Агоністичне антитіло проти notch3 і його застосування для лікування notcн3-асоційованих захворювань
Номер патенту: 102061
Опубліковано: 10.06.2013
Автори: Лі Кан, Фун Сек Чун, Ву Веньцзюань, Сінгх Санджайа, Чжоу Бінь-бін Стівен







Формула / Реферат
1. Моноклональне антитіло, яке специфічно зв′язується з амінокислотами 1384-1503 амінокислотної послідовності Notch3 (SEQ ID NO:1), де антитіло містить послідовність варіабельного важкого (VH) ланцюга, яка включає послідовності SEQ ID NO:4, SEQ ID NO:5 і SEQ ID NO:6, і послідовність варіабельного легкого (VL) ланцюга, яка включає послідовності SEQ ID NO:7, SEQ ID NO:8 і SEQ ID NO:9, або його функціонально активний фрагмент.
2. Антитіло або його фрагмент за п. 1, де послідовність VH-ланцюга включає послідовність SEQ ID NO:2.
3. Антитіло або його фрагмент за п. 1, де послідовність VL-ланцюга включає послідовність SEQ ID NO:3.
4. Антитіло або його фрагмент за п. 2, де послідовність VL-ланцюга включає послідовність SEQ ID NO:3.
5. Антитіло або його фрагмент за п. 1, де антитіло або його фрагмент специфічно зв′язується з епітопом, який включає послідовність SEQ ID NO:10.
6. Антитіло або його фрагмент за п. 1, де антитіло або його фрагмент специфічно зв′язується з епітопом, який включає послідовність SEQ ID NO:11.
7. Антитіло або його фрагмент за п. 1, де антитіло або його фрагмент має спорідненість зв′язування, що характеризується Kd від 10-8 до 10-10 М.
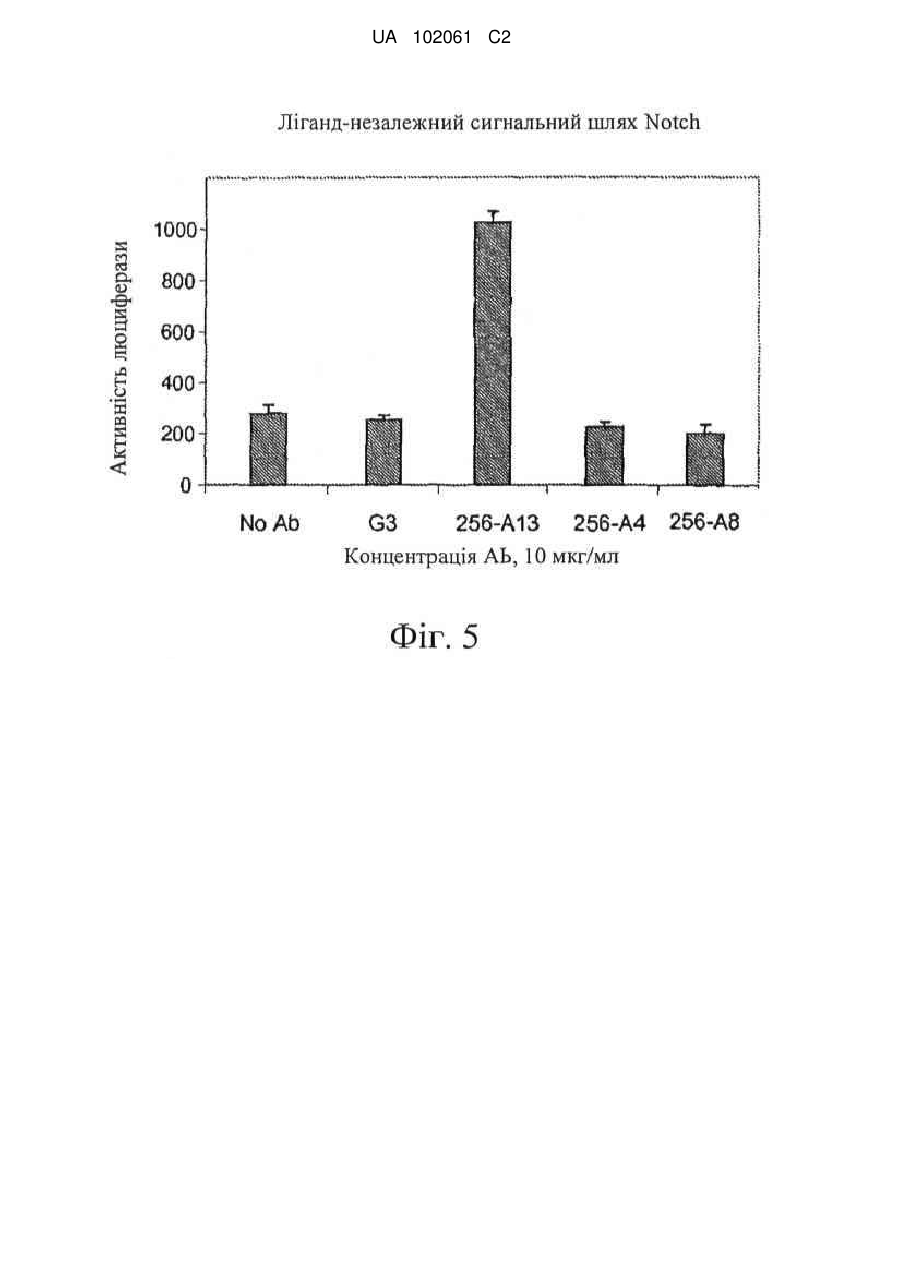
8. Антитіло або його фрагмент за п. 1, де антитіло або його фрагмент активує опосередкований Notch3 сигнальний шлях.
9. Мічене антитіло або його фрагмент, що специфічно зв′язується з амінокислотами 1384-1503 амінокислотної послідовності Notch3 (SEQ ID NO:1), яке включає антитіло або його фрагмент за будь-яким з пп. 1-8 та мітку.
10. Гуманізоване антитіло або його фрагмент, що специфічно зв′язується з амінокислотами 1384-1503 амінокислотної послідовності Notch3 (SEQ ID NO:1), яке являє собою гуманізовану форму антитіла або його фрагмента за п. 4.
11. Гуманізоване антитіло або його фрагмент за п. 10, де антитіло або його фрагмент специфічно зв′язується з епітопом, який включає послідовність SEQ ID NO:10.
12. Гуманізоване антитіло або його фрагмент за п. 10, де антитіло або його фрагмент специфічно зв′язується з епітопом, який включає послідовність SEQ ID NO:11.
13. Гуманізоване антитіло або його фрагмент за п. 10, де гуманізоване антитіло або його фрагмент має спорідненість зв′язування, що характеризується Kd від 10-8 до 10-10 М.
14 Гуманізоване антитіло або його фрагмент за п. 10, де гуманізоване антитіло або його фрагмент активує опосередкований Notch3 сигнальний шлях.
15. Нуклеїнова кислота, яка кодує антитіло або його фрагмент за п. 1.
16. Вектор експресії, який містить нуклеїнову кислоту за п. 15.
17. Клітина, яка містить нуклеїнову кислоту за п. 15 або вектор за п. 16, де клітина здатна продукувати антитіло або його фрагмент за п. 1.
18. Спосіб одержання антитіла або його фрагмента, що включає в себе культивування клітини за п. 17 в умовах, придатних для продукування антитіла і виділення продукованого антитіла або його фрагмента.
19. Нуклеїнова кислота, яка кодує антитіло або його фрагмент за п. 10.
20. Вектор експресії, який містить нуклеїнову кислоту за п. 19.
21. Клітина, яка містить нуклеїнову кислоту за п. 19 або вектор за п. 20, де клітина здатна продукувати антитіло або його фрагмент за п. 10.
22. Спосіб одержання антитіла або його фрагмента, що включає в себе культивування клітини за п. 21 в умовах, придатних для продукування антитіла і виділення продукованого антитіла або його фрагмента.
23. Застосування антитіла або його фрагмента за будь-яким з пп. 1-14 для виготовлення лікарського засобу для лікування Notch3-асоційованого захворювання або порушення.
24. Застосування за п. 23, де захворювання являє собою нейродегенеративне захворювання.
25. Застосування за п .23, де захворювання являє собою CADASIL, сімейну геміплегічну мігрень (FHM) сімейну парокситичну атаксію або синдром Алагілля.
26. Застосування антитіла або його фрагмента за будь-яким з пп. 1-14 для діагностики Notch3-асоційованого захворювання.
Текст
Реферат: Винахід належить до моноклонального антитіла, що специфічно зв'язується з амінокислотами 1384-1503 амінокислотної послідовності Notch3 (SEQ ID NO: 1). Винахід також належить до міченого антитіла, гуманізованого антитіла, нуклеїнової кислоти, вектора, клітини, що містять зазначене антитіло, способу одержання зазначеного антитіла та застосування його для виготовлення лікарського засобу для лікування Notch3-асоційованого захворювання. UA 102061 C2 (12) UA 102061 C2 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 ПЕРЕХРЕСНІ ПОСИЛАННЯ НА СПОРІДНЕНІ ЗАЯВКИ Дана заявка заявляє пріоритет на підставі попередньої заявки США № 60/852861, поданої 19 жовтня 2006, і попередньої заявки США № 60/879218, поданої 6 січня 2007, описи яких включені в даний документ у вигляді посилання у всій повноті. ГАЛУЗЬ ТЕХНІКИ ВИНАХОДУ Даний винахід стосується агоністичних антитіл проти Notch3 і їх застосування для поліпшення, лікування або профілактики Notch3-асоційованого захворювання або порушення. РІВЕНЬ ТЕХНІКИ Ген Notch вперше був описаний у 1917 р., коли було виявлено, що полотна крил штаму плодової мушки Drosophila melanogaster мають насічки (Morgan, Am Nat 51:513 (1917)). Майже сімдесят років потому ген клонували і встановили, що він кодує клітинний поверхневий рецептор, що грає ключову роль у розвитку багатьох типів клітин і тканин Drosophila (Wharton et al., Cell 43:567 (1985)). Несподівано було виявлено, що сигнальний шлях Notch являє собою механізм, опосередковуваний контактуванням клітин, і в еволюційному відношенні є консервативним від дрозофіли до людини. Виявлено, що рецептори Notch беруть участь у багатьох клітинних процесах, таких як диференціація, рішення про загибель клітини, підтримка стовбурових клітин, рухова активність клітин, проліферація й апоптоз, у різних типах клітин у процесі розвитку і гомеостазу тканин (огляд можна знайти в Artavanis-Tsakonas, et al., Science 268:225 (1995)). У ссавців виявлено чотири рецепторних білки Notch (які позначаються Notch1-Notch4) і п'ять відповідних лігандів (що позначаються дельта-подібний ліганд-1 (DLL-1), дельта-подібний ліганд-3 (DLL-3), дельта-подібний ліганд-4 (DLL-4), Jagged-1 і Jagged-2). Гени рецептора Notch ссавців кодують білки розміром 300 кДа, що розщеплюються при транспортуванні до клітинної поверхні і існують у вигляді гетеродимерів. Позаклітинний фрагмент рецептора Notch містить тридцять чотири епідермальні фактори росту (EGF)-подібних повторів і три повтори, збагачені цистеїном Notch/LIN12. Зв'язок двох розщеплюваних субодиниць здійснюється за участі послідовностей, що знаходяться безпосередньо в N-кінцевій і C-кінцевій області ділянки розщеплення, причому дві вказані субодиниці складають домени гетеродимеризації Notch (HD) (Wharton, et al., Cell 43:567 (1985); Kidd, et al., Mol Cell Biol 6:3431 (1986); Kopczynski, et al., Genes Dev 2:1723 (1988); Yochem, et al., Nature 335:547 (1988)). В даний час поки не ясно, як сигнальний шлях Notch регулюється різними рецепторами або як п'ять лігандів розрізняються за внеском в передачу сигналу або регуляцію. Розходження в передачі сигналу і/або регуляції можуть визначатися характером їхньої експресії в різних тканинах або різними умовами оточуючого середовища. Показано, що лігандні білки Notch, у тому числі Jagged/Serrate і дельта/дельта-подібні, здатні специфічно зв'язуватися з ділянкою повтору EGF і індукувати рецептор-опосередкований сигнальний шлях Notch (огляд опублікований Bray, Nature Rev Mol Cell Biol. 7:678 (2006), і Kadesch, Exp Cell Res. 260:1 (2000)). ий ий З повторів EGF 10 -12 необхідні для зв'язування ліганду з рецептором Notch, а інші повтори EGF можуть підсилювати взаємодію рецептора з лігандом (Xu, et al., J Biol Chem. 280:30158 (2005); Shimizu, et al., Biochem Biophys Res Comm. 276:385 (2000)). Хоча повтори LIN12 і домен димеризації не беруть участь безпосередньо в зв'язуванні ліганду, вони відіграють важливу роль у підтримці гетеродимерного білкового комплексу, запобігаючи ліганд-незалежне розщеплення під дією протеази й активацію рецептора (Sanche-Irizarry, et al., Mol Cell Biol. 24:9265 (2004); Vardar et al., Biochem. 42:7061 (2003)). Самопідтримка і визначення метаболічного шляху нормальних стовбурових клітин з багатьох тканин, у тому числі з тканини кишечнику і нервової тканини, залежать від сигнального шляху Notch (Fre, et al., Nature, 435: 964 (2005); van Es, et al., Nature, 435: 959 (2005); Androutsellis-Theotokis, et al., Nature, 442: 823 (2006)). Отже, агоністичне антитіло Notch3 можна використовувати для лікування дегенеративних захворювань. CADASIL (церебральна аутосомно-домінантна артеріопатія з підкірковими інфарктами і лейкоенцефалопатією) викликає інсульт і деменцію, характерні ознаки яких містять у собі підкіркові ішемічні події і мультиінфарктну деменцію. Виявлено, що CADASIL пов'язане з локалізацією мутантного гена в хромосомі 19 (Joutel, et al., Nature 383:707 (1996)). Joutel et al. ідентифікували в пацієнтів з CADASIL мутації, що викликають серйозне руйнування гена Notch 3, отже, пацієнти з CADASIL можуть мати дефектний білок Notch3. На жаль, дане захворювання, що є у високому ступені інвалідизуючим і часто летальним, в основному залишається недіагностованим, або діагностується помилково як розсіяний склероз і хвороба Альцгеймера. Проведені в даний час дослідження мають тенденцію демонструвати, що даний стан поширений набагато ширше, ніж вважалося спочатку. 1 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 Іншим прикладом Notch3-асоційованого захворювання є сімейна геміплегічна мігрень (FHM), домінантно-аутосомна форма мігрені з провісником епілептичного нападу, пов'язана з порушенням, розташованим у тій же ділянці хромосоми 19, що і ген Notch3. Слід вказати, що більш 30 % пацієнтів, що страждають від CADASIL, також страждають від мігрені з провісником епілептичного нападу. Однак останнє захворювання зустрічається приблизно тільки в 5 % населення, причому одержані результати свідчать про те, що ген Notch3 бере участь у механізмі даного стану. Подібним чином, сімейна парокситична атаксія пов'язана з геном, розташованим у тій же ділянці хромосоми 19, і можна зробити висновок, що Notch3 бере участь у розвитку даного стану. Інші стани і захворювання, пов'язані з Notch3, містять у собі синдром Алагілля (Flynn, et al., J Pathol 204:55 (2004)). В даний час проводять дослідження, щоб ідентифікувати інші захворювання і стани, пов'язані з недостатністю експресії і/або сигнального шляху Notch3. Оскільки існує велика кількість захворювань людини, пов'язаних із сигнальним шляхом Notch 3, важливо визначити нові способи профілактики і лікування даних захворювань. Даний винахід пропонує нові агоністичні антитіла проти Notch3, які можна використовувати для задоволення даної потреби медицини. СУТЬ ВИНАХОДУ Даний винахід пропонує нові агоністичні антитіла і їхні фрагменти, що специфічно зв'язуються з епітопом людського рецептора Notch3 у домені LIN12. Інший аспект даного винаходу містить у собі епітоп-зв'язувальну ділянку і антитіла, що зв'язують такий же епітоп, як і антитіла даного винаходу. Антитіла даного винаходу активують Notch3-опосередкований сигнальний шлях через рецептор Notch3 незалежно від зв'язування ліганду. Даний винахід пропонує амінокислотні послідовності варіабельних ділянок важких і легких ланцюгів антитіл і відповідні їм нуклеотидні послідовності. Інше втілення даного винаходу містить у собі послідовності CDR вказаних антитіл. Інше втілення даного винаходу містить у собі клітинні лінії і вектори, що несуть послідовності антитіл даного винаходу. Даний винахід також містить у собі епітоп, розпізнаваний агоністичними антитілами даного винаходу. Крім того, даний винахід містить у собі антитіла, що зв'язують даний епітоп. Втілення містять у собі епітоп Notch3, що містить домен Lin12, послідовність якого щонайменше на 80 %, 85 %, 90 % або 95 % ідентична SEQ ID NO: 10. Більш конкретно, епітоп Notch3 містить SEQ ID NO: 11. Даний винахід містить у собі агоністичні антитіла, які зв'язують даний епітоп. Інше втілення даного винаходу стосується застосування вказаних антитіл у виробництві лікарського засобу або композиції для лікування Notch3-асоційованих захворювань і порушень, пов'язаних, наприклад, з інактивацією рецептора. Інше втілення даного винаходу стосується застосування вказаних антитіл для лікування Notch3-асоційованих захворювань або порушень, пов'язаних, наприклад, з інактивацією рецептора, що включає в себе компенсацію вказаних дефектів, наприклад, шляхом активації сигнального шляху Notch3 незалежно від зв'язування ліганду. Notch3-асоційовані порушення можуть містити в собі, без обмеження, CADASIL, сімейну геміплегічну мігрень (FHM), сімейну парокситичну атаксію, синдром Алагілля та інші дегенеративні захворювання. КОРОТКИЙ ОПИС ФІГУР На фіг.1 зображена амінокислотна послідовність Notch3. Ділянка повтору EGF простирається від амінокислотного залишку 43 до амінокислотного залишку 1383; домен LIN12 простирається від амінокислотного залишку 1384 до амінокислотного залишку 1503; і домен димеризації простирається від амінокислотного залишку 1504 до амінокислотного залишку 1640. На фіг.2 (A-H) наведене порівняльне зображення амінокислотних послідовностей людських Notch1, Notch2, Notch3 і Notch4. На фіг.3 показаний відсоток ідентичності Notch 1, Notch 2, Notch 3 і Notch 4. На фіг.4A і 4B зображені послідовності варіабельних ділянок важкого і легкого ланцюга моноклонального антитіла MAb 256A-13 проти Notch3 (SEQ ID NO: 2), причому ділянки CDR підкреслені. На фіг.5 приведені результати люциферазного репортерного аналізу приклада 5, що демонструють активуючу дію MAb проти Notch3 на рецептор Notch3. На фіг.6 показаний вплив агоністичних антитіл Notch3 на металопротеазне розщеплення Notch3. На фіг.7 зображено епітопне картування зв'язувальної ділянки 256A-13 з конструкцій гібридних білків Notch3-Fc. 2 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 На фіг.8 наведене порівняльне зображення одержаної рекомбінантним методом послідовності, що кодує лідерний пептид Notch3, і нативної послідовності, що кодує лідерний пептид Notch3 (номер доступу в NCBI GenBank: NM_000435), що демонструє, що рекомбінантна послідовність, що кодує лідерний пептид Notch, несе зміни в нуклеотидній сполуці (8A) і обумовлює зміни трансльованої амінокислотної послідовності (8B). На фіг.8C зображений домен LIN12, а на фіг.8D зображений епітоп субдомену LIN12. На фіг.9 зображене одержання конструкції з переставленими доменами методом PCR-SOE. Прямокутники зі страліками позначають праймери ПЛР. Незафарбовані прямокутники належать до послідовності Notch3. Зафарбовані прямокутники належать до послідовності Notch1. На фіг.10 зображені амінокислотні послідовності, використовувані при епітопному картуванні домену LIN12 Notch3 з MAb 256A-13. На фіг.11 зображені аланінові скануючі пептиди, використовувані для лінійного епітопного картування 256A-13. ДОКЛАДНИЙ ОПИС ВИНАХОДУ Даний винахід не обмежується описаними тут конкретними методами, схемами, клітинними лініями, векторами або реагентами, оскільки вони можуть варіювати. Крім того, використовувана в даному документі термінологія призначена тільки для опису конкретних втілень і не обмежує обсяг даного винаходу. У даному описі й у формулі винаходу, що додається, якщо контекст не вказує інакше, однина містить у собі множину, наприклад, термін "клітина-хазяїн" містить у собі сукупність таких клітин-хазяїнів. Якщо не вказано інакше, усі використовувані тут технічні і наукові терміни, а також усі скорочення мають традиційні значення, відомі рядовим фахівцям в галузі даного винаходу. У даному описі наводяться приклади способів, пристроїв і матеріалів, однак при здійсненні даного винаходу можна використовувати будь-які способи і матеріали, подібні або еквівалентні описаним. Усі згадані тут патенти і публікації включені в даний опис у вигляді посилання в ступені, що допускається законом для опису і розкриття вказаних у них білків, ферментів, векторів, клітинхазяїнів і методів, придатних для застосування в даному винаході. Однак наведену тут інформацію не слід розглядати як допускання, що даний винахід надає право розцінювати цю інформацію як рівень техніки і протиставляти її попередньому винаходу. Визначення Терміни, використовувані за текстом даної заявки, варто інтерпретувати в їхніх традиційних значеннях, відомих рядовому фахівцю в даній галузі. Однак нижченаведені терміни автори даної заявки пропонують використовувати в конкретних, вказаних нижче значеннях. Фраза "практично ідентичний" у відношенні послідовності поліпептидного ланцюга антитіла може означати, що послідовність антитіла щонайменше на 70 %, або 80 %, або 90 %, або 95 %, ідентична порівнюваної поліпептидної послідовності. У відношенні нуклеотидної послідовності даний термін може означати, що послідовність нуклеотидів щонайменше на 85 %, або 90 %, або 95 %, або 97 % ідентична порівнюваній нуклеотидній послідовності. Термін "ідентичність" або "гомологія" варто розуміти, як відсоток амінокислотних залишків у послідовності-кандидаті, ідентичних залишкам послідовності, з яким порівнюють послідовністькандидат, обумовлений після вирівнювання послідовності і введення пробілів, якщо це необхідно для досягнення максимального відсотка ідентичності всієї послідовності, причому консервативні заміни не розглядаються як частину ідентичності послідовності. N- або C-кінцеві або подовження вставки не повинні тлумачитися як зменшення ідентичності або гомології. Способи вирівнювання і комп'ютерні програми для їх здійснення добре відомі в даній галузі. Ідентичність послідовності можна визначити за допомогою програми для аналізу послідовностей. Термін "антитіло" використовується в самому широкому сенсі, а саме, він охоплює моноклональні антитіла (у тому числі повнорозмірні моноклональні антитіла), поліклональні антитіла, і поліспецифічні антитіла (наприклад, біспецифічні антитіла), а також фрагменти антитіл, що мають бажану біологічну активність. Антитіла (Ab) і імуноглобуліни (IgG) являють собою глікопротеїни, що мають однакові структурні особливості. Тоді як антитіла мають специфічність зв'язування відносно конкретної мішені, імуноглобуліни містять у собі й антитіла, і інші антитіло-подібні молекули, що не мають специфічність відносно мішені. Антитіла даного винаходу можуть належати до будь-якого типу (наприклад, IgG, IgE, IgM, IgD, IgA і IgY), класу (наприклад, IgG1, IgG2, IgG3, IgG4, IgA1 і IgA2) або підкласу. Нативні антитіла та імуноглобуліни, як правило, являють собою гетеротетрамерні глікопротеїни розміром приблизно 150000 Дальтон, що складаються з двох ідентичних легких (L) ланцюгів і двох ідентичних важких (H) ланцюгів. Кожен важкий ланцюг містить на одному кінці варіабельний домен (V H), за 3 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 яким іде ряд константних доменів. Кожен легкий ланцюг містить на одному кінці варіабельний домен (VL), а на іншому кінці константний домен. У даному описі термін "антитіло проти Notch3" стосується антитіла, що специфічно зв'язується з людським Notch3, активуючи сигнальний шлях Notch 3 незалежно від ліганду. Термін "варіабельний" у застосуванні до домену антитіла означає, що послідовності деяких ділянок варіабельних доменів сильно розрізняються у різних антитіл і відповідають за специфічне зв'язування конкретного антитіла з конкретною мішенню. Однак протягом варіабельних доменів антитіл варіабельність розподіляється не рівномірно. Вона концентрується в трьох сегментах, що називаються ділянки, які визначають комплементарність (CDR; тобто, CDR1, CDR2 і CDR3), або гіперваріабельні ділянки, що присутні у варіабельних доменах як легкого, так і важкого ланцюгів. Більш консервативні фрагменти варіабельних доменів називають каркасними ділянками (FR). Усі варіабельні домени нативних важких і легких ланцюгів містять чотири ділянки FR, які, як правило, мають β-складчасту конфігурацію і з'єднуються трьома CDR, що утворюють петлеподібну сполуку і, у деяких випадках, частину βскладчастої структури. CDR одного ланцюга містяться разом у безпосередній близькості з ділянками FR і разом з CDR іншого ланцюга беруть участь у формуванні ділянки зв'язування антитіла з мішенню (див. Kabat, et al. Sequences of Proteins of Immunological Interest, National Institute of Health, Bethesda, Md. (1987)). Якщо не вказано інакше, у даному описі для нумерації амінокислотних залишків імуноглобулінів використовується система нумерації амінокислотних залишків імуноглобулінів Kabat, et al. Термін "фрагмент антитіла" стосується фрагмента повнорозмірного антитіла, як правило, ділянки, що зв'язує мішень, або до варіабельної ділянки. Приклади фрагментів антитіл містять у собі фрагменти F(ab), F(ab'), F(ab') 2 і Fv. Фраза "функціональний фрагмент або аналог" антитіла стосується сполуки, що має кількісну біологічну активність, подібну біологічній активності повнорозмірного антитіла. Наприклад, функціональний фрагмент або аналог антитіла проти Notch3 здатний зв'язуватися з рецептором Notch3, запобігаючи або суттєво знижуючи здатність рецептора зв'язуватися з лігандами або ініціювати передачу сигналу. У даному описі термін "функціональний фрагмент" у застосуванні до антитіл стосується фрагментів Fv, F(ab) і F(ab') 2. Фрагмент "Fv" являє собою димер, що складається з варіабельного домену одного важкого і варіабельного домену одного легкого ланцюга, що знаходяться в тісному нековалентному зв'язку (димер VH-VL). У даній конфігурації три CDR кожного варіабельного домену взаємодіють, утворюючи ділянку зв'язування з мішенню на поверхні димеру VH-VL. Разом шість CDR визначають здатність антитіла специфічно зв'язуватися з мішенню. Однак навіть один варіабельний домен (або половина Fv, що містить тільки три CDR, специфічні до мішені) має здатність розпізнавати і зв'язувати мішень, хоча і з більш низькою спорідненістю, ніж ціла зв'язувальна ділянка. Фрагменти антитіл "одноланцюжковий Fv" або "sFv" містять VH і VL домени антитіла, причому вказані домени знаходяться в одному поліпептидному ланцюзі. Як правило, поліпептид Fv також містить поліпептидний лінкер між доменами VH і VL, що дозволяє sFv формувати структуру, необхідну для зв'язування мішені. Термін "діатіла" стосується маленьких фрагментів антитіл, що містять дві антигензв'язувальні ділянки, причому вказані фрагменти в одному поліпептидному ланцюзі містять варіабельний домен важкого ланцюга (VH), з'єднаний з варіабельним доменом легкого ланцюга (VL). Шляхом застосування лінкера, що є занадто коротким, щоб дозволити спаритися двом доменам, розташованим на одному ланцюзі, домени змушують спаровуватися з комплементарними доменами іншого ланцюга з формуванням двох антиген-зв'язувальних ділянок. Фрагмент F(ab) містить константний домен легкого ланцюга і перший константний домен (CH1) важкого ланцюга. Фрагменти F(ab') відрізняються від фрагментів F(ab) додаванням декількох залишків за карбоксильним кінцем домену СН1 важкого ланцюга, у тому числі, одного або декількох залишків цистеїну в шарнірній ділянці антитіла. Фрагменти F(ab') одержують шляхом розщеплення дисульфідного зв'язку, утвореного залишками цистеїну шарнірної ділянки, у продукті розщеплення пепсином F(ab') 2. Фахівцям у даній галузі відомі та інші хімічні способи сполуки фрагментів антитіл. Термін "моноклональне антитіло" у даному описі стосується антитіла, одержаного з популяції практично гомогенних антитіл, тобто, з популяції, окремі антитіла якої є ідентичними за винятком можливих природних мутацій, що можуть бути присутнім у мінорних кількостях. Моноклональні антитіла відповідно до даного опису містять у собі "химерні" антитіла (імуноглобуліни), в яких частина важкого і/або легкого ланцюга ідентична або гомологічна відповідній послідовності антитіла, одержаного від тваринного конкретного виду або 4 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 приналежного до конкретного або класу підкласу антитіл, а інша частина ланцюга (ланцюгів) ідентична або гомологічна відповідній послідовності антитіла, одержаного від тварини іншого виду або приналежного до іншого або класу підкласу антитіл, а також фрагменти таких антитіл, що мають бажану біологічну активність (патент США № 4816567; і Morrison, et al., Proc Natl Acad Sci USA 81:6851 (1984)). Моноклональні антитіла мають високу специфічність і спрямовані проти однієї ділянки мішені. Крім того, на відміну від традиційних (поліклональних) препаратів антитіл, що зазвичай складаються з різних антитіл, спрямованих проти різних детермінант (епітопів), кожне моноклональне антитіло спрямоване проти однієї детермінанти мішені. Крім специфічності перевагою моноклональних антитіл є можливість їх синтезу в гібридомної культурі, що дозволяє уникнути забруднення іншими імуноглобулінами. Визначення "моноклональний" вказує на те, що антитіло одержане з практично гомогенної популяції антитіла, і не повинно тлумачитися як віднесення до будь-якого конкретного способу одержання. Наприклад, моноклональні антитіла, придатні для застосування в даному винаході, можна виділити з фагових бібліотек антитіл, використовуючи добре відомі методи. Вихідні моноклональні антитіла для застосування відповідно до даного винаходу можна одержати за допомогою гібридомного методу, вперше описаного Kohler, et al, Nature 256:495 (1975), або за допомогою рекомбінантних методів. "Гуманізовані" форми нелюдських (наприклад, мишачих) антитіл являють собою химерні імуноглобуліни, ланцюги або імуноглобулінів їхні фрагменти (такі як Fv, Fab, Fab', F(ab') 2 або інші мішень-зв'язувальні субпослідовності антитіл), що містять мінімальну послідовність нелюдського імуноглобуліну. Як правило, практично усі варіабельні домени гуманізованого антитіла з щонайменше одного або, найчастіше, двох, містять усі або майже всі ділянки CDR, що відповідають нелюдському імуноглобуліну, і всі або майже всі ділянки FR, що відповідають матричній послідовності людського імуноглобуліну. Гуманізоване антитіло також може містити щонайменше частину константної ділянки γFc) імуноглобуліну, як правило, вибраного з людської матриці імуноглобуліну. Терміни "клітина", "клітинна лінія" і "клітинна культура" містять у собі потомство. Варто розуміти, що не все потомство може бути цілком ідентичним батьківським клітинам за вмісту ДНК внаслідок присутності навмисних або ненавмисних мутацій. В обсяг даного винаходу також входить варіантне потомство, що має такі ж функції або біологічні властивості, як і вихідна трансформована клітина. "Клітини-хазяїни", використовувані в даному винаході, зазвичай являють собою прокаріотичні або еукаріотичні клітини. "Трансформацію" клітинного організму, або клітини клітинної лінії за допомогою ДНК здійснюють шляхом введення ДНК у клітину-мішень або у вигляді позахромосомного елемента, або за допомогою вбудовування в хромосому, за допомогою способу, що дозволяє зберегти здатність ДНК до реплікації. "Трансфекція" клітини або організму молекулою ДНК являє собою поглинання ДНК, наприклад, вектора експресії, клітиною або організмом незалежно від того, чи експресується фактично будь-яка кодуюча послідовність, чи ні. Терміни "трансфікована клітинахазяїн" і "трансформована клітина-хазяїн" належать до клітини, в яку введена ДНК. Клітина, що називається "клітина-хазяїн", може бути прокаріотичною або еукаріотичною. Типові прокаріотичні клітини-хазяїни містять у собі різні штами E. coli. Типові еукаріотичні клітинихазяїни являють собою клітини ссавців, такі як клітини яєчника китайського хом'ячка або людські клітини. Послідовність ДНК, що вводиться,може належати до того ж виду, що і клітина-хазяїн, або до іншого виду, або вона може являти собою гібридну послідовність ДНК, що містить декілька чужорідних і декілька гомологічних ДНК. Термін "вектор" стосується конструкції ДНК, що містить послідовність ДНК, функціонально зв'язану з придатною контрольною послідовністю, здатною забезпечувати експресію ДНК у придатному хазяїні. Такі контрольні послідовності містять у собі промотор, що забезпечує транскрипцію, необов'язкову послідовність оператора, що контролює таку транскрипцію, послідовність, що кодує придатні ділянки зв'язування мРНК рибосомою, і послідовності, що контролюють термінацію транскрипції і трансляції. Вектор може являти собою плазміду, фагову частку або просто потенційну геномну вставку. Після трансформації в придатний хазяїн вектор може реплікуватися і функціонувати незалежно від генома хазяїна, або в деяких випадках, він може інтегруватися в геном як такий. У даному описі терміни "плазміда" і "вектор" іноді використовуються як взаємозамінні, оскільки плазміда являє собою найбільш широко використовувану форму вектора. Однак передбачається, що даний винахід містить у собі та інші, відомі в даній галузі, форми векторів, що виконують еквівалентну функцію. "Ссавець", що піддається лікуванню, може являти собою будь-яку тварину, що стосується класу ссавців, і містить у собі людину, ручних і сільськогосподарських тварин, відмінних від 5 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 людини приматів, а також диких тварин, спортивних тварин або домашніх тварин, таких як собаки, коні, кішки, корови та ін. Термін "маркер" у даному описі стосується детектованої сполуки детектованої композиції, які можуть бути кон'юговані безпосередньо або через інші сполуки з молекулою або білком, наприклад, антитілом. Маркер сам може бути детектованим (наприклад, радіоізотопні мітки або флуоресцентні мітки) або, як у випадку ферментного маркера, може каталізувати детектовану хімічну зміну субстратної сполуки або субстратної композиції. У даному описі термін "тверда фаза" означає неводну основу, до якої може прилипати антитіло даного винаходу. Приклади твердих фаз, що входять в обсяг даного винаходу, містять у собі фази, одержані частково або цілком зі скла (наприклад, скло з контрольованим розміром пор), полісахариди (наприклад, агароза), поліакриламіди, полістирол, полівініловий спирт і кремнійорганічні сполуки. У деяких утіленнях, залежно від контексту, до твердої фази може відноситися ямка аналітичного планшета; в інших втіленнях тверда фаза може являти собою колонку для очищення (наприклад, колонку для афінної хроматографії). У даному описі термін "Notch3-опосередковане порушення" стосується стану або захворювання, що характеризується дефектною або зниженою експресією рецептора Notch3. А саме, даний термін може містити в собі стани, асоційовані з дегенеративними захворюваннями, такі як CADASIL, FHM, сімейна парокситична атаксія, синдром Аллагіля та інші дегенеративні захворювання. Рецептор NOTCH 3 як імуноген, що викликає утворення антитіл Розчинні мішені або їхні фрагменти можна використовувати як імуногени для одержання антитіл. Антитіло направлене проти мішені, яка представляє інтерес. Переважно, мішенню є біологічно важливий поліпептид, і введення антитіла ссавцю, що страждає від захворювання або порушення, може робити сприятливу терапевтичну дію на ссавця. Як імуноген для одержання антитіл можна використовувати цілі клітини. Імуноген можна одержати за допомогою рекомбінантних або синтетичних способів. Імуноген також можна виділити з природного джерела. У випадку трансмембранних молекул, таких як рецептори, як імуноген можна використовувати їхні фрагменти (наприклад, позаклітинний домен рецептора). Альтернативно, як імуноген можна використовувати клітини, що експресують трансмембранні молекули. Такі клітини можна одержати з природного джерела (наприклад, з лінії ракових клітин) або шляхом трансформації вихідних клітин з використанням рекомбінантних методів, що забезпечують підвищену експресію трансмембранної молекули. Фахівцям у даній галузі відомі та інші форми імуногенів, які можна використовувати для одержання антитіл. Альтернативно, ген або кДНК, що кодує людський рецептор Notch3, можна клонувати у плазміді або іншому векторі експресії і експресувати у будь-якій системі експресії за допомогою способів, добре відомих фахівцям у даній галузі. Способи клонування й експресії рецептора Notch3, а також кодуючої його нуклеотидної послідовності, описані в літературі (див., наприклад, патенти США №№ 5821332 і 5759546). Внаслідок виродженості генетичного коду можна використовувати велику кількість нуклеотидних послідовностей, що кодують рецепторний або білок поліпептиди Notch3. Нуклеотидну послідовність можна варіювати шляхом добору поєднань, виходячи з можливих варіантів кодонів. Дані поєднання одержують відповідно до триплетного генетичного коду, використовуваним у нуклеотидній послідовності, що кодує природний рецептор Notch3, причому можна розглядати всі одержані варіації. Будьякий з вказаних поліпептидів можна використовувати для імунізації тварини з метою одержання антитіл, що зв'язують людський рецептор Notch3. Внаслідок високої консервативності амінокислотної послідовності Notch3 як імуногенів для одержання антитіл можна використовувати також рекомбінантні білки Notch3 інших видів. Порівняння людського і мишачого Notch3 показує, що понад 90 % амінокислотних послідовностей двох видів є ідентичними. Імуногенний рецептор Notch3 за необхідності можна експресувати у вигляді гібридного білка, що містить рецептор Notch3, з'єднаний з гібридизаційним фрагментом. Гібридизаційний фрагмент часто полегшує очищення білка, наприклад, дозволяє виділити й очистити гібридний білок методом афінної хроматографії, і, крім того, його також можна використовувати для підвищення імуногенності. Гібридні білки можна одержати шляхом культивування рекомбінантної клітини, трансформованої гібридною нуклеотидною послідовністю, що кодує білок із приєднаним по карбоксильному або аміно-кінцю гібридизаційним фрагментом. Гібридизаційні фрагменти можуть містити в собі, без обмеження, ділянки Fc імуноглобулінів, глутатіон-S-трансферазу, β-галактозидазу, полігістидиновий фрагмент, здатний зв'язуватися з двовалентним іоном металу, і білок, що зв'язує мальтозу. 6 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Рекомбінантний рецепторний білок Notch3, як описано в прикладі 1, використовують для імунізації мишей з метою одержання гібридом, що продукують моноклональні антитіла даного винаходу. Приклади поліпептидів містять у собі повнорозмірну послідовність SEQ ID NO: 1 і її фрагменти, або їхні варіанти. Одержання антитіл Антитіла даного винаходу можна одержати за допомогою будь-якого придатного способу, відомого в даній галузі. Антитіла даного винаходу можуть містити в собі поліклональні антитіла. Способи одержання поліклональних антитіл відомі досвідченим фахівцям (Harlow, et al., Antibodies: a Laboratory Manual, Cold spring Harbor Laboratory Press, 2nd ed. (1988)), зміст даної публікації включений в даний опис у вигляді посилання у всій повноті). Наприклад, як описано в прикладі 1, імуноген можна вводити різним тваринам-хазяїнам, що включають у себе, без обмеження, кроликів, мишей, щурів і ін., індукуючи продукцію сироватки, що містить поліклональні антитіла, специфічні до антигена. Введення імуногена може містити в собі одну або декілька ін'єкцій імунізуючого засобу і, за необхідності, ад'юванту. Для посилення імунологічної відповіді можна використовувати різні ад'юванти, що залежать від виду хазяїна і містять у собі, без обмеження, ад'ювант Фрейнда (повний і неповний), мінеральні гелі, такі як гідроксид алюмінію, поверхнево-активні речовини, такі як лізолецитин, плюронілові поліоли, поліаніони, пептиди, масляні емульсії, гемоціанін лімфи равлика, динітрофенол і потенційно корисні людські ад'юванти, такі як BCG (бацила Кальметта-Герена) і Corynebacterium parvum. Інші приклади придатних для використання ад'ювантів містять у собі ад'ювант MPL-TDM (монофосфорил ліпіду A, синтетичний дикориноміколат трегалози). Схеми імунізації добре відомі в даній галузі і можуть проводитися будь-якими відомими в даній галузі способами, що викликають імунну відповідь у вибраної тварини-хазяїна. Ад'юванти також добре відомі в даній галузі. Як правило, імуноген (разом з ад'ювантом або без ад'юванту) вводять ссавцю шляхом декількох підшкірних або внутрішньочеревинних ін'єкцій, або внутрішньом'язово або внутрішньовенно. Імуноген може містити в собі поліпептид Notch3, гібридний білок або їхні варіанти. Залежно від природи поліпептиду (тобто, від ступеня гідрофобності, ступеня гідрофільності, стабільності, сумарного заряду, ізоелектричної точки та ін.), іноді корисно кон'югувати імуноген з білком, що свідомо є імуногенним для імунізованого ссавця. Таке кон'югування проводять або хімічними способами шляхом дериватизації активних хімічних функціональних груп на підлягаючих кон'югуванню імуногенах або імуногенних білках з утворенням ковалентного зв'язку, або за допомогою способів одержання гібридних білків, або інших способів, відомих фахівцям у даній галузі. Приклади таких імуногенних білків містять у собі, без обмеження, гемоціанін лімфи равлика, овальбумін, сироватковий альбумін, бичачий тиреоглобулін, інгібітор соєвого трипсину і різні пептиди T-хелперів. Як вказано вище, для посилення імунної відповіді можна використовувати різні ад'юванти. Антитіла даного винаходу містять у собі моноклональні антитіла. Моноклональні антитіла являють собою антитіла, що розпізнають одну антигенну ділянку. Їхня однакова специфічність робить моноклональні антитіла набагато більш корисними, ніж поліклональні антитіла, що зазвичай містять антитіла, що розпізнають декілька різних антигенних ділянок. Моноклональні антитіла можна одержати з використанням гібридомної технології, такої як описана в Kohler, et al., Nature 256:495 (1975); Патент США № 4376110; Harlow, et al., Antibodies: A Laboratory Manual, Cold spring Harbor Laboratory Press, 2nd ed. (1988) і Hammerling, et al., Monoclonal Antibodies and T-Cell Hybridomas, Elsevier (1981), методів рекомбінантних ДНК або інших методів, відомих фахівцям у даній галузі. Інші приклади методів, які можна використовувати для одержання моноклональних антитіл, містять у собі, без обмеження, метод гібридом людських Bклітин (Kosbor, et al., Immunology Today 4:72 (1983); Cole, et al., Proc Natl Acad Sci USA 80:2026 (1983)), і метод EBV-гібридом (Cole, et al., Monoclonal Antibodies and Cancer Therapy, pp. 77-96, Alan R. Liss (1985)). Такі антитіла можуть належати до будь-якого класу імуноглобулінів, у тому числі IgG, IgM, IgE, IgA, IgD, а також до будь-якого підкласу. Гібридоми, що продукуються MAb даного винаходу, можна культивувати in vitro або in vivo. У гібридомному методі хазяїна, такого як миша, гуманізована миша, миша з людською імунною системою, хом'як, кролик, верблюд або будь-якого іншого придатного тварину-хазяїна імунізують, щоб викликати утворення лімфоцитів, що продукують або здатних продукувати антитіла, що специфічно зв'язуються з білком, використовуваним для імунізації. Альтернативно, лімфоцити можна імунізувати in vitro. Потім лімфоцити гібридизують із клітинами мієломи, використовуючи придатний гібридизуючий агент, такий як поліетиленгліколь, і одержують гібридомні клітини (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp.59103 (1986)). 7 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Як правило, для одержання гібридом, що продукують антитіла, використовують або лімфоцити периферичної крові ("PBL"), якщо потрібні людські клітини, або клітини селезінки або лімфатичного вузла, якщо потребуються клітини ссавця, відмінного від людини. Потім лімфоцити гібридизують з іморталізованою клітинною лінією, використовуючи придатний гібридизуючий агент, такий як поліетиленгліколь, і одержують гібридомні клітини (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp. 59-103 (1986)). Іморталізовані клітинні лінії, як правило, являють собою трансформовані клітини ссавців, зокрема, мієломні клітини гризунів, корів або людину. Зазвичай використовують лінію клітин мієломи щура або миші. Гібридомні клітини можна культивувати в придатному поживному середовищі, яке переважно містить одну або декілька речовин, що інгібують ріст або виживання негібридизованих іморталізованих клітин. Наприклад, якщо батьківські клітини не містять фермент гіпоксантингуанін-фосфорибозилтрансферазу (HGPRT або HPRT), поживне середовище для гібридом зазвичай містить гіпоксантин, аміноптерин і тимідин ("середовище HAT"), речовини, що запобігають росту HGPRT-дефіцитних клітин. Переважними іморталізованими клітинними лініями є лінії, що ефективно гібридизуються, зберігають стабільний високий рівень продукції антитіл вибраними антитіло-продукуючими клітинами і є чутливими до такого середовища, як HAT. До вказаних мієломних клітинних лініях належать лінії клітин мишачої мієломи, наприклад, одержані з мишачих пухлин MOPC-21 і MPC11, які можна одержати від Salk Institute Cell Distribution Center, San Diego, Calif. U.S. Application No., і клітини SP2/0 або X63-Ag8-653, які можна одержати від Американської колекції типових культур, Rockville, Md. USA. Також описано, що для одержання людських моноклональних антитіл можна використовувати лінії клітин людської мієломи і мишачо-людської гетеромієломи (Kozbor, J Immunol 133:3001 (1984); Brodeur, et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc, pp.51-63 (1987)). Крім того, можна використовувати лінію клітин мишачої мієломи NSO (Европейська колекція клітинних культур, Salisbury, Wilshire, UK). Культуральне середовище, в якому вирощують гібридомні клітини, аналізують на присутність моноклональних антитіл проти Notch3. Специфічність зв'язування моноклональних антитіла, що продукуються гібридомними клітинами, можна визначити методом імунопреципітації або за допомогою аналізу зв'язування in vitro, такого як радіоімунологічний аналіз (RIA) або імуноферментний твердофазний аналіз (ELISA). Такі методи відомі в даній галузі і належать до сфери компетенції відповідних фахівців. Спорідненість зв'язування моноклонального антитіла проти Notch3 можна визначити, наприклад, за допомогою аналізу Скетчарда (Munson, et al., Anal Biochem 107:220 (1980)). Після ідентифікації гібридомних клітин, що продукують антитіла з бажаними специфічністю, афіннстю і/або активністю, клони можна субклонувати методом серійних розведень і вирощувати стандартними методами (Goding, Monoclonal Antibodies: Principles and Practice, Academic Press, pp.59-103 (1986)). Придатні для даної мети культуральні середовища містять у собі, наприклад, модифіковану по Дульбекко середовище Голка (D-MEM) або середовище RPMI-1640. Крім того, гібридомні клітини можна вирощувати in vivo в організмі тварини у вигляді асцитних пухлин. Моноклональні антитіла, що секретуються субклонами, можна відокремити або виділити з культурального середовища, асцитної рідини або сироватки за допомогою традиційних методів очищення імуноглобулінів, таких як, наприклад, застосування білок A-сефарози, хроматографія на гідроксилапатиті, ексклюзійна гель-хроматографія, гель-електрофорез, діаліз або афінна хроматографія. У даній галузі існує ряд способів одержання моноклональних антитіл і, отже, дане винахід не обмежується тільки їх продукцією в гібридомах. Наприклад, моноклональні антитіла можна одержати з використанням методів рекомбінантних ДНК, таких як описані в патенті США № 4816567. У даному контексті термін "моноклональні антитіло" стосується антитіла, одержаного з одного еукаріотичного, фагового або прокаріотичного клону. ДНК, що кодує моноклональні антитіла даного винаходу, можна легко виділити і секвенувати за допомогою традиційних способів (наприклад, з використанням олігонуклеотидних зондів, здатних специфічно зв'язуватися з генами, що кодують важкі і легкі ланцюги мишачих антитіл, або такі ланцюги з людських, гуманізованих або інших джерел) (Innis, et al. In PCR Protocols. A Guide to Methods and Applications, Academic (1990), Sanger, et al., Proc Natl Acad Sci 74:5463 (1977)). Клітини гібридоми служать джерелом ДНК. Після виділення ДНК можна помістити у вектори експресії, якими потім трансфікують клітини-хазяїни, такі як клітини E. coli, клітини NSO, мавпячі клітини COS, клітини яєчника китайського хом'ячка (CHO), або клітини мієломи, що зазвичай не продукують імуноглобулінові білки, забезпечуючи синтез моноклональних антитіл у рекомбінантних клітинах-хазяїнах. ДНК також можна модифікувати, наприклад, шляхом 8 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 введення послідовностей, що кодують константні домени людських важких і легких ланцюгів замість гомологічних мишачих послідовностей (патент США № 4816567; Morrison, et al., Proc Natl Acad Sci USA 81:6851 (1984)) або шляхом ковалентного приєднання до послідовності, що кодує імуноглобулін, повнорозмірної або часткової послідовності, що кодує поліпептид, відмінний від імуноглобуліну. Такий, відмінний від імуноглобуліну, поліпептид може використовуватися замість константних доменів антитіла даного винаходу або замість варіабельних доменів однієї антиген-зв'язувальної ділянки антитіла даного винаходу з одержанням химерного бівалентного антитіла. Антитіла можуть бути моновалентними. Способи одержання моновалентних антитіл добре відомі в даній галузі. Наприклад, один спосіб містить у собі рекомбінантну експресію легкого ланцюга і модифікованого важкого ланцюга імуноглобуліну. Як правило, важкий ланцюг укорочують по будь-якій точці ділянки Fc, щоб запобігти поперечного зв'язування важких ланцюгів. Альтернативно, щоб запобігти поперечного зв'язування, відповідні залишки цистеїну заміняють на інші амінокислотні залишки або видаляють. Фрагменти антитіла, що розпізнають конкретні епітопи, можна одержати за допомогою відомих методів. Зазвичай такі фрагменти одержують шляхом протеолітичного розщеплення інтактного антитіла (див., наприклад, Morimoto, et al., J Biochem Biophys Methods 24:107 (1992); Brennan, et al., Science 229:81 (1985)). Наприклад, фрагменти Fab і F(ab') 2 дані винаходи можна одержати шляхом протеолітичного розщеплення молекул імуноглобуліну, використовуючи такі ферменти, як папаїн (з одержанням фрагментів Fab) або пепсин (з одержанням фрагментів F(ab')2). Фрагменти F(ab')2 містять варіабельну ділянку, константна ділянка легкого ланцюга і домен CH1 важкого ланцюга. Однак дані фрагменти можуть безпосередньо продукуватися рекомбінантними клітинами-хазяїнами. Наприклад, фрагменти антитіл можна виділити з фагової бібліотеки антитіл. Альтернативно, фрагменти F(ab') 2-SH можна безпосередньо виділити з E. coli і хімічно з'єднати з одержанням фрагментів F(ab') 2 (Carter, et al., Bio/Technology 10:163 (1992). Відповідно до іншого підходу, фрагменти F(ab')2 можна безпосередньо виділити з культури рекомбінантних клітин-хазяїнів. Кваліфікованим фахівцям відомі та інші способи одержання фрагментів антитіл. В інших втіленнях вибране антитіло являє собою одноланцюжковий фрагмент Fv (Fv) (патентна заявка PCT WO 93/16185). У деяких випадках, що включають у себе застосування антитіл у людей in vivo і детекційні аналізи in vitro, переважно використовувати химерні, гуманізовані або людські антитіла. Химерне антитіло являє собою молекулу, у якій різні фрагменти антитіл одержані від тварин різних видів, таку як антитіло, що містить варіабельну ділянку мишачого моноклонального антитіла і константну ділянку людського імуноглобуліну. Способи одержання химерних антитіл відомі в даній галузі. Див., наприклад, Morrison, Science 229:1202 (1985); Oi, et al., BioTechniques 4:214 (1986); Gillies, et al., J Immunol Methods 125:191 (1989); патенти США №№ 5807715; 4816567; і 4816397, що включені в даний опис посиланнями у всій повноті. Гуманізоване антитіло має більш високий ступінь гомології відносно людського імуноглобуліну, ніж моноклональні антитіла, одержані з тварин. Гуманізація являє собою спосіб одержання химерного антитіла, в якому фрагмент, замінений на відповідну нелюдську послідовність, суттєво менше інтактного людського варіабельного домену. Гуманізовані антитіла являють собою молекули антитіл, що продукуються тваринами відмінних від людини видів, що зв'язують цільовий антиген і містять одну або декілька гіперваріабельних ділянок (CDR) відмінних від людини видів і каркасні ділянки (FR) молекули людського імуноглобуліну. Найчастіше, щоб змінити, переважно поліпшити, зв'язування антигена, каркасні залишки в людських каркасних ділянках заміняють на відповідні залишки антитіла-донора CDR. Вказані заміни в каркасній ділянці визначають за допомогою добре відомих у даній галузі способів, наприклад, шляхом моделювання взаємодій залишків CDR і каркасної ділянки, що дозволяє ідентифікувати залишки каркасної ділянки, що беруть участь у зв'язуванні антигена, і порівняння послідовностей, що дозволяє ідентифікувати незвичайні залишки каркасної ділянки в конкретних положеннях. Див., наприклад, патент США № 5585089; Riechmann, et al., Nature 332:323 (1988), що цілком включений у даний опис у вигляді посилання. Антитіла можна гуманізувати за допомогою ряду способів, відомих у даній галузі, таких як щеплення CDR (EP 239400; публікація PCT WO 91/09967; патенти США №№ 5225539; 5530101; і 5585089), або облицювання нанесення нового покриття (EP 592106; EP 519596; Padlan, Molecular Immunology 28:489 (1991); Studnicka, et al., Protein Engineering 7:805 (1994); Roguska, et al., Proc Natl Acad Sci USA 91:969 (1994)) і перетасування ланцюгів (патент США № 5565332). Як правило, гуманізоване антитіло містить один або декілька амінокислотних залишків з нелюдського джерела. Такі нелюдські амінокислотні залишки, що зазвичай беруть з "імпортного" варіабельного домену, найчастіше називають "імпортними" залишками. 9 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Гуманізацію в основному можна проводити за способом Winter і співавторів (Jones, et al., Nature 321:522 (1986); Riechmann, et al., Nature 332:323 (1988); Verhoeyen, et al., Science 239:1534 (1988)) шляхом використання нелюдських CDR або послідовностей CDR замість відповідних послідовностей людського антитіла. Відповідно, такі "гуманізовані" антитіла являють собою химерні антитіла (патент США № 4816567), в яких фрагмент, замінений на відповідну нелюдську послідовність, суттєво менше інтактного людського варіабельного домену. Фактично гуманізовані антитіла звичайно являють собою людські антитіла, в яких декілька залишків CDR і, необов'язково, декілька залишків FR замінені на аналогічні ділянки антитіл гризунів. Крім того, важливо, щоб гуманізовані антитіла зберігали підвищену спорідненість до антигена та інші корисні біологічні властивості. Для досягнення даної мети, відповідно до переважного способу, гуманізовані антитіла одержують після аналізу вихідної послідовності і різних концептуальних гуманізованих продуктів з використанням тривимірних моделей вихідних і гуманізованих послідовностей. Тривимірні моделі імуноглобулінів є широко доступними і відомі фахівцям у даній галузі. Існують комп'ютерні програми, що ілюструють і відображають можливі тривимірні конформаційні структури вибраних імуноглобулінових послідовностей-кандидатів. Аналіз одержаних зображень дозволяє зробити висновок про можливу роль деяких залишків у функціонуванні імуноглобулінової послідовності-кандидаті, тобто, ідентифікувати залишки, що впливають на здатність імуноглобуліну-кандидата зв'язувати антиген. За допомогою даного способу можна вибрати і скомбінувати залишки FR реципієнтної і імпортної послідовностей таким чином, щоб максимізувати бажані характеристики антитіла, такі як підвищена спорідненість до цільового антигена (антигенів), хоча безпосередній і найдужчий вплив на зв'язування антигена роблять залишки CDR. При одержанні гуманізованих антитіл вибір людських варіабельних доменів, як легкого, так і важкого ланцюга, має велике значення для зниження антигенності. Відповідно до так що називається методу "найкращої відповідності" послідовність варіабельного домену нелюдського антитіла піддають скринінгу проти цілої бібліотеки відомих людських послідовностей варіабельних доменів. Потім людську послідовність, що найкраще відповідає послідовності нелюдського вихідного антитіла приймають як людський FR гуманізованого антитіла (Sims, et al., J Immunol 151:2296 (1993); Chothia, et al., J Mol Biol 196:901 (1987)). В іншому способі використовують конкретну каркасну ділянку, одержану з консенсусної послідовності всіх людських антитіл конкретної підгрупи легких або важких ланцюгів. Ту саму каркасну ділянку можна використовувати для декількох різних гуманізованих антитіл (Carter, et al., Proc Natl Acad Sci USA 89:4285 (1992); Presta, et al., J Immunol 151:2623 (1993)). Для терапевтичного лікування пацієнтів-людей бажано використовувати цілком людські антитіла. Людські антитіла можна одержати за допомогою ряду відомих у даній галузі способів, що містять у собі описані вище методи фагових дисплеїв з використанням бібліотек антитіл, одержаних з послідовностей людських імуноглобулінів. Див. також патенти США №№ 4444887 і 4716111; і публікації PCT WO 98/46645, WO 98/50433, WO 98/24893, WO 98/16654, WO 96/34096, WO 96/33735 і WO 91/10741; які включені в даний опис у вигляді посилання у всій повноті. Для одержання людських моноклональних антитіл також можна використовувати методи Cole, et al. і Boerder, et al. (Cole, et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Riss (1985); and Boerner, et al., J Immunol 147:86 (1991)). Людські антитіла також можна одержати з використанням трансгенних мишей, що не здатні експресувати функціональні ендогенні імуноглобуліни, але можуть експресувати гени людських імуноглобулінів. Наприклад, комплекс генів важкого і легкого ланцюгів імуноглобуліну можна ввести довільним чином або шляхом гомологічної рекомбінації в ембріональні стовбурові клітини миші. Альтернативно, в ембріональні стовбурові клітини миші крім генів людських важких і легких ланцюгів можна ввести гени людської варіабельної ділянки, константної ділянки і D-ділянки. Гени мишачих важких і легких ланцюгів імуноглобуліну можна привести в нефункціональний стан роздільно або одночасно з введенням локусу людського імуноглобуліну шляхом гомологічної рекомбінації. Зокрема, гомологічна делеція ділянки JH запобігає продукції гомологічних антитіл. Модифіковані ембріональні стовбурові клітини вирощують і вводять методом мікроін'єкції в бластоцисти, одержуючи химерних мишей. Потім химерних мишей схрещують і одержують гомозиготне потомство, що експресує людські антитіла. Див., наприклад, Jakobovitis, et al., Proc Natl Acad Sci USA 90:2551 (1993); Jakobovitis, et al., Nature 362:255 (1993); Bruggermann, et al., Year in Immunol 7:33 (1993); Duchosal, et al., Nature 355:258 (1992)). Трансгенних мишей імунізують звичайним способом вибраним антигеном, наприклад, повнорозмірним поліпептидом даного винаходу або його фрагментом. Моноклональні антитіла, направлені проти антигена, можна одержати з імунізованих трансгенних мишей за допомогою традиційної гібридомної технології. Трансгени людських імуноглобулінів, що містяться в 10 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 трансгенних мишах, піддаються реаранжуванню в процесі диференціації B-клітин і потім переключенню класу і соматичній мутації. Таким чином, за допомогою даного методу можна одержати антитіла IgG, IgA, IgM і IgE, придатні для терапевтичного застосування. Огляд за вказаною технологією одержання людських антитіл можна знайти в Lonberg, et al., Int Rev Immunol 13:65-93 (1995). Докладне обговорення даної технології одержання людських антитіл і людських моноклональних антитіл, а також способи одержання таких антитіл можна знайти, наприклад, у публікаціях PCT WO 98/24893; WO 92/01047; WO 96/34096; WO 96/33735; Європейському патенті № 0598877; патентах США №№ 5413923; 5625126; 5633425; 5569825; 5661016; 5545806; 5814318; 5885793; 5916771; і 5939598, що включені в даний опис в усій повноті. Крім того, компанії, такі як Abgenix, Inc. (Freemont, Calif.), Genpharm (San Jose, Calif.) і Medarex, Inc. (Princeton, N.J.) можуть виготовляти на замовлення людські антитіла проти вибраного антигена за допомогою технології, подібної описаній вище. Крім того, людські MAb можна одержати шляхом імунізації мишей, яким були трансплантовані людські лейкоцити периферичної крові, спленоцити або кістковий мозок (наприклад, методи XTL Trioma). Цілком людські антитіла, що розпізнають обраний епітоп, можна одержати за допомогою методу, що називається "керована селекція". У даному способі вибране нелюдське моноклональне антитіло, наприклад, мишаче антитіло, використовують для керування селекцією цілком людського антитіла, що розпізнає такий самий епітоп (Jespers, et al., Bio/technology 12:899 (1988)). Крім того, антитіла проти поліпептидів даного винаходу, у свою чергу, можна використовувати для одержання антиідіотипічних антитіл, що "імітують" поліпептиди даного винаходу, за допомогою способів, добре відомих фахівцям у даній галузі (див., наприклад, Greenspan, et al., FASEB J 7:437 (1989); Nissinoff, J Immunol 147:2429 (1991)). Наприклад, антитіла, що зв'язуються з поліпептидом і конкурентно інгібують мультимеризацію поліпептиду і/або зв'язування поліпептиду даного винаходу з лігандом, можна використовувати для одержання антиідіотипів, що "імітують" мультимеризацію поліпептиду і/або зв'язувальний домен, і, як наслідок, зв'язуються з поліпептидом і/або його лігандом і нейтралізують їх. Такі нейтралізуючі антиідіотипи або фрагменти Fab таких антиідіотипів можна використовувати в терапевтичних режимах для нейтралізації ліганду поліпептиду. Наприклад, такі антиідіотипічні антитіла можна використовувати для зв'язування поліпептиду даного винаходу, і/або для зв'язування його лігандів/рецепторів, блокуючи в такий спосіб його біологічну активність. Антитіла даного винаходу можуть являти собою біспецифічні антитіла. Біспецифічні антитіла являють собою моноклональні, переважно людські або гуманізовані антитіла, що здатні специфічно зв'язуватися щонайменше із двома різними антигенами. У даному винаході одна зі специфічностей зв'язування може бути направлена на Notch3, а інша - на інший антиген, що переважно представляє собою білок клітинної поверхні, рецептор, субодиницю рецептора, тканиноспецифічний антиген, вірусний білок, білок вірусної оболонки, бактеріальний білок або бактеріальний поверхневий білок і ін. Способи одержання біспецифічних антитіл добре відомі. Зазвичай рекомбінантний спосіб одержання біспецифічних антитіл заснований на спільній експресії двох пар важких/легких ланцюгів імуноглобуліну, де два важкі ланцюги мають різну специфічність (Milstein, et al., Nature 305:537 (1983)). Внаслідок випадкового реаранжування важких і легких ланцюгів імуноглобуліну, вказані гібридоми (квадрогібридоми) можуть продукувати суміш з десяти різних молекул антитіл, з яких тільки одна має правильну біспецифічну структуру. Виділення правильної молекули зазвичай проводять методом афінної хроматографії. Подібні способи описані в WO 93/08829 і Traunecker, et al., EMBO J 10:3655 (1991). Варіабельні домени антитіл з бажаною специфічністю зв'язування (ділянки, за якими антитіло з'єднується з антигеном) можна гібридизувати з послідовностями константних доменів імуноглобулінів. Для гібридизації переважно використовують константний домен важкого ланцюга імуноглобуліну, що містить щонайменше частину шарнірної ділянки, ділянки CH2 і CH3. Він може містити перший константний домен важкого ланцюга (CH1), що включає в себе ділянку, необхідну для зв'язування легкого ланцюга, що присутній щонайменше в одному з гібридів. ДНК, що кодують гібриди важкого ланцюга імуноглобуліну і, при бажанні, легкий ланцюг імуноглобуліну, вставляють в окремі вектори експресії, що спільно трансфікують в організм придатного хазяїна. Більш докладний опис способів одержання біспецифічних антитіл можна знайти, наприклад, у Suresh, et al., Meth In Enzym 121:210 (1986). Гетерокон'юговані антитіла також входять в обсяг даного винаходу. Гетерокон'юговані антитіла складаються з двох ковалентно зв'язаних антитіл. Такі антитіла, наприклад, можна використовувати для націлювання клітин імунної системи на небажані клітини (патент США № 4676980). Такі антитіла можна одержати in vitro за допомогою способів, відомих в галузі 11 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 синтетичної білкової хімії, наприклад, з використанням поперечно-зшивальних реагентів. Наприклад, імунотоксини можна одержати за допомогою дисульфідної обмінної реакції або шляхом утворення тіоефірного зв'язку. Приклади реагентів, придатні для таких реакцій, містять у собі імінотіолат і метил-4-меркаптобутилімідат, а також реагенти, розкриті, наприклад, у патенті США № 4676980. Також можна одержати однодоменні антитіла проти Notch3. Способи одержання таких антитіл описані в WO9425591 на прикладі антитіла, одержаного з важкого ланцюга IgG Camelidae, крім того, у US20030130496 описане виділення однодоменного повнорозмірного людського антитіла з фагових бібліотек. Крім того, можна одержати одноланцюжкові пептидні зв'язувальні молекули, що містять з'єднані ділянки Fv важкого і легкого ланцюга. Одноланцюжкові антитіла ("scFv") і спосіб їх одержання описані в патенті США № 4946778. Альтернативно, за допомогою подібних методів можна сконструювати і експресувати Fab. Усі повністю або частково людські антитіла є менш імуногенними, ніж цілком мишачі MAb, причому фрагменти й одноланцюжкові антитіла також є менш імуногенними. Антитіла або фрагменти антитіл можна виділити з фагових бібліотек антитіл, одержаних за допомогою способів, описаних у McCafferty, et al., Nature 348:552 (1990). Clarkson, et al., Nature 352:624 (1991) і Marks, et al., J Mol Biol 222:581 (1991), описують відповідно виділення мишачих і людських антитіл з використанням фагових бібліотек. Наступні публікації описують одержання високоафінних (нм діапазон) людських антитіл шляхом перетасування ланцюгів (Marks, et al., Bio/Technology 10:779 (1992)), а також комбінаторну інфекцію і рекомбінацію in vivo як спосіб одержання дуже великих фагових бібліотек (Waterhouse, et al., Nuc Acids Res 21:2265 (1993)). Вказані методи є конкурентноздатними альтернативами традиційним гібридомним способам одержання моноклональних антитіл. Крім того, можна модифікувати ДНК, наприклад, шляхом введення послідовностей, що кодують людські константні домени важкого і легкого ланцюгів, замість гомологічних мишачих послідовностей (патент США № 4816567; Morrison, et al., Proc Natl Acad Sci USA 81:6851 (1984)). Іншою альтернативою при одержанні гібридом є застосування в більшому ступені електричної гібридизації, ніж хімічної гібридизації. Даний метод добре розроблений. Замість гібридизації трансформацію B-клітин з одержанням іморталізованих клітин можна провести, наприклад, з використанням вірусу Епштейна-Барр або трансформуючого гена. Див., наприклад, "Continuously Proliferating Human Cell Lines Synthesizing Antibody of Predetermined Specificity", Zurawaki, et al., in Monoclonal Antibodies, ed. by Kennett, et al, Plenum Press, pp.19-33. (1980)). MAb проти Notch3 можна одержати шляхом імунізації гризунів (наприклад, мишей, щурів, хом'яків і морських свинок) білком Notch3, гібридним білком або його фрагментами, експресованими в еукаріотичних або прокаріотичних системах. Для імунізації можна використовувати і інших тварин, наприклад, відмінних від людини приматів, трансгенних мишей, експресуючих імуноглобуліни, і мишей з важким комбінованим імунодефіцитом (SCID), яким були трансплантовані людські B-лімфоцити. Гібридоми можна одержати за допомогою традиційних способів шляхом гібридизації B-лімфоцитів з мієломними клітинами імунізованих тварин (наприклад, Sp2/0 і NSO), як описано раніше (Kohler, et al., Nature 256:495 (1975)). Крім того, антитіла проти Notch3 можна одержати після скринінга рекомбінантних одноланцюжкових Fv або бібліотек Fab людських В-лімфоцитів з використанням фагових дисплеїв. Специфічність MAb у відношенні Notch3 можна проаналізувати за допомогою методів ELISA, вестернімуноблоттинга, або інших імунохімічних методів. Інгібувальну дію антитіла на активацію комплементу можна оцінити за допомогою гемолітичних аналізів класичного шляху комплементу з використанням сенсибілізованих червоних кров'яних клітин курей або овець. Гібридоми з позитивних ямок клонують методом серійних розведень. Антитіла очищають і аналізують їхню специфічність до людського Notch3 за допомогою описаних вище методів. Ідентифікація антитіл проти NOTCH-3 Даний винахід пропонує агоністичні моноклональні антитіла, що активують Notch3опосередкований сигнальний шлях незалежно від ліганду. Зокрема, антитіла даного винаходу зв'язуються з Notch3 і активують його. Антитіла даного винаходу містять у собі антитіло, що позначається 256A-13. Даний винахід також містить у собі антитіла, що зв'язуються з таким же епітопом, як і 256A-13. Антитіла-кандидати проти Notch3 аналізують за допомогою методів імуноферментного твердофазного аналізу (ELISA), вестерн-імуноблоттинга, або інших імунохімічних методів. Аналізи, використовувані для характеристики окремих антитіл, описані в прикладах. Антитіла даного винаходу містять у собі, без обмеження, поліклональні, моноклональні, моновалентні, біспецифічні, гетерокон'юговані, поліспецифічні, людські, гуманізовані або 12 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 химерні антитіла, одноланцюжкові антитіла, однодоменні антитіла, фрагменти Fab, фрагменти F(ab'), фрагменти, одержувані з використанням експресійної бібліотеки Fab, антиідіотипічні (анти-Id) антитіла (які включають у себе, наприклад, анти-Id антитіла проти антитіл даного винаходу), а також епітоп-зв'язувальні фрагменти вказаних вище антитіл. Антитіла можуть являти собою людські антиген-зв'язувальніфрагменти антитіл даного винаходу, що містять у собі, без обмеження, Fab, Fab' і F(ab') 2, Fd, одноланцюжкові Fv (scFv), одноланцюжкові антитіла, дисульфід-зв'язані Fv (sdFv) і однодоменні антитіла, що містять або VL, або VH домен. Антиген-зв'язувальні фрагменти антитіл, що включають в себе одноланцюжкові антитіла, можуть містити тільки варіабельну ділянку (ділянки), або варіабельну ділянку в поєднанні з цілими шарнірними ділянками, доменами CH1, CH2 і CH3 або їх частинами. Даний винахід також охоплює антиген-зв'язувальні фрагменти, що містять будь-які поєдання варіабельної ділянки (ділянок) із шарнірною ділянкою, доменами CH1, CH2 і CH3. Антитіла даного винаходу можуть бути одержані з будь-якого тваринного джерела, що включає в себе птахів і ссавців. Переважно, антитіла можуть бути одержані від людини, відмінного від людини примата, гризуна (такого як миша і щур), осла, вівці, кролика, кози, морської свинки, верблюда, коня або курки. У даному описі "людські" антитіла" містять у собі антитіла, що мають амінокислотну послідовність людського імуноглобуліну, у тому числі антитіла, виділені з бібліотек людських імуноглобулінів або із тварин, трансгенних по одному або декільком людським імуноглобулінам, і не експресуючих ендогенні імуноглобуліни, як описано нижче і, наприклад, у патенті США № 5939598, Kucherlapati, et al. Антитіла даного винаходу можуть бути моноспецифічними, біспецифічними, триспецифічними або поліспецифічними. Поліспецифічні антитіла можуть бути специфічними до різних епітопів Notch3, або вони можуть бути специфічними до Notch3 і до гетерологічного епітопа, такого як гетерологічний поліпептид або твердофазний носій. Див., наприклад, публікації PCT WO 93/17715; WO 92/08802; WO 91/00360; WO 92/05793; Tutt, et al., J Immunol 147:60 (1991); патенти США №№ 4474893; 4714681; 4925648; 5573920; 5601819; Kostelny, et al., J Immunol 148:1547 (1992). Антитіла даного винаходу можна описати або охарактеризувати з погляду епітопа (епітопів) або фрагмента (фрагментів) Notch3, які вони розпізнають і специфічно зв'язують. Епітоп (епітопи) або поліпептидний фрагмент (фрагменти) можна охарактеризувати, як вказано в даному документі, наприклад, по N-кінцевих і C-кінцевих положеннях, по кількості суміжних амінокислотних залишків, або їх можна знайти в таблицях і кресленнях. Антитіла даного винаходу також можна описати або охарактеризувати з погляду їхньої перехресної реактивності. В обсяг даного винаходу також входять антитіла, що зв'язують поліпептиди Notch3 і є щонайменше на 95 % щонайменше на 90 % щонайменше на 85 % щонайменше на 80 % щонайменше на 75 % щонайменше на 70 % щонайменше на 65 % щонайменше на 60 % щонайменше на 55 %, і щонайменше на 50 % ідентичними Notch3 (ідентичність розраховується за допомогою відомих у даній галузі й описаних у даному документі способів). Антитіла проти Notch3 також можуть зв'язуватися з іншими білками, такими як антитіла проти Notch3 з видів, відмінних від виду, проти якого направлене використовуване -7 антитіло проти Notch3, при цьому KD може бути менше, ніж приблизно 10 M, менше, ніж -6 -5 приблизно 10 M, або менше, ніж приблизно 10 M. В окремих втіленнях антитіла даного винаходу можуть перехресно взаємодіяти з мавпячими гомологами людського Notch3 і їх відповідними епітопами. В одному втіленні вищевказана здатність до перехресної взаємодії стосується одного конкретного антигенного або імуногенного поліпептиду або до поєднання (поєднань) конкретних антигенних і/або імуногенних поліпептидів, розкритих у даному описі. Даний винахід також охоплює антитіла, що зв'язують поліпептиди, які кодуються полінуклеотидами, що у жорстких умовах гібридизуються з полінуклеотидом, що кодує Notch3. Антитіла даного винаходу також можуть бути описані або охарактеризовані з погляду спорідненості зв'язування відносно поліпептиду даного винаходу. Переважна спорідненість зв'язування характеризується рівноважною константою дисоціації або KD, що знаходиться в -8 -15 -8 -12 -8 -10 -10 -12 інтервалі від 10 до 10 M, від 10 до 10 M, від 10 до 10 M, або від 10 до 10 M. Даний винахід також пропонує антитіла, що конкурентно інгібують зв'язування антитіл з епітопом даного винаходу, що визначають за допомогою будь-якого відомого у даній галузі способу визначення конкурентного зв'язування, наприклад, за допомогою описаних тут імунологічних аналізів. У переважних втіленнях антитіло конкурентно інгібує зв'язування з епітопом щонайменше на 95 % щонайменше на 90 % щонайменше на 85 % щонайменше на 80 % щонайменше на 75 % щонайменше на 70 % щонайменше на 60 %, або щонайменше на 50 %. 13 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Вектори і клітини-хазяїни В іншому аспекті даний винахід пропонує виділені нуклеотидні послідовності, що кодують варіант антитіла, як описано в даному документі, векторні конструкції, що містять нуклеотидну послідовність, що кодує антитіла даного винаходу, клітини-хазяїни, що містять такий вектор, а також рекомбінантні способи одержання антитіл. Щоб одержати варіант антитіла рекомбінантним способом, що кодує його нуклеїнову кислоту, виділяють і вставляють у здатний до реплікації вектор для наступних клонування (ампліфікації ДНК) або експресії. ДНК, що кодує варіант антитіла, можна легко виділити і секвенувати за допомогою традиційних способів (наприклад, шляхом використання олігонуклеотидних зондів, здатних специфічно зв'язуватися з генами, що кодують важкі і легкі ланцюги варіанта антитіла). Для одержання клітинних ліній, що експресують антитіла даного винаходу, можна використовувати стандартні методи клонування і трансформації. Вектори Існує багато векторів. Як правило, вектор містить, без обмеження, одну або декілька з послідовностей, що включають у себе: сигнальну послідовність, точку початку реплікації, один або декілька маркерних генів, енхансер, промотор і послідовність термінації транскрипції. Рекомбінантні вектори експресії, що містять нуклеотидну послідовність, яка кодує антитіла даного винаходу, можна одержати за допомогою добре відомих методів. Вектори експресії містять нуклеотидну послідовність, функціонально зв'язану з придатними нуклеотидними послідовностями, що регулюють транскрипцію або трансляцію, наприклад, одержаними з генів ссавців, мікробів, вірусів або комах. Приклади регуляторних послідовностей містять у собі промотори транскрипції, оператори, енхансери, ділянки зв'язування рибосомальної мРНК і/або інші придатні послідовності, що контролюють ініціацію і термінацію транскрипції і трансляції. Нуклеотидні послідовності є "функціонально зв'язаними", якщо регуляторна послідовність знаходиться у функціональному зв'язку з послідовністю, що кодує відповідний поліпептид. Так, нуклеотидна послідовність промотору функціонально зв'язана, наприклад, з послідовністю, що кодує важкий ланцюг антитіла, якщо нуклеотидна послідовність промотору регулює транскрипцію відповідної нуклеотидної послідовності. Крім того, вектори експресії можуть містити послідовності, що кодують відповідні сигнальні пептиди, що в природі не пов'язані з послідовностями, що кодують важкий і/або легкий ланцюг антитіла. Наприклад, нуклеотидна послідовність, що кодує сигнальний пептид (секреторну лідерну послідовність), може бути зв'язана в рамці зчитування з послідовністю, що кодує поліпептид, так, щоб антитіло секретувалося у периплазматичному просторі або у середовище. Сигнальний пептид, що є функціональним у передбачуваних клітинах-хазяїнах, збільшує позаклітинну секрецію відповідного антитіла. Сигнальний пептид може відщеплюватися від поліпептиду при секреції антитіла з клітини. Приклади таких секреторних сигналів добре відомі і містять у собі, наприклад, сигнали, описані в патентах США №№ 5698435; 5698417; і 6204023. Вектор може являти собою плазмідний вектор, одноланцюжковий або дволанцюжковий фаговий вектор, або одноланцюжковий або дволанцюжковий РНК- або ДНК-вектор. Такі вектори можна вводити в клітини у вигляді полінуклеотидів за допомогою добре відомих способів введення ДНК і РНК у клітини. Фагові і вірусні вектори також можна вводити в клітини в упакованому або інкапсульованому вигляді за допомогою добре відомих способів інфікування і трансдукції. Вірусні вектори можуть бути компетентними або дефективними по реплікації. В останньому випадку розмноження вірусу, як правило, відбувається тільки в доповнюючих клітинах-хазяїнах. Крім того, для одержання білка можна використовувати безклітинні системи трансляції з використанням РНК, одержаної з конструкцій ДНК даного винаходу. Такі вектори можуть містити в собі нуклеотидну послідовність, що кодує константну ділянку молекули антитіла (див., наприклад, публікації PCT WO 86/05807 і WO 89/01036; і патент США № 5122464), у такому векторі можна клонувати варіабельний домен антитіла з наступною експресією важкого або легкого ланцюга. Клітини-хазяїни Антитіла даного винаходу можна експресувати у будь-якій придатній клітині-хазяїні. У даному винаході як клітини-хазяїни можна використовувати прокаріотичні, дріжджові або вищі еукаритичні клітини, приклади яких містять у собі, без обмеження, мікроорганізми, такі як бактерії (наприклад, E. coli, B. subtilis), трансформовані ДНК рекомбінантного бактеріофага, векторами експресії на основі плазмідної або космідної ДНК, що містять послідовності, які кодують антитіло; дріжджі (наприклад, Saccharomyces, Pichia), трансформовані дріжджовими рекомбінантними векторами експресії, що містять послідовності, які кодують антитіло; системи клітин комах, інфікованих рекомбінантними вірусними векторами експресії (наприклад, Baculovirus), що містять послідовності, які кодують антитіло; системи рослинних клітин, 14 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 інфікованих рекомбінантними вірусними векторами експресії (наприклад, вірусом мозаїки кольоровий капусти, CaMV; вірусом тютюнової мозаїки, TMV), або трансформованих рекомбінантними плазмідними векторами експресії (наприклад, плазмідой Ti), які містять послідовності, що кодують антитіло; або системи клітин ссавців (наприклад, клітини COS, CHO, BHK, 293, 3T3), що несуть рекомбінантні експресійні конструкції, що містять промотори, одержані з генома клітин ссавців (наприклад, промотор металотіонеїну) або з вірусів ссавців (наприклад, пізній промотор аденовірусу; промотор вірусу коров'ячої віспи 7.5K). Прокаріоти, придатні для застосування як клітини-хазяїв даного винаходу, містять у собі грам-негативні або грам-позитивні організми, такі як E. coli, B. subtilis, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, Serratia і Shigella, а також Bacilli, Pseudomonas і Streptomyces. Переважним для клонування хазяїном E. coli є E. coli 294 (ATCC 31446), хоча можна використовувати та інші штами, такі як E. coli B, E. coli X1776 (ATCC 31537) і E. coli W3110 (ATCC 27325). Наведені приклади є скоріше ілюстративними, ніж обмежуючими. Вектори експресії, використовувані в прокаріотичних клітинах-хазяїнах, зазвичай містять один або декілька маркерних генів фенотипічної селекції. Маркерний ген фенотипічної селекції являє собою, наприклад, ген, що кодує білок, який відповідає за стійкість до антибіотиків, або за задоволення автотрофної вимоги. Приклади векторів експресії, які можна використовувати в прокаріотичних клітинах-хазяїнах, містять у собі вектори, одержані з комерційно доступних плазмід, таких як pKK223-3 (Pharmacia Fine Chemicals, Uppsala, Sweden), pGEM1 (Promega Biotec, Madison, Wisconsin., USA) і серії векторів pET (Novagen, Madison, Wisconsin, USA) і pRSET (Invitrogen, Carlsbad, CA) (Studier, J Mol Biol 219:37 (1991); Schoepfer, Gene 124:83 (1993)). Промоторні послідовності, зазвичай використовувані у векторах експресії рекомбінантних прокаріотичних клітин-хазяїнів, містять у собі T7, (Rosenberg, et al., Gene 56:125 (1987)), (-лактамазу (пенициллиназу), систему промотору лактози (Chang, et al., Nature 275:615 (1978); Goeddel, et al., Nature 281:544 (1979)), систему промоторів триптофану (trp) (Goeddel, et al., Nucl Acids Res 8:4057 (1980)) і промотор tac (Sambrook, et al., Molecular Cloning, A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory (1990)). Дріжджі або гіфоміцети, придатні для застосування в даному винаході, містять у собі організми з родів Saccharomyces, Pichia, Actinomycetes, Kluyveromyces, Schizosaccharomyces, Candida, Trichoderma, Neurospora, а також такі гіфоміцети, як Neurospora, Penicillium, Tolypocladium і Aspergillus. Дріжджові вектори зазвичай містять послідовність початку реплікації з дріжджової плазміди 2μ, послідовність автономної реплікації (ARS), промоторну ділянку, послідовності поліаденілування, послідовності термінації транскрипції і маркерний ген селекції. Промоторні послідовності, що підходять для дріжджових векторів, містять у собі, серед інших, промотори металотіонеїну, 3-фосфогліцераткінази (Hitzeman, et al., J Biol Chem 255:2073 (1980)) або інших гліколітичних ферментів (Holland, et al., Biochem 17:4900 (1978)), таких як енолаза, гліцеральдегід-3-фосфат дегідрогеназа, гексокіназа, піруватдекарбоксилаза, фосфофруктокіназа, глюкозо-6-фосфатізомераза, 3-фосфогліцератмутаза, піруваткіназа, триозофосфатізомераза, фосфоглюкозізомераза і глюкокіназа. Інші вектори і промотори, що підходять для проведення експресії в дріжджах, також описані в Fleer, et al., Gene 107:285 (1991). Інші промотори і вектор, що підходять для дріжджів і трансформації дріжджів, добре відомі в даній галузі. Способи трансформації дріжджів широко відомі. Приклад такого способу описаний у Hinnen, et al., Proc Natl Acad Sci 75:1929 (1978). Спосіб Hinnen заснований на селекції трансформантів Trp+ у селективному середовищі. Для експресії рекомбінантних антитіл також можна використовувати системи культур клітинхазяїнів ссавців або комах. В принципі, можна використовувати будь-які культури вищих еукаріотичних клітин, як хребетних, так і безхребетних тварин. Приклади клітин безхребетних містять у собі клітини рослин і комах (Luckow, et al., Bio/Technology 6:47 (1988); Miller, et al., Genetics Engineering, Setlow, et al., eds. Vol. 8, pp. 277-9, Plenam Publishing (1986); Mseda, et al., Nature 315:592 (1985)). Наприклад, для одержання гетерологічних систем можна використовувати системи Baculovirus. У системах комах для експресії чужорідних генів як вектор експресії можна використовувати вірус ядерного поліедрозу совки каліфорнійської люцернової (AcNPV). Вірус вирощують у клітинах Spodoptera frugiperda. Послідовність, що кодує антитіло, можна клонувати окремо в неесенційних ділянках (наприклад, гена поліедрину) вірусу під контролем промотору AcNPV (наприклад, промотору поліедрину). Інші ідентифіковані хазяїни містять у собі Aedes, Drosophila melanogaster і Bombyx mori. Ряд вірусних штамів для трансфекції є загальнодоступним, наприклад, варіант L-1 AcNPV і штам Bm-5 Bombyx mori NPV, які можна використовувати як віруси відповідно до даного винаходу, особливо, для трансфекції клітин Spodoptera frugiperda. Крім того, як клітини-хазяїни також можна використовувати культури рослинних клітин бавовни, кукурудзи, картоплі, соєвих бобів, петунії, томатів і тютюну. 15 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Застосування клітин хребетних, а також розмноження клітин хребетних у культурі (тканинній культурі) є рутинними процедурами. Див. Tissue Culture, Kruse, et al., eds., Academic Press (1973). Прикладами придатних ліній клітин-хазяїнів ссавців є клітини нирки мавпи; клітини нирки ембріона людини; клітини нирки дитинчати хом'яка; клітини яєчника китайського хом'ячка/-DHFR (CHO, Urlaub, et al., Proc Natl Acad Sci USA 77:4216 (1980)); мишачі клітини Сертолі; клітини карциноми шийки матки людини (HELA); клітини нирки собаки; клітини легені людини; клітини печінки людини; клітини пухлини молочної залози миші; і клітини NSO. Клітини-хазяїни трансформують вищевказаними векторами, щоб забезпечити продукцію антитіла, і культивують у традиційному поживному середовищі, модифікованому, залежно від обставин, так, щоб індукувати промотори, послідовності, що контролюють транскрипцію і трансляцію, здійснювати селекцію трансформантів або ампліфікувати гени, що кодують цільові послідовності. Зазвичай використовують промоторні і енхансерні послідовності, одержані з вірусу поліоми, аденовірусу 2, вірусу мавп 40 (SV40) і людського цитомегаловірусу (CMV). Послідовності ДНК із генома вірусу SV40 можна використовувати для одержання інших генних елементів, використовуваних для експресії послідовності структурного гена в клітинах-хазяїнах ссавців, що містять у собі точку початку реплікації SV40, ранній і пізній промотор, енхансер, точку сплайсинга і ділянки поліаденілування. Особливо корисними є вірусні ранній і пізній промотори, оскільки і той і інший можна легко одержати з вірусного генома у вигляді фрагмента, що також може містити вірусну точку початку реплікації. Типові вектори експресії для застосування в клітинах-хазяїнах ссавців є комерційно доступними. Клітини-хазяїни, використовувані для одержання варіанта антитіла даного винаходу, можна культивувати в різних середовищах. Для культивування клітин-хазяїнів можна використовувати комерційно доступні середовища, такі як середовище Хема F10 (Sigma, St Louis, MO), мінімальне есенційне середовище (MEM, Sigma, St Louis, MO), RPMI-1640 (Sigma, St Louis, MO) і модифіковане за Дульбекко середовище Голка (DMEM, Sigma, St Louis, MO). Крім того, як культуральне середовище для клітин-хазяїнів можна використовувати кожне із середовищ, описаних у Ham, et al., Meth Enzymol 58:44 (1979), Barnes, et al., Anal Biochem 102:255 (1980), і в патентах США №№ 4767704; 4657866; 4560655; 5122469; 5712163; або 6048728. За необхідності кожне з вказаних середовищ може бути доповнене гормонами і/або іншими факторами росту (такими як інсулін, трансферин або епідермальний фактор росту), солями (такими як X-хлориди, де X означає натрій, кальцій, магній; і фосфати), буферами (такими як HEPES), нуклеотидами (такими як аденозин і тимідин), антибіотиками (такими як лікарський TM засіб GENTAMYCIN ), мікроелементами (неорганічними сполуками, кінцеві концентрації яких зазвичай знаходяться в мікромолярному діапазоні) і глюкозою або еквівалентним джерелом енергії. Також можуть бути присутніми у відповідних концентраціях будь-які інші необхідні добавки, відомі фахівцям у даній галузі. Експресію проводять в умовах культивування, що включають у себе температуру, pН і т.п., що раніше використовувалися для вибраних клітинхазяїнів і відомі рядовим фахівцям у даній галузі. Полінуклеотиди, що кодують антитіла Даний винахід також пропонує полінуклеотиди або нуклеїнові кислоти, наприклад, ДНК, що містять нуклеотидну послідовність, що кодує антитіло даного винаходу, а також їхні фрагменти. Приклади полінуклеотидів містять у собі полінуклеотиди, що кодують ланцюги антитіл, які містять одну або декілька з описаних у даному документі амінокислотних послідовностей. Даний винахід також охоплює полінуклеотиди, що гібридизуються в жорстких умовах або в умовах зниженої жорсткості з полінуклеотидами, що кодують антитіло даного винаходу. Одержати полінуклеотиди і визначити їх нуклеотидну послідовність можна за допомогою будь-якого відомого у даній галузі способу. Наприклад, якщо нуклеотидна послідовність антитіла відома, полінуклеотид, що кодує антитіло, можна зібрати з хімічно синтезованих олігонуклеотидів (наприклад, як описано в Kutmeier, et al., Bio/Techniques 17:242 (1994)), коротко говорячи, даний спосіб містить у собі синтез олігонуклеотидів, які перекриваються, що містить фрагменти послідовності, яка кодує антитіло, відпалювання і лігування вказаних олігонуклеотидів, і потім ампліфікацію лігованих олігонуклеотидів методом ПЛР. Альтернативно, полінуклеотид, що кодує антитіло, можна одержати з нуклеїнової кислоти, виділеної з придатного джерела. Якщо клон, що містить нуклеїнову кислоту, що кодує конкретне антитіло, не доступний, але послідовність молекули антитіла відома, нуклеїнову кислоту, що кодує імуноглобулін, можна синтезувати або хімічно одержати з придатного джерела (такого як бібліотека кДНК антитіла або бібліотека кДНК, одержана з, або нуклеїнова кислота, переважно + полі A РНК, виділена з будь-яких тканин або клітин, експресуючих антитіло, таких як клітини гібридоми, вибрані для експресії антитіла даного винаходу) шляхом ампліфікації методом ПЛР із використанням синтетичних праймерів, здатних гібридизуватися з 3'- і 5'-кінцями 16 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовності, або шляхом клонування з використанням олігонуклеотидного зонда, специфічного до конкретної послідовності, щоб ідентифікувати, наприклад, клон кДНК із бібліотеки кДНК, що кодує антитіло. Потім ампліфіковані нуклеїнові кислоти, одержані методом ПЛР, можна клонувати у здатних до реплікації векторах клонування за допомогою будь-яких відомих у даній галузі методів. Після визначення нуклеотидної послідовності і відповідної амінокислотної послідовності антитіла нуклеотидну послідовність антитіла можна модифікувати за допомогою добре відомих у даній галузі способів модифікації нуклеотидної послідовності, таких як, наприклад, методи рекомбінантних ДНК, направлений мутагенез, ПЛР та ін. (див., наприклад, методи, описані в Sambrook, et al., Molecular Cloning, A Laboratory Manual, 2nd ed, Cold Spring Harbor Laboratory (1990); Ausubel, et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons (1998), що включені в даний опис у вигляді посилання у всій повноті), щоб одержати антитіла з різними амінокислотними послідовностями, наприклад, щоб здійснити амінокислотні заміни, делеції і/або інсерції. У конкретному втіленні амінокислотна послідовність варіабельних доменів важкого і/або легкого ланцюга можна аналізувати, щоб ідентифікувати послідовності CDR за допомогою добре відомих методів, наприклад, шляхом порівняння з відомими амінокислотними послідовностями інших варіабельних ділянок важкого і легкого ланцюга, щоб визначити гіперваріабельні ділянки послідовності. Використовуючи рутинні методи рекомбінантних ДНК, один або декілька CDR можна вставити в каркасні ділянки, наприклад, у людські каркасні ділянки, щоб гуманізувати нелюдське антитіло, як вказано вище. Каркасні ділянки можуть являти собою природні або консенсусні каркасні ділянки, переважно, людські каркасні ділянки (перелік людських каркасних ділянок можна знайти, наприклад, у Chothia, et al., J Mol Biol 278: 457 (1998)). Переважно, полінуклеотид, одержаний у результаті поєднання каркасних ділянок і CDR, кодує антитіло, що специфічно зв'язує поліпептид даного винаходу. Переважно, як вказано вище, у каркасних ділянках можна зробити одну або декілька амінокислотних замін, що, переважно, поліпшують зв'язування антитіла з антигеном. Крім того, такі способи можна використовувати для здійснення замін або делецій одного або декількох залишків цистеїну варіабельної ділянки, що беруть участь в утворенні внутрішньоланцюжкового дисульфідного зв'язку, з одержанням молекул антитіла, що втратили один або декілька внутрішньоланцюжкових дисульфідних зв'язків. Інші модифікації полінуклеотидів входять в обсяг даного винаходу і належать до компетенції фахівців у даній галузі. Крім того, можна використовувати способи, розроблені для одержання "химерних антитіл" (Morrison, et al., Proc Natl Acad Sci 81:851 (1984); Neuberger, et al., Nature 312:604 (1984); Takeda, et al., Nature 314:452 (1985)) шляхом сплайсинга генів молекули мишачого антитіла з відповідною специфічністю до антигена і генів молекули людського антитіла з відповідною біологічною активністю. Як вказано вище, химерне антитіло являє собою молекулу, що містить фрагменти, одержані з тваринних різних видів, такі як варіабельна ділянка мишачого MAb і константна ділянка людського імуноглобуліну, прикладом такої молекули є гуманізоване антитіло. Альтернативно, методи, описані для одержання одноланцюжкових антитіл (патент США № 4946778; Bird, Science 242:423 (1988); Huston, et al., Proc Natl Acad Sci USA 85:5879 (1988); і Ward, et al., Nature 334:544 (1989)), можна адаптувати для одержання одноланцюжкових антитіл. Одноланцюжкові антитіла одержують шляхом сполуки фрагментів ділянок Fv важкого і легкого ланцюга через амінокислотний місток з одержанням одноланцюжкового поліпептиду. Також можна використовувати способи зборки функціональних фрагментів Fv у E.coli (Skerra, et al., Science 242:1038 (1988)). Способи одержання антитіл проти NOTCH3 Антитіла даного винаходу можна одержати за допомогою відомих у даній галузі способів синтезу антитіл, зокрема, шляхом хімічного синтезу, або, переважно, за допомогою методів рекомбінантної експресії. Щоб здійснити рекомбінантну експресію антитіла даного винаходу, або його фрагмента, похідного або аналога (наприклад, важкого або легкого ланцюга антитіла даного або винаходу одноланцюжкового антитіла даного винаходу), потрібно сконструювати вектор експресії, що містить полінуклеотид, що кодує антитіло або фрагмент антитіла. Після одержання полінуклеотиду, що кодує молекулу антитіла, за допомогою технології рекомбінантних ДНК можна одержати вектор, що забезпечує продукцію антитіла. Конструюють вектор експресії, що містить послідовності, які кодують антитіло, і придатні послідовності, що контролюють транскрипцію і трансляцію. Дані способи містять у собі, наприклад, методи рекомбінації ДНК in vitro, синтетичні методи і методи генетичної рекомбінації in vivo. 17 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Вектор експресії переносять у клітину-хазяїна за допомогою традиційних методів, після чого трансфіковані клітини культивують звичайним способом, одержуючи антитіло даного винаходу. В одному аспекті даного винаходу вектори, що кодують важкі і легкі ланцюги, можна спільно експресувати у клітині-хазяїні з одержанням цілої молекули імуноглобуліну, як описано нижче. Як описано вище, для експресії молекул антитіла даного винаходу можна використовувати ряд систем хазяїн-вектор експресії. Такі системи хазяїн-вектор експресії являють собою засоби, за допомогою яких можна продукувати і потім очищувати кодуючи послідовності, що представляють інтерес, але вони також являють собою клітини, які, після трансформації або трансфекції відповідною кодуючою нуклеотидною послідовністю, можуть експресувати молекулу антитіла даного винаходу in situ. Для експресії рекомбінантної молекули антитіла, особливо, для експресії цілої рекомбінантної молекули антитіла зазвичай використовують бактеріальні клітини, такі як E.coli, і еукаріотичні клітини. Наприклад, клітини ссавців, такі як CHO, у поєднанні з вектором, що містить, наприклад, основний проміжний ранній промотор гена з людського цитомегаловірусу, є ефективною системою експресії антитіл (Foecking, et al., Gene 45:101 (1986); Cockett, et al., Bio/Technology 8:2 (1990)). Крім того, можна вибрати штам клітини-хазяїна, що модулює експресію введеної послідовності або модифікує і процесує генний продукт бажаним чином. Такі модифікації (наприклад, глікозилування) і процесинг (наприклад, розщеплення) білкових продуктів можуть дуже впливати на функціонування білка. Різні клітини-хазяїни мають характерні і специфічні механізми посттрансляційних процесинга і модифікації білків і генних продуктів. Щоб забезпечити правильні модифікацію і процесинг експресованого чужорідного білка, потрібно вибрати придатні клітинні лінії або системи хазяїнів. Для даної мети можна використовувати еукаріотичні клітини-хазяїни, що мають клітинний механізм, який забезпечує відповідний процесинг первинного транскрипта, а також відповідне глікозилування і фосфорилування генного продукту. Такі клітини-хазяїни ссавців містять у собі, без обмеження, клітини CHO, COS, 293, 3T3 або клітини мієломи. Щоб досягти довгострокову продукцію рекомбінантних білків з високим виходом переважно використовувати стабільну експресію. Наприклад, методом генної інженерії можна одержати клітинні лінії, що стабільно експресують молекулу антитіла. Клітини-хазяїни переважніше трансформувати не векторами експресії, що містять вірусні точки початку реплікації, а ДНК, що містить придатні елементи регуляції експресії (такі як промотор, енхансер, послідовності термінації транскрипції, ділянки поліаденілування та ін.) і маркер селекції. Після введення чужорідної ДНК рекомбінантні клітини залишають рости протягом одного-двох днів у збагаченому середовищі, що потім змінюють на середовище для селекції. Маркер селекції, що присутній у рекомбінантній плазміді, надає стійкість до умов селекції, дозволяє плазміді стабільно інтегруватися в хромосоми клітини і вирощувати клітини до утворення осередків, які, у свою чергу, можна клонувати і вирощувати з одержанням клітинних ліній. Даний спосіб можна успішно використовувати для одержання клітинних ліній, що експресують молекулу антитіла. Такі рекомбінантні клітинні лінії, зокрема, можна використовувати для скринінга й аналізу сполук, які безпосередньо або побічно взаємодіють з молекулою антитіла. Можна використовувати ряд систем селекції, що включає в себе, без обмеження, застосування генів тимідинкінази вірусу простого герпесу (Wigler, et al., Cell 11:223 (1977)), гіпоксантин-гуанін-фосфорибозилтрансферази (Szybalska, et al., Proc Natl Acad Sci USA 48:202 (1992)) і аденін-фосфорибозилтрансферази (Lowy, et al., Cell 22:817 (1980)) у поєднанні з tk-, hgprt- або aprt-клітинами, відповідно. Крім того, селекцію можна проводити на основі стійкості до антиметаболітів з використанням наступних генів: dhfr, що надає стійкість до метотрексату (Wigler, et al., Proc Natl Acad Sci USA 77:357 (1980); О'Hаrе, et al., Proc Natl Acad Sci USA 78:1527 (1981)); gpt, що надає стійкість до мікофенолової кислоти (Mulligan, et al., Proc Natl Acad Sci USA 78:2072 (1981)); neo, що надає стійкість до аміноглікозиду G-418 (Wu, et al., Biotherapy 3:87 (1991)); і hygro, що надає стійкість до гігроміцину (Santerre, et al., Gene 30:147 (1984)). Для селекції бажаного рекомбінантного клону можна використовувати способи, широко відомі в галузі технології рекомбінантних ДНК, такі способи описані, наприклад, у Ausubel, et al., eds., Current Protocols in Molecular Biology, John Wiley & Sons (1993); Kriegler, Gene Transfer and Expression, A Laboratory Manual, Stockton Press (1990); і в главах 12 і 13, Dracopoli, et al., eds, Current Protocols in Human Genetics, John Wiley & Sons (1994); Colberre-Garapin, et al., J Mol Biol 150:1 (1981), що включені в даний опис у вигляді посилання у всій повноті. Рівень експресії молекули антитіла можна підвисити шляхом ампліфікації вектора (огляд опублікований Bebbington, et al., "The use of vectors based on gene amplification for the expression of cloned genes in mammalian cells", DNA Cloning, Vol.3. Academic Press (1987)). Якщо маркер з векторної системи, що експресує антитіло, здатний ампліфікуватися, підвищення рівня 18 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 інгібітору, що присутні у культурі клітини-хазяїна, призводить до збільшення кількості копій маркерного гена. Оскільки ампліфікована ділянка зв'язана з геном антитіла, продукція антитіла також збільшується (Crouse, et al., Mol Cell Biol 3:257 (1983)). Клітини-хазяїна можна спільно трансфікувати двома векторами експресії даного винаходу, де перший вектор кодує поліпептид, що входить до складу важкого ланцюга, а другий вектор кодує поліпептид, що входить до складу легкого ланцюга. Два вектори можуть містити ідентичні маркери селекції, що забезпечують однаковий рівень експресії поліпептидів важкого і легкого ланцюга. Альтернативно, можна використовувати один вектор, що кодує і забезпечує експресію поліпептидів і важкого і легкого ланцюга. У таких випадках, щоб уникнути надлишку токсичного вільного важкого ланцюга, послідовність, що кодує легкий ланцюг, варто розташовувати перед послідовністю, що кодує важкий ланцюг (Proudfoot, Nature 322:52 (1986); Kohler, Proc Natl Acad Sci USA 77:2197 (1980)). Послідовності, що кодують важкі і легкі ланцюги, можуть містити кДНК або геномну ДНК. Після одержання молекули антитіла даного винаходу шляхом продукції в організмі тварини, хімічного синтезу або рекомбінантної експресії, її можна очистити за допомогою будь-якого відомих у даній галузі способу очищення молекул імуноглобуліну, такого як хроматографія (наприклад, іонообмінна, афінна, зокрема, заснована на тому, що спорідненість до конкретного антигена перевищує спорідненість до білка A, і ексклюзійна хроматографія), центрифугування, використання різниці в розчинностях, або за допомогою будь-якого іншого стандартного способу очищення білків. Крім того, щоб полегшити очищення, антитіла даного або винаходу їхні фрагменти можна гібридизувати з гетерологічними поліпептидними послідовностями, описаними в даному документі, або відомими в даній галузі. Даний винахід охоплює антитіла, гібридизовані з поліпептидом за допомогою рекомбінантних методів, або кон'юговані з поліпептидом за допомогою хімічних методів (кон'югація може бути як ковалентна, так і нековалентна). Гібридизовані або кон'юговані антитіла даного винаходу можна використовувати для полегшення очищення. Див., наприклад, публікацію PCT WO 93/21232; EP 439095; Naramura, et al., Immunol Lett 39:91 (1994); патент США № 5474981; Gillies, et al., Proc Natl Acad Sci USA 89:1428 (1992); Fell, et al., J Immunol 146:2446 (1991), що включені в даний опис у вигляді посилання у всій повноті. Щоб полегшити очищення, антитіла даного винаходу або їхні фрагменти також можна гібридизувати з маркерними послідовностями, такими як пептиди. У переважних втіленнях маркерна амінокислотна послідовність являє собою гексагістидиновий пептид, наприклад, маркер, що постачається у векторі pQE (QIAGEN, Inc., Chatsworth, CA), однак можна використовувати і інші послідовності, багато з яких є комерційно доступними. Як описано в Gentz, et al., Proc Natl Acad Sci USA 86:821 (1989), для традиційного очищення гібридного білка використовують, наприклад, гексагістидин. Інші пептидні маркери, використовувані для очищення, містять у собі, без обмеження, маркер "HA", що відповідає епітопа білка гемагглютиніну вірусу грипу (Wilson, et al., Cell 37:767 (1984)) і маркер "flag". Очищення антитіла При використанні рекомбінантних методів варіант антитіла може продукуватися у внутрішньоклітинний або периплазматичний простір, або секретуватися безпосередньо в середовище. Якщо варіант антитіла продукується всередині клітини, на першій стадії за допомогою центрифугування або ультрафільтрації можна видалити дисперсний дебрис, що складається або з клітин-хазяїнів, або з лізованих фрагментів. Carter, et al., Bio/Technology 10:163 (1992) описують спосіб виділення антитіл, що секретуються в периплазматичний простір E.coli. Коротко говорячи, клітинну масу відтаюють у присутності ацетату натрію (pН 3,5), EDTA і фенілметилсульфонілфториду (PMSF) протягом приблизно 30 хвилин. Клітинний дебрис можна видалити центрифугуванням. Якщо варіант антитіла секретується в середовище, супернатанти таких систем експресії спочатку концентрують, використовуючи комерційно доступні фільтри для концентрування білків, наприклад, пристрій для ультрафільтрації Amicon або Millipore Pellicon. На всіх описаних вище стадіях можна використовувати інгібітори протеаз, такі як PMSF, щоб інгібувати протеоліз, і антибіотики, щоб запобігти ріст випадкових забруднюючих організмів. Композицію антитіла, одержану з клітин, можна очистити, наприклад, за допомогою хроматографії на гідроксилапатиті, гель-електрофорезу, діалізу й афінної хроматографії, причому афінна хроматографія є переважним методом очищення. Можливість застосування білка A як афінного ліганду залежить від виду і ізотипу домену Fc імуноглобуліну, присутнього у варіанті антитіла. Білок A можна використовувати для очищення антитіла, що містить важкі ланцюги людських IgG1, IgG2 або IgG4 (Lindmark, et al., J Immunol Meth 62:1 (1983)). Для очищення всіх мишачих ізотипів і людського IgG3 рекомендують використовувати білок G (Guss, et al., EMBO J 5:1567 (1986)). Як основу, до якої приєднують афінний ліганд, найчастіше 19 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 використовують агарозу, однак можна використовувати і інші основи. Механічно стабільні основи, такі як скло з контрольованим розміром пор або полі(стиролдивініл)бензол, дозволяють використовувати більш високі швидкості потоку і більш короткий час обробки, ніж агароза. Якщо варіант антитіла містить домен CH3, для очищення використовують смолу Bakerbond ABXTM (J. T. Baker; Phillipsburg, N.J.). Також можна використовувати інші методи очищення білків, такі як фракціонування на іонообмінній колонці, осадження етанолом, ВЕРХ на зверненій фазі, TM хроматографія на оксиді кремнію, хроматографія на гепарин-вмісній SEPHAROSE , хроматографія на аніоно- або катіонообмінній смолі (наприклад, на колонці з поліаспарагіновою кислотою), хроматофокусування, SDS-PAGE, і осадження сульфатом амонію, залежно від одержуваного варіанта антитіла. Після попередньої стадії (стадій) очищення суміш, що містить варіант антитіла, що представляє інтерес, і забруднюючі домішки, можна піддавати хроматографії гідрофобних взаємодій при низьких значеннях pН, використовуючи для елюювання буфер, що має pН у діапазоні 2,5-4,5, переважно, при низьких концентраціях солей (наприклад, приблизно в діапазоні 0-0,25 M). Фармацевтична композиція Терапевтичні композиції поліпептиду або антитіла можна одержати для збереження у вигляді ліофілізованих композицій або водних розчинів шляхом змішування поліпептиду, що має бажаний ступінь чистоти, з необов'язковими "фармацевтично прийнятними" носіями, ексципієнтами або стабілізаторами, зазвичай використовуваними в даній галузі (усі вони мають загальну назву "ексципієнти"), що містять у собі, забуферувальні засоби, стабілізуючі засоби, консерванти, засоби, що надають розчину ізотонічність, неіонні детергенти, антиоксиданти та інші різнорідні добавки. См. Rеmіngtоn's Pharmaceutical Sciences, 16th edition, Osol, Ed. (1980). Такі добавки повинні бути нетоксичними для реципієнтів у використовуваних дозах і концентраціях. Забуферувальні засоби допомагають підтримувати pН в інтервалі, близькому до фізіологічних умов. Переважно вони присутні в концентраціях, що варіюють від приблизно 2 мм до приблизно 50 мм. Забуферувальні засоби, що підходять для застосування в даному винаході, містять у собі органічні і неорганічні кислоти і їхньої солі, такі як цитратні буфери (наприклад, суміш цитрату мононатрію і цитрату динатрію, суміш лимонної кислоти і цитрату тринатрію, суміш лимонної кислоти і цитрату мононатрію та ін.), сукцинатні буфери (наприклад, суміш бурштинової кислоти і сукцинату мононатрію, наприклад, суміш бурштинової кислоти і гідроксиду натрію, суміш бурштинової кислоти і сукцинату динатрію та ін.), тартратні буфери (наприклад, суміш винної кислоти і тартрату натрію, суміш винної кислоти і тартрату калію, суміш винної кислоти і гідроксиду натрію та ін.), фумаратні буфери (наприклад, суміш фумарової кислоти і фумарату мононатрію та ін.), фумаратні буфери (наприклад, суміш фумарової кислоти і фумарату мононатрію, суміш фумарової кислоти і фумарату динатрію, суміш фумарату мононатрію і фумарату динатрію та ін.), глюконатні буфери (наприклад, суміш глюконової кислоти і глюконату натрію, суміш глюконової кислоти і гідроксиду натрію, суміш глюконової кислоти і глюконату калію та ін.), оксалатні буфери (наприклад, суміш щавлевої кислоти й оксалату натрію, суміш щавлевої кислоти і гідроксиду натрію, суміш щавлевої кислоти й оксалату калію та ін.), лактатні буфери (наприклад, суміш молочної кислоти і лактату натрію, суміш молочної кислоти і гідроксиду натрію, суміш молочної кислоти і лактату калію та ін.) і ацетатні буфери (наприклад, суміш оцтової кислоти й ацетату натрію, суміш оцтової кислоти і гідроксиду натрію та ін.). Крім того, можна згадати фосфатні буфери, гістидинові буфери і солі триметиламіну, такі як Tris. Щоб уповільнити ріст мікробів, можна додати консерванти в кількостях, що варіюють від 0,2 % до 1 % (мас/об). Консерванти, що підходять для застосування в даному винаході, містять у собі фенол, бензиловий спирт, мета-крезол, метилпарабен, пропілпарабен, октадецилдиметилбензиламонію хлорид, галогеніди бензалконію (наприклад, хлорид, бромід, йодид), хлорид гексонію і алкілпарабени, такі як метил- або пропілпарабен, катехол, резорцин, циклогексанол і 3-пентанол. Щоб забезпечити ізотонічність рідких композицій даного винаходу, можна додати засоби, що надають розчину ізотонічність, іноді називані "стабілізатори", що містять у собі багатоатомні цукрові спирти, переважно, цукрові спирти, що містять три або більше гідроксильні групи, такі як гліцерин, еритритол, арабіт, ксиліт, сорбіт і маніт. Стабілізатори належать до великої категорії ексципієнтів, що об'єднує речовини, які мають різні функції, від агентів-наповнювачів до добавок, які сприяють солюбілізації терапевтичного засобу або запобігають його денатурації або прилипанню до стінки контейнера. До типових стабілізаторів належать багатоатомні цукрові спирти (перераховані вище); амінокислоти, такі як 20 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 аргінін, лізин, гліцин, глутамін, аспарагін, гістидин, аланін, орнітин, L-лейцин, 2-фенілаланін, глутамінова кислота, треонін і ін., органічні цукри або цукрові спирти, такі як лактоза, трегалоза, стахіоза, маніт, сорбіт, ксиліт, рибіт, міоінозит, галактитол, гліцерин і т.п., у тому числі, циклічні спирти, такі як інозитол; поліетиленгліколь; амінокислотні полімери; сірковмісні відновлювальні засоби, такі як сечовина, глутатіон, тіоктова кислота, тіогліколят натрію, тіогліцерин, альфамонотіогліцерин і тіосульфат натрію; низькомолекулярні поліпептиди (тобто 10 залишків); білки, такі як людський сироватковий альбумін, бичачий сироватковий альбумін, желатин або імуноглобуліни; гідрофільні полімери, такі як полівінілпіролідон, моносахариди, такі як ксилоза, маноза, фруктоза, глюкоза; дисахариди, такі як лактоза, мальтоза, сахароза, і трисахариди, такі як рафіноза; а також полісахариди, такі як декстран. Стабілізатори можуть бути присутніми у кількості від 0,1 до 10000 масових часток відносно маси активного білка. Неіонні поверхнево-активні речовини або детергенти (також відомі як "зволожувальні засоби") можна додати для полегшення солюбілізації терапевтичного засобу, а також для захисту терапевтичного білка від агрегації, викликаної перемішуванням, дані засоби також запобігають денатурації білка, що виникає при впливі на композицію тиску поверхневого зсуву. Придатні неіонні поверхнево-активні речовини містять у собі полісорбати (20, 80 і ін.), поліоксамери (184, 188 і ін.), поліоли Pluronic.RTM., прості моноефіри поліоксіетилену і сорбітану (TWEEN-20®, TWEEN-80® і ін.). Неіонні поверхнево-активні речовини можуть бути присутніми у кількостях, що варіюють від приблизно 0,05 мг/мл до приблизно 1,0 мг/мл, переважно від приблизно 0,07 мг/мл до приблизно 0,2 мг/мл. Інші різнорідні ексципієнти містять у собі агенти-наповнювачі (наприклад, крохмаль), хелатувальні засоби (наприклад, EDTA), антиоксиданти (наприклад, аскорбінова кислота, метіонін, вітамін E) і співрозчинники. Композиція даного винаходу також може містити декілька активних сполук, якщо це необхідно для лікування конкретних симптомів, переважно такі сполуки мають взаємодоповнюючі активності, що не роблять несприятливого впливу одна на одну. Наприклад, іноді бажано додатково використовувати імунодепресивний засіб. Такі молекули можуть бути присутніми у поєданні в кількостях, ефективних для досягнення передбачуваної мети. Активні інгредієнти також можна укласти в мікрокапсулу, одержану, наприклад, шляхом коацервації або міжфазної полімеризації у випадку гідроксиметилцелюлозної або желатинової мікрокапсули і полі(метилметакрилатної) мікрокапсули, відповідно, у колоїдні системи доставки лікарських засобів (такі як ліпосоми, альбумінові мікросфери, мікроемульсії, наночастинки і нанокапсули), або в макроемульсії. Такі методи описані в Rеmіngtоn's Pharmaceutical Sciences, 16th edition, Osal, Ed. (1980). Композиції, призначені для введення in vivo, повинні бути стерильними. Стерилізацію можна легко здійснити, наприклад, шляхом фільтрації через стерильні фільтраційні мембрани. Можна використовувати препарати з уповільненим вивільненням. Придатні приклади препаратів з уповільненим вивільненням містять у собі напівпроникні матриці з твердих гідрофобних полімерів, що містять варіант антитіла, дані матриці можуть знаходитися у вигляді профільованих виробів, таких як або плівки мікрокапсули. Приклади матриць з уповільненим вивільненням містять у собі складні поліефіри, гідрогелі (такі як полі(2-гідроксіетилметакрилат), полі(вініловий спирт)), полілактиди (патент США № 3773919), співполімери L-глутамінової кислоти і етил-L-глутамату, що не деградує етиленвінілацетат, деградуючі співполімери TM молочної кислоти і гліколевої кислоти, такі як LUPRON DEPOT (придатні для ін'єкцій мікросфери, що складаються із співполімеру молочної кислоти і гліколевої кислоти і лейпроліду ацетату), і полі-D-(-)-3-гідроксимасляна кислота. Хоча полімери, такі як етиленвінілацетат і молочна кислота-гліколева кислота, здатні вивільнювати молекули протягом 100 днів, деякі гідрогелі вивільняють білки протягом більш коротких періодів. Якщо поміщені в капсулу антитіла залишаються в організмі протягом тривалого часу, вони можуть денатурувати або агрегувати під дією вологи при 37C, що може призвести до втрати біологічної активності і можливих змін імуногенності. Залежно від задіяного механізму можна використовувати раціональні стратегії стабілізації. Наприклад, якщо показано, що механізм агрегації містить у собі утворення міжмолекулярних зв'язків S-S у результаті тіодисульфідної взаємодії, препарат можна стабілізувати шляхом модифікації сульфгідрильних залишків, ліофілізації з кислих розчинів, контролювання вмісту вологи, застосування придатних добавок і розробки специфічних полімерних матриксних композицій. Кількість терапевтичного поліпептиду, антитіла або його фрагмента, ефективна для лікування конкретного порушення або стану, залежить від природи порушення або стану і може бути визначена за допомогою стандартних клінічних методів. По можливості, перед тестуванням на людях бажано одержати криву доза-відповідь для фармацевтичних композицій даного винаходу спочатку in vitro і потім на придатних тваринних модельних системах. 21 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 У переважному втіленні водний розчин терапевтичного поліпептиду, або антитіла його фрагмента вводять шляхом підшкірної ін'єкції. Кожна доза може варіювати від приблизно 0,5 мкг до приблизно 50 мкг на кілограм маси тіла, або, більш переважно, від приблизно 3 мкг до приблизно 30 мкг на кілограм маси тіла. Дозувальний режим для підшкірного введення може варіювати від одного разу на місяць до щоденного введення, залежно від ряду клінічних факторів, що включають у себе тип захворювання, тяжкість захворювання і сприйнятливість суб'єкта до терапевтичного засобу. Терапевтичне застосування антитіл проти NOTCH-3 Антитіла даного винаходу передбачається використовувати для лікування ссавців. В одному втіленні антитіло вводять відмінному від людини ссавцю, наприклад, з метою одержання передклінічних даних. Приклади відмінних від людини ссавців, що підлягають лікуванню, містять у собі відмінних від людини приматів, собак, кішок, гризунів і інших ссавців, на яких проводять передклінічні випробування. Таких ссавців можна використовувати для одержання тваринних моделей захворювання, що підлягає лікуванню антитілом, або для дослідження токсичності представляючого інтерес антитіла. В всіх описаних втіленнях дослідження з впливу підвищення дози можна проводити на ссавці. Антитіло, що вводиться або окремо в поєднанні з іншим фактором (факторами), можна використовувати як терапевтичний засіб. Даний винахід стосується способів лікування антитілами, що містять у собі введення антитіла даного винаходу тварині, ссавцю або людині, для лікування Notch3-опосередкованого захворювання, порушення або стану. Тварина або суб'єкт може являти собою ссавця, що потребує конкретного лікування, такий як ссавець, у якого діагностоване конкретне порушення, наприклад, пов'язане з Notch3. Антитіла, направлені проти Notch3, можна використовувати для лікування дегенеративних захворювань і інших Notch3-асоційованих захворювань, що включають у себе CADASIL, FHM, синдром Алагілля, неврологічні і дегенеративні порушення у ссавців, таких як, без обмеження, корови, свині, коні, кури, кішки, собаки, відмінні від людини примати та ін., а також у людей. Наприклад, шляхом введення терапевтично прийнятної дози антитіла або антитіл проти Notch3 даного винаходу, або суміші антитіл даного винаходу, або їх поєднання з іншими антитілами з різних джерел, симптоми захворювання можна поліпшити або запобігти у ссавця, що піддається лікуванню, особливо, людини. Терапевтичні сполуки даного винаходу містять у собі, без обмеження, антитіла даного винаходу (у тому числі їхні фрагменти, аналоги і похідні, як описано в даному документі) і нуклеїнові кислоти, що кодують антитіла даного винаходу, як описано нижче (у тому числі їхні фрагменти, аналоги і похідні, а також антиідіотипічні антитіла, як описано в даному документі). Антитіла даного винаходу можна використовувати для лікування, пригнічення або профілактики захворювань, порушень або станів, пов'язаних з аберантною експресією і/або активністю Notch3, що містять у собі, без обмеження, будь-яке одне або декілька з захворювань, порушень або станів, описаних у даному документі. Лікування і/або профілактика захворювань, порушень або станів, пов'язаних з аберантною експресією і/або активністю Notch3, містить у собі, без обмеження, полегшення щонайменше одного симптому, пов'язаного з зазначеними захворюваннями, порушеннями або станами. Антитіла даного винаходу можна ввести до складу фармацевтично прийнятних композицій за допомогою способів, відомих у даній галузі або описаних у даному документі. Антитіла проти Notch3 даного винаходу можна використовувати для терапевтичного лікування ряду захворювань. Даний винахід пропонує спосіб профілактики або лікування Notch3-опосередкованих захворювань у ссавців. Даний спосіб містить у собі введення ссавцю ефективної для профілактики або лікування захворювання кількості антитіла проти Notch3. Антитіло проти Notch3 зв'язується з Notch3 і пригнічує його функціонування. Сигнальний шлях Notch3 пов'язаний з різними захворюваннями, такими як CADASAL, FHM, сімейна парокситична атаксія, синдром Алагілля та інші дегенеративні захворювання і неврологічні порушення (Joutel, et al., Nature 383:707 (1996); Flynn, et al., J Pathol 204:55 (2004)). Вважають, що антитіла проти Notch3 можна ефективно використовувати для лікування вищевказаних захворювань. Кількість антитіла, ефективна для лікування, пригнічення і профілактики захворювання або порушення, пов'язаного з аберантною експресією і/або активністю Notch3, можна визначити за допомогою стандартних клінічних методів. Доза залежить від типу захворювання, що підлягає лікуванню, тяжкості і перебігу захворювання, від того, з якою метою, профілактичною або терапевтичною, вводять антитіло, попередньої терапії, клінічної історії пацієнта і відповіді на антитіло, а також від рішення лікуючого лікаря. Щоб поліпшити стан захворювання, антитіло можна вводити в режимі лікування, залежно від захворювання, наприклад, в одній або декількох дозах протягом від одного до декількох днів, або дози антитіла можна вводити періодично 22 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 протягом тривалого часу, щоб пригнітити розвиток захворювання і запобігти рецидив. Крім того, оптимальні інтервали доз можна визначити за допомогою аналізів in vitro. Точна доза, використовувана в композиції, також залежить від способу введення, тяжкості захворювання або порушення, і встановлюється відповідно до висновку лікуючого лікаря і стану кожного пацієнта. Ефективні дози можна визначити на підставі кривих доза-відповідь, одержаних in vitro або на тваринних модельних системах. Антитіла вводять пацієнту в дозі, що зазвичай складає від 0,1 мг/кг до 150 мг/кг маси тіла пацієнта. Переважно, доза, що вводиться пацієнту, складає від 0,1 мг/кг до 20 мг/кг маси тіла пацієнта, більш переважно, від 1 мг/кг до 10 мг/кг маси тіла пацієнта. Як правило, людські антитіла мають більш тривалий період напівжиття в організмі людини, ніж антитіла з інших видів внаслідок імунної відповіді на чужорідні поліпептиди. Як правило, людські антитіла найчастіше можна використовувати в більш низьких дозах і вводити їх можна рідше. Крім того, дозу і частоту введення антитіл даного винаходу можна зменшити шляхом підвищення споживання і проникання в тканині (наприклад, у мозок) антитіл шляхом модифікацій, таких як, наприклад, ліпідизація. При багаторазовому введенні протягом декількох днів або довше, залежно від стану, обробку продовжують до бажаного пригнічення симптомів захворювання. Однак можна використовувати інші режими дозування. Результат даної терапії можна легко відслідковувати за допомогою традиційних методів і аналізів. Композицію варіанта антитіла можна одержувати, дозувати і вводити відповідно до успішної медичної практики. Фактори, що враховуються в даному контексті, містять у собі конкретне порушення, що піддається лікуванню, конкретний ссавець, що піддається лікуванню, клінічний стан окремого пацієнта, причину порушення, ділянку доставки засобу, спосіб введення, режим уведення, а також інші фактори, відомі лікарям. "Терапевтично ефективна кількість" варіанта антитіла, що вводиться, залежить від таких факторів і являє собою мінімальну кількість, необхідну для профілактики, поліпшення або лікування захворювання або порушення. Варіант антитіла може необов'язково входити до складу композиції поряд з одним або декількома засобами, використовуваними в даний час для профілактики або лікування розглянутого порушення. Ефективна кількість таких інших засобів залежить від кількості антитіла, що присутня у композиції, типу порушення або способу лікування, а також від інших факторів, вказаних вище. Як правило, їх використовують у таких же дозах за допомогою таких же способів введення, як вказано вище в даному документі, або в кількостях, що складають приблизно від 1 до 99 % від раніше використовуваних доз. Антитіла даного винаходу можна вводити окремо або в поєднанні з іншими типами лікування. У переважному аспекті антитіло є практично очищеним (наприклад, практично не містить речовин, що обмежують його або дію роблять небажані побічні ефекти). Відомі різні системи доставки, які можна використовувати для введення антитіла даного винаходу, у тому числі, шляхом ін'єкції, наприклад, ліпосоми, що містять сполуку, мікрочастинки, мікрокапсули, рекомбінантні клітини, здатні експресувати сполуку, рецептор-опосередкований ендоцитоз (див., наприклад, Wu, et al., J Biol Chem 262:4429 (1987)), конструкції нуклеїнової кислоти в складі ретровірусного або іншого вектора та ін. Антитіло проти Notch3 можна вводити ссавцю будь-яким прийнятним способом. Способи введення містять у собі, без обмеження, парентеральний, підшкірний, внутрішньочеревинний, внутрішньолегеневий, внутрішньоназальний, епідуральний, інгаляцію і пероральний, і, якщо це необхідно для імунодепресивного лікування, введення в ділянку ураженої тканини. Парентеральні інфузії містять у собі внутрішньом'язове, внутрішньошкірне, внутрішньовенне, внутрішньоартеріальне або внутрішньочеревинне введення. Антитіла або композиції можна вводити за допомогою будь-якого традиційного способу, наприклад, шляхом інфузії або болюсної ін'єкції, шляхом абсорбції через епітеліальну або шкірно-слизову вистілки (такі як слизова оболонка порожнини рота, ректальна і кишкова слизові оболонки та ін.), причому їх можна вводити разом з іншими біологічно активними засобами. Введення може бути системним або локальним. Крім того, іноді бажано вводити терапевтичні антитіла або композиції даного винаходу в центральну нервову систему за допомогою будь-якого придатного способу, що включає в себе внутрішньошлуночкове і внутрішньооболонкове введення; внутрішньошлуночкове введення можна здійснювати за допомогою внутршіньошлуночкового катетера, наприклад, приєднаного до резервуара, такого як резервуар Ommaya. Антитіло також можна вводити шляхом імпульсної інфузії, особливо, зі зменшенням доз антитіла. Переважно, введення здійснюють шляхом ін'єкцій, найбільше переважно, внутрішньовенних або підшкірних ін'єкцій, зазвичай, залежно від того, чи є введення коротким або хронічним. 23 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 Крім того, можна використовувати легеневе введення, наприклад, шляхом застосування інгалятора або розпилювача і композиції, що містить засіб, який забезпечує аерозолізацію. Антитіло також можна вводити в легені пацієнта у вигляді сухої порошкоподібної композиції (див., наприклад, патент США № 6514496). У конкретному втіленні може бути бажано вводити терапевтичні антитіла або композиції даного винаходу локально в ділянку, що потребує лікування; цього можна досягти, наприклад, без обмеження, за допомогою локальної інфузії, місцевого нанесення, ін'єкції, за допомогою катетера, за допомогою свіч або за допомогою імплантату, причому вказаний імплантат складається з пористої, не пористої або гелеподбіної речовини, і містить у собі мембрани, такі як силіконові мембрани, або волокна. Переважно, при введенні антитіла даного винаходу слід додержуватися обережності при застосуванні речовин, на яких білок не абсорбується. В іншому втіленні антитіло може доставлятися у везикулі, зокрема, у ліпосомі (див. Langer, Science 249:1527 (1990); Treat, et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein, et al., eds., pp. 353-365 (1989); Lopez-Berestein, там же., pp. 317-27; див. у цілому там же). У наступному втіленні антитіло можна доставляти в системі з контрольованим вивільненням. В одному втіленні можна використовувати нагнітач (див. Langer, Science 249:1527 (1990); Sefton, CRC Crit Ref Biomed Eng 14:201 (1987); Buchwald, et al., Surgery 88:507 (1980); Saudek, et al., N Engl J Med 321:574 (1989)). В іншому втіленні можна використовувати полімерні матеріали (див. Medical Applications of Controlled Release, Langer, et al., eds., CRC Press (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen, et al., eds., Wiley (1984); Ranger, et al., J Macromol Sci Rev Macromol Chem 23:61 (1983); див. також Levy, et al., Science 228:190 (1985); During, et al., Ann Neurol 25:351 (1989); Howard, et al., J Neurosurg 71:105 (1989)). У наступному втіленні систему з контрольованим вивільненням можна помістити поблизу терапевтичної мішені. Даний винахід також пропонує фармацевтичні композиції. Такі композиції містять терапевтично ефективну кількість антитіла і фізіологічно прийнятний носій. У конкретному втіленні термін "фізіологічно прийнятний" стосується засобу, затвердженому контрольним органом Федерального уряду або уряду штату, або перерахованому у Фармакопеї США або інших широко відомих фармакопеях для застосування на тваринах, більш конкретно, на людях. Термін "носій" стосується розріджувача, ад'юванта, ексиципієнта або середовища, що вводять разом з терапевтичним засобом. Такі фізіологічні носії можуть являти собою стерильні рідини, такі як вода й масла, у тому числі нафтового, тваринного, рослинного або синтетичного походження, такі як арахісова олія, соєва олія, мінеральне масло, кунжутна олія і т.п. Вода являє собою переважний носій, якщо фармацевтичну композицію вводять внутрішньовенно. Сольові розчини і водні розчини декстрози і гліцерину також можна використовувати як рідкі носії, особливо, для ін'єктованих розчинів. Придатні фармацевтичні ексципієнти містять у собі крохмаль, глюкозу, лактозу, сахарозу, желатин, солод, рис, борошно, крейду, силікагель, стеарат натрію, моностеарат гліцерину, тальк, хлорид натрію, сухе зняте молоко, гліцерин, пропіленгліколь, воду, етанол і т.п. При необхідності композиція також може містити мінорні кількості зволожуючих або емульгувальних засобів, або забуферувальні засоби. Дані композиції можуть знаходитися у вигляді розчинів, суспензій, емульсій, таблеток, пігулок, капсул, порошків, композицій з уповільненим вивільненням і т.п. Композиція може знаходитися у вигляді свічі, що містить традиційні зв'язувальні і носії, такі як тригліцериди. Пероральні композиції можуть містити в собі стандартні носії, такі як маніт, лактоза, крохмаль, стеарат магнію, сахарин натрію, целюлоза, карбонат натрію та ін., що мають фармацевтичну категорію чистоти. Приклади придатних носіїв описані в "Rеmіngtоn's Pharmaceutical Sciences", E. W. Martin. Такі композиції містять ефективну кількість антитіла, переважно, в очищеному виді, поряд з кількістю носія, що підходить для одержання форми для відповідного введення пацієнту. Композиція повинна відповідати способу введення. В одному втіленні композицію одержують відповідно до рутинних способів як фармацевтичну композицію, адаптовану для внутрішньовенного введення людям. Як правило, композиції для внутрішньовенного введення являють собою розчини в стерильному ізотонічному водному буфері. За необхідності композиція також може містити солюбілізувальний засіб і місцевий анестетик, такий як лігнокаїн, щоб зменшити біль у ділянці ін'єкції. Як правило, інгредієнти поставляються по окремості або в суміші у вигляді одиничної дозованої форми, наприклад, у вигляді сухого ліофілізованого порошку або безводного концентрату в герметично закритому контейнері, такому як ампула або саше, із вказуванням кількості активного засобу. Якщо композицію вводять шляхом інфузії, її можна дозувати за допомогою сулії для інфузії, що містить стерильну воду або стерильний фізіологічний розчин 24 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 фармацевтичної категорії чистоти. Якщо композицію вводять шляхом ін'єкції, може надаватися ампула зі стерильною водою для ін'єкції або зі стерильним фізіологічним розчином для змішування інгредієнтів перед введенням. Даний винахід також надає фармацевтичну упаковку або набір, що містить один або декілька контейнерів, заповнених одним або декількома інгредієнтами фармацевтичних композицій даного винаходу. З таким контейнером (контейнерами) може бути необов'язково пов'язане повідомлення у формі, встановленій державним органом, що контролює виробництво, застосування або продаж фармацевтичних засобів або біологічних продуктів, що свідчить про те, що даний орган дозволяє виробництво, застосування або продаж засобу для введення людині. Промислові вироби В іншому втіленні даний винахід пропонує промисловий виріб, що містить речовини, використовувані для лікування описаних вище порушень. Промисловий виріб містить у собі контейнер і етикетку. Придатні вироби містять у собі, наприклад, сулії, флакони, шприци і пробірки. Контейнери можуть бути виготовлені з різних матеріалів, наприклад, зі скла або пластмаси. Контейнер містить композицію, ефективну для профілактики або лікування стану, і може мати отвір для стерильного доступу (наприклад, контейнер може являти собою мішок для внутрішньовенного розчину або флакон, що має пробку, яку можна проколоти голкою для підшкірних ін'єкцій). Активний засіб у композиції являє собою антитіло. Етикетка, на контейнері або приєднана до контейнера, вказує, що композиція призначена для лікування визначеного стану. Промисловий виріб може додатково містити в собі другий контейнер, що містить фармацевтично прийнятний буфер, такий як забуферений фосфатом фізіологічний розчин і розчин декстрози. Він також може містити інші компоненти, необхідні з погляду виробника і споживача, у тому числі, інші буфери, розріджувачі, фільтри, голки, шприци і листки-вкладиші в упаковці з інструкціями з застосування. Генна терапія на основі антитіл В іншому аспекті даний винахід пропонує спосіб генної терапії, в якому нуклеїнові кислоти, що містять послідовності, які кодують антитіла або їхні функціональні похідні, вводять для лікування, пригнічення або профілактики захворювання або порушення, пов'язаного з аберантною експресією і/або активністю Notch3. Генна терапія стосується терапії, проведеної шляхом введення суб'єкту експресованої або здатної експресуватися нуклеїнової кислоти. У даному втіленні даного винаходу нуклеїнові кислоти продукують кодований ними білок, що опосередковує терапевтичний ефект. Відповідно до даного винаходу можна використовувати будь-які доступні способи генної терапії. Приклади способів описані нижче. Загальні огляди з способів генної терапії можна знайти в Goldspiel, et al., Clinical Pharmacy 12:488 (1993); Wu, et al., Biotherapy 3:87 (1991); Tolstoshev, Ann Rev Pharmacol Toxicol 32:573 (1993); Mulligan, Science 260:926 (1993); Morgan, et al., Ann Rev Biochem 62:191 (1993); May, TIBTECH 11:155 (1993). В одному аспекті сполука містить у собі нуклеотидні послідовності, що кодують антитіло, причому вказані нуклеотидні послідовності є частиною векторів експресії, що забезпечують експресію антитіла або його фрагментів, або химерних білків на основі антитіла, або важких або легких ланцюгів антитіла в придатному хазяїні. Зокрема, такі нуклеотидні послідовності містять промотори, функціонально зв'язані з ділянкою, що кодує антитіло, причому вказаний промотор є або індукованим, або конститутивним і, необов'язково, тканиноспецифічним. В іншому конкретному втіленні використовують молекули нуклеїнових кислот, в яких послідовності, що кодують антитіло, а також будь-які інші необхідні послідовності фланкуються ділянками, що стимулюють гомологічну рекомбінацію в цільовій ділянці генома, забезпечуючи, таким чином, внутрішньохромосомну експресію нуклеїнових кислот, що кодують антитіло (Koller, et al., Proc Natl Acad Sci USA 86:8932 (1989); Zijlstra, et al., Nature 342:435 (1989)). У конкретних втіленнях експресована молекула антитіла являє собою одноланцюжкове антитіло; альтернативно, нуклеотидні послідовності містять послідовності, що кодують і важкі і легкі ланцюги антитіла, або їхні фрагменти. Доставка нуклеїнових кислот пацієнту може бути або безпосередньою, при якій пацієнту безпосередньо вводять нуклеїнову кислоту, або вектори, що несуть нуклеїнову кислоту, або непрямою, при якій клітини спочатку трансформують нуклеїновою кислотою in vitro, а потім трансплантують пацієнту. Описані способи відомі як генна терапія in vivo або ex vivo, відповідно. У конкретному втіленні нуклеотидні послідовності безпосередньо вводять в організм, де вони експресуються і продукують кодований продукт. Таке введення можна здійснити за допомогою кожного з численних способів, відомих у даній галузі, наприклад, шляхом включення їх до складу придатного вектора експресії нуклеїнової кислоти з наступним його введенням таким чином, щоб послідовності стали внутрішньоклітинними, наприклад, шляхом інфікування 25 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 дефектними або послабленими ретровірусами або іншими вірусними векторами (див. патент США № 4980286), або шляхом безпосереднього введення оголеної ДНК, або шляхом бомбардування мікрочастинками (наприклад, з використанням генної гармати; Biolistic, Dupont), або шляхом покриття ліпідами або клітинними поверхневими рецепторами або трансформуючими засобами, поміщення в ліпосоми, мікрочастинки або мікрокапсули, або шляхом введення нуклеотидних послідовностей, зв'язаних з пептидом, здатним проникати в ядро, або шляхом введення нуклеотидних послідовностей, зв'язаних з лігандом, здатним піддаватися рецептор-опосередкованому ендоцитозу (див., наприклад, Wu, et al., J Biol Chem 262:4429 (1987)) (яке можна використовувати для направлення до типів клітин, специфічно експресуючих рецептори) та ін. В іншому втіленні можна одержати комплекси нуклеїнова кислота-ліганд, де ліганд містить зливаючий вірусний пептид, що руйнує ендосоми і дозволяє нуклеїновій кислоті уникнути лізосомної деградації. У наступному втіленні нуклеїнова кислота, в результаті її напрямленості на конкретний рецептор, може специфічно поглинатися клітинами in vivo і експресуватися (див., наприклад, публікації PCT WO 92/06180; WO 92/22635; WO92/20316; WO93/14188, WO 93/20221). Альтернативно нуклеїнову кислоту можна ввести всередину клітини і впровадити в ДНК клітину-хазяїна з наступною експресією шляхом гомологічної рекомбінації (Koller, et al., Proc Natl Acad Sci USA 86:8932 (1989); Zijlstra, et al., Nature 342:435 (1989)). У конкретному втіленні використовують вірусні вектори, що містять нуклеотидні послідовності, що кодують антитіло даного винаходу. Наприклад, можна використовувати ретровірусний вектор (див. Miller, et al., Meth Enzymol 217:581 (1993)). Вказані ретровірусні вектори містять компоненти, необхідні для правильного укладання вірусного генома та інтеграції його в ДНК клітину-хазяїна. Нуклеотидні послідовності, що кодують антитіло, яке призначається для застосування в генній терапії, клонують в одному або декількох векторах, що забезпечують доставку гена пацієнту. Більш докладну інформацію по ретровірусним векторам можна знайти в Boesen, et al., Biotherapy 6:291 (1994), де описане застосування ретровірусного вектора для доставки гена mdrl у кровотворні стовбурові клітини, щоб зробити дані клітини більш стійкими до хіміотерапії. Інші посилання ілюструють застосування ретровірусних векторів у генній терапії: Clowes, et al., J Clin Invest 93:644 (1994); Kiem, et al., Blood 83:1467 (1994); Salmons, et al., Human Gene Therapy 4:129 (1993); і Grossman, et al., Curr Opin Gen and Dev 3:110 (1993). У даному винаході також можна використовувати аденовіруси. Відповідно до даного винаходу аденовіруси є особливо ефективним засобом доставки антитіл до епітелію дихальних шляхів. У природі аденовіруси інфікують епітелій дихальних шляхів. Іншими мішенями для систем доставки на основі аденовірусів є печінка, центральна нервова система, ендотеліальні клітини і м'язи. Перевага аденовірусів полягає в тому, що вони здатні інфікувати клітини, які не поділяються. Kozarsky, et al., Curr Opin Gen Dev 3:499 (1993) опублікували огляд з генної терапії на основі аденовірусів. Bout, et al., Human Gene Therapy 5:3 (1994) продемонстрували застосування аденовірусних векторів для перенесення генів в епітелій дихальних шляхів макакрезус. Інші випадки застосування аденовірусів у генній терапії описані в Rosenfeld, et al., Science 252:431 (1991); Rosenfeld, et al., Cell 68:143 (1992); Mastrangeli, et al., J Clin Invest 91:225 (1993); публікації PCT WO94/12649; Wang, et al., Gene Therapy 2:775 (1995). Для застосування в генній терапії також пропонуються адено-асоційовані віруси (AAV) (Walsh, et al., Proc Soc Exp Biol Med 204:289 (1993); патенти США №№ 5436146; 6632670; і 6642051). Інший підхід до генної терапії містить у собі перенесення гена в клітини тканинної культури за допомогою таких методів, як електропорація, ліпофекція, трансфекція, опосередкована фосфатом кальцію або вірусна інфекція. Як правило, спосіб перенесення містить у собі перенесення у клітини маркера селекції. Потім клітини піддають селекції, відбираючи клітини, що поглинули і експресують ген, який переноситься. Такі клітини згодом вводять пацієнту. У даному втіленні нуклеїнову кислоту вводять в клітину до введення в організм, одержуючи рекомбінантну клітину. Таке введення можна здійснювати за допомогою будь-якого відомого в даній галузі способу, що включає в себе, без обмеження, трансфекцію, електропорацію, мікроін'єкцію, інфікування вірусним або бактеріальним вектором, що містить нуклеотидні послідовності, гібридизацію клітин, хромосома-опосередковане перенесення гена, опосередковане мікропорами перенесення гена, сферопластну гібридизацію та ін. У даній галузі відомо багато способів введення чужорідних генів в клітини (див., наприклад, Loeffler, et al., Meth Enzymol 217:599 (1993); Cohen, et al., Meth Enzymol 217:618 (1993); Cline, Pharmac Ther 29:69 (1985)), які можна використовувати згідно з даним винаходом, за умови, що вони не порушують важливих функцій, пов'язаних з розвитком, а також фізіологічних функцій клітинреципієнтів. Спосіб повинен забезпечувати стабільну трансфекцію нуклеїнової кислоти в клітину 26 UA 102061 C2 5 10 15 20 25 30 35 40 45 50 55 60 так, щоб нуклеїнова кислота могла експресуватися клітиною і переважно успадковуватися і експресуватися потомством даної клітини. Одержані рекомбінантні клітини можна доставити пацієнту за допомогою різних способів, відомих у даній галузі. Рекомбінантні клітини крові (наприклад, кровотворні стовбурові клітини або попередники кровотворних клітин) переважно вводять внутрішньовенно. Кількість клітин, необхідна для застосування, залежить від бажаного ефекту, стану пацієнта та ін., і може визначатися фахівцем у даній галузі. Клітини, в які можна ввести нуклеїнову кислоту з метою генної терапії, охоплюють будь-які бажані доступні типи клітин і містять у собі, без обмеження, епітеліальні клітини, ендотеліальні клітини, кератиноцити, фібробласти, м'язові клітини, гепатоцити; клітини крові, такі як Tлімфоцити, B-лімфоцити, моноцити, макрофаги, нейтрофіли, еозинофіли, мегакаріоцити, гранулоцити; різні стовбурові клітини або клітини-попередники, зокрема, кровотворні стовбурові клітини і попередники кровотворних клітин, наприклад, одержані з кісткового мозку, крові пуповини, периферичної крові, ембріональної печінки та ін. В одному втіленні клітина, використовувана для генної терапії, є аутологічною відносно пацієнта. Нуклеотидні послідовності, що кодують антитіло даного винаходу, вводять у клітини так, щоб забезпечити їхню експресію клітинами або їх потомством, після чого рекомбінантні клітини вводять в організм, одержуючи терапевтичний ефект. У конкретному втіленні використовують стовбурові клітини або клітини-попередники. Відповідно до даного втілення даного винаходу можна використовувати будь-які стовбурові клітини і/або клітини-попередники, які можна виділити і підтримувати in vitro (див., наприклад публікацію PCT WO 94/08598; Stemple, et al., Cell 71:973 (1992); Rheinwald, Meth Cell Bio 21A:229 (1980); Pittelkow, et al., Mayo Clinic Proc 61:771 (1986)). ПРИКЛАДИ Приклад 1: Одержання імуногена: гібридного білка позаклітинного домену NOTCH3-FC Моноклональні антитіла проти Notch3, специфічно зв'язувальні LIN12/домен димеризації (далі називаний "LD") людського Notch3, одержують, використовуючи як імуноген рекомбінантний гібридний білок Notch3-Fc, що містить LD Notch3, по карбоксильному кінці якого приєднана ділянка гамма 1 Fc. А саме, імуноген являє собою гібридний білок (Notch3 LD/Fc), що містить амінокислотні залишки 1378-1640 LD Notch3 (див. фіг.1) і людський фрагмент γ1Fc. Контрольне антитіло одержують, використовуючи в якості імуногена ділянку повтору EGF Notch3, що містить амінокислотні залишки 43-1377. Послідовність білка Notch3 аналізують за допомогою інтерактивних дослідницьких програмних забезпечень і служб (Motif Search, http://motif.genome.ip/). РНК людської печінки і підшлункової залози (Ambion, Inc. Austin, TX) використовують як матрицю для синтезу першого ланцюга кДНК із використанням стандартного комерційно доступного набору для синтезу кДНК. кДНК, що кодує LD Notch3, і ділянку повтору EGF, ампліфікують методом ПЛР у присутності бетаїну (1-2 M) і DMSO (5 %). Синтезовані методом ПЛР фрагмент ДНК Notch3-LD (~0,8 т.о.) і фрагмент ДНК повтору Notch3-EGF (~4 т.о.) клонують у векторах експресії, що містять His-γ1Fc, у комерційно доступному векторі pSec або в комерційно доступному векторі pCD3.1, що несуть різні антибіотичні маркери. У результаті клонування одержують дві плазміди експресії, одна з яких експресує гібридний білок Notch3-LD/Fc, а інша експресує гібридний білок Notch3-EGF/Fc. Щоб полегшити конструювання плазміди і збільшити експресію різних рекомбінантних білків Notch 3, одержують олігонуклеотиди, що відповідають лідерній пептидній послідовності, що містить перші 135 пари основ нуклеотидної послідовності, що кодує Notch3. Одержані олігонуклеотиди містять деякі зміни в положеннях неоднозначного кодування, що зводяться до зменшення вмісту GC. Усі зміни в нуклеотидних послідовностях є мовчазними, тобто, не призводять до змін амінокислотних послідовностей (фіг.8A і 8B). Після спільного відпалу олігонуклеотидів рекомбінантну послідовність, що кодує лідерний пептид, з'єднують із залишком кодуючої послідовності, методом PCR-SOE (Ho, et al., Gene 77:51 (1989); Horton, et al., BioTechniques 8:528 (1990)) (див. фіг.9). Одержану послідовність, що кодує лідерний пептид, використовують у конструкціях експресії Notch3-LD/Fc і Notch3. Таким чином, обидва гібридних білка Fc містять сигнальний пептид, зв'язаний з N-кінцем, людську послідовність γ1Fc, з'єднану з C-кінцем. Амінокислотна послідовність Notch3-LD, що включає в себе лідерний пептид, наведена на фіг.8B і в SEQ ID NO: 6. Експресію гібридних білків Notch3-EGF/Fc і Notch3-LD/Fc підтверджують шляхом транзиторної трансфекції плазмід експресії Notch3 у клітини 293T (ATCC № CRL-11268, Manassas, VA) і CHO (Invitrogen, Carlsbad, CA), відповідно. Перед трансфекцією клітини культивують у поживному DMEM середовищі (Invitrogen, Carlsbad, CA), що містить 10 % фетальної телячої сироватки γFcS), 2 мM глутаміну і 1 розчин ессенціальних амінокислот, 27 UA 102061 C2 5 5 10 15 20 25 30 35 40 45 50 55 60 потім висівають у кількості приблизно 3-510 клітин на ямку в 6-ямковий планшет і вирощують протягом приблизно 24 годин. Три мікрограми кожної плазміди експресії, що несе гібридний білок Notch3, трансфікують у клітини кожної ямки, використовуючи систему для трансфекції Lipofectamine 2000 (Invitrogen, Carlsbad, CA) відповідно до інструкцій виробника. Після трансфекції клітини культивують у свіжому поживному середовищі в інкубаторі CO 2 протягом приблизно 40-48 годин, і потім аналізують експресію гібридного білка Notch3. Альтернативно, після трансфекції клітини культивують у поживному у середовищі протягом 3-4 годин, після чого середовище змінюють на DMEM, що містить 2 % FCS, і культивують протягом приблизно 60-66 годин, після чого кондиціоноване середовище відокремлюють і аналізують на секретований білок. Стабільні клітинні лінії одержують як для Notch3-LD/Fc (вектор His-Fcγ/pSec), так і для Notch3-EGF/Fc (вектор His-Fcγ/pSec). Кожну плазміду трансфікують у клітини CHO. Після трансфекції клітини культивують у поживному DMEM середовищі протягом ночі, потім середовище змінюють на середовище, що містить 800 мкг/мл гігроміцину, і культивують протягом щонайменше двох тижнів, поки клітини, що не містять плазміди експресії Notch3, не загинуть під дією антибіотиків. Кондиціоноване середовище стабільних клітинних ліній аналізують методом вестерн-блоттинга. Стабільно або транзиторно трансфіковані клітини аналізують на експресію і секрецію гібридного білка Notch3-LD/Fc або Notch3-EGF/Fc. Трансфіковані клітини, зібрані з чашок для культивування, промивають один раз фізіологічним розчином, забуференим фосфатом (PBS), ресуспендують у деіонізованій воді, змішують з рівним обсягом 2 буфера для завантаження білкового зразка (BioRad, Hercules, CA) і потім нагрівають приблизно при 100C протягом 10 хвилин. Секретований білок аналізують, використовуючи кондиціоноване середовище, змішане з рівним обсягом 2 буфера для завантаження білкового зразка, і нагрівають при 100C протягом 10 хвилин. Зразки розділяють методом SDS-PAGE, використовуючи 4-15 % градієнт. Білки переносять з гелю на мембрану PVDF (BioRad, Hercules, CA), що блокують 5 % знежиреним сухим молоком у PBST (PBS, що містить 0,05 % TWEEN-20®) протягом щонайменше однієї години перед переносом білка. Гібридні білки Notch3-EGF/Fc і Notch3-LD/Fc детектують шляхом інкубації з γFc-специфічним HRP-кон'югованим антитілом (Sigma, St Louis, MO) у блокуючому буфері, протягом однієї години при кімнатній температурі. Мембрану промивають три рази PBST і виявляють, використовуючи хемілюмінісцентний субстрат. Щоб очистити гібридний білок Notch3 домен/Fc, стабільні клітинні лінії CHO, як описано вище, культивують у DMEM, що містить 2 % FCS, протягом не більш 5 днів. Збирають один літр кондиціонованого середовища і піддають його афінній хроматографії на колонці, що містить гранули з іммобілізованим білком А. Колонку промивають PBS, зв'язані білки елююють 50 мм цитратним буфером (pН 2,8), і pН доводять до нейтральних значень шляхом додавання 1 M буфера Tris-HCl (pН 8). Чистоту білка визначають шляхом гель-аналізу білка методом SDSPAGE з використанням 4-15 % градієнта. Концентрацію білка визначають за допомогою Кумассі синього відповідно до інструкцій виробника (Pierce, Rockford, IL). За допомогою даного способу одержують міліграмові кількості очищеного білка Notch3-LD/Fc і Notch3-EGF/Fc, що використовують для імунізації й аналізів зв'язування методом ELISA. Приклад 2: Одержання Mab проти NOTCH3 Самцям мишей A/J (Harlan, Houston, TX) віком 8-12 тижнів підшкірно вводять 25 мкг Notch3EGF/Fc або Notch3-LD/Fc у повному ад'юванті Фрейнда (Difco Laboratories, Detroit, MI) у 200 мкл PBS. Через два тижні після ін'єкцій і за три дні до умертвіння мишам знову вводять внутрішньочеревинно 25 мкг такого ж антигена в PBS. У випадку кожного гібрида із селезінки імунізованих мишей одержують суспензії окремих клітин, що використовують для гібридизації з 8 8 клітинами мієломи Sp2/0; 510 клітин Sp2/0 і 510 клітин селезінки гібридизують у середовищі, що містить 50 % поліетиленгліколю (МВ 1450) (Kodak, Rochester, NY) і 5 % диметилсульфоксиду 5 (Sigma, St. Louis, MO). Потім концентрацію клітин доводять до 1,510 клітин селезінки на 200 мкл суспензії в середовищі Iscove (Invitrogen, Carlsbad, CA), доповненої 10 % фетальною бичачою сироваткою, 100 одиниць/мл пеніциліну, 100 мкг/мл стрептоміцину, 0,1 мкM гіпоксантину, 0,4 мкM аміноптерину і 16 мкМ тимідину. У кожну ямку приблизно шістдесятьох 96ямкових планшетів додають двісті мікролітрів клітинної суспензії. Приблизно через десять днів культуральні супернатанти відокремлюють і аналізують на антитіло-зв'язувальну активність методом ELISA. 96-ямкові плоскодонні планшети для мікроаналізу Immulon II (Dynatech, Laboratories, Chantilly, VA) покривають, використовуючи 100 мкл Notch3-EGF/Fc або Notch3-LD/Fc (0,1 мкг/мл) у (PBS), що містять 1 феноловий червоний і 3-4 краплі pHix/літр (Pierce, Rockford, IL), і 28
ДивитисяДодаткова інформація
Назва патенту англійськоюAnti-notch3 agonist antibodies and their use in the treatment of notch3-related diseases
Автори англійськоюLi, Kang, Zhou, Bin-bing Stephen, Wu, Wenjuan, Fung, Sek Chung, Singh, Sanjaya
Назва патенту російськоюАгонистическое антитело к notch3 и его применение для лечения notch3- ассоциированных заболеваний
Автори російськоюЛи Кан, Чжоу Бинь-бин Стивен, Ву Веньцзюань, Фун Сек Чун, Сингх Санджайа
МПК / Мітки
МПК: A61P 3/06, C07K 14/43, A61P 3/04, A61K 31/717, A61P 3/10
Мітки: застосування, лікування, notch3, notcн3-асоційованих, антитіло, захворювань, агоністичне
Код посилання
<a href="https://ua.patents.su/112-102061-agonistichne-antitilo-proti-notch3-i-jjogo-zastosuvannya-dlya-likuvannya-notcn3-asocijjovanikh-zakhvoryuvan.html" target="_blank" rel="follow" title="База патентів України">Агоністичне антитіло проти notch3 і його застосування для лікування notcн3-асоційованих захворювань</a>
Антагоністичні антитіла до notch3 та їх застосування для профілактики і лікування пов’язаних з notch3 захворювань

Номер патенту: 98632
Опубліковано: 11.06.2012
Автори: Чжоу Бінь-бін Стівен, Лі Юйчен, Фун Сек Чун, Лі Кан, Сінгх Санджая
МПК: A61K 39/395, A61P 35/00, C07K 16/28, C12N 15/13, C12N 5/12
Мітки: пов'язаних, notch3, антитіла, антагоністичні, застосування, захворювань, профілактики, лікування
Формула / Реферат:
1. Моноклональне антитіло, яке специфічно зв'язується з Notch3, причому антитіло специфічно зв'язується з конформаційним епітопом фрагмента Notch3, що складається з амінокислот 1378-1640 SEQ ID NO: 1, де антитіло інгібує передачу сигналу Notch3.2. Антитіло за п. 1, де антитіло зв'язується з амінокислотними залишками в домені LIN12 (SEQ ID NO: 9) і димеризаційному домені (SEQ ID NO: 18).3. Антитіло за п. 1, де антитіло містить...
Химерне антитіло проти cd40 людини, молекула нуклеїнової кислоти, вектор експресії, фармацевтична композиція для лікування захворювань, опосередкованих т-клітинами, яка містить химерне антитіло

Номер патенту: 71909
Опубліковано: 17.01.2005
Автори: Сайдек Ентоні В., Холленбау Даєн, Беррі Карен К., Бейорет Юрген, Уоткінс Джеффрі Д., Хьюз Уільям Д., Ву Херрен, Херріс Лінда Дж., Торн Барбара А., Аруффо Аледжандро А.
МПК: A61P 37/02, C07K 14/46, A61K 39/395, C07K 14/725, C12N 15/13, A61P 29/00, A61P 19/02, A61P 37/06, C12N 15/09, C12N 1/21, C07K 16/28, A61P 43/00
Мітки: експресії, химерне, містить, композиція, захворювань, фармацевтична, молекула, вектор, яка, т-клітинами, опосередкованих, людини, антитіло, нуклеїнової, лікування, кислоти
Формула / Реферат:
1. Варіабельна ділянка легкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:1 (Фіг. 4а).2. Варіабельна ділянка важкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:2 (Фіг. 4b).3. Химерне антитіло, що зв'язується з CD40 людини, яке містить легкий і важкий ланцюг, причому зазначений легкий ланцюг містить ...
Антитіло проти ngf і спосіб його застосування

Номер патенту: 92449
Опубліковано: 10.11.2010
Автори: Розенталь Арнон, Шелтон Девід Л., Понз Хауме
МПК: C07K 19/00, A61K 39/395, A61P 37/00, C07K 16/22
Мітки: застосування, антитіло, спосіб
Формула / Реферат:
1. Антитіло проти фактора росту нервів (NGF) або його фрагмент, що містять(a) варіабельну область важкого ланцюга, що включає:(i) ділянку CDR1 послідовності SEQ ID NO: 30;(ii) ділянку CDR2 послідовності SEQ ID NO: 31;(iii) ділянку CDR3, вибрану із групи, що складається з послідовностей SEQ ID NO: 5, 58, 60, 62 і 64; і(b) варіабельну область легкого ланцюга, що включає:(i) ділянку CDR1 послідовності...
Антитіло зі специфічною афінністю до характерної антигенної детермінанти ed-b домену фібронектину, його застосування для лікування ангіогенезу, кон’югати та діагностичний набір, до складу якого входить вказане

Номер патенту: 74134
Опубліковано: 15.11.2005
Автори: Бірхлер Манфред, Віті Франческа, Нері Даріо, Тарлі Лоренцо
МПК: A61K 47/48, A61K 49/00, C12N 15/09, C07K 16/18
Мітки: характерної, фібронектину, домену, набір, антитіло, лікування, афінністю, специфічною, складу, застосування, ангіогенезу, антигенної, входить, кон'югати, детермінанти, діагностичний, якого, вказане
Формула / Реферат:
1. Антитіло зі специфічною афінністю до характерної антигенної детермінанти ED-B домену фібронектину, де згадане антитіло має афінність до згаданої ED-B антигенної детермінанти у субнаномолярному діапазоні.2. Антитіло за п. 1, яке розпізнає ED-B(+) фібронектин.3. Антитіло за п. 1, яке має scFv формат.4. Антитіло за п. 3, яке є рекомбінантним.5. Антитіло за п. 3, де згадана афінність поліпшена шляхом індукування...
Антитіло проти гепсидину та його застосування

Номер патенту: 98983
Опубліковано: 10.07.2012
Автори: Льюнг Донмайєнн Дон, Уітчер Деррік Райан, Гейтлі Денніс Патрік, Тань Їнь, Луань Пен, Свенсон Барбара Енн, Хе Лухун, Цай Юйпін Ентоні
МПК: A61K 39/395, C07K 16/18, G01N 33/577, C12N 15/13, C12N 5/10, A61P 7/06
Мітки: антитіло, гепсидину, застосування
Формула / Реферат:
1. Ізольоване антитіло, яке зв'язує людський гепсидин-25 зі зв'язувальною спорідненістю KD приблизно 800 пМ або менше, яку визначають поверхневим плазмонним резонансом (SPR) при температурі 25 °C, і містить шість CDR, вибраних із групи, яку складають:(і) LCDR1, LCDR2, LCDR3, HCDR1, HCDR2 та HCDR3, які мають амінокислотні послідовності, представлені послідовностями SEQ ID NO:52, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:65, SEQ ID...
Попередній патент: Загородження
Наступний патент: Спосіб деемульсації рідких водонафтових шламових систем
Випадковий патент: Пристрій для вирощування кристалів із розплаву