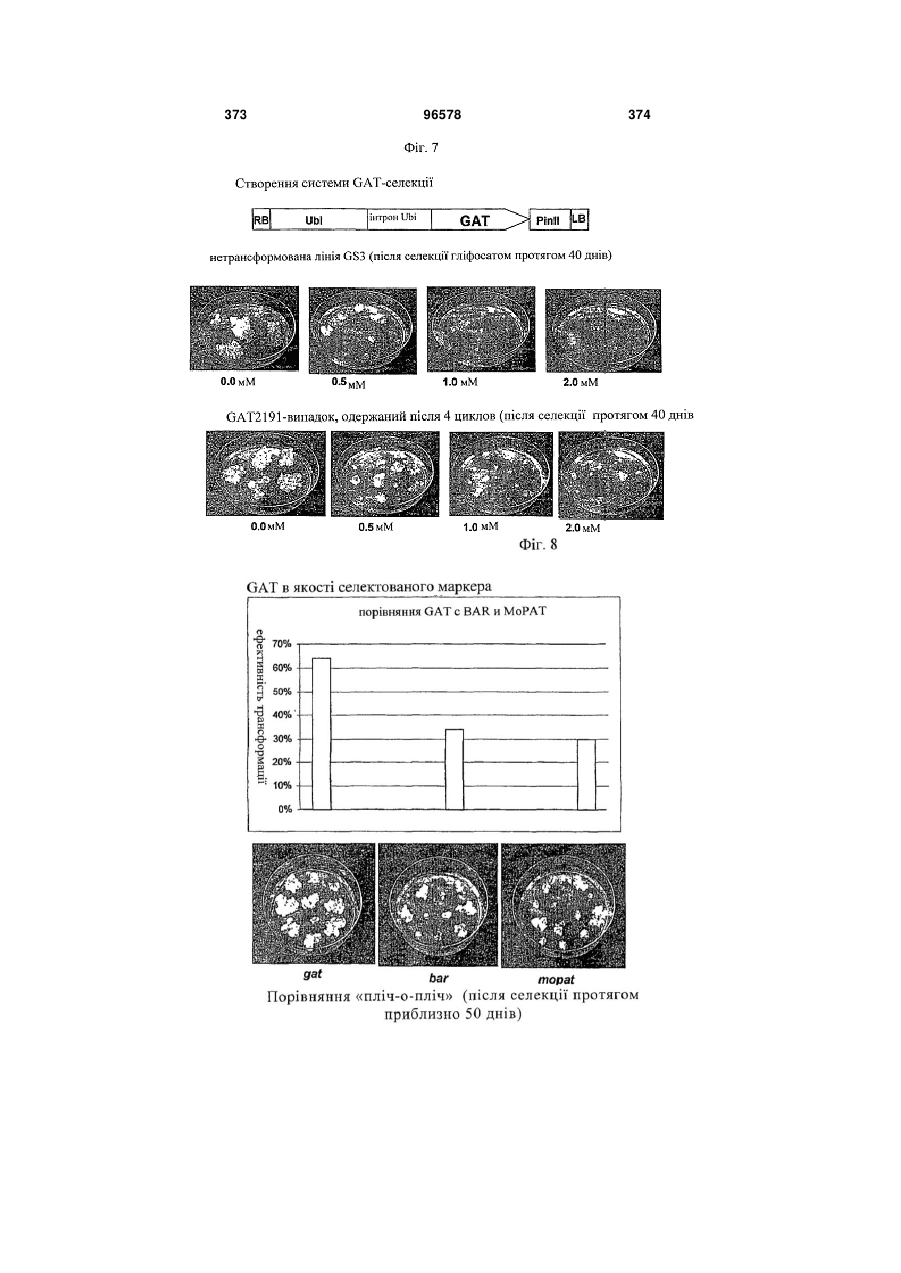
Однодольна трансгенна рослина, що містить полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату
Номер патенту: 96578
Опубліковано: 25.11.2011
Автори: Вон Джеймс Ф. Х., Касл Лінда А., Маккатчен Біллі Фред, Хатчісон Джеймс М., Чікойн Тімоті К., Вогт Марк Д., Вілкінсон Джек К., Хейзел Крістін Б., Лю Дунлун, Хекерт Мет'ю Дж., Олсон Пол Д., Хегстад Джеффрі М., Мере Вейн Дж., Чо Хьон-же, Сондерз Девід В., Грін Джеррі М., Лу Алберт Л., Гайда Ентоні Д., Джр., Мой Йорк, Клаус Джон С., Піплз Кеннет А.







































































































































































Формула / Реферат
1. Однодольна трансгенна рослина, що містить полінуклеотид, який кодує поліпептид гліфосат-N-ацетилтрансферазу, де полінуклеотид функціонально зв'язаний з
(а) промотором, який забезпечує експресію в рослині; та
(б) принаймні однією копією енхансерної послідовності, що включає
(і) нуклеотидну послідовність, представлену в SEQ ID NO: 72 або 85, або
(іі) нуклеотидну послідовність, яка принаймні на 90% ідентична послідовності, що представлена в SEQ ID NO: 72 або 85, де енхансерна послідовність модулює рівень транскрипції,
де енхансер є гетерологічним щодо промотору; та
де однодольна рослина має толерантність до гліфосату при його застосуванні в нормі витрати, ефективній для інгібування росту контрольної рослини, яка не містить полінуклеотид, що кодує гліфосат-N-ацетилтрансферазу.
2. Однодольна трансгенна рослина за п. 1, де зазначена енхансерна послідовність включає SEQ ID NO: 88 або 89.
3. Однодольна трансгенна рослина за п. 1, де однодольна рослина додатково містить полінуклеотид, який кодує толерантний до ALS поліпептид .
4. Однодольна трансгенна рослина за п. 3, у якій толерантний до інгібітору ALS поліпептид включає високостійку алель ацетолактатсинтази (HRA).
5. Однодольна трансгенна рослина за будь-яким із пп. 1-4, де полінуклеотид функціонально зв'язаний принаймні з трьома копіями енхансерної послідовності, що представлена в SEQ ID NO:72.
6. Однодольна трансгенна рослина за будь-яким із пп. 1-4, де промотор являє собою промотор убікітину або ацетолактатсинтази.
7. Однодольна трансгенна рослина за будь-яким із пп. 1-5, де енхансерна послідовність включає послідовність, яка принаймні на 90 % ідентична послідовності, що представлена в SEQ ID NO: 88 або 89, та енхансерна послідовність модулює рівень транскрипції.
8. Однодольна трансгенна рослина за будь-яким із пп. 1-7, де промотор являє собою промотор убікітину кукурудзи або промотор убікітину Arabidopsis.
9. Однодольна трансгенна рослина за будь-яким із пп. 1-8, де однодольна рослина включає конструкцію ДНК, що включає в 5’ до 3’ або 3’ до 5’ напрямку: полінуклеотид, що кодує поліпептид гліфосат-N-ацетилтрансферазу, який обумовлює толерантність до гліфосату, функціонально зв’язаний з промотором, який функціонально зв’язаний принаймні з однією копією енхансерної послідовності, де енхансерні послідовності функціонально зв’язані з другим промотором, функціонально зв’язаним з полінуклеотидом, який кодує толерантний до інгібітору ALS поліпептид, та полінуклеотиди експресуються у дивергуючому напрямку.
10. Однодольна трансгенна рослина за будь-яким із пп. 1-9, де зазначений поліпептид гліфосат-N-ацетилтрансфераза включає послідовність, представлену в SEQ ID NO: 45.
11. Трансгенна однодольна рослина за будь-яким із пп. 1-10, де однодольна рослина являє собою кукурудзу.
12. Трансгенна однодольна рослина за будь-яким із пп. 1-10, де однодольна рослина являє собою пшеницю, рис, ячмінь, сорго, цукровий очерет, просо прутоподібне або жито.
13. Трансгенна однодольна рослина за будь-яким із пп. 1-12, де копії енхансерів безпосередньо примикають одна до іншої.
14. Трансгенна однодольна рослина за будь-яким із пп. 1-13, де принаймні один із енхансерів розташований у прямій або зворотній орієнтації щодо промотору.
15. Трансгенна однодольна рослина за будь-яким із пп. 1-14, де поліпептид гліфосат-N-ацетилтрансфераза включає послідовність, яка принаймні на 90 % ідентична послідовності, що представлена в SEQ ID NO: 45, де ця послідовність має гліфосат-N-ацетилтрансферазну активність.
16. Трансгенне насіння однодольної рослини за будь-яким із пп. 1-15.
Текст
1. Однодольна трансгенна рослина, що містить полінуклеотид, який кодує поліпептид гліфосат-N-ацетилтрансферазу, де полінуклеотид функціонально зв'язаний з (а) промотором, який забезпечує експресію в рослині; та (б) принаймні однією копією енхансерної послідовності, що включає (і) нуклеотидну послідовність, представлену в SEQ ID NO: 72 або 85, або (іі) нуклеотидну послідовність, яка принаймні на 90% ідентична послідовності, що представлена в 2 (19) 1 3 96578 4 10. Однодольна трансгенна рослина за будь-яким із пп. 1-9, де зазначений поліпептид гліфосат-Nацетилтрансфераза включає послідовність, представлену в SEQ ID NO: 45. 11. Трансгенна однодольна рослина за будь-яким із пп. 1-10, де однодольна рослина являє собою кукурудзу. 12. Трансгенна однодольна рослина за будь-яким із пп. 1-10, де однодольна рослина являє собою пшеницю, рис, ячмінь, сорго, цукровий очерет, просо прутоподібне або жито. 13. Трансгенна однодольна рослина за будь-яким із пп. 1-12, де копії енхансерів безпосередньо примикають одна до іншої. 14. Трансгенна однодольна рослина за будь-яким із пп. 1-13, де принаймні один із енхансерів розташований у прямій або зворотній орієнтації щодо промотору. 15. Трансгенна однодольна рослина за будь-яким із пп. 1-14, де поліпептид гліфосат-Nацетилтрансфераза включає послідовність, яка принаймні на 90 % ідентична послідовності, що представлена в SEQ ID NO: 45, де ця послідовність має гліфосат-N-ацетилтрансферазну активність. 16. Трансгенне насіння однодольної рослини за будь-яким із пп. 1-15. Галузь техніки, до якої відноситься винахід Даний винахід відноситься до галузі молекулярної біології. Більш конкретно даний винахід відноситься до толерантності до декількох гербіцидів, пов'язаної з експресією послідовності, що обумовлює толерантність до гліфосату, у сполученні з експресією, щонайменше одного гена, що обумовлює толерантність до іншого гербіциду. Передумови створення винаходу При комерційному виробництві сільськогосподарських культур необхідно легко й швидко знищувати небажану рослинність (тобто «бур'янисту рослинність», «бур'яни») у посівах культурних рослин. Ідеальна обробка має на увазі обробку, якій можна піддавати все поле, але в результаті якої знищується тільки небажана рослинність, а культурні рослини залишаються неушкодженими. Одна з таких систем обробки включає застосування культурних рослин, толерантних до гербіциду, у результаті чого при обприскуванні гербіцидом поля толерантних до гербіциду культурних рослин, культурні рослини можуть продовжувати активно розростатися, у той час як бур'яни, що не володіють толерантністю до гербіциду, гинуть або одержують серйозні ушкодження. В ідеальному варіанті такі системи обробки повинні мати переваги, що полягають у тому, що вони дозволяють варіювати гербіцидні властивості, у результаті чого боротьба з бур'янами може найбільше ефективно забезпечувати сполучення гнучкості й економічності. Наприклад, різні гербіциди мають різну тривалість життя в польових умовах, і деякі гербіциди зберігаються й мають ефективність протягом щодо тривалих періодів часу після їхнього внесення в поле, у той час як інші гербіциди швидко розкладаються, перетворюючись в інші й/або неактивні сполуки. Ідеальна система обробки повинна дозволяти застосовувати різні гербіциди, так, щоб фермери могли здійснювати вибір гербіцидів для конкретної ситуації. Культурним рослинам можна надавати толерантність до специфічних гербіцидів шляхом створення генів у культурних рослинах, які кодують відповідні ферменти, що беруть участь у метаболізмі гербіцидів, і/або нечутливі до гербіцидів мішені. У деяких випадках ці ферменти й кодуючі їх нуклеїнові кислоти присутні в рослині. В інших ви падках їх одержують із інших організмів, таких як мікроби (див., наприклад, Padgette і ін., «New weed control opportunities: Development of soybeans with a Roundup Ready gene», 1996, і Vasil «Phosphinothricin-resistant crops», 1996, обидві статті опубліковані в Herbicide-Resistant Crops, під ред. Duke, з CRC Press, Boca Raton, Florida, cc. 5484 і cc. 85-91). До теперішнього часу створені трансгенні рослини, які експресують ряд генів, що обумовлюють толерантність до гербіцидів, які одержували з різних організмів, включаючи ген, що кодує хімерний білок щурячого цитохрома Р4507А1 і НАДФ·Н-цитохром Р450-оксиредуктазу дріжджів (Shiota і ін., Plant Physiol 106, 1994, с. 17). До інших генів, які обумовлюють толерантність до гербіцидів, відносяться ген синтази ацетоацетогідрокислоти («AHAS»), мутації в нативній послідовності якого, як було встановлено, надають експресуючим його рослинам стійкість до декількох типів гербіцидів, і який інтродукували в деякі види рослин (див., наприклад, Hattori і ін., Mol. Gen. Genet. 246, 1995, с. 419); гени глутатіонредуктази й супероксиддесмутази (Аоnо і ін., Plant Cell Physiol. 36, 1995, с. 1687); і гени різних фосфотрансфераз (Datta і ін., Plant Mol. Biol. 20, 1992, с. 619). Одним з дуже добре вивчених гербіцидів є Nфосфометилгліцин, що має загальноприйняту назва гліфосат. Гліфосат являє собою гербіцид широкого спектра дії, що знищує як широколисті, так і трав'янисті рослини, у результаті інгібування ферменту 5-єнолпіруватшикимат-3-фосфат-синтази (яку означають також як «EPSP-синтаза» або «EPSPS»), цей фермент бере участь у шляху біосинтезу, що приводить до утворення ароматичних амінокислот, гормонів і вітамінів. Були створені стійкі до гліфосату трансгенні рослини, які мають комерційно значимий рівень стійкості до гліфосату внаслідок інтродукції в них модифікованої EPSPS Agrobacterium CP4. Мішенню цього модифікованого ферменту є хлоропласт, де в присутності гліфосату триває синтез EPSP з фосфоєнолпіровиноградної кислоти («PEP») і шикимат-3-фосфату. Стійкі до гліфосату трансгенні рослини сої лінії СР4 у цей час знайшли комерційне застосування (наприклад, надходять у продаж від фірми Monsanto за назвою «Roundup Ready»). 5 Іншими гербіцидами, що представляють інтерес для комерційного виробництва культурних рослин, є гліфосинат (фосфінотрицин) і хімічні сполуки, механізм дії яких пов'язаний з інгібуванням ацетолактатсинтази (ALS), такі як гербіциди із класу сульфонілсечовин (сульфонілсечовинні гербіциди). Глуфосинат являє собою гербіцид широко спектра дії, що впливає на присутній у хлоропластах фермент глутаматсинтазу. Були отримані толерантні до глуфосинату трансгенні рослини, які несли ген bar з Streptomyces hygroscopicus. Фермент, кодуємий геном bar, володіє Nацетилюючою активністю й здатністю модифікувати детоксифікацію глуфосинату. Стійкі до глуфосинату рослини в цей час надходять на ринок (наприклад, від фірми Bayer за назвою «Liberty Link»). Сульфонілсечовинні гербіциди інгібують ріст вищих рослин шляхом блокади ацетолактатсинтази (ALS). Рослини, що несуть конкретні мутації в ALS, мають толерантність до гербіцидів, мішенню яких є ALS, у тому числі до сульфонілсечовинам. Так, наприклад, сульфонілсечовинні гербіциди, такі як синхрони (суміш хлоримурон-етилу й тифенсульфурон-метилу), можна застосовувати в сполученні з рослинами, толерантними до гербіцидів, мішенню яких є ALS, такими як соя сорту STS (толерантна до синхрони соя), що містить ознаку, що підсилює природну толерантність сої до сульфонілсечовинних гербіцидів, застосовуваних у посівах сої. Хоча в цей час на ринок надходить ряд толерантних до гербіцидів культурних рослин, одна із проблем, що виникають для багатьох вступників у продаж гербіцидів і комбінацій гербіцид/культурна рослина, полягає в тому, що конкретні гербіциди, як правило, мають неповний спектр активності відносно звичайних видів бур'янів. У постійно зростаючої популяції видів і біотипів бур'янів розвивається стійкість відносно більшості конкретних гербіцидів, які застосовувалися протягом деякого періоду часу, (див., наприклад, Tranel і Wright Weed Science 50, 2002, cc. 700-712; Owen i Zelaya, Pest Manag. Sci. 61, 2005, cc. 301-311). Описані трансгенні рослини, що володіють стійкістю більш ніж до одного гербіциду (див., наприклад, WO 2005/012515). Однак продовжує зберігатися потреба в поліпшенні всіх аспектів оброблення культурних рослин, вибору або заміни системи боротьби з бур'янами, подовженні залишкової дії гербіцидів для бур'янів і підвищенні врожайності культурних рослин. В частковості через локальні і регіональні варіації домінантних видів бур'янів, а також кращих видів культурних рослин, продовжує зберігатися необхідність у змінених системах захисту культурних рослин і регуляції чисельності бур'янів, які можна адаптувати до умов конкретного регіону, географічної зони й/або місцевості. Наприклад, продовжує зберігатися необхідність у розробці способів захисту культурних рослин і регуляції чисельності бур'янів, які дозволяють знижувати: кількість гербіцидних обробок, необхідних для боротьби з бур'янами в польових умовах; кількість гербіциду, необхідну для боротьби з бур'янами в польових умовах; кількість обробок ґрунту, необ 96578 6 хідну для вирощування культурних рослин; і/або в розробці програм, які сповільнюють або запобігають розвиток і/або появі стійких до гербіцидів бур'янів. Продовжує зберігатися необхідність у розробці способів захисту культурних рослин і регуляції чисельності бур'янів, які забезпечують спрямоване застосування конкретних комбінацій гербіцидів. Короткий виклад сутності винаходу У даному винаході запропоновані способи й композиції, призначені для одержання поліпшених рослин, які мають толерантність більш ніж до одного гербіциду або класу або підкласу гербіцидів Винахід відноситься до композицій, призначених для одержання рослин, які мають толерантність до гліфосату, а також щонайменше ще до одного іншого гербіциду або класу або підкласу гербіцидів, а також до способів їхнього застосування. Винахід відноситься також до інших композицій, призначених для одержання рослин, які містять полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату, і полінуклеотид, що кодує поліпептид, толерантний до інгібітору ALS. Один з не обмежуючих об'єм винаходу варіантів здійснення винаходу відноситься до композицій, призначених для одержання рослин, які експресують полінуклеотид, що кодує поліпептид GAT (гліфосат-N-ацетилтрансфераза) і мають толерантність щонайменше ще до одного іншого гербіциду. У деяких варіантах здійснення винаходу рослина, пропонована у винаході, експресує поліпептид GAT і поліпептид HRA. У винаході запропоновані способи боротьби з бур'янами на оброблюваних посівних площах із застосуванням рослин, пропонованих у винаході. Запропоновані також удосконалені способи трансформації. Короткий опис креслень На кресленнях показано: на Фіг. 1 - приклади конструкцій, що несуть енхансерні елементи 35S; на Фіг. 2 - схема, що ілюструє впливи енхансерів 35S на ефективність ТХ; на Фіг. 3 - схема, що ілюструє впливи енхансерів 35S на ефективність Т0; на Фіг. 4 - схема, що ілюструє впливи енхансерів 35S на кількість копій; на Фіг. 5 - таблиця, у якій представлені дані про вплив енхансерів 35S на ефективність Т2; на Фіг. 6 - аналіз для оцінки «інсектицидного» гена; на Фіг. 7 - схематичне зображення розробки системи для селекції з використанням GAT; на Фіг. 8 - дані, що демонструють, що GAT можна застосовувати в якості селетуємого маркера; на Фіг. 9 - схема, що ілюструє ефективність трансформації за допомогою GAT. Докладний опис винаходу У даному винаході запропоновані способи й композиції, призначені для одержання й застосування рослини, толерантної більш ніж до одного гербіциду або класу або підкласу гербіцидів. У деяких з варіантів здійснення винаходу рослина має толерантність як до гліфосату, так і щонайменше до одного іншого гербіциду (або класу або підкласу гербіцидів) або іншому хімічному агентові 7 (або класу або підкласу інших хімічних агентів). Такі рослини знайшли застосування, наприклад, у способах вирощування культурних рослин, що включають обробку декількома гербіцидами. Так, у винаході запропоновані поліпшені рослини, які переносять обробку гербіцидом або комбінацією гербіцидів (включаючи комбінацію гербіцидів з різними механізмами дії; тобто обробку сумішами, що володіють 2, 3, 4 або більшою кількістю механізмів гербіцидної дії), або комбінацією щонайменше одного гербіциду й щонайменше одного іншого хімічного агента, включаючи фунгіциди, інсектициди, регулятори росту рослин і т.п. Таким чином, у винаході запропоновані поліпшені способи вирощування культурних рослин, у посівах яких здійснюють вибіркове знищення бур'янів. В одному з варіантів здійснення винаходу рослини, пропоновані у винаході, містять полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату, і полінуклеотид, що кодує поліпептид, що має толерантність до інгібітору ALS. Як більш докладно описано нижче, такі рослини в контексті даного опису означають як «рослини, толерантні до гліфосату/інгібітору ALS». Рослини, пропоновані у винаході, мають модифіковану толерантність до гербіцидів і внаслідок цього в їхній присутності можна застосовувати гербіциди в нормах витрати, які повинні викликати значне ушкодження рослин, і можна також застосовувати суміші гербіцидів у більше низьких в порівнянні із загальноприйнятім! концентраціями, але які усе ще зберігають здатність до вибіркового знищення бур'янів. Крім того, рослини, толерантні до гліфосату/інгібітору ALS, пропоновані у винаході, можна застосовувати в сполученні з технологією, заснованою на застосуванні сумішей гербіцидів, і тим самим робити застосування хімічних пестицидів більше зручним, економічним й ефективним для виробника. У випадку культурних рослин, толерантних до гліфосату/інгібітору ALS, технологія, заснована на застосуванні сумішей, повинна спрощувати готування препаративних форм засобів захисту культурних рослин, наприклад, гербіцидів, мішенню яких є ALS, у формі сухих гранул, що дозволяє поставляти адаптовані суміші, створені для рішення конкретних проблем, у сполученні з культурними рослинами, толерантними до гліфосату/інгібітору ALS, пропонованими у винаході. У результаті додання вираженої толерантності до інгібітору ALS рослинам, пропонованим у винаході, це дозволяє застосовувати гербіциди, мішенню яких є ALS, оскільки відповідь культурних рослин на гербіциди знижується до мінімуму. Розробки таких гербіцидів, що володіють унікальною виборковою у сполученні з культурними рослинами, толерантними до гліфосату/інгібітору ALS, пропонованими в даному описі, у цей час можна здійснювати й адаптувати для задоволення постійно мінливих потреб у боротьбі з бур'янами. Ці досягнення дозволяють у цей час створювати величезну кількість сумішей гербіцидів, включаючи, наприклад, суміші інгібіторів ALS, які можна адаптувати для вдосконалення системи контролю чисельності бур'янів (оскільки хімічні агенти, що представляють собою інгібітори ALS, мають різні 96578 8 гербіцидні властивості), включаючи розширення спектра бур'янів, здатність до забезпечення специфічної залишкової дії, використання другого механізму дії для подолання або вповільнення розвитку стійкості бур'янів (додаткове застосування гліфосату, глуфосинату або т.п.), а також нові форми, які можна створювати або для передвсходового, або для післявсходового внесення або для їх обох. Використання сумішей дозволяє також додавати або змішувати в одному резервуарі інші агрохімічні агенти (агрохімікати, хімічні засоби захисту рослин) у загальноприйнятих, зазначених на етикетах нормах витрати, такі як додаткові гербіциди, що володіють 3-їм або 4-им механізмом дії, для заповнення «проломів» у спектрі дії, або навіть дозволяють включати фунгіциди, інсектициди, регулятори росту рослин і т.п., знижуючи тим самим вартість, пов'язану з додатковими обробками. Як буде більш докладно описано нижче, способи, пропоновані у винаході, можна адаптувати для конкретної області або регіону. Запропоновані також удосконалені способи трансформації. І. Рослини, толерантні до гліфосату/інгібітору ALS а. Толерантність до гліфосату У винаході запропоновані рослини, які містять полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату, і полінуклеотид, що кодує поліпептид, толерантний до інгібітору ALS. У способах і композиціях, пропонованих у винаході, можна застосовувати різні послідовності, що обумовлюють толерантність до гліфосату. В одному з варіантів здійснення винаходу механізм стійкості до гліфосату обумовлений експресією полінуклеотиду, що володіє трансферазною активністю. У контексті даного опису поліпептид з «трансферазною» активністю має здатність переносити ацильну групу з ацетил-Co на N гліфосату, переносити пропіонільну групу пропіоніл-Со на N гліфосату або каталізувати ацетилування аналогів гліфосату й/або метаболітів гліфосату, наприклад, амінометилфосфонової кислоти. Методи аналізу зазначеної активності описані, наприклад, в опублікованому U.S. № 2003/0083480, опублікованому U.S. № 2004/0082770 і заявці на патент США № 10/835615, зареєстрованої 29 квітня 2004 p., WO 2005/012515, WO 2002/36782 і WO 2003/092360. В одному з варіантів здійснення винаходу поліпептид із трансферазною активністю являє собою поліпептид гліфосат-N-ацетилтрансферази «GAT». У контексті даного опису поліпептид або фермент GAT містить поліпептид, що володіє гліфосат-N-ацетилтрансферазною активністю («GAT»активність), тобто здатністю каталізувати ацетилування гліфосату. У конкретних варіантах здійснення винаходу поліпептид, що володіє гліфосат-Nацетилтрансферазною активністю, може переносити ацетильную групу з ацетил-Co на N гліфосат. Крім того, деякі поліпептиди GAT переносять пропіонільну групу пропіоніл-Со на N гліфосату. Деякі поліпептиди GAT володіють також здатністю каталізувати ацетилування аналогів гліфосату й/або метаболітів гліфосату, наприклад, амінометилфосфонової кислоти. Поліпептиди GAT характеризу 9 ються їхньою структурною подібністю один з одним, наприклад, з погляду подібності послідовностей, коли здійснюють порівняння структури поліпептидів GAT один з одним. Приклади поліпептидів GAT і кодуючих їх полінуклеотидів відомі в даній області й конкретно описані, наприклад, у заявці на патент США № 10/004357, зареєстрованої 29 жовтня 2001 p., заявці на патент США № 10/427692, зареєстрованої 30 квітня 2003 р. і в заявці на патент США № 10/835615, зареєстрованої 29 квітня 2004 p., кожна з яких повністю включена в даний опис як посилання. У деяких варіантах здійснення винаходу поліпептиди GAT, застосовувані для створення рослин, пропонованих у винаході, містять амінокислотну послідовність, представлену в: SEQ ID NO:5, 14, 11, 8, 21, 27, 17, 24, 30, 35, 46, 47, 48, 49, 50, 51, 52, 53, 39, 42, 45 або 54. Кожна із цих послідовностей описана також у заявці на патент США № 10/835615, зареєстрованої 29 квітня 2004 р. У деяких варіантах здійснення винаходу застосовують відповідні GAT-полінуклеотиди, які кодують зазначені поліпептиди; ці полінуклеотидні послідовності представлені в SEQ ID NO:3, 12, 9, 6, 19, 15, 25, 22, 28, 33,4, 7, 10,13, 16, 18, 20, 23, 26, 29, 32, 34, 36, 38, 41,44, 43, 56, 31, 37, 40, 57, 58, 59, 60, 61, 62, 63 або 64. Кожна із цих послідовностей описана також у заявці на патент США № 10/835615, зареєстрованої 29 квітня 2004 р. Як буде більш докладно описано нижче, застосування фрагментів або інших відомих варіантів GAT-полінуклеотидів, зумовлюючих толерантність до гербіцидів полінуклеотидів, підпадає також під об'єм даного винаходу. У конкретних варіантах здійснення винаходу рослини, толерантні до гліфосату/інгібітору ALS, пропоновані у винаході, експресують поліпептид GAT, тобто поліпептид, що володіє гліфосатФІацетилтрансферазною активністю, у результаті якої відбувається перенос ацетильной групи з ацетил-Co на N гліфосату. Так, рослини, пропоновані у винаході, оброблені гліфосатом, містять метаболіт гліфосату N-ацетилгліфосат («NAG»). Таким чином, винахід відноситься також до рослин, які містять NAG, а також до способу одержання NAG шляхом обробки рослин, які містять ген GAT (тобто які експресують поліпептид GAT), гліфосатом. Присутність N-ацетилгліфосату може служити діагностичним маркером присутності активного гена GAT у рослині і його можна виявляти методами, відомими в даній області, наприклад, за допомогою мас-спектрометрії або імуноаналіза. Як правило, рівень NAG у рослині, що містить ген GAT, що обробляли гліфосатом, корелює з активністю гена GAT і кількістю гліфосату, яким рослину обробляли. Рослини, пропоновані у винаході, можуть містити також декілька полінуклеотидів GAT (тобто, щонайменше 1, 2, 3, 4, 5, 6 або більше). Установлено, що, якщо застосовують кілька поліпептидів GAT, те полінуклеотиди GAT можуть кодувати поліпептиди GAT з різними кінетичними параметрами, тобто варіант GAT, що має більше низьке значення Km, можна поєднувати з варіантом, що має більше високе значення kcat. У деяких варіантах здійснення винаходу різні поліпептиди можна зли 96578 10 вати із транзитною послідовністю хлоропласта або з іншою сигнальною послідовністю, що забезпечує експресію поліпептидів у різних клітинних компартментах, органелах або секрецію одного або декількох поліпептидів. Поліпептид GAT, кодуємий полінуклеотидом GAT, може мати підвищену ферментативну активність у порівнянні з раніше ідентифікованими ферментами. Ферментативну активність можна характеризувати за допомогою звичайних кінетичних параметрів, таких як kcat, KМ і kcat/KМ. kcat можна розглядати як міру швидкості ацетилування, насамперед при високих концентраціях субстрату; KМ є мірою афінності ферменту GAT до його субстратів (наприклад, ацетил-Co, пропіоніл-Со і гліфосат); і kcat/KМ є мірою каталітичної ефективності, що враховує як афінність до субстрату, так і швидкість каталізу. kcat/KM є особливою важливою завбільшки ситуації, коли концентрація субстрату щонайменше частково є обмежуючим швидкість фактором. У цілому, GAT з більше високим значенням kcat або kcat/KM є більше ефективним каталізатором, чим інший фермент GAT з більше низьким значенням kcat або kcat/KМ. GAT з більше низьким значенням Км є більше ефективним каталізатором, чим інший GAT з більше високим значенням KМ. Так, для порівняння одного GAT з іншим з позицій ефективності можна порівнювати кінетичні параметри двох ферментів. Відносна важливість kcat, kcat/KМ і KМ повинна варіюватися залежно від контексту, у якому розглядається функція GAT, наприклад, залежно від очікуваної ефективної концентрації гліфосату відносно Км для гліфосату. Активність GAT можна характеризувати також з позицій кожної із численних функціональних характеристик, включаючи, але, не обмежуючись ними, стабільність, чутливість до інгібіторів або до активації іншими молекулами. Так, наприклад, поліпептид GAT може мати більше низьке значення KМ для гліфосату в порівнянні з раніше ідентифікованими ферментами, наприклад, менш чим 1мМ, 0,9мМ, 0,8мМ, 0,7мМ, 0,6мМ, 0,5мМ, 0,4мМ, 0,3мМ, 0,2мМ, 0,1 мМ, 0,05мМ або менш. Поліпептид GAT може мати більше високе значення kcat для гліфосату, чим раніше ідентифіковані ферменти, наприклад, зна-1 -1 чення kcat щонайменше 500 хв , 1000 хв , 1100 хв 1 -1 -1 -1 -1 , 1200 хв , 1250 хв , 1300 хв , 1400 хв , 1500 хв 1 -1 -1 -1 -1 , 1600 хв , 1700 хв , 1800 хв ', 1900 хв або 2000 -1 хв або більше. Поліпептиди GAT, призначені для застосування відповідно до винаходу, можуть мати більше високе значення kcat/KМ для гліфосату, чим раніше ідентифіковані ферменти, наприклад, -1 -1 значення kcat/KМ щонайменше 1000мМ хв , -1 -1 -1 -1 -1 -1 2000мМ хв , 3000мМ хв , 4000мМ хв , 5000мМ 1 -1 -1 -1 -1 -1 -1 -1 хв , 6000мМ хв , 7000мМ хв або 8000мМ хв або більше. На активність ферментів впливає, наприклад, значення рН і концентрація солей; відповідні методи аналізу й умови відомі в даній галузі (див., наприклад, WO 2005/012515). Такі поліпшені ферменти можуть знайти конкретне застосування в методах вирощування культурних рослин у польових умовах, у яких існує ризик ушкодження культурної рослини при застосуванні конкретного гербіциду або комбінації гербіцидів 11 і/або інших сільськогосподарський хімічних агентів, якщо ферментативна активність у рослині (тобто kCat, KМ або kcat/KМ) є зниженою. Толерантні до гліфосату рослини можна одержувати також шляхом модифікації рослини з метою підвищення її здатності продукувати більше високі рівні 5-єнолпірувшшикимат-3-фосфатсинтази (EPSPS), що більш докладно описано в U.S. №№ 6248876; 5627061; 5804425; 5633435; 5145783; 4971908; 5312910; 5188642; 4940835; 5866775; 6225114;6130366; 5310667; 4535060; 4769061; 5633448; 5510471; Re. 36449; RE 37287 Ε; і 5491288; і опублікованих міжнародних заявках на патент WO 97/04103; WO 00/66746; WO 01/66704; і WO 00/66747, які повністю й для всіх цілей включені в даний опис як посилання. Стійкість до гліфосату можна також надавати рослинам, які експресують ген, що кодує фермент гліфосат-оксидоредуктазу, що більш докладно описано в U.S. №№ 5776760 і 5463175, які повністю й для всіх цілей включені в даний опис як посилання. Крім того, толерантні до гліфосату рослини можна створювати шляхом селекції мутацій, що зустрічаються в природних умовах, що обумовлюють толерантність до гліфосату. При створенні винаходу було встановлено, що в способах і композиціях, пропонованих у винаході, можна застосовувати будь-яку відому в даній області комбінацію послідовностей (тобто послідовності, що діють за допомогою того самого або різних механізмів), які обумовлюють толерантність до гліфосату, для одержання рослин і експлантантів рослин з підвищеною стійкістю до гліфосату. б. Толерантність до інгібіторів ацетолактатсинтази (ALS) Винахід відноситься до рослин, толерантних до гліфосату/інгібітору ALS, які містять полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату, і містить також полінуклеотид, що кодує толерантний до інгібітору ацетолактатасинтази (ALS) поліпептид. У контексті даного опису поняття «толерантний до інгібітору ALS поліпептид» відноситься до будь-якого поліпептиду, що при експресії в рослині обумовлює толерантність щонайменше до одного інгібітору ALS. Відомі різні інгібітори ALS, і до них відносяться, наприклад, такі гербіциди, як сульфонілсечовина, імідазолінон, тріазолопіримідини, піримідинокси(тіо)бензоати й/або сульфоніламінокарбонілтріазолінони. Відомі й інші інгібітори ALS, і вони зазначені також у даному описі. У даній області відомі мутації ALS, що відносяться до різних класів, залежно від їхньої здатності обумовлювати толерантність до сульфонілсечовинам, імідазолінонам, тріазолопіримідинам і піримідініл(тіо)бензоатам, включаючи мутації, які мають наступні характеристики: (1) обумовлюють широкий спектр толерантності до гербіцидів із всіх 4 зазначених груп; (2) обумовлюють толерантність до імідазолінонів і пирімідініл(тіо)бензоатів; (3) обумовлюють толерантність до сульфонілсечовин і тріазолопіримідинів; і (4) обумовлюють толерантність до сульфонілсечовинів і імідазолінонів. Можна застосовувати різні толерантні до інгібітору ALS поліпептиди. У деяких варіантах здійс 96578 12 нення винаходу толерантні до інгібітору ALS полінуклеотиди містять щонайменше одну нуклеотидну мутацію, що приводить до однієї амінокислотної заміни в поліпептиді ALS. У конкретних варіантах здійснення винаходу заміна зачіпає одну із семи практично консервативних областей ацетолактатсинтази (див., наприклад, Hattori і ін., Molecular Genetics and Genomes 246, 1995, cc. 419-425; Lee і ін., ЕМВО Journal 7, 1998, cc. 1241-1248; Mazur і ін., Ann. Rev. Plant Phys. 40, 1989, cc. 441-470; і U.S. № 5605011, всі документи повністю включені в даний опис як посилання. Толерантний до інгібітору ALS поліпептид може кодуватися, наприклад, локусом SuRA або SuRB ALS. У конкретних варіантах здійснення винаходу толерантний до інгібітору ALS поліпептид містить мутант ALS C3, мутант ALS HRA і мутант S4 або мутант S4/HRA або їхню будь-яку комбінацію. Відомі різні мутації ALS, які обумовлюють толерантність до різних гербіцидів і груп (і/або підгрупам) гербіцидів; див., наприклад, Tranel і Wright, Weed Science 50, 2002, cc. 700-712 (див. також, U.S. №№ 5605011, 5378824, 5141870 і 5013659, кожний з яких повністю включений у даний опис як посилання). Крім того, див. SEQ ID NO:65, що містить послідовність HRA сої; SEQ ID NO:66, що містить послідовність HRA кукурудзи; SEQ ID NO:67, що містить послідовність HRA Arabidopsis; і SEQ ID NO:86, що містить послідовність HRA, застосовувану в бавовнику. Мутація ALS HRA знайшла конкретне застосування в одному з варіантів здійснення винаходу. Мутація приводить до виробництва поліпептиду ацетолактатсинтази, що має стійкість щонайменше до одної з хімічних сполук, що є інгібіторами ALS, у порівнянні з білком дикого типу. Наприклад, рослина, експресуюча толерантний до інгібітору ALS поліпептид, може мати толерантність до доз таких гербіцидів, як сульфонілсечовина, імідазолінон, тріазолопіримідини, піримідінілокси(тіо)бензоати й/або сульфоніламінокарбонілтріазолінон, які щонайменше в 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 50, 70, 80, 100, 125, 150, 200, 500 або 1000 разів вище дози гербіциду, що може викликати ушкодження відповідної контрольної рослини. У деяких варіантах здійснення винаходу толерантний до інгібітору ALS поліпептид містить кілька мутацій. Крім того, можна створювати рослини, які несуть толерантний до інгібітору ALS поліпептид, шляхом селекції мутацій, що зустрічаються в природних умовах, які надають толерантність до гліфосату. У деяких варіантах здійснення винаходу толерантний до інгібітору ALS поліпептид обумовлює толерантність до сульфонілсечовинних і імідазолінонівих гербіцидів. Сульфонілсечовинні й імідазоліноніві гербіциди інгібують ріст вищих рослин шляхом блокади ацетолактатсинтази (ALS), що називають також синтаза ацетоацетогідрокислоти (AHAS). Наприклад, рослини, що несуть конкретні мутації в ALS (наприклад, мутації S4 і/або HRA), маю толерантність до сульфонілсечовинних гербіцидів. Одержання рослин, толерантних до сульфонілсечовин, і рослин, толерантних до імідазолінонів, більш докладно описано в U.S. №№ 5605011; 5013659; 5141870; 5767361; 5731180; 13 5304732; 4761373; 5331107; 5928937; і 5378824; і опублікованій міжнародній заявці WO 96/33270, які все повністю включено в даний опис як посилання для всіх цілей. У конкретних варіантах здійснення винаходу толерантний до інгібітору ALS поліпептид містить толерантну до сульфонамідів ацетолактатсинтазу (якої називають також толерантною до сульфонамідів синтазою ацетоацетогідрокислоти) або толерантну до імідазолінонів ацетолактатсинтазу (яку називають також толерантною до імідазолінонів синтазою ацетоацетогідрокислоти). Рослина, пропонована у винаході, що містить щонайменше одну послідовність, що обумовлює толерантність до гліфосату, і щонайменше одну послідовність, що обумовлює толерантність до інгібітору ALS, означають у даному описі як «рослина, толерантна до гліфосату/інгібітору ALS». Рослина, пропонована у винаході, що містить щонайменше один поліпептид GAT і щонайменше один поліпептид HRA, означають у даному описі як «GAT-HRA-рослина». в. Толерантність до інших гербіцидів У деяких варіантах здійснення винаходу запропоновані рослини, що володіють підвищеною толерантністю до гліфосату й щонайменше до одного гербіциду-інгібітору ALS. а також толерантністю принаймні ще до одного додаткового гербіциду. У конкретних варіантах здійснення винаходу толерантність до додаткового гербіциду пов'язана з експресією щонайменше одного поліпептиду, що надає толерантність до додаткового гербіциду. У деяких варіантах здійснення винаходу композиція, пропонована у винаході (наприклад, рослина), може містити два, три, чотири, п'ять, шість, сім або більше ознак, що обумовлюють толерантність щонайменше до одного гербіциду, у результаті рослина, пропонована у винаході, може мати толерантність щонайменше до двох, трьох, чотирьох, п'яти, шести або семи або більшої кількості різних типів гербіцидів. Так, рослина, пропонована у винаході, що має толерантність більш ніж до двох різних гербіцидів, може мати толерантність до гербіцидів, які мають різні механізми дії й/або різними місцями дії. У деяких варіантах здійснення винаходу всі ці ознаки являють собою трансгенні ознаки, хоча в інших варіантах здійснення винаходу щонайменше один із цих ознак не є трансгенним. У деяких із цих варіантів здійснення винаходу кожний ген, що обумовлює толерантність до гербіциду, надає толерантність до різних гербіцидів або класів або підкласів гербіцидів. У деяких із цих варіантів здійснення винаходу щонайменше два гени, що обумовлюють толерантність до гербіцидів, надають толерантність до того самого гербіциду або представника того самого класу або підкласу гербіцидів. Таким чином, винахід відноситься також до рослин, що несуть полінуклеотид, що кодує поліпептид, що може обумовлювати толерантність до гліфосату, і полінуклеотид, що кодує пептид, толерантний до інгібітору ALS, які можуть містити також щонайменше один полінуклеотид, зумовлюючий толерантність до додаткового гербіциду, що при експресії надає толерантність до додаткового гербіциду. Такі додаткові 96578 14 гербіциди включають, але, не обмежуючись ними, інгібітор ацетил-СоА-карбоксилази, такий як хизалофоп-п-етил, синтетичний ауксин, такий як квинхлорак, гербіцид-інгібітор протопорфиногеноксидази (РРО) (такий як сулфентразон), гербіцидінгібітор синтезу пігментів, такий як інгібітор гідроксифенілпіруват-діоксигенази (наприклад, мезотріон або сулькотріон), інгібітор фосфінотрицинацетилтрансферази або фітоендесатурази типу дифлуфенікану або інгібітор синтезу пігментів. Варто розуміти, що винахід не обмежений механізмом дії гербіциду, якщо вирішується завдання, поставлене у винаході (тобто толерантність до гліфосату й щонайменше до одного інгібітору ALS). Додаткові гербіциди, що представляють інтерес, перераховані нижче. У деяких варіантах здійснення винаходу композиції, пропоновані у винаході, додатково містять поліпептиди, що обумовлюють толерантність до гербіцидів, які інгібують фермент глутамінсинтазу. таким як фосфінотрицин або глуфосинат (наприклад, ген bar або ген pat). Глутамінсинтетаза (GS), очевидно, являє собою основний фермент, необхідний для розвитку й життя більшості рослинних кліток, і інгібітори GS токсичні для рослинних кліток. Гербіциди-аналоги глуфосинату розроблені на основі їхньої токсичної дії, пов'язаної з інгібуванням GS у рослинах. Ці гербіциди мають невибіркову дію; це означає, що вони інгібують ріст всіх різних присутніх видів рослин. Створення рослин, що містять екзогенну фосфінотрицинацетилтрансферазу, описане в U.S. №№ 5969213; 5489520; 5550318; 5874265; 5919675; 5561236; 5648477; 5646024; 6177616; і 5879903, які повністю й для всіх цілей включені в даний опис як посилання. Описана також мутантна фосфінотрицинацетилтрансфераза, що володіє зазначеною активністю. В інших варіантах здійснення винаходу композиції, пропоновані у винаході, містять також поліпептиди, що обумовлюють толерантність до гербіцидів, які інгібують протоке (протопорфиногеноксидазу). Протоке потрібно для виробництва хлорофілу, що необхідно для виживання всіх рослин. Фермент протоке служить як мішень різних сполук, що володіють гербіцидною активністю. Ці гербіциди інгібують також ріст всіх інших присутніх видів рослин. Створення рослин, що містять протоке зі зміненою активністю, які мають стійкість до гербіцидів, описані в U.S. №№ 6288306; 6282837; і 5767373; і в опублікованій міжнародній заявці WO 01/12825, які повністю й для всіх цілей включені в даний опис як посилання. У наступних варіантах здійснення винаходу композиції, пропоновані у винаході, можуть містити поліпептиди, пов'язані з іншими механізмами стійкості до гербіцидів. Наприклад, гідроксифенілпіруватдіоксигенази являють собою ферменти, які катализують реакцію, у якій парагідроксифенілпіруват (НРР) трансформується в гомогентисат. Молекули, які інгібують цей фермент і які зв'язуються з ферментом, інгібуючи трансформацію НРР у гомогентисат, застосовують як гербіциди. Рослини, що володіють стійкістю до інших конкретних гербіцидів, описані в U.S. №№ 6245968; 6268549; і 6069115; і в опублікованій мі 15 жнародній заявці WO 99/23886, які повністю й для всіх цілей включені в даний опис як посилання. Описана також мутантна гідроксифенілпіруватдіоксигеназа, що володіє зазначеною активністю. г. Фрагменти й варіанти послідовностей, які обумовлюють толерантність до гербіцидів Залежно від контексту поняття «фрагмент» відноситься до області полінуклеотиду або області амінокислотної послідовності й, природно, до кодованого білка. Фрагменти полінуклеотиду можуть кодувати фрагменти білка, які зберігають біологічну активність вихідного білка й, отже, обумовлювати толерантність до гербіциду. Так, фрагменти нуклеотидної послідовності можуть складатися щонайменше приблизно з 20 нуклеотидів, приблизно 50 нуклеотидів, приблизно 100 нуклеотидів і аж до повнорозмірного полінуклеотиду, що кодує толерантний до гербіциду поліпептид. Фрагмент толерантного до гербіциду полінуклеотиду, що кодує біологічно активну область толерантного до гербіциду поліпептиду, повинен кодувати щонайменше 15, 25, 30, 50, 100, 150, 200 або 250 суміжних амінокислот або аж до повної кількості амінокислот, що є присутнім у повнорозмірному толерантному до гербіциду поліпептиді. Біологічно активну область толерантного до гербіциду поліпептиду можна одержувати шляхом виділення області толерантного до гербіциду полінуклеотиду, експресуючого кодуючу область толерантного до гербіциду поліпептиду (наприклад, шляхом рекомбінантної експресії in vitro), і оцінки активності кодуємої області толерантного до гербіциду поліпептиду. Полінуклеотиди, що представляють собою фрагменти толерантного до гербіциду полінуклеотиду, містять щонайменше 16, 20, 50, 75, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 800, 900, 1000, 1100, 1200, 1300 або 1400 суміжних нуклеотидів або аж до кількості нуклеотидів, присутніх у повнорозмірному толерантному до гербіциду полінуклеотиді. Поняття «варіанти» відноситься до практично подібних послідовностей. Для полінуклеотидів варіант являє собою полінуклеотид, що має делеції (тобто вкорочення) на 5'- і/або 3'-кінці; делеції й/або додавання одного або декількох нуклеотидів в одному або декількох внутрішніх сайтах у нативном полінуклеотиді; і/або заміну одного або декількох нуклеотидів в одному або декількох сайтах у нативном полінуклеотиді. У контексті даного опису поняття «нативний» полінуклеотид або поліпептид відноситься до послідовності, що зустрічається в природних умовах нуклеотидної, або амінокислотній послідовності відповідно. Для полінуклеотидів консервативні варіанти включають такі послідовності, які через вродженість генетичного коду кодують амінокислотну послідовність толерантного до гербіциду поліпептиду. Варіанти, що зустрічаються в природних умовах алельні, можна ідентифікувати за допомогою добре відомих методів молекулярної біології, таких, наприклад, як полімеразна ланцюгова реакція (ПЦР) і методи гібридизації. Варіанти полінуклеотидів включають також отримані шляхом синтезу полінуклеотиди, наприклад, отримані за допомогою сайтнаправленого мутагенезу або «перестановки». Як правило, 96578 16 послідовності варіантів конкретного полінуклеотиду ідентичні щонайменше приблизно на 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% або більше послідовності зазначеного конкретного полінуклеотиду, що визначають за допомогою описаних нижче програм, призначених для порівняльного аналізу первинної структури послідовностей, і використовуваних у них параметрів. Варіанти конкретного полінуклеотиду (тобто, полінуклеотиду, з яким проводять порівняння (референс-полінуклеотиду) можна оцінювати також шляхом порівняння відсотка ідентичності послідовностей поліпептиду, кодованого варіантом полінуклеотиду, і поліпептиду, кодованого референсполінуклеотидом. Відсоток ідентичності послідовностей будь-яких двох поліпептидів можна розраховувати за допомогою програм, призначених для порівняльного аналізу первинної структури послідовностей, і параметрів які описані нижче. При оцінці будь-якої конкретної пари полінуклеотидів, пропонованих у винаході, шляхом порівняння відсотка ідентичності послідовностей на основі двох поліпептидів, які вони кодують, відсоток ідентичності послідовностей двох кодованих поліпептидів становить щонайменше приблизно 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% або більше. Поняття «варіант» білка означає білок, отриманий з нативного й/або вихідного білка шляхом делеції (так зване вкорочення) однієї або декількох амінокислот на N-кінці й/або С-кінці білка; делеції й/або додавання однієї або декількох амінокислот в один або кілька внутрішніх сайтів білка; або заміни однієї або декількох амінокислот в одному або декількох сайтах білка. Варіанти білків, що підпадають під об'єм даного винаходу, є біологічно активними, оскільки вони продовжують зберігати необхідну толерантність до гербіцидів, що буде описано нижче. Послідовність біологічно активних варіантів толерантного до гербіциду поліпептиду, пропонованого у винаході, повинна бути ідентична щонайменше приблизно на 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% або більше амінокислотної послідовності нативного білка, що визначають за допомогою програм, призначених для порівняльного аналізу первинної структури послідовностей, і використовуваних у них параметрів, які описані нижче. Біологічно активний варіант толерантного до гербіциду поліпептиду може відрізнятися лише 1-15 амінокислотними залишками, лише 1-10, наприклад 6-10, лише 5, лише 4, 3, 2 або навіть 1 амінокислотним залишком. Варіанти зумовлюючих толерантність до гербіцидів поліпептидів, а також полінуклеотиди, що кодують ці варіанти, відомі в даній області. Зумовлюючі толерантність до гербіцидів поліпептиди можна змінювати різними шляхами, включаючи амінокислотні заміни, делеції, укорочення й інсерції. Методи здійснення таких маніпуляцій є загальновідомими в даній області. Наприклад, варіанти й фрагменти зумовлюючих толерантність до гербіцидів поліпептидів можна одержувати 17 шляхом мутації в полінуклеотиді, що кодує. Методи мутагенезу й зміни полінуклеотидів добре відомі в даній області (див., наприклад, Kunkel, Proc. Nail. Acad. Sci. USA 82, 1985, cc. 488-492; Kunkel і ін., Methods in Enzymol. 154, 1987, cc. 367-382; U.S. № 4873192; Techniques in Molecular Biology, під ред. Walker і Gaastra, 1983, з MacMillan Publishing Company, New York) і процитовані в цих публікаціях посилання. Посібник зі здійснення амінокислотних замін, які не впливають на біологічну активність білка, що представляє інтерес, описано на моделі, створеної Dayhoff і ін. (Atlas of Protein Sequence and Structure, з Natl. Biomed. Res. Found., Washington, D.C., 1978), публікація включена в даний опис як посилання. Можна здійснювати консервативні заміни, такі як заміна однієї амінокислоти на іншу, що має аналогічні властивості. Фахівцеві в даній області повинно бути очевидно, що активність зумовлюючу толерантність до гербіциду поліпептиду можна оцінювати за допомогою загальноприйнятих методів скринінга. Це означає, що активність можна оцінювати, визначаючи, чи володіє трансгенна рослина підвищеною толерантністю до гербіциду, наприклад, за допомогою методу, проілюстрованого в прикладі 1, або за допомогою аналізу in vitro, наприклад, по утворенню N-ацетилфосфату із гліфосату за допомогою поліпептиду GAT (див., наприклад, WO 02/36782). Варіанти полінуклеотидів і поліпептидів містять також послідовності й білки, отримані в результаті методів мутагенезу й рекомбінації, таких як перестановка ДНК. За допомогою зазначеної процедури можна маніпулювати однією або декількома кодуючими послідовностями зумовлюючих толерантність до гербіциду поліпептиду для створення нового зумовлюючу толерантність до гербіциду поліпептиду з необхідними властивостями. Таким шляхом створюють бібліотеки рекомбінантних полінуклеотидів з популяції родинних полінуклеотидних послідовностей, що містять області, послідовності яких є практично ідентичними, і які можна гомологично рекомбінувати in vitro або in vivo. Наприклад, за допомогою зазначеного підходу мотиви послідовностей, що кодують домен, що представляє інтерес, можна поміщати шляхом перестановки між геном, що кодує зумовлюючу толерантність до гербіциду поліпептид, і іншими відомими генами, що кодують білок з поліпшеним властивістю, що представляє інтерес, таким як збільшене значення Km у випадку ферменту. Стратегії такої перестановки ДНК відомі в даній області (див., наприклад, Stemmer, Proc. Natl. Acad. Sci. USA 91, 1994, cc. 10747-10751; Stemmer, Nature 370, 1994, cc. 389-391; Crameri і ін., Nature Biotech. 15, 1997, cc. 436-438; Moore і ін., J. Mol. Biol. 272, 1997, cc. 336-347; Zhang і ін., Proc. Natl. Acad. Sci. USA 94, 1997, cc. 4504-4509; Crameri Nature 391, 1998, cc. 288-291; і U.S. №№ 5605793 і 5837458). Для опис взаємозв'язків послідовностей двох або більшої кількості полінуклеотидів або поліпептидів використовують наступні поняття: (а) «референс-послідовність», (б) «вікно порівняння», (в) 96578 18 «ідентичність послідовності» і (г) «відсоток ідентичності послідовностей». (а) У контексті даного опису поняття «референс-послідовність» відноситься до певної послідовності, застосовуваної як базис для порівняння послідовностей. Референс-послідовність може являти собою субпопуляцію або повну конкретну послідовність; наприклад, сегмент повнорозмірної кДНК або послідовності гена або повну кДНК або послідовність гена. (б) У контексті даного опису «вікно порівняння» відноситься до безперервного й конкретного сегмента полінуклеотидної послідовності, причому, полінуклеотидна послідовність у вікні порівняння може містити доповнення або делеції (тобто пролому) щодо референс-послідовності (яка не містить додавання або делеції) для оптимального порівняльного аналізу первинної структури двох полінуклеотидів. Як правило, вікно порівняння складається щонайменше з 20 послідовних нуклеотидів і оптимально 30, 40, 50, 100 або більше нуклеотидів. Фахівцям у даній області повинно бути очевидно, що для того, щоб уникнути високої подібності з референс-послідовністю через включення проломів у полінуклеотидну послідовність, як правило, уводять штраф за пролом і віднімають його з кількості збігів. Методи порівняльного аналізу первинної структури послідовностей добре відомі в даній області. Так, визначення відсотка ідентичності послідовностей двох будь-яких послідовностей можна здійснювати за допомогою математичного алгоритму. Прикладами таких математичних алгоритмів є (але, не обмежуючись ними) алгоритм, описаний в Myers і Miller CABIOS 4, 1988, cc. 11-17; локальний алгоритм для порівняльного аналізу первинної структури послідовностей, описаний в Smith і ін., Adv. Appl. Math. 2, 1981, с. 482; загальний алгоритм порівняльного аналізу первинної структури послідовностей, описаний в Needleman і Wunsch, J. Mol. Biol. 48, 1970, сс. 443-453; метод порівняльного аналізу первинної структури послідовностей, заснований на алгоритмі локального пошуку, описаний в Pearson і Lipman, Proc. Natl. Acad. Sci. 85, 1988, сс. 2444-2448; алгоритм, описаний в Karlin і Altschul, Proc. Natl. Acad. Sci. USA 87, 1990, cc. 2264-2268, модифікація якого описана в Karlin і Altschul, Proc. Natl. Acad. Sci. USA 90, 1993, cc. 5873-5877. Комп'ютерну реалізацію цих математичних алгоритмів можна застосовувати для порівняння послідовностей з метою визначення ідентичності послідовностей. Такі реалізації включають (але, не обмежуючись ними): програму CLUSTAL, що входить у програму PC/Gene (фірма Intelligenetics, Маунтин-Вью, шт. Каліфорнія)); програму ALIGN (версія 2.0) і GAP, BESTFIT, BLAST, FASTA і TFASTA, які входять у пакет програм GCG Wisconsin Genetics, версія 10 (фірма Accelrys Inc., 9685 Scranton Road, Сан-Дієго, шт. Каліфорнія, США). Порівняльні аналізи первинної структури за допомогою цих програм можна здійснювати з використанням параметрів, що задають за замовчуванням. Програма CLUSTAL докладно описана в Higgins і ін., Gene 73, 1988, сс. 237-244; Higgins і 19 ін., CABIOS 5, 1989, сс. 151-153; Corpet і ін., Nucleic Acids Res. 16, 1988, сс. 10881-90; Huang і ін., CABIOS 8, 1992, сс. 155-65; і Pearson і ін., Meth. Mol. Biol. 24, 1994, сс. 307-331. Програма ALIGN заснована на алгоритмі, запропонованому Myers і Miller,1988, вище. При порівнянні амінокислотних послідовностей за допомогою програми ALIGN можна використати таблицю ваг залишків РАМ120, штраф за довжину пролому 12 і штраф за пролом 4. Програма BLAST, описана в Altschul і ін., J. Mol. Biol. 215, 1990, с 403, заснована на алгоритмі, запропонованому Karlin і Altschul, 1990, вище. Пошук нуклеотидів за допомогою BLAST можна здійснювати з використанням програми BLASTN, у якій бал=100, довжина слова=12, для одержання нуклеотидних послідовностей, гомологічних нуклеотидної послідовностей, що кодує білок, пропонований у винаході. Пошук білків за допомогою BLAST можна здійснювати з використанням програми BLASTX, у якій бал=50, довжина слова=3, для одержання амінокислотних послідовностей, гомологічних білку або поліпептиду, пропонованому у винаході. Для порівняльного аналізу первинної структури послідовностей з використанням проломів можна застосовувати програму Gapped BLAST (в BLAST 2.0), описану в Altschul і ін., Nucleic Acids Res. 25, 1997, с. 3389. В іншому варіанті можна застосовувати програму PSI-BLAST (в BLAST 2.0) для здійснення ітеративного пошуку, що дозволяє виявляти віддалені взаємозв'язки між молекулами (див. Altschul і ін., 1997, вище). При використанні програм BLAST, Gapped BLAST, PSIBLAST, можна застосовувати параметри, що задають за замовчуванням, що відповідають програм (наприклад, BLASTN для нуклеотидних послідовностей, BLASTX для білків). Програмне забезпечення BLAST доступно наукової громадськості на сайті в мережі Інтернет NCBI. Порівняльний аналіз первинної структури послідовностей можна здійснювати вручну шляхом візуального аналізу. Якщо не зазначене інше, то дані про ідентичність/подібності послідовностей, представлені в даному описі, являють собою дані, отримані за допомогою програми GAP, версія 10, з використанням наступних параметрів: % ідентичності й % подібності нуклеотидних послідовностей з використанням ваги пролому 50 і ваги довжина слова 3 і матриці балів nwsgapdna.cmp; % ідентичності й % подібності для амінокислотних послідовностей з використанням ваги пролому 8 і довжини слова 2, і матриці балів BLOSUM62; або будь-якої еквівалентної їй програми. Під «еквівалентною програмою» розуміють будь-яку програму для порівняння послідовностей, з використанням якої для будь-яких двох розглянутих послідовностей при порівняльному аналізі первинної структури послідовностей одержують ідентичні збіги нуклеотидних або амінокислотних залишків і ідентичний відсоток ідентичності послідовностей, при порівнянні з відповідними результатами порівняльного аналізу первинної структури послідовностей, отриманими за допомогою програми GAP, версія 10. Програма GAP заснована на використанні алгоритму Needleman і Wunsch, J. Mol. Biol. 48, 1970, 96578 20 сс. 443-453, призначеного для порівняльного аналізу первинної структури двох повних послідовностей, що дозволяє максимізувати кількість збігів і мінімізувати кількість проломів. У програмі GAP розглядаються всі можливі порівняльні аналізи первинної структури послідовностей і положення проломів, і вона дозволяє здійснювати порівняльний аналіз первинної структури послідовностей з найбільшою кількістю збігів підстав і найменшою кількістю проломів. Вона дозволяє виражати штраф за створення пролому й штраф за подовження пролому в одиницях збігу підстав. Програма GAP повинна дозволяти робити «рентабельною» кожну убудовану на її основі проломів з позицій штрафу за створення пролому й кількості збігів. Якщо обраний штраф за подовження пролому перевищує нуль, то програма GAP повинна дозволяти, крім того, робити «рентабельною» кожний убудований пролом з позицій штрафу за довжину пролому, помноженого на штраф за подовження пролому. Задані за замовчуванням величини штрафу за створення пролому й величини штрафу за подовження пролому у версії 10 пакета програм GCG Wisconsin Genetics Software Package для білкових послідовностей становлять 8 і 2 відповідно. Для нуклеотидних послідовностей штраф, що задає по умовчанню, за створення пролому становить 50, а штраф, що задає по умовчанню, за подовження пролому становить 3. Штрафи за створення пролому й подовження пролому можна виражати в цілих числах від 0 до 200. Так, наприклад, штрафи за створення пролому й за подовження пролому можуть становити 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65 або більше. Програма GAP є одним із представників сімейства найбільш ефективних програм для порівняльного аналізу первинної структури послідовностей. У це сімейство входить багато представників, але жоден з інших представників не має більше високу якість. GAP дозволяє одержувати наступні чотири параметри при порівняльному аналізі первинної структури послідовностей: якість, співвідношення, ідентичність і подібність. Якість являє собою метричний параметр, максимізований для вирівнювання послідовностей. Співвідношення являє собою якість, ділену на кількість підстав у більше короткому сегменті. Відсоток ідентичності являє собою відсоток символів, які фактично збігаються. Відсоток подібності являє собою відсоток символів, які є подібними. Символи, що входять у проломі, ігноруються. Подібність оцінюють, коли величина матриці балів для пари символів більше або дорівнює 0,50, що є межею подібності. Матриця балів, застосовувана у версії 10 пакета програм GCG Wisconsin Genetics Software, являє собою матрицю BLOSUM62 (див. Henikoff і Henikoff, Proc. Natl. Acad. Sci. USA 89, 1989, с. 10915). (в) У контексті даного опису поняття «ідентичність послідовностей» або «ідентичність» відносно двох полінуклеотидних або поліпептидних послідовностей відноситься до залишків у двох послідовностях, які є однаковими при вирівнюванні для максимальної відповідності в конкретному вікні 21 порівняння. Якщо відсоток ідентичності послідовностей визначають для білків, то враховують, що положення залишків, які не є ідентичними, часто розрізняються консервативними амінокислотними замінами, при яких амінокислотні залишки замінені іншими амінокислотними залишками з подібними хімічними властивостями (наприклад, заряд або гідрофобність), і тому не змінюють функціональні властивості молекули. Коли послідовності розрізняються консервативними замінами, то відсоток ідентичності послідовностей можна регулювати шляхом корекції на консервативну природу заміни. Послідовності, що розрізняються такими консервативними замінами, відносять до «подібних послідовностей» або говорять про їх «подібність або подобу». Методи одержання зазначеного коректування добре відомі в даній області. Як правило, вони засновані на тім, що консервативну заміну оцінюють у балах як часткове, а не повна розбіжність, підвищуючи тим самим відсоток ідентичності послідовностей. Так, наприклад, якщо ідентичній амінокислоті привласнюється бал 1, а неконсервативній заміні привласнюється бал 0, те консервативній заміні привласнюється бал між нулем і одиницею. Бальну оцінку консервативних замін, розраховують, наприклад, за допомогою програми PC/GENE (фірма Intelligenetics, Маунтин-Вью, шт. Каліфорнія). (г) У контексті даного опису «відсоток ідентичності послідовностей» означає величину, що визначають шляхом порівняння двох оптимальним чином вирівняних послідовностей у вікні порівняння, при цьому фрагмент полінуклеотидної послідовності у вікні порівняння може містити доповнення або делеції (тобто пролому) у порівнянні з референс-послідовністю (яка не містить доповнень або делецій) для оптимального порівняльного аналізу первинної структури двох послідовностей. Відсоток розраховують, визначаючи кількість положень, у яких перебувають однакові нуклеотидні основи або амінокислотні залишки в обох послідовностях, у результаті чого одержують кількість співпадаючих положень, після чого здійснюють розподіл кількості співпадаючих положень на загальну кількість положень у вікні порівняння й множать результат на 100 для одержання відсотка ідентичності послідовностей. Мається на увазі, що застосування поняття «полінуклеотид» не обмежене полінуклеотидами, що містить ДНК. Фахівцеві в даній області повинно бути очевидно, що полінуклеотиди можуть містити рибонуклеотиди й комбінації рибонуклеотидів і дезоксирибонуклеотидів. Такі дезоксирибонуклеотиди й рибонуклеотиди включають як, молекули, що зустрічаються в природних умовах, так і їхні синтетичні аналоги. Так, до полінуклеотидум відносяться також всі форми послідовностей, включаючи (але, не обмежуючись ними) одноланцюгові форми, двухланцюгові форми, «шпильки» і структури типу «стебло-і-петля» і т.п. д. Толерантність до гербіцидів «Гербіцид» являє собою хімічну сполуку, що викликає тимчасову або постійну поразку рослини. Приклади деяких гербіцидів (але, не обмежуючись ними), які можна застосовувати в різних способах і 96578 22 композиціях, пропонованих у винаході, більш докладно будуть дані нижче в даному описі. Гербіцид може проникати в рослину або може впливати на рослину без проникнення в рослину або її клітки. Діюча речовина являє собою хімічну сполуку в гербіцидній препаративній формі, відповідальну насамперед за її фітотоксичність, і позначену як діюча речовина на етикетці продукту. Дані про інформації, що повинна бути представлена на етикетці продукту, можна одержати в Агентстві по захисту навколишнього середовища США (U.S. Environmental Protection Agency) і в режимі «online» на обновлюваному сайті url oaspub.epa.gov/pestlabl/ppls.own; дані про інформацію, представлену на етикетці продукту, доступні також у режимі «on-line» на сайті url www.cdms.net. Поняття «кислотний еквівалент» відноситься до рівня або кількості, вираженому в перерахуванні на гербіцидно активну батьківську (вихідну) кислоту. Наприклад, 2,4-д-кислота (2,4дихлорфеноксиоцтова кислота) часто у вигляді діючої речовини в складі препаративної форми продуктів являє собою натрієву або амінну сіль або ефір. Активний кислотний еквівалент на галон при використанні широко застосовуваної препаративної форми на основі ефіру становить 3,8 фунтів д.в./галон (приблизно 0,454 кг д.в./л), при цьому вміст діючої речовини на галон становить 6,0 фунтів д.в./галон (приблизно 0,717 кг д.в./л). У контексті даного опису поняття «агрохімікат (сільськогосподарська хімічна речовина, хімічний засіб захисту рослин)» відноситься до будь-якої хімічної речовини, застосовуваній у сільському господарстві. «Телерантний до гербіцидів» або «толерантний» або «толерантність сільськогосподарської культури» відносно обробки гербіцидом або іншою хімічною обробкою в контексті даного опису означає, що в рослини або іншого організму, обробленого конкретним гербіцидом або класом або підкласом гербіцидів або іншою хімічною речовиною або класом або підкласом іншої хімічної речовини, не виявлені помітні ушкодження або виявлені менш виражене ушкодження після обробки в порівнянні з відповідною контрольною рослиною. Рослина може мати природну толерантність до конкретного гербіциду або хімічної речовини або рослина може мати толерантність до гербіциду в результаті людського втручання, наприклад, селекції або генетичній інженерії. «Зумовлюючу толерантність до гербіциду поліпептид» («толерантний до гербіциду поліпептид») являє собою поліпептид, що обумовлює толерантність до гербіциду рослини або іншого організму, у якому відбувається його експресія (тобто він надає рослині або іншому організму толерантність до гербіциду, а «зумовлюючу толерантність до гербіциду полінуклеотид» («толерантний до гербіциду полінуклеотид») являє собою полінуклеотид, що кодує зумовлюючу толерантність до гербіциду поліпептид. Наприклад, толерантний до сульфонілсечовини поліпептид являє собою поліпептид, що обумовлює толерантність до сульфонілсечовинним гербіцидів рослини або іншого організму, у якому відбувається його експресія, толерантний до імі 23 дазолінону поліпептид являє собою поліпептид, що обумовлює толерантність до імідазолінонівих гербіцидів рослини або іншого організму, у якому відбувається його експресія; і толерантний до гліфосату поліпептид являє собою поліпептид, що обумовлює толерантність до гліфосату рослини або іншого організму, у якому відбувається його експресія. Таким чином, рослина має толерантність до гербіциду або іншій хімічній речовині, якщо при порівнянні з відповідною контрольною рослиною установлено, що її ушкодження нижче, ніж ушкодження контрольної рослини, щонайменше на 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 90%, 100%, 150%, 200%, 250%, 300%, 400%, 500%, 600%, 700%, 800%, 900% або 1000% або більше. У цьому плані рослина, що володіє толерантністю до гербіциду або іншій хімічній речовині, володіє «підвищеною толерантністю» у порівнянні з відповідною контрольною рослиною. Ушкодження, викликуване обробкою гербіцидом або іншою хімічною речовиною, оцінюють за допомогою будь-якого параметра росту рослини або параметра, розглянутого в якості прийнятного фахівцем у даній області. Ушкодження можна оцінювати візуальним оглядом і/або за допомогою статистичного аналізу прийнятних параметрів індивідуальних рослин або групи рослин. Так, для оцінки ушкодження можна застосовувати, наприклад, такі параметри як висота рослини, вага рослини, кольори листів, довжина листів, цвітіння, плідність, вимітування пестичних стовпчиків, урожайність, насінна продуктивність і т.п. Ушкодження можна оцінювати також по періоду часу, необхідному для конкретної стадії розвитку (наприклад, вимітування пестичних стовпчиків, цвітіння або скидання пилка), або періоду часу, протягом якого рослина відновлюється після обробки конкретною хімічною речовиною й/або гербіцидом. При здійсненні таких оцінок конкретні значення можна співвідносити з конкретним рівнем ушкодження, так, що можна проводити статистичний аналіз або кількісне порівняння. Застосування рангових значень для опису конкретних рівнів ушкодження відомо в даній області, і для цієї мети можна застосовувати будь-яку прийнятну систему рангів або шкалу. Наприклад, бальну оцінку пов'язаного з гербіцидом ушкодження (також називану бальною оцінкою толерантності) можна проводити, як проілюстровано в прикладі 1 з використанням шкали, представленої в таблиці 7. На цій шкалі оцінка 9 свідчить про те, що обробка гербіцидом не зробіла впливу на сільськогосподарську культуру, тобто що після гербіцидною обробки не виявленої ослаблення або ушкодження сільськогосподарської культури. Таким чином, на цій шкалі бал 9 свідчить про те, що сільськогосподарська культура не ушкоджується гербіцидом, і отже сільськогосподарська культура має толерантність до гербіциду. Як відзначалося вище, толерантність до гербіциду може бути оцінена також іншими балами з використанням цієї шкали, якщо ушкодження відповідної контрольної рослини характеризується більше низьким балом на цій шкалі або якщо група відповідних контрольних рослин характеризується 96578 24 статистично більше низьким балом відносно обробки гербіцидом у порівнянні із групою рослин, пропонованих у винаході. Ушкодження, викликуване гербіцидом або іншою хімічною речовиною, можна оцінювати в різні моменти часу після обробки рослини гербіцидом. Часте ушкодження оцінюють приблизно в той момент часу, коли в контрольної рослини виявлений максимум ушкодження. Іноді ушкодження оцінюють після періоду часу, протягом якого контрольна рослина, не оброблена гербіцидом або іншою хімічною речовиною, характеризується помітним ростом і/або розвитком щодо розміру або стадії, на якій проводять обробку. Ушкодження можна оцінювати в різний час, наприклад, через 12 год після обробки аналізуємої рослини гербіцидом або через 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 днів або трьох тижнів, чотирьох тижнів або пізніше. Для оцінки прийнятним є будь-який момент часу, якщо при цьому можна виявляти розходження відповіді на обробку між досвідченими й контрольними рослинами. Гербіцид не викликає «істотне ушкодження» рослини, коли він або не робить впливи на рослини, або коли він робить деякий вплив на рослину, з якого потім одержують рослину, або коли він робить вплив, що є шкідливим, але скомпенсованим, наприклад, вплив певного гербіциду на бур'яни. Так, наприклад, культурна рослина не є «істотно ушкодженою» гербіцидною або іншою обробкою, якщо в неї знижується менш чим на 50%, 40%, 30%, 25%, 20%, 15%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% або 1% щонайменше один прийнятний параметр, що свідчить про стан здоров'я й/або продуктивності рослини в порівнянні з відповідною контрольною рослиною (наприклад, неопрацьованою контрольною рослиною). Прийнятні параметри, які свідчать про стан здоров'я й/або продуктивності рослини включають, наприклад, висоту рослини, вагу рослини, довжину листів, час знаходження на конкретній стадії розвитку, цвітіння, урожайність, насінну продуктивність і т.п. Оцінку параметра можна здійснювати шляхом візуального огляду й/або статистичного аналізу будь-якого прийнятного параметра. Порівняння можна здійснювати візуальним оглядом і/або статистичним аналізом. Таким чином, культурна рослина не є «істотно ушкодженою» гербіцидною або іншою обробкою, якщо в неї відбувається зниження щонайменше одного параметра, але це зниження є тимчасовим по природі, і рослина повністювідновлюється протягом 1 тижня, 2 тижнів, 3 тижнів, 4 тижнів або 6 тижнів. I навпаки, рослина істотно ушкоджується в результаті гербіцидною або іншої обробки, якщо в неї відбувається зниження більш ніж на 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 150%, 170% принаймні одного прийнятного параметра, що свідчить про стан здоров'я й/або продуктивності рослини в порівнянні з відповідною контрольною рослиною (наприклад, неопрацьованим бур'яном цього ж виду). Таким чином, рослина істотно ушкоджується, якщо в неї відбувається зниження щонайменше одного параметра, і рослина не від 25 новлюється повністю протягом 1 тижня, 2 тижнів, 3 тижнів, 4 тижнів або 6 тижнів. Ушкодження рослини в результаті гербіцидної або іншої хімічної обробки може оцінювати візуальним оглядом фахівець у даній області і його можна оцінювати за допомогою статистичного аналізу прийнятних параметрів. Підлягаючу оцінці рослину означають як «досвідчену рослину». Як правило, контрольна рослина, що відповідає, являє собою рослину, що експресує такий(і) ж зумовлюючий(і) толерантність до гербіцидів поліпептид(и), що й рослина, яка підлягає оцінці відносно толерантності до гербіцидів (тобто «досвідчена рослина»), але не оброблена гербіцидом. Наприклад, при оцінці толерантності до гербіцидів рослини, пропонованої у винаході, що володіє толерантністю до гліфосату й інгібітору ALS, що відповідає контрольній рослині, повинно являти собою рослину, що експресує кожну із зазначених послідовностей, але яку не обробляють гербіцидом. У деяких випадках контрольна рослина являє собою рослину, що піддавалася такій же гербіцидною обробці, що й рослина, яка підлягає оцінці (тобто аналізована рослина), але яка не експресує фермент, призначений для забезпечення толерантності до гербіциду, що представляє інтерес, у аналізованій рослині. Фахівець у даній області може розробляти, виконувати й проводити оцінку відповідного експерименту в контрольованих умовах, призначеного для оцінки толерантності до гербіцидів рослини, що представляє, інтерес, включаючи відбір відповідних аналізованих рослин, контрольних рослин і обробок. Таким чином, у контексті даного опису «аналізована рослина або рослинна клітка» являє собою рослину або рослинну клітку, у яких була здійснена генетична зміна, що зачіпає щонайменше один ген, що представляє, інтерес, або рослина або рослинна клітка, які є нащадками рослини або клітки, змінених зазначеним чином, і які несуть цю зміну. Генетичну зміну можна інтродукувати у рослину шляхом селекції або трансформації. Поняття «генетична зміна» відноситься до гена або його мутації, що приводить до утворення фенотипу рослини, що відрізняється від фенотипу рослини, що не містить генетичну зміну. Поняття «контроль» або «контрольна рослина» або «контрольна рослинна клітка» відноситься до реперного об'єкта (об'єкту, з яким проводиться порівняння) для оцінки змін фенотипу в розглянутої рослини або рослинної клітки. Контрольна рослина або рослинна клітка може являти собою, наприклад,: (а) рослину або клітку дикого типу, тобто маючу такий же фенотип, що й вихідний матеріал для генетичної зміни, якому піддали розглянуту рослину або клітку; (б) рослину або клітку з таким же генотипом, що й вихідний матеріал, але яких трансформували «порожньою» конструкцією (тобто конструкцією, що не робить відомого впливу на ознака, що представляє інтерес, наприклад, конструкцією, що містить маркерний ген); (в) рослину або рослинну клітку, які являють собою нетрансформований ізолят (сегрегант) у потомстві розглянутої рослини або рослинної клітки; (г) рослину або рослинну клітку, які генетичні ідентичні розг 96578 26 лянутій рослині або рослинній клітці, але які не піддавали такій же обробці (наприклад, гербіцидній обробці), що й розглянута рослина або рослинна клітка; (д) сама розглянута рослина або рослинна клітка в умовах, коли не відбувається експресія гена, що представляє інтерес; або (e) сама розглянута рослина або рослинна клітка в умовах, коли їх не піддають конкретній обробці, такий, наприклад, як обробка гербіцидом або комбінацією гербіцидів і/або іншими хімічними речовинами. У деяких випадках відповідна контрольна рослина або контрольна рослинна клітка може мати генотип, відмінний від генотипу розглянутої рослини або рослинної клітки, але може мати ознаки чутливості до гербіциду вихідного матеріалу, призначеного для генетичного(них) зміни(н), які приводять до одержання розглянутої рослини або клітки (див., наприклад, Green, Weed Technology 12, 1998, cc. 474-477; Green i Ulrich, Weed Science 41, 1993, cc. 508-516). У деяких випадках відповідна контрольна рослина кукурудзи являє собою варіант NK603 (Nielson і ін., European Food Research and Technology 219, 2004, cc. 421-427 і Ridley і ін., Journal of Agriculture and Food Chemistry 50, 2002 cc. 7235-7243), елітна рослина із жорстким стеблом, рослина Р3162 (фірма Pioneer HiBred International), рослина 39Т66 (фірма Pioneer Hi-Bred International) або рослина 34М91 (фірма Pioneer Hi-Bred International). У деяких випадках відповідна контрольна рослина сої являє собою рослину сої лінії «Jack» (фірма Illinois Foundation Seed, Шампейн, шт. Іллінойс). Рослини, пропоновані у винаході, експресують поліпептид, що обумовлює толерантність до гліфосату, і щонайменше ще один поліпептид, що обумовлює толерантність до інгібітору ALS. Рослина, пропонована у винаході, володіє щонайменше однією поліпшеною властивістю в порівнянні з відповідною контрольною рослиною, таким, наприклад, як поліпшена толерантність до гербіцидів, знижене полягання, збільшена висота, знижений період часу до дозрівання й підвищена врожайність. Рослина має поліпшену властивість, коли вона статистично достовірно відрізняється від відповідної контрольної рослини, якщо це розходження відноситься до напрямку, що приводить до поліпшення в порівнянні з контрольною рослиною. Наприклад, рослина має поліпшену властивість, коли для неї характерна статистично достовірна підвищена врожайність у порівнянні з контрольною рослиною, і/або коли для неї характерне зниження ушкодження, пов'язаного з обробкою гербіцидом. Методики таких оцінок відомі в даній області. Можна застосовувати будь-який прийнятний статистичний аналіз, такий, наприклад, як дисперсійний аналіз (ANOVA) (який можна проводити з використанням вступника в продаж пакета програм фірми SAS Institute, Inc., 100 SAS Campus Drive, Кери, шт. Північна Кароліна). e. Рослини У контексті даного опису поняття «рослина» включає рослинні клітки, протопласти рослини, культури тканини рослинних кліток, з яких можна регенерувати рослину, рослинні калюси, маткове кореневище рослини, експлантати й рослинні кліт 27 ки, які перебувають в інтактномy стані в рослинах або частинах рослин, такі як зародки, пилок, насінняпочки, насіння, листи, квітки, галузі, плоди, зерна, качани, стрижні кукурудзяного качана, листові обгортки, стовбури, корінь, кінчики корінців, пильовики й т.п. Під зерном розуміють зрілі насіння, одержувані фермерами, які вирощують продукцію для ринку, для цілей, відмінних від вирощування або розмноження видів. Потомство, варіанти й мутанти регенерованих рослин також підпадають під об'єм винаходу, за умови, що ці частини містять інтродуковані полінуклеотиди. Таким чином, винахід відноситься до трансгених насінь, які продукують рослини, пропоновані у винаході. Даний винахід можна застосовувати для трансформації будь-яких видів рослин, включаючи (але, не обмежуючись ними) однодольні й двочасткові рослини. Прикладами видів рослин, що представляють інтерес, є (але, не обмежуючись ними ) кукурудза (Zea mays, що називають також «маїсом»), Brassica spp. (наприклад, В. napus, В. rара, В. juncea), насамперед види Brassica, які є джерелами масла, одержуваного з насінь (які називають також «канола»), льон (Linum spp.), люцерна (Medicago sativa), рис (Oryza sativa), жито (Secale cereale), сорго (Sorghum bicolor, Sorghum vulgare), просо (наприклад, просо американське (Pennisetum glaucum), просо культурне (Раnісum miliaceum), просо італійське (Setaria italica), просо пальчасте (Eleusine coracana), соняшник (Helianthus annum), сафлор (Carthamus tinctorius), пшениця (Triticum aestivum), соя (Glycine max), тютюн (Nicotiana tabacum), картопля (Solanum tuberosum), арахіс (Arachis hypogaea), бавовник (Gossypium barbadense, Gossypium hirsutum), солодка картопля (Ipomoea batatus), маніок (Manihot esculenta), кава (Coffea spp.), кокос (Cocos nucifera), ананас (Ananas comosus), цитрусові дерева (Citrus spp.), какао (Theobroma cacao), чай (Camellia sinensis), банан (Musa spp.), авокадо (Persea americana), смоківниця (Ficus casica), гуава (Psidium guajava), манго (Mangifera indica), олива (Olea europaea), папайя (Carica papaya), анакардія західна (Anacardium occidentale), макадамія (Macadamia integrifolia), мигдаль (Prunus amygdalus), цукровий буряк (Beta vulgaris), цукровий очерет (Saccharum spp.), вівси, ячмінь, овочеві культури, плодові культури, декоративні рослини (квіткові), цукровий очерет, хвойні дерева, Arabidopsis. Овочеві культури включають томати (Lycopersicon esculentum), салат (наприклад, Lactuca sativa), овочеву квасолю (Phaseolus vulgaris), квасолю Ліма (Phaseolus limensis), горох (Lathyrus spp.) і представників роду Cucumis, таких як огірок (С. sativus), канталупа (С. cantalupensis) і диня (С. melo). Декоративні культури включають азалію (Rhododendron spp.), гортензію (Macrophylla hydrangea), гибіскус (Hibiscus rosasanensis), троянди (Rosa spp.), тюльпани (Tulipa spp.), нарциси (Narcissus spp.), петунії (Petunia hybrida), гвоздику (Dianthus caryophyllus), пуансеттію (Euphorbia pulcherrima) і хризантему. 96578 28 Можна застосовувати також будь-які деревні рослини. Хвойні рослини, які можна застосовувати для втілення даного винаходу на практиці, включають, наприклад, соснові, такі як сосна ладанна (Pinus taeda), сосна їжакова (Pinus elliotii), сосна жовта (Pinus ponderosa), сосна скручена широкохвойна (Pinus contorta) і сосна чудова (Pinus radiata); дугласію (Pseudotsuga menziesu); тсугу західну (Tsuga canadensis); ялину ситхинську (Рісеа glauca); секвою вічнозелену (Sequoia sempervirens); справжні пихтові, такі як ялиця біла (Abies amabilis) і ялиця бальзамічна (Abies balsamea); і кедрові, такі як туя (Thuja plicata) і жовтий кедр Аляски (Chamaecyparis nootkatensis). Можна застосовувати також листяні дерева, такі як ясен, осика, бук, липа, береза, черемшина, горіх чорний, кінський каштан, каштан зубчастий, тополя волосистоплодний, дерен, в'яз, каркас, кария, гостролист, робинія, магнолія, клен, дуб, тополя, червона вільха, церцис, королівська пауловнія повстяна, сассафрас лікарський, амброзове дерево, явір, нисса, верба, липа американська. У конкретних варіантах здійснення винаходу рослини, пропоновані в даному винаході, являють собою культурні рослини (наприклад, кукурудзу (називану також «маїсом»), люцерну, соняшник, представників Brassica, сою, бавовник, сафлор, арахіс, сорго, пшеницю, просо, тютюн і т.д.). Інші цікаві рослини включають зернові культури, що дають насіння, олійні рослини й бобові рослини. Насіння, що представляють інтерес, включають насіння зернових культур, таких як кукурудза, пшениця, ячмінь, рис, сорго, жито й т.д. До олійних культур відноситься бавовник, соя, сафлор, соняшник, представники Brassica, маїс, люцерна, пальма, кокос і т.д. Бобові рослини включають боби й горохи. До бобового відносяться хьюар, ріжкове дерево, пажитник гречиський, соя, садові боби, горох коров'ячий, квасоля золотава, квасоля Ліма, кормові боби, сочевиця, нут і т.д. Інші цікаві рослини включають газонні трави, наприклад, газонні трави, що відносяться до родів Poa, Agrostis, Festuca, Lolium і Zoysia. Інші газонні трави можуть відноситься до підродини Panicoideae. До газонних трав відносяться також (але, не обмежуючись ними) грамова трава (Bouteloua gracilis (H.B.K.) Lag. Ex Griffiths); бізонова трава (Buchloe dactyloids (Nutt.) Engelm.); повзуча овсяница червона Слендера (Festuca rubra ssp. Litoralis); овсяница червона (Festuca rubra); мітлиця колоніальна (Agrostis tennis Sibth.); мітлиця болотна (Agrostis palustris Huds.); пирій пустельний (Agropyron cristatum (L.) Gaertn.); овсяница жосткувата (Festuca longifolia Thuill.); мятлик луговий (Poa pratensis L.); райграс пасовищний (Lolium perenne L.); мятлик звичайний (Poa trivialis L.); бутелоуа коротконизходящая (Bouteloua curtipendula Michx. Torr.); багаття безосний (Bromus inermis Leyss.); овсяница висока (Festuca arundinacea Schreb.); мятлик однолітній (Poa annua L.); райграс однолітній (Lolium multiflorum Lam.); мітлиця біла (Agrostis alba L.); зойсия японська (Zoysia japonica); свинорой пальчастий (бермудська трава) (Cynodon dactylon; Cynodon spp L.C. Rich; 29 Cynodon transvaalensis); мітлиця гречана (Paspalum vaginatum Swartz); зоисія (Zoysia spp Willd; Zoysia japonica і Ζ matrella var. matrella); гречка помітна (Paspalum notatum Flugge); аксонус чудесний (Axonopus affinis Chase); еремохлоа (Centipedegrass) (Eremochloa ophiuroides Munro Hack.); кикуйя (Pennisetum clandesinum Hochst Ex Chiov); мітлиця волосоподібна (Agrostis tenuis, відома також як A. capillaris); мітлиця собача (Agrostis canina); райграс пасовищний (Lolium perenne); і августинова трава (Stenotaphrum secundatum Walt. Kuntze). Інші цікаві трави включають лепешняк (Раnісum virgatum). ж. Угруповання ознак і додаткові ознаки, що представляють інтерес У деяких варіантах здійснення винаходу полінуклеотид, що обумовлює толерантність до гліфосату й толерантність до інгібіторів ALS у рослинах, пропонованих у винаході, вбудовують у групу молекул. В інших варіантах здійснення винаходу група молекул містить також щонайменше один додатковий полінуклеотид, що обумовлює толерантність до 3-тього гербіциду. Такі послідовності представлені в даному описі. В одному з варіантів здійснення винаходу послідовність обумовлює толерантність до глуфосинату, а в іншому конкретному варіанті здійснення винаходу послідовність містить доречну послідовність (пат). В інших варіантах здійснення винаходу рослини, толерантні до гліфосату/інгібітору ALS, пропоновані у винаході, містять одну або декілька інтерес ознак, що представляють інтерес і в більше конкретних варіантах здійснення винаходу послідовності рослини групують із будь-якою комбінацією полінуклеотидних послідовностей, що представляють інтерес, з метою створення рослин з необхідною комбінацією ознак. Поняття «ознака» у контексті даного опису відноситься до фенотипу, отриманому на основі конкретної послідовності або груп послідовностей. Наприклад, полінуклеотиди що обумовлюють толерантність до гербіцидів, можна групувати з будь-якими іншими полінуклеотидами, які кодують поліпептиди, що володіють пестицидною і/або інсектицидною активністю, такі як токсичні білки Bacillus thuringiensis (описані в U.S. №№ 5366892; 5747450; 5737514; 5723756; 5593,881; в Geiser і ін., Gene 48, 1986, с. 109; Lee і ін., Appl. Environ. Microbiol 69, 2003, cc. 4648-4657 (Vір3А); Galitzky і ін., Acta Crystallogr. D. Biol. Crystallogr. 57, 2001, cc. 1101-1109 (Сrу3Вb1); і Herman і ін., J. Agric. Food Chem. 52, 2004, cc. 2726-2734 (Cry1F)), лектини (Van Damme і ін., Plant Mol. Biol 24, 1994, с. 825, пентин (описаний в U.S. № 5981722) і т.п. Створені комбінації можна включати також у безліч копій кожного із являючих інтерес полінуклеотидів. В деяких варіантах здійснення винаходу толерантні до гербіцидів полінуклеотиди рослин, толерантних до гліфосату/інгібітору ALS (тобто такі рослини, які містять GAT і HRA), можна групувати з іншими полінуклеотидами, що обумовлюють ознаки толерантності до гербіцидів, для створення трансгеннщї рослини, пропонованої у винаході, з додатковими поліпшеними властивостями. Інші полінуклеотиди, зумовлюючі толерантність до гер 96578 30 біцидів, які можна застосовувати в таких варіантах здійснення винаходу, включають полінуклеотиди, що обумовлюють толерантність до гліфосату або інгібіторів ALS за допомогою інших механізмів дії, такі, наприклад, як ген, що кодує фермент гліфосат-оксидоредуктазу, що описано більш докладно в U.S. №№ 5776760 і 5463175. Інші ознаки, які можна поєднувати із зумовлюючими толерантність до гербіцидів полінуклеотидами, толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять такі послідовності, як GAT і HRA), включають ознаки, обумовлені полінуклеотидами, які надають рослині здатність продукувати, наприклад, високий рівень 5-єнолпіруватшикимат-3-фосфатсинтази (EPSPS), що більш докладно описано в U.S. №№ 6248876 В1; 5627061; 5804425; 5633435; 5145783; 4971908; 5312910; 5188642; 4940835; 5866775; 6225114 В1; 6130366; 5310667; 4535060; 4769061; 5633448; 5510471; Re. 36449; RE 37287 Ε і 5491288; і в опублікованих міжнародних заявках WO 97/04103; WO 00/66746; WO 01/66704; і WO 00/66747. Інші ознаки, які можна поєднувати із зумовлюючими толерантність до гербіцидів полінуклеотидами, толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять такі послідовності, як GAT і HRA), включають ознаки, обумовлені полінуклеотидами, які надають толерантність до сульфонілсечовини й/або імідазолінону, наприклад, описані більш докладно в U.S. №№ 5605011; 5013659; 5141870; 5767361; 5731180; 5304732; 4761373; 5331107; 5928937 і 5378824; і в опублікованій міжнародній заявці WO 96/33270. У деяких варіантах здійснення винаходи полінуклеотиди, зумовлюючі толерантність до гербіцидів, толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять такі послідовності, як GAT і HRA), можна групувати, наприклад, з гідроксифенілпіруватдіоксигеназами, які являють собою ферменти, каталізуючі реакцію, у якій wapaгідроксифенілпіруват (НРР) трансформується в гомогентисат. Молекули, які інгібують цей фермент і які зв'язуються з ферментом, інгібуючи трансформацію НРР у гомогентисат, можна застосовувати як гербіциди. Ознаки, що обумовлюють толерантність рослин до таких гербіцидів, описані в U.S. №№ 6245968 В1; 6268549; і 6069115; і в опублікованій міжнародній заявці WO 99/23886. Іншими прикладами придатних ознак толерантності до гербіцидів, які можна групувати із полінуклеотидами, зумовлюючими толерантність до гербіцидів, толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять такі послідовності, як GAT і HRA), включають ознаки, обумовлені полінуклеотидами арилоксиалканоат-діоксигенази (яка, як відомо, обумовлює толерантність до 2,4-Д і інших гербіцидів із класу феноксиауксинів, а також до арилоксифеноксипропіонатних гербіцидів, описаним, наприклад, в WO 2005/107437), і полінуклеотиди, що обумовлюють толерантність до дикамбе, що описано, наприклад, в Herman і ін., J. Biol. Chem 280, 2005, сс. 24759-24767. Другими прикладами придатних ознак толерантності до гербіцидів, які можна поєднувати із зумовлюючу толерантність до гербіцидів полінуклеотидами толерантних до гліфосату/інгібітору ALS 31 рослин (тобто рослин, що містять GAT і HRA), включають ознаки, обумовлені полінуклеотидами, які кодують екзогенну фосфінотрицинацетилтрансферазу, описану в U.S. №№ 5969213; 5489520; 5550318; 5874265; 5919675; 5561236; 5648477; 5646024; 6177616; і 5879903. Рослини, що містять екзогенну фосфінотрицинацетилтрансферазу, можуть мати підвищену толерантність до гербіцидів - похідних глуфосинату, які інгібують фермент глутамінсинтазу. Іншими прикладами придатних ознак толерантності до гербіцидів, які можна поєднувати із зумовлюючими толерантність до гербіцидів полінуклеотидами толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять GAT і HRA), включають ознаки, обумовлені полінуклеотидами, що кодують протопорфиногеноксидазу (протоке) зі зміненою активністю, які описані в U.S. №№ 6288306 В1; 6282837 В1 і 5767373; і в опублікованій міжнародній заявці WO 01/12825. Рослини, що містять такі полінуклеотиди, можуть мати підвищену толерантність до різних гербіцидів, мішенню яких є фермент протоке (які називають також «інгібіторами протокса»). Іншими прикладами придатних ознак толерантності до гербіцидів, які можна поєднувати із зумовлюючими толерантність до гербіцидів полінуклеотидами толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять GAT і HRA), включають ознаки, обумовлені полінуклеотидами, які надають толерантність щонайменше до одного гербіциду рослини, такого, наприклад, як рослина маїсу або мілкопелюк канадський. Толерантні до гербіцидів бур'янисті рослини відомі в даній області, і вони являють собою рослини з різною толерантністю до конкретних гербіцидів (див., наприклад, Green і Williams «Correlation of Corn (Zea mays) Inbred Response to Nicosulfuron and Mesotrione», 2004, постер, представлений на щорічному з'їзді WSSA у Канзас-Ситу, шт. Міссурі, 912 лютого, 2004 p.; Green, Weed Technology 12, 1998, cc. 474-477; Green і Ulrich, Weed Science 41, 1993. cc. 508-516). Ознака(і), відповідальна(і) за зазначену толерантність, можна поєднувати шляхом селекції або за допомогою інших методів із зумовлюючими толерантність до гербіцидів полінуклеотидами толерантних до гліфосату/інгібітору ALS рослин (тобто рослин, що містять GAT і HRA), з одержанням рослини, пропонованої у винаході, а також для здійснення способів їхнього застосування. Тaким чином, у винаході запропоновані рослини, які мають підвищену толерантність до гліфосату й іншим хімічним речовинам-інгібіторам ALS, а також рослини, які мають підвищену толерантність до гербіциду, до якого кожний з описаних вище ознак обумовлює толерантність. Так, у винаході запропоновані способи вирощування сільськогосподарських рослин (тобто для вибіркового контролю бур'янистої рослинності на оброблюваній посівній площі), що полягають у тому, що обробляють площу, що представляє інтерес, (наприклад, поле або оброблювану посівну площу) щонайменше одним гербіцидом, до якого рослина, пропонована у винаході, має толерантність, таким, наприклад, 96578 32 як гліфосат, хімічна сполука-інгібітор ALS, суміш хімічних сполук-інгібіторів ALS або суміш гліфосату й хімічної сполуки-інгібітору ALS. У деяких варіантах здійснення винаходу способи, пропоновані у винаході, додатково полягають у тому, що здійснюють обробку додатковими гербіцидами, до яких рослина, пропонована у винаході, має толерантність. У таких варіантах здійснення винаходу способи, пропоновані у винаході, як правило, забезпечують вибіркове знищення бур'янистої рослинності без помітного ушкодження сільськогосподарської культури. У контексті даного опису поняття «оброблювана посівна площа» відноситься до будь-якої площі, що підходить для вирощування рослини. Такі оброблювані посівні площі являють собою (але не обмежуючись ними) поле, на якому обробляють рослину (таке як поле, зайняте культурою, поклад, ділянку, засаджену деревами, окультурений ліс, поле для вирощування плодових культур і овочів і т.д.), теплицю, вегетаційну камеру й т.д. Для створення рослин, пропонованих у даному винаході, ознаки толерантності до гербіцидів можна поєднувати також щонайменше з однією іншою ознакою, що має на увазі також різні комбінації необхідних ознак, що включають (але, не обмежуючись ними) ознаки, необхідні для відгодівлі тварин, такі як високий вміст масла (див., наприклад, U.S. № 6232529); збалансований вміст амінокислот (наприклад, гордотіоніни (U.S. №№ 5990389; 5885801; 5885802 і 5703409; U.S. № 5850016); високий вміст лізину в ячмені (Williamson і ін., Eur. J. Biochem. 165, 1987, cc. 99-106 і WO 98/20122) і білки з високим вмістом метіоніну (Pedersen і ін., J. Biol. Chem. 261, 1986, с. 6279; Kirihara і ін., Gene 71, 1988, с. 359; і Musumura ін., Plant Mol. Biol. 12, 1989, с. 123)); підвищена засвоюваність (наприклад, модифіковані запасні білки (заявка на патент США сірий. № 10/053410, зареєстрована 7 листопада 2001 p.); і тіоредоксини (заявка на патент США, сірий. № 10/005429, зареєстрована 3 грудня 2001 р.)); опис яких включений в даний опис як посилання. Необхідні комбінації ознак включають також LLNC (низький вміст ліноленової кислоти; див., наприклад, Dyer і ін., Аррі. Microbiol. Biotechnol 59, 2002, cc. 224-230) і OLCH (високий вміст олеїнової кислоти; див., наприклад, Fernandez-Moya і ін., J. Agric. Food Chem 53, 2005, cc. 5326-5330). Ознаки, що представляють інтерес, толерантності до гербіцидів можна поєднувати також з іншими необхідними ознаками, такими, наприклад, як гени детоксифікації фумонисиму (U.S. № 5792931), гени авірулентності й гени, що обумовлюють стійкість до хвороб (Jones і ін., Science 266, 1994, с. 789; Martin і ін., Science 262, 1993, с. 1432; Mindrinos і ін., Cell 78, 1994, с. 1089), і ознаками, необхідними для переробки й обробки продуктів, такими як модифіковані масла (наприклад, гени десатурази жирних кислот (U.S. № 5952544; WO 94/11516)); модифіковані крохмалі (наприклад, ADPG-пірофосфосфорілази (AGPaзa), синтази крохмалю (SS), ферменти, каталізуючі розгалуження крохмалю (SBE), і ферменти, каталізуючі лінеаризацію (зниження розгалуженості) крохмалю 33 (SDBE)); і полімери або біопластики (див., наприклад, U.S. № 5602321; бета-кетотіолаза, полігідроксибутират-синтаза й ацетоацетил-СоА-редуктаза (Schubert і ін., J. Bacteriol. 170, 1988, cc. 58375847), полегшення експресії полігідроксиалканоатів (РНА)); опис яких включений в даний опис як посилання. Можна також поєднувати зумовлюючі толерантність до гербіцидів полінуклеотиди з полінуклеотидами, що обумовлюють агрономічні ознаки, такі як чоловіча стерильність (див., наприклад, U.S. № 5583210), міцність стебла, час цвітіння, або з ознаками, пов'язаними з технологією трансформації, такими як регуляція клітинного циклу або спрямований перенос генів (див., наприклад, WO 99/61619, WO 00/17364 і WO 99/25821); опис яких включений в даний опис як посилання. В іншому варіанті здійснення винаходу ознаки, що представляють інтерес, толерантності до гербіцидів можна поєднувати з послідовністю Rcgl або її біологічно активним варіантом або фрагментом. Послідовність Rcgl являє собою ген, що обумовлює стійкість до антракнозної стеблев гнилі кукурудзи (див., наприклад, US №№ 11/397153, 11/397275 і 11/397247 опис яких включений в даний опис як посилання). Ці об'єднані комбінації можна створювати за допомогою будь-якого методу, що включає (але не обмежуючись ними) селекцію рослин за допомогою будь-якої загальноприйнятої TopCrossметодології або генетичну трансформацію. Якщо послідовності згруповані шляхом генетичної трансформації рослин, то послідовності, що представляють інтерес, полінуклеотидні, можна поєднувати в будь-який час або в будь-якому порядку. Наприклад, трансгенна рослина, що містить один або кілька необхідних ознак, може застосовуватися як мішень для інтродукції додаткових ознак шляхом наступної трансформації. Ознаки можна інтродукувати одночасно відповідно до протоколу котрансформації із представляючими інтерес полінуклеотидами з використанням будь-якої комбінації касет, що трансформують. Наприклад, якщо потрібно інтродукувати дві послідовності, те дві послідовності можна включати в різні касети, що трансформують (транс) або в ту саму касету, що трансформує (цис). Експресія послідовностей може перебувати під контролем того самого промотору або різних промоторів. У деяких випадках може виявитися бажаним інтродукувати касету, що трансформує, що повинна придушувати експресію полінуклеотиду що представляє інтерес. Її можна поєднувати з будь-якою комбінацією інших гнітючих експресію касет або касет для надекспресії для створення необхідної комбінації ознак у рослині. Установлено також, що полінуклеотидні послідовності можна групувати в необхідному локусі генома за допомогою сайтспецифічної системи рекомбінації (див., наприклад, WO 99/25821, WO 99/25854, WO 99/25840, WO 99/25855 і WO99/25853, опис яких включений в даний опис як посилання). Різні представляючі інтерес зміни фенотипу включають модифікацію сполуки жирних кислот у рослині, зміна вмісту амінокислот у рослині, зміна 96578 34 механізму захисту рослини від патогенів і т.п. Для досягнення цього можна використати експресію гетерологічних продуктів або підвищену експресію ендогенних продуктів у рослинах. В іншому варіанті для досягнення зазначеного результату можна використати зниження рівня експресії в рослині одного або декількох ендогенних продуктів, зокрема, ферментів або кофакторів. Ці зміну приводять до зміни фенотипу трансформованої рослини. Гени, що представляють інтерес, визначаються ринком, інтереси якого враховуються при створенні сільськогосподарської культури. Оскільки сільськогосподарські культури, що представляють інтерес, і ринки змінюються, і нації, що розвиваються, створюють нові світові ринки, то необхідно створювати також нові сільськогосподарські культури й технології. Крім того, у міру того як загліблюються знання про агрономічні ознаки й характеристики, таких як підвищення врожайності й гетерозису, відповідно повинен змінюватися вибір генів для трансформації. Загальні категорії генів, що представляють інтерес, включають, наприклад, гени, які беруть участь у передачі інформації, такі як «цинкові пальці», які беруть участь у комунікації, такі як кінази, які беруть участь в «домашнім господарстві», наприклад, білки теплового шоку. Більше специфічні категорії трансгенів включають, наприклад, гени, що кодують важливі для агрономії ознаки, стійкість до комах, стійкість до хвороб, стійкість до гербіцидів, стерильність, характеристики зерна й призначених для продажу продуктів. Гени, що представляють інтерес, включають, як правило, гени, що беруть участь у метаболізмі масел, крохмалю, вуглеводів або живильних речовин, а також впливають на розмір зерен, рівень сахарози й т.п. Агрономічно важливі ознаки, такі як вміст масла, крохмалю й білків, можна змінювати генетично крім використання традиційних методів селекції. Модифікації включають підвищення вмісту олеїнової кислоти, насичених і ненасичених масел, підвищення рівнів лізину й сірки, введення незамінних амінокислот, а також модифікацію крохмалю. Похідні послідовностей, що кодують, можна створювати за допомогою сайтнаправленого мутагенезу, призначеного для підвищення рівня попередньо відібраних амінокислот у кодуємому поліпептиді. Наприклад, ген, що кодує поліпептид ячменя з високим вмістом лізину (BHL), одержують на основі інгібітору хімотрипсину ячменя (див. заявку на патент США, сірий. № 08/740682, зареєстровану 1 листопада 1996 p., і WO 98/20133, опис яких включений в даний опис як посилання). Інші білки являють собою богаті метіоніном рослинні білки, наприклад, з насінь соняшника (Lilley і ін., Proceedings of the World Congress on Vegetable Protein Utilization in Human Foods and Animal Feedstuffs, під ред. Applewhite, з American Oil Chemists Society, Champaign, Illinois, 1989, cc. 497502; публікація включена в даний опис як посилання); кукурудзи (Pedersen і ін., J. Biol. Chem. 261, 1986, с. 6279; Kirihara і ін., Gene 71, 1988, с. 359; обидві публікації включені в даний опис як посилання); і рису (Musumura і ін., Plant Mol. Biol 12, 1989, с.123, публікація включена в даний опис як 35 посилання). Інші агрономічно важливі гени кодують латекс, Floury 2, фактори росту, фактори, пов'язані зі зберіганням насінь, і фактори транскрипції. Гени, що обумовлюють стійкість до комах, кодують стійкість до шкідників, які викликають значну втрату врожаю, таких як, тих, що ушкоджують корінь, личинки, гусеніци подгризаючих совок, метелик кукурудзяний і т.п. Такі гени включають, наприклад, гени токсичного білка Bacillus thuringiensis (U.S. №№ 5366892; 5747450; 5736514; 5723756; 5593881; і Geiser і ін., Gene 48, 1986, с. 109) і т.п. Гени, що кодують такі ознаки, як стійкість до хвороб, являють собою детоксифікуючі гени, наприклад, гени проти фумоносина (U.S. № 5792931); гени авірулентності (avr) і стійкості до хвороб (R) (Jones і ін., Science 266, 1994, с. 789; Martin і ін., Science 262, 1993, с.1432; і Mindrinos і ін., Cell 78, 1994, с.1089) і т.п. Зумовлюючі стерильність гени можуть також кодуватися експресійними касетами і являти собою альтернативу фізичному видаленню мітелок. Прикладами застосовуваних для цієї мети генів є гени, специфічні для чоловічих тканин, і гени, що обумовлюють фенотип чоловічої стерильності, такі як QM, описані в U.S. № 5583210. Іншими генами є гени кіназ і гени, що кодують сполуки, що виявляються токсичну дію при розвитку або чоловічого, або жіночого гаметофіта. На якість зерна впливають такі ознаки, як рівні й типи масел, насичених і ненасичених, якість і кількість незамінних амінокислот, і рівні целюлози. Для кукурудзи описані модифіковані білки хордотіоніни, див. U.S. №№ 5703049, 5885801, 5885802 і 5990389. Ознаки, важливі з комерційної точки зору, можна також кодувати геном або генами, які можуть підвищувати, наприклад, виробництво крохмалю або етанолу, або забезпечують експресію білків. Іншим важливим з комерційної точки зору застосуванням трансформованих рослин є виробництво полімерів або біопластиків, наприклад, описане в U.S. № 5602321. Такі гени, як гени -кетотіолази, РНВази (полігідроксибутират-синтази) і ацетоацетил-СоА-редуктази (див. Schubert і ін., J. Bacteriol 170, 1988, сс. 5837-5847), полегшують експресію полігідроксиалканоатів (РНА). До екзогенних продуктів відносяться ферменти й продукти рослин, а також продукти з інших джерел, включаючи прокариотичні й інші еукаріотичні організми. До таких продуктів відносяться ферменти, кофактори, гормони й т.п. Можна підвищувати рівень білків, насамперед модифікованих білків з поліпшеною амінокислотною сполукою, з метою підвищення живильної цінності рослини. Для досягнення цієї мети використовують експресію зазначених білків з підвищеним вмістом амінокислот. II. Конструкції полінуклеотидів У конкретних варіантах здійснення винаходу один або декілька зумовлюючих толерантність до гербіцидів полінуклеотидів (толерантних до гербіцидів полінуклеотидів), застосовуваних у способах і композиціях, може(уть) перебувати в касеті експресії, призначеної для експресії в рослинах, що 96578 36 представляють інтерес, або інших організмах. Касета повинна включати регуляторні 5' - і 3'послідовності, функціонально пов'язані із зумовлюючим толерантність до гербіциду полінуклеотидом. Поняття «функціонально зв'язаний» відноситься до функціонального зв'язку між двома або більшою кількістю елементів. Наприклад, функціональний зв'язок між представляючим інтерес полінуклеотидом і регуляторною послідовністю (наприклад, промотором) являє собою функціональний зв'язок, що забезпечує експресію полінуклеотиду, що представляє інтерес. Функціонально зв'язані елементи можуть бути суміжними або несуміжними. Відносно стику двох областей, що кодують, білка поняття «функціонально зв'язані» означає, що області, що кодують, перебувають в одній і тій же рамці зчитування. Відносно дії енхансера поняття «функціонально зв'язаний» означає, що енхансер підсилює експресію конкретного полінуклеотиду, що представляє інтерес або полінуклеотидів. Якщо що представляючий(і) інтерес полінуклеотид або полінуклеотиди кодує(ють) поліпептид, то рівень виробництва кодованого поліпептиду підвищується. Касета може містити також щонайменше один додатковий ген для котрансфомації організму. В іншому варіанті додатковий(і) ген(и) може(уть) перебувати в декількох касетах експресії. Таку касету експресії постачають безліччю сайтів рестрикції й/або сайтів рекомбінації для вбудовування зумовлюючу толерантність до гербіциду полінуклеотидів, регуляція транскрипції яких забезпечується регуляторними областями. Касета експресії додатково може містити інші гени, включаючи інші гени селектованих маркерів. Якщо касета містить більше одного полінуклеотиду, то полінуклеотиди в касеті можуть транскрибуватися в тому самому або в різних напрямках (що називають також «дивергентною» транскрипцією). Касета експресії, що містить зумовлюючий толерантність до гербіциду полінуклеотид, повинна містити в напрямку 5'>3' транскрипції область ініціації транскрипції й трансляції (тобто промотор), полінуклеотид, що обумовлює толерантність до гербіциду (наприклад, полінуклеотид GAT, полінуклеотид, що обумовлює толерантність до інгібітору ALS, полінуклеотид HRA або їхню будь-яку комбінацію й т.д.), ділянку терминації транскрипції й трансляції (тобто термінуючу дялінку), які мають функціональну активність в інших рослинах, що представляють інтерес, і організмах. Таким чином, запропоновані також рослини, що містять такі касети експресії. Регуляторні ділянки (тобто промотори, дялінки, що регулюють транскрипцію, і ділянки терминації трансляції) і/або зумовлюючий толерантність до гербіциду полінуклеотид можуть бути нативними (тобто аналогічними) клітціхазяїнові або один одному. В іншому варіанті регуляторні області й/або зумовлюючий толерантність до гербіциду полінуклеотид, пропонований у винаході, може бути гетерологічен клітці-хазяїнові або один одному. У контексті даного опису поняття «гетерологічний» відносно послідовності означає, що послідовність отримана із чужорідного організму, або, якщо її одержують із цього ж організму, то 37 вона в значній мірі модифікована відносно її нативної форми по сполуці й/або в геномном локусі в результаті втручання людини. Наприклад, промотор, функціонально пов'язаний з гетерологічним полінуклеотидом, одержують із видів, відмінних від видів, з яких отриманий полінуклеотид, або, якщо їх одержують із тих самих (тобто аналогічних) видів, те один з них або обоє піддають значної модифікації щодо їхньої вихідної форми й/або генного локусу, або промотор не є нативним промотором для функціонально пов'язаного з ним полінуклеотиду. Хоча для експресії полінуклеотидів оптимальним є застосування гетерологічних промоторів, можна використати нативні промоторні послідовності. Такі конструкції можуть змінювати рівні експресії й/або схеми експресії кодованого поліпептиду в рослині або рослинній клітці. Рівні експресії й/або схеми експресії кодованого поліпептиду можна змінювати також з використанням додаткового регуляторного елемента, що є частиною конструкції, такого, наприклад, як енхансер. Таким чином, фенотип рослини або клітки можна змінювати, навіть при використанні нативного промотору. Термінуюча ділянка може бути нативною щодо ділянки ініціації транскрипції, може бути нативної відносно функціонально зв'язаного полінуклеотиду, що представляє інтерес, зумовлюючого толерантність до гербіциду, може бути нативною щодо рослини-хазяїна, або її можна одержувати з іншого джерела (тобто вона може бути чужорідної або гетерологичною) щодо промотору, полінуклеотиду, що представляє інтерес, що обумовлює толерантність до гербіциду, рослини-хазяїна або будь-якої їхньої комбінації. Прийнятними терминирующімі є ділянки з Ті-плазміди А. tumefaciens, такі як термінуючі ділянки октопинсинтази й нопалінсинтази, або їх можна одержувати з генів рослин, таких як ген інгібітору II Solarium tuberosum (див. також Guerineau і ін., Mol. Gen. Genet 262, 1991, cc. 141144; Proudfoot, Cell 64, 1991, cc. 671-674; Sanfacon і ін., Genes Dev 5, 1991, cc. 141-149; Mogen і ін., Plant Cell 2, 1990, cc. 1261-1272, Munroe і ін., Gene 91, 1990, cc. 151-158; Ballas і ін., Nucleic Acids Res 17, 1989, cc. 7891-7903; і Joshi і ін., Nucleic Acids Res 15, 1987, cc. 9627-9639. Для втілення на практиці винаходу можна застосовувати ряд промоторів, включаючи нативний промотор полінуклеотидної послідовності, що представляє інтерес. Промотори можна вибирати залежно від необхідного результату. Представляючі інтерес полінуклеотиди можна поєднувати з конститутивними, кращими для конкретного типу тканини (тканьоспецифічними) або іншими промоторами для експресії в рослинах. До таких конститутивних промоторів відносяться, наприклад коровий промотор промотору Rsyn7 і інші конститутивні промотори, описані в WO 99/43838 і U.S. № 6072050; коровий промотор CaMV 35S (Odell і ін., Nature 313, 1985, cc. 810812); промотор актину рису (McElroy і ін., Plant Cell 2, 1990, cc. 163-171); і промотор актину кукурудзи; промотор убікитину (див., наприклад, Christensen і ін., Plant Mol. Biol. 12, 1989, cc. 619-632; Christensen і ін., Plant Mol. Biol. 18, 1992, сс. 675 96578 38 689; Callis і ін., Genetics 139, 1995, сс. 921-39); pEMU (Last і ін., Theor. Appl. Genet. 81, 1991, сс. 581-588); MAS (Velten і ін., ЕМВО J. 3, 1984, сс. 2723-2730); промотор ALS (U.S. № 5659026) і т.п. До інших конститутивних промоторів відносяться, наприклад, промотори, описані в U.S. №№ 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463; 5608142 і 6177611. Деякі промотори підвищують рівень експресії при їхньому застосуванні в сполученні з нативною 5'нетрансльованою ділянкою й/або іншими елементами, такими, наприклад, як інтрон. Наприклад, промотор убікитину кукурудзи часто поміщають проти ходу транскрипції щодо полінуклеотиду, що представляє інтерес поряд щонайменше із частиною 5'-нетрансльованої ділянки гена убікитину, включаючи перший інтрон гена убікитину кукурудзи. Регульовані хімічними речовинами промотори можна застосовувати для модуляції експресії гена в рослині з використанням екзогенного хімічного регулятора. Залежно від завдання промотор може являти собою індукований хімічною речовиною промотор, при використанні якого обробка хімічною речовиною індукує експресію гена, або промотор може являти собою промотор, що придули хімічною речовиною, при використанні якого обробка хімічною речовиною придушує експресію гена. Індуковані хімічними речовинами промотори відомі в даній галузі й включають (але не обмежуючись ними) промотор Іn2-2 кукурудзи, що активується антидотами бензолсульфонамідних гербіцидів, промотор GST кукурудзи, що активується гідрофобними електрофільними сполуками, які застосовують у якості передвсходових гербіцидів, і промотор PR-1a тютюну, що активується саліциловою кислотою. Іншими хімічними речовинами регульованими промоторами, що представляють інтерес, є реагуючі на стероїди промотори (див., наприклад, дані про індукований глюкокортикоїдами промотор, представлені в Schena і ін., Proc. Natl. Acad. Sci. USA 88, 1991, сс. 10421-10425 і McNellis і ін., Plant J. 14(2), 1998, сс. 247-257) і індуковані тетрацикліном промотори (див., наприклад, Gatz і ін., Mol. Gen. Genet. 227, 1991, сс. 229-237 і U.S. №№ 5814618 і 5789156), які включені в даний опис як посилання. Тканьоспецифічні промотори можна застосовувати для цілеспрямованого забезпечення підвищеного рівня експресії зумовлюючого толерантність до гербіцидів поліпептиду в конкретній тканині рослини. Тканеспецифічні промотори описані в Yamamoto і ін., Plant J. 12(2), 1997, cc. 255265; Kawamata Plant Cell Physiol. 38(7), 1997, cc. 792-803; Hansen і ін., Mol. Gen Genet. 254(3), 1997, cc. 337-343; Russell і ін., Transgenic Res. 6(2), 1997, cc. 157-168; Rinehart і ін., Plant Physiol. 112(3), 1996, cc. 1331-1341; Van Camp і ін., Plant Physiol. 112(2), 1996, cc. 525-535; Canevascini і ін., Plant Physiol. 112(2), 1996, cc. 513-524; Yamamoto і ін., Plant Cell Physiol. 35(5), 1994, cc. 773-778; Lam, Results Probl. Cell Differ. 20, 1994, cc. 181-196; Orozco і ін., Plant Mol Biol. 23(6), 1993, cc. 11291138; Matsuoka і ін., Proc Natl. Acad. Sci. USA 90(20), 1993, cc. 9586-9590 і Guevara-Garcia і ін., 39 Plant J. 4(3), 1993, cc. 495-505. Такі промотори при необхідності можна модифікувати для ослаблення експресії. У даній галузі відомі кращі для листів промотори (див., наприклад, Yamamoto і ін., Plant J. 12(2), 1997, cc. 255-265; Kwon і ін., Plant Physiol. 105, 1994, cc. 357-367; Yamamoto і ін., Plant Cell Physiol. 35(5), 1994, cc. 773-778; Gotor і ін., Plant J. 3, 1993, cc. 509-518; Orozco і ін., Plant Mol. Biol. 23(6), 1993, cc. 1129-1138 і Matsuoka і ін., Proc. Natl. Acad. Sci. USA 90(20), 1993, cc. 9586-9590). У даній галузі відомі кращі для кореня промотори і їх можна вибирати з відомих з літератури промоторів або виділених de novo промоторів з різних сумісних видів (див., наприклад, Hire і ін., Plant Mol. Biol. 20(2), 1992, cc. 207-218 (специфічний для кореня ген глутамінсинтетази); Keller і Baumgartner, Plant Cell 3(10), 1991, cc. 1051-1061 (специфічний для кореня контролюючий елемент у гені GRP 1.8 квасолі звичайної); Sanger і ін., Plant Mol. Biol. 14(3), 1990, cc. 433-443 (специфічний для кореня промотор гена маннопинсинтази (MAS) Agrobacterium tumefaciens); і Міао і ін., Plant Cell 3(1), 1991, cc. 11-22 (клон повнорозмірной кДНК, що кодує цитозольну глутамінсинтетазу (GS), що експресуєся в коріннях і кореневих вузликах сої); див. також Bogusz і ін., Plant Cell 2(7), 1990, cc. 633-641, у цій публікації описані два специфічних для кореня промотору, виділених з генів гемоглобіну азотфіксуючого не належачого до бобової рослини Parasponia andersonii і родинного неазотфіксуючого не належачого до бобової рослини Trema tomentosa). Промотори цих генів зв'язували з репортерним геном -глюкуронідази й інтродукували як у не належачого до бобової рослини Nicotiana tabacum, так і в належачого до бобової рослини Lotus corniculatus, і в обох випадках зберігалася специфічна для кореня промоторна активність. Leach і Aoyagi, 1991 описали аналіз промоторів, індукуючих у корені володіючі високим рівнем експресії гени rol і rol Agrobacterium rhizogenes (див. Plant Science (Limerick) 79(1), cc. 69-76). Ці дослідники встановили, що в цих промоторах відбувається дисоціація енхансера й тканеспецифічних детермінант ДНК. Teeri і ін., 1989, використали генне злиття з lac для доказу того, що ген Т-ДНК Agrobacterium, що кодує октопинсинтазу, що має особливо високу активність в епідермісі верхівки кореня, і ген TR2', що є специфічним для кореня в інтактній рослині й стимулюється ушкодженням листової тканини, являють собою особливо кращу комбінацію характеристик для застосування в сполученні з інсектицидним або ларвицидним геном (див. ЕМВО J. 8(2), сс. 343350). Ген TR1', злитий з nptll (неоміцинфосфотрансфераза II), має аналогічні характеристики. Додаткові кращі для кореня промотори являють собою промотор гена VfENOD-GRP3 (Kuster і ін., Plant Mol. Biol 29(4), 1995, сс. 759-772); і промотор rol (Сараnа і ін., Plant Mol. Biol 25(4), 1994, сс. 681-691 (див. також U.S. №№ 5837876; 5750386; 5633363; 5459252; 5401836; 5110732 і 5023179). «Кращі для насіння» промотори являють собою як «специфічні для насіння» промотори (ці 96578 40 промотори мають активність під час розвитку насіння, такі як промотори запасних білків насіння), а також «пов'язані із проростанням насінь» промотори (ці промотори мають активність у процесі проростання насіння) (див. Thompson і ін., BioEssays 10, 1989, с. 108), публікація включена в даний опис як посилання). До таких кращих для насіння промоторів відносяться (але не обмежуючись ними) Сіm1і (індукований цитокініном перенос); с19В1 (зеїн кукурудзи розміром 19 кДа); milps (міо-інозит-1-фосфатсинтаза) (див. WO 00/11177 і U.S. № 6225529; які включені в даний опис у якості посилання). ГамМа-зеїн являє собою специфічний для ендосперму промотор. Глобулін 1 (Glb-1) являє собою репрезентативний специфічний для зародка промотор. Для двочасткових рослин специфічні для насіння промотори являють собою (але не обмежуючись ними) -фазеолін квасолі, напин, -конгліцинин, лектин сої, круциферин і т.п. Для однодольних рослин специфічні для насіння промотори являють собою (але не обмежуючись ними) зеїн кукурудзи з молекулярною масою 15 кДа, 22 кДа-зеїн, 27 кДа-зеїн, гамМа-зеїн, «воскової» (waxy), «зморщений» (shrunken 1), shrunken 2, глобин 1 і т.д. (див. також WO 00/12733, у якій описані кращі для насіння промотори генів end1 і end2; яка включена в даний опис як посилання). Другими промоторами, що представляють інтерес, є промотор SCP1 (US № 6072050), промотор НВ2 (US № 6177611) і промотор SAMS (US 20030226166 і SEQ ID NO:87 і його біологічно активні варіанти й фрагменти); кожна з публікацій включена в даний опис як посилання. Крім того, як зазначено в даному описі, у сполученні із зазначеними промоторами можна застосовувати енхансери, такі, наприклад, як інтрон убікитину (тобто інтрон 1 убікитину кукурудзи (див., наприклад, NCBIпослідовність S94464), омега-енхансер або омегаприм-енхансер (Gallie і ін., Molecular Biology of RNA под. ред Cech (Liss, New York), 1989, с.з. 237256 і Gallie і ін., Gene, 60, 1987, cc. 217-25), або енхансер 35S; кожна з публікацій включена в даний опис як посилання. Касета експресія може містити також ген селетуємого маркера для селекції трансформованих кліток. Гени селектованих маркерів застосовують для селекції трансформованих кліток або тканин. Маркерні гени являють собою гени, що кодують стійкість до антибіотиків, такі як кодують неоміцинфосфотрансферазу II (ΝΕΟ) і гігроміцинфосфотрансферазу (НРТ), а також гени, що обумовлюють стійкість до гербіцидів, таким як глуфосинатамоній, бромоксиніл. імідазолінони й 2,4дихлорфеноксиацетат (2,4-Д). До інших селектованих маркерів відносяться фенотипічні маркери, такі як -галактозидаза й флуоресцентні білки, такі як зелений флуоресцентний білок (GFP) (Su і ін., Biotechnol Bioeng 85, 2004, cc. 610-619 і Fetter і ін., Plant Cell 16, 2004, cc. 215-228), ціановий флуоресцентний білок (CYP) (Bolte і ін., J. Cell Science 117, 2004, cc. 943-954 і Kato і ін., Plant Physiol 129, 2002, cc. 913-942) і жовтий флуоресцентний білок (PhiYFP фірми Evrogen, див. Bolte і ін., J. Cell Science 117, 2004, cc. 943-54). Дані про інших селектованих маркери представлені в цілому в 41 Yarranton, Curr. Opin. Biotech. З, 1992, cc. 506-511; Christopherson і ін., Proc. Natl. Acad. Sci. USA 89, 1992, cc. 6314-6318; Yao і ін, Cell 71, 1992, cc. 6372; Reznikoff, Mol. Microbiol. 6, 1992, cc. 2419-2422; Barkley і ін. в: The Operon, 1980, cc. 177-220; Ни і ін., Cell 48, 1987, cc. 555-566; Brown і ін., Cell, 49, 1987, сс. 603-612; Figge і ін., Cell 52, 1988, cc. 713722; Deuschle і ін., Proc. Natl. Acad. Асі. USA 86, 1989, сс. 5400-5404; Fuerst і ін., Proc. Natl. Acad. Sci. USA 86, 1989, cc. 2549-2553; Deuschle і ін., Science 248, 1990, cc. 480-483; Gossen, 1993, Ph.D. Thesis, University of Heidelberg; Reines і ін., Proc. Natl. Acad. Sci. USA 90, 1993, cc. 1917-1921; Labow і ін., Mol. Cell. Biol. 10, 1990, cc. 3343-3356; Zambretti і ін., Proc. Natl. Acad. Sci. USA 89, 1992, cc. 3952-3956; Bairn і ін., Proc. Natl. Acad. Sci. USA 88, 1991, cc. 5072-5076; Wyborski і ін., Nucleic Acids Res. 19, 1991, cc. 4647-4653; Hillenand-Wissman, Topics Mol. Struc. Biol. 10, 1989, cc. 143-162; Degenkolb і ін., Antimicrob. Agents Chemother. 35, 1991, cc. 1591-1595; Kleinschnidt і ін., Biochemistry 27, 1988, cc. 1094-1104; Bonin, 1993, Ph.D. Thesis, University of Heidelberg; Gossen і ін., Proc. Natl. Acad. Sci. USA 89, 1992, cc. 5547-5551; Oliva і ін., Antimicrob. Agents Chemother 36, 1992, cc. 913-919; Hlavka і ін., Handbook of Experimental Pharmacology, т. 78, з Springer-Verlag, Berlin, 1985; Gill і ін., Nature 334, 1988, cc. 721-724. Ці публікації включені в даний опис як посилання. Перерахований вище перелік генів селектованих маркерів не слід розглядати як обмежуючий об'єм винаходу. У даному винаході можна застосовувати будь-який ген селетуємого маркера, включаючи ген GAT і/або ген HRA. У даній галузі відомі методи підвищення рівня експресії поліпептиду, пропонованого у винаході, у рослині або рослинній клітці, наприклад, шляхом вбудовування в послідовність, що кодує поліпептид, одного або двох багатих G/C кодонів (таких як GCG або GCT), що безпосередньо прімікають і перебувають у напрямку по ходу транскрипції щодо ініціюючого кодона метіоніну ATG. При необхідності полінуклеотиди можна оптимізувати для підвищення експресії в трансформованій рослині. Це означає, що можна синтезувати полінуклеотиди із заміною в послідовності, що кодує, поліпептиду одного або декількох кодонів, які являють собою рідше, що зустрічаються в рослин кодони, що кодують одну(і) і ту ж(ті ж) амінокислоту(и), що найбільше часто зустрічається(ються) у рослинах, і інтродукувати модифіковану послідовність, що кодує, у рослину або рослинну клітку й експресувати модифіковану послідовність, що кодує (див., наприклад, в Campbell і Gowri, Plant Physiol. 92, 1990, cc. 1-11 обговорення кращих для хазяїв найбільше що часто зустрічаються кодонів). У даній галузі відомі методи синтезу найбільш кращих для рослини генів (див., наприклад, U.S. №№ 5380831 і 5436391, і Murray і ін., Nucleic Acids Res. 17, 1989, cc. 477-498, які включені в даний опис як посилання). Варіанти таких модифікацій є також відмітною ознакою винаходу. Відомі інші модифікації послідовностей, що дозволяють підсилювати експресію гена в клітціхазяїні. Вони включають елімінацію послідовнос 96578 42 тей, що кодують помилкові сигнали поліаденилювання, сигнали екзон-інтронних сайтів сплайсинга, повтори типу транспозона й інші добре охарактерізовані послідовності, які можуть робити «шкідливий» вплив на експресію генів. Вміст G-C у послідовності можна регулювати до досягнення середніх рівнів, характерних для даної кліткихазяїна, шляхом розрахунку в порівнянні з відомими генами, які експресуються в клітці-хазяіні. Якщо цей можливо, то послідовність модифікують для того, щоб уникнути присутності передвіщених вторинних структур мРНК у вигляді «шпильок». Можна застосовувати також «енхансери», такі як енхансер CaMV 35S (див., наприклад, Benfey і ін., ЕМВО J. 9, 1990, сс. 1685-1696), або можна використати інші енхансери. Наприклад, можна застосовувати послідовність, представлену в SEQ ID NO:1, 72, 79, 84, 85, 88 або 89, або її біологічно активний варіант або фрагмент (див. також заявку на патент на винахід США No. ____, озаглавлену «Methods and Compositions for the Expression of a Polynucleotide of Interest», подану одночасно з даною заявкою й повністю включеною в неї як посилання. Поняття «промотор» означає регуляторну ділянку ДНК, що включає ділянку ініціації транскрипції, що у деяких варіантах містить Бокс-ТАТАбокс, що володіє здатністю «спонукувати» РНКполімеразу II ініціювати синтез РНК у відповідному сайті ініціації транскрипції для конкретної послідовності, що кодує. Промотор може також бути функціонально пов'язаний з додатковими регуляторними елементами, які впливають на транскрипцію, включаючи (але не обмежуючись ними) інтрони, 5'нетрансльовані ділянки й енхансерні елементи. У контексті даного опису мається на увазі, що «енхансерна послідовність», «енханерний домен», або «енхансерний елемент» або «енхансер», коли він функціонально пов'язаний з відповідним промотором, може модулювати рівень транскрипції функціонально зв'язаного полінуклеотиду, що представляє інтерес. Біологічно активні фрагменти й варіанти енхансерного домена можуть зберігатибіологічну активність, що модулює (підвищувальну або понижающую) рівень транскрипції, коли вони функціонально пов'язані з відповідним промотором. Полінуклеотидні фрагменти енхансерного домена або промотору можуть складатися щонайменше приблизно з 50 нуклеотидів, приблизно з 100 нуклеотидів, приблизно з 150 нуклеотидів, приблизно з 200 нуклеотидів, приблизно з 250 нуклеотидів, приблизно з 300 нуклеотидів, приблизно з 350 нуклеотидів, приблизно з 400 нуклеотидів, приблизно з 450 нуклеотидів, приблизно з 500 нуклеотидів і для енхансерного домена, пропонованого у винаході, аж до повнорозмірной нуклеотидної послідовності, пропонованої у винаході. В інших варіантах здійснення винаходу фрагмент енхансерного домена має довжину від приблизно 50 до приблизно 100, від 100 до приблизно 150, від 150 до приблизно 200, від 200 до приблизно 250, від приблизно 250 до приблизно 300, від приблизно 300 до приблизно 350, від приблизно 350 до приблизно 400, від приблизно 400 до приблизно 450, від приблизно 450 до приблизно 500, від приблиз 43 но 500 до приблизно 535 нуклеотидів. Як правило, послідовності варіантів конкретних полінуклеотидів, пропонованих у винаході, повинні бути щонайменше приблизно на 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% або більше ідентичні послідовності конкретних полінуклеотидів, що визначають за допомогою програм для порівняльного аналізу первинної структури послідовностей і параметрів, представлених у даному описі. Біологічно активний варіант енхансера або промотору може відрізнятися від послідовності лише 1-15 залишками нуклеїнової кислоти, лише 1-10, наприклад 6-10, лише 10, 9, 8, 7, 6, 5, 4, 3, 2 залишками або навіть 1 залишком нуклеїнової кислоти. Такі активні варіанти й фрагменти повинні зберігати здатність модулювати транскрипцію. Безліч копій енхансерного домена або його активних варіантів або фрагментів можна функціонально зв'язувати із промотором. У конкретних варіантах здійснення винаходу химерна ділянка, що здійснює регуляторний контроль транскрипції, містить щонайменше 1, 2, 3, 4, 5, 6, 7 або більше копій енхансерного домена. В інших варіантах здійснення винаходу застосовуваний енхансерний домен не містить послідовність, представлену в SEQ ID NO:5. Крім того, енхансер може перебувати в будь-якій орієнтації (тобто смисловій або антисмисловій (зворотної)). Відстань між промотором і енхансерим доменом може варіюватися в межах, при яких химерна ділянка, що здійснює регуляторний контроль транскрипції, продовжує регулювати транскрипцію функціонально зв'язаного полінуклеотиду, що представляє інтерес необхідним чином. Наприклад, енхансерний домен може перебувати на відстані щонайменше від приблизно 10000 до приблизно 15000, від приблизно 10000 до приблизно 9000, від приблизно 9000 до приблизно 8000, від приблизно 8000 до приблизно 7000, від приблизно 7000 до приблизно 6000, від приблизно 6000 до приблизно 5000, від приблизно 5000 до приблизно 4000, від приблизно 4000 до приблизно 3000, від приблизно 3000 до приблизно 2000, від приблизно 2000 до приблизно 1000, від приблизно 1000 до приблизно 500, від приблизно 500 до приблизно 250, від приблизно 250 нуклеотидів до положення, що безпосередньо прімікає до промотору. Установлено також, що одну або кілька копій енхансера можна поміщати проти ходу транскрипції (5') щодо промотору або в іншому варіанті одна або кілька копій енхансера може бути локалізована в 3'напрямку щодо промотору. У конкретних варіантах здійснення винаходу при локалізації в 3'-напрямку щодо промотору енхансер розташований по ходу транскрипції щодо сайту терминації. В інших варіантах здійснення винаходу один або декілька енхансерів можуть перебувати або в 5'>3'-орієнтації (як показане в SEQ ID NO:1 або 72), або в 3'>5'орієнтації. Якщо застосовують декілька енхансерів, то енхансери можна поміщати в конструкції в таке положення щодо промотору, щоб досягати необхідного впливу на експресію. Наприклад, енхансери можуть безпосередньо прімікати друг до друга 96578 44 або між ними може перебувати щонайменше від 1 до 100, від 100 до 300, від 300 до 500, від 500 до 1000 нуклеотидів. Установлено також, що енхансер, застосовуваний відповідно до винаходу, можна розташовувати в конструкції ДНК між першим і другим промотором і у функціональному зв'язку з ними. У таких варіантах здійснення винаходу енхансер дозволяє модулювати експресію в протилежних напрямках як щодо першого, так і другого промотору. Наприклад, такі конструкції ДНК містять (але не обмежуючись ними) в 5'>3'- або в 3'>5'напрямку: перший полінуклеотид, що представляє інтерес, функціонально пов'язаний з першим промотором, що функціонально зв'язаний щонайменше з однією копією енхансера, пропонованого у винаході, що функціонально зв'язаний із другим промотором, що функціонально пов'язаний із другим представляючим інтерес полінуклеотидом. У конкретних варіантах здійснення винаходу енхансерна послідовність є гетерологичною щодо першої й другий енхансерної послідовності. В інших варіантах здійснення винаходу перший промотор функціонально пов'язаний з полінуклеотидом, що кодує інгібітор ALS, а другий промотор функціонально зв'язаний полінуклеотидом, що кодує поліпептид, що обумовлює толерантність до гліфосату. Такі полінуклеотиди представлені в даному описі. Касета експресії може містити додатково 5'лідерні послідовності. Функцією таких лідерних послідовностей може бути посилення трансляції. Лідери трансляції відомі в даній галузі і являють собою: лідери пикорновірусів, наприклад, лідер EMCV (5'-ділянка, що некодує, вірусу енцефаломіокардита) (Elroy-Stein і ін., Proc. Natl. Acad. Sci. USA 86, 1989, cc. 6126- 6130); лідери потивірусів, наприклад, лідер TEV (вірус тютюнового гравірування) (Gallie і ін., Gene 165(2), 1995, сс. 233-238), лідер MDMV (вірус строкатої карликовости кукурудзи) (Virology 154, сс. 9-20) і лідер білка, що зв'язує важкий ланцюг людського імуноглобуліну (Ві) (Macejak і ін., Nature 353, 1991, сс. 90-94); нетрансльований лідер мРНК оболонкового білка вірусу мозаїки люцерни (AMV RNA 4) (Jobling і ін., Nature 325, 1987, сс. 622-625); лідер вірусу тютюнової мозаїки (TMV) (Gallie i ін., в: Molecular Biology of RNA, під ред. Cech (Liss, New York), 1989, cc. 237256); і лідер вірусу хлорозної плямистості кукурудзи (MCMV) (Lommel і ін., Virology 81, 1991, сс. 382-385), див., також Della-Cioppa і ін., Plant Physiol. 84, 1987, сс. 965-968. При створенні касети експресії можна маніпулювати з різними полінуклеотидними фрагментами для того, що послідовності перебували в правильній орієнтації й при необхідності в правильній рамці зчитування. Для цієї мети можна застосовувати адаптери або лінкери для зв'язку фрагментів або можна здійснювати інші маніпуляції для одержання прийнятних сайтів рестрикції, видалення надлишкового матеріалу, наприклад, можна видаляти сайти рестрикції або т.п. Для цієї мети можна використати мутагенез in vitro, праймерную репарацію, рестрикцію, віджиг, повторні заміни, наприклад, трансзиції й трансверсії. Стандартні методи рекомбінантної ДНК і молекулярного клонування, 45 застосовувані при створенні даного винаходу, добре відомі в даній галузі й докладно описані, наприклад, в Sambrook і ін., Molecular Cloning: A Laboratory Manual, з Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1989, (керівництво відомо також за назвою «Maniatis»). У деяких варіантах здійснення винаходу здійснюють спрямований перенос полінуклеотиду, що представляє інтерес в хлоропласт для експресії. При такій методиці, якщо полінуклеотид, що представляє інтерес, не убудований безпосередньо в хлоропласт, то касета експресії повинна додатково містити нуклеїнову кислоту, що кодує транзитний пептид для переносу генного продукту, що представляє інтерес, у хлоропласти. Такі транзитні пептиди відомі в даній галузі (див., наприклад, Von Heijne і ін., Plant Mol. Biol. Rep. 9, 1991, сс. 104126; Clark і ін., J. Biol. Chem. 264, 1989, сс. 1754417550; Della-Cioppa і ін., Plant Physiol. 84, 1987, сс. 965-968; Romer і ін., Biochem. Biophys. Res. Comraun. 196, 1993, сс. 1414-1421 і Shah і ін., Science 233, 1986, сс. 478-481). Послідовності, що забезпечують спрямований перенос у хлоропласт, відомі в даній галузі й до них відносяться хлоропластна мала субодиниця рібулозо-1,5-біфосфаткарбоксилази (Rubisco) (de Castro Silva Filho і ін., Plant Mol. Biol. 30, 1996, сс. 769-780; Schnell і ін., J. Biol. Chem. 266(5), 1991, сс. 3335-3342); 5-(єнолпірувіл)шикимат-3-фосфатсинтаза (EPSPS) (Archer і ін., J. Bioenerg. Biomemb. 22(6), 1990, сс. 789-810); триптофансинтаза (Zhao і ін., J. Biol. Chem. 270(11), 1995, сс. 6081-6087); пластоцианін (Lawrence і ін., J. Biol. Chem. 272(33), 1997, сс. 20357-20363); хоризматсинтаза (Schmidt і ін., J. Biol. Chem. 268(36), 1993, сс. 27447-27457) і, що зв'язується з а, що запасає світлову енергію хлорофілом,/b білок (LHBP) (Lamppa і ін., J. Biol. Chem. 263, 1988, сс. 1499614999) (див. також Von Heijne і ін., Plant Mol. Biol. Rep. 9, 1991, сс. 104-126; Clark і ін., J. Biol. Chem. 264, 1989, сс. 17544-17550; Della-Cioppa і ін., Plant Physiol. 84, 1987, сс. 965-968; Romer і ін., Biochem. Biophys. Res. Commun. 196, 1993, сс. 1414-1421 і Shah і ін., Science 233, 1986, сс. 478-481). Методи трансформації хлоропластів добре відомі в даній галузі (див., наприклад, Svab і ін., Proc. Natl. Acad. Sci. USA 87, 1990, сс. 8526-8530; Svab і Maliga, Proc. Natl. Acad. Sci. USA 90, 1990, cc. 913-917; Svab і Maliga, EMBO J. 12, 1993, cc. 601-606). Метод заснований на введенні за допомогою генної рушниці часток ДНК, що містять селектований маркер і забезпечують спрямований перенос ДНК у пластидний геном шляхом гомологичной рекомбінації. Крім того, можна здійснювати пластидну трансформацію шляхом трансактивації присутнього у пластиді трансгена за допомогою тканеспецифічної експресії кодуємої у ядрі й спрямованої до пластид РНК-полімерази. Така система описана в McBride і ін., Proc. Natl. Acad. Sci. USA 91, 1994, cc. 7301-7305. Експресію представляющих інтерес полінуклеотидів, які підлягають спрямованому переносу в хлоропласт, можна оптимізувати для здійснення експресії в хлоропласті з урахуванням розходжень у найбільше що часто зустрічаються кодонах між 96578 46 ядром рослини й цієї органеллой. При такій методиці полшуклеотид, що представляє інтерес, можна синтезувати з використанням кращих для хлоропласта кодонів (див., наприклад, U.S. №5380831, включений у даний опис як посилання). Поняття «ген» відноситься до полінуклеотиду, що експресує специфічний білок, як правило, що включає регуляторні послідовності, розташовані перед (5'-послідовності, що некодують) і розташовані після (3'-послідовності, що некодують) послідовності, що кодує (тобто ділянки послідовності, що кодує специфічний білок). Поняття «нативний ген» відноситься до гена, що зустрічається в природних умовах, як правило, з його власними регуляторними послідовностями. «Трансген» означає ген, що інтродукували в геном шляхом процедури трансформації. Таким чином, «трансгенна рослина» являє собою рослине, що містить трансген, де трансген інтродукували в конкретну рослину шляхом трансформація або селекції; так, під це визначення підпадають нащадки спочатку трансформованої рослини. III. Методи інтродукції Рослини, пропоновані у винаході, створюють шляхом інтродукції поліпептиду або полінуклеотиду в рослину. Поняття «інтродукція» означає вбудовування в рослину полінуклеотиду або поліпептиду таким чином, що послідовність перебуває усередині клітки рослини. Способи, пропоновані у винаході, не повинні залежати від конкретного методу інтродукції послідовності в рослину, вони тільки повинні забезпечувати вбудовування полінуклеотиду або поліпептиду усередину щонайменше однієї клітки рослини. Методи інтродукції полінуклеотидів або поліпептидів у рослини відомі в даній галузі й включають (але, не обмежуючись ними) методи стабільної трансформації, методи короткочасної трансформації, методи опосередковуваної вірусом трансформації й селекцію. Поняття «стабільна трансформація» означає, що відбувається інтеграція нуклеотидної конструкції, інтродукованої у рослину в геном рослини, і вона може передаватися в спадщину його потомству. Поняття «короткочасна трансформація» означає, що полінуклеотид інтродукують у рослину й не інтегрують у геном рослини, або що в рослину інтродукують поліпептид. Протоколи трансформації, а також протоколи інтродукції поліпептидних або полінуклеотидних послідовностей у рослини можуть варіюватися залежно від типу рослини або рослинної клітки (тобто однодольна або двочасткова рослина), що підлягає трансформації. Прийнятними методами інтродукції поліпептидів і полінуклеотидів у рослинну клітку є мікроін'єкція (Crossway і ін., Biotechniques 4, 1986, сс. 320-334), електропорація (Riggs і ін., Proc. Natl. Acad. Sci. USA 83, 1986, сс. 5602-5606, опосередковувана Agrobacterium трансформація (U.S. № 5563055 і U.S. № 5981840), безпосередній перенос генів (Paszkowski і ін., ЕМВО J. З, 1984, сс. 2717-2722) і балістичне прискорення часток (див., наприклад, U.S. № 4945050; U.S. № 5879918; U.S. №№ 5886244 і 5932782; Tomes і ін., в: Plant Cell, Tissue, and Organ Culture: 47 Fundamental Methods, під ред. Gamborg and Phillips, з Springer-Verlag, Berlin, 1995; McCabe і ін., Biotechnology 6, 1988, сс. 923-926); і Lec1трансформація (WO 00/28058). Методи трансформації описані також в Weissinger і ін., Ann. Rev. Genet. 22, 1988, сс. 421-477; Sanford і ін., Particulate Science and Technology 5, 1987, сс. 2737 (лук); Christou і ін., Plant Physiol. 87, 1988, сс. 671-674 (соя); McCabe і ін., Bio/Technology 6, 1988, сс. 923-926 (соя); Finer і McMullen, In Vitro Cell Dev. Biol. 27P, 1991, cc. 175-182 (соя); Singh і ін., Theor. Appl. Genet. 96, 1998, cc. 319-324 (соя); Datta і ін., Biotechnology 8, 1990, cc. 736-740 (рис); Klein і ін., Proc. Natl. Acad. Sci. USA 85, 1988, cc. 4305-4309 (кукурудза); Klein і ін. Biotechnology 6, 1988, cc. 559-563 (кукурудза); U.S. №№ 5240855; 5322783 і 5324646; Klein і ін., Plant Physiol. 91, 1988, cc. 440444 (кукурудза); Fromm і ін., Biotechnology 8, 1990, cc. 833-839 (кукурудза); протоколи, опубліковані в електронному виді на сайті «IP.com» під постійними ідентифікаторами публікацій IPCOM000033402D, IPCOM000033402D і IPCOM000033402D і доступні на сайті в мережі Інтернет «IP.com» (бавовник); Hooykaas-Van Slogteren і ін., Nature (London) 311, 1984, сс. 763764; U.S. №№ 5736369 (злаки); Bytebier і ін., Proc. Natl. Acad. Sci. USA 84, 1987, cc. 5345-5349 (лілейні); De Wet і ін. в: The Experimental Manipulation of Ovule Tissues, під ред. Chapman і ін., з Longman, New York, 1985, cc 197-209 (пилок); Kaeppler і ін., Plant Cell Reports 9, 1990, cc. 415-418 і Kaeppler і ін., Theor. Appl. Genet. 84, 1992, cc. 560-566 (трансформація, здійснювана за допомогою пензлика); D'Halluin і ін., Plant Cell 4, 1992, сс. 1495-1505 (електропорація); Li і ін., Plant Cell Reports 12, 1993, сс. 250-255 і Christou і Ford, Annals of Botany 75, 1995, cc. 407-413 (рис); Osjoda і ін., Nature Biotechnology 14, 1996, cc. 745-750 (кукурудза за допомогою Agrobacterium tumefaciens); всі ці публікації включені в даний опис як посилання. У конкретних варіантах здійснення винаходу, що обумовлюють толерантність до гербіцидів або інші необхідні послідовності можна вносити в рослину за допомогою різних методів короткочасної трансформації. Такі методи короткочасної трансформації включають (але, не обмежуючись ними) інтродукцію поліпептидів або його варіантів або фрагментів безпосередньо в рослину або інтродукцію транскрипта в рослину. Такі методи включають, наприклад, мікроін'єкцію або бомбардування частками (див, наприклад, Crossway і ін., Mol Gen. Genet. 202, 1986, cc. 179-185; Nomura і ін., Plant Sci. 44, 1986, cc. 53-58; Hepler і ін., Proc. Natl. Acad. Sci. 91, 1994, cc. 2176-2180 і Hush і ін., The Journal of Cell Science 107, 1994, cc. 775-784, всі ці публікації включені в даний опис як посилання. В іншому варіанті здійснення винаходу зумовлюючим толерантність до гербіциду полінуклеотидом можна короткочасно трансформувати рослину за допомогою відомих у даній галузі методик. Такі методики засновані на використанні вірусної векторної системи й на осадженні полінуклеотиду таким чином, щоб запобігати наступне вивільнення ДНК. Так, транскрипція пов'язаної із часткою ДНК може відбуватися, але частота, з якої відбу 96578 48 вається її вивільнення для інтеграції в геном, різко знижується. Такі методи передбачають застосування часток, на яке нанесене покриття з поліетиленіміну (РЕІ; фірма Sigma № Р3143). В інших варіантах здійснення винаходу полінуклеотиди можна інтродукувати у рослини, приводячи рослини в контакт із вірусом або вірусними нуклеїновими кислотами. Як правило, такі методи передбачають включення нуклеотидної конструкції в молекулу вірусної ДНК або РНК. Відомо, поліпептид, що представляє інтерес, можна синтезувати спочатку у вигляді частини вірусного поліпротеїну, що потім можна процесувати шляхом протеолізу in vivo або in vitro з одержанням необхідного білка. Крім того, установлено, що придатні промотори можуть являти собою промотори, застосовувані для транскрипції за допомогою вірусних РНКполімераз. Методи інтродукції полінуклеотидів з використанням молекул вірусної ДНК або РНК у рослини й експресії кодованих ними поліпептидів відомі в даній галузі (див., наприклад, U.S. №№ 5,889191, 5889190, 5866785, 5589367, 5316931 і Porta і ін., Molecular Biotechnology 5, 1996, cc. 209221; які включені в даний опис як посилання). У даній галузі відомі методи цілеспрямованої інсерції полінуклеотиду в певне положення в рослинному геномі. В одному з варіантів здійснення винаходу для інсерції полінуклеотиду в необхідне положення в геномі застосовують сайтепецифічну систему рекомбінації (див., наприклад, WO 99/25821, WO 99/25854, WO 99/25840, WO 99/25855 і WO 99/25853, які всі включені в даний опис як посилання). У цілому, полінуклеотид може перебувати в касеті для переносу, фланкований двома не придатними для рекомбінації (нерекомбіногенними) сайтами рекомбінації. Касету для переносу інтродукують у рослину, у геном якого стабільно включений сайт-мішень, що фланкований двома нерекомбіногенними сайтами рекомбінації, які відповідають сайтам касети для переносу. Застосовують відповідну рекомбіназу й касету для переносу інтегрують у сайт-мішень. Тим самим відбувається інтеграція полінуклеотиду, що представляє інтерес, в певне положення на хромосомі рослинного генома. Трансформовані клітки можна вирощувати з одержанням рослин за допомогою загальноприйнятих методів (див., наприклад, McCormick і ін., Plant Cell Reports 5, 1986, cc. 81-84). Потім ці рослини можна вирощувати й запилювати або тією же самою трансформованою лінією, або іншими лініями, і в потомстві, що утворилося, ідентифікувати конститутивну експресію необхідної фенотипічної ознаки. Можна вирощувати два або більша кількості покоління для гарантії того, що необхідна фенотипічна ознака стабільно підтримується й передається в спадщину, і потім збирають насіння для гарантії того, що досягнуто появу необхідної фенотипічної ознаки. Таким чином, даний винахід відноситься до трансформованого насіння (позначеному також як «трансгенне насіння»), що несе полінуклеотид, пропонований у винаході, наприклад, касету експресії, пропоновану у винаході, стабільно включену в його геном. 49 У конкретних варіантах здійснення винаходу поліпептид або полінуклеотид, що представляє інтерес, інтродукують у рослинну клітку. Потім рослинну клітку, у яку інтродукована послідовність, пропонована у винаході, відбирають у допомогою методів, відомих фахівцеві в даній галузі, таких як (але, не обмежуючись ними) аналіз методом Саузерн-блотингу, секвенування ДНК, ПЦР-аналіз або фенотипічний аналіз. Рослину або рослинну клітку, змінену або модифіковану відповідно до вищевикладених варіантів здійснення винаходу, вирощують в умовах, що дозволяють утворитися рослині, протягом проміжку часу, достатнього для модуляції концентрації й/або активності поліпептидів у рослині. Умови, що дозволяють утворитися рослині, добре відомі в даній галузі й у цілому будуть описані нижче. Установлено також, що рівень і/або активність інтерес, що представляє, поліпептиду можна модулювати за допомогою полінуклеотиду, що не має здатність до безпосереднього контролю в трансформованій рослині експресії білка або РНК. Наприклад, полінуклеотиди, пропоновані у винаході, можна застосовувати для створення полінуклеотидних конструкцій, які можна застосовувати в методах зміни або мутації геномной нуклеотидної послідовності в організмі. Такі полінуклеотидні конструкції включають (але, не обмежуючись ними) вектори РНК:ДНК, мутаційні вектори РНК:ДНК, репараційні вектори РНК:ДНК, змішані дуплексні олігонуклеотиди, самокомплементарні олігонуклеотиди РНК:ДНК і рекомбіногенні олігонуклеотидні основи. Такі нуклеотидні конструкції й методи їхнього застосування відомі в даній галузі (див. U.S. №№ 5565350; 5731181; 5756325; 5760012; 5795972 і 5871984; які всі включені в даний опис як посилання). Вони описані також в WO 98/49350, WO 99/07865, WO 99/25821 і в Beetham і ін., Proc. Natl. Acad. Sci. USA 96, 1999, cc. 8774-8778; які включені в даний опис як посилання. При створенні винаходу було встановлено, що способи, пропоновані в даному винаході, не залежать від включення повного полінуклеотиду в геном, якщо тільки рослина або його клітка не змінюються в результаті інтродукції полінуклеотиду в клітку. В одному з варіантів здійснення винаходу геном може змінюватися після інтродукції полінуклеотиду в клітку. Наприклад, полінуклеотид або його будь-яку частину можна включати в геном рослини. Зміни генома, пропоновані в даному винаході, включають (але, не обмежуючись ними) додавання, делеції й заміни нуклеотидів у геномі. Хоча способи, пропоновані в даному винаході, не залежать від додавань, делецій і замін будь-якої конкретної кількості нуклеотидів, при створенні винаходу встановлено, що такі додавання, делеції або заміни зачіпають щонайменше один нуклеотид. Рослини, пропоновані у винаході, можна одержувати будь-яким прийнятним методом, включаючи селекцію. Селекцію рослин можна застосовувати для інтродукції необхідних ознак (наприклад, стабільно включений трансген або генетичний варіант або генетична зміна, що представляє інтерес) у конкретну лінію, що представляє інтерес, 96578 50 рослини, і її можна здійснювати за допомогою кожного з декількох відомих шляхів. Чистопорідну селекцію починають зі схрещування двох генотипів, що представляють інтерес, таких як елітна лінія й одна інша елітна інбредна лінія, що несе один або кілька необхідних ознак (тобто в яку стабільно включений полінуклеотид, що представляє інтерес, що має змінену активність і/або рівнем експресії інтерес, що представляє, поліпептиду й т.д.), що(е) доповнює(ють) елітну лінію, що представляє інтерес, рослини. Якщо два вихідних батьки не несуть всі необхідні ознаки, то для селекції запиленням можна використати інші джерела. При використанні методу родовідної селекції рослинам вищої якості дають самозапилюватися й відбирають наступні одне за іншим дочірні покоління. У послідовних дочірніх поколіннях умова гетерозиготности дозволяє одержувати гомозиготні лінії в результаті самозапилення й селекції. Як правило, при використанні методу родовідної селекції дають самозапилюватися п'яти або більшій кількості послідовних дочірніх поколінь і здійснюють наступний шлях селекції: F1F2; F2F3; F3F4; F4F5 і т.д. Після здійснення достатньої кількості близкородинних схрещувань послідовні дочірні покоління можуть служити для створення інбредної лінії з підвищеним виробництвом насінь. У конкретних варіантах здійснення винаходу інбредна лінія несе гомозиготні алелі в приблизно 95% локусів або більше. У даній галузі відомі численні методики, які можна застосовувати для полегшення й прискорення процесу селекції (наприклад, поворотне схрещування), включаючи, наприклад, застосування теплиці або вегетативної камери з подовженими циклами день/ніч, аналіз молекулярних маркерів для ідентифікації необхідного потомства й т.п. Поповз застосування поворотного схрещування для створення конверсії, беккросування можна застосовувати також у сполученні із чистопорідною селекцією для модифікації елітної лінії, що представляє інтерес, і гібрида, отриманого з використанням модифікованої елітної лінії. Як обговорювалося раніше, поворотне схрещування можна застосовувати для переносу одного або декількох конкретних необхідних ознак з однієї лінії, батькадонора, в інбредну лінії, називану рекурентним батьком, що має в цілому гарні агрономічні характеристики, але в якої відсутній(ні) бажана ознака або ознаки. Однак таку ж процедуру можна застосовувати для вбудовування генотипу нащадка в генотип рекуррентного батька, але який у той же час зберігає багато компонентів нерекуррентного батька в результаті припинення процесу поворотного схрещування на ранній стадії й продовження процесу самозапилення й селекції. Наприклад, створюють гібрид першого покоління (F1), такий як призначений для продажу гібрид. Цей призначений для продажу гібрид можна піддавати поворотному схрещуванню з однієї з його батьківських ліній, одержуючи ВС1 або ВС2. Потомству дають самозапилюватися й відбирають, у результаті знову створена інбредна лінія має багато ознак рекуррентного батька, а також ще кілька необхідних ознак нерекуррентного батька. Цей підхід підви 51 щує цінність і силу рекуррентного батька, що дозволяє застосовувати його для створення нових гібридів і селекції. Таким чином, одним з варіантів здійснення даного винаходу є спосіб одержання конверсії на основі поворотного схрещування інбредної лінії, що представляє інтерес, що полягає в тому, що схрещують рослину інбредної лінії, що представляє інтерес, з рослиною-донором, що містить щонайменше один мутантний ген або трансген, що обумовлює необхідну ознаку (наприклад, толерантність до гербіцидів), відбирають рослину з F1покоління, що містить мутантний ген або трансген, що обумовлює необхідну ознаку, і здійснюють поворотне схрещування відібраної рослини F1покоління з рослиною інбредної лінії, що представляє інтерес. Цей спосіб може додатково включати стадію одержання профілю молекулярного маркера інбредної лінії, що представляє інтерес, і застосування профілю молекулярного маркера для відбору потомства рослини з необхідною ознакою й профілем молекулярного маркера інбредної лінії, що представляє інтерес. Зазначений спосіб можна використати також для одержання насіння гібрида F1 шляхом введення кінцевої стадії схрещування необхідної конверсії ознаки інбредної лінії, що представляє інтерес, з іншою рослиною для одержання насіння гібрида F1, що містить мутантний ген або трансген, що обумовлює необхідна ознака. Рекуррентна селекція являє собою метод, застосовуваний у програми селекції рослин для поліпшення популяції рослин. Метод полягає в перехресному запиленні індивідуальних рослин один одним з одержанням потомства. Потомство вирощують і потомство вищої якості, включаючи індивідуальну рослину, полусибси, повні сибси, потомство, отримане в результаті самозапилення, і топкроси, відбирають за допомогою кожного із численних методів селекції. Здійснюють перехресне запилення відібраного потомства один одним, одержуючи потомство іншої популяції. Цю популяцію висаджують і знову рослини вищої якості відбирають для перехресного запилення один одним. Рекуррентна селекція являє собою циклічний процес і її можна повторювати необхідну кількість разів. Метою рекуррентнох селекції є поліпшення ознак популяції. Поліпшену популяцію потім можна використати як джерело матеріалу для селекції для одержання інбредних ліній, які можна застосовувати в гібридах або як батьків для штучного культивара. Штучний культивар являє собою потомство, отримане в результаті интеркросса декількох відібраних инбредов. Масова селекція являє собою коштовний метод, коли його використовують у сполученні із селекцією, посиленої відбором молекулярного маркера. При масовій селекції насіння окремих рослин відбирають на основі фенотипу й/або генотипу. Ці відібрані насіння потім поєднують і застосовують для вирощування наступного покоління. Метод відбору «Akarp» (один з методів масової селекції) вимагає вирощування популяції рослин на ділянках великого розміру, що дозволяє рослинам самозапилюватися, зібрані насіння поєднують у бункері й потім використовують зразок 96578 52 насінь, зібраних у бункер, для вирощування рослини наступного покоління. Як частина цієї програми селекції можна використати замість самозапилення пряме запилення. Відбір мутацій є одним із численних методів, які можна застосовувати для інтродукції нових ознак в елітну лінію. Рослинник-селекціонер може використати спонтанні мутації або штучно індуковані мутації як джерело розмаїтості. Метою штучного мутагенезу є підвищення частоти мутації, що приводить до одержання необхідної ознаки. Частоту мутації можна підвищувати за допомогою численних різних шляхів (факторів), включаючи температуру, тривале зберігання насінь, умови культивування тканини, опромінення, наприклад рентгенівськими променями, гама-променями (наприклад, кобальтом 60 або цезієм 137), нейтронами (продукт ядерного розпаду урану 235 в атомному реакторі), бета-випромінювання (випромінюване радіоізотопами, такими як фосфор 32 або вуглець 14) або ультрафіолетове випромінювання (переважно при довжині хвилі від 2500 до 2900 нм), або хімічні мутагени (такі як основні аналоги (5-бромурацил), родинні сполуки (8етоксикофеїн)), антибіотики (стрептонигрин), алкілуючі агенти (сульфовані похідні гірчичного газу, азотсодержащие похідні гірчичного газу, епоксиди, етиленаміни, сульфати, сульфонати, сульфони, лактони), азиди, гідроксиламін, азотиста кислота або акридини. Після того, як необхідна ознака отримана в результаті мутагенезу, ознаку можна включати в існуючу зародкову плазму за допомогою традиційних методів селекції, таких як поворотне схрещування. Докладно відбір мутацій описаний в: «Principals of Cultivar Development» Fehr, з Macmillan Publishing Company, 1993, публікація включена в даний опис як посилання. Крім того, мутації, створені в інших лініях, можна застосовувати для одержання конверсії, створеної шляхом поворотного схрещування, елітних ліній, які несуть таку мутацію. IV. Методи модуляції експресії У деяких варіантах здійснення винаходу модулюють активність і/або рівень експресії поліпептиду (тобто підвищують або знижують). Можна досягати підвищення рівня й/або активності поліпептиду, вбудовуючи поліпептид у рослину. Як описано нижче, у даній галузі відомо багато методів вбудовування поліпептиду в рослину, які включають (але, не обмежуючись ними) безпосередню інтродукцію поліпептиду в рослину, інтродукцію в рослину (короткочасно або стабільно) полшуклеотидної конструкції, що кодує поліпептид, що володіє необхідною активністю. Установлено також, що в способах, пропонованих у винаході, можна застосовувати полінуклеотид, що не має здатність у трансформованій рослині контролювати експресію білка або РНК. Таким чином, рівень і/або активність поліпептиду можна модулювати, змінюючи ген, що кодує поліпептид, або його промотор (див., наприклад, Kmiec, U.S. № 5565350; Zarling і ін., PCT/US93/03868). Таким чином, описані мутантні рослини, які несуть мутації гена, де мутації підвищують рівень експресії гена або підвищують активність кодованого поліпептиду. 53 В інших варіантах здійснення винаходу активність і/або рівень поліпептиду знижують або елімінують шляхом інтродукції в рослину полінуклеотиду, що інгібує рівень або активність поліпептиду. Полінуклеотид може інгібувати експресію поліпептиду безпосередньо шляхом припинення трансляції відповідної матричної РНК або побічно шляхом кодування поліпептиду, що інгібує транскрипцію або трансляцію гена, що кодує білок. Методи інгібування або елімінації експресії гена в рослині добре відомі в даній галузі, і будь-які такі методи можна застосовувати згідно із даним винаходом для інгібування експресії гена в рослині. В інших варіантах здійснення винаходу активність поліпептиду знижують або елімінують шляхом трансформації рослинної клітки послідовністю, що кодує поліпептид, інгібуючий активність поліпептиду. В інших варіантах здійснення винаходу активність поліпептиду можна знижувати або елімінувати шляхом порушення гена, що кодує поліпептид. Під об'єм винаходу підпадають мутантні рослини, що несуть мутації в гені, що представляє інтерес, де мутації знижують експресію гена або інгібують активність кодованого поліпептиду. Зниження активності специфічних генів (яку називають також мовчанням гена або супресією гена) потрібно для рішення деяких аспектів створення рослин методом генної інженерії. Багато методик, що приводять до одержання генів, що мовчать, відомі фахівцеві в даній галузі, включаючи (але, не обмежуючись ними) антисмислову технологію (див., наприклад, Sheehy і ін., Proc. Natl. Acad. Sci. USA 85, 1988, cc. 8805-8809; і U.S. №№ 5107065; 5453566 і 5759829); косупресію (див., наприклад, Taylor, Plant Cell 9, 1997, с. 1245; Jorgensen, Trends Biotech. 8(12), 1990, cc. 340-344; Flavell, 1988, cc. 91, 1994, cc. 3490-3496; Finnegan і ін., Bio/Technology 12, 1994, cc. 883-888 і Neuhuber і ін., Mol. Gen. Genet. 244, 1994, cc. 230-241); інтерференцію РНК (Napoli і ін., Plant Cell 2, 1990, cc. 279-289; U.S. № 5034323; Sharp, Genes Dev. 13, 1999, cc. 139-141; Zamore і ін., Cell 101, 2000, cc. 25-33 і Montgomery і ін., Proc. Natl. Acad. Sci. USA 95, 1998, cc. 15502-15507), індуковане вірусом мовчання гена (Burton і ін., (2000) Plant Cell 12, 2000, cc. 691-705 і Baulcombe, Curr. Op. Plant Bio. 2, 1999, cc. 109-113); специфічні для РНК-мішені рібозімі (Haseloff і ін., (1988) Nature 334, 19888, cc. 585-591); структури типу «шпильки» (Smith і ін., (2000) Nature 407, 2000, сс. 319-320; WO 99/53050; WO 02/00904; WO 98/53083; Chuang і Meyerowitz, Proc. Natl. Acad. Sci. USA 97, 2000, сс. 4985-4990; Stoutjesdijk і ін., Plant Physiol. 129, 2002, сс. 17231731; Waterhouse і Helliwell, Nat. Rev. Genet. 4, 2003, cc. 29-38; Pandolfini і ін., ВМС Biotechnology 3, с. 7, опублікований патент США № 20030175965; Panstruga і ін., Mol. Biol. Rep. 30, 2003, сс. 135-140; Wesley і ін., Plant J. 27, 2001, сс. 581-590; Wang і Waterhouse, Сurr. Opin. Plant Biol. 5, 2001, сс. 146-150; опублікований патент США № 20030180945; і WO 02/00904, всі публікації включені в даний опис як посилання); рібозімі (Steinecke і ін., ЕМВО J. 11, 1992, с 1525 і Perriman і ін., Antisense Res. Dev. 3, 1993, сс. 253); цілеспрямовану опосередковувану олігонуклеотидом 96578 54 модифікацію (див., наприклад, WO 03/076574 і WO 99/25853); молекули, мішенню яких є «Zn-пальці» (див., наприклад, WO 01/52620; WO 03/048345 і WO 00/42219); транспозонове мічення (Maes і ін., Trends Plant Sci. 4, 1999, сс. 90-96; Dharmapuri і Sonti, FEMS Microbiol. Lett. 179, 1999, cc. 53-59; Meissner і ін., Plant J. 22, 2000, cc. 265-274; Phogat і ін., J. Biosci. 25, 2000, cc. 57-63; Walbot, Curr. Opin. Plant Biol. 2, 2000, cc. 103-107; Gai і ін., Nucleic Acids Res. 28, 2000, cc. 94-96; Fitzmaurice і ін., Genetics 153, 1999, cc. 1919-1928; Bensen і ін., Plant Cell 7, 1995, cc. 75-84; Mena і ін., Science 274, 1996, cc. 1537-1540 і U.S. № 5962764); кожна з публікацій включена в даний опис як посилання; і інші методи або комбінації вищевказаних методів, які відомі фахівцям у даній галузі. Установлено, що можна створювати антисмислові конструкції, комплементарні щонайменше частини матричної РНК (мРНК) полінуклеотиду, що представляє інтерес. Антисмислові нуклеотиди конструюють для гібридизації з відповідної мРНК. Можна створювати модифікації антисмислових послідовностей, якщо послідовності гібридизуються й впливають на експресію відповідної мРНК. У такому методі можна застосовувати антисмислові конструкції, послідовність яких ідентична щонайменше на 70%, оптимально на 80%, більш оптимально на 85% відповідним антисмисловим послідовностям. Крім того, можна застосовувати фрагменти антисмислових нуклеотидів для порушення експресії генів-мішені. Як правило, можна застосовувати послідовності, що складаються щонайменше з 50 нуклеотидів, 100 нуклеотидів, 200 нуклеотидів, 300, 400, 450, 500, 550 або більшої кількості нуклеотидів. Можна застосовувати також полінуклеотиди, що перебувають у смисловій орієнтації, для придушення експресії ендогенних генів у рослинах. Методи придушення експресії генів у рослинах з використанням полінуклеотидів у смисловій орієнтації, відомі в даній галузі. Методи, як правило, включають трансформацію рослин конструкцією ДНК, що містить промотор, що забезпечує експресію в рослині, функціонально зв'язаний щонайменше із частиною полінуклеотиду, що відповідає транскрипту ендогенного гена. Звичайно така нуклеотидна послідовність практично ідентична послідовності транскрипта ендогенного гена, як правило, більш ніж приблизно на 65%, 85% або 95% (див., U.S. №№ 5283184 і 5034323; які включені в даний опис як посилання). Таким чином, для зниження або елімінації активності поліпептиду можна застосовувати багато методів. Більше одного методу можна використати для зниження активності того самого поліпептиду. Крім того, для зниження або елімінації активності поліпептиду можна застосовувати комбінації методів. В одному з варіантів здійснення винаходу рівень експресії поліпептиду можна оцінювати безпосередньо, наприклад, визначаючи рівень полінуклеотиду або поліпептиду або відомого метаболіту в рослині (наприклад, визначаючи рівень N-ацетилгліфосату («NAG») у рослині, що містить ген GAT), або побічно, наприклад, шляхом оцінки в утримуючої його рослини 55 ознаки, обумовленого поліпептидом, наприклад, стійкості до гербіцидів. V. Способи боротьби з бур'янистою рослинністю У винаході запропоновані способи боротьби з бур'янистою рослинністю на оброблюваній посівній площі, попередження розвитку або появи стійкості до гербіцидів у бур'янистих рослин на оброблюваній посівній площі, одержання сільськогосподарської культури й підвищення схоронності сільськогосподарської культури. Поняття «боротьба» і його похідні, наприклад, «боротьба з бур'янистою рослинністю» відносяться до одному або декількох шляхам інгібування росту, проростання, розмноження й/або розростання; і/або знищення, видалення, винищування або зниження іншим способом зустрічальності й/або активності бур'янистої рослинності. Стійкі до гліфосату/інгібітору ALS рослини, пропоновані у винаході, мають модифіковану толерантність до гербіцидів і тому їх можна обробляти одним або декількома гербіцидам у нормах витрати, які можуть у значній мірі ушкоджувати ι контрольні рослини, і їх можна обробляти також комбінаціями гербіцидів у більше низьких концентраціях у порівнянні зі звичайно застосовуваними, які ще збережуть здатність до вибіркового знищення бур'янистих рослин. Крім того, толерантні до гліфосату/інгібітору ALS рослини, пропоновані у винаході, можна застосовувати в сполученні з технологією, заснованої на застосуванні сумішей гербіцидів, і тим самим підвищувати зручність, економічність і ефективної для виробника. Способи, пропоновані у винаході, полягають у тому, що засівають оброблювану посівну площу насіннями сільськогосподарської рослини, толерантної до гліфосату/інгібітору ALS, або засівають рослинами, пропонованими у винаході, і в конкретних варіантах здійснення винаходу, обробляють будь-яку сільськогосподарську рослину, частина сільськогосподарської рослини, бур'яниста рослина або оброблювана посівна площа гербіцидом, що представляє інтерес, в ефективній кількості. При створенні винаходу встановлено, що гербіцид можна вносити до або після посадки сільськогосподарської культури на оброблювану посівну площу. Така гербіцидна обробка може включати обробку гліфосатом, хімічною сполукою-інгібітором ALS або їхньою комбінацією. У конкретних варіантах здійснення винаходу суміш хімічної сполукиінгібітору ALS у сполученні із гліфосатом наносять на толерантну до гліфосату/інгібітору ALS рослина, при цьому ефективна концентрація щонайменше двох хімічних сполук-інгібіторів ALS повинна в значній мірі ушкоджувати відповідну контрольну рослину. В одному з не обмежуючих об'єм винаходу варіантів здійснення винаходу гербіцид являє собою щонайменше один із сульфоніламінокарбонілтріазолінонів; тріазопіримідинів; піримідініл(тіо)бензоатів; імідазолінонів; тріазінів; і/або похідних фосфінової кислоти. Відповідно до іншого не обмежуючий об'єм винаходу варіанту здійснення винаходу комбінація гербіцидів містить гліфосат, імазапир, хлоримурон-етил, хизалофоп і фомесафен, де ефективне 96578 56 кількість переноситься сільськогосподарською рослиною й контрольними бур'янами. Як буде представлено нижче в даному описі, можна застосовувати ці гербіциди в будь-якій ефективній кількості. У конкретних варіантах здійснення винаходу зазначена комбінація гербіцидів містить гліфосат в ефективній кількості, що становить від приблизно 1110 до приблизно 1130 м д.в./гектар; імазапир в ефективній кількості, що становить від приблизно 7,5 до приблизно 27,5 м д.в./гектар; хлоримуронетил в ефективній кількості, що становить від приблизно 7,5 до приблизно 27,5 м д.в./гектар; хізалофоп в ефективній кількості, що становить від приблизно 50 до приблизно 70 м д.в./гектар; і фомесафен в ефективній кількості, що становить від 240 до приблизно 260 м д.в./гектар. В інших варіантах здійснення винаходу застосовують щонайменше комбінацію двох гербіцидів, де до складу комбінації не входить гліфосат. В інших варіантах здійснення винаходу на рослину наносять щонайменше один інгібітор ALS і гліфосат. Більше докладні дані, що стосуються різних комбінацій гербіцидів, які можна застосовувати в способах, пропонованих у винаході, і їхні обговорення будуть представлені нижче. В одному з варіантів здійснення винаходу спосіб боротьби з бур'янами полягає в тому, що засівають площу насіннями толерантного до гліфосату/інгібітору ALS сільськогосподарської рослини або рослинами й обробляють сільськогосподарську рослину, частину сільськогосподарської рослини, насіння сільськогосподарської рослини або оброблювану посівну площу гербіцидом в ефективній кількості, де ефективна кількість являє собою I) кількість, що не переноситься першою контрольною сільськогосподарською рослиною при нанесенні на першу контрольну сільськогосподарську рослину, частину сільськогосподарської рослини, насіння або оброблювану посівну площу, де перша контрольна сільськогосподарська рослина експресує перший полінуклеотид, що обумовлює толерантність до гліфосату, і не експресує другий полінуклеотид, що кодує толерантний до інгібітору ALS поліпептид; II) кількість, що не переноситься другою контрольною сільськогосподарською рослиною при нанесенні на другу контрольну сільськогосподарську рослину, частину сільськогосподарської рослини, насіння або оброблювану посівну площу, де друга контрольна сільськогосподарська рослина експресує другий полінуклеотид і не експресує перший полінуклеотид; і III) кількість, що переноситься при нанесенні на толерантну до гліфосату/інгібітору ALS сільськогосподарська рослина, частина сільськогосподарської рослини, її насіння або оброблювану посівну площу. Гербіцид може являти собою комбінацію гербіцидів, що або включає, або не включає гліфосат. У конкретних варіантах здійснення винаходу комбінація гербіцидів містить хімічні сполуки-інгібітори ALS, що буде більш докладно описано нижче. В іншому варіанті здійснення винаходу спосіб боротьби з бур'янами полягає в тому, що засівають площа насіннями або рослинами толерантної 57 до гліфосату/інгібітору ALS сільськогосподарської культури, і наносять на сільськогосподарську рослину, частину цієї сільськогосподарської рослини, його насіння або оброблювана посівна площа гербіцид в ефективній кількості, де ефективна кількість являє собою кількість, що перевищує рекомендовану на етикетці по застосуванню норму витрати для зазначеної сільськогосподарської рослини, де ця ефективна кількість переноситься при нанесенні на толерантну до гліфосату/інгібітору ALS сільськогосподарська рослина, частину сільськогосподарської рослини, її насіння або оброблювану посівна площа. Застосовуваний гербіцид може являти собою комбінацію гербіцидів, що або включає, або не включає гліфосат. У конкретних варіантах здійснення винаходу комбінація гербіцидів містить хімічні сполуки-інгібітори ALS, що буде більш докладно описано нижче. Інші гербіциди і їхні комбінації, які можна застосовувати в різних способах, пропонованих у винаході, будуть більш докладно описані нижче. Відповідно до іншому не обмежуючий об'єм винаходу варіанту здійснення винаходу застосовуваний у кожному із представлених у даному описі способів гербіцид не являє собою гліфосат, хлоримурон-метил, римсульфурон, трибенуронметил або тифенсуфурон-метил. а. Типи гербіцидів Для обробки толерантного до гліфосату/ інгібітору ALS сільськогосподарської рослини, частини рослини або оброблюваної посівної площі, що містить зазначену культурну рослину можна застосовувати будь-який гербіцид. Класифікація гербіцидів (тобто поділ гербіцидів на класи й підкласи) добре відома в даній галузі й включає класифікацію HRAC (Діючий комітет зі стійкості до гербіцидів) і WSSA (Американське суспільство по вивченню бур'янів) (див. також Retzinger і MallorySmith, Weed Technology 11, 1997, cc. 384-393). Скорочена версія HRAC-класифікації (з урахуванням відповідної WSSA-групи) наведена нижче в таблиці 1. Гербіциди можна класифікувати по їхньому механізмі й/або місцю дії й можна класифікувати також за часом їхнього внесення (наприклад, передвсходове або післявсходове), за методом їхнього застосування (наприклад, листова обробка або ґрунтове внесення) або по тому, як вони проникають у рослину або вражають рослину. Наприклад, трифенсульфурон-метил і трибенурон-метил наносять на листя сільськогосподарської рослини (наприклад, кукурудзи) і, як правило, вони метаболізуються в ньому, а римсульфурон і хлоримуронетил, як правило, проникають як через корінь, так і через листя рослини. Поняття «механізм дії», як правило, відноситься до метаболическому або фізіологічного процесу в рослині, у результаті якого гербіцид інгібує або іншим способом ушкоджує рослина, а поняття «місце дії», як правило, відноситься до фізіологічної локалізації або біохімічного сайту в рослині, на який робить вплив гербіцид або з яким він безпосередньо взаємодіє. Гербіциди можна класифікувати різними шляхами, у тому числі по їхньому механізмі дії й/або місцю дії (див., наприклад, таблицю 1). 96578 58 Часто зумовлюючий толерантність до гербіциду ген, що надає толерантність експресуючій його рослині до конкретного гербіциду або іншій хімічній речовині, може обумовлювати толерантність також до інших гербіцидів або хімічних речовин цього ж класу або підкласу, наприклад, класу або підкласу, представленого в таблиці 1. Таким чином, у деяких варіантах здійснення винаходу трансгенна рослина, пропонована у винаході, має толерантність більш ніж до одного гербіциду або хімічної речовини цього ж класу або підкласу, такому, наприклад, як інгібітор РРО, похідне сульфосечовини або синтетичний ауксин. Як правило, рослини, пропоновані в даному винаході, можуть переносити обробку різними типами гербіцидів (тобто гербіцидами з різним механізмом дії й/або різним місцем дії), а також більше високим концентраціями гербіцидів у порівнянні з відомими раніше для рослин, забезпечуючи тим самим поліпшені стратегії контролю за чисельністю бур'янистої рослинності в порівнянні з рекомендованими для зниження зустрічальності й поширення толерантності бур'янів до гербіцидів. Для ефективної боротьби з бур'янами можна застосовувати конкретні комбінації гербіцидів. Таким чином, у винаході запропоновані трансгенні культурні рослини, які можна відбирати для застосування в рослинництві з урахуванням поширення толерантності до гербіцидів видів бур'янистих рослин на площі, на якій повинно вирощувати трансгенну сільськогосподарську рослину. У даній галузі відомі методи оцінки толерантності до гербіцидів різних видів бур'янів. Методи контролю за чисельністю бур'янистої рослинності також відомі в даній галузі, такі, наприклад, як ротація з використанням сільськогосподарської культури, толерантної до гербіциду, до якого місцеві види бур'янів не мають толерантність. Деякі із зазначених органів і громадських організацій публікують дані про випадки виникнення й особливостях толерантності до гербіцидів бур'янів, включаючи Діючий комітет зі стійкості до гербіцидів (HRAC), Американське суспільство по вивченню бур'янів, різні державні агентства (див., наприклад, дані про толерантність до гербіцидів різних широколиственних бур'янів у довіднику Illinois Agricultural Pest Management Handbook, 2004), і фахівець у даній галузі може використати цю інформацію для рішення питання про те, яку сільськогосподарську культуру й комбінацію гербіцидів можна використати в конкретному регіоні. Зазначені організації публікують також ради й посібника з попередження розвитку й/або виникнення й контролю поширення толерантних до гербіцидів бур'янів (див., наприклад, Owen і Hartzler (2004), 2005 Herbicide Manual for Agricultural Professionals, публікація WC 92 Revised (Iowa State University Extension, Iowa State University of Science and Technology, Ames, Iowa); Weed Control for Corn, Soybeans, and Sorghum, розділ 2 «2004 Illinois Agricultural Pest Management Handbook» (University of Illinois Extension, University of Illinois at Urbana-Champaign, Illinois)); Weed Control Guide for Field Crops, MSU Extension Bulletin E434 (Michigan State University, East Lansing, Michigan)). 59 96578 Таблиця 1 Скорочена версія HRAC-класифікації гербіцидів І. Інгібітори ALS (група 2 по WSSA-класифікаціі) А. Сульфонілсечовини 1. Азимсульфурон 2. Хлоримурон-етил 3. Метсульфурон-метил 4. Никосульфурон 5. Римсульфурон 6. Сульфометурон-метил 7. Трифенсульфурон-метил 8. Трибенурон-метил 9. Амідосульфурон 10. Бенсульфурон-метил 11. Хлорсульфурон 12. Циносульфурон 13. Циклосульфамурон 14. Етаметсульфурон-метил 15. Етоксисульфурон 16. Флазасульфурон 17. Флупирсульфурон-метил 18. Форамсульфурон 19. Імазосульфурон 20. Іодосульфурон-метил 21. Мезосульфурон-метил 22. Оксасульфурон 23. Прімісульфурон-метил 24. Просульфурон 25. Піразосульфурон-етил 26. Сульфосульфурон 27. Тріасульфурон 28. Трифоксифлоксисульфурон 29. Трифлусульфурон-метил 30. Тритосульфурон 31. Галосульфурон-метил 32. Флуцетосульфурон Б. Сульфоніламінокарбонілтріазолінони 1. Флукарбазон 2. Прокарбазон В. Тріазолопіримідини 1. Клорансулам-метил 2. Флуметсулам 3. Диклосулам 4. Флорасулам 5. Метосулам 6. Пенокссулам 7. Пірокссулам Г. Піримідінілокси(тіо)бензоати 1. Біспірибак 2. Пірифталід 3. Пірибензоксим 4. Піритіобак 5. Пирімінобак-метил Д. Імідазолінони 1. Імазапир 2. Імазетапир 3. Імазахин 4. Імазапик 5. Імазаметабенз-метил 60 6. Імазамокс II. Інші гербіциди - діючі речовини/додаткові механізми дії А. Інгібітори ацетил-СоА-карбоксилази (АССаза) (група 1 по WSSA-класифікації) 1. Арилоксифеноксипропіонати («FOP») а. Хизалофоп-П-етил б. Дихлофоп-метил в. Клодинафоп-пропаргіл г. Феноксапроп-П-етил д. Флуазифоп-П-бутил е. Пропахизафоп ж. Галоксифоп-П-метил з. Цигалофоп-бутил и. Хизалофоп-П-етил 2. Циклогександіони («DIM») а. Аллоксидим б. Бутоксидим в. Клетодим г. Циклоксидим д. Сетоксидим е. Тепралоксидим ж. Тралкоксидим Б. Інгібітори системи фотосинтезу II, група 31 по HRAC-класифікації/група 5 по WSSA-класифікації 1. Тріазіни а. Аметрин б. Атразін в. Цианазін г. Десметрин д. Диметаметрин е. Прометон ж. Прометрин з. Пропазин и. Сімазін к. Симетрин л. Тербуметон м. Тербутилазін н. Тербутрин о. Триетазін 2. Тріазінони а. Гексазінон б. Метрибузин в. Метамітрон 3. Тріазолінон а. Амікарбазон 4. Урацили а. Бромацил б. Ленацил в. Тербацил 5. Піридазінони а. Піразон 6. Фенілкарбамати а. Десмедифам б. Фенметифам В. Інгібітори системи фотосинтезу - II, група 32 по HRAC-класифікації/група 7 по WSSA-класифікації 1. Сечовини а. Флуометурон б. Лінурон в. Хлоробромурон
ДивитисяДодаткова інформація
Назва патенту англійськоюMonocotyledonous transgenic plant containing polynucleotide encoding polypeptide providing tolerance to glyphosate
Автори англійськоюMCCUTCHEN BILLY FRED, CASTLE LINDA, CHICOINE TIMOTHY K, CHO HYEON-JE, CLAUS JON S, GREEN JERRY M, GUIDA ANTHONY D, HAZEL CHRISTINE B, HEKERT METYU J., HEGSTAD JEFFREY M, HUTCHISON JAMES M, LIU DONGLONG, LU ALBERT L, MEHRE WAYNE J, MOY YORK, OLSON PAUL D, PEEPLES KENNETH A, SAUNDERS DAVID W, VOGT MARK D, WILKINSON JACK Q, WONG JAMES F H
Назва патенту російськоюОднодольное трансгенное растение, содержащее полинуклеотид, который кодирует полипептид, обуславливающий толерантность к глифосату
Автори російськоюМаккатчен Билли Фред, Касл Линда А., Чикойн Тимоти К., Чо Хьон-же, Клаус Джон С., Грин Джерри М., Гайда Энтони Д., Джр., Хейзел Кристин Б., Хекерт Метью Дж., Хегстад Джеффри М., Хатчисон Джеймс М., Лю Дунлун, Лу Алберт Л., Мере Вейн Дж., Мой Йорк, Олсон Пол Д., Пиплз Кеннет А., Сондерз Девид В., Вогт Марк Д., Вилкинсон Джек К., Вон Джеймс Ф. Х.
МПК / Мітки
МПК: C12N 15/82, A01H 5/00
Мітки: кодує, поліпептид, рослина, обумовлює, гліфосату, трансгенна, однодольна, містить, толерантність, полінуклеотид
Код посилання
<a href="https://ua.patents.su/188-96578-odnodolna-transgenna-roslina-shho-mistit-polinukleotid-shho-kodueh-polipeptid-shho-obumovlyueh-tolerantnist-do-glifosatu.html" target="_blank" rel="follow" title="База патентів України">Однодольна трансгенна рослина, що містить полінуклеотид, що кодує поліпептид, що обумовлює толерантність до гліфосату</a>
Поліпептид вірусу гепатиту с корейського типу (khcv), кднк khcv, полінуклеотид, що кодує поліпептид khcv, рекомбінантний експресуючий вектор, клітина, трансформована вектором, поліпептид, який містить епітоп, с

Номер патенту: 43310
Опубліковано: 17.12.2001
Автори: Чо Чунг Мюнг, Лі Йонг Беом, Со Хонг Сеоб, Лім Кук Джін, Кім Чан Хюнг, Парк Янг Ву, Янг Джає Янг, Чой Деог Янг, Кім Сунг Таєк
МПК: C07K 14/00, C07K 16/00, C12P 21/08, C07K 16/08, C12Q 1/70, C12N 15/09, C07K 14/02, C07K 19/00, C07K 14/10, C07K 14/18, G01N 33/576, G01N 33/53, C12N 1/21, C12N 1/19, C12P 21/02, C12N 15/51
Мітки: типу, вектор, гепатиту, епітоп, рекомбінантний, кднк, вектором, khcv, корейського, вірусу, експресуючий, кодує, поліпептид, містить, полінуклеотид, клітина, трансформована
Формула / Реферат:
1. Полипептид вируса гепатита С корейского типа (KHCV), обладающий следующей аминокислотной последовательностью: фрагмент, обладающий специфичностью KHCV, или его производные.2. Полипептид KHCV по п. 1, отличающийся тем, что аминокислоты 828-933 полипептида, охарактеризованного в п. 1, заменены любым полипептидом, выбранным из группы, включающей полипептиды, обладающие следующими...
Ізольований або рекомбінантний полінуклеотид, що кодує поліпептид, який має активність гліфосат-n-ацетилтрансферази (gat)

Номер патенту: 86918
Опубліковано: 10.06.2009
Автори: Міншулл Джеремі, Дак Ніколас Б., Чен Янг Хонг, Гайвер Лорайн Дж., Сайхл Ден, Кембл Роджер, Айві Крістіна, Касл Лінда А., МакКатчен Біллі Ф., Паттен Філліп А.
МПК: C12N 9/10, C12N 5/14, A01H 5/00, C12N 15/54, C12N 15/82
Мітки: має, поліпептид, гліфосат-n-ацетилтрансферази, полінуклеотид, gat, ізольований, активність, кодує, рекомбінантний
Формула / Реферат:
1. Ізольований або рекомбінантний полінуклеотид, що кодує поліпептид, який має активність глiфосат-N-ацетилтрансферази, що включає нуклеотидну послідовність, що кодує послідовність амінокислот, яка має щонайменше 60 % iдентичностi послiдовностi по всій довжині послідовності, вибраної з групи, що включає SEQ. ID. No.: 300, SEQ. ID. No.: 445 i SEQ. ID. No.: 457.2. Ізольований або рекомбінантний полінуклеотид, що кодує поліпептид, який...
Виділений полінуклеотид, що кодує злитий протеїн з пестицидною активністю, та трансгенна клітина-хазяїн

Номер патенту: 84666
Опубліковано: 25.11.2008
Автори: Шваб Джордж Е., Кардіно Гай А., Стамп Лайза, Фінстад Лі Стейсі, Майкаелс Трейсі Елліс, Шнепф Х.Ернст, Херман Род А., Кнут Марк, Поллард Майкл Р., Діель Паула, Нарва Кеннет Е., Доджіллоу Джоанна
МПК: C12N 15/09, C12N 15/32, C07K 14/235, C12N 5/10, C12N 1/15, C12R 1/07, A01N 63/02, C12N 1/21, C12N 15/82
Мітки: полінуклеотид, пестицидною, активністю, виділений, клітина-хазяїн, протеїн, трансгенна, злитий, кодує
Формула / Реферат:
1. Виділений полінуклеотид, що кодує злитий протеїн, що включає першу амінокислотну послідовність і другу амінокислотну послідовність, де згадана перша амінокислотна послідовність є амінокислотною послідовністю поліпептиду, що має молекулярну вагу 45 kDa, і згадана друга амінокислотна послідовність є амінокислотною послідовністю поліпептиду, що має молекулярну вагу 15 kDa, де згадані поліпептиди є токсичними по відношенню до кореневого черва...
Полінуклеотид, що кодує поліпептид потенціалзалежного ворітного іонного каналу людини (canion), поліпептид, антитіло, спосіб ідентифікації кандидатного модулятора canion-поліпептиду, спосіб лікування біполярног

Номер патенту: 79927
Опубліковано: 10.08.2007
Автори: Чумаков Ілья, Коен Даніель, Сімон Анн-Марі, Абдеррахім Хаді
МПК: A61P 25/18, A61P 25/00, C07K 16/28, C12N 15/12, C12N 15/63, C07K 14/705, A61K 39/395, G01N 33/50
Мітки: ворітного, полінуклеотид, каналу, кандидатного, антитіло, модулятора, лікування, потенціалзалежного, поліпептид, кодує, canion-поліпептиду, біполярног, ідентифікації, canion, спосіб, іонного, людини
Формула / Реферат:
1. Виділений, очищений або рекомбінантний полінуклеотид, який кодує поліпептид потенціалзалежного ворітного іонного каналу людини (CanIon),a) що складається з будь-якої з нуклеотидних послідовностей, представлених як SEQ ID NO: 1-3; абоb) що складається з:і) нуклеотидної послідовності, представленої як SEQ ID NO: 4,іі) послідовності, комплементарної будь-якій з послідовностей (а); абоііі) полінуклеотиду,...
Трансгенна рослинна клітина, що включає поліпептид з активністю гліфосат-n-ацетилтрансферази

Номер патенту: 94688
Опубліковано: 25.05.2011
Автори: Чен Янг Хонг, Дак Ніколас Б., Гайвер Лорайн Дж., Касл Лінда А., МакКатчен Біллі Ф., Сайхл Ден, Міншулл Джеремі, Айві Крістіна, Кембл Роджер, Паттен Філліп А.
МПК: C12N 5/14, C12N 9/10, A01H 5/00, C12N 15/54, C12N 15/82
Мітки: активністю, включає, гліфосат-n-ацетилтрансферази, рослинна, поліпептид, трансгенна, клітина
Формула / Реферат:
1. Трансгенна рослинна клітина, що включає гетерологiчний поліпептид з активністю глiфосат-N-ацетилтрансферази, де вказана рослинна клітина включає метаболічний продукт гліфосату, який являє собою N-ацетилгліфосат після обробки гліфосатом.2. Трансгенна рослинна клітина за п. 1, де рослинна клітина виявляє підвищену стійкість до глiфосату порівняно з клітиною дикої рослини того ж виду, лінією або культурним сортом.3. Трансгенна...
Наступний патент: Кришка-контейнер для автономного міксингу
Випадковий патент: Спосіб недетермінованого криптографічного перетворення блоків даних