніми найчастіше виступають суми певних температур, суми опадів за певний період, рівень максимальних чи мінімальних температур тощо. У даній групі методів прогнозування найбільшою мірою розроблені термофенологічні, за якими настання прогнозованого явища визначають на основі показників тільки термічного режиму. Серед зазначених вище відомий метод фенотермограм А.С. Подольського[11]. Уданому випадку прогноз дат настання певних фенофаз складається за термічними номограмами, побудованими на основі середнього багаторічного ходу температури повітря. Основним недоліком методу фенотермограм є неврахування дії Інших факторів, крім температурного, зокрема світлоіого. У зв'язку з цим, а також через те, що "номограми будуються для кожного району окремо за місцевими кліматичними показниками для використання у випадку перенесення рослин з одних фізико-географічних районів у Інші метод не придатний. Відомий метод прогнозу строків цвітіння плодових культур в Середній Азії [12, стор. 111-112]. Метод базується на зв'язку середньої швидкості розвитку дерев у зимово-весняний період Із середньою температурою повітря за цей період, що описується формулою де - середня швидкість розвитку; t -середня температура повітря за період з моменту першого переходу температури через 5°С; В - нижня межа ефективних температур, °С (для різних порід неоднакова); А - сума ефективних температур, необхідних для зацвітання тієї чи Іншої породи, °С; n -тривалість періоду від дати переходу середньодобової температури через певний рівень до дати зацвітання, дні. За даним методом прийнято різні суми ефективних температур для весен після теплих і після холодних зим. Метод має регіональне призначення, вплив фотоперіоду на розвиток росин у ньому не враховується. Відомий метод прогнозу цвітіння яблуні в Узбецькій РСР [12, С 112-116]. Метод базується на використанні сум ефективних температур понад 5°С, необхідних для початку цвітіння яблуні і сум мінімальних температур від 1 січня до дати переходу середньодобової температури через 5°С. "Метод має регіональне призначення, розраховані за ним суми ефективних температур, необхідні для початку цвітіння яблуні, у різних районах Узбекистану різко відрізняються одна від одної. Вплив фотоперіоду на розвиток рослин уданому методі не враховується. Відомий метод прогнозу цвітіння яблуні в Туркменській РСР [12, с 116-117]. Метод базується на використанні сум ефективних температур понад 0°С. Розрахунки проводяться за тією ж формулою, що і в описаному вище методі прогнозу строків цвітіння плодових культур в Середній Азії. Метод має регіональне призначення. Розраховані для різних районів Туркменистану суми ефективних температур, необхідні для зацвітання яблуні, Істотно різняться між собою. Вплив фотоперіоду на розвиток рослин уданому методі не враховується. Відомий метод прогнозу строків цвітіння плодових культур в Грузинській РСР [12. с 117-118]. Для груші і яблуні в основу методу покладено зв'язок строків цвітіння цих культур із середньомісячною температурою березня, а для персика і абрикоса зв'язок строків цвітіння з датами переходу середньої добової температури повітря через 5°С. Метод має регіональне призначення. Вплив фотоперіоду на розвиток рослин у ньому не враховано Відомий метод прогнозу розвивання перших листків шовковиці в Грузинській РСР [12, с 121] В основу методу покладено зв'язок дати розвивання перших листків шовковиці з кількістю днів від 1 жовтня до переходу середньої добової температури повітря через 10°С. Метод має регіональне призначення; вплив фотоперіоду на розвиток рослин у ньому не враховано Відомий метод прогнозу строків появи перших і п'ятих листків у шовковиці в Узбецькій РСР [12, 120-121] Метод базується на використанні сум ефективних температур понад 5°С Він має регіональне призначення, суми ефективних температур, необхідні для появи листків у шовковиці, у різних районах Узбекистану Істотно різняться між собою. Вплив фотоперіоду на розвиток рослин у методі не враховано. Відомий метод прогнозу строків появи перших і п'ятих листків у шовковиці в Туркменській РСР [12, с 118119]. Метод базується на використанні зв'язку середньої швидкості розвитку дерев у зимово-весняний період Із середньою добовою температуою повітря за цей період. Цей зв'язок описується тією ж формулою, яка використана в описаному вище методі прогнозу цвітіння плодових культур в Середній Азії [12, с 111-112]: де середня швидкість розвитку; t - середня температура повітря за період з моменту першого переходу температури через 5°С; В - нижня межа ефективних температур, °С; А - сума ефективних температур, °С; η - тривалість періоду від дати переходу середньодобової температури через певний рівень до дати появи листків, дні. Метод має регіональне призначення може використовуватися в південних районах Туркменистану. Прототипом запропонованого способу може бути метод прогнозу строків цвітіння плодових культур в Європейській частині СССР [12, с. 108-111] (метод сум температур). Метод базується на використанні сум ефективних температур понад 5°С. Дата зацвітання плодових культур визначається за формулою де n = тривалість періоду від переходу середньої добової температури повітря через 5°С до зацвітання, дні; St > 5 – сума ефективних температур понад 5°С, при якій починається цвітіння (константа); Τ - середня температура повітря за період прогнозування п, °С Використання даного методу обмежується певним регіоном Методі недостатньо точним для випадків, пов'язаних з перенесенням рослин з одних фізико-географічних районів в інші, оскільки вплив фотоперіоду на розвиток рослин у ньому не враховується В основу даного винаходу покладено завдання розробити такий спосіб визначення дат настання вегетації у таксонів деревних рослин (видів, популяцій, сортів тощо) у разі перенесення їх з одних фізико-географічних районів у Інші, в якому поряд з показником температурного режиму, зокрема сумою ефективних температур понад біологічний нуль у заданому районі враховувався б і фотоперіод, що дало б можливість скоротити термін виконання робіт і підвищити їх точність порівняно з Існуючими способами. Визначення дати початку вегетації у таксонів рослин проводять на основі багаторічних (не менше як 4річних) паралельних спостережень за їх фенологічним станом і метеорологічними показниками у вихідній місцевості (за останні можуть бути використані дані найближчої метеостанції, розташованої в аналогічних топокліматичних умовах). Спостереження проводять за групою екземплярів окремого таксона (чим більше екземплярів рослин охоплено спостереженнями, тим вища їх точність; бажано, щоб екземплярів було не менше як 50 шт.). Періодично фіксують кількість екземплярів таксона, що вступила у вегетацію. За кумулятою фенологічного процесу за кожний рік графічно (шляхом інтерполяції) або розрахунковим методом, наведеним у роботі [2], знаходять дату початку вегетації (у вегетацію вступило 5 або 10% екземплярів таксона). Визначають тривалість дня на дату початку вегетації (за "Астрономическим ежегодником ... на ... год" [1]). Визначають суму ефективних температур понад біологічний нуль для даного таксону на дату початку вегетації. За паралельними багаторічними даними складають рівняння і будують графік зв'язку тривалості дня і сум ефективних температур на початок вегетації в досліджуваного таксона. Треба зазначити, що тривалість дня і сума ефективних температур понад біологічний нуль щодо впливу на проходження фенологічних процесів у рослин є факторами, які компенсують один одного. Тому, чим вищим є від’ємний кореляційний зв'язок між ними, тим точніше знайдено точки відліку суми ефективних температур: дату початку відліку і біологічний нуль. Пошук останніх показників спрямовують таким чином, щоб отримати якнайвищий від'ємний зв'язок між тривалістю дня і сумою ефективних температур. (Див. рис. 1, де точками (1-6) позначено тривалість дня і суму ефективних температур на дату початку вегетації досліджуваного таксона в окремі роки у вихідному районі; і лінія зв'язку тривалості дня і суми ефективних температур на дату початку вегетації таксона у вихідному районі, описується рівнянням виду: у = ао + а1х, де у - сума ефективних температур понад біологічний нуль, °С; x - тривалість дня, год; II - лінія середньорічного ходу наростання суми ефективних температур понад біологічний нуль в залежності від тривалості дня, тобто в залежності і від дат, у заданому районі). За графіком зв'язку тривалості дня і сум ефективних температур на початок вегетації таксона у вихідному районі і ходом наростання сум ефективних температур у заданому районі знаходять тривалість дня (за точкою перетину ліній I і II на рис.1), а отже і середньорічну дату початку вегетації в таксона, якби він був перенесений у заданий район. Можливість реалізації способу підтверджується даними фенологічних спостережень за популяційними культурами дуба скельного на Розточчі (Лелехівське лісництво навчально-виробничого лісокомбінату УкрДЛТУ). Культури закладені 1990 року семирічними саджанцями, У них представлені чотири популяції дуба скельного з різних фізико-географічних районів України: розтоцька (місцева), словечансько-овруцька (Із Словечансько-Овруцької височини), придністровська (з Подільського Придністров'я), могилів-подільська (з Могилів-подільської рівнини). Фенологічні спостереження проводилиси у 1991-1996 pp. Ділянка, де закладено популяційні культури, має підвищене, але майже рівне місцеположення, її висота над рівнем моря 33S м. Вона являє собою зруб площею близько 6 га, горизонт з півдня і заходу повністю відкритий, зі сходу трохи закритий стіною молодого листяного деревостану, а з півночі - стіною стиглого мішаного деревостану. При складанні рівнянь зв'язку тривалості дня і сум ефективних температур на дату початку вегетації в популяцій дуба скельного було використано кліматичні показники метеорологічної станції, розташованої за 20 км від дослідної ділянки в аеропорту м. Львова. Метеоплощадка цієї станції має рівне, майже повністю відкрите місцеположення і абсолютну висоту 319 м. Тому її кліматичні показники, безсумнівно, тісно корелюють з показниками дослідної ділянки. За ознаку вступання деревець дуба у вегетацію було прийнято наявність у них однієї або більше розпукнутих бруньок. У кожній з популяцій спостереженнями було охоплено від 58 до 102 деревець. Дані фенологічних спостережень за вступанням популяцій дуба скельного у вегетацію наведено у табл.1. З даними табл.1 розрахунковим способом, викладеним у роботі [2], для популяцій дуба скельного знайдено щорічні дати початку вегетації (вступило у фенофазу 10% екземплярів). На ці дати для району Розточчя визначено тривалість дня 1 суми ефективних температур понад 3°С, а також понад 5°С. Підрахунок сум ефективних температур понад 5°С проводили прийнятим у фенології способом [13, с 14-15]. Враховуючи усі середньодобові температури повітря, які перевищували 5°С, у т.ч. і за період до дати переходу середньодобових температур повітря через 5°С. Відлік сум ефективних температур понад 3°С розпочинали від тієї дати, на яку сума ефективних максимальних температур повітря понад 10°С, обчислена від дати переходу середньодобової температури повітря через 2°С, досягала 20°С. На нашу думку, такий підхід є правильний, бо дуже ймовірно, що саме певний рівень максимальних температур виводить рослини Із стану спокою, а далі їх розвиток проходить у тісному зв'язку із середньодобовими температурами. Сума ефективних максимальних температур у 20°С знайдена емпіричним шляхом: при меншій або при більшій сумі цих температур обернений зв'язок між тривалістю дня і сумою ефективних температур понад 3°С послаблювався. Показники початку вступання у вегетацію популяцій дуба скельного наведені в табл.2. За цими даними побудовано рівняння зв'язку тривалості дня і суми ефективних температур понад 3°С на дату вступання популяцій у вегетацію. Графічно цей зв'язок для чотирьох популяцій дуба скельного показано на рис. 2: на а) для словечансько-овруцької популяції, б) для розтоцької, в) придністровської, г) могилів-подільської, де цифрами позначено: 1 - дані 1991 року, 2 -1992 року, 3-1993 року, 4-1994 року, 5 -1995 року, 6 - 1996 року. Для усіх популяцій зазначений зв'язок є сильним, коефіцієнти кореляції достовірні на 95%-ному рівні значущості (t ф - 3,69 - 11,83; t0.05 = 2,78)· Щодо суми ефективних температур понад 5°С, обчислених відомим методом [13, с. 14-15], і тривалості дня, то кореляційний зв'язок між ними загалом неоднозначний - коефіцієнт кореляції коливається для різних популяцій у межах від +0,293 до -0,548. Параметри рівнянь наведено у табл.3. З даних табл.2 і 3, а також рис. 1 видно, що Із зростанням тривалості дня необхідна для настання вегетації у популяцій дуба скельного сума ефективних температур понад 3°С зменшується. Тобто зазначені фактори у даному випадку за своїм впливом на настання вегетації у рослин взаємно компенсують один одного. Як видно з табл.2, популяції дуба скельного в умовах Розточчя істотно відрізняються одна від одної як за щорічними, так і за середніми багаторічними датами початку вегетації, тривалістю дня і сумами ефективних температур понад 3°С на ці дати. Популяції з південних районів - придністровська і могилів-подільська розпочинають вегетацію у середньому, відповідно, на 2,6 і 4,7 дні раніше і при меншій тривалості дня, ніж місцева розтоцька, а популяція з північного району-словечансько-овруцька - на 0,5 дня пізніше і при більшій тривалості дня. Придністровська і могилів-подільська популяції розпочинають вегетацію при сумах ефективних температур понад 3°С, відповідно, в 1,36 і 1,70 раза менших, ніж місцева, а словечансько-овруцька в 1,06 раза більшій. Але це не означає, що популяції з півдня порівняно з місцевою популяцією є менш теплолюбив!, а популяція з півночі є більш теплолюбивою -найімовірніше, могло б бути навпаки. Наведені ж дані ще раз підтверджують наявність взаємокомпенсуючого впливу на настання вегетації у рослин тривалості дня і температурного фактора - у нашому випадку сум ефективних температур понад 3°С. Як відомо, після весняного рівнодення (23 березня) і до осіннього рівнодення (22 вересня), у географічних пунктах північної півкулі у вищих широтах тривалість дня ε більшою, ніж у нижчих. Тобто, на будь-яку дату зазначеного періоду у північних районах тривалість дня є більшою, ніж у південних. А настання одних і тих же температурних показників у весняну частину зазначеного періоду в північних районах припадає на більш пізні дати, ніж у південних, тобто, обов'язково на більшу тривалість дня. Тому при переміщенні на північ, тобто у райони з більшою тривалістю дня, рослини розпочинають вегетацію при менших сумах ефективних температур І, навпаки, при переміщенні на південь - у райони з меншою тривалістю дня - рослини вступають у вегетацію при більших сумах ефективних температур. Це явище добре простежується у нашому досліді (табл.2). Взаємокомпенсуючу роль тривалості дня і температурного показника (у даному випадку не сум ефективних температур, а середньодобової температури повітря) добре висвітлюють дані, наведені у табл.4. Суми ефективних температур необхідно обчислювати за результатами щоденних метеорологічних спостережень, останні у розпорядженні автора є тільки для району Розточчя. Середньодобову температуру повітря можна знаходити за графіком її середнього багаторічного ходу. Вона, звичайно, тісно корелює Із сумами ефективних температур. Як видно з табл.4, місце походження словечансько-овруцької популяції дуба скельного знаходиться на 1°23' північніше від місця вирощування, а місця походження придністровської і могилів-подільської популяцій відповідно, на 1°09 ' і 1°43' південніше. Для розтоцької популяції географічна широта місця походження і місця вирощування одна і та ж. За даними табл.4, на Розточчі місцева популяція вступає у вегетацію при середньодобовій температурі 8.1°С, популяції з півдня - при дещо вищій. Але це не. може означати, що південні популяції менш теплолюбиві, ніж розтоцька, а популяція з півночі - більш теплолюбива. Вважаємо за можливе припустити, що усі популяції дуба скельного практично однаково вибагливі до тепла. У такому разі у місцях їх походження початок вегетації у них наставав би при тій самій середньодобовій температурі повітря, що й в розтоцької популяцій на Розточчі, тобто, при 81°C, I відповідній на дату досягнення цієї температури тривалості дня. Як видно з табл.4, на дату досягнення середньодобової температури повітря 8.1°С тривалість дня у місці походження могилів-подільської популяції становить 13,70 год. На Розточчі ж ця популяція вступає у вегетацію при нижчій від зазначеної середньодобовій температурі повітря, але при більшій, ніж 13,70 год, тривалості дня. Аналогічне можна сказати і про придністровську популяцію. У місці походження словечансько-овруцької популяції середньодобова температура 8.1 °С настає при тривалості дня 14.20 год. На Розточчі ця популяція вступає у вегетацію при тривалості дня 14.16 год і середньодобовій температурі повітря 8,2°С. Таким чином, і результати багаторічних фенологічних спостережень, проведених на Розточчі, і розрахунки, які стосуються переміщення популяцій дуба скельного у широтному напрямку, свідчать про те, що настання вегетації у рослин великою мірою залежить від тривалості дня і показників температурного режиму. Зазначені фактори взаємно компенсують один одного за своїм впливом на ростові процеси у рослин, і це проявляється не тільки щодо популяцій дуба скельного, а й інших популяцій і видів рослин. У роботі Л.С. Плотникової і В.Н. НІлова [10] наводяться дані, за якими у видів спіреї (спостереження проводили за шістнадцятьма видами) в Архангельську вегетація починається на 6-18 днів пізніше, ніж у Москві (тобто при більшій травалості дня - Р.К.), при меншій (в 1,3-3,5 рази) сумі ефективних температур- Тобто, за своїм впливом на вступання рослин у вегетацію тривалість дня і сума ефективних температур взаємно компенсують одне одного. Pivert Francouse, Jacques Monique, Bonnet-Masimbert Μ., Miginiak Є. [17J виявили, що збільшення фотоперіоду, так само як і температури повітря й величини світлової енергії, прискорює і стимулює інтенсивність розпукування бруньок у дугласії. Про вплив фотоперіоду поряд із сумою ефективних температур на початок росту у видів роду Juniperus і Pinus вказують Северотока 1.1. і Зайцев Г.К. [14]. Про регуляторну роль фотоперіоду і температури щодо вступання у вегетацію сіянців сосни звичайної зазначається у роботі Дормлінга [16]. Про Істотний вплив фотоперіоду на настання фенофаз, у т.ч. і розпукування бруньок, у 7 видів вишні зазначається у роботі Губіної Є.М. [3]. А Б.С. Мошков [8] обгрунтовано стверджує, що регуляторна роль фотоперіодів простежується у рослин в будь-яких фізіологічних процесах і явищах (І репродуктивних, і ростових). Отже, настання вегетації у деревних рослин відбувається під впливом як температурного фактора, так і фотоперіоду (тривалості дня). Врахування останнього дає можливість підвищити точність розрахунків початку вегетації у деревних рослин, особливо у разі переміщення їх з одних фізико-географічних районів в інші. Используемая литература 1. Астрономический ежегодник ... на ... год. - М.-Л.: Изд-во АН СССР, 19... 2. Буторина Т.Н., Крутовская Л.П., Молоков В.А., Полякова В.И. Опыт применения фенологического интегрального метода при геоботанических и почвенных исследованиях // Фенологические методы изучения лесных биогеоценозов. - Красноярск, 1975. - С. 21-53. 3. Губина Е.М. Влияние радиационного и термического режимов на динамику сезонного развития растений на примере рода Cerasus Juss. // Фенологическая индикация и фенопрогнозирование. Тезисы докладов на V Всесоюзном совещании (Алма-Ата, май 1984 года). - Л., 1984. - С. 24. 4. Лапин П.И. Сезонный ритм развития древесных растений и его значение для интродукции // Бюлл. Гл. бот. сада. 1967. Вып. 65. С. 13-18. 5. Лапин П.И., Калуцкий К.К., Калуцкая О.Н. Интродукция лесных пород. - М.: Лесная пром-сть, 1979. - 224 с. 6. Лапин П.И., Сиднева СВ.. Определение перспективности растений для интродукции по данным фенологии // Бюлл. Гл. бот. сада, 1969. - Вып. 69. - С. 14-21. 7. Мауринь A.M., Тардов Б.Н. Биологическое прогнозирование. - Рига, 1975. - 279 с. 8. Мошков Б.С. Актиноритмизм растений. - М.: ВО "Агропромиздат". - 207 с. 9. Плотникова Л.С. Научные основы интродукции и охраны древесных растений флоры СССР. - М.: Наука, 1988. - 264 с. 10. Плотникова Л.С, Нилов В.Н. Сезонный ритм развития таволги в Москве и Архангельске // Бюлл. Гл. бот. сада. 1982. -Вып. 65.-С. 17-27. 11. Подольский А.С. Фенологический прогноз (Математический прогноз в экологии). Изд. 2-е, доп. и перераб. - М.: Колос, 1974. 12. Руководство по агрометеорологическим прогнозам. Т.2. Технические, овощные, плодовые, субтропические культуры, травы, пастбищная растительность, отгонное животноводство / Под ред. Ю.С. Мельника, Н.В. Гулиновой, Г.А. Побетовой, С.А. Бедарева. -Л.: Гидрометеоиздат, 1984. - 264 с. 13. Руководство по составлению агрометеорологических ежегодников. Изд. 3-е, перераб. и доп. Обнинск. 1969. - 59 с. 14. Северотока И.И., Зайцев Г.К. Взаимосвязи некоторых фенофаз и факторов среды у хвойных в Туркменистане // Научно-технический прогресс в пустыне. Тезисы докл. 5 Всесоюзной науч. конф., Ашхабад, 14-16 окт. 1986. - Ашхабад, 1986. - С 128-130. 15. Шульц Г.. Общая фенология. - Л,: Наука. 1981.- 188 с. 16. Dormllng I. The role of photoperiod and temperature in the Induction and the release of dormancy in Pinus sylvestris L seedlings // Ann. sci. forest. 1986. v. 43. P. 228-232. 17." Plvert Francouse, Jacques Monique, Bonnet-Maslmbert M., Migtniak E. Comportement des bourgeons de Pseudotsuga menzlesii (Mirb.) Franko Cultives en conditions controlees // Ann. sci. forest., 1986. - V. 43. - № 4. - P. 521-532.
ДивитисяДодаткова інформація
МПК / Мітки
МПК: A01G 23/00
Мітки: визначення, деревних, фізико-географічних, інші, вегетації, початку, перенесення, разі, районів, рослин, одних, дат, спосіб
Код посилання
<a href="https://ua.patents.su/9-22867-sposib-viznachennya-dat-pochatku-vegetaci-u-derevnikh-roslin-u-razi-perenesennya-kh-z-odnikh-fiziko-geografichnikh-rajjoniv-v-inshi.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення дат початку вегетації у деревних рослин у разі перенесення їх з одних фізико-географічних районів в інші</a>
Спосіб визначення посухостійкості деревних рослин

Номер патенту: 16333
Опубліковано: 29.08.1997
Автори: Кайбияйнен Лев Константинович, Надеждіна Надія Євстафійовна
Мітки: спосіб, визначення, рослин, посухостійкості, деревних
Формула / Реферат:
Способ определения засухоустойчивости древесных растений, включающий измерение биофизического показателя, по значению которого оценивают засухоустойчивость сортов, отличающийся тем, что, с целью повышения точности определения за счет исключения влияния на него условий окружающей среды, в качестве биофизического показателя измеряют значение водного потенциала листьев растений при максимальной относительной скорости потока пасоки, при этом...
Спосіб визначення критичного водного дефіциту у деревних рослин унаслідок повітряної засухи

Номер патенту: 16328
Опубліковано: 29.08.1997
Автори: Разнополов Олег Миколайович, Надеждіна Надія Євстафійовна
МПК: A01G 7/00
Мітки: критичного, водного, рослин, деревних, визначення, унаслідок, дефіциту, спосіб, повітряної, засухи
Формула / Реферат:
Способ определения критического водного дефицита в древесных растениях вследствие воздушной засухи, включающий измерение относительной скорости потока пасоки, отличающийся тем, что, с целью повышения точности и чувствительности определения потребности древесных пород в проведении освежительных или освежительно-увлажнительных поливов, относительную скорость потока определяют непрерывно или дискретно в течение нескольких часов до восхода...
Спосіб прогнозування років масового плодоношення деревних рослин

Номер патенту: 17716
Опубліковано: 20.05.1997
Автори: Мельник Анатолій Степанович, Шляхта Янош Михайлович
МПК: A01G 17/00
Мітки: спосіб, прогнозування, рослин, років, деревних, масового, плодоношення
Формула / Реферат:
Спосіб прогнозування років масового плодоношення деревних рослин, який включає визначення строків масового формування генеративних органів рослин, який відрізняється тим, що з цією метою використовуються пагони рослин починаючи від останнього року і до 4-6 річного віку, у яких проводять поміри річних приростів за діаметром і довжиною І визначають мінімальне співвідношення рослин.
Спосіб щеплення деревних рослин

Номер патенту: 17529
Опубліковано: 06.05.1997
Автор: Пленкіна Владлена Федорівна
МПК: A01H 1/06
Мітки: рослин, спосіб, деревних, щеплення
Формула / Реферат:
Способ прививки древесных растений, включающий выполнение надреза и помещение в него прививаемого материала, отличающийся тем, что в качестве прививаемого материала используют нижнюю часть молодых дичков - ствол с корнем.
Спосіб щеплення деревних рослин живцем

Номер патенту: 16752
Опубліковано: 29.08.1997
Автор: Панков Володимир Вікторович
МПК: A01G 1/06
Мітки: щеплення, рослин, спосіб, живцем, деревних
Формула / Реферат:
Способ прививки древесных растений черенком, включающий выполнение надрезов на подвое, соответствующих диаметру привойного черенка, соединение привоя и подвоя с совмещением камбиальных слоев, отличающийся тем, что, с целью снижения трудозатрат и увеличения процента приживаемости прививочных компонентов при выращивании плодовых культур, надрезы на подвое выполняют в форме гнезда цилиндрической формы с диаметром, соответствующим диаметру...
Попередній патент: Спосіб визначення зусиль в нерухомому кінці талевого канату бурової установки
Наступний патент: Спосіб приготування глиняної пасти та пристрій для його здійснення
Випадковий патент: Амперометричний ферментний біосенсор для визначення вмісту гліцерину у алкогольних напоях