Спосіб визначення зиготності bm3 мутантного алеля і алеля comt дикого типу з використанням тканини рослини кукурудзи
Номер патенту: 113613
Опубліковано: 27.02.2017
Автори: ван Опдорп Натан, Вей Чень, Чаннабасаварадхя Чандра-Шекара, Кумпатла Сіва П.

























Формула / Реферат
1. Спосіб визначення зиготності bm3 мутантного алеля і алеля COMT дикого типу з використанням тканини рослини кукурудзи, при цьому спосіб включає в себе:
одержання зразка ізольованої геномної ДНК з тканини рослини кукурудзи;
здійснення контакту в умовах високої жорсткості ізольованої геномної ДНК з
першою молекулою нуклеїнової кислоти, що має нуклеотидну послідовність, яка містить від 10 до 35 суміжних нуклеотидів комплементу bm3 полінуклеотиду з послідовністю SEQ ID NO: 10, де перша молекула нуклеїнової кислоти, не здатна гібридизуватися з послідовністю SEQ ID NO: 11 в умовах високої жорсткості; і
другою молекулою нуклеїнової кислоти, що має нуклеотидну послідовність, яка містить від 10 до 35 суміжних нуклеотидів комплементу COMT полінуклеотиду з послідовністю SEQ ID NO: 11, де друга молекула нуклеїнової кислоти не здатна гібридизуватися з послідовністю SEQ ID NO: 10 в умовах високої жорсткості; і
виявлення гібридизації між першою і другою молекулою нуклеїнової кислоти і геномної ДНК, тим самим визначаючи зиготність bm3 мутантного алеля і алеля COMT дикого типу в геномній ДНК.
2. Спосіб за п. 1, де контактування виділеної геномної ДНК з першою і другою молекулою нуклеїнової кислоти в умовах високої жорсткості включає контактування виділеної геномної ДНК з
(а) молекулою нуклеїнокислотного праймера, що має нуклеотидну послідовність, яка ідентична комплементу першого bm3 полінуклеотиду, що складається з від 10-35 суміжних нуклеотидів послідовності SEQ ID NO: 10; і
(b) молекулою нуклеїнокислотного праймера, що має нуклеотидну послідовність, яка ідентична комплементу другого іншого bm3 полінуклеотиду, що складається з від 10-35 суміжних нуклеотидів послідовності SEQ ID NO: 10, де спосіб додатково включає контактування виділеної геномної ДНК з
(с) молекулою нуклеїнокислотного праймера, що має нуклеотидну послідовність, яка ідентична комплементу першого COMT полінуклеотиду, що складається з від 10-35 суміжних нуклеотидів послідовності SEQ ID NO: 11; і
(d) молекулою нуклеїнокислотного праймера, що має нуклеотидну послідовність, яка ідентична комплементу другого іншого COMT полінуклеотиду, що складається з від 10-35 суміжних нуклеотидів послідовності SEQ ID NO: 11;
де спосіб додатково включає:
ампліфікацію від праймерів (а) і (b) нуклеотидної послідовності між першим і другим bm3 полінуклеотидом, якщо виділена геномна ДНК містить перший і другий bm3 полінуклеотид; і
ампліфікацію від праймерів (с) і (d) нуклеотидної послідовності між першим і другим COMT полінуклеотидом, якщо виділена геномна ДНК містить перший і другий COMT полінуклеотид, де реакційна суміш для ампліфікації містить:
першу молекулу нуклеїнокислотного зонда, яка помічена першим репортером, що має нуклеотидну послідовність, яка є комплементом від 10-35 суміжних нуклеотидів ампліфікованої нуклеотидної послідовності геномної ДНК між комплементом праймера (а) і комплементом праймера (b), де перший нуклеїнокислотний зонд не здатний гібридизуватися з SEQ ID NO: 11 в умовах високої жорсткості, і
другу молекулу нуклеїнокислотного зонда, яка помічена другим іншим репортером, що має нуклеотидну послідовність, яка є комплементом від 10-35 суміжних нуклеотидів ампліфікованої нуклеотидної послідовності геномної ДНК між комплементом праймера (с) і комплементом праймера (d), де другий нуклеїнокислотний зонд не здатний гібридизуватися з SEQ ID NO: 10 в умовах високої жорсткості; і
де визначення зиготності bm3 мутантного алеля і алеля COMT дикого типу в ізольованій геномній ДНК включає виявлення рівнів першого і другого репортерів.
3. Спосіб за п. 2, в якому перший репортер і другий репортер є флуоресціюючими барвниками з відмінними спектрами збудження/емісії.
4. Спосіб за п. 3, в якому першим репортером є FAM, а другим репортером є VIC.
5. Спосіб за п. 1, в якому молекула нуклеїнокислотного праймера згідно з (а), (b), (c) або (d) має довжину від 15 до 30 нуклеотидів.
6. Спосіб за п. 1, в якому молекула нуклеїнокислотного праймера згідно з (а) вибрана з групи, яка складається з послідовностей SEQ ID NO: 4, SEQ ID NO: 5 і SEQ ID NO: 6.
7. Спосіб за п. 1, в якому перша молекула нуклеїнокислотного праймера помічена першим флуоресціюючим барвником, і друга молекула нуклеїнокислотного праймера помічена другим флуоресціюючим барвником зі спектром збудження/емісії, який відрізняється від спектра збудження/емісії першого флуоресціюючого барвника.
8. Спосіб за п. 7, в якому або перша, або друга молекула нуклеїнової кислоти помічена FAM, і інша молекула нуклеїнової кислоти помічена VIC.
9. Спосіб за п. 2, в якому реакція ампліфікації є ПЛР-реакцією.
10. Спосіб за п. 2, в якому реакція ампліфікації є реакцією, що не основана на ПЛР.
11. Спосіб надійної і передбачуваної інтрогресії ознаки низького вмісту лігніну в зародкову плазму рослини, при цьому вказаний спосіб включає в себе:
схрещування рослини, що має bm3 мутантний алель з іншою рослиною;
одержання зразка ізольованої геномної ДНК з рослини-потомка, одержаного при схрещуванні;
здійснення контакту ізольованої геномної ДНК з молекулою нуклеїнової кислоти, що має нуклеотидну послідовність, яка ідентична від 10 до 35 суміжних нуклеотидів комплементу bm3 полінуклеотиду з послідовністю SEQ ID NO: 10, де молекула нуклеїнової кислоти не здатна гібридизуватися з SEQ ID NO: 11 в умовах високої жорсткості; і
відбір потомства від схрещування, яке має bm3 мутантний алель, через розмноження рослини, з якої одержано зразок, що містить геномну ДНК, яка гібридизується в умовах високої жорсткості з молекулою нуклеїнової кислоти, з одержанням таким чином генетично сконструйованої рослини, при цьому шляхом інтрогресії ознака низького вмісту лігніну введена в зародкову плазму генетично сконструйованої рослини.
Текст
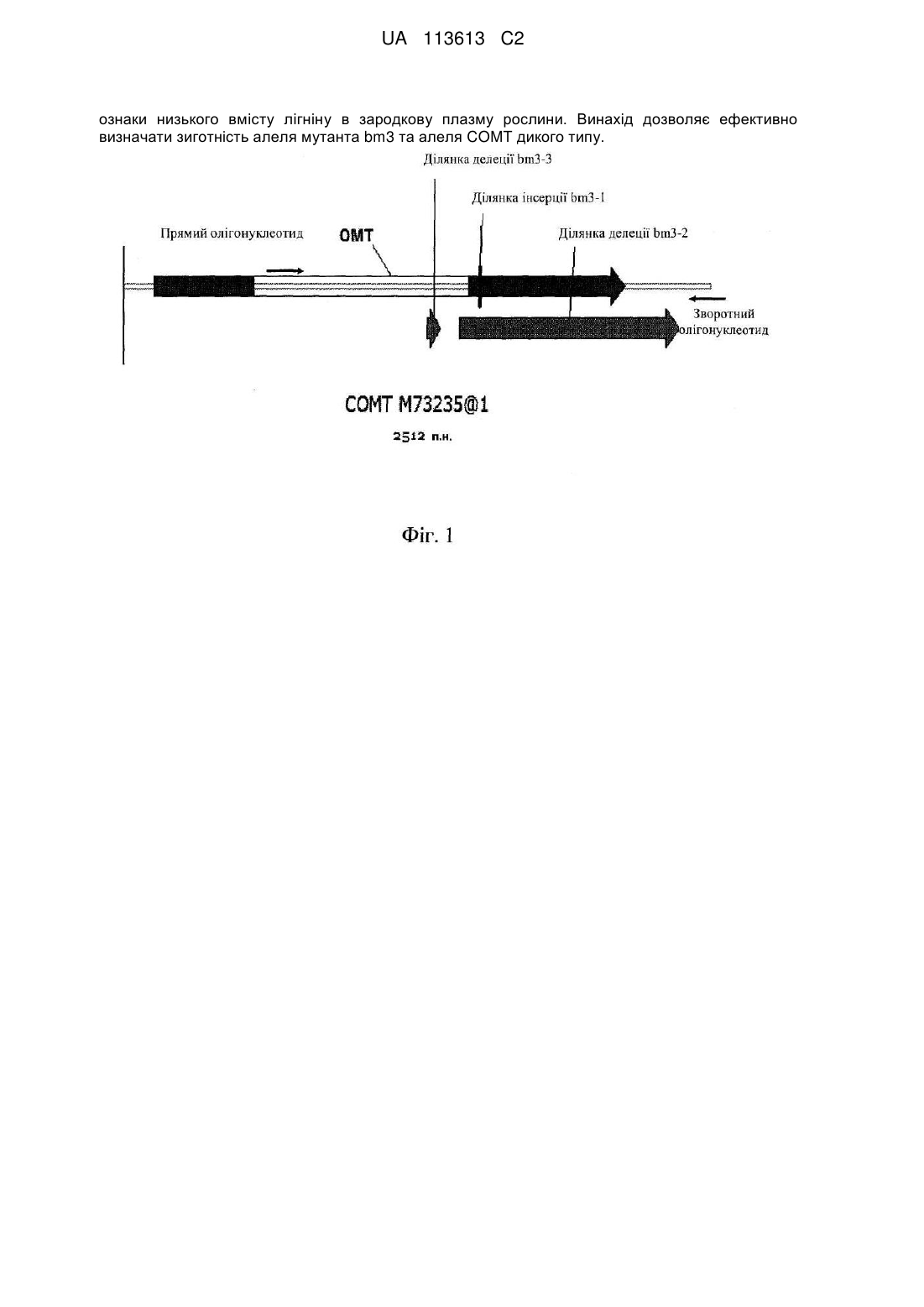
Реферат: Даний винахід пропонує удосконалення процесу селекції кукурудзи, зокрема стосується способу визначення зиготності алеля мутанта bm3 та алеля COMT дикого типу з використанням тканин рослини кукурудзи. Також запропоновано спосіб надійної і передбачуваної інтрогресії UA 113613 C2 (12) UA 113613 C2 ознаки низького вмісту лігніну в зародкову плазму рослини. Винахід дозволяє ефективно визначати зиготність алеля мутанта bm3 та алеля COMT дикого типу. UA 113613 C2 5 10 15 20 25 30 35 40 45 50 55 60 Дана заявка заявляє пріоритет по даті подачі попередньої заявки на видачу патенту США з реєстраційним номером 61/334073, поданої 12 травня 2010, "Use of brown midrib-3 gene specific markers in maize for trait introgression". ГАЛУЗЬ ТЕХНІКИ, ДО ЯКОЇ НАЛЕЖИТЬ ВИНАХІД Даний опис, загалом, стосується селекції рослин. Пропонуються способи визначення зиготності рослин, що містять мутації коричневої середньої жилки (brown midrib 3, bm3). Способи, розкриті в описі, крім того, застосовні для удосконалення способу селекції BMRвмісних ліній рослин. РІВЕНЬ ТЕХНІКИ Лігніни є універсальними компонентами в рослинах, які поперечно зв'язуються з вуглеводами, такими як геміцелюлози, в клітинній стінці. Полімери лігніну знижують розщеплення волокон у жуйних, і ступінь лігніфікації може бути зворотно пропорційним перетравлюваності кормових культур. Cherney et al. (1991) Adv. Agron. 46:157-98. Кукурудза, що містить мутацію коричневої середньої жилки brown midrib (BMR) має червонувато-коричневу пігментацію середньої жилки листа, що асоційовано зі значно зниженим вмістом лігніну, зміненим складом лігніну і поліпшеною перетравлюваністю. Було ідентифіковано щонайменше чотири незалежних мутації BMR у кукурудзи. Kuc et al. (1968) Phytochemistry 7:1435-6. У випадку всіх таких мутацій, названих "bm1, bm2, bm3 і bm4", є знижений вміст лігніну в порівнянні з контрольною кукурудзою. Мутації bm3 включають інсерції (bm3-1), делеції (bm3-2) та інсерції/делеції (bm3-3) в гені О-метилтрансферази кофеїнової кислоти (COMT, наприклад, GenBank Accession No. M73235). Morrow et al. (1997) Mol. Breeding 3:351-7; Vignols et al. (1995) Plant Cell 7:407-16. Ген COMT контролює ферментативні активності, залучені до біосинтезу лігніну. COMT використовує S-аденозилметіонін для трансметилювання кофеїнової кислоти, що приводить до утворення ферулової кислоти. У кінцевому результаті з ферулової кислоти утворюються коніфериловий спирт і синапіловий спирт. Поєднання коніферилового, ферулового і синапілового спиртів в присутності вільних радикалів приводить до продукції лігніну. Мутації Bm3 були охарактеризовані, і вважається, що вони інгібують трансметилювання кофеїнової кислоти внаслідок інактивації генів. Guillet-Claude et al. (2004) Theor. Appl. Genet. 110: 126-35; Piquemal et al. (2002) Plant Physiology 130: 1-11; He et al. (2003) Crop Sci. 43: 2240-51; Morrow et al. (1997) Mol. Breeding 3: 351-7; Vignols et al. (1995) Plant Cell 7: 407-16. Однак не розроблено швидкого способу для специфічного виявлення і тестування зиготності конкретної рослини по локусу bm3. СУТЬ ВИНАХОДУ У даному описі розкриті способи високопродуктивної основаної на ПЛР молекулярної характеристики сортів кукурудзи BMR (наприклад, мутантів bm3), які можуть значно поліпшити процес селекції BMR-вмісних ліній. Розкриті способи визначення зиготності зразка рослинної тканини, і отже рослини, з якої одержаний зразок, за допомогою визначення присутності або відсутності алелей мутанта bm3 і COMT дикого типу. Таким чином, пропонується аналіз зиготності на основі ПЛР з реєстрацією в кінцевій точці з використанням зондів, які працюють за принципом гідролізу (який в даному описі вказаний як аналіз TaqMan®), який специфічно виявляє і тестує стан зиготності в локусі bm3. Розкриті аналізи, в яких використовують біплекси олігонуклеотидів, специфічних до мутанта bm3 і до відповідних послідовностей дикого типу, в одному і тому ж аналізі. КОРОТКИЙ ОПИС КРЕСЛЕНЬ Фіг. 1 включає схематичне представлення гена COMT з мутаціями bm3 і олігонуклеотиди, сконструйовані для часткової ампліфікації гена COMT (~1,8 т. п. н.). На фіг. 2 зображена ампліфікація неповної послідовності гена COMT з дванадцяти ліній bm3 і трьох ліній не-bm3 (ліній дикого типу). ПЛР-продукти візуалізували в 2 % E-Gel® і потім екстрагували, використовуючи 0,8 % E-Gel®, для подальшого клонування у векторі pCR4TOPO®. На фіг. 3 показане вирівнювання консенсусних послідовностей мутантів bm3 і генів COMT дикого типу. Три раніше описаних делеційних/інсерційних мутацій підкреслені. На фіг. 4 включає схематичне представлення гена COMT з праймерами, специфічними до мутації bm3 та інтактного гена COMT. (А) Ген COMT з пунктирними лініями означає делецію bm3. Специфічні для гена COMT праймери знаходяться в межах сайту делеції. (В) Мутантний ген bm3. Специфічні для bm3 олігонуклеотиди знаходяться в місці з'єднання, де прямий і зворотні олігонуклеотиди, які фланкують сайт делеції, і зонд охоплюють делетовану область. Фіг. 5a і 5b містять графіки ПЛР-ампліфікації в реальному часі, де показані відносні одиниці флуоресценції (RFU) для bm3 (a) з використанням FAM і (b) для COMT дикого типу з 1 UA 113613 C2 5 10 15 20 25 30 35 40 45 50 55 60 використанням VIC (замінювали Hex через обмеження приладу для ПЛР в реальному часі). Експонентну фазу ампліфікації спостерігали з 22 по 36 цикл як у випадку гена bm3, так і у випадку гена COMT дикого типу. На фіг. 6 показані визначення генотипів, здійснювані з використанням аналізу, що розрізняє алелі на основі вимірювання у відносних одиницях флуоресценції FAM для алеля 1 (гомозиготний bm3) і VIC (Hex, що замінюється через обмеження приладу) у випадку алеля 2 (гомозиготний COMT дикого типу) в циклі 30. На фіг. 7 показані визначення зиготності bm3, здійснювані з використанням аналізу TaqMan® з реєстрацією в кінцевій точці. Після завершення ПЛР і реєстрації флуоресценції одержували графік розподілу як описано нижче: SOB1 = сигнал вище фону FAM (сигнал зразка вище середнього фонового сигналу при 535 нм), SOB2 = сигнал вище фону VIC (сигнал зразка вище середнього фонового сигналу при 560 нм). Визначення генотипу здійснювали, використовуючи SOB1/SOB2
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of brown midrib-3 gene specific markers in maize for trait introgression
Автори англійськоюWei, Chen, van Opdorp, Nathan, Channabasavaradhya, Chandra-Shekara, Kumpatla, Siva P.
Автори російськоюВей Чень, ван Опдорп Натан, Чаннабасаварадхя Чандра-Шекара, Кумпатла Сива П.
МПК / Мітки
МПК: A01H 5/00, C12N 15/29, C12Q 1/68
Мітки: алеля, дикого, визначення, тканини, кукурудзи, типу, використанням, спосіб, зиготності, мутантного, рослини
Код посилання
<a href="https://ua.patents.su/27-113613-sposib-viznachennya-zigotnosti-bm3-mutantnogo-alelya-i-alelya-comt-dikogo-tipu-z-vikoristannyam-tkanini-roslini-kukurudzi.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення зиготності bm3 мутантного алеля і алеля comt дикого типу з використанням тканини рослини кукурудзи</a>
Спосіб визначення зиготності рослини кукурудзи, що містить об’єкт das-40278-9 aad-1 кукурудзи

Номер патенту: 109882
Опубліковано: 26.10.2015
Автори: Чжоу Нін, Грін Томас Уілльям, Цуй Юнсінь Корі, Новак Стефен
МПК: C12N 15/10, C12Q 1/68, A01H 1/04, C12N 15/82
Мітки: спосіб, кукурудзи, das-40278-9, зиготності, об`єкт, aad-1, визначення, рослини, містить
Формула / Реферат:
1. Спосіб визначення зиготності рослини кукурудзи, що містить об'єкт DAS-40278-9 AAD-1 кукурудзи, що містить SEQ ID NO:1, де вказаний об'єкт містить трансгенну конструкцію, що містить ген AAD-1, де вказана трансгенна конструкція фланкована 5¢-фланкуючою геномною ДНК кукурудзи і 3¢-фланкуючою геномною ДНК кукурудзи, де вказана 5'-фланкуюча геномна ДНК кукурудзи складається із залишків 1-1873 SEQ ID NO:1 і вказана 3'-фланкуюча...
Спосіб визначення зиготності події у рослини сої, що включає aad-12-подію pdab4468-0416 у сої

Номер патенту: 109644
Опубліковано: 25.09.2015
Автори: Чжоу Нін, Новак Стефен, Грін Томас, Цуй Юнсінь Корі
МПК: C12N 15/82, C07H 21/04, C12Q 1/68, A01H 1/04, C12P 19/34
Мітки: спосіб, pdab4468-0416, визначення, aad-12-подію, включає, події, сої, рослини, зиготності
Формула / Реферат:
1. Спосіб визначення зиготності події у рослини сої, що включає AAD-12-подію pDAB4468-0416 у сої, яка включає SEQ ID NO: 1, де вказана подія містить трансгенну конструкцію, що включає ген AAD-12, і вказана трансгенна конструкція фланкована 5'-фланкуючою геномною ДНК сої і 3'-фланкуючою геномною ДНК сої, де вказана 5'-фланкуюча ДНК включає залишки 1-2730 SEQ ID NO: 1, вказана 3'-фланкуюча ДНК включає залишки 9122-10212 SEQ ID NO: 1, і вказана...
Спосіб визначення типу взаємної дії факторів середовища на рослини

Номер патенту: 23658
Опубліковано: 11.06.2007
Автори: Шемет Сергій Анатолійович, Феденко Володимир Савелійович
МПК: A01G 7/00
Мітки: рослини, дії, факторів, середовища, визначення, взаємної, спосіб, типу
Формула / Реферат:
Спосіб визначення типу взаємної дії факторів середовища на рослини, що включає тестування об'єкта за дії окремих факторів та їх комбінацій в діапазоні доз, встановлення дозових змін тестового показника, діагностику ефекту взаємодії на основі співставлення залежностей показника за умов комбінованого впливу та його відсутності, який відрізняється тим, що встановлюють співвідношення інкременту функції тестового показника при взаємодії факторів...
Спосіб визначення типу цитоплазми кукурудзи

Номер патенту: 73707
Опубліковано: 10.10.2012
Автори: Сліщук Георгій Іванович, Сиволап Юрій Михайлович, Волкова Наталія Едуардівна
МПК: A01H 5/00
Мітки: кукурудзи, спосіб, типу, визначення, цитоплазми
Формула / Реферат:
Спосіб визначення типу цитоплазми кукурудзи включає ПЛР-аналіз регіонів мітохондріонів у зразках кукурудзи, який відрізняється тим, що проводять ідентифікацію химерної відкритої рамки зчитування Orf355, atp6/atp9-регіону та химерного гену T-urf мітохондріонів S-, О- та Т-типу цитоплазми, відповідно.
Прилад для дослідження закономірностей згину та зламу складових рослини кукурудзи

Номер патенту: 73813
Опубліковано: 10.10.2012
Автори: Ракул Олександр Іванович, Пилип Володимир Єгорович, Бондаренко Олександр Володимирович
МПК: G01N 3/08
Мітки: закономірностей, складових, зламу, кукурудзи, дослідження, прилад, згину, рослини
Формула / Реферат:
1. Прилад для дослідження закономірностей згину та зламу складових рослини кукурудзи, який складається з платформи, стійок, опори, перемички з гвинтовим механізмом, динамометру та згинаючого робочого органу, який відрізняється тим, що опори мають П-подібну форму та встановлені в отворах платформи прямокутної форми, а наявність тарировочної шкали та гвинтового механізму опор дає можливість встановлення останніх на необхідній для досліджень...
Попередній патент: Сполуки, композиції та способи для зниження або усунення гіркого смаку
Наступний патент: Спосіб експлуатації кисневої продувальної фурми в металургійній ємності і вимірювальна система для визначення використовуваних при цьому сигналів вимірювань
Випадковий патент: Безредукторна вертикальноосьова вітроенергетична установка