Подія 5307 кукурудзи
Номер патенту: 110320
Опубліковано: 25.12.2015
Автори: Преарі Анна Андервуд, Мехджі Моез Раджабалі, Де Фрамонд Аннік Жанна, Нью Стівен





















































































































































































































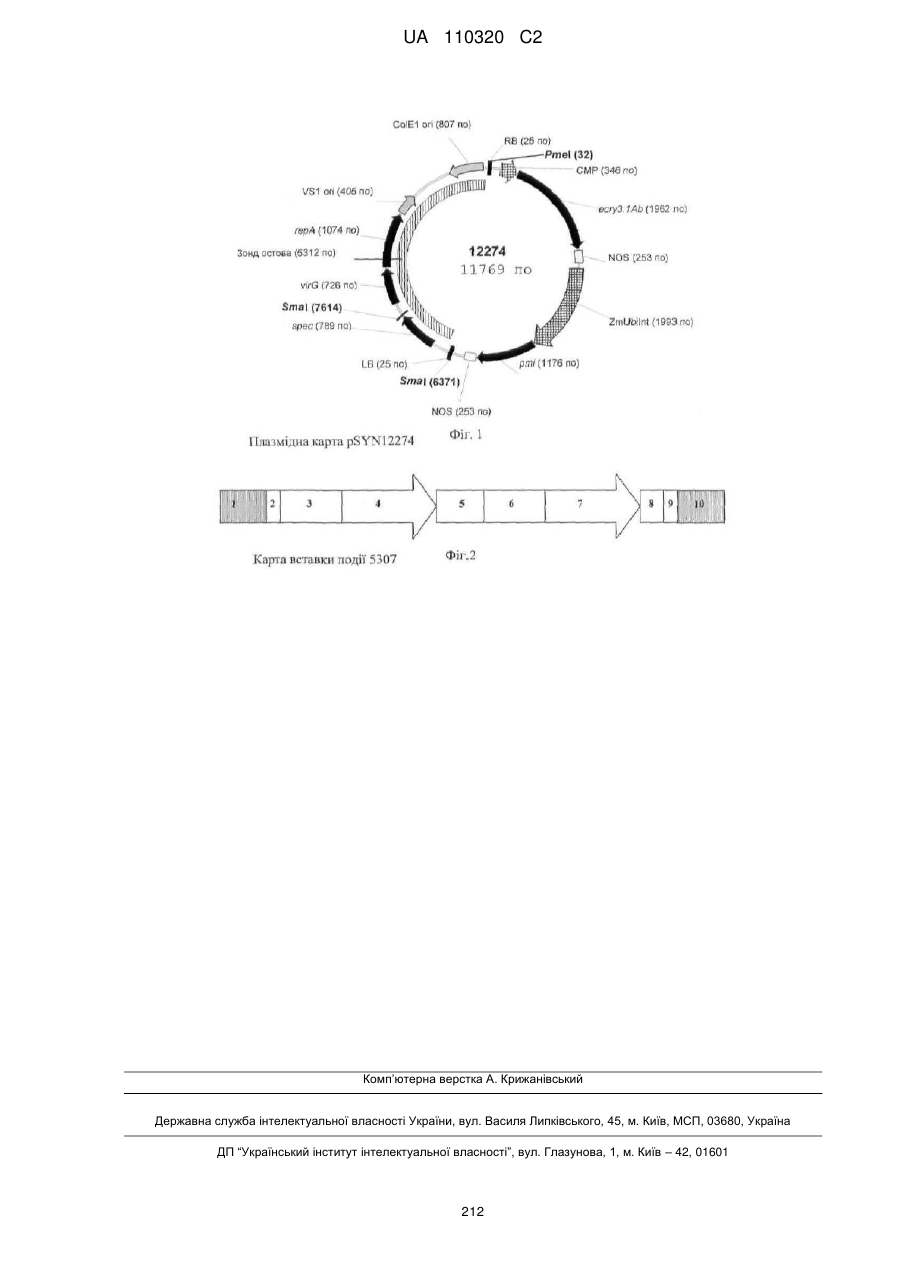
Формула / Реферат
1. Клітина, отримана з трансгенної рослини кукурудзи, що включає подію 5307 для захисту від комах, де зазначена клітина включає SEQ ID NO: 1 або SEQ ID NO: 2.
2. Трансгенна рослина кукурудзи, її клітини та тканини, що включають SEQ ID NO: 1 або SEQ ID NO: 2.
3. Молекула нуклеїнової кислоти, що включає SEQ ID NO: 1, SEQ ID NO: 3 або SEQ ID NO: 4.
4. Молекула нуклеїнової кислоти за п. 3, де молекула нуклеїнової кислоти включена в кукурудзяне насіння, депоноване в АТСС під інвентарним номером PTA-9561.
5. Амплікон, що включає молекулу нуклеїнової кислоти за п. 3 або п. 4.
6. Пара полінуклеотидних праймерів, що включає перший полінуклеотидний праймер і другий полінуклеотидний праймер, які функціонують разом у присутності ДНК-матриці події 5307 кукурудзи в зразку з продукуванням амплікона, діагностичного для події 5307 кукурудзи, де перша праймерна послідовність являє собою або є комплементарною до геному рослини кукурудзи, що фланкує точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, і друга полінуклеотидна праймерна послідовність являє собою або є комплементарною до гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, де
а) перший полінуклеотидний праймер включає щонайменше, 10 суміжних нуклеотидів з положення 1-1348, як викладено в SEQ ID NO: 5, або їх комплементарні ланцюги; або щонайменше 10 суміжних нуклеотидів з положення 1-1093, як викладено в SEQ ID NO:6, або їх комплементарні ланцюги; і
б) другий полінуклеотидний праймер включає щонайменше 10 суміжних нуклеотидів з положення 1-6206, як викладено в SEQ ID NO: 7, або їх комплементарні ланцюги,
де вказана подія 5307 кукурудзи характеризується наявністю нуклеотидних послідовностей, як вказано в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO:4, або їх комплементарних ланцюгів,
і де вказана подія 5307 кукурудзи включена в кукурудзяне насіння, депоноване в АТСС під інвентарним номером PTA-9561.
7. Спосіб виявлення наявності молекули нуклеїнової кислоти, яка специфічна події 5307, у зразку, що включає нуклеїнові кислоти кукурудзи, при цьому спосіб включає:
a) виділення молекули нуклеїнової кислоти з кукурудзи;
b) об'єднання молекули нуклеїнової кислоти з парою праймерних послідовностей нуклеїнової кислоти SEQ ID NO: 8 - SEQ ID NO: 15 або SEQ ID NO: 69 - SEQ ID NO: 72 з SEQ ID NO: 16 - SEQ ID NO: 68, або їх комплементарними ланцюгами;
c) проведення реакції ампліфікації нуклеїнової кислоти, яка дає в результаті амплікон; і
d) виявлення амплікона.
8. Спосіб виявлення наявності молекули нуклеїнової кислоти, яка специфічна події 5307, у зразку, що включає нуклеїнові кислоти кукурудзи, при цьому спосіб включає:
a) виділення молекули нуклеїнової кислоти з кукурудзи;
b) об'єднання молекули нуклеїнової кислоти з парою праймерних послідовностей нуклеїнової кислоти SEQ ID NO: 82 і SEQ ID NO: 83, SEQ ID NO: 85 і SEQ ID NO: 86, SEQ ID NO: 87 і SEQ ID NO: 89, поряд з їхніми послідовностями-зондами SEQ ID NO: 84, SEQ ID NO: 87 і SEQ ID NO: 90, відповідно;
c) проведення реакції ампліфікації нуклеїнової кислоти, яка дає в результаті амплікон, що включає зонд; і
d) виявлення зонда.
9. Молекула ДНК, що включає амплікон, отриманий способом за п. 7.
10. Спосіб підтвердження відсутності молекули нуклеїнової кислоти, яка специфічна події 5307, у зразку, що включає нуклеїнові кислоти кукурудзи, при цьому спосіб включає:
a) виділення геномної ДНК із кукурудзи;
b) об'єднання молекули нуклеїнової кислоти з парою праймерних послідовностей нуклеїнової кислоти SEQ ID NO: 8 - SEQ ID NO: 15 або SEQ ID NO: 69 - SEQ ID NO: 72 з SEQ ID NO: 16 - SEQ ID NO: 68 або їх комплементарними ланцюгами, поряд з парою праймерів нуклеїнової кислоти нативного гена кукурудзи як позитивного контролю;
c) проведення реакції ампліфікації нуклеїнової кислоти, яка не дає в результаті амплікон, специфічний події 5307, і дає в результаті амплікон, специфічний позитивному контролю нативного гена кукурудзи; та
d) виявлення амплікона, специфічного позитивному контролю нативного гена кукурудзи.
11. Біологічний зразок, отриманий з рослини кукурудзи, тканини, насіння або клітини події 5307, де зразок включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна SEQ ID NO: 1 або SEQ ID NO:2.
12. Біологічний зразок за п. 11, де послідовність є такою, що виявляється в екстракті за допомогою способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти.
13. Біологічний зразок за п. 11, де зразок вибраний із групи, що складається з кукурудзяного борошна, кукурудзяної мучки, кукурудзяної патоки, кукурудзяної олії, кукурудзяного крохмалю та круп, зроблених повністю або частково з вмістом кукурудзяних побічних продуктів.
14. Екстракт, отриманий з біологічного зразка рослини кукурудзи, насіння, тканини або клітини події 5307, при цьому зазначений екстракт включає нуклеотидну послідовність, яка комплементарна SEQ ID NO: 1 або SEQ ID NO: 2.
15. Екстракт за п. 14, де послідовність є такою, що виявляється в екстракті за допомогою способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти.
16. Екстракт за п. 14, де зразок вибраний із групи, що складається з кукурудзяного борошна, кукурудзяного борошна, кукурудзяної мучки, кукурудзяної патоки, кукурудзяної олії, кукурудзяного крохмалю та круп, зроблених повністю або частково з вмістом кукурудзяних побічних продуктів.
17. Спосіб добору за допомогою маркера за ознакою стійкості до комах у кукурудзи, що включає:
a) виділення молекули нуклеїнової кислоти з кукурудзи;
b) об'єднання молекули нуклеїнової кислоти з парою праймерних послідовностей і зондами нуклеїнової кислоти, SEQ ID NO: 91 - SEQ ID NO: 102 або їх комплементарними ланцюгами;
c) проведення реакції ампліфікації нуклеїнової кислоти, яка дає в результаті амплікон;
d) виявлення амплікона; та
e) добір рослини для цілей селекції стійкої до комах кукурудзи.
18. Спосіб виявлення наявності молекули нуклеїнової кислоти, що відповідає події 5307 кукурудзи, у зразка, при цьому спосіб включає:
a) взаємодію зразка зі зондом, який гібридизується в умовах високої жорсткості з геномною ДНК із події 5307 кукурудзи та не гібридизується в умовах високої жорсткості із ДНК контрольної рослини кукурудзи;
b) вплив на зразок і зонд гібридизаційними умовами високої жорсткості; та
c) виявлення гібридизації зонда із ДНК.
19. Набір для виявлення наявності нуклеїнових кислот події 5307 кукурудзи в біологічному зразку, при цьому набір включає щонайменше одну молекулу ДНК, що включає достатню довжину суміжних нуклеотидів, яка безпосередньо являє собою або комплементарна SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 або SEQ ID NO: 7, яка функціонує як ДНК-праймер або зонд, специфічний для події 5307 кукурудзи.
20. Маїсовий хромосомний сайт-мішень, розташований на хромосомі 5 між молекулярним маркером umc1475, де umc1475 включає SEQ ID NO: 104, і молекулярним маркером uaz190, де uaz190 включає SEQ ID NO: 107, і де сайт-мішень включає гетерологічну нуклеїнову кислоту.
21. Маїсовий хромосомний сайт-мішень, розташований на хромосомі 5 між нуклеотидом 1 і нуклеотидом 161752 SEQ ID NO: 103, де сайт-мішень включає гетерологічну нуклеїнову кислоту.
22. Маїсовий хромосомний сайт-мішень за п. 20 або п. 21, де зазначений сайт-мішень розташований між нуклеотидами 75908 і 75946 SEQ ID NO: 103.
23. Маїсовий хромосомний сайт-мішень за п. 20 або п. 21, де зазначений сайт-мішень фланкований на 5' нуклеотидами 55980-75908 SEQ ID NO: 103 і фланкований на 3' нуклеотидами 75946-95946 SEQ ID NO: 103.
24. Спосіб створення трансгенної рослини маїсу, що включає вставлення гетерологічної нуклеїнової кислоти в положення на хромосомі 5, розташоване між молекулярним маркером umc1475, де umc1475 включає SEQ ID NO: 104, і молекулярним маркером uaz190, де uaz190 включає SEQ ID NO: 107.
25. Спосіб за п. 24, де зазначена гетерологічна нуклеїнова кислота вставлена на хромосомі 5 між нуклеотидами 75908 і 75946 SEQ ID NO: 103.
26. Спосіб за п. 24, де зазначена вставлена гетерологічна нуклеїнова кислота фланкована на 5' нуклеотидами 5454-25454 SEQ ID NO: 103 і фланкована на 3' нуклеотидами 25513-45513 SEQ ID NO: 103.
Текст
Реферат: Винахід належить до трансгенної події кукурудзи 5307. Також винахід належить до послідовностей ДНК рекомбінантних конструктів, вставлених у геном кукурудзи, і геномних послідовностей, що фланкують сайт вставки, що в результаті дало подію 5307. Крім того, даний винахід додатково належить до аналізів виявлення наявності послідовностей ДНК події 5307, рослин кукурудзи та насіння кукурудзи, що включають такий генотип. UA 110320 C2 (12) UA 110320 C2 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 Галузь винаходу [0001] Даний винахід, загалом, стосується галузі молекулярної біології рослин, трансформації рослин і селекції рослин. Більш конкретно, даний винахід стосується стійких до комах трансгенних рослин кукурудзи, що включають новий трансгенний генотип, і способів виявлення присутності ДНК рослини кукурудзи в зразку та їх композицій. Передумови [0002] Шкідники рослин є головним чинником у втраті важливих світових сільськогосподарських культур. Приблизно 8 мільярдів доларів втрачається щороку тільки в США через зараження шкідниками-нессавцями, включаючи комах. Види кукурудзяних жуків розглядаються як найбільш згубні шкідники кукурудзи. Важливі види кукурудзяних жуківшкідників включають Diabrotica virgifera virgifera, західний кукурудзяний жук; D. longicornis barberi, північний кукурудзяний жук, D. undecimpunctata howardi, південний кукурудзяний жук, і D. virgifera zeae, мексиканський кукурудзяний жук. [0003] З кукурудзяним жуком борються, головним чином, шляхом інтенсивного застосування хімічних пестицидів. Таким чином, можна досягти гарного контролю кукурудзяного жука, але ці хімікати також можуть іноді впливати на корисні організми. Іншою проблемою, що виникає в результаті широкого застосування хімічних пестицидів, є виникнення стійких різновидів комах. Це частково послаблялося за допомогою різних практик керування стійкістю, але існує зростаюча необхідність в альтернативних стратегіях боротьби зі шкідниками. Одна така альтернатива включає експресію чужорідних генів, що кодують інсектицидні білки в трансгенних рослинах. Цей підхід забезпечив ефективні засоби захисту проти обраних комах-шкідників, і були запущені у виробництво трансгенні рослини, що експресують інсектицидні токсини, дозволяючи фермерам зменшити застосування хімічних інсектицидів. [0004] На експресію чужорідних генів у рослин може впливати їх хромосомне положення, можливо, обумовлене структурою хроматину та близькістю транскрипційних регуляторних елементів до сайту інтеграції [дивися, наприклад, Weising et al., 1988, "Foreign Genes in Plants," Ann. Rev. Genet. 22:421-477]. Отже, звичайним є одержання сотень різних подій і скринінг цих подій заради однієї події, яка має бажані рівні та патерни експресії трансгена для комерційних цілей. Подія, яка має бажані рівні або патерни експресії трансгена, є придатною для інтрогресії трансгена в інші генетичні фони за допомогою статевого ауткросингу із застосуванням загальноприйнятих способів селекції. Потомство таких гібридів зберігає властивості експресії трансгена вихідного трансформанта. Ця стратегія застосовується для забезпечення надійної генної експресії в ряді різновидів, добре адаптованих до місцевих умов вирощування. [0005] Було б вигідним мати можливість виявляти присутність конкретної події для того, щоб визначати, чи містить потомство статевого схрещування трансген інтересу. До того ж, спосіб виявлення конкретної події був би корисним для відповідності правилам, що вимагають схвалення перед випуском на ринок і маркування харчових продуктів, наприклад, отриманих з рекомбінантних сільськогосподарських рослин. Можна виявляти присутність трансгена за допомогою будь-якого добре відомого способу виявлення нуклеїнової кислоти, включаючи, але без обмежень, термальну ампліфікацію (полімеразна ланцюгова реакція (ПЛР)) і застосування полінуклеотидних праймерів або гібридизації ДНК із застосуванням зондів нуклеїнової кислоти. Зазвичай, заради простоти та однаковості реагентів і методик для застосування при виявленні конкретного ДНК-конструкта, який був застосований для трансформування різних сортів рослин, ці способи виявлення в основному зосереджуються на часто використовуваних генетичних елементах, наприклад, промоторах, термінаторах і маркерних генах, тому що для багатьох ДНК-конструктів, ділянка кодуючої послідовності є взаємозамінною. У результаті, такі способи можуть не бути придатними для розрізнення між конструктами, які відрізняються тільки стосовно кодуючої послідовності. До того ж такі способи можуть не бути придатними для розрізнення між різними подіями, особливо тими, які отримані із застосуванням того ж ДНКконструкта, якщо відома послідовність хромосомної ДНК, суміжна із вставленою гетерологічною ДНК ("фланкуюча ДНК"). [0006] Даний винахід включає стійку до комах подію трансгенної кукурудзи, яка має вбудований у її геном ген FR8a, розкритий у міжнародній публікації WO 08/121633, опублікованій 9 жовтня 2008 р., яка включена в даний опис посиланням, що кодує FR8a інсектицидний токсин, придатний у боротьбі з комахами-шкідниками Diabrotica spp. Подія трансгенної кукурудзи також має вбудований у її геном ген PMI, що кодує фермент фосфоманоза-ізомеразу (PMI), розкритий у патенті США № 5767378, який включено у даний опис посиланням, придатний у якості селективного маркера, який дозволяє рослині використовувати манозу в якості джерела вуглецю. Даний винахід, крім того, включає нові виділені послідовності нуклеїнових кислот, які є унікальними для події трансгенної кукурудзи, 1 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 придатні для ідентифікації події трансгенної кукурудзи та задля виявлення нуклеїнових кислот з події трансгенної кукурудзи в біологічному зразку, а також набори, що включають реагенти, необхідні для застосування при виявленні цих нуклеїнових кислот у біологічному зразку. Короткий опис [0007] Даний винахід представляє подію трансгенної кукурудзи, позначену 5307, яка включає новий трансгенний генотип, що включає ген FR8a і ген PMI, який надає стійкість до комах і здатність використовувати манозу в якості джерела вуглецю, відповідно, події 5307 кукурудзи та її потомству. Даний винахід також забезпечує трансгенні рослини кукурудзи, що включають генотип даного винаходу, насіння від трансгенних рослин кукурудзи, що включає генотип даного винаходу, та способи одержання трансгенної рослини кукурудзи, що включає генотип даного винаходу, за допомогою схрещування інцухтованої рослини кукурудзи, що включає генотип даного винаходу, з власне собою або іншою лінією кукурудзи відмінного генотипу. Трансгенні рослини кукурудзи даного винаходу можуть мати, по суті, усі з морфологічних і фізіологічних властивостей відповідної ізогенної нетрансгенної рослини кукурудзи додатково до наданих рослині кукурудзи новим генотипом даного винаходу. Даний винахід також забезпечує композиції та способи виявлення присутності нуклеїнових кислот з події 5307 на основі послідовності ДНК рекомбінантних касет експресії, вставлених у геном кукурудзи, що дає в результаті подію 5307, і геномних послідовностей, що фланкують сайт вставки. Подія 5307 може бути, крім того, охарактеризована за допомогою аналізу рівнів експресії білків FR8a і PMI, а також випробуванням ефективності проти кукурудзяного жука. [0008] Відповідно до одного аспекту даний винахід забезпечує переважно виділену молекулу нуклеїнової кислоти, що включає, щонайменше, 10 суміжних нуклеотидів гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, і, щонайменше, 10 суміжних нуклеотидів ДНК геному рослини кукурудзи, що фланкують точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи. Переважно, виділена молекула нуклеїнової кислоти згідно із цим аспектом може містити, щонайменше, 20 або, щонайменше, 50 суміжних нуклеотидів гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, і, щонайменше, 20 або, щонайменше, 50 суміжних нуклеотидів ДНК геному рослини кукурудзи, що фланкує точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи. [0009] Згідно з іншим аспектом даний винахід забезпечує переважно виділену молекулу нуклеїнової кислоти, що включає, щонайменше, одну з'єднуючу послідовність події 5307, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2 і їх комплементарних ланцюгів. З'єднуюча послідовність охоплює з'єднання між гетерологічною ДНК, що включає касети експресії, вставлені в геном кукурудзи, і ДНК із геному кукурудзи, що фланкує сайт вставки, і є діагностичною для події 5307. [0010] Згідно з іншим аспектом даний винахід забезпечує переважно виділену нуклеїнову кислоту, що зв'язує гетерологічну молекулу ДНК із геномом рослини кукурудзи в події 5307 кукурудзи, що включає, послідовність від приблизно 11 до приблизно 20 суміжних нуклеотидів, вибраних із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2 і їх комплементарних ланцюгів. [0011] Згідно з іншим аспектом даний винахід забезпечує переважно виділену молекулу нуклеїнової кислоти, що включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 і їх комплементарних ланцюгів. [0012] Згідно з іншим аспектом даного винаходу забезпечено амплікон, що включає молекулу нуклеїнової кислоти даного винаходу. [0013] Згідно ще одному аспекту даного винаходу забезпечені праймери фланкуючих послідовностей для виявлення події 5307. Такі праймери фланкуючих послідовностей містять переважно виділену послідовність нуклеїнової кислоти, що включає, щонайменше, 10-15 суміжних нуклеотидів з нуклеотидів 1-1348, як викладено в SEQ ID NO: 5 (довільно позначену в даному описі як 5' фланкуюча послідовність), або їх комплементарні ланцюги, також розкриті як SEQ ID NO: 111. В одному варіанті здійснення даного аспекту праймери фланкуючих послідовностей вибирають із групи, яка складається з SEQ ID NO: 9 – SEQ ID NO: 14 і їх комплементарних ланцюгів. [0014] В іншому аспекті даного винаходу праймери фланкуючих послідовностей включають переважно виділену послідовність нуклеїнової кислоти, що включає, щонайменше, 10-15 суміжних нуклеотидів з нуклеотидів 1-1093, як викладено в SEQ ID NO: 6 (довільно позначену в даному описі як 3' фланкуюча послідовність), або їх комплементарні ланцюги. В одному варіанті здійснення даного аспекту праймери фланкуючих послідовностей вибирають із групи, яка складається з SEQ ID NO: 69 – SEQ ID NO: 72 і їх комплементарних ланцюгів. 2 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 [0015] Згідно з іншим аспектом даного винаходу забезпечено пари праймерів, які придатні, наприклад, для ампліфікації нуклеїнової кислоти. Такі пари праймерів включають перший праймер, що включає нуклеотидну послідовність, щонайменше, з 10-15 суміжних нуклеотидів завдовжки, яка безпосередньо являє собою або комплементарна одній з вищеописаних геномних фланкуючих послідовностей (SEQ ID NO: 5 або SEQ ID NO: 6), і другий праймер, що включає нуклеотидну послідовність, щонайменше, з 10-15 суміжних нуклеотидів гетерологічної ДНК, вставленої в геном події 5307. Другий праймер переважно включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна послідовності вставки, суміжною з геномною фланкуючою послідовністю ДНК рослини, як викладено в SEQ ID NO: 7. В одному варіанті здійснення даного аспекту праймери послідовності вставки вибирають із групи, що складається з SEQ ID NO: 15 – SEQ ID NO: 68 і їх комплементарних ланцюгів. [0016] Згідно з іншим аспектом даного винаходу забезпечено способи виявлення присутності ДНК, що відповідає події 5307 у біологічному зразку. Такі способи включають: (a) взаємодію зразка, що включає ДНК, з парою праймерів, що, при застосуванні в реакції ампліфікації нуклеїнової кислоти з геномною ДНК із події 5307 кукурудзи, продукує амплікон, який є діагностичним для події 5307 кукурудзи; (b) виконання реакції ампліфікації нуклеїнової кислоти, внаслідок чого продукується амплікон; і (c) виявлення амплікона. В одному варіанті здійснення даного аспекту амплікон включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 і їх комплементарних ланцюгів. [0017] Згідно з іншим аспектом даний винахід забезпечує способи виявлення присутності ДНК, що відповідає події 5307, у біологічному зразку. Такі способи включають: (a) взаємодію зразка, що включає ДНК, із зондом, який гібридизується в умовах високої жорсткості з геномною ДНК із події 5307 кукурудзи та не гібридизується в умовах високої жорсткості з ДНК контрольної рослини кукурудзи; (b) вплив на зразок і зонд гібридизаційними умовами високої жорсткості; та (c) виявлення гібридизації зонда з ДНК. Виявлена гібридизована послідовність ДНК включає, щонайменше, одну полінуклеотидну послідовність, що включає SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5, SEQ ID NO: 6 і їх комплементарні ланцюги. [0018] Згідно з іншим аспектом даного винаходу забезпечено набір для виявлення нуклеїнових кислот події 5307 у біологічному зразку. Набір включає, щонайменше, одну послідовність ДНК, що включає достатню довжину полінуклеотидів, яка безпосередньо являє собою або комплементарна SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5 або SEQ ID NO: 6, де послідовності ДНК придатні в якості праймерів або зондів, які гібридизуються з виділеною ДНК з події 5307, і які при ампліфікації або гібридизації з послідовністю нуклеїнової кислоти в зразку з наступним виявленням амплікона або гібридизацією з мішеневою послідовністю є діагностичними для присутності послідовностей нуклеїнової кислоти з події 5307 у зразку. Набір, крім того, включає інші матеріали, необхідні, щоб уможливити способи гібридизації або ампліфікації нуклеїнової кислоти. [0019] В іншому аспекті даний винахід забезпечує спосіб виявлення білка події 5307 кукурудзи в біологічному зразку, що включає: (a) екстрагування білка із зразка тканини події 5307 кукурудзи; (b) проведення аналізу виділеного білка із застосуванням імунологічного способу, що включає антитіло, специфічне для інсектицидного або селективного маркерного білка, що продукується подією 5307; і (c) виявлення зв'язування зазначеного антитіла з інсектицидним або селективним маркерним білком. [0020] В іншому аспекті даний винахід забезпечує біологічний зразок, отриманий з рослини кукурудзи, тканини або насіння події 5307, де зразок включає нуклеїнову кислоту, що включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна послідовності, вибраній з групи, що складається з SEQ ID NO: 1 і SEQ ID NO: 2, і де послідовність виявляється в зразку із застосуванням способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти. В одному варіанті здійснення даного аспекту зразок вибирають із групи, що складається з кукурудзяного борошна, кукурудзяної мучки, кукурудзяної патоки, кукурудзяної олії, кукурудзяного крохмалю та круп, виготовлених повністю або частково з вмістом кукурудзяних побічних продуктів. [0021] В іншому аспекті даний винахід забезпечує екстракт, отриманий з рослини кукурудзи, тканини або насіння події 5307, що включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна нуклеотидній послідовності, вибраній з групи, що складається з SEQ ID NO: 1 і SEQ ID NO: 2. В одному варіанті здійснення даного аспекту послідовність виявляється в екстракті із застосуванням способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти. В іншому варіанті здійснення даного аспекту зразок вибирають із групи, що складається з кукурудзяного борошна, кукурудзяної мучки, кукурудзяної патоки, 3 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 кукурудзяної олії, кукурудзяного крохмалю та круп, виготовлених повністю або частково з вмістом кукурудзяних побічних продуктів. [0022] Згідно з іншим аспектом даного винаходу забезпечено рослини кукурудзи та насіння, що включають молекули нуклеїнової кислоти даного винаходу. В одному варіанті здійснення даного винаходу кукурудзяна насінина події 5307 була депонована в Американську колекцію типових культур (ATCC) відповідно до Будапештської угоди 15 жовтня 2008 р. Приклад зазначеного насіння є депонованим під інвентарним № ATCC: PTA-9561. [0023] Згідно з іншим аспектом даний винахід забезпечує спосіб одержання рослини кукурудзи, стійкої до, щонайменше, враження кукурудзяним жуком, що включає: (a) статеве схрещування першої батьківської рослини кукурудзи із другою батьківською рослиною кукурудзи, де перша або друга батьківська рослина кукурудзи включає ДНК події 5307 кукурудзи, таким чином, одержуючи безліч рослин-нащадків першого покоління; (b) добір рослини-нащадка першого покоління, яка є стійкою, щонайменше, до враження кукурудзяним жуком; (c) самозапилення рослини-нащадка першого покоління, таким чином, одержуючи безліч рослин-нащадків другого покоління; (d) добір з рослин-нащадків другого покоління рослини, яка є, щонайменше, стійкою до враження кукурудзяним жуком; де рослини-нащадки другого покоління включають нуклеотидну послідовність, вибрану з групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 і SEQ ID NO: 4. [0024] Згідно з ще одним аспектом даний винахід забезпечує спосіб одержання насіння кукурудзи, що включає схрещування першої батьківської рослини кукурудзи з другою батьківською рослиною кукурудзи та збирання отриманого в результаті насіння кукурудзи першого покоління, де перша або друга батьківська рослина кукурудзи є інцухтованою рослиною кукурудзи даного винаходу. [0025] Згідно з іншим аспектом даний винахід забезпечує спосіб одержання насінин гібридної кукурудзи, що включає етапи: (a) висівання насінин першої інхухт-лінії кукурудзи згідно з даним винаходом і насінин другої інцухт-лінії кукурудзи, що має відмінний генотип; (b) вирощування рослини кукурудзи, отриманої в результаті зазначеного висівання, до появи квіток; (c) видалення недоспілих маточок квіток рослини кукурудзи однієї з інцухт-ліній кукурудзи; (d) забезпечення здійснення запилення іншої інцухт-лінії, та (e) збирання гібридного насіння, утвореного таким шляхом. [0026] Згідно з іншим аспектом даного винаходу даний винахід забезпечує спосіб добору рослин і насінин кукурудзи, що включають молекули нуклеїнової кислоти події 5307 на хромосомі 5. В одному варіанті здійснення даного винаходу поліморфні маркери застосовуються, щоб добирати або відслідковувати послідовності, специфічні для події 5307 кукурудзи. Даний винахід забезпечує спосіб добору послідовностей, специфічних для події 5307 кукурудзи, що включає етапи: (a) виявлення послідовності поліморфного маркера; (b) планування аналізу з метою виявлення маркера; (c) проведення аналізу послідовностей нуклеїнової кислоти кукурудзи з багатьох ліній кукурудзи та (d) добір ліній кукурудзи на основі послідовностей з нуклеотидами, специфічними для події 5307 кукурудзи. [0027] Згідно з іншим аспектом даного винаходу даний винахід забезпечує сайт на хромосомі 5 для націленої інтеграції гетерологічної нуклеїнової кислоти. Даний винахід забезпечує спосіб добору послідовностей, специфічних для події 5307 кукурудзи, для націленої інтеграції, що включає етапи: (a) розробки гомологічних послідовностей на основі сайту вставки або послідовності вектора; (b) застосування цих гомологічних послідовностей у мішеневому локусі; (c) застосування нуклеаз «цинкові пальці» для створення розриву в мішеневому локусі, та (d) вставлення гетерологічної донорної молекули в нуклеотиди, специфічні для події 5307 кукурудзи або послідовності вектора pSYN12274. Приклад цієї техніки продемонстрований в Shukla et al. (Nature vol. 459, 21 May 2009). [0028] Попередні й інші аспекти даного винаходу стануть більш очевидними з наступного докладного опису. Опис послідовностей у переліку послідовностей SEQ ID NO: 1 є 5’ з'єднанням геном-вставка. SEQ ID NO: 2 є 3’ з'єднанням геном-вставка. SEQ ID NO: 3 є 5’ послідовністю геном + вставка. SEQ ID NO: 4 є 3’ послідовністю вставка + геном. SEQ ID NO: 5 є 5’ послідовністю геном + вставка. SEQ ID NO: 6 є 3’ фланкуючою послідовністю генома кукурудзи. SEQ ID NO: 7 є вставкою повної довжини події 5307. SEQ ID No: 8-14 є 5’ праймерами фланкуючої послідовності, придатними в даному винаході. SEQ ID No: 15-68 є праймерами трансгенної вставки 5307, придатними в даному винаході. 4 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 SEQ ID No: 69-72 є 3’ праймерами фланкуючої послідовності, придатними в даному винаході. SEQ ID No: 73-75 є FR8a TAQMAN праймерами та зондом. SEQ ID No: 76-78 є PMI TAQMAN праймерами та зондом. SEQ ID No: 79-81 є ZmAdh TAQMAN праймерами та зондом. SEQ ID No: 82-90 є праймерами та зондами, специфічними для події 5307, придатними в даному винаході. SEQ ID No: 91-102 є геномними праймерами та зондами кукурудзи, придатними в даному винаході. SEQ ID NO: 103 є послідовністю AC202955 хромосоми 5, де N є будь-якою основою “А”, “Т”, “G” або “C”. SEQ ID NO: 104 є маркерною ділянкою umc1475. SEQ ID No: 105-106 є праймерами umc1475. SEQ ID NO: 107 є маркерною ділянкою uaz190. SEQ ID NO: 108-109 є праймерами uaz190. SEQ ID NO: 110 є зворотною комплементарною SEQ ID NO: 103, послідовністю AC202955 хромосоми 5, де N є будь-якою основою “А”, “Т”, “G” або “C”. SEQ ID NO: 111 є 5’ фланкуючою послідовністю генома кукурудзи. Короткий опис графічних матеріалів Фігура 1 ілюструє вектор експресії рослини, позначений pSYN12274. На карті плазміди позначені сайти рестрикції SmaI і PmeI, що застосовуються для Саузерн-аналізу. Фігура 2 являє собою графічну карту, що ілюструє організацію елементів, включаючи гетерологічні послідовності нуклеїнової кислоти, вставлені в геном кукурудзи для утворення події 5307, і яка визначає відносні положення, в яких вставлені послідовності нуклеїнової кислоти зв'язані з геномними послідовностями ДНК кукурудзи, що фланкують кінці вставлених гетерологічних послідовностей ДНК. 1=5’фланкуючий геном рослини (SEQ ID NO: 5); 2= права гранична ділянка; 3= промотор CMP; 4= ген FR8a; 5= термінатор NOS; 6= промотор ZmUbINT; 7= ген PMI; 8= термінатор NOS; 9=ліва гранична ділянка (відрізки 2-9 містяться в SEQ ID NO: 7); і 10=3’ фланкуючий геном рослини (SEQ ID NO: 6). Визначення [0029] Представлено наступні визначення та способи, щоб краще визначити даний винахід і допомогти фахівцям в даній галузі в здійсненні даного винаходу. Якщо не зазначене інше, вирази, використовувані в даному описі, слід розуміти відповідно до загальноприйнятого використання фахівцями у відповідній галузі. Визначення загальних виразів у молекулярній біології можна також знайти в Rieger et al., Glossary of Genetics: Classical and Molecular, 5th edition, Springer-Verlag: New York, 1994. [0030] Як використовується в даному документі, вираз "ампліфікований" означає конструювання множинних копій молекули нуклеїнової кислоти або множинних копій, комплементарних до молекули нуклеїнової кислоти, із застосуванням, щонайменше, однієї з молекул нуклеїнової кислоти у якості шаблону. Системи ампліфікації включають систему полімеразної ланцюгової реакції (ПЛР), систему лігазної ланцюгової реакції (LCR), ампліфікацію на основі послідовності нуклеїнової кислоти (NASBA, Cangene, Міссиссауга, Онтаріо), Q-бета репліказні системи, систему транскрипційної ампліфікації (TAS) і ампліфікацію із заміщенням ланцюгів (SDA). Дивись, наприклад, Diagnostic Molecular Microbiology: Principles and Applications, D. H. Persing et al., Ed., American Society for Microbiology, Washington, D.C. (1993). Продукт ампліфікації називається амплікон. [0031] “Біологічний зразок” є рослиною, рослинним матеріалом або продуктами, що включають рослинний матеріал. Вираз “рослина” призначений для включення тканин рослини кукурудзи (Zea mays) на будь-якій стадії зрілості, а також клітин, тканин, органів, узятих з або отриманих з будь-якої такої рослини, включаючи, без обмеження, будь-які насінини, листи, стебла, квітки, корені, окремі клітини, гамети, клітинні культури, культури тканин або протопласти. “Рослинний матеріал”, як використовується в даному документі, стосується матеріалу, який отриманий або вилучений з рослини. Продукти, що включають рослинний матеріал, відносяться до продовольства, корму або інших продуктів, які виробляють із застосуванням рослинного матеріалу або можуть бути забруднені рослинним матеріалом. Зрозуміло, що в контексті даного винаходу такий біологічний зразок тестується на присутність нуклеїнових кислот, специфічних для події 5307 кукурудзи, що має на увазі присутність нуклеїнових кислот у зразках. Таким чином, способи, що згадуються в даному описі для ідентифікації події 5307 кукурудзи в біологічних зразках, стосуються ідентифікації в біологічних зразках нуклеїнових кислот, які отримані з рослини події 5307 кукурудзи та є діагностичними для 5 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 події 5307. [0032] "Кодуюча послідовність" є послідовністю нуклеїнової кислоти, яка транскрибується в РНК, таку як мРНК, рРНК, тРНК, мяРНК, змістовна РНК або антизмістовна РНК. Переважно, РНК потім транслюється в організмі з утворенням білка. [0033] “Набір для виявлення”, як використовується в даному документі, стосується набору, що застосовується для виявлення присутності або відсутності ДНК із рослин події 5307 кукурудзи в зразку, що включає зонди та праймери нуклеїнової кислоти даного винаходу, які специфічно гібридизуються в умовах високої жорсткості з мішеневою послідовністю ДНК, і інші матеріали, необхідні, щоб уможливити способи гібридизації або ампліфікації нуклеїнової кислоти. [0034] Як використовується в даному документі, вираз трансгенна "подія" стосується рекомбінантної рослини, отриманої за допомогою трансформації та регенерації однієї рослинної клітини гетерологічною ДНК, наприклад, касетою експресії, яка включає ген інтересу. Вираз “подія” стосується вихідного трансформанта та/або потомства трансформанта, які включають гетерологічну ДНК. Вираз “подія” також стосується потомства, отриманого шляхом статевого ауткросингу між трансформантом і іншою лінією кукурудзи. Навіть після повторного зворотного схрещування з рекурентнтим батьком вставлена ДНК і фланкуюча ДНК із трансформованого батька присутня у потомстві схрещування в тому ж хромосомному положенні. У нормі, трансформація рослинної тканини утворює безліч подій, кожна з яких представляє вставку ДНК-конструкта в різні положення в геномі рослинної клітини. На основі експресії трансгена або інших бажаних характеристик добирають конкретну подію. Таким чином, “подія 5307”, “5307 подія” або “5307”, як використовується в даному документі, означає вихідний 5307 трансформант і/або потомство 5307 трансформанта, включаючи будь-яку рослину, отриману від них. [0035] "Касета експресії", як використовується в даному документі, означає молекулу нуклеїнової кислоти, здатну керувати експресією конкретної нуклеотидної послідовності у відповідній клітині-хазяїні, що включає промотор, функціонально зв'язаний з нуклеотидною послідовністю інтересу, яка є функціонально зв'язаною із сигналами термінації. Вона також типово включає послідовності, необхідні для правильної трансляції нуклеотидної послідовності. Касета експресії може також включати послідовності, що не є необхідними для прямої експресії нуклеотидної послідовності інтересу, але чия присутність обумовлена придатними сайтами рестрикції для видалення касети експресії з вектора експресії. Касета експресії, що включає нуклеотидну послідовність інтересу, може бути химерною, що означає, що, щонайменше, один з її компонентів є гетерологічним відносно, щонайменше, одного з її інших компонентів. Касета експресії може також бути такою, що зустрічається в природі, але була отримана в рекомбінантній формі, придатній для гетерологічної експресії. Проте, типово касета експресії є гетерологічною відносно хазяїна, тобто конкретна послідовність нуклеїнової кислоти касети експресії не зустрічається в природі в клітині-хазяїні та має бути введена в клітину-хазяїна або предкові клітини-хазяїна за допомогою процесу трансформації, відомого в рівні техніки. Експресія нуклеотидної послідовності в касеті експресії може перебувати під контролем конститутивного промотору або індуцибельного промотору, який ініціює транскрипцію тільки тоді, коли клітина-хазяїн зазнає деякого конкретного зовнішнього стимулу. У випадку багатоклітинного організму, такого як рослина, промотор може також бути специфічним для конкретної тканини, або органу, або стадії розвитку. Касета експресії, або її фрагмент, може також бути названа “вставленою послідовністю” або “послідовністю вставки”, коли трансформована в рослину. [0036] "Ген" є визначеною ділянкою, яка розташована в геномі і яка, крім вищезгаданої кодуючої послідовності нуклеїнової кислоти, включає інші, у першу чергу регуляторні, послідовності нуклеїнової кислоти, відповідальні за контроль експресії, тобто транскрипції та трансляції кодуючої частини. Ген також може містити інші 5' і 3' нетрансльовувані послідовності та термінуючі послідовності. Додатковими елементами, які можуть бути присутніми, є, наприклад, інтрони. [0037] "Ген інтересу" стосується будь-якого гена, який при переносі в рослину надає рослині бажану характеристику, таку як стійкість до антибіотика, стійкість до вірусу, стійкість до комах, стійкість до хвороби або стійкість до інших шкідників, гербіцидна витривалість, поліпшена поживна цінність, поліпшені показники у виробничому процесі або змінена репродуктивна здатність. "Ген інтересу" може також бути таким, який переносять в рослини для одержання комерційно цінних ферментів або метаболітів у рослині. [0038] “Генотип”, як використовується в даному документі, є генетичним матеріалом, успадкованим від батьківських рослин кукурудзи, не все з якого обов'язково експресується в 6 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 рослинах-нащадках кукурудзи. 5307 генотип стосується гетерологічного генетичного матеріалу, трансформованого в геном рослини, а також генетичного матеріалу, фланкуючого вставлену послідовність. [0039] "Гетерологічна" послідовність нуклеїнової кислоти є послідовністю нуклеїнової кислоти, не зв'язаною в природі із клітиною-хазяїном, у яку вона введена, включаючи множинні копії, що не зустрічаються в природі, послідовності нуклеїнової кислоти, що зустрічається в природі. [0040] "Гомологічна" послідовність нуклеїнової кислоти є послідовністю нуклеїнової кислоти, зв'язаною в природі із клітиною-хазяїном, у яку вона введена. [0041] Вираз "виділений", коли використовується по відношенню до нуклеїнової кислоти, стосується послідовності нуклеїнової кислоти, яка ідентифікована та відділена від, щонайменше, однієї нуклеїнової кислоти-забруднювача, з якою вона зазвичай зв'язана в її природному джерелі. Виділена нуклеїнова кислота присутня у формі або положенні, яке відрізняється від того, у якому вона перебуває в природі. На відміну від цього, невиділені нуклеїнові кислоти, такі як ДНК і РНК, перебувають у стані, у якому вони існують у природі. Виділена нуклеїнова кислота може бути в трансгенній рослині й усе ще вважатися “виділеною”. [0042] “Функціонально-зв'язаний” стосується поєднання послідовностей нуклеїнової кислоти на одному фрагменті нуклеїнової кислоти так, що функціонування однієї впливає на функціонування іншої. Наприклад, промотор є функціонально-зв'язаним з кодуючою послідовністю або функціональною РНК, коли він здатний впливати на експресію цієї кодуючої послідовності або функціональної РНК (тобто кодуюча послідовність або функціональна РНК перебуває під транскрипційним контролем промотору). Кодуючі послідовності у змістовній або антизмістовній орієнтації можуть бути функціонально-зв'язаними з регуляторними послідовностями. [0043] “Праймери”, як використовується в даному документі, є виділеними нуклеїновими кислотами, які відпалюються з комплементарним мішеневим ланцюгом ДНК за допомогою гібридизації нуклеїнової кислоти з утворенням гібриду між праймером і мішеневим ланцюгом ДНК, потім подовжується уздовж мішеневого ланцюга ДНК полімеразою, такою як ДНКполімераза. Пари праймерів або набори можна використовувати для ампліфікації молекули нуклеїнової кислоти, наприклад, за допомогою полімеразної ланцюгової реакції (ПЛР) або інших загальноприйнятих способів ампліфікації нуклеїнової кислоти. [0044] “Зонд” є виділеною нуклеїнової кислотою, до якої прикріплена загальноприйнята мітка, що виявляється, або репортерна молекула, така як радіоактивний ізотоп, ліганд, хемілюмінісцентний засіб або фермент. Такий зонд є комплементарним до ланцюга мішеневої нуклеїнової кислоти, у випадку даного винаходу до ланцюга геномної ДНК із події кукурудзи, M5307. Геномна ДНК події 5307 може бути з рослини кукурудзи або зі зразка, який включає ДНК із події. Зонди згідно з даним винаходом включають не тільки дезоксирибонуклеїнові або рибонуклеїнові, а також поліаміди й інші зондові матеріали, які специфічно зв'язуються з мішеневою послідовністю ДНК і можуть використовуватися для виявлення присутності цієї мішеневої послідовності ДНК. [0045] Праймери та зонди звичайно складають 10-15 нуклеотидів або більше завдовшки. Праймери та зонди також можуть складати, щонайменше, 20 нуклеотидів або більше завдовшки, або, щонайменше, 25 нуклеотидів або більше, або, щонайменше, 30 нуклеотидів або більше завдовшки. Такі праймери та зонди специфічно гібридизуються з мішеневою послідовністю при умовах гібридизації високої жорсткості. Праймери та зонди за даним винаходом можуть мати повну комплементарність послідовності з мішеневою послідовністю, хоча зонди, які відрізняються від мішеневої послідовності і зберігають здатність гібридизуватися з мішеневими послідовностями, можуть бути розроблені загальноприйнятими способами. [0046] "Жорсткі умови" або "жорсткі умови гібридизації" включають посилання на умови, при яких зонд буде гібридизуватися з його мішеневою послідовністю у виявно більшій мірі, ніж з іншими послідовностями. Жорсткі умови є залежними від мішеневої послідовності та будуть відрізнятися залежно від структури полінуклеотиду. Шляхом контролювання жорсткості умов гібридизації та/або відмивання можуть бути ідентифіковані мішеневі послідовності, які є на 100% комплементарними зонду (гомологічне зондування). Альтернативно, жорсткі умови можуть бути відрегульовані, щоб дозволити деякі розбіжності в послідовностях так, що виявляються нижчі ступені подібності (гетерологічне зондування). Довші послідовності гібридизуються, зокрема, при вищих температурах. Великий посібник з гібридизації нуклеїнових кислот знаходиться в Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular BiologyHybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier: New York; і Current Protocols in Molecular 7 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 Biology, Chapter 2, Ausubel et al., Eds., Greene Publishing and Wiley-Interscience: New York (1995), а також Sambrook et al. (2001) Molecular Cloning: A Laboratory Manual (5th Ed. Cols Spring Harbor Laboratory, Cold Spring Harbor, NY). [0047] Специфічність зазвичай залежить від постгібридизаційних відмивань, при цьому вирішальними факторами є іонна сила та температура розчину остаточного відмивання. Взагалі, вибрані умови гібридизації високої жорсткості та відмивання на приблизно 5ºC нижче термічної точки плавлення (Tm) для специфічної послідовності при визначеній іонній силі та pH. Tm є температурою (при визначеній іонній силі та pH), при якій 50% мішеневої послідовності гібридизується з ідеально збіжним зондом. Зазвичай, при умовах високої жорсткості зонд буде гібридизуватися з його мішеневою підпослідовністю, а не з іншими послідовностями. [0048] Прикладом умов гібридизації високої жорсткості для гібридизації комплементарних нуклеїнових кислот, які мають більш ніж 100 комплементарних залишків, на фільтрі в Саузернабо Нозерн-блотинзі є 50% формамід з 1 мг гепарину при 42ºC, при цьому гібридизацією проводять протягом ночі. Прикладом умов відмивання дуже високої жорсткості є 0,15 M NaCl при 72ºC протягом приблизно 15 хвилин. Прикладом умов відмивання високої жорсткості є відмивання 0,2 x SSC при 65ºC протягом 15 хвилин (дивись, Sambrook, нижче, для опису SSC буфера). [0049] Зразкові умови гібридизації для даного винаходу включають гібридизацію в 7% SDS, 0,25 M NaPO4 pH 7,2 при 67 °C протягом ночі з наступними двома відмиваннями в 5% SDS, 0,20 M NaPO4 pH 7,2 при 65 °C протягом 30 хвилин на кожне відмивання та двома відмиваннями в 1% SDS, 0,20 M NaPO4 pH 7,2 при 65 °C протягом 30 хвилин на кожне відмивання. Зразковим середовищем жорсткого відмивання для дуплексу, наприклад, з більш ніж 100 нуклеотидів, є 1 x SSC при 45ºC протягом 15 хвилин. Зразковим відмиванням низької жорсткості для дуплексу, наприклад, з більш ніж 100 нуклеотидів, є 4-6 x SSC при 40ºC протягом 15 хвилин. [0050] Для зондів з від приблизно 10 до 50 нуклеотидів умови високої жорсткості типово включають концентрації солі менше приблизно 1,0 M іону Na, типово концентрацію іону Na (або інших солей) від приблизно 0,01 до 1,0 M при pH від 7,0 до 8,3, і температура становить типово, щонайменше, приблизно 30ºC. Умови високої жорсткості можуть також бути досягнуті додаванням дестабілізуючих засобів, таких як формамід. Взагалі, співвідношення сигналу до шуму становить 2x (або вище), ніж те, яке спостерігалося для незв'язаного зонда в конкретному гібридизаційному аналізі, що показує виявлення специфічної гібридизації. Нуклеїнові кислоти, які не гібридизуються одна з одною в умовах високої жорсткості є все ще, по суті, ідентичними, якщо білки, які вони кодують, є, по суті, ідентичними. Це трапляється, наприклад, коли копія нуклеїнової кислоти створена із застосуванням максимальної виродженості кодону, дозволеної генетичним кодом. [0051] Далі наведені зразкові набори умов гібридизації/відмивання, які можуть бути застосовані для гібридизації нуклеотидних послідовностей, які є, по суті, ідентичними до еталонних нуклеотидних послідовностей даного винаходу: еталонна нуклеотидна послідовність переважно гібридизується з еталонною нуклеотидною послідовністю в 7% додецилсульфаті натрію (SDS), 0,5 M NaPO4, 1 мM EDTA при 50 °C з відмиванням в 2 x SSC, 0,1% SDS при 50 °C, більш бажано в 7% додецилсульфаті натрію (SDS), 0,5 M NaPO4, 1 мM EDTA при 50 °C з відмиванням в 1 x SSC, 0,1% SDS при 50 °C, ще більш бажано в 7% додецилсульфаті натрію (SDS), 0,5 M NaPO4, 1 мM EDTA при 50 °C з відмиванням в 0,5 x SSC, 0,1% SDS при 50 °C, переважно в 7% додецилсульфаті натрію (SDS), 0,5 M NaPO4, 1 мM EDTA при 50 °C з відмиванням в 0,1 x SSC, 0,1% SDS при 50 °C, більш переважно в 7% додецилсульфаті натрію (SDS), 0,5 M NaPO4, 1 мM EDTA при 50 °C з відмиванням в 0,1 x SSC, 0,1% SDS при 65 °C. Послідовності даного винаходу можуть бути виявлені із застосуванням усіх вищеописаних умов. Для цілей визначення даного винаходу застосовують умови високої жорсткості. [0052] "Трансформація" є процесом уведення гетерологічної нуклеїнової кислоти в клітинуабо організм-хазяїн. Зокрема, "трансформація" означає стабільну інтеграцію молекули ДНК у геном організму інтересу. [0053] "Трансформований/трансгенний/рекомбінантний" стосуються організму-хазяїна, такого як бактерія або рослина, у який була введена гетерологічна молекула нуклеїнової кислоти. Молекула нуклеїнової кислоти може бути стабільно інтегрована в геном хазяїна, або молекула нуклеїнової кислоти може також бути присутньою у якості позахромосомної молекули. Така позахромосомна молекула може бути такою, що самореплікується. Трансформовані клітини, тканини або рослини, як розуміють, охоплюють не тільки кінцевий продукт процесу трансформації, але також їх трансгенне потомство. "Нетрансформований", "нетрансгенний" або "нерекомбінантний" хазяїн стосується організму дикого типу, наприклад, бактерії або рослини, який не містить гетерологічної молекули нуклеїнової кислоти. Як використовується в даному 8 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 документі, "трансгенний" стосується рослини, рослинної клітини або маси структурованих або неструктурованих рослинних клітин, що мають інтегровану, за допомогою добре відомих технік генетичної маніпуляції та вставлення генів, нуклеїнову кислоту, що представляє ген інтересу, у геном рослини, і типово в хромосому клітинного ядра, мітохондрію або іншу органелу, що містить хромосоми, у локус, що відрізняється від того, або в кількості копій більшій, ніж та, що в нормі присутня в нативній рослині або рослинній клітині. Трансгенні рослини, отримані в результаті маніпуляції та вставлення таких послідовностей нуклеїнової кислоти, у порівнянні з мутаціями, що зустрічаються в природі, утворюють рослину, що не зустрічається в природі, або рослину з генотипом, що не зустрічається в природі. Техніки трансформації рослин і рослинних клітин є добре відомими в даній галузі та можуть включати, наприклад, електропорацію, мікроін'єкцію, опосередковану Agrobacterium трансформацію та балістичну трансформацію. [0054] У даному описі використана номенклатура для основ ДНК і амінокислот, як викладено в 37 C.F.R. § 1.822. Докладний опис [0055] Даний винахід стосується генетично покращеної лінії кукурудзи, яка продукує білок захисту від комах, FR8a, і фермент фосфоманоза-ізомераза (PMI), який дозволяє рослині використовувати манозу в якості джерела вуглецю. Даний винахід зокрема спрямований на трансгенну подію кукурудзи, позначену як подія 5307, що включає новий генотип, а також на композиції та способи виявлення нуклеїнових кислот із цієї події в біологічному зразку. Даний винахід, крім того, спрямований на рослини кукурудзи, що включають генотип події 5307, на трансгенне насіння від рослин кукурудзи та на способи одержання рослини кукурудзи, що включає генотип події 5307, шляхом схрещування інцухтованої кукурудзи, що включає генотип події 5307 із власне собою або з іншою лінією кукурудзи. Рослини кукурудзи, що включають генотип події 5307 даного винаходу, придатні в боротьбі з твердокрилими комахамишкідниками, включаючи Diabrotica virgifera virgifera, західний кукурудзяний жук, D. virgifera zeae, мексиканський кукурудзяний жук, і D. longicornis barberi, північний кукурудзяний жук. Рослини кукурудзи, що включають генотип події 5307 даного винаходу, також здатні використовувати манозу в якості джерела вуглецю. [0056] В одному варіанті здійснення даний винахід охоплює трансгенне насіння кукурудзи події 5307 рослини кукурудзи. При цьому приклад зазначеного насіння депоновано в АТСС під інвентарним № PTA-9561. Трансгенне насіння події 5307 включає SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5 або SEQ ID NO: 6 і їх комплементарні ланцюги. Ці послідовності визначають точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи. В іншому варіанті здійснення даний винахід охоплює переважно виділену молекулу нуклеїнової кислоти, що включає SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 або SEQ ID NO: 4. В іншому варіанті здійснення даний винахід охоплює переважно виділену молекулу нуклеїнової кислоти, де молекула нуклеїнової кислоти включена в насіння кукурудзи, депоноване в ATCC під інвентарним № PTA-9561. [0057] В одному варіанті здійснення даний винахід охоплює молекулу нуклеїнової кислоти, переважно виділену, що включає, щонайменше, 10 або більше (наприклад, 15, 20, 25 або 50) суміжних нуклеотидів гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, і, щонайменше, 10 або більше (наприклад, 15, 20, 25 або 50) суміжних нуклеотидів ДНК генома рослини кукурудзи, що фланкує точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи. Також включені нуклеотидні послідовності, які включають 10 або більше нуклеотидів суміжної послідовності вставки з події 5307 і, щонайменше, один нуклеотид фланкуючої ДНК із події 5307, суміжний з послідовністю вставки. Такі нуклеотидні послідовності є діагностичними для події 5307. Ампліфікація нуклеїнової кислоти геномної ДНК із події 5307 продукує амплікон, що ключає такі діагностичні нуклеотидні послідовності. [0058] В іншому варіанті здійснення даний винахід охоплює молекулу нуклеїнової кислоти, переважно виділену, що включає нуклеотидну послідовність, яка включає, щонайменше, одну з'єднуючу послідовність події 5307, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 і їх комплементарних ланцюгів, де з'єднуюча послідовність охоплює з'єднання між гетерологічною касетою експресії, вставленою в геном кукурудзи, і ДНК із генома кукурудзи, що фланкує сайт вставки, і є діагностичною для події. [0059] В іншому варіанті здійснення даний винахід охоплює переважно виділену нуклеїнову кислоту, що зв'язує гетерологічну молекулу ДНК із геномом рослини кукурудзи в події 5307 кукурудзи, що включає послідовність з від приблизно 11 до приблизно 20 суміжних нуклеотидів, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2 і їх комплементарних ланцюгів. [0060] В іншому варіанті здійснення даний винахід охоплює молекулу нуклеїнової кислоти, 9 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 переважно виділену, що включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 і їх комплементарних ланцюгів. [0061] В одному варіанті здійснення даного винаходу забезпечено амплікон, що включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 і їх комплементарних ланцюгів. [0062] В іншому варіанті здійснення даний винахід охоплює праймери фланкуючої послідовності для виявлення події 5307. Такі праймери фланкуючої послідовності включають виділену послідовність нуклеїнової кислоти, що включає, щонайменше, 10-15 суміжних нуклеотидів з нуклеотидів 1-1348 з SEQ ID NO: 5 (довільно позначена в даному описі як 5' фланкуюча послідовність) або їхні комплементарні ланцюги, також розкриті як SEQ ID NO: 111. В одному аспекті цього варіанта здійснення праймери фланкуючої послідовності вибрані із групи, яка складається з SEQ ID NO: 8 – SEQ ID NO: 14 і їх комплементарних ланцюгів. Фланкуючі послідовності можуть бути подовжені із включенням послідовностей хромосоми 5 з конкретним акцентом на нуклеотид, включений з SEQ ID NO: 103, придатний у виявленні послідовностей, зв'язаних з подією 5307 кукурудзи. У контексті SEQ ID NO: 103 “N” визначається як будь-яка основа “А”, “Т”, “G” або “C”. SEQ ID NO: 110 є зворотною комплементарною послідовністю цієї послідовності. У контексті SEQ ID NO: 110 “N” визначається як будь-яка основа “А”, “Т”, “G” або “C”. [0063] В іншому варіанті здійснення даний винахід охоплює праймери фланкуючої послідовності, які містять, щонайменше, 10-15 суміжних нуклеотидів з нуклеотидів 1-1093 SEQ ID NO: 6 (довільно позначена в даному описі як 3' фланкуюча послідовність) або їх комплементарні ланцюги. В одному аспекті цього варіанта здійснення праймери фланкуючої послідовності вибирають із групи, яка складається з SEQ ID NO: 69 – SEQ ID NO: 72 і їх комплементарних ланцюгів. [0064] У ще одному варіанті здійснення даний винахід охоплює пару полінуклеотидних праймерів, що включає перший полінуклеотидний праймер і другий полінуклеотидний праймер, які функціонують разом у присутності ДНК-матриці події 5307 кукурудзи в зразку із продукуванням амплікона, діагностичного для події 5307 кукурудзи, де перша праймерна послідовність безпосередньо являє собою або комплементарна геному рослини кукурудзи, що фланкує точку вставки гетерологічної послідовності ДНК, вставленої в геном рослини кукурудзи події 5307 кукурудзи, і друга полінуклеотидна праймерна послідовність безпосередньо являє собою або комплементарна вставленій у геном рослини кукурудзи гетерологічній послідовності ДНК події 5307 кукурудзи. [0065] В одному аспекті цього варіанта здійснення перший полінуклеотидний праймер включає, щонайменше, 10 суміжних нуклеотидів з положення 1-1348 SEQ ID NO: 5 або їх комплементарні ланцюги. У наступному аспекті цього варіанта здійснення перший полінуклеотидний праймер включає нуклеотидну послідовність, викладену в SEQ ID NO: 8 – SEQ ID NO: 14, або їх комплементарні ланцюги. В іншому аспекті цього варіанта здійснення перший полінуклеотидний праймер менш 10 суміжних нуклеотидів з положення 1-1093 SEQ ID NO: 6 або їх комплементарні ланцюги. В іншому аспекті цього варіанта здійснення перший полінуклеотидний праймер включає нуклеотидну послідовність, викладену в SEQ ID NO: 69 – SEQ ID NO: 72, або їх комплементарні ланцюги. У ще одному аспекті цього варіанта здійснення другий полінуклеотидний праймер включає, щонайменше, 10 суміжних нуклеотидів SEQ ID NO: 7 або їх комплементарні ланцюги. У наступному аспекті цього варіанта здійснення другий полінуклеотидний праймер включає нуклеотидну послідовність, викладену в SEQ ID NO: 15 SEQ ID NO: 68, або їх комплементарні ланцюги. [0066] В іншому аспекті цього варіанта здійснення перший полінуклеотидний праймер, який викладено в SEQ ID NO: 8, і другий полінуклеотидний праймер, який викладено в SEQ ID NO: 41, функціонують разом у присутності ДНК-матриці події 5307 кукурудзи в зразку із продукуванням амплікона, діагностичного для події 5307 кукурудзи, як описано в Прикладі 4. В іншому аспекті цього варіанта здійснення перший полінуклеотидний праймер, який викладено в SEQ ID NO: 69, і другий полінуклеотидний праймер, який викладено в SEQ ID NO: 72, функціонують разом у присутності ДНК-матриці події 5307 кукурудзи в зразку із продукуванням амплікона, діагностичного для події 5307 кукурудзи, як описано в Прикладі 4. [0067] У межах спеціальних знань добре було б отримати додаткову послідовність, що далі виходить у геномну послідовність, що фланкує той або інший кінець вставлених гетерологічних послідовностей ДНК, для застосування в якості праймерної послідовності, яку можна використовувати в таких парах праймерів для ампліфікації послідовностей, які є діагностичними для події 5307. Для цілей даного розкриття фраза "що далі виходить у геномну послідовність, яка фланкує той або інший кінець вставлених гетерологічних послідовностей ДНК" стосується, 10 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 зокрема, послідовного просування від кінців вставлених гетерологічних послідовностей ДНК, точок, у яких вставлені послідовності ДНК прилягають до нативної геномної послідовності ДНК, і виходу в геномну ДНК конкретної хромосоми, у яку було вставлено гетерологічні послідовності ДНК. Переважно, праймерна послідовність, що відповідає або комплементарна частині послідовності вставки, повинна служити запалом транскрипційному подовженню ланцюга, що утворюється, ДНК або РНК до найближчого з'єднання фланкуючої послідовності. Отже, праймерна послідовність, що відповідає або комплементарна частині геномної фланкуючої послідовності, повинна служити запалом транскрипційному подовженню ланцюга ДНК або РНК, що утворюється, до найближчого з'єднання фланкуючої послідовності. Праймерна послідовність може безпосередньо являти собою або може бути комплементарною гетерологічній послідовності ДНК, вставленій в хромосому рослини, або геномній фланкуючій послідовності. Фахівець у даній галузі легко визнає користь того, що праймерна послідовність або повинна являти собою, або повинна бути комплементарною послідовності, як викладено у вставленій гетерологічній послідовності ДНК або як викладено в SEQ ID NO: 3 або SEQ ID NO: 4, залежно від природи продукту, який бажають одержати за допомогою застосування гніздового набору праймерів, призначених для застосування в ампліфікації конкретної фланкуючої послідовності, що включає з'єднання між геномною послідовністю ДНК і вставленою гетерологічною послідовністю ДНК. Більш того, фахівець у даній галузі зможе розробити праймери для великох кількості генів нативної кукурудзи з метою розробки позитивного контролю. Один такий приклад є геном Adh1 кукурудзи, де приклади придатних праймерів для продукування амплікона за допомогою ампліфікації нуклеїнової кислоти викладені як SEQ ID NO: 79 і SEQ ID NO: 80. [0068] В іншому варіанті здійснення даний винахід охоплює спосіб виявлення присутності ДНК, що відповідає події 5307, у біологічному зразку, де спосіб включає: (a) взаємодію зразка, що включає ДНК, із зондом, який гібридизується в умовах високої жорсткості з геномною ДНК із події 5307 кукурудзи та не гібридизується в умовах високої жорсткості з ДНК контрольної рослини кукурудзи; (b) вплив на зразок і зонд гібридизаційними умовами високої жорсткості та (c) виявлення гібридизації зонда із ДНК. В одному аспекті цього варіанта здійснення амплікон включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 і їх комплементарних ланцюгів. [0069] В іншому варіанті здійснення даний винахід включає спосіб виявлення наявності ДНК, що відповідає події 5307, у біологічному зразку, де спосіб включає: (a) взаємодію зразка, що включає ДНК, із зондом, який гібридизується в умовах високої жорсткості з геномною ДНК із події 5307 кукурудзи та не гібридизується в умовах високої жорсткості з ДНК контрольної рослини кукурудзи; (b) вплив на зразок і зонд гібридизаційними умовами високої жорсткості та (c) виявлення гібридизації зонда із ДНК. Виявлення може здійснюватися будь-якими засобами, добре відомими у даній галузі, включаючи, але без обмеження, флуоресцентний, хемолюмінісцентний, радіологічний, імунологічний або інший. У випадку, у якому гібридизація передбачається для застосування в якості засобу для ампліфікації конкретної послідовності для одержання амплікона, який є діагностичним для події 5307 кукурудзи, одержання та виявлення будь-якими засобами, добре відомими у даній галузі, амплікона, як передбачається, вказує на передбачувану гібридизацію з мішеневою послідовністю, якщо використовують один зонд або праймер, або послідовностями, якщо використовують два або більше зондів або праймерів. Вираз "біологічний зразок", як мають на увазі, включає зразок, який містить або, як припускають, містить нуклеїнову кислоту, що включає від п'яти до десяти нуклеотидів на будь-якому боці точки, у якій один або інший із двох термінальних кінців вставленої гетерологічної послідовності ДНК контактує з геномною послідовністю ДНК у межах хромосоми, у яку вставили гетерологічну послідовність ДНК, у даному описі також відомі як з'єднуючі послідовності. До того ж, з’єднюча послідовність включає до двох нуклеотидів, якими є перший нуклеотид у межах фланкуючої геномної ДНК, суміжний з і ковалентно приєднаний до першого нуклеотиду в межах вставленої гетерологічної послідовності ДНК. [0070] У ще одному варіанті здійснення даний винахід включає набір для виявлення наявності нуклеїнових кислот події 5307 у біологічному зразку, де набір включає, щонайменше, одну молекулу нуклеїнової кислоти достатньої довжини суміжних нуклеотидів, гомологічних або комплементарних нуклеотидній послідовності, вибраній з групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 і SEQ ID NO: 4, яка функціонує як ДНК-праймер або зонд, специфічний для події 5307, і інші матеріали, необхідні, щоб уможливити гібридизацію або ампліфікацію нуклеїнової кислоти. Можна використовувати різноманітні способи виявлення, включаючи TAQMAN (Perkin Elmer), термічну ампліфікацію, лігазну ланцюгову реакцію, саузернгібридизацію, способи твердофазного імуноферментного аналізу, колориметричні та 11 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 флуоресцентні способи виявлення. Зокрема, даний винахід забезпечує набори для виявлення наявності мішеневої послідовності, тобто, щонайменше, одного із з'єднань вставки ДНК із геномною ДНК рослини кукурудзи в події 5307, у зразку, що містить геномну нуклеїнову кислоту з події 5307. Набір складається з, щонайменше, одного полінуклеотиду, здатного до зв'язування із сайтом-мішенню або, по суті, суміжно із сайтом-мішенню, і, щонайменше, одного засобу для виявлення зв'язування полінуклеотиду із сайтом-мішенню. Засоби виявлення можуть бути флуоресцентними, хемілюмінісцентними, колориметричними або ізотопними і можуть бути спряжені з, щонайменше, імунологічними способами виявлення зв'язування. Також передбачено набір, який може виявляти наявність сайту-мішені в зразку, тобто, щонайменше, одного із з'єднань ДНК вставки з геномною ДНК рослини кукурудзи в події 5307, використовуючи дві або більше полінуклеотидних послідовностей, які разом здатні до зв'язування з нуклеотидними послідовностями, суміжними з або в межах приблизно 100 пар основ, або в межах приблизно 200 пар основ, або в межах приблизно 500 пар основ, або в межах приблизно 1000 пар основ мішеневої послідовності, і які можуть бути подовжені в напрямку одна до одної з утворенням амплікона, який включає, щонайменше, сайт-мішень. [0071] В іншому варіанті здійснення даний винахід включає спосіб виявлення білка події 5307 у біологічному зразку, при цьому спосіб включає: (a) виділення білка зі зразка тканини події 5307 кукурудзи; (b) аналіз виділеного білка за допомогою імунологічного способу, що включає антитіло, специфічне до інсектицидного або селектовного маркерного білка, що продукується подією 5307; і (c) виявлення зв'язування зазначеного антитіла з інсектицидним або селектовним маркерним білком. [0072] Інший варіант здійснення даного винаходу охоплює рослину кукурудзи або його частини, що включають генотип трансгенної події 5307, де генотип включає нуклеотидну послідовність, викладену в SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, або їх комплементарні ланцюги. В одному аспекті цього варіанта здійснення рослина кукурудзи являє собою рослину з інцухт-ліній кукурудзи CG5NA58, CG5NA58A, CG3ND97, CG5NA01, CG5NF22, CG4NU15, CG00685, CG00526, CG00716, NP904, NP948, NP934, NP982, NP991, NP993, NP2010, NP2013, NP2015, NP2017, NP2029, NP2031, NP2034, NP2045, NP2052, NP2138, NP2151, NP2166, NP2161, NP2171, NP2174, NP2208, NP2213, NP2222, NP2275, NP2276, NP2316, BCTT609, AF031, H8431, 894, BUTT201, R327H, 2044BT і 2070BT. Фахівець даної галузі, однак, визнає, що генотип події 5307 може бути інтрогресований у будь-який сорт рослини, який може бути виведений з кукурудзою, включаючи дикі види маїсу, і, таким чином, переважні інцухт-лінії цього варіанта здійснення не призначені для обмеження. [0073] В іншому варіанті здійснення даний винахід охоплює рослину кукурудзи, що включає, щонайменше, першу та другу послідовність ДНК, з'єднану разом з утворенням суміжної нуклеотидної послідовності, де перша послідовність ДНК знаходиться в межах з'єднуючої послідовності та включає, щонайменше, приблизно 10-15 суміжних нуклеотидів, вибраних із групи, що складається з нуклеотидів SEQ ID NO: 5, SEQ ID NO: 6 і їх комплементарних ланцюгів, де друга послідовність ДНК знаходиться в межах гетерологічної послідовності ДНК вставки, вибраної із групи, що складається з SEQ ID NO: 15 – SEQ ID NO: 68 і їх комплементарних ланцюгів; і де перша та друга послідовності ДНК є придатними в якості нуклеотидних праймерів або зондів для виявлення наявності послідовностей нуклеїнових кислот події 5307 кукурудзи в біологічному зразку. В одному аспекті цього варіанта здійснення нуклеотидні праймери застосовують у способі ампліфікації ДНК для ампліфікації мішеневої послідовності ДНК з матриці ДНК, виділеної з рослини кукурудзи, і рослина кукурудзи ідентифікується від інших рослин кукурудзи за допомогою продукування амплікона, що відповідає послідовності ДНК, що включає SEQ ID NO: 1 або SEQ ID NO: 2. [0074] Рослини кукурудзи даного винаходу можна додатково охарактеризувати тим, що розщеплення геномної ДНК рослини рестрикційними ендонуклеазами SmaI і PmeI дає в результаті єдину смугу гібридизації з використанням зонда повної довжини в умовах високої жорсткості. Наведений у прикладі в даному описі являє собою зонд повної довжини, що включає нуклеотидну послідовність, викладену в SEQ ID NO: 7. [0075] В одному варіанті здійснення даний винахід забезпечує рослину кукурудзи, де генотип події 5307 надає рослині кукурудзи стійкість до комах або здатність використовувати манозу. В одному аспекті цього варіанта здійснення генотип, що надає стійкість до комах рослині кукурудзи, включає ген FR8a. В іншому аспекті цього варіанта здійснення генотип, що надає рослині кукурудзи здатність використовувати манозу, включає ген PMI. [0076] В одному варіанті здійснення даний винахід забезпечує біологічний зразок, отриманий з рослини кукурудзи, тканини або насіння події 5307, де зразок включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна послідовності, вибраній із 12 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 і SEQ ID NO: 4, і де послідовність виявляється в зразку за допомогою способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти. Таким чином, генетична послідовність функціонує як засіб виявлення. В одному аспекті цього варіанта здійснення зразок є вибраним з кукурудзяного борошна, кукурудзяної мучки, кукурудзяної патоки, кукурудзяної олії, кукурудзяного крохмалю та круп, зроблених повністю або частково з вмістом кукурудзяних продуктів. [0077] В іншому варіанті здійснення даний винахід забезпечує екстракт, отриманий з рослини кукурудзи, тканині або насіння події 5307, що включає нуклеотидну послідовність, яка безпосередньо являє собою або комплементарна послідовності, вибраній із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 і SEQ ID NO: 4. Приклад такого насіння депоновано в ATCC під інвентарним № PTA-9561. В одному аспекті цього варіанта здійснення послідовність виявляють в екстракті з використанням способу ампліфікації нуклеїнової кислоти або гібридизації нуклеїнової кислоти. В іншому аспекті цього варіанта здійснення зразок вибрано з кукурудзяного борошна, кукурудзяної патоки, кукурудзяної олії, кукурудзяного крохмалю та круп, зроблених повністю або частково з вмістом кукурудзяних продуктів. [0078] У ще одному варіанті здійснення даний винахід забезпечує спосіб одержання рослини кукурудзи, стійкої, щонайменше, до ураження кукурудзяним жуком, що включає: (a) статеве схрещування першої батьківської рослини кукурудзи із другою батьківською рослиною кукурудзи, де зазначена перша або друга батьківська рослина кукурудзи включає ДНК події 5307 кукурудзи, таким чином, одержуючи велику кількість рослин-нащадків першого покоління; (b) добір рослини-нащадка першого покоління, яка є стійкою, щонайменше, до ураження кукурудзяним жуком; (c) самозапилення рослини-нащадка першого покоління, таким чином, одержуючи велику кількість рослин-нащадків другого покоління; і (d) добір з рослин-нащадків другого покоління рослини, яка є, щонайменше, стійкою до ураження кукурудзяним жуком; де рослини-нащадки другого покоління включають нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO:3 і SEQ ID NO: 4. [0079] В іншому варіанті здійснення даний винахід забезпечує спосіб одержання гібридного насіння кукурудзи, що включає: (a) посів насіння першої інцухт-лінії кукурудзи, що включає нуклеотидну послідовність, вибрану із групи, що складається з SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 і SEQ ID NO: 4, і насіння другої інцухт-лінії, що має відмінний генотип; (b) вирощування рослин кукурудзи, одержуваних у результаті зазначеного посіву, до часу появи квіток; (c) видалення недоспілих маточок зазначених квіток рослин однієї з інцухт-ліній кукурудзи; (d) статеве схрещування однієї з однією двох відмінних інцухт-ліній; і (e) збирання гібридного насіння, отриманого таким чином. В одному аспекті цього варіанта здійснення перша інцухт-лінія кукурудзи забезпечує жіночі батьківські форми. В іншому аспекті цього варіанта здійснення перша інцухт-лінія кукурудзи забезпечує чоловічі батьківські форми. Даний винахід також включає гібридну насінину, отриману способом за варіантом здійснення, і гібридну рослину, вирощену з насінини. [0080] В іншому варіанті здійснення даний винахід забезпечує спосіб добору маркерів, пов'язаних з подією 5307 кукурудзи, що включає: (a) скринінг послідовностей хромосоми 5 події 5307 кукурудзи, (b) порівняння їх з нетрансгенними послідовностями NP2222, (c) порівняння послідовностей з метою виявлення варіацій послідовностей, (d) застосування цих варіацій послідовностей як засобу для створення маркерів, пов'язаних з подією 5307 кукурудзи, (e) застосування маркерів для скринінга ліній, і (f) виявлення маркера, що підтверджує наявність послідовностей події 5307 кукурудзи на хромосомі 5. [0081] Фахівець у даній галузі визнає, що трансгенний генотип даного винаходу може бути інтрогресований за допомогою селекції в інші лінії кукурудзи, що включають відмінні трансгенні генотипи. Наприклад, інцухтованну рослину кукурудзи, що включає трансгенний генотип даного винаходу, можна схрестити з інцухтованою рослиною кукурудзи, що включає трансгенний генотип події Bt11 стійкості до лускокрилих, яка відомо в даній галузі, таким чином, одержуючи кукурудзяну насінину, яка включає як трансгенний генотип даного винаходу, так і трансгенний генотип Bt11. Приклади інших трансгенних подій, які можна схрестити з інцухтованою рослиною даного винаходу, включають події витривалості до гербіциду гліфосату GA21 і NK603, подію витривалості до гліфосату/стійкості до лускокрилих комах MON802, подію стійкості до лускокрилих комах DBT418, подію стійкості до лускокрилих комах DAS-06275-8, подію стійкості до лускокрилих комах MIR162, подію чоловічої стерильності MS3, подію витривалості до фосфінотрицину B16, подію стійкості до лускокрилих комах MON 80100, події витривалості до фосфінотрицину T14 і T25, подію стійкості до лускокрилих комах 176, подію стійкості до твердокрилих комах MIR604 і подію стійкості до твердокрилих комах MON863, всі з яких відомі в 13 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 даній галузі. Крім того, буде визнано, що можна здійснити інші комбінації із трансгенним генотипом даного винаходу, і, таким чином, ці приклади не повинні розглядатися як обмежуючі. [0082] Фахівець у даній галузі також визнає, що трансгенну кукурудзяну насінину, що включає трансгенний генотип даного винаходу, можна обробляти різними хімікатами для обробки насіння, включаючи інсектициди, для посилення або синергії інсектицидної активності білка FR8a. Наприклад, трансгенну кукурудзяну насінину даного винаходу можна обробити комерційним інсектицидом Cruiser®. Таку комбінацію можна застосовувати для підвищення спектра активності та для підвищення ефективності білка, який експресується, та хімікату. Селекція [0083] Трансгенний генотип даного винаходу може бути інтрогресований у будь-яку інцухтовану рослину або гібрид кукурудзи за допомогою селекційних методик, визнаних у даній галузі техніки. Метою селекції рослини є об'єднання в одному сорті або гібриді різних бажаних ознак. Для польових культур ці ознаки можуть включати стійкість до комах і захворювань, витривалість до гербіцидів, витривалість до жари та посухи, зменшення часу дозрівання врожаю, більш високу врожайність і кращу агрономічну якість. При механізованому збиранні багатьох культур важлива однорідність характеристик рослин, таких як початок розвитку та встановлення густоти стояння рослин, швидкість росту, визрівання та висота рослини й качана. [0084] Польові культури селектують за допомогою методик, які використовують спосіб запилення рослин. Рослина є самозапилювальною, якщо пилок з однієї квітки переносять на ту саму або іншу квітку тієї ж рослини. Рослина є перехресно запилювальною, якщо пилок беруть з квітки на іншій рослині [0085] Рослини, які піддавали самозапиленню та добору за типом протягом багатьох поколінь, стають гомозиготними майже за всіма генними локусами і дають однорідну популяцію потомства розведення гомозигот. Схрещування між двома різними гомозиготними лініями дає однорідну популяцію гібридних рослин, які можуть бути гетерозиготними за багатьма генними локусами. Схрещування двох рослин, кожна з яких гетерозиготна за рядом генних локусів, буде давати популяцію гібридних рослин, які відризняються генетично та не будуть однорідними. [0086] Кукурудзу можна селектувати методиками як самозапилення, так і перехресного запилення. Кукурудза має окремі чоловічі та жіночі квітки на одній рослині, розташовані на волоті та качані, відповідно. Природне запилення відбувається в кукурудзи при здуванні вітром пилку з волоті на шовки, які викидаються з верхівок качанів. [0087] Надійний спосіб контролю чоловічої фертильності в рослин забезпечує можливість покращеної селекції рослин. Це особливо актуально для створення кукурудзяних гібридів, що покладається на будь-яку систему чоловічої стерильності. Існує кілька варіантів контролю чоловічої фертильності, доступних для селекціонерів, таких як ручне або механізоване видалення недоспілих маточок (або видалення волоті), цитоплазматична чоловіча стерильність, генетична чоловіча стерильність, гаметоциди та подібне. [0088] Гібридну кукурудзяну насінину, як правило, одержують за допомогою системи чоловічої стерильності, що включає ручне або механізоване видалення волоті. Смуги, що чергуються, двох типів інцухтованих рослин кукурудзи висівають у поле та волоті, що несуть пилок, видаляють в одних з інцухтованих рослин (жіночих). За умови, що існує достатня ізоляція від джерел пилку чужорідної кукурудзи, качани інцухтованих рослин з вилученими волотями будуть запліднені тільки від іншої інцухтованої рослини (чоловічої), і отримане насіння є, отже, гібридним та буде утворювати гібридні рослини. [0089] Трудомісткого та часом ненадійного процесу видалення волоті можна уникнути застосуванням одного з багатьох способів надання генетичної чоловічої стерильності в даній галузі, кожний з яких зі своїми власними перевагами та недоліками. У цих способах застосовують ряд підходів, таких як доставка в рослину гена, що кодує цитотоксичну речовину, пов'язану із промотором, специфічним для чоловічої тканини, або антизмістовною системою, у якій ідентифіковано ген, критичний для фертильності, і вставляють у рослину антизмістовний такому гену (дивись: Fabinjanski, et al. EPO 89/3010153.8 публікація № 329308 і РСТ заявка PCT/CA90/00037, опублікована як WO 90/08828). Створення інцухт-ліній кукурудзи [0090] Застосування інцухтованих рослин з чоловічою стерильністю є лише одним з факторів при одержанні гібридів кукурудзи. Методики селекції рослин, відомі в даній галузі та застосовувані в плані з селекції рослин кукурудзи, включають, але не обмежуються, періодичний добір, зворотне схрещування, педігрі селекцію, добір, посилений поліморфізмом довжин рестрикційних фрагментів, добір за допомогою маркера та трансформацію. Створення гібридів кукурудзи в плані з селекції рослин кукурудзи вимагає, в основному, створення гомозиготних інцухт-ліній, схрещування цих ліній і оцінювання кросів. Способи педігрі селекції та 14 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 періодичного добору використовують для створення інцухт-ліній із селекційних популяцій. Плани з селекції рослин кукурудзи поєднують генетичні фони із двох або більш інцухт-ліній або різних інших джерел зародкової плазми в селекційні пули, з яких створюють нові інцухт-лінії шляхом самозапилення та добору бажаних фенотипів. Нові інцухтовані рослини схрещують з іншими інцухт-лініями та гібриди від цих схрещувань оцінюють задля визначення таких, що мають комерційний потенціал. Селекція рослин і створення гібридів, як здійснюється на практиці в плані з селекції рослин кукурудзи, є дорогими та витратними за часом процесами. [0091] Педігрі селекція починається зі схрещування двох генотипів, кожен з яких може мати одну або більше бажаних характеристик, яких не вистачає в іншому або які доповнюють інший. Якщо двоє вихідних батьків не дають усіх бажаних характеристик, у селекційну популяцію можна включити інші джерела. У педігрі способі старші рослини самозапилюють і добирають у наступних поколіннях. У наступних поколіннях гетерозиготний стан заступається однорідними лініями в результаті самозапилення й добору. Як правило, на практиці використовують педігрі спосіб селекції п'яти або більш поколінь самозапилення та добору: F 1 F2; F2 F3; F3 F4; F4 F5 і т.д. [0092] Селекція періодичним добором, наприклад, зворотне схрещування, може використовуватися для поліпшення інцухт-лінії та гібрида, який створюють за допомогою таких інцухтованих рослин. Зворотне схрещування можна використовуватися для переносу конкретної бажаної ознаки від однієї інцухтованої рослини або джерела в інцухтовану рослину, якій не вистачає такої ознаки. Це можна виконати, наприклад, спочатку шляхом схрещування старшої інцухтованої рослини (рекурентний батько) з донорною інцухтованою рослиною (нерекурентний батько), яка несе відповідний ген(гени) для ознаки, яку розглядають. Потомство цього схрещування потім зворотно схрещують зі старшим рекурентним батьком з наступним добором в отриманого потомства за бажаною ознакою, яка підлягає перенесенню від нерекурентного батька. Через п'ять або більш поколінь зворотного схрещування з добором за бажаною ознакою потомство буде гомозиготним за локусами, що контролюють характеристику, яку переносять, але буде схоже на старшого батька фактично за всіма іншими генами. Покоління останнього зворотного схрещування потім самозапилюють з одержанням чистого селекційного потомства за геном(генами), який переносять. Гібрид, створений з інцухтованих рослин, що містять перенесений ген(гени), є, по суті, таким же як і гібрид, створений від тих самих інцухтованих рослин без перенесеного гена(генів). [0093] Елітні інцухт-лінії, тобто чисті селекційні гомозиготні інцухт-лінії, можна також використовувати в якості вихідних матеріалів для селекційних або початкових популяцій, з яких можна створити інші інцухт-лінії. Ці інцухт-лінії, що походять від елітних інцухт-ліній, можна створити за допомогою способів педігрі селекції та селекції періодичним добором, описаних раніше. Наприклад, якщо використовують селекцію зворотними схрещуванням для створення таких похідних ліній у плані з селекції рослин кукурудзи, то можна використовувати елітні інцухтовані рослини у якості батьківської лінії, або вихідного матеріалу, або початкової популяції, і вони можуть бути або в якості донора, або в якості рекурентного батька. Створення гібридів кукурудзи [0094] Простий гібрид кукурудзи походитьвід схрещування двох інцухт-ліній, кожна з яких має генотип, який доповнює генотип іншої. Гібридне потомство першого покоління позначають F1. При створенні комерційних гібридів у плані з селекції рослин кукурудзи необхідні тільки гібридні рослини F1. Переважні гібриди F1 є більш сильними, ніж їх інцухтовані батьки. Ця гібридна сила, або гетерозис, може проявлятися в багатьох полігенних ознаках, включаючи підвищений вегетативний ріст і покращену врожайність. [0095] Створення гібрида кукурудзи в плані з селекції рослин кукурудзи включає три етапи: (1) добір рослин з різних пулів зародкової плазми для початкових селекційних схрещувань; (2) самозапилення відібраних рослин із селекційних схрещувань у декількох поколіннях з одержанням ряду інцухт-ліній, які, хоча й відрізняються один від одного, передають ознаки потомству і є високо однорідними; та (3) схрещування відібраних інцухт-ліній з іншими інцухтлініями з одержанням гібридного потомства (F1). Під час процесу інцухта в кукурудзи сила ліній зменшується. Силу відновлюють при схрещуванні двох різні інцухт-ліній з одержанням гібридного потомства (F1). Важливим наслідком гомозиготності та гомогенності інцухт-ліній є те, що гібрид між визначеною парою інцухтованих рослин буде завжди однаковим. Після ідентифікації інцухтованих рослин, які дають переважаючий гібрид, гібридне насіння можна відтворювати нескінченно, доки зберігається гомогенність інцухтованих батьків. Більша частина гібридної сили, що виявлялася гібридами F 1, втрачається в наступному поколінні (F2). Отже, насіння від гібридів не використовують для посадкового матеріалу. [0096] Одержання гібридного насіння вимагає виключення або інактивації пилку, утвореного 15 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 жіночою батьківською формою. Неповне видалення або інактивація пилку дає можливість самозапилення. Таке ненавмисно самозапилене насіння може бути ненавмисно зібране та упаковане з гібридним насінням. [0097] Після висівання насіння можна ідентифікувати та відібрати такі самозапилені рослини. Такі самозапилені рослини будуть генетично еквівалентні жіночій інцухт-лінії, використаній для одержання гібрида. [0098] Як очевидно для фахівця в даній галузі, вищезгадані є лише деякими з різних способів, за допомогою яких інцухтована рослина даного винаходу може бути отримана фахівцями, що прагнуть інтрогресувати трансгенний генотип даного винаходу в інші лінії кукурудзи. Доступні інші засоби, і вищенаведені приклади є лише ілюстративними. Приклади [0099] Даний винахід буде далі описаний за допомогою посилання на наступні докладні приклади. Ці приклади наведені лише з метою ілюстрації та не призначені обмежувати, якщо не визначене інше. Стандартні методики рекомбінантної ДНК і молекулярного клонування, використовувані в даному описі, добре відомі в даній галузі й описуються в Ausubel (ed.), Current Protocols in Molecular Biology, John Wiley and Sons, Inc. (1994); J. Sambrook, et al., Molecular Cloning: A Laboratory Manual, 3d Ed., Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press (2001); і в T.J. Silhavy, M.L. Berman і L.W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984). Приклад 1. Трансформація та добір події 5307 [00100] Подію 5307 одержали за допомогою опосередкованої Agrobacterium трансформації кукурудзяної (Zea mays) інцухт-лінії NP2222. Незрілі ембріони трансформували, у цілому, як описано в Negrotto et al. (Plant Cell Reports 19: 798-803, 2000), що включено в даний опис посиланням, за допомогою фрагмента ДНК із плазміди pSYN12274 (Фігура 1). pSYN12274 містить нуклеотидну послідовність, що включає тандемні касети експресії. Перша касета експресії включає CMP промоторну послідовність (патент США № 7166770), функціонально зв'язану з кодуючою послідовністю FR8a, додатково функціонально зв'язаною з 3’ кінцевою послідовністю термінації транскрипції та поліаденілування нопалінсинтази. Друга касета експресії включає промотор убіквинтину маїсу (ZmUbiInt) (Christensen et al. 1992 PMB 18: 675), функціонально зв'язаного з кодуючою послідовністю PMI, додатково функціонально зв'язаною з 3’ кінцевою послідовністю термінації транскрипції та поліаденілування нопалінсинтази. [00101] Незрілі ембріони вилучили з качанів віком 8-12 днів і промили свіжим середовищем при підготовці для трансформації. Ембріони змішали із суспензією клітин Agrobacterium, що містять вектор трансформації pSYN12274, струшували на вортексі протягом 30 секунд і дали інкубуватися протягом додаткових 5 хвилин. Надлишок розчину Agrobacterium аспірували та ембріони потім перемістили на чашки, що містили неселективне культуральне середовище. Ембріони сумісно культивували з Agrobacterium, що залишилися, при 22°C протягом 2-3 днів у темряві. Ембріони перенесли на культуральне середовище, доповнене тикарциліном (100 мг/мл) і нітратом срібла (1,6 мг/л), і інкубували в темряві протягом 10 днів. Ембріони, що продукували ембріогений калюс, перенесли на середовище для клітинних культур, що містить манозу. [00102] Регенеровані проростки протестували за допомогою ПЛР-аналізу TAQMAN® (див. Приклад 2) на наявність як PMI, так і FR8a генів, а також на відсутність гена стійкості до антибіотика спектиноміцину (spec). Рослини, позитивні за обома трансгенами та негативні за геном spec, перенесли в теплицю для подальшого розведення. Позитивні події ідентифікували та піддали скринінгу за допомогою біоаналізів проти кукурудзяного жука. Інсектицидні події відрізняли за числом копій за допомогою аналізу TAQMAN. Подію 5307 вибрали для подальшого аналізу на основі того, що вона мала одну копію трансгенів, гарну експресію білка, як ідентифікували за допомогою твердофазного імуноферментного аналізу, та кращу інсектицидну активність проти кукурудзяного жука в порівнянні з іншими подіями, створеними з тим же конструктом. [00103] T0 подію 5307 зворотно схрестили з інцухт-лінією кукурудзи NP2460, створивши популяцію T1. Рослини T1 самозапилили для створення покоління T 2 і цей процес повторили для створення покоління T3. Вдалися до перевірки за потомством рослин T 3 для ідентифікації гомозиготних (обернених) родин. Обернені за подією 5307 інцухтовані рослини NP2460 схрестили з іншими елітними інцухт-лініями для створення гібридів, використовуваних у подальших дослідженнях. Приклад 2. Виявлення події 5307 за допомогою ПЛР TAQMAN [00104] Аналіз TAQMAN проводили, головним чином, як описано в Ingham et al. (Biotechniques, 31:132-140, 2001), включеному в даний опис посиланням. Коротко, геномну ДНК 16 UA 110320 C2 5 10 15 20 виділили з листів трансгенних і нетрансгенних рослин кукурудзи за допомогою набору для виділення геномної ДНК Puregene® (Gentra Systems, Мінеаполіс, Мінесота), головним чином, за інструкцією виробника, за винятком того, що всі етапи проводили в 1,2 мл 96-ямкових планшетах. Висушений залишок ДНК ресупендували в TE-буфері (10 мМ Tris-HCl, pH 8,0, 1 мМ EDTA). [00105] Реакції ПЛР TAQMAN проводили в 96-ямкових планшетах. Для внутрішнього контролю гена кукурудзи сконструювали праймери та зонди, специфічні до гена алкогольдегідрогенази (Adh) Zea mays (інвентарний номер Genbank AF044295). Фахівець у даній галузі зрозуміє, що можна використовувати інші гени кукурудзи як внутрішні контролі. Реакції об'єднали для одночасної ампліфікації FR8a і Adh або PMI і Adh. Для кожного зразка створили вихідну суміш шляхом об'єднання 20 мкл екстрагованої геномної ДНК із 35 мкл 2 x TAQMAN Universal PCR Master Mix (Applied Biosystems), доповненої праймерами до кінцевої концентрації 900 нМ кожного, зондами до кінцевої концентрації 100 нМ кожного та водою до 70 мкл кінцевого об’єму. Цю суміш розподілили на три повторності по 20 мкл кожна в 96-ямкові планшети для ампліфікації та запечатали оптично прозорою теплоізолюючою плівкою (Marsh Bio Products). ПЛР проводили в приладі ABI Prism 7700 з використанням наступних параметрів ампліфікації: 2 хв. при 50 °C і 10 хв. при 95 °C з наступними 35 циклами 15 секунд при 95 °C і 1 хв. при 60 °C. [00106] Результати аналізу TAQMAN продемонстрували, що подія 5307 мала одну копію гена FR8a і одну копію гена PMI. [00107] Приклади придатних комбінацій послідовностей праймер/зонд, які були використані, являють собою: Назва праймера FR8 a-прямий FR8 a-зворотний FR8 a-зонд Pmi-Прямий Pmi-Зворотний Pmi-Зонд Zmadh-267 прямий Zmadh-337 зворотний Zmadh-316 зонд 25 Праймерна послідовність 5’- TACGAGAGCTGGGTGAACTTCA-3’ 5’-CGATCAGGTCCAGCACGG -3’ 5’-CCGCTACCGCCGCGAGATGA-3’ (5’ мітка = FAM, 3’ мітка = TAMRA) 5’-CCGGGTGAATCAGCGTTT-3’ 5’-GCCGTGGCCTTTGACAGT-3’ 5’-TGCCGCCAACGAATCACCGG-3’ (5’ мітка = FAM, 3’ мітка = TAMRA) 5’-GAACGTGTGTTGGGTTTGCAT-3’ 5’-TCCAGCAATCCTTGCACCTT-3’ 5’-TGCAGCCTAACCATGCGCAGGGTA-3’ (5’ мітка= TET, 3’ мітка = TAMRA) SEQ ID NO: SEQ ID NO: 73 SEQ ID NO: 74 SEQ ID NO: 75 SEQ ID NO: 76 SEQ ID NO: 77 SEQ ID NO: 78 SEQ ID NO: 79 SEQ ID NO: 80 SEQ ID NO: 81 [00108] Аналізи TAQMAN PM1271, MIC5307a і MIC5307b розроблені в якості специфічного для події аналізу, який охоплює 3’ з'єднуючу послідовність. [00109] Приклади придатних комбінацій послідовностей праймер/зонд, які були використані, являють собою: Назва праймера PM 1277-прямий PM 1277-зворотний PM 1277-зонд MIC5307 a-прямий MIC5307 a-зворотний MIC5307 a-зонд MIC5307 b-прямий MIC5307 b-зворотний MIC5307 b-зонд Праймерна послідовність 5’-GCCGTATCCGCAATGTGTTA-3’ 5’-GGCCCAGGGAAGAGGGTATAT-3’ 5’-AAGTTGTCTAAGCGTCAAT-3’ (5’ мітка = TET, 3’ мітка = TAMRA) 5’-TGTCTAAGCGTCAATTTGTTTACACC-3’ 5’-TTTGCCAGTGGGCCCA-3’ 5’-ACAATATACCCTCTTCCCTGGGCCAGG-3’ (5’ мітка= TET, 3’ мітка = TAMRA) 5’-GCCGTATCCGCAATGTGTTA-3’ 5’-AAGTTGTCTAAGCGTCAAT-3’ 5’-GGCCCAGGGAAGAGGGTATAT-3’ (5’ мітка= TET, 3’ мітка = TAMRA) SEQ ID NO: SEQ ID NO: 82 SEQ ID NO: 83 SEQ ID NO: 84 SEQ ID NO: 82 SEQ ID NO: 83 SEQ ID NO: 84 SEQ ID NO: 82 SEQ ID NO: 83 SEQ ID NO: 84 30 Приклад 3. Виявлення події 5307 за допомогою саузерн-блотинга [00110] Геномну ДНК, використовувану для аналізу за Саузерном, виділили зоб'єднаної в пул тканини листя десяти рослин, що представляють покоління шостого зворотного 17 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 схрещування (BC6) події 5307, за допомогою, головним чином, способу Thomas et al. (Theor. Appl. Genet. 86:173-180, 1993), який включено в даний опис посиланням. Усі рослини, використані для виділення ДНК, окремо проаналізували за допомогою ПЛР TAQMAN (як описано в Прикладі 2) для підтвердження наявності однієї копії гена FR8a і гена PMI. Для негативних сегрегантних контролів виділили ДНК із об'єднаної в пул тканини листя п'яти рослин, що представляють покоління BC4 події 5307. Ці негативні сегрегантні рослини окремо проаналізували за допомогою ПЛР TAQMAN, і аналізи були негативними на наявність гена FR8a і гена PMI, але, як і очікувалося, були позитивними за внутрішнім контролем аналізу, ендогенним геном Adh маїсу. [00111] Аналіз за Саузерном виконали за допомогою традиційних методик молекулярної біології. Геномну ДНК (7,5 мкг) дворазово розщепили за допомогою рестрикційих ферментів SmaI і PmeI, які мали єдині сайти впізнавання в межах Т-ДНК-вставки події 5307 із плазміди pSYN12274 (Фігура 1). Цей підхід передбачає визначення числа копій елементів, що відповідають конкретному зонду, використовуваному для кожного аналізу за Саузерном, які були включені в подію 5307. Це дає в результаті одну смугу гібридизації на копію елемента, що присутній в події 5307. Після електрофорезу в агарозному гелі та лужного переносу на мембрану Nytran® гібридизації проводили за допомогою специфічних до елементів створених за допомогою ПЛР зондів повної довжини. Зонд повної довжини, використовуваний у саузернблотингах, включає нуклеотидні послідовності, викладені в SEQ ID NO: 7. Зонд помітили 32P за допомогою випадкового запалювання із застосуванням системи Rediprime™ II (Amersham Biosciences, кат. № RPN1633). [00112] Використовували наступні умови гібридизації високої жорсткості: 1-2 мільйона імп./хв./мл додали в PerfectHyb (Sigma), доповнену 100 мкг/мл ДНК тимусу теляти (Invitrogen), попередньо нагріту до 65 °C. Попередня гібридизація проходить у тому ж розчині, який описано вище, при тій же температурі протягом ночі або протягом, щонайменше, однієї години. Гібридизацію проводять при 65 °C протягом 3 годин з наступним 2X відмиванням в 2X SSC, 0,1% SDS протягом 20 хвилин при 65 °C і 2X в 0,1X SSC, 0,1% SDS протягом 20 хвилин при 65 °C. [00113] У кожний аналіз за Саузерном включили три контрольні зразки: (1) ДНК із негативного (нетрансформованого) сегреганта, використовувана для ідентифікації будь-яких ендогенних послідовностей Zea mays, які можуть перехресно гібридизуватися зі специфічним до елемента зондом; (2) ДНК із негативного сегреганта, у яку вводять деяку кількість pSYN12274, розщепленої SmaI-PmeI, яка дорівнює кількості однієї копії за довжиною зонда, для демонстрації чутливості експерименту при виявленні одиничної копії гена в геномі Zea mays; і (3) плазміда pSYN12274, розщеплена SmaI-PmeI, яка дорівнює кількості однієї копії за довжиною зонда, у якості позитивного контролю для гібридизації, а також для демонстрації чутливості експерименту. [00114] Дані гібридизації надають підкріплювальний доказ для підтвердження ПЛР-аналізу TAQMAN, що подія 5307 включає єдину копію генів FR8a і PMI, і що подія 5307 не містить будьяких векторних остовних послідовностей, що присутні в pSYN12274. Як очікувалося і для FR8a, і для PMI зондів, розщеплення SmaI-PmeI дало в результаті єдину смугу гібридизації правильного розміру, демонструючи, що в події 5307 є присутньою єдина копія кожного гена. До того ж, для остовного зонда відсутність гібридизації демонструє відсутність будь-яких остовних послідовностей вектора pSYN12274, які включаються в подію 5307 під час процесу трансформації. Приклад 4. Секвенування T-ДНК-вставки [00115] Нуклеотидну послідовність всієї трансгенної ДНК-вставки, що присутня у події 5307, визначили для демонстрації повної цілісності вставки, суміжності функціональних елементів і задля виявлення будь-яких змін окремих пар основ. Вставку події 5307 ПЛР-ампліфікували із ДНК, отриманої з покоління BC5, у вигляді двох окремих фрагментів, що перекриваються. Кожний фрагмент ампліфікували за допомогою одного полінуклеотидного праймера, гомологічного рослинним геномним послідовностям, що фланкують вставку події 5307, і одного полінуклеотидного праймера, гомологічного гену FR8a. Для створення 5’ фрагмента перший полінуклеотидний праймер, гомологічний 5’ фланкуючій послідовності, SEQ ID NO: 8 – SEQ ID NO: 15, об'єднали з другим полінуклеотидним праймером, гомологічним вставленій ДНК гена FR8a, SEQ ID NO: 33 – SEQ ID NO: 41, промотором убіквинтина, SEQ ID NO: 42 – SEQ ID NO: 53 або геном PMI, SEQ ID NO: 54 – SEQ ID NO: 60. Для створення 3’ фрагмента перший полінуклеотидний праймер, гомологічний 3’ фланкуючій послідовності, SEQ ID NO: 69 – SEQ ID NO: 72, об'єднали з другим полінуклеотидним праймером, гомологічним вставленій ДНК у гені FR8a, SEQ ID NO: 9 – SEQ ID NO: 17, промотором убіквинтина, SEQ ID NO: 18 – SEQ ID NO: 26 18 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 або геном PMI, SEQ ID NO: 27 – SEQ ID NO: 32. [00116] ПЛР-ампліфікацію проводили із застосуванням розширеної ПЛР-системи високої точності (Roche, кат. № 1732650) і наступними параметрами ампліфікації: 2 хв. при 94 °C на 1 цикл з наступними 10 циклами 15 сек при 94 °C, 30 сек при 55-65 °C і 5 хв. при 68 °C, з наступними 20 циклами 15 сек при 94 °C, 30 сек при 55-65 °C і 5 хв+5 сек/цикл 72 °C з наступним 1 циклом 7 хв. при 72 °C. [00117] Амплікон, отриманий у результаті ПЛР-ампліфікації за допомогою SEQ ID NO: 8 і SEQ ID NO: 41, включав 5’ з'єднуючу послідовність (SEQ ID NO: 1). Амплікон, отриманий у результаті ПЛР-ампліфікації за допомогою SEQ ID NO: 69 і SEQ ID NO: 72, включав 3’ з'єднуючу послідовність (SEQ ID NO: 2). Кожний фрагмент секвенування окремо клонували у вектор pCR® -XL-TOPO (Invitrogen, кат. № K4700-20), ідентифікували та секвенували три окремі клони для кожного фрагмента. Секвенування провели за допомогою аналізатора ABI3730XL з використанням хімії dGTP ABI BigDye® 1.1 або Big Dye 3.1 (для матриць, багатих на GC). Аналіз послідовностей виконали за допомогою пакета Phred, Phrap і Consed з Вашингтонського університету та провели із частотою помилок менш ніж 1 на 10000 основ (Ewing і Green, 1998). Кінцеву консенсусну послідовність визначили шляхом об'єднання даних послідовностей від шести окремих клонів (три на кожен фрагмент секвенування) для створення однієї консенсусної послідовності вставки події 5307. Для подальшого підтвердження будь-яких невідповідностей окремих пар основ між вставкою події 5307 і плазмідою pSYN12274 за допомогою такої ж методології, як описано раніше, ампліфікували невеликі (приблизно 300-500 п.о.) ПЛР-продукти, специфічні до будь-яких ділянок, де спостерігали невідповідність пар основ у початковій консенсусній послідовності. Для всіх передбачуваних невідповідностей пар основ у вставці події 5307 пряме секвенування ПЛР-продукту дало в результаті одиничні чисті піки у всіх розглянутих парах основ, вказуючи на те, що ці невідповідності ймовірно присутні у вставці події 5307. Провели вирівнювання за допомогою програми ClustalW з наступними параметрами: матриця підрахунку blosum55, штраф за відкриття гепа 15, штраф за подовження гепа 6,66 (Thompson et al, 1994, Nucleic Acids Research, 22, 4673-4680). [00118] Дані консенсусної послідовності для Т-ДНК-вставки події 5307 демонструють, що вся цілісність вставки та суміжність функціональних елементів у вставці, як передбачалося в pSYN12274, збереглися. Приклад 5. Аналіз фланкуючої послідовності ДНК [00119] ДНК-послідовність генома кукурудзи, що фланкує гетерологічну ДНК, вставлену в геном рослини кукурудзи події 5307, одержали за допомогою технології OmniPlex™, головним чином описаної в Kamberov et al (Proceedings of SPIE, Tools for Molecular Analysis and HighThroughput Screening, 4626:1-12, 2002), що включена в даний опис посиланням. [00120] 5’ і 3’ фланкуючі послідовності та з'єднуючі послідовності підтверджували за допомогою стандартних ПЛР-процедур. 5’ фланкуючі та з'єднуючі послідовності підтвердили із застосуванням першого полінуклеотидного праймера, викладеного в SEQ ID NO: 8 – SEQ ID NO: 14 у поєднанні з другим полінуклеотидним праймером, викладеним в SEQ ID NO: 33 – SEQ ID NO: 41. 3’ фланкуючі та з'єднуючі послідовності підтвердили із застосуванням першого полінуклеотидного праймера, викладеного в SEQ ID NO: 69 – SEQ ID NO: 72 у поєднанні з другим полінуклеотидним праймером, викладеним в SEQ ID NO: 27 – SEQ ID NO: 32. Фахівець у даній галузі зрозуміє, що можна використовувати інші праймерні послідовності для підтвердження фланкуючих і з'єднуючих послідовностей. [00121] Як виявили, вставка події 5307 фланкована на правій границі (5’ фланкуюча послідовність) геномною послідовністю кукурудзи, що показана в SEQ ID NO: 5, і фланкована на лівій границі (3’ фланкуюча послідовність) геномною послідовністю кукурудзи, що показана в SEQ ID NO: 6. 5’ з'єднуюча послідовність викладена в SEQ ID NO: 1. 3’ з'єднуюча послідовність викладена в SEQ ID NO: 2. Сайт інтеграції вставки вектора pSYN12274 включений в SEQ ID NO: 103 або його зворотний комплементарний ланцюг SEQ ID NO: 110, залежно від орієнтації використовуваної нуклеїнової кислоти. Приклад 6. Виявлення білка події 5307 за допомогою твердофазного імуноферментного аналізу. [00122] Для характеристики діапазону експресії білків FR8a (діючий інсектицидний початок) і фосфоманозоізомерази (PMI) (селектовний маркер) у рослин події 5307 визначили концентрації білка FR8a і PMI за допомогою твердофазного імуноферментного аналізу в деяких тканинах рослин. Гібриди були гемізиготнимиза трансгенами у події 5307, тоді як інцухтована рослина була гомозиготною за трансгенами. [00123] Цілі рослини та окремі частини (за винятком пилку) подрібнили в дрібний порошок за допомогою обробки з використанням будь-якого з кавомолки, блендера, дробарки Grindomix™ 19 UA 110320 C2 5 10 15 20 25 30 35 40 45 50 55 60 (Brinkmann Instruments; Уестбурі, Нью Йорк, США), ступки з товкачиком або млина, або комбінації цих пристроїв. Усю обробку виконали в присутності або сухого льоду, або рідкого азоту. Зразки добре перемішали для забезпечення гомогенності. Цілий зразок тканини рослини або репрезентативну субпробу зберегли для аналізу, забезпечуючи достатній розмір зразка для архівного сховища резервних зразків тканин рослин. Визначили процентну суху вага кожного зразка та оброблені зразки зберігали при приблизно -80 °C до ліофілізації. [00124] Екстрагували зразки свіжої тканини (за винятком пилку та силосу) і цілої рослини. Для кожного проаналізованого зразка 1,0 г аліквотну пробу подрібненого в порошок свіжого матеріалу зважили в 15-мл поліпропіленовій пробірці, суспендували в 3 мл буфера для екстракції [50 мM CAPS, 0,1 M NaCl, 2 мM EDTA, 1 мM дитіотреїтолу, 1 мM 4-(1аміноетил)бензолсульфонілфториду HCl, 1 мM лейпептину, pH 10] і екстрагували за допомогою гомогенізатора Autogizer® (Tomtek; Хамден, Коннектикут, США). Після центрифугування протягом 15 хвилин при 10000 x g при 4ºC супернатант використовували для аналізу FR8a і PMI із застосуванням твердофазного імуноферментного аналізу. Після обробки йодацетамідом, як описано Hill і Straka (1988), сумарний білок в екстрактах піддали кількісному аналізу із застосуванням реагенту для аналізу білків BCA™ (Pierce; Рокфорд, Іллінойс, США). [00125] Екстракти пилку одержали шляхом суспендування пилку 1:30 (процентне співвідношення ваги до об’єму) у буфері для екстракції. Через 30 хвилин на льоді суспензії пилку зруйнували за допомогою трикратного пропущення через французький прес при приблизно 15000 фунтів на квадратний дюйм із наступним центрифугуванням при 14000 x g протягом 5 хвилин при 4ºC. За допомогою твердофазного імуноферментного аналізу здійснили аналізи Cry3A055 і PMI на супернатантах, як описано нижче. Сумарний білок піддали кількісному аналізу, як описано вище. [00126] Екстракти силосу одержали шляхом суспендування силосу 1:25 (процентне співвідношення ваги до об’єму) в 2X буфері для екстракції. Через 30 хвилин на льоді суспензії силосу екстрагували за допомогою гомогенізатора Polytron® від Brinkmann (Brinkmann; Уестбурі, штат Нью Йорк, США). Після центрифугування протягом 15 хвилин при 10000 x g при 4ºC супернатант використовували для аналізу FR8a і PMI із застосуванням твердофазного імуноферментного аналізу. Сумарний білок піддали кількісному аналізу, як описано вище. Кількісний аналіз FR8a [00127] Екстракти, отримані, як описано вище, кількісно проаналізували на FR8a за допомогою твердофазного імуноферментного аналізу (Tijssen, 1985) із застосуванням очищеного імуноафінною хроматографією моноклонального антитіла до mCry3A і очищеного імуноафінною хроматографією поліклонального антитіла до Cry1Ab. Нижню межу кількісного аналізу твердофазного імуноферментного аналізу за методом «подвійного сандвіча» оцінювали за найнижчою концентрацією чистого еталонного білка, що лежить на лінійній частині стандартної кривої, максимальним об’ємом контрольного екстракту, який можна проаналізувати без фонових перешкод, і відповідною вагою зразка, який представлений аліквотною пробою. [00128] Рівні білка FR8a, що визначаються кількісно, виявили у всіх рослинних тканинах, отриманих від події 5307. У більшості випадків результати представлені як середні п'яти повторних зразків тканини. Рівні контрольних зразків були нижче межі кількісного аналізу для всіх тканин. [00129] У всіх стадіях росту середні рівні FR8a, обмірювані в листах, коріннях і пилку, варіювали в діапазоні приблизно 18-29 мкг/г сирої ваги (77-113 мкг/г сухої ваги), приблизно 1,84,1 мкг/г сирої ваги (22-41 мкг/г сухої ваги) і приблизно
ДивитисяДодаткова інформація
Автори російськоюDe Framond, Annick, Jeanne, Meghji, Moez, Rajabali, New, Stephen, Prairie, Anna, Underwood
МПК / Мітки
МПК: A01H 5/00, C12N 15/82
Код посилання
<a href="https://ua.patents.su/214-110320-podiya-5307-kukurudzi.html" target="_blank" rel="follow" title="База патентів України">Подія 5307 кукурудзи</a>
Спосіб визначення зиготності рослини кукурудзи, що містить об’єкт das-40278-9 aad-1 кукурудзи

Номер патенту: 109882
Опубліковано: 26.10.2015
Автори: Цуй Юнсінь Корі, Новак Стефен, Чжоу Нін, Грін Томас Уілльям
МПК: C12N 15/82, C12Q 1/68, A01H 1/04, C12N 15/10
Мітки: містить, кукурудзи, спосіб, das-40278-9, об`єкт, визначення, рослини, aad-1, зиготності
Формула / Реферат:
1. Спосіб визначення зиготності рослини кукурудзи, що містить об'єкт DAS-40278-9 AAD-1 кукурудзи, що містить SEQ ID NO:1, де вказаний об'єкт містить трансгенну конструкцію, що містить ген AAD-1, де вказана трансгенна конструкція фланкована 5¢-фланкуючою геномною ДНК кукурудзи і 3¢-фланкуючою геномною ДНК кукурудзи, де вказана 5'-фланкуюча геномна ДНК кукурудзи складається із залишків 1-1873 SEQ ID NO:1 і вказана 3'-фланкуюча...
Спосіб визначення схожості насіння кукурудзи

Номер патенту: 86727
Опубліковано: 10.01.2014
Автори: Черчель Владислав Юрійович, Кирпа Микола Якович, Стюрко Марина Олександрівна
МПК: A01C 1/02
Мітки: спосіб, насіння, схожості, визначення, кукурудзи
Формула / Реферат:
Спосіб визначення схожості насіння кукурудзи, що включає висівання насіння кукурудзи в ростильник з ґрунтом, витримування насіння кукурудзи в холодильній шафі, витримування насіння кукурудзи в термостаті до появи сходів, визначення схожості насіння, який відрізняється тим, що витримування насіння кукурудзи в холодильній шафі при температурі від +8 до +10 °C проводять протягом трьох діб, витримування в термостаті при температурі...
Спосіб індукції калусогенезу в культурі незрілих зародків кукурудзи

Номер патенту: 56325
Опубліковано: 10.01.2011
Автори: Піралов Григорій Рафаїлович, Сатарова Тетяна Миколаївна, Абраімова Ольга Євгеніївна, Деркач Катерина Вікторівна
МПК: A01H 4/00
Мітки: індукції, спосіб, калусогенезу, незрілих, кукурудзи, зародків, культури
Формула / Реферат:
Спосіб індукції калусогенезу в культурі незрілих зародків кукурудзи, що включає запилення донорних рослин кукурудзи, відбір та стерилізацію качанів, асептичне видалення, експлантацію та культивування незрілих зародків на індуктивному середовищі N6 з додаванням інозиту, L-проліну, сахарози, нітрату срібла, 2,4-дихлорфеноксіоцтової кислоти, який відрізняється тим, що до складу живильного середовища додають суміш амінокислот, до складу якої...
Спосіб адаптації рослин-регенерантів кукурудзи у ґрунті

Номер патенту: 73966
Опубліковано: 10.10.2012
Автори: Абраімова Ольга Євгеніївна, Сатарова Тетяна Миколаївна, Гармаш Світлана Миколаївна, Деркач Катерина Вікторівна
МПК: A01H 4/00
Мітки: ґрунті, кукурудзи, спосіб, адаптації, рослин-регенерантів
Формула / Реферат:
Спосіб адаптації рослин-регенерантів кукурудзи у ґрунті, що включає запилення донорних рослин кукурудзи, відбір і стерилізацію качанів, асептичне видалення та експлантацію незрілих зародків, індукцію калусогенезу, регенерацію, укорінення рослин-регенерантів, висаджування їх у ґрунт, який відрізняється тим, що висаджування рослин-регенерантів у ґрунт є безпосереднім і ґрунт зволожують розчином біогумату, що є екстрактом з біогумусу,...
Спосіб експлантації незрілих зародків кукурудзи in vitro

Номер патенту: 58347
Опубліковано: 11.04.2011
Автори: Сатарова Тетяна Миколаївна, Деркач Катерина Вікторівна, Піралов Григорій Рафаїлович, Абраімова Ольга Євгеніївна
МПК: A01H 4/00
Мітки: кукурудзи, спосіб, експлантації, зародків, незрілих, vitro
Формула / Реферат:
Спосіб експлантації незрілих зародків кукурудзи in vitro, що включає зривання качана у відповідній стадії розвитку з донорської рослини, зняття обгорток качана, відокремлення зернівок від стрижня качана, стерилізацію зернівок, вилучення зародків із зернівок, перенесення зародків на живильне середовище, який відрізняється тим, що зернівки стерилізують і зародки вилучають із зернівок, коли зернівки знаходяться на стрижні качана.
Попередній патент: Зрівняльний вентиль
Наступний патент: Водна резорцин-формальдегід-латексна дисперсія, її застосування, спосіб її одержання, волокна з поліпшеною адгезивністю, спосіб їх одержання та їх застосування
Випадковий патент: Автомобільне бензинове паливо для двигунів внутрішнього згоряння