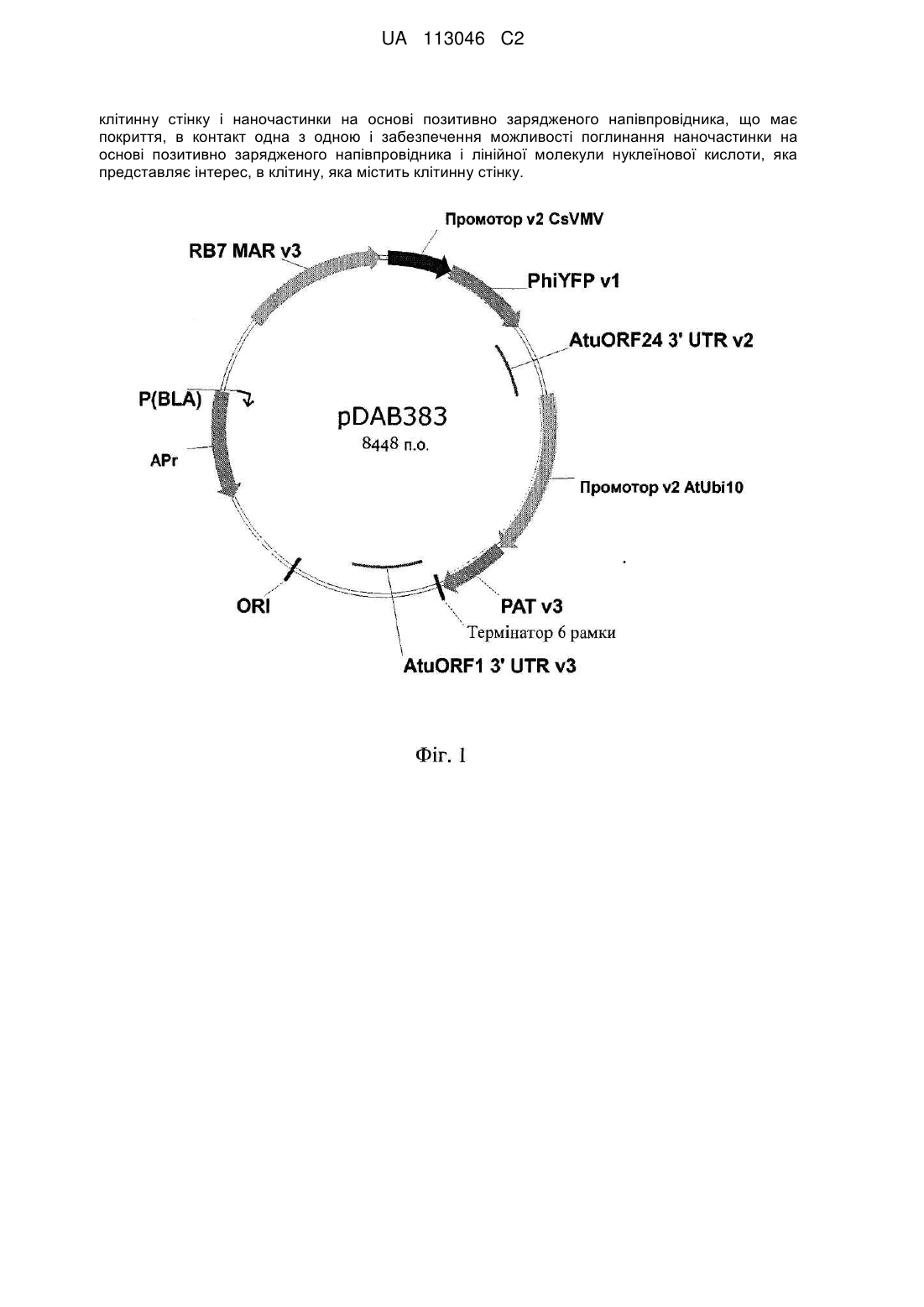
Спосіб введення лінійної молекули нуклеїнової кислоти в клітину рослини, яка має клітинну стінку, із застосуванням пегильованих напівпровідникових наночастинок
Номер патенту: 113046
Опубліковано: 12.12.2016
Автори: Семьюел Джаякумар Пон, Йо Керм Й., Бероуз Френк Дж., Самбоджу Нарасімха Чарі, Веб Стивен Р.







Формула / Реферат
1. Спосіб введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину рослини, яка має клітинну стінку, за яким:
надають клітину рослини, яка має клітинну стінку;
покривають наночастинку на основі позитивно зарядженого напівпровідника поліетиленгліколем;
покривають наночастинку на основі позитивно зарядженого напівпровідника лінійною молекулою нуклеїнової кислоти, яка представляє інтерес;
приводять клітину, яка має клітинну стінку, і наночастинку на основі позитивно зарядженого напівпровідника, що має покриття, в контакт одна з одною; і
надають можливість поглинання наночастинки на основі позитивно зарядженого напівпровідника і лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину, яка містить клітинну стінку.
2. Спосіб за п. 1, за яким додатково надають можливість поглинання наночастинки на основі позитивно зарядженого напівпровідника в компартмент клітини рослини, яка містить клітинну стінку.
3. Спосіб за п. 2, за яким додатково покривають наночастинку на основі позитивно зарядженого напівпровідника білком, який націлює на субклітинний компартмент.
4. Спосіб за п. 3, де компартмент вибраний із групи, яка складається з цитозолю, ядра, тонопластів, пластиди, етіопласту, хромопласту, лейкопласту, елайопласту, протеїнопласту, амілопласту, хлоропласту і просвіту подвійної мембрани.
5. Спосіб за п. 1, де клітина рослини, яка має клітинну стінку, являє собою клітину рослини з комерційних видів зернових рослин.
6. Спосіб за п. 5, де клітина рослини, яка має клітинну стінку, вибрана з групи, яка складається з клітин тютюну, моркви, маїсу, каноли, ріпаку, бавовнику, пальми, арахісу, сої, Oryza sp., Arabidopsis sp., Ricinus sp. і цукрової тростини.
7. Спосіб за п. 6, де клітина рослини походить із тканини, вибраної з групи, яка складається із зародка, меристеми, калусу, пилка, листя, пиляків, коріння, кінчиків коріння, квіток, насіння, стручків і стебел.
8. Спосіб за п. 1, де клітина рослини, яка має клітинну стінку, являє собою культивовану клітину.
9. Спосіб за п. 1, де напівпровідникова наночастинка на основі позитивно зарядженого напівпровідника являє собою квантову точку.
10. Спосіб за п. 1, за яким додатково проводять дериватизацію поверхні наночастинки на основі позитивно зарядженого напівпровідника.
11. Спосіб за п. 1, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, містить послідовність нуклеїнової кислоти, вибрану з групи, яка складається з молекул ДНК, РНК, RNAi і генів.
12. Спосіб за п. 11, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, містить ген.
13. Спосіб за п. 12, де ген являє собою ген чужорідного білка, агрономічний ген або маркерний ген.
14. Спосіб за п. 1, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, отримана шляхом розщеплення молекули нуклеїнової кислоти нуклеазою.
15. Спосіб за п. 14, де молекула нуклеїнової кислоти, розщеплена нуклеазою, вибрана з групи, яка складається з плазміди, косміди, штучних хромосом, штучних хромосом дріжджів і штучних хромосом бактерій.
16. Спосіб за п. 11, за яким додатково відбирають клітини, у які стабільно інтегрована молекула, яка представляє інтерес.
17. Спосіб за п. 16, де відібрані клітини є регенерованими клітинами.
18. Спосіб за п. 17, за яким додатково проводять регенерацію рослини з регенерованих клітин.
19. Спосіб введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в рослинний матеріал, за яким:
надають рослинний матеріал, де рослинний матеріал вибраний із групи, яка включає клітини рослини, тканини рослини і рослини;
покривають наночастинку на основі позитивно зарядженого напівпровідника поліетиленгліколем;
покривають наночастинку на основі позитивно зарядженого напівпровідника лінійною молекулою нуклеїнової кислоти, яка представляє інтерес;
приводять клітину рослини, яка має клітинну стінку, і наночастинки, на основі позитивно зарядженого напівпровідника, що має покриття, в контакт одна з одною; і
надають можливість поглинання наночастинки і лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в рослинний матеріал.
20. Спосіб за п. 19, де рослинний матеріал являє собою тканину рослини, вибрану з групи, яка складається з зародка, тканини меристеми, калусу, пилка, листя, пиляків, коріння, кінчиків коріння, квіток, насіння, стручків і стебел.
Текст
Реферат: Винахід належить до способу введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину рослини, яка має клітинну стінку, який включає надання клітини рослини, яка має клітинну стінку, покриття наночастинки на основі позитивно зарядженого напівпровідника поліетиленгліколем, покриття наночастинки на основі позитивно зарядженого напівпровідника лінійною молекулою нуклеїнової кислоти, яка представляє інтерес, приведення клітини, яка має UA 113046 C2 (12) UA 113046 C2 клітинну стінку і наночастинки на основі позитивно зарядженого напівпровідника, що має покриття, в контакт одна з одною і забезпечення можливості поглинання наночастинки на основі позитивно зарядженого напівпровідника і лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину, яка містить клітинну стінку. UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Заява про пріоритет Дана заявка заявляє пріоритет за датою подачі попередньої заявки на патент США "Доставляння лінійної молекули днк у рослини для стабільної трансформації за допомогою пегильованих квантових точок" з порядковим номером 61/362224, поданої 7 липня 2010 року. Рівень техніки Наночастинки мають унікальні властивості, які були використані для доставляння ДНК у конкретні тваринні клітини. Наночастинки мають унікальні властивості, які використовувалися для доставляння ДНК у конкретні тваринні клітини. Було знайдено, що коли певні наночастинки, покриті ДНК, інкубуються з клітинами, які не мають клітинної стінки, клітини поглинають наночастинки і починають експресувати гени, кодовані на ДНК. Напівпровідникові наночастинки (наприклад, квантові точки (QD)) у діапазоні розмірів 3-5 нм також були використані як носії для доставляння молекул у клітини. ДНК і білки можуть бути зв'язані з деякими лігандами, прикріпленими до поверхні QD. Див., наприклад, Patolsky, F. et al., J. Am. Chem. Soc. 125:13918 (2003). Покриті карбоновою кислотою або аміном QD можуть бути зшиті з молекулами, які містять тіолову групу, див., наприклад, Dubertret B. et al., Science 298:1759 (2002); Akerman et al., Proc. Natl. Acad. Sci. U.S.A. 99:12617 (2002); Mitchell et al., J. Am. Chem. Soc. 121:8122 (1999)), або N-гідроксисукцинімільну (NHS) ефірну групу, з використанням стандартних протоколів біокон'югації. Див., наприклад, Pinaud, et al., Am. Chem. Soc. 126:6115 (2004); Bruchez, et al., Science 281:2013 (1998). Альтернативним шляхом для приєднання молекул до поверхні QD є спосіб кон'югації покритих стрептавідином QD з біотинільованими білками, олігонуклеотидами або антитілами. Див., наприклад, Dahan, et al., Science 302:442 (2003); Pinaud, et al., J. Am. Chem. Soc. 126:6115 (2004); Wu, et al., Nature Biotechnol. 21:41 (2003); Jaiswal, et al., Nature Biotechnol. 21:47 (2003); і Mansson, et al., Biochem. Biophys. Res. Commun. 314:529 (2004). Доставляння чужорідних молекул нуклеїнових кислот у рослини є складною задачею через наявність стінок у рослинних клітин. Використовувані способи для генетичної трансформації рослин основані на інвазивному доставлянні. Клітинна стінка в клітинах рослин є бар'єром для доставляння екзогенно внесених молекул. Для досягнення доставляння генів і малих молекул у клітини рослин із клітинною стінкою були використані численні інвазивні способи, такі як біолістичне доставляння (генна гармата), мікроін'єкція, електропорація й опосередкована Agrobacterium трансформація, але доставляння білків було досягнуто тільки з використанням мікроін'єкції. Коли бажане доставляння молекул нуклеїнових кислот у клітини рослин, клітинну стінку видаляють перед додаванням частинок до протопластів рослини (див. Torney, et al., Nature Nanotechnol. 2:295-300 (2007). Опис винаходу Тут описані способи і композиції для використання наночастинок і лінеаризованих молекул нуклеїнових кислот для введення молекули, яка представляє інтерес, у рослинну клітину, яка має клітинну стінку. Деякі втілення розкритих способів можуть використовуватися для продукування стабільно трансформованих генетично модифікованих фертильних рослин. У деяких втіленнях характерні властивості лінійних молекул нуклеїнових кислот забезпечують доставляння специфічних, які представляють інтерес, генних послідовностей без послідовностей сторонніх нуклеїнових кислот, які можуть мати регуляторні функції для цільового трансгенного організму. У деяких втіленнях наночастинки можуть включати молекули пегильованої лінійної нуклеїнової кислоти. У конкретних втіленнях наночастинки можуть бути напівпровідниковими наночастинками, такими як квантові точки (QD). У деяких втіленнях молекулою лінійної нуклеїнової кислоти може бути лінеаризована плазмідна ДНК. В інших втіленнях молекули лінійної нуклеїнової кислоти можуть включати кодуючі послідовності фосфінотрицин-Nацетилтрансферази (PAT) і/або жовтого флуоресцентного білка (YFP). Також розкриті способи введення молекули, яка представляє інтерес, у клітину рослини, яка має клітинну стінку, де способи можуть включати надавання клітини рослини, яка має клітинну стінку; покриття поверхні наночастинки ПЕГ для отримання ПЕГильованих наночастинок; покриття ПЕГильованих наночастинок щонайменше однією молекулою лінійної нуклеїнової кислоти, яка представляє інтерес; приведення в контакт одна з одною клітини рослини, яка має клітинну стінку, з ПЕГильованими наночастинками, покритими молекулою(ами) лінійної нуклеїнової кислоти, яка представляє інтерес; і надавання можливості поглинання наночастинки і молекули(молекул) лінійної нуклеїнової кислоти, яка представляє інтерес, у клітину, яка має клітинну стінку. Додатково розкриті способи інтогресії генетичної ознаки в рослину. У деяких втіленнях спосіб може включати надавання рослинної клітини; покриття поверхні наночастинок ПЕГ для продукування ПЕГильованих наночастинок; покриття ПЕГильованих наночастинок засобами 1 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 експресії генетичної ознаки в рослині; приведення в контакт одна з одною клітини рослини з ПЕГильованими наночастинками, покритими засобами експресії генетичної ознаки в рослині; і забезпечення поглинання клітиною рослини наночастинки з засобами експресії генетичної ознаки в рослині для отримання трансформованої клітини рослини; регенерацію цілої рослини з трансформованої рослинної клітини; і репродукцію рослини. У деяких втіленнях генетичну ознаку, яка може бути інтрогресована відповідно до способів винаходу, включають ознаку, вибрану без обмеження з: чоловічої фертильності; стійкості до гербіцидів; стійкості до комах; і стійкості до бактеріального, грибкового захворювання і/або вірусного захворювання. Також розкриті способи за винаходом можуть бути використані для трансформації рослин in planta. У деяких втіленнях рослина може бути вибрана з рослин роду Arabidopsis, наприклад, A. thaliana. У конкретному втіленні, рослина, трансформована in planta, може бути вибрана з екотипу Колумбія рослин A. thaliana. Додатково розкритими є генетично модифіковані (ГМ) рослинні клітини і способи їхнього отримання, де клітини рослин мають одну або декілька нуклеїнових кислот, введених у них за допомогою способів даного винаходу. У деяких втіленнях плазміда, яка містить щонайменше один ген, який представляє інтерес, і селектований маркер, може бути введена в клітину рослини, яка має клітинну стінку, за допомогою наночастинки відповідно до даного винаходу. У додаткових варіантах втіленнях можуть бути відібрані стабільні трансформанти, які мають стабільно інтегрований щонайменше один ген, який представляє інтерес, і/або селектований маркер. В альтернативних втіленнях клітина рослини, яка вже містить ген, який представляє інтерес, може бути розмножена для отримання інших клітин, які містять молекулу, яка представляє інтерес. В інших варіантах втіленнях клітина рослин, яка вже містить молекулу, яка представляє інтерес, може бути регенерованою клітиною, яка може бути використана для регенерації цілої рослини, яка включає молекулу, яка представляє інтерес. Додатково розкритими є способи отримання регенерованих клітин рослин, які містять молекулу, яка представляє інтерес, для застосування в тканинних культурах. Ця тканинна культура може бути здатною регенерувати рослини, які мають по суті той же самий генотип, що і регенеровані клітини. Регенеровані клітини в таких тканинних культурах можуть бути зародками, протопластами, меристематичними клітинами, калусом, пилком, листям, пилком, корінням, кінчиками коріння, квітами, насінням, стручками або стеблами. Крім того, деякі втілення забезпечують рослини, регенеровані з тканинних культур цього винаходу. Додатково розкритими є способи генерування стабілізованих ліній рослин, які містять бажану генетичну ознаку або молекулу нуклеїнової кислоти, яка представляє інтерес, де бажана генетична ознака або молекула нуклеїнової кислоти, що представляє інтерес, можуть спочатку вводитися за рахунок поглинання наночастинки через клітинну стінку рослини. Способи генерування стабілізованих ліній рослин добре відомі фахівцю зі звичайною кваліфікацією в даній галузі і можуть включати, без обмеження, такі способи, як самозапилення, зворотне схрещування, отримання гібридів, схрещування з популяціями і т. п. Таким чином, також розкриті рослини і клітини рослин, які містять бажану генетичну ознаку або молекулу нуклеїнової кислоти, яка представляє інтерес, вперше введені в клітину рослини (або в її попередники) поглинанням наночастинок через клітинну стінку. Клітини рослини, які містять бажану генетичну ознаку або молекулу нуклеїнової кислоти, яка представляє інтерес, вперше введені в рослину або в клітину (або в її попередники) поглинанням наночастинок через клітинну стінку, можуть бути використані в схрещуваннях з іншими клітинами рослин, які відрізняються, для отримання гібридних клітин першої генерації (F 1), насіння і/або рослин з бажаними характеристиками. Крім ілюстративних аспектів і варіантів втілень, описаних вище, додаткові аспекти і втілення стануть очевидними у світлі наступних описів. Короткий опис фігур Фіг. 1 включає графічний опис нелінеаризованої плазміди pDAB3831. Фіг. 2 включає графічний опис плазміди pDAB3831, лінеаризованої за допомогою ферменту рестрикції KpnІ. Фіг. 3 включає вирівнювання послідовностей між послідовністю ДНК фосфінотрицин-Nацетилтрансферази (PAT) із наночастинок з лінійною ДНК трансформованого генома Arabidopsis і послідовністю PAT з бази даних NCBI. Список послідовностей SEQ ID NO: 1 показує послідовність прямого праймера, використовуваного для ампліфікації гена YFP: TGTTCCACGGCAAGATCCCCTACG. 2 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 SEQ ID NO:2 показує послідовність зворотного праймера, використовуваного для ампліфікації гена YFP: TATTCATCTGGGTGTGATCGGCCA. SEQ ID NO:3 показує послідовність прямого праймера, використовуваного для ампліфікації гена PAT: GGAGAGGAGACCAGTTGAGATTAG. SEQ ID NO:4 показує послідовність зворотного праймера, використовуваного для ампліфікації гена PAT: AGATCTGGGTAACTGGCCTAACTG. Найкращі варіанти здійснення винаходу I. Огляд деяких втілень Способи за винаходом, які забезпечують неінвазивну передачу генів можуть бути дуже корисними для створення генетично модифікованих рослин з бажаними генетичними ознаками. Неінвазивна передача генів може сприяти специфічному націлюванню і коректуванню молекулярних сайтів всередині клітин для застосування в таких сферах, як ведення бажаних показників, досягнення підсумкового результату й отримання бажаних агрономічних ознак культур рослин. Описані способи, як альтернатива для ГМО, також можуть бути корисні для транзієнтної трансформації рослин, для розширення технології інтрогресії генетичних ознак і додавання деревам або овочевим культурам стійкості до патологій, де технологія в даний час обмежена. Нещодавня заявка на патент США No. 60/978059 розкриває неінвазивні засоби доставляння ДНК, основані на наночастинках, використовуючи різні, які несуть корисне навантаження наночастинки, у тому числі для доставляння кільцевої плазмідної ДНК, і зрозуміло демонструє стабільну інтеграцію трансгенів у насіння T 1 рослин Arabidopsis. Отримані так трансгенні рослини, які містять кільцеву плазмідну ДНК, демонструють бажані толерантні до гербіцидів фенотипи, і показують високий рівень толерантності при одночасному обприскуванням у полі щонайменше чотириразовою кількістю глюфосинату амонію. Заявка США No. 60/978059 показала, серед іншого, генетичну трансформацію в Arabidopsis за рахунок застосування позитивно заряджених наночастинок золота з кільцевою плазмідною ДНК. Дана робота описує, зокрема, застосування лінійних молекул нуклеїнових кислот для стабільної генетичної трансформації рослин. Заявка США No.60/978059 описує, зокрема, доставляння плазмідної ДНК, опосередковану позитивно зарядженими наночастинками. Однак дотепер не повідомлялося про досягнення стабільної геномної інтеграції трансгена при використанні для доставляння лінійних плазмід. Цей винахід описує застосування лінійних молекул нуклеїнової кислоти з використанням позитивно заряджених наночастинок для стабільної генетичної трансформації рослин. Молекулярний аналіз показується експресію PAT разом з YFP у трансгенних рослинах Arabidopsis T1, трансформованих геном pat і геном yfp способами за винаходом. Tрансгенні рослини T1 є фертильними і продукують насіння. Це насіння може використовуватися для розмноження, і поряд з молекулярним і білковим аналізами може бути виконаний сегрегаційний аналіз. Молекули лінійних нуклеїнових кислот мають характерні властивості, які відрізняють їх від кільцевої плазмідної ДНК. Наприклад, молекули лінійних нуклеїнових кислот можуть мати чітко визначену генну касету без кістяка несучого вектора і без селектованого маркера бактеріального антибіотика. Гербіцид глюфосинат амонію (GLA), може бути розпилений у полі з рівнем концентрації для скринінгу трансгенів. Саджанці Arabidopsis T 1, отримані з використанням способів за винаходом, показали стійкість до гербіцидів при п'ятикратному рівні дозування глюфосинату в полі, наприклад, у режимі "через день", починаючи із семи днів після появи сходів. Геномні ДНК цих трансгенних рослин були проаналізовані на предмет присутності pat і yfp методом ПЛР, і результати показали присутність цільових послідовностей pat і yfp у ДНК. Секвенування отриманих продуктів ПЛР показало наявність правильних трансгенних послідовностей pat і yfp у Arabidopsis T1, отриманих з використанням способів за винаходом. II. Терміни В описі і таблицях, які йдуть далі, використовується ряд термінів. Для того, щоб забезпечити зрозуміле й однакове розуміння опису і формули винаходу, у тому числі обсягу домагань, який повинен підтримуватися таким термінам, представлені наступні визначення. Зворотне схрещування. Як використовується тут, термін "зворотне схрещування" може бути процесом, у якому селекціонер багаторазово схрещує гібридне потомство з одним з батьків, наприклад, гібрид першої генерації F1 з одним з батьківських генотипів цього гібрида F1. 3 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Зародок. Як використовується тут, термін "зародок" може стосуватися маленької рослини, яка міститься в зрілому насінні. Наночастинка. Як використовується тут, термін "наночастинка" може стосуватися мікроскопічної частинки, яка має щонайменше один нанометровий розмір, наприклад, менше 100 нм. Наночастинки, які підходять для застосування в даному винаході, можуть мати розмір 1 нм-0,84 мкм. Одним із класів наночастинок є "квантова точка" (QD). Квантова точка може мати медіанний діаметр 1 нм-10 нм, переважно 2-4 нм. Інші варіанти наночастинок включають, без обмеження: наночастинки золота; покриті золотом наночастинки; пористі наночастинки; мезопористі наночастинки; наночастинки діоксиду кремнію; полімерні наночастинки, включаючи дендримери; вольфрамові наночастинки; желатинові наночастинки, нанооболонки; наноядра; наносфери; нанострижні; магнітні наночастинки і їхні комбінації. Серед доступних наночастинок, люмінесцентні напівпровідникові нанокристали (QD) забезпечують багато відомих застосувань при біологічній візуалізації і зондуванні. Їхня корисність є наслідком комбінації унікальних Фото-фізичних характеристик і розмірів, порівняним з розміром великого білка. Гідродинамічний радіус гідрофільних QD з CdSe-ZnS варіює від 5 нм (для нанокристалів, кепіруючих молекулярні ліганди) до 20 нм для нанокристалів, інкапсульованих у блок-співполімери. Один QD може бути кон'югований з декількома біомолекулами (наприклад, з антитілами, пептидами і молекулами нуклеїнових кислот), з отриманням багатофункціональних біокон'югатів QD з розширеною авідністю. Крім того, їхня сильна стійкість до хімічної і фотодеградації може в цілому забезпечити довгостроковий флуоресцентний моніторинг специфічних біологічних процесів. Див. Nie і Emory, Science 275: 1 102-106 (1997). Множина схем нековалентних кон'югацій, основаних на самозбиранні через афінність металів і біотин-авідинового зв'язування, можуть одночасно використовуватися в рамках того самого комплексу, без необхідності додаткового очищення, приводячи до отримання багатофункціональних біокон'югатів QD, які є стабільними навіть у внутрішньоклітинних середовищах. Див. Yezhelyev et al., J. Am. Chem. Soc. 130(28):9006-12 (2008). Використовуючи в середньому по 10 YFP плюс номінальна кількість, яка дорівнює 50, які проникають у клітину пептидів (CPP) на одну QD, може бути досягнуте внутрішньоклітинне доставляння білкового навантаження з молекулярною вагою щонайменше 300 кДа і з просторовим розміром 150 ангстрем. Див. там же. Навантаження, яке доставляється, для QD-bPE кон'югатів мають набагато більший діапазон розмірів і молекулярну вагу; наприклад, доставлені зборки, з розрахунку в середньому 2,5 стрептавідин-b-PE на кон'югат, мають молекулярну вагу, яка в цілому перевищує 103 кДа і просторові розміри наближаються до 500 ангстремів. Молекулярна вага і розмір може бути значно збільшений, якщо використовуються кон'югати з більш високими b-PE валентностями. Молекула нуклеїнової кислоти. Полімерні форми нуклеотидів, які можуть включати як смислові, так і антисмислові ланцюги РНК, кДНК, геномної ДНК, штучних хромосом (ACE), і синтетичні форми і суміші вищевказаних полімерів. Нуклеотид стосується рибонуклеотиду, дезоксирибонуклеотиду або модифікованої форми обох типів нуклеотидів. Термін "молекула нуклеїнової кислоти", як використовується тут, є синонімом термінів "нуклеїнова кислота" і "полінуклеотид". Молекула нуклеїнової кислоти звичайно має щонайменше 10 основ по довжині, якщо не зазначено інше. Цей термін включає одноланцюжкові і дволанцюжкові форми ДНК. Молекула нуклеїнової кислоти може включати як окремо, так і разом, природні і модифіковані нуклеотиди, які зв'язані між собою природними і/або які не зустрічаються в природі нуклеотидними зв'язками. Функціонально зв'язаний. Перша послідовність нуклеїнової кислоти функціонально зв'язана з другою послідовністю нуклеїнової кислоти, коли перша послідовність нуклеїнової кислоти знаходиться у функціональному взаємозв'язку з другою послідовністю нуклеїнової кислоти. Наприклад, промотор є функціонально зв'язаним з кодуючою послідовністю, якщо промотор впливає на транскрипцію або експресію кодуючої послідовності. При рекомбінантному продукуванні, послідовності функціонально зв'язаних нуклеїнових кислот можуть бути суміжними, і, при необхідності, можуть з'єднувати дві кодуючи області білка у тій же рамці зчитування. Однак для того, щоб бути функціонально зв'язаними, нуклеїнові кислоти можуть і не бути суміжними. Пегильований. Як використовується в даному документі, термін "Пегильований" може стосуватися наночастинок (наприклад, квантових точок), коли поверхні наночастинок були змінені за допомогою поліетиленгліколю (PEG, ПЕГ) для поліпшення біосумісності. Пегильовані наночастинки можуть бути додатково покриті різними націлювальними лігандами, наприклад, пептидами й антитілами, для поліпшення ефективності доставляння до конкретних клітин і тканин. ПЕГ був кон'югований з наночастинками і з різними лікарськими речовинами, 4 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 ліпосомами і полімерними міцелами, для того, щоб, наприклад, продовжити час циркуляції наночастинок з покриттям у крові, шляхом зниження неспецифічної адсорбції білків за рахунок ефекту просторової стабілізації. Квантова точка. Як використовується в даному документі, термін "квантова точка" (QD), також іноді раніше відомі як нанокристали, може стосуватися напівпровідникової наноструктури, яка обмежує рух електронів зони провідності і дірок валентної зони, або екситонів (зв'язаних пар електронів зони провідності і дірок валентної зони) у всіх трьох просторових напрямках. Це обмеження може бути зумовлене, наприклад, електростатичними потенціалами (генерованими зовнішніми електродами, за рахунок легування, деформацією, домішками і т. п.), присутністю межі розподілу між різними напівпровідниковими матеріалами (наприклад, у нанокристалічних системах ядро-оболонка), присутністю напівпровідникової поверхні (наприклад, у напівпровідникового нанокристала) або їх комбінаціями. Квантова точка може мати дискретний квантований енергетичний спектр. Відповідні хвильові функції можуть бути просторово локалізованими в квантовій точці, але тягнуться на багато періодів кристалічної решітки. Квантова точка містить мале кінцеве число (наприклад, близько 1-100) електронів зони провідності, дірок зони валентності або екситонів (тобто має кінцеве число елементарних електричних зарядів). Квантові точки є особливим класом напівпровідникових матеріалів, які можуть бути кристалами, які складаються з матеріалів елементів періодичних груп II-VI, III-V або IV-VI. Їхні розміри можуть варіюватися, наприклад, від 2 до 10 нанометрів (10-50 атомів) у діаметрі. У деяких втіленнях квантові точки можуть бути виготовлені у вигляді ядра й оболонки із селеніду кадмію і сульфіду цинку (CdSe/ZnS), і вони мають ряд корисних електричних і оптичних властивостей, які відрізняються за своїм характером від характеристик вихідного об'ємного матеріалу. Наночастинки у вигляді квантових точок були досліджені як засіб для візуалізації in vivo і in vitro, через високий квантовий вихід, високого коефіцієнта молярного поглинання і високої стійкості до Фотознебарвлення. Стійкий до гліфосату. Стійкість до дози гліфосату стосується здатності рослини виживати (тобто рослина не може загинути) під дією даної дози гліфосату. У деяких випадках толерантні рослини можуть тимчасово пожовкнути або іншим способом виявити яке-небудь ушкодження, індуковане гліфосатом (наприклад, надлишкове пагоноутворення і/або уповільнення росту), але потім вони відновлюються. Стабілізований. Як використовується в даному документі, термін "стабілізований" може стосуватися характеристик рослини, які репродуцибельно передаються від однієї генерації до наступної генерації інбредних рослин одного сорту. Трансген. Як використовується в даному документі, термін "трансген" може стосуватися послідовності екзогенної нуклеїнової кислоти. Як один з прикладів, трансген являє собою послідовність гена (наприклад, гена стійкості до гербіциду), ген, який кодує промислово або фармацевтично корисна сполука, або ген, який кодує бажану сільськогосподарську ознаку. У ще одному прикладі трансген являє собою послідовність антисмислової нуклеїнової кислоти, де експресія послідовності антисмислової нуклеїнової кислоти інгібує експресію послідовності цільової нуклеїнової кислоти. Трансген може містити регуляторні послідовності, функціонально зв'язані з трансгеном (наприклад, промотор). У деяких втіленнях молекула нуклеїнової кислоти, яка представляє інтерес, яка повинна бути введена трансформацією за допомогою наночастинок, являє собою трансген. Однак в інших втіленнях, молекула нуклеїнової кислоти, яка представляє інтерес являє собою послідовність ендогенної нуклеїнової кислоти, для якої бажані додаткові геномні копії послідовності ендогенної нуклеїнової кислоти, або молекулою нуклеїнової кислоти, яка знаходиться в антисмисловій орієнтації стосовно цільової молекули нуклеїнової кислоти в організмі хазяїна. Поглинання. Як використовується в даному документі, термін "поглинання" може стосуватися транслокації частинки, такої як наночастинка (наприклад, квантової точки), через клітинну стінку або клітинну мембрану, де транслокація має місце не тільки як результат імпульсу, який додається цій частинці чимось іншим, ніж клітина, у яку ця частинка поглинається. Необмежувальні приклади пристроїв або способів, які викликають транслокацію частинки через клітинну стінку або клітинну мембрану тільки в результаті імпульсу, який додається частинці, являють собою біолістику, генну гармату, мікроін'єкцію і/або введення методом проколу за допомогою наноматеріалів (імпалефекція). III. Доставляння молекул ДНК за допомогою наночастинок для стабільної трансформації клітин рослин А. Огляд 5 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Цей винахід описує, зокрема, нові способи трансформації рослин, використовуючи перенесення за допомогою наночастинок лінеаризованої плазмідної ДНК для генетичної трансформації і розвитку стабільних трансгенних рослин. Способи стосовно деяких втілень даного винаходу можуть надати не тільки швидку генерацію трансгенного організму, але також і додаткові можливості для бажаних геномних модифікацій, у порівнянні з іншими способами трансформації. Втілення винаходу привели до стабільно трансформованої рослини, про яку повідомляється вперше, отриману за допомогою доставляння лінеаризованої плазмідної ДНК, опосередкованої наночастинками. Розкриті способи генетичної модифікації являють собою відмовлення від традиційних способів генетичної трансформації рослин, і можуть бути дуже корисними для генерації трансгенних культур рослин. B. Молекули ДНК З появою молекулярних біологічних методик, які забезпечили виділення і характеризацію генів, які кодують конкретний білок або РНК-продукти (наприклад, інтерферуючі РНК (RNAi), вчені в сфері біології рослин відчували сильний інтерес до створення генома клітин, які містять і експресують чужорідні гени, або додаткові або модифіковані варіанти нативних або ендогенних генів (можливо під керуванням різних промоторів), для того, щоб, наприклад, визначеним способом змінити генетичні ознаки клітини. Такі чужорідні додаткові і/або модифіковані гени згадані тут загальним терміном "трансгени." Трансгени можуть, наприклад, кодувати білок, який представляє інтерес, або бути транскрибовані в RNAi. За останні п'ятнадцять - двадцять років були розроблені декілька методів для продукування трансгенних клітин, і даний винахід у специфічних втіленнях стосується варіантів трансформованих клітин, і методів їхнього продукування за допомогою введення в клітини рослини, які мають клітинну стінку, одну або декілька лінійних молекул нуклеїнової кислоти за рахунок поглинання наночастинки через стінку клітини. У деяких втіленнях винаходу трансген може міститися в лінеаризованому векторі експресії. Трансформація клітини може включати конструювання вектора експресії, який буде функціонувати в конкретній клітині. Такий вектор може включати послідовність нуклеїнової кислоти, яка включає ген під контролем регулюючого елемента або оперативно зв'язаний з регулюючим елементом (наприклад, із промотором, енхансером, послідовністю термінації або їх комбінацією). Так, вектор експресії може містити одну або декілька таких функціонально зв'язаних комбінацій ген/регулюючий елемент. Вектор(и) може бути у формі плазміди, і може використовуватися окремо або в комбінації з іншими плазмідами, для того щоб забезпечити трансформацію клітини, використовуючи методи трансформації як описано тут, щоб включити трансген(и) у генетичний матеріал клітини рослини, яка містить клітинну стінку. В інших втіленнях, молекула нуклеїнової кислоти, яка представляє інтерес, може бути лінійною молекулою нуклеїнової кислоти. Лінійні молекули нуклеїнової кислоти можуть бути отримані, наприклад, розщепленням кільцевої плазміди ендонуклеазої рестрикції. Ендонуклеази рестрикції розщеплюють плазміду на одному або декількох сайтах розпізнавання в межах нуклеотидної послідовності плазміди. Таким чином, можуть бути створені плазміди, які забезпечують продукування однієї або декількох специфічних лінійних молекул нуклеїнової кислоти, за рахунок розщеплення специфічної ендонуклеазої рестрикції. Альтернативно, дана нуклеотидна послідовність плазміди може бути проаналізована з метою пошуку сайтів розпізнавання однієї або декількома специфічними ендонуклеазами рестрикції, які забезпечують продукування однієї або декількох специфічних лінійних молекул нуклеїнової кислоти (нуклеїнових кислот). Шляхом вибору сайтів рестрикції, які розщеплюються по специфічних положеннях у межах кільцевої плазміди або лінійної молекули нуклеїнової кислоти, можуть бути отримані лінійні молекули нуклеїнової кислоти, у яких відсутня одна або декілька послідовностей молекули нуклеїнової кислоти-попередника. Наприклад, може бути отримана лінійна молекула нуклеїнової кислоти в який відсутні чужорідні нуклеїнокислотні послідовності (наприклад, остов несучого вектора, маркери селекції, такі як бактеріальні маркери селекції і не є необхідними послідовності нуклеїнової кислоти, які є гомологічними геномної ДНК цільової клітини). Альтернативно, лінійна молекула нуклеїнової кислоти може бути синтезована так, щоб у ній були відсутні чужорідні нуклеїнокислотні послідовності. В інших втіленнях, де молекула, яка представляє інтерес, включає один або декілька генів, ген(и) може (можуть) бути домінантною або рецесивним алелем. Наприклад, ген(и) може (можуть) додати такі ознаки, як стійкість до гербіциду, стійкість до комах, стійкість до бактеріальних захворювань, стійкість до грибів, стійкість до вірусних захворювань, чоловіча фертильність, чоловіча стерильність, поліпшена харчова якість і промислове використання. Гени, які додають ці й інші ознаки, відомі в даній галузі техніки, і будь-який ген може бути введений у клітину, яка містить клітинну стінку, відповідно до способів винаходу. 6 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Експресуючі вектори для лінеаризації і поглинання за допомогою наночастинок: Маркерні гени Експресуючі вектори можуть необов'язково включати щонайменше один генетичний маркер, наприклад, функціонально зв'язаний з регуляторним елементом, який дозволяє трансформованим клітинам, які містять цей маркер, витягатися або негативним відбором (тобто інгібуванням росту клітин, які не містять цей ген селектованого маркера) або позитивним відбором (тобто скринінгом продукту, кодованого цим генетичним маркером). Багато генів селектованих маркерів для трансформації добре відомі в даній галузі техніки і включають, наприклад, без обмеження: гени, які кодують ферменти, які метаболічно детоксифікують селективний хімічний агент, який може бути антибіотиком або гербіцидом; або гени, які кодують змінену мішень, яка може бути нечутлива до інгібітору. Специфічні способи позитивного добору також відомі в даній галузі техніки. Один ген селектованого маркера, який може бути придатним для трансформації рослин деякими молекулами нуклеїнових кислот, являє собою ген неоміцинфосфотрансферази II (nptII), необов'язково під контролем регуляторних сигналів рослини, який додає стійкість до канаміцину. Див., наприклад, Fraley et al., Proc. Natl. Acad. Sci. U.S.A., 80:4803 (1983). Інший ген селектованого маркера, який може бути використаний, являє собою ген гігроміцинтрансферази, який додає стійкість до антибіотика гігроміцину. Див., наприклад, Vanden Elzen et al., Plant Mol. Biol., 5:299 (1985). Додаткові гени селектованих маркерів, які можуть бути використані в способах за винаходом, включають гени селектованих маркерів бактеріального походження, наприклад, ті, котрі додають стійкість до антибіотиків, таких як гентаміцинацетилтрансфераза, стрептоміцинфосфотрансфераза, аміноглікозид-3'-аденілтрансфераза і блеоміцин. Див. Hayford et al., Plant Physiol. 86:1216 (1988), Jones et al., Mol. Gen. Genet, 210:86 (1987), Svab et al., Plant Mol. Biol. 14:197 (1990), і Hille et al., Plant Mol. Biol. 7:171 (1986). Інші гени селектованих маркерів додають стійкість до гербіцидів, таких як гліфосат, глюфосинат або бромоксиніл. Див. Comai et al., Nature 317:741-744 (1985), Gordon-Kamm et al., Plant Cell 2:603-618 (1990) і Stalker et al., Science 242:419-423 (1988). Інші гени селектованих маркерів, які можуть бути використані для трансформації рослин, включають гени небактеріального походження. Ці гени включають, наприклад, без обмеження, дигідрофолатредуктазу миші, 5-енолпірувілшикімат-3-фосфатсинтазу рослин і ацетолактатсинтазу рослин. Див. Eichholtz et al., Somatic Cell Mol. Genet. 13:67 (1987), Shah et al., Science 233:478 (1986), і Charest et al., Plant Cell Rep. 8:643 (1990). Інший клас генів маркерів, які підходять для трансформації рослин, може знадобитися для скринінгу приблизно трансформованих клітин рослин, а не для прямого генетичного відбору трансформованих клітин на стійкість до токсичної речовини, такої як антибіотик. Ці гени зокрема застосовні для кількісного визначення або візуалізації просторової картини експресії гена в конкретних тканинах, і вони часто називаються "репортерними генами", оскільки вони можуть бути злиті з геном або регуляторною послідовністю гена для дослідження експресії гена. Гени, звичайно використовувані для скринінгу трансформованих клітин, включають, без обмеження, β-глюкуронідазу (GUS), β-галактозидазу, люциферазу і хлорамфеніколацетилтрансферазу. Див. Jefferson, R. A., Plant Mol. Biol. Rep. 5:387 (1987), Teeri et al., EMBO J. 8:343 (1989), Koncz et al., Proc. Natl. Acad. Sci. U.S.A. 84:131 (1987), і DeBlock et al., EMBO J. 3:1681 (1984). Зовсім недавно, гени, які кодують флуоресцентні білки (наприклад, GFP, EGFP, EBFP, ECFP і YFP), були використані як маркери при експресії генів у прокаріотичних і еукаріотичних клітинах. Див. Chalfie et al., Science 263:802 (1994). Таким чином, флуоресцентні білки і мутанти флуоресцентних білків можуть бути використані як маркери при скринінгу в деяких втіленнях винаходу. Експресуючі вектори для поглинання за допомогою наночастинки: Промотори Гени, включені в експресуючі вектори, можуть запускатися нуклеотидною послідовністю, яка містить регуляторний елемент, наприклад, промотор. Деякі типи промоторів добре відомі в галузі техніки трансформації, також як і інші регуляторні елементи, які можуть бути використані самостійно або в комбінації з промоторами. Промотор являє собою область ДНК, яка може знаходитися ліворуч (у положенні "апстрим") від старту транскрипції, і може бути включене впізнавання і зв'язування РНК полімерази і/або інших білків, для того, щоб ініціювати транскрипцію. "Промотор рослини" може бути промотором, який здатний ініціювати транскрипцію в клітинах рослин. Приклади промоторів під контролем розвитку включають промотори, які переважно ініціюють транскрипцію в конкретних тканинах, таких як листя, коріння, насіння, волокна, судини ксилеми, трахеїди або склеренхіма. Такі промотори називаються "тканинопереважними". Промотори, які ініціюють транскрипцію 7 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 тільки в конкретних тканинах, називають "тканиноспецифічними". Промотор, специфічний для "типу клітини", спочатку запускає експресію у визначених типах клітин в одному або декількох органах, наприклад, у клітинах судин коріння або листя. "Індуцибельний" промотор може являти собою промотор, який може бути під контролем навколишнього середовища. Приклади умов навколишнього середовища, які можуть впливати на транскрипцію під індуцибельними промоторами, включають, без обмеження, анаеробні умови або наявність світла. Тканиноспецифічні, тканинопереважні, специфічні для типу клітин і індуцибельні промотори утворюють клас "неконститутивних промоторів". "Конститутивний промотор" являє собою промотор, який може бути активним при більшості умов навколишнього середовища. 1. Індуцибельні промотори Індуцибельний промотор може бути функціонально зв'язаний з експресованим у клітині геном. Необов'язково, індуцибельний промотор може бути функціонально зв'язаний з нуклеотидною послідовністю, яка кодує сигнальну послідовність, яка може бути функціонально зв'язана з експресованим у клітині геном. У випадку індуцибельного промотору, швидкість транскрипції збільшується у відповідь на індукуючий агент. Будь-який індуцибельний промотор може бути використаний у втіленні даного винаходу. Див. Ward et al., Plant Mol. Biol. 22:361-366 (1993). Приклади індуцибельних промоторів включають, без обмеження, промотори системи ACEI, яка відповідає на мідь (Mett et al., Proc. Natl. Acad. Sci. U.S.A. 90:4567-4571 (1993)); промотор гена In2 маїсу, який відповідає на антидот до бензолсульфамідних гербіцидів (Hershey et al., Mol. Gen Genetics 227:229-237 (1991) і Gatz et al., Mol. Gen. Genetics 243:32-38 (1994)); і промотор репресора Tet з Tn1O (Gatz et al., Mol. Gen. Genetics 227:229-237 (1991)). Особливо корисним індуцибельним промотором може бути промотор, який відповідає на індукуючий агент, на який рослини в звичайних умовах не відповідають. Прикладом такого індуцибельного промотору є індуцибельний промотор гена стероїдного гормону, транскрипційна активність якого може бути індукована глюкокортикостероїдним гормоном. Див. Schena et al., Proc. Natl. Acad. Sci. U.S.A. 88:0421 (1991). 2. Конститутивні промотори Конститутивний промотор може бути функціонально зв'язаний з експресованим у клітині геном, або конститутивний промотор може бути функціонально зв'язаний з нуклеотидною послідовністю, яка кодує сигнальну послідовність, яка може бути функціонально зв'язана з експресованим у клітині геном. Різні конститутивні промотори можуть бути використані в даному винаході. Приклади конститутивних промоторів включають, без обмеження, промотори вірусів рослин, такі як промотор 35S з CaMV (Odell et al., Nature 313:810-812 (1985)); промотори генів актину рису (McElroy et al., Plant Cell 2:163-171 (1990)); промотор убіквітину (Christensen et al., Plant Mol. Biol. 12:619-632 (1989) і Christensen et al., Plant Mol. Biol. 18:675-689 (1992)); промотор pEMU (Last et al., Theor. Appl. Genet. 81:581-588 (1991)); промотор MAS (Velten et al., EMBO J. 3:2723-2730 (1984)); і промотор гістону Н3 маїсу (Lepetit et al., Mol. Gen. Genetics 231:276-285 (1992) і Atanassova et al., Plant Journal 2 (3):291-300 (1992)); і промотор ALS, XbaІ/NcoI-фрагмент (по 5’кінцю) структурного гена ALS3 Brassica napus (або нуклеотидна послідовність, подібна із зазначеним XbaІ/NcoI-фрагментом. Див. міжнародну публікацію PCT WO 96/30530. 3. Тканиноспецифічні або тканинопереважні промотори Тканиноспецифічний промотор може бути функціонально зв'язаний з експресованим у клітині геном. Необов'язково, цей тканиноспецифічний промотор може бути функціонально зв'язаний з нуклеотидною послідовністю, яка кодує сигнальну послідовність, яка може бути функціонально зв'язана з експресованим у клітині геном. Рослини, трансформовані геном, який представляє інтерес, функціонально зв'язаним із тканиноспецифічним промотором, можуть продукувати білковий продукт цього трансгена винятково або переважно в конкретній тканині. Будь-який тканиноспецифічний або тканинопереважний промотор може бути застосований у даному винаході. Приклади тканиноспецифічних або тканинопереважних промоторів включають, без обмеження: кореневопереважний промотор, наприклад, промотор гена фазеоліну (Murai et al., Science 23:476-482 (1983) і Sengupta-Gopalan et al., Proc. Natl. Acad. Sci. U.S.A. 82:3320-3324 (1985)); листоспецифічний і світлоіндукований промотор, наприклад, промотор cab або rubisco (Simpson et al., EMBO J. 4(11):2723-2729 (1985) і Timko et al., Nature 318:579-582 (1985)); специфічний для пильцевиків промотор, наприклад, промотор LAT52 (Twell et al., Mol. Gen. Genetics 217:240-245 (1989)); специфічний для пилка промотор, наприклад, промотор ZM13 (Guerrero et al., Mol. Gen. Genetics 244:161-168 (1993)), або мікроспоропереважний промотор, наприклад, промотор apg (Twell et al., Sex. Plant Reprod. 6:217-224 (1993)). 8 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 Транспорт продукованого трансгенами білка в субклітинний компартмент, такий як хлоропласт, вакуоль, пероксісома, гліоксисома, клітинна стінка або мітохондрія, або для секреції в апопласт, може бути виконаний за допомогою функціонального зв'язування нуклеотидної кодуючої послідовності сигнальної послідовності 5'- і/або 3'-кінцевої області гена, який кодує білок, який представляє інтерес. Послідовності, які націлюють на 5'- і/або 3'-кінець структурного гена, у ході синтезу і процесінгу білка, можуть визначати, наприклад, де може бути остаточно компартменталізований кодований білок. Альтернативно, такі білки, які націлюють на субклітинний компартмент, можуть бути безпосередньо зв'язані з наночастинкою, для направлення наночастинки, покритої молекулою, яка представляє інтерес, у бажаний субклітинний компартмент. Наявність сигнальної послідовності може направити поліпептид або у внутрішньоклітинну органеллу, або в субклітинний компартмент, або для секреції в апопласт. Багато сигнальних послідовностей є відомими в даній галузі техніки. Див., наприклад, Becker et al., Plant Mol. Biol. 20:49 (1992), Close, P.S., Master's Thesis, Iowa State University (1993), Knox, C, et al., Plant Mol. Biol. 9:3-17 (1987), Lerner et al., Plant Physiol. 91:124-129 (1989), Fontes et al., Plant Cell 3:483-496 (1991), Matsuoka et al., Proc. Natl. Acad. Sci. 88:834 (1991), Gould et al., J. Cell. Biol. 108:1657 (1989), Creissen et al., Plant J. 2:129 (1991), Kalderon, et al., Cell 39:499-509 (1984), Steifel, et al., Plant Cell 2:785-793 (1990). Гени чужорідних білків і агрономічні гени Трансгенні рослини, відповідно до втілень даного винаходу, можуть продукувати чужорідний білок у комерційних кількостях. Таким чином, методи добору і розмноження трансформованих рослин дають множину трансгенних рослин, які збирають загальновідомим способом. Потім чужорідний білок може бути екстрагований із тканини, яка представляє інтерес, або з загальної біомаси. Екстракція білка з біомаси рослини може бути виконана відомими способами, які обговорюються, наприклад, у роботі Heney і Orr, Anal. Biochem. 114:92-96 (1981). У деяких аспектах винаходу, рослинний матеріал, який забезпечує комерційну продукцію чужорідного білка, може бути рослиною, тканиною рослини або клітиною рослини. Для трансгенних рослин, які демонструють більш високі рівні експресії, може бути створена генетична карта, наприклад, за допомогою традиційних аналізів RFLP, PCR і SSR, яка ідентифікує приблизне розташування хромосом в об'єднаній молекулі ДНК. У цьому зв'язку, див. приклади методологій у Glick і Thompson, Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton 269:284 (1993). Інформація з картування, яка стосується розташування хромосом, може бути корисна, наприклад, для патентного захисту трансгенної рослини, як об'єкта винаходу, або для оцінки біологічної безпеки. Якщо можуть бути розпочаті несанкціоновані розмноження і схрещування з іншою зародковою плазмою, ця карта області інтеграції може бути порівняна з аналогічними картами для підозрюваних рослин, для того, щоб визначити, чи є в останньої спільне походження з рослиною, яка є об'єктом винаходу. Порівняння карт можуть включати гібридизації, RFLP, PCR, SSR і секвенування, всі з яких є загальноприйнятими способами. Аналогічно, агрономічні гени можуть бути експресовані в трансформованих клітинах або у їхньому потомстві. Більш конкретно, рослини можуть бути генетично сконструйовані за допомогою способів цього винаходу для експресії різних фенотипів, які представляють агрономічний інтерес. Приклади генів, які можуть бути використані в цьому відношенні, включають, без обмеження, гени, згруповані нижче в окремі категорії. 1. Гени, які додають стійкість до шкідників або захворювань: A) Гени стійкості до патологій рослин. Захист рослин часто активуються специфічною взаємодією між продуктом гена стійкості до патології (R) рослини і продуктом відповідного гена авірулентності (Avr) патогену. Сорти рослин можуть бути трансформовані клонованими генами стійкості для сконструйованих рослин, які є стійкими до специфічних штамів патогенів. Див., наприклад, Jones et al., Science 266:789 (1994) (клонування гена Cf-9 для стійкості томата до Cladosporium fulvum); Martin et al., Science 262:1432 (1993) (ген Pto томата, для стійкості до вражаючого томат Pseudomonas syringae, який кодує протеїнкіназу); Mindrinos et al., Cell 78:1089 (1994) (ген RSP2 для стійкості до Pseudomonas syringae). B) Ген, який додає стійкість до шкідника, такого як соєва цистоутворююча нематода (Heterodera glycines). Див., наприклад, міжнародну публікацію РСТ WO 96/30517; міжнародну публікацію PCT WO 93/19181. C) Білок Bacillus thuringiensis, його похідне або синтетичний поліпептид, який моделює його. Див., наприклад, Geiser et al., Gene 48:109 (1986) (клонування і нуклеотидна послідовність гена Bt δ-ендотоксину). Більш того, молекули ДНК, які кодують гени δ-ендотоксину, можуть бути 9 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 придбані в American Type Culture Collection (Manassas, Va.), наприклад, під номерами доступу АТСС 40098, 67136, 31995 і 31998. D) Лектин. Див., наприклад, Van Damme et al., Plant Molec. Biol. 24:25 (1994) (нуклеотидні послідовності деяких генів лектина, який зв'язує манозу Clivia miniata. E) Білок, який зв'язує вітамін, наприклад, авідин. Див. міжнародну публікацію РСТ US93/06487 (застосування авідину і гомологів авідину як ларвіцидів проти комах-шкідників). F) Інгібітор ферменту, наприклад, інгібітор протеази або протеїнази, або інгібітор амілази. Див., наприклад, Abe et al., J. Biol. Chem. 262:16793 (1987) (нуклеотидна послідовність інгібітору цистеїнпротеїнази), Huub et al., Plant Molec. Biol. 21:985 (1993) (нуклеотидна послідовність кДНК, яка кодує інгібітор I протеїнази тютюну), Sumitani et al., Biosci. Biotech. Biochem. 57:1243 (1993) (нуклеотидна послідовність інгібітору альфа-амілази Streptomyces nitrosporeus) і патент США No. 5494813. G) Специфічний для комах гормон або феромон, наприклад, екдистероїдний або ювенільний гормон, його варіант, міметик на його основі, або його антагоніст або агоніст. Див., наприклад, Hammock et al., Nature 344:458 (1990) (експресії бакуловірусом клонованої естерази ювенільного гормону, інактиватора ювенільного гормону). H) Специфічний для комах пептид або нейропептид, який після експресії порушує фізіологію ураженого шкідника. Див., наприклад, Regan, J. Biol. Chem. 269:9 (1994) (експресійне клонування дає ДНК, яка кодує рецептор діуретичного гормону комахи), і Pratt et al., Biochem. Biophys. Res. Comm. 163:1243 (1989) (алостатин може бути ідентифікований у Diploptera puntata). Див. також патент США No. 5266317 (гени, які кодують паралітичні нейротоксини, специфічні для комах). I) Специфічна для комах отрута, продукована у природі змією, осою або будь-яким іншим організмом. Див., наприклад, Pang et al., Gene 116: 165 (1992) (гетерологічна експресія в рослинах гена, який кодує комахотоксичний для скорпіона пептид). J) Фермент, відповідальний за наднакопичення монотерпену, сесквітерпену, стероїду, гідроксамової кислоти, похідного фенілпропаноїду або іншої небілкової молекули з інсектицидною активністю. K) Фермент, який бере участь у модифікації, включаючи посттрансляційну модифікацію, біологічно активної молекули, наприклад: гліколітичний фермент; протеолітичний фермент; ліполітичний фермент; нуклеаза; циклаза; трансаміназа; естераза; гідролаза; фосфатаза; кіназа; фосфорилаза; полімераза; еластаза; хітиназа; або глюканаза, незалежно від того, чи є вони природними або синтетичними. Див. міжнародну публікацію РСТ WO 93/02197 (нуклеотидна послідовність гена калази). Молекули ДНК, які містять послідовності, які кодують хітиназу, можуть бути отримані, наприклад, з АТСС під номерами доступу 39637 і 67152. Див. також Kramer et al., Insect Biochem. Molec. Biol. 23:691 (1993) (нуклеотидна послідовність кДНК, яка кодує хітиназу тютюнового бражника) і Kawalleck et al., Plant Molec. Biol. 21:673 (1993) (нуклеотидна послідовність гена поліубіквітину ubi-4-2 петрушки). L) Молекула, яка стимулює трансдукцію сигналу. Див. наприклад, Botella et al., Plant Molec. Biol. 24:757 (1994) (нуклеотидні послідовності для кДНК-клонів кальмодуліну квасолі золотавої (Phaseolus aureus)) і Griess et al., Plant Physiol. 104:1467 (1994) (нуклеотидна послідовність кДНК-клону кальмодуліну маїсу). M) Пептид з гідрофобним моментом. Див. міжнародну публікацію PCT WO 95/16776 (пептидні похідні тахиплезину, які інгібують патогени грибів рослин) і міжнародну публікацію РСТ WO 95/18855 (синтетичні антимікробні пептиди, які додають стійкість до захворювань). N) Мембранна пермеаза, утворювач каналів або блокатор каналів. Див., наприклад, Jaynes et al., Plant Sci 89:43 (1993) (гетерологічна експресія аналога цекропін-β-літичного пептиду для додавання трансгенним рослинам тютюну стійкості до Pseudomonas solanacearum). O) Вірус-інвазивний білок або зроблений з нього комплексний токсин. Наприклад, нагромадження вірусних білків оболонки в трансформованих клітинах рослин додає стійкість до вірусної інфекції і/або до розвитку захворювання, яке викликається вірусом, з якого може бути отриманий ген білка оболонки, а також родинними вірусами. Див. Beachy et al., Ann. rev. Phytopathol. 28:451 (1990). Опосередкована білком оболонки стійкість була додана рослинам після трансформації проти вірусу хвороби мозаїки люцерни, вірусу хвороби мозаїки огірка, вірусу смугастості тютюну, вірусу картоплі Х, вірусу картоплі Y, вірусу гравірування тютюну, вірусу погремковості тютюну і вірусу мозаїки тютюну. Див. там же. P) Специфічне для комах антитіло або імунотоксин, зроблений з нього. Так, антитіло, націлене на критичну метаболічну функцію в кишечнику комахи, може інактивувати фермент, який знаходиться під його впливом, призводячи до загибелі комахи. Порівн. Taylor et al., Abstract #497, Seventh Іnt'l Symposium on Molecular Plant-Microbe Interactions (Edinburgh, Scotland) (1994) 10 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 (ферментативна інактивація в трансгенному тютюні за рахунок продукування одноланцюжкових фрагментів антитіл). Q) Вірус-специфічне антитіло. Див., наприклад, Tavladoraki et al., Nature 366:469 (1993) (трансгенні рослини, експресуючі гени рекомбінантних антитіл, захищені від вірусної атаки). R) Затримуючий розвиток білок, продукований у природі патогеном або паразитом. Наприклад, ендо-1,4-D-полігалактуронази грибів полегшують колонізацію грибів і вивільнення поживних речовин рослин солюбілізацією гомо-α-1,4-D-галактуронази клітинних стінок рослини. Див. Lamb et al., Bio/Technology 10:1436 (1992). Див. також Toubart et al., Plant J. 2:367 (1992) (клонування і характеризація гена, який кодує білок, інгібуючий ендополігалактуроназу бобів). S) Затримуючий розвиток білок, продукований у природі рослиною. Див. наприклад, Logemann et al., Bio/Technology 10:305 (1992) (трансгенні рослини, експресуючі ген, інактивуючий рибосоми ячменю, мають збільшену стійкість до грибних захворювань). 2. Гени, які додають стійкість до гербіциду: A) Гербіцид, який інгібує точку росту або меристему, наприклад, імідазолінон або сульфонілсечовина. Прикладами генів у цій категорії є гени, які кодують мутантний білок ALS і AHAS, як описано, наприклад, Lee et al., EMBO J. 7:1241 (1988), і Miki et al., Theor. Appl. Genet. 80:449 (1990), відповідно. B) стійкість до гліфосату, яка додається, наприклад, мутантними генами 5енолпірувілшикімат-3-фосфатсинтази (EPSP) (за рахунок введення рекомбінантних нуклеїнових кислот і/або різних форм мутагенезу нативних генів EPSP in vivo), генами aroА і генами гліфосатуцетилтрансферази (GAT), відповідно); інші фосфонові сполуки, такі як глюфосинат (гени фосфінотрицинацетилтрансферази (РАТ) з Streptomyces species, включаючи Streptomyces hygroscopicus і Streptomyces viridichromogenes), і піридинокси- або фенокси-пропіонові кислоти і циклогексони (гени, які кодують інгібітори ACC-ази). Див., наприклад, патент США No. 4940835 і патент США No. 6248876 (нуклеотидні послідовності форм EPSP, які можуть додавати рослині стійкість до гліфосату). Молекула ДНК, яка кодує мутантний ген aroА, може бути отримана під номером доступу АТСС 39256. Див. також патент США No. 4769061 (нуклеотидна послідовність мутантного гена aroА). Європейська заявка на патент No. 0333033 і патент США No. 4975374 розкривають нуклеотидні послідовності генів глутамінсинтетази, які можуть додавати стійкість до гербіцидів, таких як L-фосфінотрицин. Нуклеотидна послідовність ілюстративних генів PAT надана в європейській заявці No. 0242246 і в роботі DeGreef et al., Bio/Technology 7:61 (1989) (отримання трансгенних рослин, які експресують химерні гени bar, які кодують активність РАТ). Приклади генів, які додають стійкість до феноксипропіонових кислот і циклогексонів, таких як сетоксидим і галоксифоп, включають гени Асс1-S1, Acc1-S2 і Асс1-S3, описані Marshall et al., Theor. Appl. Genet. 83:435 (1992). Гени GAT, здатні додавати стійкість до гліфосату, описані, наприклад, у WO 2005012515. Гени, які додають стійкість до 2,4-D, фопу і піридилоксіауксинових гербіцидів, описані, наприклад, у WO 2005107437. C) Гербіцид, який інгібує фотосинтез, такий як триазин (гени psbА і гени gs+) або бензонітрил (ген нітрилази). Див., наприклад, Przibila et al., Plant Cell 3:169 (1991) (трансформація Chlamydomonas плазмідами, які кодують мутантні гени psbА). Нуклеотидні послідовності для генів нітрилази розкриті в патенті США No. 4810648, а молекули ДНК, які містять ці гени, доступні під номерами доступу АТСС 53435, 67441 і 67442. Див. також Hayes et al., Biochem. J. 285: 173 (1992) (клонування й експресія ДНК, яка кодує глутатіон-Sтрансферазу). 3. Гени, які додають або вносять внесок в ознаку доданої цінності, такої як: A) Модифікований метаболізм жирних кислот, наприклад, шляхом трансформації рослини антисмисловим геном стеарил-АСР-десатурази для збільшення вмісту стеаринової кислоти в цій рослині. Див. Knultzon et al., Proc. Natl. Acad. Sci. U.S.A. 89:2624 (1992). B) Зменшений вміст фітату. Введення гена, який кодує фітазу, може підсилити руйнування фітату, додаючи тим самим додатковий вільний фосфат трансформованій рослині. Див. наприклад, Van Hartingsveldt et al., Gene 127:87 (1993) (нуклеотидна послідовність гена фітази Aspergillus niger). Можливе введення гена для зменшення вмісту фітату. Наприклад, у маїсі це може бути виконано клонуванням з наступним повторним введенням ДНК, асоційованої з єдиним алелем, який може бути відповідальним за мутанти маїсу, які характеризуються низькими рівнями фітинової кислоти. Див. Raboy et al., Maydica 35:383 (1990). C) Модифікований склад вуглеводів, вироблений, наприклад, трансформацією рослин геном, який кодує фермент, який змінює характер розгалуження крохмалю. Див. Shiroza et al., J. Bacteol. 170:810 (1988) (нуклеотидна послідовність гена фруктозилтрансферази мутантів Streptococcus), Steinmetz et al., Mol. Gen. Genet. 20:220 (1985) (ген левансахарази), Pen et al., Bio/Technology 10:292 (1992) (α -амілаза), Elliot et al., Plant Molec. Biol. 21:515 (1993) 11 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 (нуклеотидні послідовності генів інвертази томата), Sogaard et al., J. Biol. Chem. 268:22480 (1993) (ген β-амілази ячменю) і Fisher et al., Plant Physiol. 102:1045 (1993) (фермент II розгалуження крохмалю ендосперму маїсу). C. Наночастинки Стосовно деяких втілень, винахід надає способи введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину, яка включає клітинну стінку (наприклад, клітину рослини). У деяких втіленнях спосіб може включати розміщення наночастинки, покритої лінійною молекулою нуклеїнової кислоти, яка представляє інтерес, у контакті з клітиною, і надавання можливості поглинання наночастинки через клітинну стінку. У конкретних втіленнях наночастинка може оборотно або необоротно містити, бути покритою, або іншим способом зв'язаною з і/або нести лінійну молекулу нуклеїнової кислоти, яка представляє інтерес. У деяких втіленнях лінійна молекула нуклеїнової кислоти, яка представляє інтерес, може бути введена в наночастинки перед контактом із клітиною рослини, яка має клітинну стінку, або одночасно, із введенням наночастинки в клітину рослини, яка має клітинну стінку. Приклади наночастинок, які можуть використовуватися у втіленнях даного винаходу, включають без обмеження, квантові точки, окремо або в комбінації з напівпровідниковими наночастинками; позитивно заряджені наночастинки; наночастинки золота; покриті золотом наночастинки, пористі наночастинки, мезопористі наночастинки, наночастинки діоксиду кремнію, полімерні наночастинки, включаючи дендримери, вольфрамові наночастинки, желатинові наночастинки, нанооболонки, наноядра, наносфери, нанострижні і магнітні наночастинки. Стосовно втілень даного винаходу, клітина рослини, яка має клітинну стінку, може бути будь-якою клітиною рослини, яка включає інтактну і цілу клітинну стінку. Приклади клітин, які мають клітинну стінку, включають, без обмеження, водорості, тютюн, моркву, маїс, канолу, рапс, бавовник, пальму, арахіс, сою, цукрову тростину, Oryza sp., Arabidopsis sp. і Ricinus sp. Втілення винаходу можуть включати клітини, які мають клітинну стінку, з будь-якої тканини або з будь-якого місця, де вони можуть бути знайдені, включаючи без обмеження: у зародках, меристематичних клітинах, калусі, пилку, включаючи гаплоїдні і диплоїдні мікроспори, листях, пильцевиках, корінні, кінчиках коріння, квітах, насінні, стручках, стеблах і тканинній культурі. У втіленнях винаходу лінійна молекула нуклеїнової кислоти, яка представляє інтерес, може бути будь-якою молекулою нуклеїнової кислоти, яку можна доставити стосовно даного винаходу в клітину рослини, яка має клітинну стінку. Молекули нуклеїнової кислоти, які представляють інтерес, можуть включати, без обмеження: ДНК; РНК; молекули RNAi; гени; плазміди; косміди; YAC (штучної хромосоми грибів) і BAC (штучної хромосоми бактерій). Молекули нуклеїнової кислоти, які представляють інтерес, можуть бути введені в клітину рослини, яка має клітинну стінку разом з, наприклад, але без обмеження, поліпептидами, ферментами, гормонами, глікопептидами, цукрами, жирами, сигнальними пептидами, антитілами, вітамінами, месенджерами, вторинними месенджерами, амінокислотами, цАМФ, лікарськими речовинами, гербіцидами, фунгіцидами, антибіотиками і/або з їх комбінаціями. У конкретних втіленнях винаходу поверхня наночастинки може бути функціоналізована, що може, наприклад, забезпечувати націлене поглинання або допускати оборотне або необоротне зв'язування інших речовин з поверхнею наночастинки. Як необмежуючий приклад, поверхня наночастинки (наприклад, квантової точки) може бути функціоналізована моношаром, який самозбирається наприклад, алкантіолатами, які можуть бути додатково функціоналізовані або дериватизовані. У додатковому необмежуючому прикладі поверхня наночастинки може бути дериватизована лінкерами, які самі потім можуть бути додатково функціоналізовані або дериватизовані. В одному втіленні наночастинка може бути ПЕГильована. В іншому втіленні наночастинка може містити або може бути мультифункціоналізована одним або декількома ядрами (активними або неактивними), стеричним (просторовим) покриттям (активним або інертним), який розщеплюється зв'язком і/або націлювальними молекулою або лігандом. Наночастинки, такі як квантові точки, можуть бути функціоналізовані ПЕГом, використовуючи протокол Dubertret et al., Science 298:1759 (2002), або відповідно до цього протоколу, зміненим за розсудом кваліфікованого фахівця. Наприклад, квантові точки CdSe/ZnS, покриті TOPO (три-октилфосфіноксид) можуть бути суспендовані в хлороформі з PEG-PE, з наступним випарюванням розчинника і солюбілізацією з водою одержуваних ПЕГильованих квантових точок. В аспектах винаходу наночастинка може поглинатися в різні частини клітин. Приклади місць розташування, в які може поглинатися наночастинка, включають, без обмеження, цитозоль, ядро, тонопласти, пластиди, етіопласти, хромопласти, лейкопласти, елайопласти, протеїнопласти, амілопласти, хлоропласти і просвіт подвійної мембрани. В інших втіленнях, 12 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 поглинання наночастинки в клітину, яка містить клітинну стінку, може здійснюватися за допомогою симпластного або апопластного шляху. D. Стабільно трансформовані клітини рослини Стабільно трансформована клітина рослини за винаходом може бути будь-якою клітиною рослини, здатною до того, щоб бути трансформованою лінійною молекулою нуклеїнової кислоти, яка представляє інтерес, шляхом трансформації, опосередкованою наночастинками. Відповідно, клітина рослини може бути виділена або культивована з дводольної або однодольної рослини. Клітина рослини може також бути представлена в тканині рослини або в цілій рослині. Необмежуючі приклади стабільно трансформованих клітин дводольних рослин за винаходом включають клітини: люцерни; бобів; броколі; капусти; моркви; цвітної капусти; селери; китайської капусти; бавовнику; огірка; баклажану; салату; дині; гороху; перцю; арахісу; картоплі; гарбуза; редису/редьки; рапсу; шпинату; сої; гарбуза; цукрового буряка; соняшника; тютюну; томата і кавуна. Необмежуючі приклади стабільно трансформованих клітин однодольних рослин за винаходом включають клітини зернових, цибулі, рису, сорго звичайного, пшениці, жита, проса, цукрової тростини, вівса, тритикале, проса лозиноподібного і газонної трави. Трансгенні рослини відповідно до винаходу можуть бути регенеровані зі стабільно трансформованих клітин рослини, отриманих способами за винаходом. Такі рослини можуть використовуватися або вирощуватися будь-яким способом, у якому бажана присутність молекул нуклеїнової кислоти, які представляють інтерес. Відповідно, трансгенні рослини inter alia можуть бути отримані методами генної інженерії для того, щоб мати одину або декілька бажаних генетичних ознак, будучи трансформованими лінійними молекулами нуклеїнової кислоти за допомогою опосередкованої наночастинками трансформації, з посадкою і культивацією будьяким способом, відомим фахівцю в даній галузі техніки. Наступні приклади надані для ілюстрації окремих специфічних ознак і/або втілень. Приклади не повинні розглядатися як такі, що обмежують даний винахід окремими ознаками й ілюстративними втіленнями. Приклади Приклад 1: Отримання наночастинок для трансформації клітини рослини Отримання плазмідної ДНК ДНК плазміди pDAB3831, Фіг. 1, була виділена і підготовлена для трансформації рослини, опосередкованою лінійною ПЕГильованою квантовою точкою(PQD)/ДНК. Ця плазміда містить селектований ген маркера PAT, який запускається промотором 10 Arabidopsis Ubiquitin (AtUbi10), і ген жовтого флуоресцентного білка Philadium (JPhiYFP), який запускається промотором вірусу прожилкової мозаїки маніоки (CsVMV). Штам Escherichia coli, який містить плазміду, був інокульований і вирощений до появи мутності в бульйоні Luria-Bertani, який містить ампіцилін при 37 °C. ДНК була виділена з використанням набору Qiagen Plasmid MidiPrep kit (Qiagen, Valencia, CA). Виділена ДНК лінеаризувалась за допомогою розщеплення ферментом рестрикції. Для розщеплення ДНК використовувався фермент рестрикції KpnІ, одержуючи, у такий спосіб лінеаризовану плазмідну ДНК. Отримання лінійних ДНК-PQD комплексів Квантові точки були отримані з Ocean Nanotechnology (Springdale, AZ). Квантові точки CdSe/ZnS, покриті 2 мг TOPO (триоктилфосфіноксид), були суспендовані в хлороформі з 0,15 мг (5,5 мкмМ) PEG-PE (1,2-діацил-sn-гліцеро-3-фосфоетаноламін-N-[метокси-полі(етиленгліколь)]) (Avanti Polar Lipids, Alabaster, AL), з наступним випарюванням розчинника і солюбілізації з водою. Кон'югування з ПЕГ було виконано для захисту від цитотоксичності. PQD були кон'юговані з лінійної плазмідної ДНК. 2 мг PQD з 4 мг HS-PEG-OCH3 (Prochimia, Zacisze, Польща) були суспендовані протягом ночі при приблизно 60-70 °C. Розчинник був вилучений у вакуумній печі. Потім залишок був суспендований у 1 мл води (18M). Остання стадія супроводжувалася переходом осаду червоного кольору в жовтогарячий, оптично прозорий чистий розчин. Для покриття H3CO-PEG-SH-QD лінеаризованої плазмідної ДНК для експериментів по трансформації, 0,02 мг очищеної лінеаризонної плазмідної ДНК (pDAB3831) інкубували з отриманими кон'югатами PQD у 2 мл води протягом 2 годин при 23 °C у темряві. Torney, et al., Nature Nanotechnol. 2:295-300 (2007). Приклад 2: Трансформація квіткових зародків Arabidopsis Для генетичної трансформації in planta, плазміда pDAB3831 була розщеплена ферментом рестрикції KpnІ, для лінеаризації ДНК. Після розщеплення ферментом рестрикції, лінеаризована ДНК використовувалася при визначеному співвідношенні ПЕГ, QD і лінійної ДНК. Лінеаризована ДНК була змішана з квантовими точками і ПЕГ, і інкубувалась протягом 30 хвилин. Потім, нанокомплекс, який складається з наночастинок і розчину лінеаризованої 13 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 плазмідної ДНК, був змішаний з робочим розчином, який містить 5 %-ий розчин сахарози у воді з 0,02-0,04 % Silwet-L77. Рослинний матеріал для трансформації in planta Синхронізоване проростання насіння є важливим для гарантії однорідності розвитку квіток на рослинах T0. Насіння A. thaliana cv. Columbia були суспендовані в розчині агару 0,1 % (мас./об.) і інкубували при 4 °C протягом 48 годин, до повної стратифікації. Було відважено 60 мг насіння і перенесено в пробірку на 15 мл. Для рівномірного розподілу насіння у пробірку додавали 13 мл 0,1 % (мас./об.) розчин агару, і її піддавали перемішуванню обертанням. Це привело до суспензії з концентрацією насіння з розрахунку 4,6 мг на 1 мл 0,1 % (мас./об.) розчин агару (або приблизно 230 насінин/мл). Були підготовлені 6 пробірок (72 мл суспензії) для висівання в 4 піддонах, які вміщують 18 чашок (3 1/2 дюйми) у кожному піддоні. Суспензію насіння інкубували протягом 48 годин при 4 °C для завершення стратифікації. У кожну чашку індивідуально засівали 1,0 мл суспензії стратифікованого насіння на одну чашку. Після засівання всіх чашок, піддони були закриті куполами для підтримки ґрунту в зволоженому стані при рості рослин. Купола були вилучені через 5 днівпісля дати висівання. Після проростання насіння і появи рослин, вони були поміщені для росту в камеру Conviron® (моделі CMP4030 і CMP3244, Controlled Environments Limited, Winnipeg, Manitoba, Canada) на режим довгого дня 2 (16 годин освітлення/8 годин темряви), при інтенсивності світла 120-150 мкмМоль/м ·сек (по еквіваленту світлової енергії, поглиненої рослиною), при постійній температурі (22 °C) і вологості (40-50 %). Через 10-14 днів після висівання рослин їх піддавали зрошенню розчином Hoagland's і потім DI водою, для того, щоб тримати ґрунт зволоженим, але не мокрим. Через 4 тижні після дати висівання рослин у них були зрізані квіти для інтенсифікації росту вторинних квіток. Рослини були готові до процесу трансформації на 5-му тижні після висівання. Трансформації in planta Трансформація A. thaliana cv. Columbia за допомогою лінійної ДНК/PQD була виконана з використанням модифікованого протоколу Clough–Bent. Див., Clough and Bent, Plant J. 16:73543 (1998). Було приготовлено 20 мл суспензії комплексу лінійної ДНК/PQD при концентрації 0,5 мг лінійної ДНК і 4 нМ PQD, яка використовувалася для обробки рослин Arabidopsis (головним чином незрілі суцвіття, які включають невелику кількість запліднених стручків). Перед зануренням рослин, до суспензії комплексу лінійної ДНК/PQD був доданий Silwet L-77 до концентрації 0,05 % (мас./об.) (250 мкл/500 мл) - 0,005 %, і розчин був добре перемішаний. Надземні частини рослини були занурені на 30 секунд у розчин лінійної ДНК/PQD з обережним перемішуванням. Оброблені рослини були поставлені у висхідне положення у тінь на 30 хвилин при 22-24 °C. Рослини були розміщені в камеру Conviron в умови, як описано вище, для надавання можливості дозрівання і збору насіння. Для скринування загального врожаю насіння від рослин T0 використовувалися лотки для селекції розміром 10,5×21×1 дюймів, з розрахунку приблизно 10000 насінин на лоток. Для гарантії того, що обприскування при селекції виконано коректно, використовували дві групи контролю: рослини Col-0 як негативний контроль трансформанта, і рослини Columbia Col0 дикого типу, збагачені гомозиготними насінинами для селектованого маркера PAT (фосфінотрицин-N-ацетилтрансферази), як позитивний контроль трансформанта. Насіння, з метою синхронізації, було стратифіковане в розчині агару з концентрацією 0,1 % (мас./об.) протягом 48 годин до висівання. Для забезпечення щільності висівання 10000 насінин на лоток для селекції, до розчину агару з концентрацією 0,1 % (мас./об.) було додано 200 мг насіння, і розчин був підданий перемішуванню обертанням доти, поки насіння не було рівномірно суспендоване. Потім, стратифіковане насіння висівали на лотках для селекції, які були заповнені ґрунтовою сумішшю Sunshine LP5 і одержували для зволоження розчин Hоаglаnd's шляхом подачі розчину знизу. Для ефективного обприскування при селекції рослин важливо, щоб 40 мл суспендованого насіння було рівномірно висіяно в лоток для селекції. Після висівання, кожен лоток для селекції був закритий куполом, і рослини вирощувалися для селекції. Купола були вилучені приблизно через 5 днів після висівання. Приклад 3: Аналіз трансформованого Arabidopsis Селекція трансформованих рослин Свіжозібраному насінню T1 давали можливість висохнути протягом 7 днів при кімнатній температурі. Насіння T1 висівали в лотки розміром 26,5×51 см для проростання, при цьому в кожен висівалася аліквота 200 мг стратифікованого насіння T1 (приблизно 10000 насінин), які були раніше суспендовані в 40 мл 0,1 % (мас./об.) розчину агару, і зберігалися при 4 °C протягом 2 днів, через вимоги завершення періоду спокою і для гарантії синхронного проростання насіння. 14 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Ґрунтова суміш Sunshine LP5 була покрита чистим вермікулітом і піддавалася зволоженню розчином Hоаglаnd's шляхом подачі розчину знизу з наступним видаленням надлишку вологи за рахунок сили тяжіння. Кожна аліквота стратифікованого насіння об'ємом 40 мл рівномірно наносилася піпеткою на вермікуліт і закривалася куполами для збереження вологості протягом 4-5 днів. Купола були вилучені за 1 день до первинної селекції трансформантів з використанням обприскування глюфосатом у післясходовий період. Через сім днів після посадки (DAP) (від сім'ядолі і стадії 2-4 листків, відповідно) рослини T1 послідовно обприскували п'ять разів протягом п'яти днів 0,2 %-им розчином (мас./об.) гербіциду Liberty (200 м глюфосинату по активній речовині на 1 л, Bayer Crop Sciences, Kansas City, MO) при об'ємі обприскування 10 мл на лоток (703 л/га) з використанням наконечника DeVilbiss для розпилення стисненим повітрям, щоб забезпечити доставляння ефективної кількості глюфосинату при нормі 280 г/га глюфосинату по активній речовині на одне обприскування. Рослини, які вижили, (активно зростаючі рослини), були ідентифіковані через 4-7 днів після заключного обприскування і пересаджені в індивідуальні 3-дюймові горщики з поживним середовищем для росту в горщиках (Metro Mix 360). Пересаджені рослини були покриті куполами для збереження вологості на 3-4 дні, і вони були вміщені для росту в камеру при 22 °C, як і до того, або були перенесені безпосередньо в теплицю. Згодом купола були зняті, і рослини були розміщені в теплиці (22±5 °C; відносна вологість 50±30 %; світловий режим: 14 годин освітлення/10 годин темряви; освітленість (природне + додаткове світло) по еквіваленті 2 поглиненої рослиною світлової енергії - 500 мкмМоль/м ·сек). Молекулярні і біохімічні дослідження Були виконані молекулярні аналізи по інтеграції PAT і YFP у геном рослини. Продукти ампліфікації ПЛР були секвеновані і порівняні з трансгенними послідовностями. Молекулярний аналіз і доказ геномної інтеграції трансгенів у потомство T1 Arabidopsis thaliana (cv Columbia) Геномна ДНК із трансгенних рослин A. thaliana була витягнута з матеріалу листка 6тижневих рослин з використанням набору Plant DNAZOL™ (Invitrogen) відповідно до інструкцій виробника. Для виявлення трансгенів PAT і YFP були сконструйовані праймери для ПЛР. Приклади YFP представлені як SEQ ID NO: 1 і SEQ ID NO: 2. Праймери PAT представлені як SEQ ID NO: 3 і SEQ ID NO: 4. ПЛР ампліфікація геномної ДНК трансгенів Реакції ампліфікації ПЛР для PAT і YFP були виконані з використанням набору TaKaRa ExTaq™ (Takara, Otsu, Shiga, Japan). Генні продукти були ампліфіковані в повному об'ємі реакції, яка складає 50 мкл. Реакція ПЛР містила 100 нанограмів геномної матриці ДНК, реакційних буферів 1 X ExTaq, 0,2 мМ dNTP, 10 пМ кожного прикладу і 0,025 Од/мкл ExTaq. Використовувалися наступні умови ПЛР: 1 цикл при 96 °C протягом 5 хвилин, і 31 цикл при наступних умовах: 94 °C протягом 15 секунд, 65 °C протягом 30 секунд, 72 °C протягом 1 хвилини і заключна ампліфікація при 72 °C протягом 7 хвилин. Продукт ампліфікації ПЛР був проаналізований електрофорезом на 0,8 % гелі агарози і був візуалізований фарбуванням бромістим етидієм. Фрагменти ДНК були виділені з гелю агарози, використовуючи набір для очищення з гелю QIAEX™ II (Qiagen, Valencia, CA). Фрагменти з ПЛР були секвеновані, використовуючи прямий праймер для PAT (SEQ ID NO: 3) і зворотний праймер для YFP (SEQ ID NO: 1), з використанням поліпшеної методики секвенування по Sanger (MWG Biotech., Huntsville, AL). Дані по послідовностях були проаналізовані з використанням програмного забезпечення Sequencher. Результати секвенування ампліконів PAT і YFP із ПЛР відповідали очікуваній нуклеотидній послідовності для цих генів. Ці результати зрозуміло показують, що послідовності PAT і YFP з pDAB3831 були стабільно інтегровані в геномну ДНК Arabidopsis при використанні протоколу трансформації ПЕГильованою квантовою точкою і лінійною ДНК. Генні продукти для PAT і YFP (жовта флуоресцентна мітка, Evrogen) із ПЛР були ампліфіковані в загальному об'ємі реакції 50 мкл суміші, яка містить 100 нанограмів матричної геномної ДНК, реакційний буфер 1Х ExTaq (TaKaRa Bio), 0,2 мМ dNTP, 10 пМ праймера і 0,025 Од/мкл ExTaq. Використовувалися наступні умови ПЛР: 1 цикл при 96 °C протягом 5 хвилин і 31 цикл по наступній програмі ПЛР: 94 °C, 15 секунд; 65 °C, 30 секунд; 72 °C, 1 хв. Заключна ампліфікація була виконана при 72 °C протягом 7 хвилин, для повного синтезу продукту ПЛР. Зображення гелю були отримані з використанням системи візуалізації BioRad Gel Imagining System. Див. Фіг. 1 і 2. Ампліфіковані фрагменти були виділені з гелю, використовуючи набір для очищення з гелю (Qiagen, Valencia, CA) відповідно до інструкцій виробника. Фрагменти з ПЛР були секвеновані, використовуючи прямий праймер для PAT і прямий праймер для YFP, з використанням поліпшеної методики секвенування по Sanger (MWG 15 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 Biotechnologies, Inc.), а послідовності були проаналізовані з використанням програмного забезпечення Sequencher. Дані результати зрозуміло показують, що послідовності PAT і YFP були доставлені за допомогою позитивно зарядженої наночастинки, яка містить лінеаризовану ДНК, отриману в Прикладі 1, і в такий спосіб забезпечують доказ стабільної інтеграції трансгенів у геномну ДНК рослин T1 Arapidopsis. Приклад 4: Опосередковане наночастинками доставляння лінійних молекул нуклеїнової кислоти в культивовані клітини рослини Підготовка рослинного матеріалу у вигляді одиничної клітини Наприклад, використовуються як клітини BY2, так і клітини NT1. Клітини BY2 не мають зеленого кольору і являють собою швидкозростаючу лінію клітин тютюну. Клітини NT1 являють собою фотоавтотрофні клітини, виділені з тютюну. За три - чотири дні до трансформації, однотижневу суспензійну культуру субкультивували у свіжому середовищі шляхом перенесення 2 мл культури NT1 або BY2 у 40 мл середовища NTIB або LSBY2, яке містить 50 нм DAS-PMTI-1 (інгібітор мікроканальців) і 0,5-0,1 % (об./об.) диметилсульфоксиду, у колбу на 250 мл. Одиничні клітини були зібрані або через чотири дні або через сім днів після обробки інгібітором мікроканальців. Використовувані одиничні клітини BY2 були проаналізовані за допомогою проточного цитометра Beckman Flow для підрахунку життєздатних клітин. Клітини досліджені з використанням мікроскопа диференційно-інтерференційного контрасту (DIC), оснащеного конфокальною системою відображення, для встановлення того, що одиничні клітини включають великі кількості пластид (амілопластів), розподілених по всій цитоплазмі клітин. Клітини субкультивували один раз у 14 днів, шляхом перенесення 1 мл суспензії з 3,0 Од оптичної 600 густини при OD . Культивовані клітини використовуються як цільові клітини для трансформації. Препарат наночастинок і обробка клітин Плазмідна ДНК виділена і підготовлена для трансформації рослини, опосередкованою лінійною ПЕГильованою квантовою точкою(PQD)/ДНК. Плазміда містить ген селектованого маркера PAT, який запускається промотором 10 Arabidopsis Ubiquitin (AtUbi10) і ген жовтого флуоресцентного білка Philadium (JPhiYFP), який запускається промотором вірусу прожилкової мозаїки маніоки (CsVMV). Штам Escherichia coli, який містить плазміду, був інокульований і вирощений до появи мутності в бульйоні Luria-Bertani, який містить ампіцилін при 37 °C. ДНК була видалена з використанням набору Qiagen Plasmid Midi-Prep kit (Qiagen, Valencia, CA). Видалена ДНК лінеаризувалась за допомогою розщеплення ферментом рестрикції. Для розщеплення ДНК використовувався фермент рестрикції KpnІ, одержуючи, у такий спосіб лінеаризовану плазмідну ДНК. Квантові точки були отримані з Ocean Nanotechnology (Springdale, AZ). Квантові точки CdSe/ZnS, покриті 2 мг TOPO (триоктилфосфіноксид), були суспендовані в хлороформі з 0,15 мг (5,5 мкмм) PEG-PE (1,2-діацил-sn-гліцеро-3-фосфоетаноламін-N-[метокси-полі(етиленгліколь)]) (Avanti Polar Lipids, Alabaster, AL), з наступним випарюванням розчинника і солюбілізації з водою. Кон'югування з ПЕГ було виконано для захисту від цитотоксичності. PQD були кон'юговані з лінійної плазмідної ДНК. 2 мг PQD з 4 мг HS-PEG-OCH3 (Prochimia, Zacisze, Польща) були суспендовані протягом ночі при приблизно 60-70 °C. Розчинник був вилучений у вакуумній печі. Потім залишок був суспендований у 1 мл води (18M). Остання стадія супроводжувалася переходом осаду червоного кольору в жовтогарячий, оптично прозорий чистий розчин. Для покриття H3CO-PEG-SH-QD лінеаризованої плазмідної ДНК для експериментів по трансформації, 0,02 мг очищеної лінеаризонної плазмідної ДНК (pDAB3831) інкубували з отриманими кон'югатами PQD у 2 мл води протягом 2 годин при 23 °C у темряві. Torney, et al., Nature Nanotechnol. 2:295-300 (2007). PQD при концентрації 1-3 мкл/мл були додані в 24-ямковий мікротитрувальний планшет до 500 мкл клітин, і планшет піддавали обережному обертанню в шейкері протягом 20 хвилин у темряві. Наночастинки пройшли через клітинні стінки. Приклад 5: Трансформація Arabidopsis, опосередкована мультифункціоналізованою наночастинкою in planta Трансформація Arabidopsis in planta може бути виконана з використанням модифікованого протоколу Clough-Bent (1998). Концентрація ДНК на мультифункціоналізованій наночастинці разом з молекулами хоумінгу, такими як домен білкової трансдукції (PTD) і одиницями сигналу ядерної локалізації (NLS), оптимізована для того, щоб досягти підвищеної ефективності трансформації. Рослинний матеріал: Здорові рослини Arabidopsis вирощені в режимі тривалого дня в горщиках із ґрунтом до цвітіння. Перші стрілки були підрізані для стимуляції проліферації 16 UA 113046 C2 5 10 15 20 25 30 35 40 45 50 55 60 множини вторинних стрілок. Рослини були готові через приблизно 4-6 днів після підрізування. Arabidopsis thaliana екотипу Columbia (Col-0) використовувалися як базове тло (рослина T0) для трансформації квіток in planta. Синхронізоване проростання насіння є важливим для того, щоб гарантувати однорідність розвитку квіток на рослинах T0. Насіння дикого типу суспендували в 0,1 % розчині агару і інкубували при 4 °C протягом 48 годин для завершення стратифікації. 60 мг насіння зважували на папері і переносили в пробірки на 15 мл. Додавали 13 мл 0,1 %-ого розчину агару і пробірки піддавали перемішуванню обертанням, доти, поки насіння не буде рівномірно дисперговане. Це привело до суспензії з концентрацією насіння з розрахунку 4,6 мг на 1 мл розчину (або приблизно 230 насіння/мл). Були підготовлені 6 пробірок (72 мл суспензії) для висівання в 4 піддонах, які вміщають 18 чашок (3 1/2 дюйми) у кожному піддоні, і в 2 загальних чашках. Суспензію насіння інкубували протягом 48 годин при 4 °C для завершення стратифікації. У кожну чашку індивідуально засівали 1,0 мл суспензії стратифікованого насіння на одну чашку. Після засівання всіх чашок, піддони були закриті куполами для підтримки ґрунту в зволоженому стані при рості рослин. Купола були вилучені через 5 днів після дати висівання. Після проростання насіння і появи рослин, вони були вміщені для росту в камеру Conviron® (моделі CMP4030 і CMP3244, Controlled Environments Limited, Winnipeg, Manitoba, Canada) на режим довгого дня (16 годин освітлення/8 годин темряви), при інтенсивності світла 120-150 2 мкмМоль/м ·сек (по еквіваленту світлової енергії, поглиненої рослиною), при постійній температурі (22 °C) і вологості (40-50 %). Через 10-14 днів після висівання рослин їх піддавали зрошенню розчином Hоаglаnd's і потім DI водою, для того, щоб тримати ґрунт зволоженим, але не мокрим. Через 4 тижні після дати висівання рослин у них були зрізані квіти для інтенсифікації росту вторинних квіток. Рослини були готові до процесу трансформації на 5-му тижні після висівання. Підготовка нанокон'югатів для обробки квіток: Наночастинки/квантові точки в діапазоні розмірів 2-120 нм були вибрані для обробки, і мультифункціоналізовані фрагментом очищеної pDAB3138 і одиницями пептиду для хоумінгу згідно з Derufus et al. (2007). Квантові точки з максимумами емісії при 655 або 705 нм, і модифіковані за допомогою ПЕГ з аміногрупами, були отримані з Quantum Dot Corporation (ITK amino). Концентрації QD вимірювали по оптичному поглинанню при 595 нм, використовуючи коефіцієнти екстинкції, надані постачальником. Використані крос-лінкери являли собою сульфо-LC-SPDP (сульфосукцинімідил 6-(3'-[2-піридилдитіо]-пропіонамідо)гексаноат) (Pierce) і сульфо-SMCC (сульфосукцинімідил-4-(N-малеїмідометил)циклогексан-1-карбоксилат) (Sigma). Модифіковані аміном QD кон'югували з тіол-вмісною плазмідною ДНК і пептидами хоумінга, використовуючи сульфо-LC-SPDP і сульфо-SMCC як крос-лінкери. QD ресуспендували в розчині 50 мМ фосфату натрію, 150 нМ хлористого натрію, pН 7,2, використовуючи фільтри Amicon Ultra-4 (з відсіченням 100 кДа). ДО QD додавали крос-лінкер (1000-кратний надлишок) з надаванням можливості реагувати протягом 1 години. Зразки відфільтровували на колонці NAP-5 під дією сили тяжіння (для видалення надлишку крос-лінкера) в аналогічний буфер, доповнений 10 мМ EDTA. Лінеаризовану плазміду ДНК pDAB3831 обробляли 0,1М DTT протягом 1 години й відфільтровували на колонці NAP-5 у EDTA-вмісний буфер. Як правило, використовували пептиди у формі ліофілізованого порошку. Пептиди і лінеаризована плазмідна ДНК були додані до відфільтрованих QD з надаванням можливості реагувати протягом ночі при 4 °C. Використовуючи три фільтри Amicon, продукт був відфільтрований фосфатно-фізіологічним розчином по Dulbecco (PBS), двічі з високосольовим буфером (1,0М хлористого натрію, 100 мМ цитрат натрію, pН 7,2), і знову двічі з PBS. Відмивання з високосольовим буфером необхідні для видалення електростатично зв'язаних ДНК і пептидів, які не вилучені відмиванням тільки одним PBS. Сульфо-SMCC має на одному кінці N-гідроксисукцинімідний ефір (NHS), який реагує з модифікованими аміном QD, з утворенням амідного зв'язку, і малеїмідної групи на іншому кінці, яка реагує з тіольованою плазмідною ДНК, з утворенням тіоефіру. Сульфо-LC-SPDP також містить аміно-реактивний N-гідроксисукцинімідний ефір (NHS), який швидко реагує з будь-якою молекулою, яка містить первинний амін, утворюючи, таким чином, стійкий амідний зв'язок. Трансформація in planta і скринування стійких рослин T1: Була отримана суспензія з підсумковим об'ємом 250-500 мл, яка містить наночастинки, пептиди хоумінгу і розчин кон'югата плазмідної ДНК (NHpD), і рослини Arabidopsis (головним чином незрілі квіткові групи з деякою кількістю запліднених стручків) використовували для подальшої обробки. До занурення рослин, до розчину кон'югатів NHpD був доданий Silwet L-77 до концентрації 0,05 % (250 мкл/500 мл) – 0,005 %, і розчин був добре перемішаний. Надземні частини рослини були занурені в розчин кон'югатів NHpD на 2-30 секунд з обережним перемішуванням. Оброблені рослини тримали під куполом або покриттям протягом 16-24 годин при 22-24 °C. Рослини були перенесені в камеру Conviron для надавання можливості росту рослин до дозрівання і збору насіння. Для 17 UA 113046 C2 5 10 15 20 25 скринування загального врожаю насіння від рослин T0 використовувалися лотки для селекції (розміром 10,5×21×1 дюймів), які містять приблизно 10000 насінин на лоток. Для гарантії того, що обприскування при селекції виконано коректно, використовували дві групи контролю: рослини Col-0 як негативний контроль трансформанту, і рослини Columbia Col-0 дикого типу, збагачені гомозиготним насінням для селектованого маркера PAT (фосфінотрицин-Nацетилтрансферази), як позитивний контроль трансформанту. Насіння, з метою синхронізації, було стратифіковано в розчині агару з концентрацією 0,1 % (мас./об.) протягом 48 годин до висівання. Для забезпечення щільності посіву 10000 насінин на лоток для селекції, до розчину агару з концентрацією 0,1 % (мас./об.) було додано 200 мг насіння, і розчин був підданий перемішуванню обертанням доти, поки насіння не було рівномірно суспендоване. Потім, стратифіковане насіння висівали на лотках для селекції, які були заповнені ґрунтовою сумішшю Sunshine LP5 і одержували для зволоження розчин Hоаglаnd's шляхом подачі розчину знизу. Для ефективного обприскування при селекції рослин важливо, щоб 40 мл суспендованого насіння було рівномірно висіяне в лоток для селекції. Після висівання, кожен лоток для селекції був закритий куполом, і рослини вирощувалися для селекції в умовах, зазначених вище. Купола були вилучені приблизно через 5 днів після висівання. Розсаду піддавали обприскуванню через п'ять днів після висівання і ще раз через десять днів після висівання, шляхом обприскування паростків 0,2 %-им (мас./об.) розчин (20 мкл/10 мл dН2O) глюфосинату амонію (гербіцид Liberty® від Bayer Crop Sciences), при об'ємі обприскування 10 мл на лоток (703 л/га), з використанням наконечника DeVilbiss для розпилення стисненим повітрям, щоб забезпечити доставляння ефективної кількості глюфосинату при нормі 280 г глюфосинату на гектар на одне обприскування (GPA). Кількість гербіциду Liberty® для готування розраховувалося в такий спосіб: (703 л за об'ємом, який розпорошується, на гектар=280 GPA). (280 г (за кислотою) на гектар) x (1 га/703 л)×(1 л/200 м глюфосинату (за кислотою)=0,20 %-ий розчин (або 20 мкл/10 мл). 10 мл розчину для кожного лотка, який піддається обприскуванню, переносили піпеткою в сцинтиляціний флакон на 20 мл. Обприскування виконували з використанням горизонтального і вертикального переміщення. Рослини, стійкі до гербіциду, були ідентифіковані через чотири - сім днів після другого обприскування. 30 18 UA 113046 C2 19 UA 113046 C2 ФОРМУЛА ВИНАХОДУ 5 10 15 20 25 30 35 40 45 50 55 1. Спосіб введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину рослини, яка має клітинну стінку, за яким: надають клітину рослини, яка має клітинну стінку; покривають наночастинку на основі позитивно зарядженого напівпровідника поліетиленгліколем; покривають наночастинку на основі позитивно зарядженого напівпровідника лінійною молекулою нуклеїнової кислоти, яка представляє інтерес; приводять клітину, яка має клітинну стінку, і наночастинку на основі позитивно зарядженого напівпровідника, що має покриття, в контакт одна з одною; і надають можливість поглинання наночастинки на основі позитивно зарядженого напівпровідника і лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в клітину, яка містить клітинну стінку. 2. Спосіб за п. 1, за яким додатково надають можливість поглинання наночастинки на основі позитивно зарядженого напівпровідника в компартмент клітини рослини, яка містить клітинну стінку. 3. Спосіб за п. 2, за яким додатково покривають наночастинку на основі позитивно зарядженого напівпровідника білком, який націлює на субклітинний компартмент. 4. Спосіб за п. 3, де компартмент вибраний із групи, яка складається з цитозолю, ядра, тонопластів, пластиди, етіопласту, хромопласту, лейкопласту, елайопласту, протеїнопласту, амілопласту, хлоропласту і просвіту подвійної мембрани. 5. Спосіб за п. 1, де клітина рослини, яка має клітинну стінку, являє собою клітину рослини з комерційних видів зернових рослин. 6. Спосіб за п. 5, де клітина рослини, яка має клітинну стінку, вибрана з групи, яка складається з клітин тютюну, моркви, маїсу, каноли, ріпаку, бавовнику, пальми, арахісу, сої, Oryza sp., Arabidopsis sp., Ricinus sp. і цукрової тростини. 7. Спосіб за п. 6, де клітина рослини походить із тканини, вибраної з групи, яка складається із зародка, меристеми, калусу, пилку, листя, пиляків, коріння, кінчиків коріння, квіток, насіння, стручків і стебел. 8. Спосіб за п. 1, де клітина рослини, яка має клітинну стінку, являє собою культивовану клітину. 9. Спосіб за п. 1, де напівпровідникова наночастинка на основі позитивно зарядженого напівпровідника являє собою квантову точку. 10. Спосіб за п. 1, за яким додатково проводять дериватизацію поверхні наночастинки на основі позитивно зарядженого напівпровідника. 11. Спосіб за п. 1, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, містить послідовність нуклеїнової кислоти, вибрану з групи, яка складається з молекул ДНК, РНК, RNAi і генів. 12. Спосіб за п. 11, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, містить ген. 13. Спосіб за п. 12, де ген являє собою ген чужорідного білка, агрономічний ген або маркерний ген. 14. Спосіб за п. 1, де лінійна молекула нуклеїнової кислоти, яка представляє інтерес, отримана шляхом розщеплення молекули нуклеїнової кислоти нуклеазою. 15. Спосіб за п. 14, де молекула нуклеїнової кислоти, розщеплена нуклеазою, вибрана з групи, яка складається з плазміди, косміди, штучних хромосом, штучних хромосом дріжджів і штучних хромосом бактерій. 16. Спосіб за п. 11, за яким додатково відбирають клітини, у які стабільно інтегрована молекула, яка представляє інтерес. 17. Спосіб за п. 16, де відібрані клітини є регенерованими клітинами. 18. Спосіб за п. 17, за яким додатково проводять регенерацію рослини з регенерованих клітин. 19. Спосіб введення лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в рослинний матеріал, за яким: надають рослинний матеріал, де рослинний матеріал вибраний із групи, яка включає клітини рослини, тканини рослини і рослини; покривають наночастинку на основі позитивно зарядженого напівпровідника поліетиленгліколем; покривають наночастинку на основі позитивно зарядженого напівпровідника лінійною молекулою нуклеїнової кислоти, яка представляє інтерес; 20 UA 113046 C2 5 приводять клітину рослини, яка має клітинну стінку, і наночастинки, на основі позитивно зарядженого напівпровідника, що має покриття, в контакт одна з одною; і надають можливість поглинання наночастинки і лінійної молекули нуклеїнової кислоти, яка представляє інтерес, в рослинний матеріал. 20. Спосіб за п. 19, де рослинний матеріал являє собою тканину рослини, вибрану з групи, яка складається з зародка, тканини меристеми, калусу, пилку, листя, пиляків, коріння, кінчиків коріння, квіток, насіння, стручків і стебел. 21 UA 113046 C2 22 UA 113046 C2 Комп’ютерна верстка Г. Паяльніков Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 23
ДивитисяДодаткова інформація
Назва патенту англійськоюLinear dna molecule delivery using pegylated quantum dots for stable transformation in plants
Автори англійськоюSamboju, Narasimha Chary, Yau, Kerrm Y., Burroughs, Frank G., Samuel, Jayakumar Pon, Webb, Steven R.
Автори російськоюСамбоджу Нарасимха Чари, Йо Керрм Й., Берроуз Фрэнк Дж., Сэмьюел Джаякумар Пон, Уэбб Стивен Р.
МПК / Мітки
МПК: C12N 15/87, C12N 15/82, B82B 3/00
Мітки: стінку, застосуванням, наночастинок, спосіб, нуклеїнової, клітину, яка, пегильованих, має, кислоти, клітинну, лінійної, рослини, молекули, напівпровідникових, введення
Код посилання
<a href="https://ua.patents.su/25-113046-sposib-vvedennya-linijjno-molekuli-nuklenovo-kisloti-v-klitinu-roslini-yaka-maeh-klitinnu-stinku-iz-zastosuvannyam-pegilovanikh-napivprovidnikovikh-nanochastinok.html" target="_blank" rel="follow" title="База патентів України">Спосіб введення лінійної молекули нуклеїнової кислоти в клітину рослини, яка має клітинну стінку, із застосуванням пегильованих напівпровідникових наночастинок</a>
Спосіб введення молекули функціоналізованої лінійної нуклеотидної касети у клітину, що містить клітинну стінку

Номер патенту: 112758
Опубліковано: 25.10.2016
Автори: Семьюел Джаякумар Пон, Йо Керрм Й., Самбоджу Нарасімха Чарі, Уебб Стівен Р., Берроуз Френк Дж.
МПК: B82B 3/00, C12N 15/82, C12N 15/87
Мітки: клітинну, функціоналізованої, спосіб, касеті, містить, введення, лінійної, молекули, стінку, клітину, нуклеотидної
Формула / Реферат:
1. Спосіб введення молекули функціоналізованої лінійної нуклеотидної касети, що представляє інтерес, у клітину рослини, що має клітинну стінку, причому спосіб включає в себе:надання рослинної клітини, що має клітинну стінку;покривання наночастинки на основі квантової точки молекулою функціоналізованої лінійної нуклеотидної касети, що представляє інтерес, де наночастинка на основі квантової точки містить функціональну групу, яка...
Спосіб визначення послідовності молекули нуклеїнової кислоти (варіанти) та набір для використання при визначенні послідовності нуклеїнової кислоти

Номер патенту: 48119
Опубліковано: 15.08.2002
Автор: Дрманач Радоє
МПК: G01N 37/00, C12N 15/09, C12Q 1/68, G01N 33/53
Мітки: молекули, визначення, варіанти, кислоти, використання, набір, спосіб, послідовності, визначенні, нуклеїнової
Формула / Реферат:
1. Способ определения последовательности молекулы нуклеиновой кислоты, отличающийся тем, что проводят гибридизацию молекулы с комплементарными последовательностями олигонуклеотидов из двух наборов малых олигонуклеотидных зондов известной последовательности, причем первый набор зондов прикреплен к твердой подложке, а второй набор зондов представляет собой меченые зонды, находящиеся в растворе, создают ковалентные связи между гибридизированным...
Молекули нуклеїнової кислоти, які кодують wrinkled1-подібні поліпептиди

Номер патенту: 89960
Опубліковано: 25.03.2010
Автори: Міттендорф Фолькер, Бхатт Гаріма, Хертель Хайко, Шенк Карін Дж.
МПК: C12N 9/02, C07H 21/04, A01H 1/06
Мітки: кислоти, кодують, молекули, wrinkled1-подібні, поліпептиди, нуклеїнової
Формула / Реферат:
1. Виділена нуклеїнова кислота, що кодує протеїн ліпідного метаболізму (LMP), яка містить полінуклеотидну послідовність, вибрану з групи, яка включає:a) полінуклеотидну послідовність, як визначено в SEQ ID NО:4, SEQ ID NО:5, SEQ ID NО:7, SEQ ID NО:8, SEQ ID NО:10, SEQ ID NО:11, SEQ ID NО:13, SEQ ID NО:14, SEQ ID NО:16, SEQ ID NО:17, SEQ ID NО:19, SEQ ID NО:20, SEQ ID NО:22, SEQ ID NО:23, SEQ ID NО:25, SEQ ID NО:26, SEQ ID NО:28, SEQ...
Невірусний вектор для введення нуклеїнової кислоти та спосіб його одержання, конденсований комплекс поліпептид/нуклеїнова кислота та спосіб вбудовування нуклеїнової кислоти, що представляє інтерес, в еукаріотич

Номер патенту: 77393
Опубліковано: 15.12.2006
Автори: Меттьюз Девід, Руссель Віллі, Пєрузель Ерік, Тагава Тошіакі, Міллєр Ендрю Девід, Мюррей Карл, Менвелл Мішель, Алтон Ерік
МПК: C12N 15/34, C07K 14/075, A61P 25/28, C12N 15/88, A61K 38/00, C12N 15/09, A61K 9/127, A61P 25/00, A61K 48/00, A61P 43/00, A61P 35/02, A61P 35/00, A61K 47/48
Мітки: інтерес, одержання, спосіб, еукаріотич, нуклеїнової, комплекс, невірусний, вектор, кислота, введення, кислоти, представляє, вбудовування, конденсований
Формула / Реферат:
1. Невірусний вектор для введення нуклеїнової кислоти, який містить конденсований комплекс поліпептид/нуклеїнова кислота і катіонний ліпід, де комплекс включає(а) нуклеотидну послідовність (NOI), що представляє інтерес; і(б) один або декілька аденовірусних поліпептидів або їх похідних, які упаковують нуклеїнову кислоту, де поліпептиди або їх похідні (І) мають здатність зв'язуватися з NOI; і (II) мають здатність конденсувати NOI;...
Спосіб одержання розчинних цукрів з генетично модифікованих рослин, що експресують ферменти, деградуючі клітинну стінку

Номер патенту: 112643
Опубліковано: 10.10.2016
Автори: Чжан Дунчен, Бугрі Олєг, Рааб Р. Майкл
МПК: C12N 9/42, C12N 15/82, C12N 9/24, C10L 5/44, C12P 19/14
Мітки: стінку, одержання, експресують, генетично, модифікованих, цукрів, ферменти, спосіб, рослин, деградуючі, розчинних, клітинну
Формула / Реферат:
1. Спосіб одержання розчинних цукрів з одержаного способами генної інженерії рослинного матеріалу, який включає:попередню обробку шляхом змішування одержаного способами генної інженерії рослинного матеріалу з перетворюючим у пульпу складом, який включає бісульфіт амонію і карбонат амонію, з утворенням суміші з рН в діапазоні від 5,0 до 10,0, де одержаний способами генної інженерії рослинний матеріал включає першу полінуклеотидну...
Попередній патент: Система та спосіб освітлення багатоярусних кліткових батарей для несучок
Наступний патент: Спосіб електростартерного запуску двигуна внутрішнього згоряння та пристрій для його здійснення
Випадковий патент: Спосіб прогнозування гематологічної токсичності хіміотерапії у хворих на рак грудної залози