Рослина пшениці з підвищеною резистентністю до імідазолінонових гербіцидів








Формула / Реферат
1. Рослина пшениці, яка містить множинні IMI нуклеїнові кислоти, які походять із різних геномів, причому рослина пшениці має підвищену резистентність до імідазолінонового гербіциду порівняно з різновидами рослини дикого типу, і де принаймні одна з множинних IMI нуклеїнових кислот вибрана із групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
2. Рослина пшениці за п. 1, яка відрізняється тим, що множинні IMI нуклеїнові кислоти вибирають із групи, яка складається з Imi1 нуклеїнової кислоти, Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
3. Рослина пшениці за п. 1, яка відрізняється тим, що принаймні одна з множинних IMI нуклеїнових кислот кодує білок, який має мутацію в консервативній амінокислотній послідовності, вибраній з групи, яка складається з Домену A, Домену B, Домену C, Домену D та Домену E.
4. Рослина пшениці за п. 3, яка відрізняється тим, що консервативна амінокислотна послідовність є Доменом E.
5. Рослина пшениці за п. 4, яка відрізняється тим, що мутація забезпечує заміщення серину аспарагіном в IMI білку порівняно з білком AHAS дикого типу.
6. Рослина пшениці за п. 1, яка відрізняється тим, що одна з IMI нуклеїнових кислот включає полінуклеотидну послідовність, яка кодує поліпептид SEQ ID NO:2.
7. Рослина пшениці за п. 1, яка відрізняється тим, що містить дві IMI нуклеїнові кислоти.
8. Рослина пшениці за п. 7, яка відрізняється тим, що містить Imi1 нуклеїнову кислоту та Imi3 нуклеїнову кислоту.
9. Рослина пшениці за п. 8, яка відрізняється тим, що Imi1 нуклеїнова кислота включає полінуклеотидну послідовність, яка кодує поліпептид SEQ ID NO:2.
10. Рослина пшениці за п. 1, яка відрізняється тим, що містить три IMI нуклеїнові кислоти.
11. Рослина пшениці за будь-яким з пп. 1-5, яка відрізняється тим, що рослина є трансгенною.
12. Рослина пшениці за будь-яким з пп. 1-5, яка відрізняється тим, що рослина не є трансгенною.
13. Рослина пшениці за п. 12, яка відрізняється тим, що рослина отримана або походить від рослини з номером патентного депонування ATCC PTA-3955.
14. Рослина пшениці за п. 1, яка відрізняється тим, що імідазоліноновий гербіцид вибирають із групи, яка складається з 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-3-хінолінкарбонової кислоти, 5-етил-2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-метилнікотинової кислоти та суміші метил 6-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-м-толуату і метил 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-п-толуату.
15. Рослина пшениці за п. 1, яка відрізняється тим, що імідазоліноновий гербіцид є 5-етил-2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотиновою кислотою.
16. Рослина пшениці за п. 1, яка відрізняється тим, що імідазоліноновий гербіцид є 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)-нікотиновою кислотою.
17. Рослинна частина рослини пшениці за п. 1, яка відрізняється тим, що рослинна частина включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
18. Рослинна клітина рослини пшениці за п. 1, яка відрізняється тим, що рослинна клітина включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
19. Насіння, вироблене рослиною пшениці за п. 1, яке відрізняється тим, що насіння включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
20. Насіння за п. 19, яке відрізняється тим, що це насіння є розведенням гомозигот для підвищеної резистентності до імідазолінонового гербіциду порівняно з насінням сорту рослини пшениці дикого типу.
21. Рослина пшениці , яка містить IMI нуклеїнову кислоту, причому рослина пшениці має підвищену резистентність до імідазолінонового гербіциду порівняно з різновидами рослини дикого типу, і де IMI нуклеїнова кислота вибрана з групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
22. Рослина пшениці за п. 21, яка відрізняється тим, що імідазоліноновий гербіцид вибирають із групи, яка складається з 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-3-хінолінкарбонової кислоти, 5-етил-2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)-нікотинової кислоти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-метилнікотинової кислоти та суміші метил 6-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-м-толуату і метил 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-п-толуату.
23. Рослина пшениці за п. 21, яка відрізняється тим, що імідазоліноновий гербіцид є 5-етил-2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-нікотиновою кислотою.
24. Рослина пшениці за п. 21, яка відрізняється тим, що імідазоліноновий гербіцид є 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)-нікотиновою кислотою.
25. Рослинна частина рослини пшениці за п. 21, яка відрізняється тим, що рослинна частина включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
26. Рослинна клітина рослини пшениці за п. 21, яка відрізняється тим, що рослинна клітина включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
27. Насіння, вироблене рослиною пшениці за п. 21, яке відрізняється тим, що насіння включає принаймні одну з IMI нуклеїнових кислот, вказаних в а) - г).
28. Насіння за п. 27, яке відрізняється тим, що це насіння є розведенням гомозигот для підвищеної резистентності до імідазолінонового гербіциду порівняно з насінням сорту рослини пшениці дикого типу.
29. Рослина пшениці за п. 21, яка відрізняється тим, що рослина є трансгенною.
30. Рослина пшениці за п. 21, яка відрізняється тим, що рослина не є трансгенною.
31. Виділена IMI нуклеїнова кислота, яка відрізняється тим, що нуклеїнова кислота включає полінуклеотид, вибраний з групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
32. Виділена IMI нуклеїнова кислота за п. 31, яка відрізняється тим, що ця нуклеїнова кислота включає полінуклеотид SEQ ID NO:1.
33. Виділена IMI нуклеїнова кислота за п. 31, яка відрізняється тим, що ця нуклеїнова кислота включає полінуклеотид, який кодує поліпептид SEQ ID NO:2.
34. Спосіб контролю над бур'янами в оточенні рослин пшениці, який включає нанесення імідазолінонового гербіциду на бур'яни та рослини пшениці, причому рослина пшениці має підвищену резистентність до імідазолінонового гербіциду порівняно з сортом рослини пшениці дикого типу, причому рослина містить множинні IMI нуклеїнові кислоти, причому нуклеїнові кислоти походять із різних геномів, і де принаймні одну з множинних IMI нуклеїнових кислот вибирають із групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
35. Спосіб за п. 34, який відрізняється тим, що множинні IMI нуклеїнові кислоти вибирають із групи, яка складається з Imi1 нуклеїнової кислоти, Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
36. Спосіб за п. 35, який відрізняється тим, що рослина містить Imi1 нуклеїнову кислоту та Imi3 нуклеїнову кислоту.
37. Спосіб контролю над бур'янами в оточенні рослин пшениці, який включає нанесення імідазолінонового гербіциду на бур'яни та рослини пшениці, причому рослина пшениці має підвищену резистентність до імідазолінонового гербіциду порівняно з сортом рослини пшениці дикого типу, і рослина містить IMI нуклеїнову кислоту, вибрану з групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
38. Спосіб за п. 37, який відрізняється тим, що IMI нуклеїнову кислоту вибирають із групи, яка складається з Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
39. Спосіб зміни толерантності рослини до імідазолінонового гербіциду, який включає зміну експресії множинних IMI нуклеїнових кислот, причому нуклеїнові кислоти походять із різних геномів, і де принаймні одну з множинних IMI нуклеїнових кислот вибирають із групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
40. Спосіб за п. 39, який відрізняється тим, що множинні IMI нуклеїнові кислоти вибирають із групи, яка складається з Imi1 нуклеїнової кислоти, Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
41. Спосіб за п. 40, який відрізняється тим, що рослина містить Imi1 нуклеїнову кислоту та Imi3 нуклеїнову кислоту.
42. Спосіб зміни толерантності рослини до імідазолінонового гербіциду, який включає зміну експресії IMI нуклеїнової кислоти, де IMI нуклеїнова кислота вибрана з групи, яка складається з:
a) полінуклеотиду, який включає SEQ ID NO:1,
б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в).
43. Спосіб за п. 42, який відрізняється тим, що IMI нуклеїнову кислоту вибирають із групи, яка складається з Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
44. Спосіб одержання трансгенної рослини з підвищеною резистентністю до імідазолінонового гербіциду, який включає:
a) трансформацію рослинної клітини одним або кількома векторами експресії, які включають множинні IMI нуклеїнові кислоти, причому нуклеїнові кислоти походять із різних геномів, і де принаймні одна з множинних IMI нуклеїнових кислот вибрана з групи, яка складається з:
і) полінуклеотиду, який включає SEQ ID NO:1,
іі) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
ііі) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
іv) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від і) до ііі), та
б) вирощування з рослинної клітини трансгенної рослини з підвищеною резистентністю до імідазолінонового гербіциду порівняно з сортом рослини дикого типу.
45. Спосіб за п. 44, який відрізняється тим, що множинні IMI нуклеїнові кислоти вибирають із групи, яка складається з Imi1 нуклеїнової кислоти, Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
46. Спосіб за п. 45, який відрізняється тим, що рослина містить Imi1 нуклеїнову кислоту та Imi3 нуклеїнову кислоту.
47. Спосіб одержання трансгенної рослини з підвищеною резистентністю до імідазолінонового гербіциду, який включає:
a) трансформацію рослинної клітини вектором експресії, який включає IMI нуклеїнову кислоту, причому ІМІ нуклеїнова кислота вибрана з групи, яка складається з:
і) полінуклеотиду, який включає SEQ ID NO:1,
іі) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2,
ііі) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і
іv) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від і) до ііі), та
б) вирощування з рослинної клітини трансгенної рослини з підвищеною резистентністю до імідазолінонового гербіциду порівняно з сортом рослини дикого типу.
48. Спосіб за п. 47, який відрізняється тим, що IMI нуклеїнову кислоту вибирають із групи, яка складається з Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кислоти.
Текст
1. Рослина пшениці, яка містить множинні IMI нуклеїнові кислоти, які походять із різних геномів, причому рослина пшениці має підвищену резистентність до імідазолінонового гербіциду порівняно з різновидами рослини дикого типу, і де принаймні одна з множинних IMI нуклеїнових кислот вибрана із групи, яка складається з: a) полінуклеотиду, який включає SEQ ID NO:1, 2 (19) 1 3 87261 4 12. Рослина пшениці за будь-яким з пп. 1-5, яка метил-5-оксо-2-імідазолін-2-іл)-нікотинової кисловідрізняється тим, що рослина не є трансгенною. ти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)13. Рослина пшениці за п. 12, яка відрізняється 5-(метоксиметил)-нікотинової кислоти, 2-(4тим, що рослина отримана або походить від росізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5лини з номером патентного депонування ATCC метилнікотинової кислоти та суміші метил 6-(4PTA-3955. ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-м14. Рослина пшениці за п. 1, яка відрізняється тим, толуату і метил 2-(4-ізопропіл-4-метил-5-оксо-2що імідазоліноновий гербіцид вибирають із групи, імідазолін-2-іл)-п-толуату. яка складається з 2-(4-ізопропіл-4-метил-5-оксо-223. Рослина пшениці за п. 21, яка відрізняється імідазолін-2-іл)-нікотинової кислоти, 2-(4-ізопропілтим, що імідазоліноновий гербіцид є 5-етил-2-(44-метил-5-оксо-2-імідазолін-2-іл)-3ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)хінолінкарбонової кислоти, 5-етил-2-(4-ізопропіл-4нікотиновою кислотою. метил-5-оксо-2-імідазолін-2-іл)-нікотинової кисло24. Рослина пшениці за п. 21, яка відрізняється ти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)тим, що імідазоліноновий гербіцид є 2-(4-ізопропіл5-(метоксиметил)-нікотинової кислоти, 2-(44-метил-5-оксо-2-імідазолін-2-іл)-5ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5(метоксиметил)-нікотиновою кислотою. метилнікотинової кислоти та суміші метил 6-(425. Рослинна частина рослини пшениці за п. 21, ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-мяка відрізняється тим, що рослинна частина вклютолуату і метил 2-(4-ізопропіл-4-метил-5-оксо-2чає принаймні одну з IMI нуклеїнових кислот, вкаімідазолін-2-іл)-п-толуату. заних в а) - г). 15. Рослина пшениці за п. 1, яка відрізняється тим, 26. Рослинна клітина рослини пшениці за п. 21, що імідазоліноновий гербіцид є 5-етил-2-(4яка відрізняється тим, що рослинна клітина вклюізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)чає принаймні одну з IMI нуклеїнових кислот, вканікотиновою кислотою. заних в а) - г). 16. Рослина пшениці за п. 1, яка відрізняється тим, 27. Насіння, вироблене рослиною пшениці за п. 21, що імідазоліноновий гербіцид є 2-(4-ізопропіл-4яке відрізняється тим, що насіння включає приметил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)наймні одну з IMI нуклеїнових кислот, вказаних в нікотиновою кислотою. а) - г). 17. Рослинна частина рослини пшениці за п. 1, яка 28. Насіння за п. 27, яке відрізняється тим, що це відрізняється тим, що рослинна частина включає насіння є розведенням гомозигот для підвищеної принаймні одну з IMI нуклеїнових кислот, вказаних резистентності до імідазолінонового гербіциду в а) - г). порівняно з насінням сорту рослини пшениці дико18. Рослинна клітина рослини пшениці за п. 1, яка го типу. відрізняється тим, що рослинна клітина включає 29. Рослина пшениці за п. 21, яка відрізняється принаймні одну з IMI нуклеїнових кислот, вказаних тим, що рослина є трансгенною. в а) - г). 30. Рослина пшениці за п. 21, яка відрізняється 19. Насіння, вироблене рослиною пшениці за п. 1, тим, що рослина не є трансгенною. яке відрізняється тим, що насіння включає при31. Виділена IMI нуклеїнова кислота, яка відрізнянаймні одну з IMI нуклеїнових кислот, вказаних в ється тим, що нуклеїнова кислота включає полінуа) - г). клеотид, вибраний з групи, яка складається з: 20. Насіння за п. 19, яке відрізняється тим, що це a) полінуклеотиду, який включає SEQ ID NO:1, насіння є розведенням гомозигот для підвищеної б) полінуклеотиду, який кодує поліпептид, що резистентності до імідазолінонового гербіциду включає SEQ ID NO:2, порівняно з насінням сорту рослини пшениці диков) полінуклеотиду, який кодує поліпептид, що має го типу. послідовність амінокислот, принаймні на 90 % іде21. Рослина пшениці , яка містить IMI нуклеїнову нтичну до SEQ ID NO:2, причому поліпептид має кислоту, причому рослина пшениці має підвищену заміщення серин на аспарагін в Домені Е, і резистентність до імідазолінонового гербіциду г) полінуклеотиду, комплементарного будь-якому з порівняно з різновидами рослини дикого типу, і де полінуклеотидів від a) до в). IMI нуклеїнова кислота вибрана з групи, яка скла32. Виділена IMI нуклеїнова кислота за п. 31, яка дається з: відрізняється тим, що ця нуклеїнова кислота вклюa) полінуклеотиду, який включає SEQ ID NO:1, чає полінуклеотид SEQ ID NO:1. б) полінуклеотиду, який кодує поліпептид, що 33. Виділена IMI нуклеїнова кислота за п. 31, яка включає SEQ ID NO:2, відрізняється тим, що ця нуклеїнова кислота вклюв) полінуклеотиду, який кодує поліпептид, що має чає полінуклеотид, який кодує поліпептид SEQ ID послідовність амінокислот, принаймні на 90 % ідеNO:2. нтичну до SEQ ID NO:2, причому поліпептид має 34. Спосіб контролю над бур'янами в оточенні розаміщення серин на аспарагін в Домені Е, і слин пшениці, який включає нанесення імідазоліг) полінуклеотиду, комплементарного будь-якому з нонового гербіциду на бур'яни та рослини пшениці, полінуклеотидів від a) до в). причому рослина пшениці має підвищену резис22. Рослина пшениці за п. 21, яка відрізняється тентність до імідазолінонового гербіциду порівнятим, що імідазоліноновий гербіцид вибирають із но з сортом рослини пшениці дикого типу, причому групи, яка складається з 2-(4-ізопропіл-4-метил-5рослина містить множинні IMI нуклеїнові кислоти, оксо-2-імідазолін-2-іл)-нікотинової кислоти, 2-(4причому нуклеїнові кислоти походять із різних геізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-3номів, і де принаймні одну з множинних IMI нуклеїхінолінкарбонової кислоти, 5-етил-2-(4-ізопропіл-4 5 87261 6 нових кислот вибирають із групи, яка складається пресії IMI нуклеїнової кислоти, де IMI нуклеїнова з: кислота вибрана з групи, яка складається з: a) полінуклеотиду, який включає SEQ ID NO:1, a) полінуклеотиду, який включає SEQ ID NO:1, б) полінуклеотиду, який кодує поліпептид, що б) полінуклеотиду, який кодує поліпептид, що включає SEQ ID NO:2, включає SEQ ID NO:2, в) полінуклеотиду, який кодує поліпептид, що має в) полінуклеотиду, який кодує поліпептид, що має послідовність амінокислот, принаймні на 90 % ідепослідовність амінокислот, принаймні на 90 % ідентичну до SEQ ID NO:2, причому поліпептид має нтичну до SEQ ID NO:2, причому поліпептид має заміщення серин на аспарагін в Домені Е, і заміщення серин на аспарагін в Домені Е, і г) полінуклеотиду, комплементарного будь-якому з г) полінуклеотиду, комплементарного будь-якому з полінуклеотидів від a) до в). полінуклеотидів від a) до в). 35. Спосіб за п. 34, який відрізняється тим, що 43. Спосіб за п. 42, який відрізняється тим, що IMI множинні IMI нуклеїнові кислоти вибирають із грунуклеїнову кислоту вибирають із групи, яка склапи, яка складається з Imi1 нуклеїнової кислоти, дається з Imi2 нуклеїнової кислоти та Imi3 нуклеїImi2 нуклеїнової кислоти та Imi3 нуклеїнової киснової кислоти. лоти. 44. Спосіб одержання трансгенної рослини з під36. Спосіб за п. 35, який відрізняється тим, що ровищеною резистентністю до імідазолінонового слина містить Imi1 нуклеїнову кислоту та Imi3 нукгербіциду, який включає: леїнову кислоту. a) трансформацію рослинної клітини одним або 37. Спосіб контролю над бур'янами в оточенні рокількома векторами експресії, які включають мнослин пшениці, який включає нанесення імідазоліжинні IMI нуклеїнові кислоти, причому нуклеїнові нонового гербіциду на бур'яни та рослини пшениці, кислоти походять із різних геномів, і де принаймні причому рослина пшениці має підвищену резистеодна з множинних IMI нуклеїнових кислот вибрана нтність до імідазолінонового гербіциду порівняно з з групи, яка складається з: сортом рослини пшениці дикого типу, і рослина і) полінуклеотиду, який включає SEQ ID NO:1, містить IMI нуклеїнову кислоту, вибрану з групи, іі) полінуклеотиду, який кодує поліпептид, що яка складається з: включає SEQ ID NO:2, a) полінуклеотиду, який включає SEQ ID NO:1, ііі) полінуклеотиду, який кодує поліпептид, що має б) полінуклеотиду, який кодує поліпептид, що послідовність амінокислот, принаймні на 90 % ідевключає SEQ ID NO:2, нтичну до SEQ ID NO:2, причому поліпептид має в) полінуклеотиду, який кодує поліпептид, що має заміщення серин на аспарагін в Домені Е, і послідовність амінокислот, принаймні на 90 % ідеіv) полінуклеотиду, комплементарного будь-якому нтичну до SEQ ID NO:2, причому поліпептид має з полінуклеотидів від і) до ііі), та заміщення серин на аспарагін в Домені Е, і б) вирощування з рослинної клітини трансгенної г) полінуклеотиду, комплементарного будь-якому з рослини з підвищеною резистентністю до імідазополінуклеотидів від a) до в). лінонового гербіциду порівняно з сортом рослини 38. Спосіб за п. 37, який відрізняється тим, що IMI дикого типу. нуклеїнову кислоту вибирають із групи, яка скла45. Спосіб за п. 44, який відрізняється тим, що дається з Imi2 нуклеїнової кислоти та Imi3 нуклеїмножинні IMI нуклеїнові кислоти вибирають із грунової кислоти. пи, яка складається з Imi1 нуклеїнової кислоти, 39. Спосіб зміни толерантності рослини до імідаImi2 нуклеїнової кислоти та Imi3 нуклеїнової кисзолінонового гербіциду, який включає зміну екслоти. пресії множинних IMI нуклеїнових кислот, причому 46. Спосіб за п. 45, який відрізняється тим, що ронуклеїнові кислоти походять із різних геномів, і де слина містить Imi1 нуклеїнову кислоту та Imi3 нукпринаймні одну з множинних IMI нуклеїнових кислеїнову кислоту. лот вибирають із групи, яка складається з: 47. Спосіб одержання трансгенної рослини з підa) полінуклеотиду, який включає SEQ ID NO:1, вищеною резистентністю до імідазолінонового б) полінуклеотиду, який кодує поліпептид, що гербіциду, який включає: включає SEQ ID NO:2, a) трансформацію рослинної клітини вектором в) полінуклеотиду, який кодує поліпептид, що має експресії, який включає IMI нуклеїнову кислоту, послідовність амінокислот, принаймні на 90 % ідепричому ІМІ нуклеїнова кислота вибрана з групи, нтичну до SEQ ID NO:2, причому поліпептид має яка складається з: заміщення серин на аспарагін в Домені Е, і і) полінуклеотиду, який включає SEQ ID NO:1, г) полінуклеотиду, комплементарного будь-якому з іі) полінуклеотиду, який кодує поліпептид, що полінуклеотидів від a) до в). включає SEQ ID NO:2, 40. Спосіб за п. 39, який відрізняється тим, що ііі) полінуклеотиду, який кодує поліпептид, що має множинні IMI нуклеїнові кислоти вибирають із групослідовність амінокислот, принаймні на 90 % ідепи, яка складається з Imi1 нуклеїнової кислоти, нтичну до SEQ ID NO:2, причому поліпептид має Imi2 нуклеїнової кислоти та Imi3 нуклеїнової кисзаміщення серин на аспарагін в Домені Е, і лоти. іv) полінуклеотиду, комплементарного будь-якому 41. Спосіб за п. 40, який відрізняється тим, що роз полінуклеотидів від і) до ііі), та слина містить Imi1 нуклеїнову кислоту та Imi3 нукб) вирощування з рослинної клітини трансгенної леїнову кислоту. рослини з підвищеною резистентністю до імідазо42. Спосіб зміни толерантності рослини до імідалінонового гербіциду порівняно з сортом рослини золінонового гербіциду, який включає зміну ексдикого типу. 7 87261 8 48. Спосіб за п. 47, який відрізняється тим, що IMI дається з Imi2 нуклеїнової кислоти та Imi3 нуклеїнуклеїнову кислоту вибирають із групи, яка скланової кислоти. Ця заявка заявляє пріоритет попередньої заявки США №60/311,282, поданої 9 серпня 2001р. Даний винахід у цілому стосується рослин, які мають підвищену резистентність до імідазолінонових гербіцидів. Тобто даний винахід стосується пшеничних культур, одержаних шляхом мутагенезу та кросбридингу і перетворення, які мають підвищену резистентність до імідазолінонових гербіцидів. Синтаза ацетогідроксіоцтової кислоти (AHAS; EC 4.1.3.18) є першим ферментом, який каталізує біохімічний синтез амінокислот з розгалуженими ланцюгами валіну, лейцину та ізолейцину [Singh В. K., 1999 Biosynthesis of valine, leucine and isoleucine in: Singh В. K. (Ed) Plant amino acids. Marcel Dekker Inc. New York, New York. Pg 227247]. AHAS є місцем дії чотирьох різних за структурою груп гербіцидів, включаючи сульфонілсечовини [LaRossa RA and Falco SC, 1984 Trends Biotechnol 2:158-161], імідазолінони [Shaner et al., 1984 Plant Physiol 76:545-546], триазолопіримідини [Subramanian and Gerwick, 1989 Inhibition of acetolactate synthase by triazolopyrimidines in (ed) Whitaker JR, Sonnet PE Biocatalysis in agricultural biotechnology. ACS Symposium Series, American Chemical Society. Washington, D.C. Pg 277-288] та піримідилоксибензоати [Subramanian et al., 1990 Plant Physiol 94: 239-244]. Імідазолінонові та сульфонілсечовинні гербіциди широко застосовують у сучасному сільському господарстві завдяки їхній ефективності при дуже низьких нормах внесення та відносній нетоксичності для тварин. Шляхом інгібування активності AHAS ці групи гербіцидів перешкоджають ростові та розвиткові чутливих до них рослин, включаючи багато видів бур'янів. Прикладами імідазолінонових гербіцидів серійного виробництва є PURSUITÒ (імазетапір), SCEPTERÒ (імазахін) та ARSENALÒ (імазапір). Приклади сульфонілсечовинних гербіцидів є хлорсульфурон, метсульфурон-метил, сульфометуронметил, хлоримурон-етил, трифенсульфуронметил, трибенурон-метил, бенсульфурон-метил, нікосульфурон, етаметсульфурон-метил, римсульфурон, трифлусульфурон-метил, триасульфурон, примісульфурон-метил, циносульфурон, амідосульфурон, флузасульфурон, імазосульфурон, піразосульфурон-етил та галосульфурон. Завдяки їх високій ефективності та низькій токсичності, імідазоліноновим гербіцидам віддають перевагу, коли йдеться про нанесення шляхом розпилення над широкими площами рослинних культур. Можливість розпилення гербіциду над великими площами рослинних культур знижує витрати, пов'язані зі створенням плантацій та доглядом за ними і зменшує потребу в підготуванні місця до застосування таких хімікатів. Розпилення над потрібними толерантними видами в результаті також забезпечує можливість досягнення максимального врожаю потрібних культур завдяки відсутності конкуруючих видів. Однак, можливість застосування таких способів розпилення залежить від наявності резистентних до імідазолінону видів потрібних рослин на площі розпилення. Серед основних сільськогосподарських культур деякі види зернобобових, такі як соя, мають природну резистентність до імідазолінонових гербіцидів завдяки їх здатності до швидкого засвоєння гербіцидних сполук [Shaner and Robinson, 1985 Weed Sci. 33:469-471]. Інші культури, такі як кукурудза [Newhouse et al., 1992 Plant Physiol. 100:882886] та рис [Barrette et al., 1989 Crop Safeners for Herbicides, Academic Press New York, pp. 195-220], певною мірою є чутливими до імідазолінонових гербіцидів. Розбіжності в чутливості до імідазолінонових гербіцидів залежать від хімічної природи конкретного гербіциду та характеру метаболізму сполуки від токсичної до нетоксичної форми в кожній рослині [Shaner et al., 1984 Plant Physiol. 76:545-546; Brown et al., 1987 Pestic. Biochm. Physiol. 27:24-29]. Інші фізіологічні розбіжності рослин, такі як абсорбція та транслокація, також відіграють важливу роль у чутливості [Shaner and Robinson, 1985 Weed Sci. 33:469-471]. Культурні сорти, резистентні до імідазолінонів, сульфонілсечовин та триазолопіримідинів, успішно виводилися з застосуванням насіння, мікроспор, пилку та мутагенезу калюсу в Zea mays, Arabidopsis thaliana, Brassica napus, Glycine max, and Nicotiana tabacum [Sebastian et al., 1989 Crop Sci. 29:1403-1408; Swanson et al., 1989 Theor. Appl. Genet. 78:525-530; Newhouse et al., 1991 Theor. Appl. Genet. 83:65-70; Sathasivan et al., 1991 Plant Physiol. 97:1044-1050; Mourand et al., 1993 J. Heredity 84: 91-96]. В усіх випадках резистентність забезпечував єдиний, частково домінантний ядерний ген. Раніше також були виведені чотири резистентні до імідазолінону пшеничні культури після мутагенезу насіння Triticum aestivum L. cv Fidel [Newhouse et al., 1992 Plant Physiol. 100:882-886]. Дослідження успадкування підтвердили, що єдиний, частково домінантний ген забезпечував резистентність. На основі алельних досліджень автори дійшли висновку, що мутації в чотирьох ідентифікованих лініях містилися в одному місці. Один з генів резистентності культури Fidel було позначено як FS-4 [Newhouse et al., 1992 Plant Physiol. 100:882-886]. Комп'ютерне моделювання тривимірної конформації комплексу AHAS-інгібітора передбачає кілька амінокислот у запропонованій кишені зв'язування інгібітора як місця, де викликані мутації можуть забезпечувати вибіркову резистентність до імідазолінонів [Ott et al., 1996 J. Mol. ВіоІ. 263:359368) Пшеничні культури, одержані за допомогою 9 87261 10 деяких із цих раціонально спланованих мутацій у лотою. ІМІ нуклеїнова кислота може бути, напризапропонованих місцях зв'язування ферменту клад, lmi2 або lmi3 нуклеїновою кислотою. AHAS, фактично інгібували специфічну резистентІМІ нуклеїнові кислоти даного винаходу моність до єдиного класу гербіцидів [Ott et al., 1996 J. жуть включати нуклеотидну послідовність, вибрану Mol. ВіоІ. 263:359-368)]. з групи, яка складається з: полінуклеотиду SEQ ID Про резистентність рослин до імідазоліноноNO:1; полінуклеотиду SEQ ID NO:3; полінуклеотивих гербіцидів також повідомлялося в багатьох дної послідовності, яка кодує поліпептид SEQ ID патентах. У патентах США №№ 4,761,373, NO:2; полінуклеотидної послідовності, яка кодує 5,331,107, 5,304,732, 6,211,438, 6,211,439 та полі пептид SEQ ID NO:4, полінуклеотиду, який 6,222,100 у загальних рисах описано застосування включає принаймні 60 послідовних нуклеотидів зміненого гена AHAS для викликання резистентнобудь-якого з вищезгаданих полінуклеотидів; та сті до гербіцидів у рослинах і докладно описано полінуклеотиду, комплементарного будь-якому з деякі резистентні до імідазолінону лінії кукурудзи. вищезгаданих полінуклеотидів. У патенті США №5,013,659 описано рослини, які Рослини даного винаходу можуть бути трансвиявляють резистентність до гербіцидів і мають генними або нетрансгенними. Прикладами нетрамутації у принаймні одній амінокислоті в одній або нсгенних пшеничних культур з підвищеною резискількох збережених ділянках. Описані в ньому мутентністю до імідазолінонових гербіцидів є тації кодують або перехресну резистентність до пшенична культура, яка має Номер патентного імідазолінонів та сульфонілсечовин або специфічдепонування АТСС РТА-3953 або РТА-3955; або ну до сульфонілсечовини резистентність, але спемутантна, рекомбінантна або піддана генній інжецифічна до імідазолінону резистентність не описунерії похідна рослини з номером патентного депоється. Крім того, в патенті США №5,731,180 та нування АТСС РТА-3953 або РТА-3955; або будьпатенті США №5,767,361 обговорюється виділеякого потомства рослини з номером патентного ний ген, який має єдине амінокислотне заміщення депонування АТСС РТА-3953 або РТА-3955; або в амінокислотній послідовності AHAS однодольних рослина, яка є потомством будь-якої з цих рослин. дикого типу, яка в результаті забезпечує специфіКрім композицій даного винаходу, забезпечучну до імідазолінону резистентність. ється кілька способів. Авторами описано способи До цього часу в роботах існуючого рівня технізміни толерантності рослини до імідазолінонового ки не було описано резистентних до імідазолінону гербіциду, які включають зміну експресії ІМІ нуклепшеничних культур, які містять більше одного зміїнової кислоти в рослині. Описано також способи неного гена AHAS. Так само в роботах існуючого одержання трансгенної рослини, яка має підвищерівня техніки не було описано резистентних до ну толерантність до імідазолінонового гербіциду, імідазолінону пшеничних культур, які містять муякі включають перетворення рослинної клітини тації в геномах, крім геному, від якого походить ген вектором експресії, що включає одну або кілька FS-4. Таким чином, дана галузь потребує ідентиІМІ нуклеїнових кислот, та вирощування рослини з фікації генів резистентності до імідазолінону з дорослинної клітини. Винахід також охоплює спосіб даткових геномів. Дана галузь також потребує контролю над бур'янами в оточенні пшеничної пшеничних культур, які мають підвищену резистекультури, який включає нанесення імідазолінононтність до гербіцидів, таких як імідазолінон, і місвого гербіциду на бур'яни та пшеничну культуру, тять більше одного зміненого гена AHAS. Потрібні причому пшенична культура має підвищену резистакож способи контролю над ростом бур'янів потентність до імідазолінонового гербіциду порівняблизу від таких пшеничних культур. Ці композиції но з сортом пшеничної культури дикого типу, і рота способи дозволяють застосовувати технології слина містить одну або кілька ІМІ нуклеїнових розпилення при нанесенні гербіцидів на площі з кислот. У деяких оптимальних варіантах втілення пшеничними культурами. цих способів рослини включають множинні ІМІ Даний винахід забезпечує пшеничні культури, нуклеїнові кислоти, які містяться в різних геномах які включають ІМІ нуклеїнові кислоти, причому пшениці. пшенична культура має підвищену резистентність Фігура 1 є таблицею, на якій показано резульдо імідазолінонового гербіциду порівняно з різнотати оцінки резистентності однієї рослини до імавидами рослини дикого типу. Пшеничні культури замоксу в популяціях батьківської рослини та поможуть містити одну, дві, три або більше ІМІ нуккоління F1, одержаного в результаті взаємного леїнових кислот. В одному варіанті втілення пшесхрещування між резистентними лініями та CDC нична культура включає кілька ІМІ нуклеїнових Teal. Цифри означають кількість рослин, занесекислот, які містяться в різних геномах. В оптиманих до кожного фенотипічного класу. Батьківські льному варіанті ІМІ нуклеїнові кислоти кодують лінії позначено жирним шрифтом. Кількість батьбілки, які включають мутацію в консервативній ківських ліній включає лінії, зараховані до популяамінокислотній послідовності, вибраній з групи, яка цій F2. складається з Домену А, Домену В, Домену С, ДоФігура 2 є таблицею, на якій показано реакцію мену D та Домену Е. У ще кращому варіанті мутана імазамокс у популяціях F2 та BCF1, одержаних у ція міститься в консервативному Домені Ε або результаті схрещування між резистентними лініяконсервативному Домені С, Забезпечуються також ми та CDC Teal, і результати тестів Chi-square одчастини рослин та насіння рослин, отримані від нолокусних та дволокусних моделей (15А´Teal) описаних авторами пшеничних культур. В іншому для контролю резистентності. Символи, викорисваріанті втілення пшенична культура включає ІМІ тані на Фігурі 2, означають: а - значення Ρ Chiнуклеїнову кислоту, яка не є lmi1 нуклеїновою кисsquare (1 df) представляє ймовірність того, що відхилення від випробуваного співвідношення зу 11 87261 12 лінонових гербіцидів, тоді як BW755 містить FS4 мовлюються лише випадком. Значення Ρ Chiмутантний ген. squares, більші за 0,05, вказують на те, що знаФігура 7 є таблицею з порівнянням витримучення, які спостерігалися, не мали значних відмінвання ураження трьома генотипами пшениці, обностей від очікуваних значень; b - значення Ρ Chiробленими імазамоксом у кількості 10Х або 30Х. square, яке представляє ймовірність того, що відНорма 1Х становить 20г/га. BW755 містить FS4 хилення між популяціями F2, одержаними в ремутантний ген. 15А/11А є основною кількістю змізультаті взаємного схрещування між CDC Teal та шаного потомства від схрещування Teal11А та резистентними лініями, зумовлюються лише випаTeal15A. Популяція ще не була гомозиготною в дком. Значення Chi-square, більші за 0,05, вказуусіх трьох неалельних локусах. ють на те, що взаємні популяції F2 були гомогенФігура 8 показує лінійне розташування посліними, і дані між двома взаємними популяціями довності ДНК часткових генів пшениці Als1 та lmi1, були об'єднані; с - CDC Teal використовували як ампліфікованих із геномної ДНК: CDC Teal (рядок повторну батьківську рослину; d - випробувані 2; SEQ ID NO:15 і SEQ ID NO:16), BW755 (рядок 3; співвідношення ґрунтувалися на результатах поSEQ ID NO:17 і SEQ ID NO:18), TeallMI 10A (рядок коління F2; і e - значення Ρ Chi-square (1 df) для 4; SEQ ID NO:19 і SEQ ID NO:20), TeallMI 11A (ряспіввідношення BCF1. док 5; SEQ ID NO:21 і SEQ ID NO:22) і TeallMI 15A Фігура 3 є таблицею, на якій показано резуль(рядок 6; SEQ ID NO:23 і SEQ ID NO:24). Часткові тати оцінки резистентності до імазамоксу в групах послідовності суміщали з повною послідовністю F2:3 у результаті схрещування між резистентними гена рису ALS (рядок 1; SEQ ID NO:13 і SEQ ID лініями та CDC Teal, і результати тестів Chi-square NO:14), отриманого з банку генів (інвентарний нооднолокусних та двофокусних моделей (15А´Teal) мер АВО49822) і переносили на білкові послідовдля контролю резистентності. Символи, викорисності (представлені над послідовностями ДНК). тані на Фігурі 3, означають: а - випробувані співП'ять висококонсервативних амінокислотних довідношення сегрегації в групі ґрунтувалися на ременів, що містять мутації, які забезпечують резисзультатах популяцій F2 та BCF1; b - значення Ρ тентність до інгібіторів AHAS, позначено жирним Chi-square (2 df), яке представляє ймовірність того, шрифтом. Заміщення гуанідину аденіном у BW755, що відхилення від випробуваного співвідношення TeallMI 10А та TeallMI 15A ведуть до заміщення зумовлюються лише випадком. Значення Ρ Chiсерину аспарагіном (серин627 у рисі) в домені squares, більші за 0,05, вказують на те, що знаIPSGG (Домен Е) Als1 гена. Відповідно, гени резичення, які спостерігалися, не мали значних відмінстентності, присутні у рослинах BW755, TealІМІ ностей від очікуваних значень. 10А та TeallMI 15A, були вказані як частина mi1 Фігура 4 є таблицею, на якій показано резулькласу. Ці гени резистентності Teal вказуються автати оцінки резистентності однієї рослини до іматорами як ТеаІІMI1 10А та ТеаІІMI1 15A. замоксу в популяціях F2 у результаті взаємного Фігура 9 показує лінійне розташування послісхрещування між резистентними лініями. Співвіддовності ДНК часткових генів пшениці Als2 та lmi2, ношення згідно з тестом Chi-square ґрунтувалися ампліфікованих із геномної ДНК: CDC Teal (рядок на результатах у групах F2 та F2:3, одержаних від 2; SEQ ID NO:25 і SEQ ID NO:26), BW755 (рядок 3; схрещування між резистентними лініями та CDC SEQ ID NO:27 і SEQ ID NO:28), TeallMI 10A (рядок Teal. Випробуване співвідношення 15:1 стосується 4; SEQ ID NO:29 і SEQ ID NO:30), TeallMI 11A (рядволокусної моделі, а випробуване співвідношендок 5; SEQ ID NO:31 і SEQ ID NO:32) і TeallMI 15A ня 63:1 стосується трилокусної моделі. Символ (рядок 6; SEQ ID NO:33 і SEQ ID NO:34). Часткові "а", використаний на Фігурі 4, означає: значення Ρ послідовності AHAS суміщали з повною послідовChi-square (1 df), яке представляє ймовірність того, ністю AHAS рису (рядок 1; SEQ ID NO:13 і SEQ ID що відхилення від випробуваного співвідношення NO:14), отриманою з банку генів (інвентарний нозумовлюються лише випадком. Значення Ρ Chiмер АВ049822) і переносили в білкові послідовноsquares, більші за 0,05, вказують на те, що знасті (представлені над послідовностями ДНК). П'ять чення, які спостерігалися, не мали значних відмінвисококонсервативних амінокислотних доменів, ностей від очікуваних значень. що містять мутації, які забезпечують резистентФігура 5 є таблицею, на якій показано резульність до інгібіторів AHAS, позначено жирним шритати оцінки імазамоксу резистентність у групах F2:3 фтом. Заміщення гуанідину аденіном у TeallMI 11А у результаті сегрегуючого взаємного схрещування в результаті веде до заміщення серину аспарагіміж резистентними лініями. Символи, використані ном (серин627 у рисі) в домені IPSGG Als2 гена. на Фігурі 5, означають: а - випробувані співвідноВідповідно, ген резистентності, присутній у рослині шення сегрегації в групі ґрунтувалися на результаTeallMI 11А, було вказано як частину класу Іті2 тах досліджених популяцій F2; b - значення Ρ Chiнуклеїнових кислот. Цей ген резистентності Teal square (2 df), яке представляє ймовірність того, що вказано авторами як ТеаІІМІ2 11А. відхилення від випробуваного співвідношення зуФігура 10 показує часткову послідовність ДНК мовлюються лише випадком. Значення Ρ ChiTeallM11 15А (SEQ ID NO:1) та виведену з неї аміsquares, більші за 0,05, вказують на те, що знанокислотну послідовність (SEQ ID NO:2). чення, які спостерігалися, не мали значних відмінФігура 11 показує часткову послідовність ДНК ностей від очікуваних значень. ТеаІІМІ2 11А (SEQ ID NO:3) та виведену з неї аміФігура 6 є таблицею з порівнянням відсотка нокислотну послідовність (SEQ ED NO:4). неінгібованої in vitro активності AHAS у чотирьох Фігура 12 показує нуклеіновокислотну послідолініях пшениці у присутності зростаючих концентвність Teal ALS1 ORF дикого типу (SEQ ID NO:5), рацій імідазолінонового гербіциду імазамокс. Teal є лінією дикого типу без толерантності до імідазо 13 87261 14 Teal ALS2 ORF (SEQ ID NO:6) Teal ALS3 ORF порівняно з різновидами рослини дикого типу. (SEQ ID NO:7). Вжитий авторами термін "ІМІ нуклеїнова кислота" Фігура 13 є схематичним зображенням консестосується нуклеїнової кислоти, яка є мутованою рвативних амінокислотних послідовностей в AHAS від нуклеїнової кислоти AHAS у пшеничній культурі генах, пов'язаних із резистентністю до різних інгібідикого типу, і яка надає підвищеної резистентності торів AHAS. Конкретну амінокислотну ділянку, яка до імідазолінону рослині, в якій вона транскрибувідповідає за резистентність, позначено підкресється. Нуклеїнові кислоти AHAS Teal дикого типу ленням. [Modified from Devine, M. D. and Eberlein, С показано в SEQ ID NO:5 (Teal ALS1 ORF), SEQ ID V., 1997 Physiological, biochemical and molecular NO:6 (Teal ALS2 ORF) і SEQ ID NO:7 (Teal ALS3 aspects of herbicide resistance based on altered ORF). В одному варіанті втілення пшенична кульtarget sites in Herbicide Activity: Toxicity, тура включає множинні ІМІ нуклеїнові кислоти. Biochemistry, and Molecular Biology, IOS Press Вжитий в описі ІМІ нуклеїнових кислот термін Amsterdam, p. 159-185]. "множинні" стосується ІМІ нуклеїнових кислот, які Даний винахід стосується пшеничних культур, мають різні нуклеотидні послідовності і не стосучастин пшеничних культур та клітин пшеничних ється простого збільшення кількості однієї ІМІ нуккультур з підвищеною резистентністю до імідазолеїнової кислоти. Наприклад, ІМІ нуклеїнові кислолінонових гербіцидів. Даний винахід також охопти можуть бути різними через те, що вони лює насіння, вироблене описаними авторами походять із різних геномів або містяться в різних пшеничними культурами, та способи контролю над геномах пшениці. бур'янами в оточенні описаних авторами пшеничПшеничні культури даного винаходу можуть них культур. Слід розуміти, що вжита в описі та мати множинні ІМІ нуклеїнові кислоти з різних геформулі форма однини може означати як однину, номів, оскільки ці рослини можуть містити більше так і множину, залежно від контексту, в якому її одного геному. Наприклад, пшенична культура вжито. Так, наприклад, посилання на "клітину" моTriticum aestivum містить три геноми, які іноді наже означати використання принаймні однієї клітизивають геномами А, В та D. Оскільки AHAS є пони. трібним метаболічним ферментом, то вважається, Вжитий авторами термін "пшенична культура" що геном має принаймні один ген, який кодує фестосується рослини, яка належить до роду рмент AHAS, який зазвичай спостерігається з інTriticum. Пшеничні культури даного винаходу мошими метаболічними ферментами в гексаплоїдній жуть належати до роду Triticum, включаючи, крім пшениці, підданій картуванню. Нуклеїнова кислота інших, Т. aestivum, Т. turgidum, Т. timopheevii, Т. AHAS у кожному геномі може відрізнятися і зазвиmonococcum, Т. zhukovskyi та T. urartu і їх гібриди. чай відрізняється своєю нуклеотидною послідовніПрикладами підвиду Т. aestivum, які охоплюються стю від нуклеїнової кислоти AHAS в іншому геномі. даним винаходом, є aestivum (пшениця звичайна), Спеціаліст у даній галузі може визначити геном compaction (карликова пшениця), macha (пшениця походження кожної нуклеїнової кислоти AHAS, маха), vavilovi (пшениця Вавілова), spelta та застосовуючи генетичний кросинг та/або інші споsphaecrococcum (пшениця спельта). Прикладами соби секвенування або гідроліз екзонуклеази, які підвиду Т. turgidum, які охоплюються даним винавідомі спеціалістам, а також описані нижче у Приходом, є turgidum, carthlicum, dicoccon, durum, кладі 2. З точки зору цього винаходу, ІМІ нуклеїноpaleocolchicum, polonicum, turanicum та ві кислоти, які походять із одного з геномів А, В dicoccoides. Прикладами підвиду Т. monococcums, або D, розпізнають і позначають як нуклеїнові кисякі охоплюються даним винаходом, є monococcum лоти lmi1, lmi2 або lmi3. (einkom) та aegilopoides. В одному варіанті втіленАвтори не стверджують, що будь-який конкреня даного винаходу пшенична культура належить тний клас ІМІ нуклеїнової кислоти співвідноситься до виду Triticum aestivum species, точніше, сорту з будь-яким конкретним геномом А, В або D. НаCDC Teal. приклад, автори не стверджують, що lmi1 нуклеїТермін "пшенична культура" охоплює пшеничнові кислоти співвідносяться з нуклеїновими кисні культури на будь-якій стадії стиглості або розвилотами геному А, що lmi2 нуклеїнові кислоти тку, а також будь-які тканини або органи (частини співвідносяться з нуклеїновими кислотами геному рослин), взяті або одержані від будь-якої подібної В і т. д. Позначення lmi1, lmi2 та lmi3 лише вказурослини, якщо в контексті прямо не вказано іншоють, що ІМІ нуклеїнові кислоти в межах кожного го. До частин рослин належать, крім інших, стебла, такого класу не виділяються незалежно, тоді як дві коріння, квітки, насінні зачатки, тичинки, листя, ІМІ нуклеїнові кислоти різних класів виділяються зародки, ділянки меристеми, тканина калюсу, незалежно і, таким чином, можуть походити з різкультури пиляка, гаметофіти, спорофіти, пилок, них геномів пшениці. Клас lmi1 нуклеїнових кислот мікроспори, протопласти та ін. Даний винахід тавключає FS-4 ген, як описано в роботі Newhouse et кож охоплює насіння, вироблене пшеничними al. [1992 Plant Physiol. 100:882-886], та ген ТеаІІMI1 культурами даного винаходу. В одному варіанті 15А, детальніше описаний нижче. Клас lmi2 нуклевтілення насіння одержують шляхом розведення їнових кислот включає ген ТеаІІМІ2 11А, описаний гомозигот для підвищеної резистентності до іміданижче. Кожен Imi клас може включати члени інших золінонового гербіциду порівняно з насінням сорту видів пшениці. Отже, кожен Imi клас включає ІМІ пшеничної культури дикого типу. нуклеїнові кислоти, які відрізняються за своєю нуУ даному винаході описано пшеничну культуклеотидною послідовністю, але, незважаючи на ру, яка включає одну або кілька ІМІ нуклеїнових це, позначаються як такі, що походять або міскислот, причому пшенична культура має підвищетяться в одному геномі пшениці, на основі досліну резистентність до імідазолінонового гербіциду 15 87261 16 ших, з-поміж, 2-(4-ізопропіл-4-метил-5-оксо-2дження успадкування, як описано нижче у Приклаімідазолін-2-іл)-нікотинов кислота, 2-(4-ізопропіл)дах і відомо спеціалістам у даній галузі. 4-метил-5-оксо-2-імідазолін-2-іл)-3Відповідно, даний винахід охоплює пшеничну хінолінкарбонової кислоти, 5-етил-2-(4-ізопропіл-4культуру, яка включає одну або кілька ІМІ нуклеїметил-5-оксо-2-імідазолін-2-іл)-нікотинової кислонових кислот, причому пшенична культура має ти, 2-(4-ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)підвищену резистентність до імідазолінонового 5-(метоксиметил)-нікотинової кислоти, 2-(4гербіциду порівняно з різновидами рослини дикого ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5типу, і одну або кілька ІМІ нуклеїнових кислот виметилнікотинової кислоти, та суміші метил 6-(4бирають із групи, яка складається з нуклеїнової ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-(тікислоти Imi1, Imi2 та lmi3. В одному варіанті втітолуату і метил 2-(4-ізопропіл-4-метил-5-оксо-2лення рослина містить lmi1 нуклеїнову кислоту та імідазолін-2-іл)-m-толуату. Перевагу віддають заlmi3 нуклеїнову кислоту. В оптимальному варіанті стосуванню 5-етил-2-(4-ізопропіл-4-метил-5-оксовтілення lmi1 нуклеїнова кислота включає полінук2-імідазолін-2-іл)-нікотинової кислоти та 2-(4леотидну послідовність, показану в SEQ ID NO:1. ізопропіл-4-метил-5-оксо-2-імідазолін-2-іл)-5В іншому варіанті втілення рослина містить lmi2 (метоксиметил)-нікотинової кислоти. Особливу нуклеїнову кислоту. В оптимальному варіанті втіперевагу віддають застосуванню 2-(4-ізопропіл-4лення lmi2 нуклеїнова кислота включає полінуклеметил-5-оксо-2-імідазолін-2-іл)-5-(метоксиметил)отидну послідовність, показану в SEQ ID NO:3. нікотинової кислоти. Вжитий авторами по відношенню до нуклеїноВ одному варіанті втілення пшенична культура вих кислот термін "із" стосується нуклеїнової кисмістить дві ІМІ нуклеїнові кислоти, причому нуклеїлоти, яка "міститься у" або "походить із" конкретнові кислоти походять із або містяться в різних ного геному. Термін "міститься у" стосується геномах пшениці. В оптимальному варіанті однією нуклеїнової кислоти, яка міститься в межах конкз двох нуклеїнових кислот є lmi1 нуклеїнова кислоретного геному. Також вжитий авторами по віднота, краще - якщо вона включає полінуклеотидну шенню до геному термін "походить із" стосується послідовність SEQ ID NO:1. В іншому варіанті втінуклеїнової кислоти, видаленої або виділеної з лення пшенична культура містить одну ІМІ нуклеїцього геному. Термін "виділений" детальніше винову кислоту, причому нуклеїнова кислота вклюзначено нижче. чає полінуклеотидну послідовність SEQ ID NO:1 В іншому варіанті втілення пшенична культура або SEQ ID NO:3. У ще одному варіанті втілення включає ІМІ нуклеїнову кислоту, причому ця нуклепшенична культура містить три або більше ІМІ їнова кислота не є lmi1 нуклеїновою кислотою. нуклеїнових кислот, причому кожна нуклеїнова Термін "не lmi1" стосується ІМІ нуклеїнової кислокислота походить із свого геному. В оптимальному ти, яка не належить до lmi1 класу, як описано виваріанті принаймні одна з трьох ІМІ нуклеїнових ще. Приклади нуклеїнових кислот lmi1 класу покакислот включає полінуклеотидну послідовність, зано в рядках 3, 4 та 5 на Фігурі 8. Один приклад вибрану з групи, яка складається з SEQ ID NO:1 і не lmi1 нуклеїнової кислоти показано в рядку 5 на SEQ ID NO:3. Фігурі 8. Відповідно, в оптимальному варіанті втіВ оптимальному варіанті втілення даного вилення пшенична культура містить ІМІ нуклеїнову находу одна або кілька ІМІ нуклеїнових кислот, які кислоту, яка включає полінуклеотидну послідовмістяться в рослині, кодують амінокислотну посліність, яка кодує поліпептид SEQ ID NO:4. Полінукдовність, яка включає мутацію в домені, збережелеотидна послідовність може включати послідовному серед кількох білків AHAS. Ці консервативні ність, показану в SEQ ID NO:3. домени вказуються авторами як Домен А, Домен Даний винахід включає пшеничні культури, які В, Домен С, Домен D та Домен Е. Фігура 13 покамістять одну, дві, три або більше ІМІ нуклеїнових зує загальне розташування кожного домену в білку кислот, причому пшенична культура має підвищеAHAS. Вжитий авторами термін Домен А містить ну резистентність до імідазолінонового гербіциду амінокислотну послідовність AITGQVPRRMIGT порівняно з різновидами рослини дикого типу. ІМІ (SEQ ID NO:8); Домен В містить амінокислотну нуклеїнові кислоти можуть включати нуклеотидну послідовність QWED (SEQ ID NO:9); Домен С міспослідовність вибрану з групи, яка складається з тить амінокислотну послідовність полінуклеотиду SEQ ID NO:1; полінуклеотиду SEQ VFAYPGGASMEIHQALTRS (SEQ ID NO:10); Домен ID NO:3; полінуклеотидної послідовності, яка кодує D містить амінокислотну послідовність AFQETP поліпептид SEQ ED NO:2; полінуклеотидної послі(SEQ ID NO:11); Домен Ε містить амінокислотну довності, яка кодує поліпептид SEQ ID NO:4, поліпослідовність IPSGG (SEQ ID NO:12). Даний винануклеотиду, який включає принаймні 60 послідовхід також передбачає можливість незначних відних нуклеотидів будь-якого з вищезгаданих хилень у консервативних доменах, наприклад, у полінуклеотидів; та полінуклеотиду, комплементакуколі сериновий залишок у Домені Ε заміщено рного будь-якому з вищезгаданих полінуклеотидів. аланіновим залишком. Імідазоліноновий гербіцид може бути вибраВідповідно, даний винахід охоплює пшеничну ний, крім інших, з-поміж, PURSUITÒ (імазетапір), культуру, яка включає ІМІ нуклеїнову кислоту, яка CADREÒ (імазапіку), RAPTORÒ (імазамокс), кодує амінокислотну послідовність, яка має мутаSCEPTERÒ (імазахін), ASSERTÒ (імазетабензу), цію в консервативному Домені, вибраному з групи, ARSENALÒ (імазапір), похідної будь-якого з вищеяка складається з Домену А, Домену В, Домену С, згаданих гербіцидів або суміші двох або більшої Домену D та Домену Е. В одному варіанті втілення кількості вищезгаданих гербіцидів, наприклад, імапшенична культура містить ІМІ нуклеїнову кислоту, запіру/імазамоксу (ODYSSEYÒ). Точніше, імідазояка кодує амінокислотну послідовність, яка має ліноновий гербіцид може бути вибраний, крім ін 17 87261 18 мутацію в Домені Е. В інших оптимальних варіантидну та виведену амінокислотну послідовності, тах втілення мутації в консервативних доменах які відповідають генові TealMI1 15A, показано в трапляються в місцях, позначених таким підкресSEQ ID NO:1 і SEQ ID NO:2, відповідно. Єдиним ленням: AITGQVPRRMIGT (SEQ ID NO:8); OWED типом послідовностей, не включених до SEQ ID (SEQ ID NO:9); VFAYPGGASMEIHQALTRS (SEQ ID NO:1 і SEQ ID NO:2, є послідовності, які кодують і NO:10); AFQETP (SEQ ID NO:11) та IPSGG (SEQ відповідають приблизно 100-150 парам основ на 5' ID NO:12). Оптимальним заміщенням є аспарагін кінці і приблизно 5 парам основ на 3' кінці кодуючої замість серину в Домені E (SEQIDNO:12). ділянки. Описані авторами пшеничні культури можуть Окремі депонування 2500 зразків насіння сорбути або трансгенними пшеничними культурами, тів пшениці TeallMI 11A та TeallMI 15A було зробабо нетрансгенними пшеничними культурами. лено в Американському зібранні типових культур, Вжитий авторами термін "трансгенний" стосується Манассас, Вірджинія, 3 січня 2002р. Ці депонуванбудь-якої рослини, рослинної клітини, калюсу, роня було зроблено згідно з умовами Будапештської слинної тканини або частини рослини, що містить, угоди стосовно депонування мікроорганізмів. Деповністю або частково, принаймні один рекомбінапонування було зроблено на термін не менше нтний полінуклеотид. У багатьох випадках рекомтридцяти років і не менше п'яти років після отрибінантний полінуклеотид, частково або повністю, є мання АТСС останнього запиту про надання депостабільно включеним у хромосому або стійкий нованого зразка. Депоновані зразки насіння отрипозахромосомний елемент таким чином, що він мали номери патентного депонування РТА-3953 передається наступним поколінням. З точки зору (TeallMI 11A) та РТА-3955 (TeallMI 15A). винаходу термін "рекомбінантний полінуклеотид" Даний охоплює пшеничну культуру, яка має стосується полінуклеотиду, який було змінено, номер патентного депонування РТА-3953 або перебудовано або модифіковано шляхом генної РТА-3955; мутантну, рекомбінантну або піддану інженерії. Прикладами є будь-який клонований генній інженерії похідну рослини з номером патенполінуклеотид або полінуклеотиди, які є зв'язанитного депонування РТА-3953 або РТА-3955; будьми або приєднаними до гетерологічних послідовяке потомство з номером патентного депонування ностей. Термін "рекомбінантний" не стосується РТА-3953 або РТА-3955; та рослину, яке є потомсзмін полінуклеотидів, які є результатом природних твом будь-якої з цих рослин. В оптимальному ваподій, таких як спонтанні мутації, або неспонтанноріанті втілення пшенична культура даного винахого мутагенезу з наступною селекцією. Рослини, які ду додатково має характеристики резистентності містять мутації, що виникають внаслідок неспондо гербіцидів, як у рослини з номером . патентного танного мутагенезу та селекції, вказуються автодепонування РТА-3953 або РТА-3955. рами як нетрансгенні рослини і охоплюються даДаний винахід також охоплює описані автораним винаходом. У варіантах втілення, в яких ми гібриди сортів пшениці TeallMI 11A та TeallMI пшенична культура є трансгенною і містить мно15A. Приклад 5 демонструє ТеаІІMI11A/ТеаІІМІ15А жинні ІМІ нуклеїнові кислоти, нуклеїнові кислоти гібриди з підвищеною резистентністю до імідазоліможуть походити з різних геномів або одного генонового гербіциду. Даний винахід також охоплює ному. Або ж у варіантах втілення, в яких пшенична гібриди сортів пшениці TeallMI 11A або TeallMI культура є нетрансгенною і містить множинні ІМІ 15As та іншого сорту пшениці. До інших сортів нуклеїнові кислоти, нуклеїнові кислоти містяться в пшениці належить, крім інших, Т. aestivum L. cv різних геномах. Fidel та будь-який сорт пшениці, який містить муПрикладом нетрансгенного сорту пшеничної тантний ген FS-1, FS-2, FS-3 або FS-4. [див. патент культури, що містить одну ІМІ нуклеїнову кислоту, США №6,339,184 та патентну заявку США є сорт рослини, депонований в АТСС під номером №08/474,832]. В оптимальному варіанті втілення патентного депонування РТА-3953 і вказаний авпшенична культура є гібридом між сортом TeallMI торами як сорт пшениці ТеаІІМІ 11A. Сорт пшениці 11A та сортом Fidel FS-4. Гібриди TeallMI 11A/FS-4 ТеаІІМІ 11А містить lmi2 нуклеїнову кислоту. Частмістять Imi1 нуклеїнову кислоту та lmi2 нуклеїнову кову нуклеотидну та виведену амінокислотну покислоту. Гібрид TeallMI 11A та сорту Fidel, який слідовності, які відповідають генові ТеаІІМІ2 11A, містить FS-4 ген, охоплюється даним винаходом і показано в SEQ ID NO:3 і SEQ ID NO:4, відповідно. є депонованим в Американському зібранні типових Єдиним типом послідовностей, не включених до культур, Манассас, Вірджинія, 3 січня 2002р. Це SEQ ID NO:3 і SEQ ID NO:4, є послідовності, які депонування було зроблено згідно з умовами Букодують і відповідають сигнальній послідовності, дапештської угоди стосовно депонування мікрооряка відщеплюється від зрілого білка ТеаІІМІ2 11А. ганізмів. Депонування було здійснено на термін не Відповідно, SEQ ID NO:4 являє собою повну вивеменше тридцяти років і не менше п'яти років після дену послідовність зрілого білка ТеаІІМІ2 11A. отримання АТСС останнього запиту про надання Прикладом сорту пшеничної культури, який мідепонованого зразка. Депоноване насіння отристить дві ІМІ нуклеїнові кислоти в різних геномах, є мало номер патентного депонування РТА-3954. сорт рослини, депонований в АТСС під номером Терміни "сорт" та "різновид" стосуються групи патентного депонування РТА-3955 і вказаний аврослин у межах виду, які мають певну кількість торами як сорт пшениці ТеаІІМІ 15А. Сорт пшениці спільних характеристик або особливостей, які спеТеаІІМІ 15А містить lmi1 та lmi3 нуклеїнові кислоти ціалістами вважаються достатніми для відрізнення lmi1 нуклеїнова кислота включає мутацію, яка веде одного сорту або різновиду від іншого сорту або до заміни серину на аспарагін у кодованому нею різновиду. Жоден з термінів не передбачає, що всі ІМІ білку. Мутовані гени AHAS позначено авторами рослини будь-якого даного сорту або різновиду є як ТеаІІММ 15А та ТеаІІМІ3 15А. Часткову нуклеогенетично ідентичними на рівні повного гена або 19 87261 20 на молекулярному рівні, або що будь-яка дана ланцюга або 2'-гідрокси у групі рибози РНК. Антирослина є гомозиготною в усіх локусах. Сорт або смислові полінуклеотиди та рибозими можуть різновид вважають "розведенням гомозигот" для складатися повністю з рибонуклеотидів, або моконкретної особливості, якщо в разі, коли гомозижуть містити змішані рибонуклеотиди та деоксиготний сорт або різновид є самозапилюваним, усе рибонуклеотиди. Полінуклеотиди винаходу одерпотомство має цю особливість. У даному винаході жують будь-якими способами, включаючи особливість виникає внаслідок мутації в гені AHAS одержання геномних препаратів, кДНК-препаратів, пшеничної культури або насіння. in vitro синтез, RT-ПЛР та in vitro або in vivo трансСлід розуміти, може включати дикий або некрипцію. мутований ген AHAS додатково до ІМІ гена. Як "Виділеною" молекулою нуклеїнової кислоти є описано у Прикладі 4, передбачається, що сорт молекула, яка є практично відокремленою від інпшениці TeallMI 11A містить мутацію лише в одноших молекул нуклеїнових кислот, присутніх у приму з множинних AHAS ізоферментів, і що сорт родному джерелі нуклеїнової кислоти (тобто попшениці ТеаІІМІ 15A містить мутацію лише у двох слідовностей, які кодують інші поліпептиди). В із множинних AHAS ізоферментів. Отже, даний оптимальному варіанті "виділена" нуклеїнова кисвинахід охоплює пшеничну культуру, яка включає лота є вільною від деяких із послідовностей, які у одну або кілька ІМІ нуклеїнових кислот додатково природі є фланкуючими на нуклеїновій кислоті до однієї або кількох диких або немутованих нук(тобто послідовностей, розміщених на 5' та 3' кінлеїнових кислот AHAS. цях нуклеїнової кислоти) в її природному репліконі. Крім пшеничних культур, даний винахід охопНаприклад, клоновану нуклеїнову кислоту вважалює виділені ІМІ білки та нуклеїнові кислоти. Нукють виділеною. У різних варіантах втілення виділеїнові кислоти включають полінуклеотид, вибралена молекула ІМІ нуклеїнової кислоти може місний з групи, яка складається з полінуклеотиду SEQ тити менше, ніж приблизно 5kb, 4kb, 3kb, 2kb, 1kb, ID NO:1; полінуклеотиду SEQ ID NO:3; полінуклео0,5kb або 0,1kb нуклеотидних послідовностей, які у тидної послідовності, яка кодує поліпептид SEQ ID природі є фланкуючими на молекулі нуклеїнової NO:2; а полінуклеотидної послідовності, яка кодує кислоти в геномній ДНК клітини, з якої походить поліпептид SEQ ID NO:4, полінуклеотиду, який нуклеїнова кислота (наприклад, клітини Triticum включає принаймні 60 послідовних нуклеотидів aestivum). Нуклеїнову кислоту також вважають будь-якого з вищезгаданих полінуклеотидів; та виділеною, якщо її було змінено через втручання полінуклеотиду, комплементарного будь-якому з людини або поміщено в локус або місце, яке не є її вищезгаданих полінуклеотидів. В оптимальному природним місцем, або якщо її було введено у варіанті втілення ІМІ нуклеїнова кислота включає клітину шляхом агроінфекції або біолістики. Крім полінуклеотидну послідовність, яка кодує поліпептого, "виділена" молекула нуклеїнової кислоти, тид SEQ ID NO:2 або SEQ ID NO:4. В іншому оптака як молекула кДНК, може бути вільною від тимальному варіанті втілення ІМІ нуклеїнова кисдеякого іншого клітинного матеріалу, з яким вона лота включає полінуклеотидну послідовність SEQ пов'язана у природі, або культурального середоID NO:1 або SEQ ID NO:3. вища, якщо її одержують рекомбінантними спосоТермін "білок AHAS" стосується білка синтази бами, або хімічних попередників або інших хімічацетогідроксіоцтової кислоти, а термін "ІМІ білок" них речовин, якщо її синтезують хімічним шляхом. стосується будь-якого білка AHAS, який є мутоваПрямо виключаються з визначення "виділені ним від білка AHAS дикого типу і який надає більнуклеїнові кислоти": природні хромосоми (такі як шої резистентності до імідазолінону рослині, росрозтягнуті хромосоми), бібліотеки штучних хромолинній клітині, частині рослини, насінню рослини сом, геномні бібліотеки та бібліотеки кДНК, які ісабо рослинній тканині, якщо він у них експресуєтьнують або як in vitro препарат нуклеїнової кислоти, ся. В оптимальному варіанті втілення ІМІ білок або як препарат трансфікованої/трансформованої включає поліпептид SEQ ID NO:2 або SEQ ID клітини-хазяїна, причому клітини-хазяї є або in vitro NO:4. також вжиті авторами терміни "нуклеїнова гетерогенним препаратом, або вирощеними на кислота" та "полінуклеотид" стосуються РНК або пластині як гетерогенна популяція окремих колоДНК, яка є лінійною або розгалуженою, одно- або ній. Також прямо виключаються вищезгадані біблідволанцюговою, або їх гібриду. Термін також охоотеки, в яких зазначена нуклеїнова кислота склаплює гібриди РНК/ДНК. Ці терміни також охоплюдає менше 5% від кількості нуклеїновокислотних ють нетрансльовану послідовність, розміщену на вставок у векторних молекулах. Також прямо ви3' та 5' кінцях кодуючої ділянки гена: принаймні ключаються геномні ДНК цілих клітин або препаблизько 1000 нуклеотидів послідовності перед 5' рати РНК цілих клітин (включаючи препарати цілих кінцем кодуючої ділянки, і принаймні близько 200 клітин, які є механічно зрізаними або підданими нуклеотидів послідовності після 3' кінця кодуючої ферментативному гідролізові). Крім того, прямо ділянки гена. Менш поширені основи, такі як іновиключаються препарати цілих клітин, які існують зин, 5-метилцитозин, 6-метиладенін, гіпоксантин або як in vitro препарат, або як гетерогенна суміш, та інші, також можуть бути використані для антивідокремлена шляхом електрофорезу, в якій нуксмислового, dsPHK- та рибозимного спарювання. леїнову кислоту згідно з винаходом не було піддаНаприклад, було виявлено, що полінуклеотиди, які но подальшому відокремленню від гетерологічних містять С-5 пропінові аналоги уридину та цитидинуклеїнових кислот у середовищі електрофорезу ну, зв'язуються з РНК з високою спорідненістю і є (наприклад, подальшому відокремленню шляхом сильними антисмисловими інгібіторами експресії вирізання однієї смуги з гетерогенної сукупності генів. Також здійснюють інші модифікації, наприсмуг в агарозному гелі або шляхом блотування на клад, модифікацію фосфодіестерного основного нейлон). 21 87261 22 Молекулу нуклеїнової кислоти даного винахорослини дикого типу. Способи кількісного визнаду, наприклад, молекулу нуклеїнової кислоти, яка чення підвищеної резистентності до імідазоліномістить нуклеотидну послідовність SEQ ID NO:1, нових гербіцидів представлено нижче у ПриклаSEQ ED NO:3 або її частину, виділяють, застосодах. Біологічно активні частини ІМІ білка вуючи стандартні способи молекулярної біології та включають пептиди, кодовані полінуклеотидними представлену авторами інформацію про послідовпослідовностями, які включають SEQ ID NO:1 або ність. Наприклад, ІМІ кДНК Т. aestivum виділяють з SEQ ID NO:3, які включають менше амінокислот, бібліотеки Т. aestivum, застосовуючи, повністю або ніж ІМІ білок повної довжини, і надають підвищеної частково, послідовність SEQ ID NO:1 або SEQ ID резистентності до імідазолінонового гербіциду NO:3. Крім того, молекула нуклеїнової кислоти, яка після експресії в рослині. Як правило, біологічно включає, повністю або частково, SEQ ID NO:1 або активні частини (наприклад, пептиди, які мають у SEQ ID NO:3, може бути виділена шляхом полімедовжину, наприклад, 5, 10, 15, 20, 30, 35, 36, 37, разної ланцюгової реакції з застосуванням оліго38, 39, 40, 50, 100 або більше амінокислот) вклюнуклеотидних праймерів, побудованих на основі чають домен або мотив з принаймні однією активцієї послідовності. Наприклад, мРНК виділяють із ністю ІМІ білка. Крім того, інші біологічно активні рослинних клітин [наприклад, шляхом процедури частини, в яких делетовано інші ділянки поліпепвиділення гуанідин тіоціанату, описаної в роботі тиду, одержують рекомбінантними способами і Chirgwin et al., 1979 Biochemistry 18:5294-5299], а оцінюють на наявність описаних авторами однієї кДНК одержують, застосовуючи зворотну трансабо кількох активностей. В оптимальному варіанті криптазу (наприклад, Moloney MLV зворотну біологічно активні частини ІМІ білка включають транскриптазу, яку отримують від Gibco/BRL, один або кілька консервативних доменів, вибраних Bethesda, MD; або AMV зворотну транскриптазу, з групи, яка складається з Домену А, Домену В, яку отримують від Seikagaku America, Inc., St. Домену С, Домену D та Домену Е, причому консеPetersburg, FL). Синтетичні олігонуклеотидні рвативний домен містить мутацію. праймери для ампліфікації полімеразної ланцюгоВинахід також забезпечує химерні або злиті вої реакції можуть бути побудовані на основі нукІМІ поліпептиди. Вжитий авторами термін ІМІ "хилеотидної послідовності, показаної в SEQ ID NO:1 мерний поліпептид" або "злитий поліпептид" охопабо SEQ ID NO:3. Молекула нуклеїнової кислоти лює ІМІ поліпептид, функціонально зв'язаний з невинаходу може бути ампліфікована з застосуванІМІ поліпептидом. Термін "не-ІМІ поліпептид" стоням кДНК або, в альтернативному варіанті, геномсується поліпептиду, який має амінокислотну поної ДНК як матриці та відповідних олігонуклеотидслідовність, яка не має значної ідентичності ІМІ них праймерів згідно зі стандартними способами поліпептидові, наприклад, поліпептиду, який не є ампліфікації ПЛР. Ампліфіковану таким чином моІМІ ізоферментом, причому цей пептид виконує лекулу нуклеїнової кислоти клонують у відповідний відмінну від ІМІ поліпептиду функцію. По відновектор і характеризують шляхом аналізу послідовшенню до злитого поліпептиду термін "функціонаності ДНК. Крім того, олігонуклеотиди, які відповільно зв'язаний" означає, що ІМІ поліпептид та недають нуклеотидній послідовності ІΜІ, можуть бути ІМІ поліпептид є злитими один з одним таким чиодержані стандартними способами синтезу, наном, що обидві послідовності виконують передбаприклад, із застосуванням автоматизованого синчену функцію, характерну для даної послідовності. тезатора ДНК. Не-ІМІ поліпептид може бути злитий з N-кінцем ІМІ нуклеїнові кислоти даного винаходу моабо С-кінцем ІМІ поліпептиду. Наприклад, в одножуть включати послідовності, які кодують ІМІ білок му варіанті втілення злитий поліпептид є GST-IMI (тобто "кодуючі ділянки"), а також 5' нетрансльозлитим поліпептидом, у якому ІМІ послідовність є вані послідовності та 3' нетрансльовані послідовзлитою з С-кінцем GST послідовності. Такі злиті ності. В альтернативному варіанті молекули нуклеполіпептиди можуть сприяти очищенню рекомбіїнових кислот даного винаходу можуть включати нантних ІМІ поліпептидів. В іншому варіанті втілише кодуючі ділянки ІМІ гена, або можуть містити лення злитий поліпептид є ІМІ поліпептидом, який повні геномні фрагменти, виділені з геномної ДНК. містить гетерологічну сигнальну послідовність на Кодуючу ділянку цих послідовностей позначають своєму N-кінці. У деяких клітинах-хазяях (наприяк "ORF-позицію". Крім того, молекула нуклеїнової клад, клітинах-хазяях ссавців) експресія та/або кислоти винаходу може включати частину кодуюсекреція ІМІ поліпептиду може бути посилена зачої ділянки ІМІ гена, наприклад, фрагмент, який вдяки застосуванню гетерологічної сигнальної може бути використаний як зонд або праймер. послідовності. Нуклеотидні послідовності, визначені за клонуванВиділена молекула нуклеїнової кислоти, яка ням ІМІ гена з T. Aestivum, дозволяють утворювати кодує ІМІ поліпептид, що має послідовність, ідензонди та праймери, призначені для застосування в тичну поліпептидові, який кодується полінуклеотирозпізнанні та/або клонуванні ІМІ гомологів в інших дною послідовністю SEQ ID NO:1 або SEQ ID типах клітин та організмів, а також ІМІ гомологів з NO:3, може бути створена шляхом введення одноінших пшеничних культур та споріднених видів. го або кількох нуклеотидних заміщень, додавань Частина кодуючої ділянки також може кодувати або делецій у нуклеотидну послідовність SEQ ID біологічно активний фрагмент ІМІ білка. NO:1 або SEQ ID NO:3 таким чином, щоб одне або Вжитий авторами термін "біологічно активна кілька амінокислотних заміщень, додавань або частина" ІМІ білка охоплює частину, наприклад, делецій були введені в кодований поліпептид. Мудомен/мотив ІМІ білка, який у разі вироблення в тації вводять у послідовність SEQ ID NO:1 або рослині підвищує резистентність рослини до імідаSEQ ID NO:3 стандартними способами, такими як золінонового гербіциду порівняно з різновидами сайт-специфічний мутагенез та ПЛР 23 87261 24 опосередкований мутагенез. В оптимальному ваідентичності двох нуклеїнових кислот виставляють ріанті консервативні амінокислотні заміщення показник "gap opening penalty" 15 і показник "gap здійснюють в одному або кількох прогнозованих extension penalty" 6,66. Для визначення відсотка замінних амінокислотних залишках. ідентичності двох поліпептидів виставляють пока"Консервативне амінокислотне заміщення" є зник "gap opening penalty" 10 і показник "gap таким, при якому амінокислотний залишок заміщуextension penalty" 0,1. Усі інші параметри встановється амінокислотним залишком, який має подіблено за умовчанням. Слід розуміти, що для визнаний боковий ланцюг. Спеціалістами було визначечення ідентичності послідовності, якщо порівнюно групи амінокислотних залишків, які мають ють послідовність ДНК з послідовністю РНК, подібні бокові ланцюги. До цих груп належать амітимідиновий нуклеотид є еквівалентом урациловонокислоти з основними боковими ланцюгами (наго нуклеотиду. В оптимальному варіанті виділені приклад, лізин, аргінін, гістидин), кислотними боІМІ поліпептиди, які охоплюються даним винахоковими ланцюгами (наприклад, аспарагінова дом, є принаймні приблизно на 50-60%, краще кислота, глутамінова кислота), незарядженими принаймні приблизно на 60-70%, ще краще - приполярними боковими ланцюгами (наприклад, глінаймні приблизно на 70-75%, 75-80%, 80-85%, 85цин, аспарагін, глутамін, серин, треонін, тирозин, 90% або 90-95%, найкраще - принаймні приблизно цистеїн), неполярними боковими ланцюгами (нана 96%, 97%, 98%, 99% або більше ідентичними приклад, аланін, валін, лейцин, ізолейцин, пролін, повній амінокислотній послідовності, кодованій фенілаланін, метіонін, триптофан), бетаполінуклеотидною послідовністю, показаною в розгалуженими боковими ланцюгами (наприклад, SEQ ID NO:1 або SEQ ID NO:3. В іншому варіанті треонін, валін, ізолейцин) та ароматичними боковтілення виділені ІМІ поліпептиди, які охоплюютьвими ланцюгами (наприклад, тирозин, фенілалася даним винаходом, є принаймні приблизно на нін, триптофан, гістидин). Таким чином, прогнозо50-60%, краще - принаймні приблизно на 60-70%, ваний замінний амінокислотний залишок в ІМІ ще краще - принаймні приблизно на 70-75%, 75поліпептиді оптимально заміщується іншим аміно80%, 80-85%, 85-90% або 90-95%, найкраще кислотним залишком з тієї самої групи бокових принаймні приблизно на 96%, 97%, 98%, 99% або ланцюгів. В альтернативному варіанті втілення більше ідентичними повній амінокислотній послімутації вводять випадково по всій ІМІ кодуючій довності, показаній у SEQ ID NO:2 або SEQ ID послідовності або її частині, наприклад, шляхом NO:4. насиченого мутагенезу, і одержані в результаті Крім того, можуть бути створені оптимізовані мутанти відбирають на наявність описаної автоІМІ нуклеїнові кислоти. В оптимальному варіанті рами ІМІ активності для розпізнання мутантів, які оптимізована ІМІ нуклеїнова кислота кодує ІМІ зберігають ІМІ активність. Після мутагенезу посліполіпептид, який модулює толерантність рослини довності SEQ ID NO:1 або SEQ ID NO:3, кодований до імідазолінонових гербіцидів, ще краще - якщо поліпептид може бути експресований рекомбінанпідвищує толерантність рослини до імідазолінонотно, і активність поліпептиду може бути визначена вого гербіциду після його переекспресії в рослині. шляхом аналізу резистентності до імідазолінону в Вжитий авторами термін "оптимізований" стосурослини, яка експресує поліпептид, як описано ється нуклеїнової кислоти, підданої генній інжененижче у Прикладах. рії для посилення її експресії в даній рослині або Для визначення відсотка ідентичності послідотварині. Для забезпечення оптимізованих для ровності двох амінокислотних послідовностей (наслини ІМІ нуклеїнових кислот послідовність ДНК приклад, SEQ ID NO:2 або SEQ ID NO:4 та їх мугена модифікують, щоб вона 1) включала кодони, тантної форми) послідовності суміщують для оптимальні для високоекспресованих рослинних оптимального порівняння (наприклад, у послідовгенів; 2) включала А+Т, який практично відповідає ність одного поліпептиду вставляють розриви для за складом нуклеотидної основи тому, що міститьоптимального суміщення з іншим поліпептидом). ся в рослинах; 3) утворювала ініціюючу послідовПісля цього порівнюють амінокислотні залишки у ність рослини, 4) не містила послідовностей які відповідних позиціях амінокислот. Якщо позиція в викликають дестабілізацію, неналежне поліаденіодній послідовності (наприклад, SEQ ID NO:2 або лювання, деградацію та термінацію РНК, або які SEQ ID NO:4) займається тим самим амінокислотутворюють шпильки вторинної структури або місця ним залишком, що й відповідна позиція в іншій сплайсингу РНК. Посилення експресії ІМІ нуклеїпослідовності (наприклад, мутантній формі SEQ ID нових кислот у рослинах досягають шляхом застоNO:2 або SEQ ID NO:4), то молекули в цій позиції є сування частоти розподілу використання кодонів у ідентичними. Таке саме порівняння здійснюють рослинах взагалі або в конкретній рослині. Спосоміж двома послідовностями нуклеїнових кислот. би оптимізації експресії нуклеїнових кислот у росВідсоток ідентичності послідовності між двома линах описано в [ЕРА 0359472; ЕРА 0385962; запослідовностями залежить від кількості ідентичних явці РСТ № WO 91/16432; патенті США позицій, спільних для цих послідовностей (тобто №5,380,831; патенті США №5,436,391; роботах відсоток ідентичності послідовностей = кількість Perlack et al., 1991 Proc. Natl. Acad. Sci. USA ідентичних позицій/загальна кількість пози88:3324-3328; та Murray et al., 1989 Nucleic Acids Res. 17:477-498]. цій´100). З точки зору винаходу відсоток ідентичВжитий авторами термін "частота переважного ності послідовностей між двома нуклеїновими кисвикористання кодонів" стосується переваги, яку лотами або поліпептидними послідовностями віддає конкретна клітина-хазяїн при використанні визначають, застосовуючи пакет програм Vector нуклеотидних кодонів для визначення даної аміноNTI 6.0 (PC) (InforMax, 7600 Wisconsin Ave., кислоти. Для визначення частоти використання Bethesda, MD 20814). Для визначення відсотка 25 87261 26 конкретного кодону в гені кількість випадків цього Термін "антисмислова" з точки зору винаходу кодону в гені ділять на загальну кількість випадків стосується нуклеїнової кислоти, яка включає поліусіх кодонів, які вказують на ту саму амінокислоту нуклеотид, який є достатньо комплементарним, в гені. Так само частота переважного використанповністю або частково, генові, первинному трансня кодонів, яку виявляє клітина-хазяїн, може бути криптові або процесованій мРНК, щоб впливати на розрахована через середнє значення частоти пеекспресію ендогенного гена. "Комплементарними" реважного використання кодонів у великій кількосполінуклеотидами є ті, які здатні до спарювання ті генів, експресованих клітиною-хазяїном. В оптиоснов згідно зі стандартними правилами комплемальному варіанті цей аналіз обмежують генами, ментарності Вотсона-Кріка. Зокрема, пурини ствоякі сильно експресуються клітиною-хазяїном. Відрюють пари основ з піримідинами для утворення соток відхилення частоти переважного викорискомбінації гуаніну, спареного з цитозином (G:C) та тання кодонів для синтетичного гена від частоти аденіну, спареного або з тиміном (А:Т) у разі ДНК, використання клітиною-хазяїном розраховують або аденіну, спареного з урацилом (A:U) у разі спочатку шляхом визначення відсотка відхилення РНК. Слід розуміти, що два полінуклеотиди мочастоти використання одного кодону від частоти жуть гібридизуватися один з одним, навіть якщо використання клітиною-хазяїном з наступним одевони не є повністю комплементарними один одноржанням середнього показника відхилення по всіх му, якщо кожен має принаймні одну ділянку, праккодонах. Як визначено авторами, цей розрахунок тично комплементарну іншій. Термін "антисмисловключає унікальні кодони (тобто ATG і TGG). Тобва нуклеїнова кислота" охоплює експресійні касети то загальне середнє відхилення використання коодноланцюгової РНК, а також дволанцюгової ДНК, донів оптимізованого гена від показника клітинаякі можуть бути транскрибовані для утворення хазяїна розраховують, користуючись рівнянням антисмислової РНК. "Активними" антисмисловими 1А=n=1ΖХп - ΥnΧn разів 100 Ζ, де Χn=частота виконуклеїновими кислотами є молекули антисмислористання для кодону n у клітині-хазяїні; Υn=частота вої РНК, здатні селективно гібридизуватися з первикористання для кодону n у синтетичному гені, n винним транскриптом або мРНК, що кодує поліпепредставляє окремий кодон, який визначає аміноптид, який має принаймні 80% ідентичність кислоту, і загальна кількість кодонів дорівнює Ζ. послідовності з поліпептидом, кодованим полінукЗагальне відхилення частоти використання кодолеотидною послідовністю SEQ ID NO:1 або SEQ ID нів, А, для всіх амінокислот в оптимальному варіаNO:3. нті становить менше, ніж приблизно 25%, краще Крім вищеописаних ІМІ нуклеїнових кислот та менше, ніж приблизно 10%. поліпептидів, даний винахід охоплює ці нуклеїнові Таким чином, ІМІ нуклеїнова кислота може букислоти та поліпептиди, приєднані до компонента. ти оптимізована таким чином, щоб її частота розДо цих компонентів, крім інших, належать компоподілу використання кодонів відхилялася в оптиненти виявлення, компоненти гібридизації, компомальному варіанті не більше, ніж на 25% від ненти очищення, компоненти доставления, компочастоти високоекспресованих рослинних генів, у ненти реакції, компоненти зв'язування та ін. ще кращому варіанті - не більше, ніж приблизно на Типовою групою нуклеїнових кислот, які мають 10%. Крім того, враховують відсотковий вміст G+C приєднані компоненти, є зонди та праймери. Зонвиродженої третьої основи (однодольні підтримуди та праймери, як правило, включають практично ють G+C у цій позиції, на відміну від дводольних). ізольований олігонуклеотид. Олігонуклеотид, як Визнано також, що XCG (де X є А, Т, С, або G) правило, включає ділянку нуклеотидної послідовнуклеотид є найменшим оптимальним кодоном у ності, яка гібридизується за жорстких умов з придводольних, тоді як ХТА кодону уникають як однонаймні приблизно 12, краще - приблизно 25, ще дольні, так і дводольні. Оптимізовані ІМІ нуклеїнові краще - приблизно 40, 50 або 75 послідовними кислоти цього винаходу також бажано мати індекнуклеотидами смислового ланцюга послідовності, си уникнення дублетів CG і ТА, наближені до індевказаної в SEQ ID NO:1 або SEQ ID NO:3, антиксів вибраної рослини-хазяїна (тобто Triticum смисловою послідовністю послідовності, вказаної aestivum). Краще, якщо ці індекси відхиляються від в SEQ ID NO:1 або SEQ ID NO:3, або їх природниіндексу хазяїна не більше, ніж приблизно 10-15%. ми мутантами. Праймери на основі нуклеотидної Крім молекул нуклеїнових кислот, які кодують послідовності SEQ ID NO:1 або SEQ ID NO:3 заописані вище ІМІ поліпептиди, інший аспект винастосовують у ПЛР для клонування ІМІ гомологів. ходу стосується виділених молекул нуклеїнових Зонди на основі ІМІ нуклеотидних послідовностей кислот, які є антисмисловими до них. Вважається, "застосовують для виявлення транскриптів або що антисмислові полінуклеотиди інгібують генну геномних послідовностей, які кодують ті ж самі або експресію полінуклеотиду-мішені шляхом специгомологічні поліпептиди. В оптимальних варіантах фічного зв'язування з полінуклеотидом-мішенню і втілення зонд також включає приєднану до нього впливу на транскрипцію, сплайсинг, перенесення, мітку, наприклад, мітку, яка може бути радіоізототрансляцію та/або стійкість полінуклеотиду-мішені. пом, флуоресцентною сполукою, ферментом або В існуючому рівні техніки описано способи спрякофактором ферменту. Такі зонди застосовують як мування антисмислового полінуклеотиду на хрочастину випробувального комплекту геномного мосомну ДНК, на первинний транскрипт РНК або маркера для розпізнання клітин, які експресують на процесовану мРНК. В оптимальному варіанті ІМІ поліпептид, наприклад, шляхом вимірювання ділянки-мішені включають місця сплайсингу, кодорівня ІМІ-кодуючої нуклеїнової кислоти, у зразку ни ініціації трансляції, кодони термінації трансляції клітин, наприклад, виявлення рівня ІМІ мРНК або та інші послідовності в межах відкритої рамки зчивизначення, чи був геномний ІМІ ген мутованим чи тування. видаленим. 27 87261 28 Винахід також забезпечує виділений рекомбіють експресію нуклеотидної послідовності лише в нантний вектор експресії, який включає ІМІ нуклеїдеяких клітинах-хазяях або за певних умов. Спецінову кислоту, як описано вище, причому вектор алістам стане зрозуміло, що побудова вектора експресії у клітині-хазяїні в результаті забезпечує експресії може залежати від таких чинників, як підвищену резистентність до імідазолінонового вибір клітини-хазяїна, яка підлягає трансформації, гербіциду порівняно з різновидом клітини-хазяїна рівень експресії потрібного поліпептиду і т. д. Векдикого типу. Вжитий авторами термін "вектор" стотори експресії згідно з винаходом вводять у клітисується молекули нуклеїнової кислоти, здатної ни-хазяї для утворення поліпептидів або пептидів, транспортувати іншу нуклеїнову кислоту, з якою її включаючи злиті поліпептиди або пептиди, кодобуло з'єднано. Одним типом вектора є "плазміда", вані нуклеїновими кислотами, як описано автораяка означає кільцеву дволанцюгову петлю ДНК, у ми (наприклад, ІМІ поліпептиди, злиті поліпептиди яку можуть бути ліговані додаткові сегменти ДНК. і т. д.). Іншим типом вектора є вірусний вектор, у якому В оптимальному варіанті втілення даного видодаткові сегменти ДНК можуть бути ліговані у находу ІМІ поліпептиди експресуються в рослинах вірусний геном. Деякі вектори здатні до самостійта рослинних клітинах, таких як клітини однокліної реплікації у клітині-хазяїні, в яку вони є введетинних рослин (наприклад, водоростей) [див. ними (наприклад, бактеріальні вектори, які мають Falciatore et al., 1999 Marine Biotechnology 1(3):239бактеріальне походження реплікації, та епісомні 251, та посилання, що містяться в цій роботі] та вектори ссавців). Інші вектори (наприклад, неепіклітини вищих рослин (наприклад, сперматофітів, сомні вектори ссавців) вводять у геном клітинитаких як хлібні злаки). ІМІ полінуклеотид може бути хазяїна після введення у клітину-хазяїн, а отже, "введений" у рослинну клітину будь-яким спосореплікують разом з геномом хазяїна. Крім того, бом, включаючи трансфекцію, трансформацію або деякі вектори здатні спрямовувати експресію генів, трансдукцію, електропорацію, бомбардування часз якими вони є функціонально зв'язаними. Такі тинками, агроінфекцію, біолістику і т. ін. Одним з вектори названо авторами "векторами експресії". відомих спеціалістам способів трансформації є Взагалі, вектори експресії, які застосовують у техзанурення рослини під час цвітіння в розчин нологіях рекомбінантних ДНК, часто мають форму Agrobacteria, в якому Agrobacteria містить ІМІ нукплазмід. У даному описі терміни "плазміда" та "велеїнову кислоту, з наступним розведенням трансктор" можуть вживатися поперемінно, оскільки формованих гамет. плазміда є найпоширенішою формою вектора. Інші відомості про придатні способи трансфоОднак, винахід охоплює й інші форми векторів рмації або трансфекції клітин-хазяїв, включаючи експресії, такі як вірусні вектори (наприклад, ретрослинні клітини, можна знайти в [Sambrook, et al. ровіруси з недостатньою реплікацією, аденовіруси (Molecular Cloning: A Laboratory Manual. 2nd, ed., та аденоасоційовані віруси), які виконують рівноCold Spring Harbor Laboratory, Cold Spring Harbor цінні функції. Laboratory Press, Cold Spring Harbor, NY, 1989) та Рекомбінантні вектори експресії згідно з винаінших лабораторних посібниках, таких як Methods ходом включають нуклеїнову кислоту винаходу у in Molecular Biology, 1995, Vol. 44, Agrobacterium формі придатної для експресії нуклеїнової кислоти protocols, ed: Gartland and Davey, Humana Press, у клітині-хазяїні, що означає, що рекомбінантні Totowa, New Jersey]. Оскільки підвищена резистевектори експресії включають одну або кілька регунтність до імідазолінонових гербіцидів є загальною ляторних послідовностей, вибраних на основі кліособливістю, успадкування якої є бажаним для тин-хазяїв, які мають бути застосовані для експребагатьох видів рослин, таких як кукурудза, пшенисії, які є функціонально зв'язаними з ця, жито, овес, тритикале, рис, ячмінь, соя, арахіс, нуклеїновокислотною послідовністю, яка підлягає бавовна, рапс та канола, маніхот, перець, соняшекспресії. По відношенню до рекомбінантного векник та чорнобривці, пасльонові рослини, такі як тора експресії "функціонально зв'язаний" означає, картопля, тютюн, баклажан та томати, види віки, що дана нуклеотидна послідовність є зв'язаною з горох, люцерна, чагарникові рослини (кава, какао, регуляторною(ими) послідовністю (послідовностячай), види верби, дерева (олійна пальма, кокос), ми) таким чином, що це дозволяє експресію нукбагаторічні трави та фуражні культури, ці хлібні леотидної послідовності (наприклад, в in vitro злаки також є оптимальними рослинами, які мотранскрипційній/трансляційній системі або у клітижуть бути об'єктами генної інженерії як ще один ні-хазяїні, коли вектор є введеним у клітинуваріант втілення даного винаходу. До фуражних хазяїн). Термін "регуляторна послідовність" охопкультур належать, крім інших, пирій, Phalaris лює промотори, енхансери та інші елементи контarundinacea, Bromopsis inermis, дике жито, ролю експресії (наприклад, сигнали поліаденілюBluegrass, Dactillis glomerata L., люцерна, Salfoin, вання). Такі регуляторні послідовності описано, Lotus comiculatus L., шведська конюшина, червона наприклад, у роботі [Goeddel, Gene Expression конюшина та буркун. Technology: Methods in Enzymology 185, Academic В одному варіанті втілення даного винаходу Press, San Diego, CA (1990), або в роботі Gruber трансфекції ІМІ полінуклеотиду в рослину досягаand Crosby, Methods in Plant Molecular Biology and ють шляхом Agrobacterium-опосередкованого пеBiotechnology, eds. Glick and Thompson, Chapter 7, ренесення гена. Agrobacterium-опосередкована 89-108, CRC Press: Boca Raton, Florida, включаючи трансформацію рослин здійснюють, застосовуючи, посилання, які в них містяться]. До регуляторних наприклад, GV3101(pMP90) [Koncz and Schell, послідовностей належать ті, які спрямовують кон1986 Mol. Gen. Genet. 204:383-396] або LBA4404 ститутивну експресію нуклеотидної послідовності в (Clontech) штам Agrobacterium tumefaciens. багатьох типах клітин-хазяїв, та ті, які спрямовуТрансформацію здійснюють стандартними спосо 29 87261 30 бами трансформації та регенерації [Deblaere et al., ендогенним геном AHAS у мікроорганізмі або рос1994 Nucl. Acids. Res. 13:4777-4788; Gelvin, лині. Додаткова молекула фланкуючої нуклеїнової Stanton B. and Schiiperoort, Robert A, Plant кислоти AHAS має достатню довжину для успішної Molecular Biology Manual, 2nd Ed. - Dordrecht : гомологічної рекомбінації ендогенним геном. Як Kluwer Academic Publ., 1995. - in Sect., Ringbuc правило, вектор включає від кількох сотень пар Zentrale Signatur: BT11-P ISBN 0-7923-2731-4; основ до тисяч основ фланкуючих ДНК (як на 5', Glick, Bernard R. and Thompson, John E., Methods так і на 3' кінцях) [див., наприклад, Thomas, К. R., in Plant Molecular Biology and Biotechnology, Boca and Capecchi, Μ. R., 1987 Cell 51:503, де описано Raton : CRC Press, 1993 360 S., ISBN 0-8493-5164вектори гомологічної рекомбінації, або Strepp et 2]. Наприклад, рапс трансформують шляхом al., 1998 PNAS, 95(8):4368-4373, де описано рекотрансформації сім'ядолі або гіпокотиля [Moloney ef мбінацію на основі кДНК у Physcomitrella patens]. al., 1989 Plant cell Report 8:238-242; De Block ef al., Однак, оскільки ІМІ ген зазвичай відрізняється від 1989 Plant Physiol. 91:694-701]. Застосування ангена AHAS у дуже небагатьох амінокислотах, у тибіотиків для вибору Agrobacterium та рослини фланкуючій послідовності не завжди є необхідзалежить від бінарного вектора та штаму ність. Вектор гомологічної рекомбінації вводять у Agrobacterium, який застосовують для трансформікроорганізм або рослинну клітину (наприклад, мації. Вибір рапсу зазвичай здійснюють, застосочерез поліетиленгліколь-опосередковану ДНК) і вуючи канаміцин як маркер для селекції рослин. відбирають клітини, в яких введений ІМІ ген було Agrobacterium-опосередковане перенесення генів гомологічно рекомбіновано ендогенним геном у льон здійснюють, застосовуючи, наприклад, споAHAS, застосовуючи відомі спеціалістам способи. сіб, описаний у роботі [Mlynarova et al., 1994 Plant В іншому варіанті втілення одержують рекомCell Report 13:282-285]. Крім того, трансформацію бінантні мікроорганізми, які містять вибрані систесої здійснюють, застосовуючи, наприклад, спосіб, ми, що дозволяють здійснювати регульовану ексописаний у європейському патенті №0424047, папресію введеного гена. Наприклад, включення ІМІ тенті США №5,322,783, європейському патенті гена у вектор з поміщенням його під контроль Іас№0397 687, патенті США №5,376,543 або патенті оперона дозволяє експресію ІМІ гена лише у приСША №5,169,770. Трансформації кукурудзи досясутності IPTG. Такі регуляторні системи добре гають шляхом бомбардування частинками, опосевідомі спеціалістам. редкованого поліетиленгліколем поглинання ДНК Чи є він присутнім у позахромосомному невідабо за допомогою волокон карбіду кремнію. [Див., творюваному векторі, чи у векторі, введеному у наприклад, Freeling and Walbot "The maize хромосому, ІМІ полінуклеотид в оптимальному handbook" Springer Verlag: New York (1993) ISBN 3варіанті містяться в експресійній касеті рослини. 540-97826-7]. Конкретний приклад трансформації Експресійна касета рослини в оптимальному варікукурудзи міститься в патенті США №5,990,387, а анті містить регуляторні послідовності, здатні виконкретний приклад трансформації пшениці міскликати експресію гена в рослинних клітинах, які є титься в заявці РСТ № WO 93/07256. функціонально зв'язаними таким чином, що кожна Згідно з даним винаходом, введений ІМІ поліпослідовність може виконувати свою функцію, нануклеотид може стійко утримуватись у рослинній приклад, термінацію транскрипції сигналами поліклітині, якщо його введено в нехромосомний автоаденілювання. Оптимальними сигналами поліаденомний реплікон або введено у хромосоми рослин. нілювання є ті, що походять із т-ДНК Agrobacterium В альтернативному варіанті введений ІМІ полінукtumefaciens, такі як ген 3, відомий як октопін синлеотид може бути присутній на позахромосомному таза Ті-плазміди pTiACHS [Gielen et al., 1984 невідтворюваному векторі і може бути короткочасEMBO J. 3:835] або їх функціональні еквіваленти, но експресованим або короткочасно активним. В але придатними є також усі інші термінатори, фунодному варіанті втілення може бути створений кціонально активні в рослинах. Оскільки експресія гомологічний рекомбінантний мікроорганізм, у рослинних генів дуже часто не обмежується на якому ІМІ полінуклеотид є введеним у хромосому, транскрипційних рівнях, експресійна касета рослистворюється вектор, який містить принаймні часни в оптимальному варіанті містить інші функціотину гена AHAS, у який введено делецію, доданально зв'язані послідовності, такі як перехідні вання або заміщення для того, щоб змінити, наенхансери, наприклад, надлишкова послідовність приклад, функціонально розірвати, ендогенний ген (overdrive sequence), яка містить 5'AHAS і створити ІМІ ген. Для створення точкової нетрансльовану лідерну послідовність вірусу тюмутації через гомологічну рекомбінацію, використюнової мозаїки, що збільшує пропорцію поліпептовують ДНК-РНК гібриди, застосовуючи спосіб, тиду в РНК [Gallic et al., 1987 Nucl. Acids Research відомий як химерапластія [Cole-Strauss et al., 1999 15:8693-8711]. Прикладами рослинних векторів Nucleic Acids Research 27(5): 1323-1330 and Kmiec, експресії можуть бути ті, які детально описано в 1999 Gene therapy American Scientist 87(3):240роботах [Becker, D. et al., 1992 New plant binary 247]. Спеціалістам також добре відомі інші процеvectors with selectable markers located proximal to дури гомологічної рекомбінації у виді Triticum, моthe left border, Plant Mol. Biol. 20:1195-1197; Bevan, жливість застосування яких авторами передбачаM.W., 1984 Binary Agrobacterium vectors for plant ється. transformation, Nucl. Acid. Res. 12:8711-8721; and У векторі гомологічної рекомбінації до ІМІ гена Vectors for Gene Transfer in Higher Plants; in: на його 5' та 3' кінцях може приєднуватися додатTransgenic Plants, Vol. 1, Engineering and Utilization, кова молекула нуклеїнової кислоти гена AHAS, що eds.: Kung and R. Wu, Academic Press, 1993, S. 15дозволяє здійснення гомологічної рекомбінації між 38]. екзогенним ІМІ геном, що міститься у векторі, та 31 87261 32 Експресія рослинного гена має бути функціоспецифічні до пелюсток, специфічні до чашолистнально пов'язана з відповідним промотором, який ка, специфічні до квітконіжки, специфічні до стручзабезпечує вчасну, клітино- або тканиноспецифічка, специфічні до суплідь, специфічні до коріння ну експресію гена. До промоторів, застосовуваних промотори та інші. Специфічні до насіння промов експресійних касетах згідно з винаходом, може тори в оптимальному варіанті експресуються під належати будь-який промотор, здатний ініціювати час розвитку насіння або проростання. Наприклад, транскрипцію в рослинній клітині. До таких промоспецифічні до насіння промотори можуть бути торів належать, крім інших, ті, які можуть бути специфічними до ембріонів, специфічними до енодержані з рослин, рослинних вірусів та бактерій, досперму та специфічними до оболонки насіння. які містять гени, що експресуються в рослинах, [Див. Thompson et al. 1989 BioEssays 10:108]. Притаких як Agrobacterium та Rhizobium. кладами специфічних до насіння промоторів є, Промотор може бути конститутивним, індуцикрім інших, целюлоза-синтаза (сеІА), Сіm1, гаммабельним, специфічним до стадії розвитку, специзеїн, глобулін-1, зеїн кукурудзи 19kD (cZ19BI) та фічним до типу клітин, тканиноспецифічним або інші. органоспецифічним. Конститутивні промотори є До інших придатних тканиноспецифічних або активними за більшості умов. Прикладами констиорганоспецифічних промоторів належать промотутивних промоторів є CaMV 19S та 35 S промототор напін-гена з рапсу [Патент США №5,608,152], ри [Odell et al. 1985 Nature 313:810-812], sX CaMV USP-промотор із Vicia faba [Baeumlein et al., 1991 35S промотор [Kay et al. 1987 Science 236:1299Mol Gen Genet. 225(3):459-67], олеозин-промотор 1302] Sep1 промотор, промотор актину рису із Arabidopsis [Заявка РСТ № WO 98/45461], фазе[McElroy et al. 1990 Plant Cell 2:163-171], промотор олін-промотор із Phaseolus vulgaris [Патент США актину Arabidopsis, промотор убіхітану [Christensen №5,504,200], Все4-промотор із Brassica [Заявка et al. 1989 Plant Molec Biol. 18:675-689]; pEmu [Last РСТ № WO 91/13980] або легумін В4 промотор ef al. 1991 Theor Appl Genet. 81:581-588], 35S про[LеВ4; Baeumlein et al., 1992 Plant Journal, 2(2):233мотор вірусу мозаїки ранника, Smas промотор 9], а також промотори, які забезпечують специфіч[Velten ef al. 1984 EMBO J. 3:2723-2730], GRP1-8 ну до насіння експресію в однодольних рослинах, промотор, промотор дегідрогенази цинамілового таких як кукурудза, ячмінь, пшениця, жито, рис і т. спирту [Патент США №5,683,439], промотори з Тд. Придатними промоторами є промотор 1pt2 або ДНК Agrobacterium, такі як манопін синтаза, нопаIpt1-гена з ячменю [Заявка PCX № WO 95/15389 та лін синтаза та октопін синтаза, промотор малої Заявка PCX № WO 95/23230] або описані в заявці субодиниці рибулоза біфосфат карбоксилази PCX № WO 99/16890 (промотори з гордеїн-гена (ssuRUBISCO) промотор та ін. ячменю, глютелін-гена рису, оризин-гена рису, Індуцибельні промотори є активними за певпроламін-гена рису, гліадин-гена пшениці, глютених навколишніх умов, таких як наявність або віднін-гена пшениці, глютенін-гена вівса, касиринсутність поживної речовини або метаболіту, тепло гена сорго та секалін-гена жита). або холод, світло, напад патогенів, анаеробні умоДо інших промоторів, які застосовують в ексви та ін. Наприклад, hsp80 промотор із Brassica пресійних касетах винаходу, належать, крім інших, індукується термічним ударом, PPDK промотор головний промотор a/b-зв'язувального білка хлоіндукується світлом, PR-1 промотор з тютюну, рофілу, промотори гістону, Ар3 промотор, промоArabidopsis та кукурудзи індукуються інфекцією тор конгліцину, промотор напіну, промотор соєвого патогену, a Adh1 промотор індукується гіпоксією та лектину, промотор зеїну кукурудзи 15kD, промотор впливом холоду. Експресії рослинного гена також зеїну 22kD, промотор зеїну 27kD, промотор gможе сприяти індуцибельний промотор [див. Gatz, зеїну, промотори waxy, shrunken 1, shrunken 2 та 1997 Annu. Rev. Plant Physiol. Plant Mol. Biol. 48:89bronze, Zm13 промотор [Патент США №5,086,169], 108]. Хімічно індуцибельні промотори є особливо промотори полігалактуронази кукурудзи (PG) [Папридатними, якщо потрібна специфічна до часу тенти США №№ 5,412,085 та 5,545,546] і SGB6 експресія гена. Прикладами таких промоторів є промотор [Патент США №5,470,359], а також синіндукований саліциловою кислотою промотор [Затетичні або інші природні промотори. явка РСТ № WO 95/19443], індукований тетрацикДодаткової гнучкості в контролюванні експресії ліном промотор [Gatz et al., 1992 Plant J. 2:397-404] гетерологічного гена в рослинах досягають шлята індукований етанолом промотор [Заявка РСТ № хом застосування доменів зв'язування ДНК та WO 93/21334]. елементів відповіді з гетерологічних джерел (тобто Специфічні до стадії розвитку промотори в опдоменів зв'язування ДНК з нерослинних джерел). тимальному варіанті експресуються на певних Прикладом такого гетерологічного домену зв'язустадіях розвитку. До тканино- та органоспецифічвання є домен зв'язування ДНК LexA [Brent and них промоторів належать ті, які в оптимальному Ptashne, Cell 43:729-736 (1985)]. варіанті експресуються в певних тканинах або орІнший аспект винаходу стосується клітинганах, таких як листя, коріння, насіння або ксилехазяїв, у які було введено рекомбінантний вектор ма. Прикладами тканиноспецифічних та органоекспресії згідно з винаходом. Терміни "клітинаспецифічних промоторів є, крім інших, специфічні хазяїн" та "рекомбінантна клітина-хазяїн" вживадо плодів, специфічні до насінних зачатків, специються авторами поперемінно. Слід розуміти, що фічні до чоловічих тканин, специфічні до насіння, такі терміни стосуються не лише конкретної клітиспецифічні до шкірки, специфічні до бульб, спени, але вони також стосуються потомства або моцифічні до стебел, специфічні до оплодня та спежливого потомства такої клітини. Оскільки в нацифічні до листя, специфічні до приймочок, спеступних поколіннях можуть траплятися певні зміни цифічні до пилку, специфічні до пиляка, через мутації або вплив навколишнього середо 33 87261 34 вища, таке потомство фактично не може бути ідевідмінних від ІМІ хімічних речовин, краще - менше, нтичним батьківській клітині, але все одно охопніж приблизно 20% хімічних попередників або відлюється вжитим авторами терміном. Клітинамінних від ІМІ хімічних речовин, ще краще - менхазяїн може бути будь-якою прокаріотною або еуше, ніж приблизно 10% хімічних попередників або каріотною клітиною. Наприклад, ІМІ полінуклеотид відмінних від ІМІ хімічних речовин, найкраще - меможе бути експресований у бактеріальних клітинше, ніж приблизно 5% хімічних попередників або нах, таких як С glutamicum, клітини комах, грибкові відмінних від ІМІ хімічних речовин. В оптимальних клітини або клітини ссавців (такі як клітини яєчника варіантах втілення виділені поліпептиди або їх китайського хом'яка (СНО) або клітини COS), вобіологічно активні частини не містять забруднююдорості, війчасті, рослинні клітини, грибки або інші чих поліпептидів з того організму, з якого походить мікроорганізми, такі як С glutamicum. Інші придатні ІМІ поліпептид. Як правило, такі поліпептиди одеклітини-хазяї є відомими спеціалістам. ржують шляхом рекомбінантної експресії, наприКлітину-хазяїн згідно з винаходом, таку як проклад, ІМІ поліпептиду Triticum aestivum у рослинах, каріотна або еукаріотна клітина-хазяїн у культурі, які не є Triticum aestivum, або мікроорганізмах, використовують для вироблення (тобто експресії) таких як С glutamicum, війчасті, водорості або гриІМІ полінуклеотиду. Відповідно, винахід також забки. безпечує способи вироблення ІМІ поліпептидів з Послідовності ІМІ полінуклеотиду та поліпепвикористанням клітин-хазяїв згідно з винаходом. В тиду згідно з винаходом мають різне застосування. одному варіанті втілення спосіб включає культивуНуклеїновокислотні та амінокислотні послідовності вання клітини-хазяїна згідно з винаходом (у яку даного винаходу застосовують для трансформації було введено рекомбінантний вектор експресії, рослин, таким чином, модулюючи резистентність який кодує ІМІ поліпептид, або в геном якої було рослин до імідазолінонових гербіцидів. Відповідно, введено ген, який кодує поліпептид дикого типу винахід забезпечує спосіб одержання трансгенної або ІМІ поліпептид) у придатному середовищі до рослини, яка має підвищену толерантність до імівироблення ІМІ поліпептиду. В іншому варіанті дазолінонового гербіциду, включаючи (а) трансвтілення спосіб також включає виділення ІМІ поліформацію рослинної клітини одним або кількома пептидів із середовища або клітини-хазяїна. Інший векторами експресії, які включають одну або кільаспект винаходу стосується виділених ІМІ поліпепка ІМІ нуклеїнових кислот, і (б) вирощування з ротидів та їх біологічно активних частин. "Виділений" слинної клітини трансгенної рослини з підвищеною або "очищений" поліпептид або його біологічно резистентністю до імідазолінонового гербіциду активна частина є вільними від деякого клітинного порівняно з сортом рослини дикого типу. В одному матеріалу, якщо одержуються способами рекомбіваріанті втілення множинні ІМІ нуклеїнові кислоти нантних ДНК, або хімічних попередників або інших походять із різних геномів. Даний винахід також хімічних речовин, якщо хімічно синтезуються. Виохоплює способи одержання трансгенної рослини, раз "практично вільний від клітинного матеріалу" яка має підвищену толерантність до імідазоліноохоплює препарати ІМІ поліпептиду, в яких полінового гербіциду, включаючи (а) трансформацію пептид є відокремленим від деяких із клітинних рослинної клітини вектором експресії, який вклюкомпонентів клітин, у яких він природним або речає ІМІ нуклеїнову кислоту, причому нуклеїнова комбінантним шляхом виробляється В одному кислота не є lmi1 нуклеїновою кислотою, і (б) виваріанті втілення вираз "практично вільний від рощування з рослинної клітини трансгенної росликлітинного матеріалу" охоплює препарати ІМІ пони з підвищеною резистентністю до імідазоліноноліпептиду, що мають менше, ніж приблизно 30% вого гербіциду порівняно з сортом рослини дикого (сухої маси) відмінного від ІМІ матеріалу (який татипу. кож вказується авторами як "забруднюючий поліДаний винахід включає способи зміни толерапептид"), краще - менше, ніж приблизно 20% віднтності рослини до імідазолінонового гербіциду, мінного від ІМІ матеріалу, ще краще - менше, ніж які включають зміну експресії однієї або кількох ІМІ приблизно 10% відмінного від ІМІ матеріалу, найнуклеїнових кислот. В оптимальному варіанті нуккраще - менше, ніж приблизно 5% відмінного від леїнові кислоти містяться в різних геномах або ІМІ матеріалу. походять із різних геномів. Резистентність рослини Якщо ІМІ поліпептид або його біологічно актидо імідазолінонового гербіциду може бути підвивна частина виробляється рекомбінантно, вони щена або знижена, що досягається посиленням також в оптимальному варіанті є практично вільабо послабленням експресії ІМІ полінуклеотиду, ними від культурального середовища, тобто кульвідповідно. В оптимальному варіанті резистенттуральне середовище складає менше, ніж приблиність рослини до імідазолінонового гербіциду підзно 20%, краще - менше, ніж приблизно 10%, вищують шляхом посилення експресії ІМІ полінукнайкраще - менше, ніж приблизно 5% об'єму прелеотиду. Експресія ІМІ полінуклеотиду може парату поліпептиду. Вираз "практично вільний від змінюватися будь-якими відомими спеціалістам хімічних попередників або інших хімічних речовин" способами. Застосовують способи посилення ексохоплює препарати ІМІ поліпептиду, в яких поліпресії ІМІ полінуклеотидів, у яких рослина є або пептид є відокремленим від хімічних попередників трансгенною, або нетрансгенною. У випадках, коабо інших хімічних речовин, які беруть участь у ли рослина є трансгенною, рослина може бути синтезі поліпептиду. В одному варіанті втілення трансформована вектором, який містить будь-яку вираз "практично вільний від хімічних попередниз вищеописаних ІМІ кодуючих нуклеїнових кислот, ків або інших хімічних речовин" охоплює препараабо ж, рослина може бути трансформована проти ІМІ поліпептиду, що мають менше, ніж приблизмотором, який спрямовує експресію ендогенних но 30% (сухої маси) хімічних попередників або ІМІ полінуклеотидів у рослині. Винахід передба 35 87261 36 чає, що такий промотор може бути тканиноспеци10-400г аі/га-1, краще - 20-160г аі/га-1, найкраще фічним або регульованим розвитком. В альтерна40-80г аі/га-1. Вжитий авторами термін "витримувативному варіанті нетрансгенні рослини можуть ти" нанесення імідазолінонового гербіциду ознамати експресію ендогенного ІМІ полінуклеотиду, чає, що рослина не знищується або не зазнає яка змінюється включенням природного промотоураження від такого нанесення. ра. Експресія полінуклеотидів, які включають SEQ Крім того, авторами пропонується спосіб контID NO:1 або SEQ ID NO:3 у заданих рослинах моролю над бур'янами в оточенні пшеничної культуже здійснюватися, крім іншого, за допомогою: (а) ри, який включає нанесення імідазолінонового конститутивного промотора, (б) хімічно індукованогербіциду на бур'яни та пшеничну культуру, приго промотора та (в) переекспресії підданого інжечому пшенична культура має підвищену резистеннерії промотора, наприклад, похідними від цинкотність до імідазолінонового гербіциду порівняно з вих пальців факторами транскрипції [Greisman and сортом пшеничної культури дикого типу, і рослина Pabo, 1997 Science 275:657]. містить одну або кілька ІМІ нуклеїнових кислот. В В оптимальному варіанті втілення транскриподному варіанті втілення рослина містить множинцію ІМІ полінуклеотиду модулюють, застосовуючи ні ІМІ нуклеїнові кислоти, які містяться в різних похідні від цинкових пальців фактори транскрипції геномах або походять із різних геномів. В іншому (ZFP), які описано у [Greisman and Pabo, 1997 варіанті втілення рослина містить відмінну від Іті1 Science 275:657], і які виробляються Sangamo нуклеїнову кислоту. Завдяки забезпеченню пшеBiosciences, Inc. Ці ZFP включають як домен розпіничних культур з підвищеною резистентністю до знання ДНК, так і функціональний домен, який імідазолінону, існує широкий вибір придатних до викликає активацію або пригнічення заданої нуклезастосування композицій для захисту пшеничних їнової кислоти, такої як ІМІ нуклеїнова кислота. культур від бур'янів з метою активізації росту росОтже, створюють активуючі і пригнічувальні ZFP, лин та зниження конкурентної боротьби за поживні які специфічно розпізнають промотори ІМІ полінуречовини. Імідазоліноновий гербіцид застосовують клеотиду, які було описано вище і які застосовують окремо для досходового, післясходового, переддля посилення або послаблення експресії ІМІ попосівного та посівного контролю над бур'янами на лінуклеотиду в рослині, таким чином, модулюючи площах, які оточують описані авторами пшеничні резистентність рослини до гербіцидів. культури, або ж застосовують композицію імідазоЯк було детальніше описано вище, рослини, лінонового гербіциду, яка містить інші додатки. які одержують способами даного винаходу, моІмідазоліноновий гербіцид також застосовують для жуть бути однодольними або дводольними. Рособробки насіння. До додатків, які містяться в комлини вибирають, наприклад, з-поміж кукурудзи, позиції імідазолінонового гербіциду, належать інші пшениці, жита, вівса, тритикале, рису, ячменю, сої, гербіциди, детергенти, ад'юванти, засоби розсіюарахісу, бавовни, рапсу, каноли, маніхоту, перцю, вання, засоби прилипання, стабілізатори або інші соняшника, чорнобривців, пасльонових рослин, подібні агенти. Композиція імідазолінонового геркартоплі, тютюну, баклажану, томатів, видів віки, біциду може бути рідкою або сухою композицією і гороху, люцерни, кави, какао, чаю, видів верби, може включати, крім іншого, сипкі порошки, емульолійної пальми, кокосу, багаторічних трав та фуговані концентрати та рідкі концентрати. Імідазоліражних культур. До фуражних культур належать, ноновий гербіцид та гербіцидні композиції нанокрім інших, пирій, Phalaris arundinacea, Bromopsis сять традиційними способами, наприклад, шляхом inermis, дике жито, Bluegrass, Dactillis glomerata L, розпилення, поливання, зрошування або іншими люцерна, Salfoin, Lotus corniculatus L, шведська подібними способами. конюшина, червона конюшина та буркун. В оптиУ заявці містяться посилання на різні публікамальному варіанті втілення рослиною є пшенична ції. Описи всіх цих публікацій та посилання, навекультура. У кожному з вищеописаних способів родені в цих публікаціях у їх повному обсязі, включеслинна клітина включає, крім іншого, протопласт, но авторами шляхом посилання в цю заявку для гаметопродукуючу клітину та клітину, яка регенебільш повного опису рівня техніки, якого стосуєтьрується в цілу рослину. Вжитий авторами термін ся цей винахід. "трансгенний" стосується будь-якої рослини, росСлід також розуміти, що вищенаведене стосулинної клітини, калюсу, рослинної тканини або ється оптимальних варіантів втілення даного вичастини рослини, що містить, повністю або частконаходу, і що існує можливість внесення численних во, принаймні один рекомбінантний полінуклеотид. змін без відхилення від обсягу винаходу. Винахід У багатьох випадках рекомбінантний полінуклеодалі пояснюється на представлених нижче притид, частково або повністю, є стабільно включекладах, які не повинні розглядатись як такі, що ним у хромосому або стійкий позахромосомний якимось чином обмежують його обсяг. Навпаки, елемент таким чином, що він передається наступслід чітко розуміти, що існує можливість звернення ним поколінням. до різних інших варіантів його втілення, модифікаЯк описано вище, даний винахід пропонує цій та еквівалентів, які, по ознайомленню з предкомпозиції та способи підвищення резистентності ставленим описом, стануть зрозумілими спеціалісдо імідазолінону у пшеничної культури або насіння там без відхилення від сутності даного винаходу порівняно з різновидами рослини або насінням та/або обсягу формули винаходу, що додається. дикого типу. В оптимальному варіанті втілення Приклади резистентність до імідазолінону у пшеничної кульПриклад 1 тури або насіння підвищується таким чином, що Мутагенез та відбір резистентних ліній пшенирослина'або насіння може витримувати нанесення ці імідазолінонового гербіциду в кількості приблизно 37 87261 38 Приблизно 40000 зразків насіння Triticum 8´16, на стадії двох листків, використовуючи шлаaestivum L. cv CDC Teal [Hughes and Hucl, 1993 нговий розпилювач, калібрований таким чином, Can. J. Plant Sci. 73:193-197] піддавали мутагенещоб розпилювати 100л га-1. Імазамокс наносили зові, застосовуючи змінені процедури, описані в на рослини при нормі 20г аі/га-1, застосовуючи нароботі [Washington and Sears (1970)]. Насіння посадку 8001 EVS під тиском 275кПа. До розчину передньо вимочували в дистильованій воді протягербіциду перед нанесенням додавали поверхнегом чотирьох годин, після чого обробляли 0,3% во-активну речовину для злиття (0,05% (об'EMS протягом шести годин. Насіння постійно проєм/об'єм)). Через п'ятнадцять днів після нанесення мивали водопровідною водою протягом семи гогербіциду рослини розбивали по групах на основі дин і залишали висихати протягом приблизно чопервинної реакції і поділяли на резистентні, протирьох годин перед висіванням у полі. Рослини М1 міжні або чутливі. Резистентні рослини після герзмішували ι насіння збирали в загальну масу. біцидної обробки були фенотипічно неушкоджеПриблизно 2´106 рослин М2 вирощували в полі ними, тоді як проміжні рослини характеризувалися затримкою росту перших двох листків, потемніннаступного року і обприскували на стадії двох лисням (темно-зеленим забарвленням) листя та потків імазамоксом при нормі 40г аі/га-1 в об'ємі аеявою колеоптильних відростків. Чутливі рослини розолю 100л га-1. До розчину аерозолю додавали характеризувалися відсутністю розвитку нових допоміжну речовину для злиття 0,05%(об'листків, широким хлорозом листя і, зрештою, загиєм/об'єм). Відбирали шість ліній, резистентних до беллю рослини. Для менделевського аналізу сегімазамоксу, які позначали як лінії 1А, 9А, 10А, 11A, регуючих популяцій рослини поділяли на резисте15А та 16А. Покоління М3 та М4 вирощували в кантні та чутливі категорії і випробували на мері для вирощування і відбирали рослини, резисадекватність різним 1-генним, 2-генним та 3тентні до імазамоксу при нормі 20г аі/га-1. Резистегенним моделям аналіз chi-square. Для даних по нтні рослини відбирали в поколінні M5 після рослинах F2 та BCF1 проміжні реакції було вклюнанесення 40г аі/га-1 в полі. Насіння М5 було гомочено до категорії резистентної реакції. Для регузиготним за характеристиками, оскільки випробулювання значення chi-square застосовували коревання потомства не виявляло сегрегації щодо рекцію за Yates для безперервності, коли в аналізі зистентності до імазамокс. chi-square користувалися лише одним ступенем Приклад 2 свободи [Steele and Torrie 1980 Principles and Способи, які застосовують для визначення усprocedures of statistics. McGraw-Hill, New York, New падкування та алелізму ІМІ генів York, pp. 633]. Для визначення генетичного контролю резисПриклад3 тентності до імазамоксу в шести лініях пшениці Результати, що стосуються успадкування ІМІ здійснювали взаємне схрещування між шістьма генів гомозиготними резистентними М6 лініями та CDC Усі резистентні батьківські рослини давали Teal (чутливим до імазамоксу). Випадково відібраоднаковий фенотип при розпилюванні 20г аі/га-1 ні рослини F1 від кожного схрещування піддавали зворотному схрещуванню з CDC Teal для утвоімазамоксу. (Фігура 1). Взаємне схрещування між рення зворотно схрещених (BC)F1 популяцій. Для резистентними лініями та чутливими батьківськидослідження алелізму здійснювали всі можливі ми рослинами (CDC Teal) у результаті давало ровзаємні схрещування між шістьма мутантами та слини F1, які витримували нанесення імазамоксу SWP965001 (Grandin/3*Fidel-FS-4). SWP965001 є (Фігура 1), що вказувало на те, що резистентність лінією ярової пшениці, яка є гомозиготною для FSдо імазамокс є ядерною, а не цитоплазматичною 4 алелі. Батьківські генотипи вирощували в камері характеристикою. За винятком гібриду 15А´Teal, для вирощування з 16-годинним фотоперіодом і рослини F1, одержані в результаті схрещування 24°С денним та 16°С нічним температурним рекожної з резистентних ліній з CDC Teal, виявляли жимом. Колоски, які з'являлися на ¾, піддавали проміжну реакцію (Фігура 1). Оскільки рослини F1 емаскуляції, а потім запилювали через 2-3 дні пісбули фенотипічно проміжними між двома батьківля емаскуляції. Випадково відібрані рослини F2 з ськими рослинами, було зроблено висновок, що усіх сегрегуючих гібридів змішували для одержанрезистентність до імазамоксу в цих лініях була ня груп F2:3. Батьківські рослини, F1, BCF1, F2 та частково домінантною характеристикою (Фігура 1). групи F2:3 випробували на реакцію на імазамокс. Генетичний аналіз резистентності до імідазоліноУсі експерименти здійснювали в камері для виронів та сульфонілсечовини в Arabidopsis thaliana щування з 16-годинним фотоперіодом і 23°С ден[Haughn and Somerville, 1986 Mol. Gen. Genet. ним та 16°С нічним температурним режимом. Усі 204:430-434] lea mays [Newhouse et al., 1991 Theor. експерименти здійснювали на повністю випадковій Appl. Genet. 83:65-70], Brassica napus [Swanson et основі. В експериментах із залученням груп F2:3 al., 1989 Theor. Appl. Gen. 78:525-530] та Glycine намагалися забезпечити рандомізацію як у межах max [Sebastian et al., 1989 Crop Sci. 29:1403-1408] групи, так і між групами. Популяції F1 та F2 відбитакож вказував на присутність єдиного, частково рали в такому самому експерименті разом з батьдомінантного ядерного гена. ківськими генотипами та CDC Teal як контрольниЧотирнадцять рослини F1, одержаних у реми зразками. Обидві популяції, BCF1 та F2:3, зультаті схрещування 15А´Teal, зараховували до відбирали у двох окремих експериментах разом з резистентних (Фігура 1). Оцінка популяцій F2 від відповідними батьківськими генотипами як контроцього схрещування показала, що два незалежно льними зразками. сегреговані локуси були пов'язані з забезпеченням Обробку гербіцидами застосовували до росрезистентності в цьому генотипі (Фігура 2). Оскільлин, які вирощували на платформах з комірками ки аі/га-1 має нести два гетерозиготні резистентні 39 87261 40 Nicotiana tabacum резистентність до інгібіторів локуси, можна очікувати, що спостерігатиметься AHAS успадковується як єдиний частково домінарезистентна реакція. Якщо кожен з цих локусів нтний ядерний ген [Newhouse et al. 1991; окремо забезпечує часткове домінування, то в сумі Newhouse et al. 1992; Chaleff and Ray, 1984 два гетерозиготні локуси мають забезпечувати Science 223:1148-1151; Sathasivan et al., 1991 Plant резистентну реакцію. Swanson et al. (1989) комбіPhysiol. 97:1044-1050]. Резистентність рослин до нували дві напівдомінантні алелі резистентності інгібуючих AHAS гербіцидів відбувається здебільдо імідазолінону з Brassica napus, які представляшого через одиночну мутацію в гені, що кодує феють два роз'єднані гени, для одержання гібриду F1, рмент AHAS [Harms et al. 1992, Mol. Gen. Genet. який переважав у резистентності до імідазолінону 233:427-435; Winder and Spalding, 1988 Mol. Gen. будь-яку з окремих гетерозиготних ліній. Автори Genet. 238:394-399]. дійшли висновку, що механізми резистентності є Дані для популяції F2, одержаної в результаті сукупними, і вищий рівень резистентності спостерігають у лініях, які несуть більше однієї алелі резисхрещування Teal´15Α, відповідали співвідношенстентності. ню 15:1 резистентності:чутливості, вказуючи на Аналіз цитоплазматичної спадковості здійснюсегрегацію двох незалежно сегрегованих локусів вали в поколінні F2 шляхом випробування рівномі(Фігура 2). Якщо це так, то групи F2:3 мають сегрерності відхилення від співвідношення сегрегації гуватися і відповідати співвідношенню групи F2:3 між двома взаємними популяціями F2. Аналіз chi7:8:1 резистентності:сегрегації:чутливості. Групи square не виявив значного відхилення між взаємF2:3 від схрещування 15А´Teal не відповідали очіними популяціями, підтвердивши відсутність цитокуваному співвідношенню 7:8:1 (Фігура 3), підтверплазматичної спадковості (Фігура 2). Оскільки циджуючи результати для популяцій F2 та BCF1, які топлазматична спадковість була відсутня, дані свідчили про те, що резистентність у 15А забезпедвох взаємних популяцій комбінували і розраховучується двома незалежними локусами. Наскільки вали загальний показник chi-square по об'єднаних відомо винахідникові, це перший повідомлений даних F2 (Фігура 2). випадок, коли дві незалежно сегреговані резистенЗа винятком Teal´15Α, усі популяції F2, одертні до імідазолінону алелі були ідентифіковані в жані в результаті схрещувань резистентних та одній лінії після мутагенезу насіння. чутливих рослин, відповідали співвідношенню 3:1 Приклад 4 резистентності до чутливості, вказуючи на сегреРезультати, що стосуються алелізму ІМІ генів гацію єдиного головного гена резистентності до Для визначення алельних взаємозв'язків генів імазамоксу (Фігура 2). Коли рослини F1 схрещуварезистентності досліджували всі можливі схрещули з чутливими батьківськими рослинами, одержавання між резистентними лініями. Не спостерігали ні в результаті популяції BCF1 відповідали співвідчутливих рослин у популяціях F2, одержаних у реношенню 1:1 резистентності:чутливості, зультаті взаємного схрещування між лініями підтверджуючи однолокусну гіпотезу (Фігура 2). SWP965001, 1А, 9А, 10А, 15Ата 16А (Фігура 4). Дані щодо популяції F2 від схрещування 15А´Teal Оскільки ці популяції не сегрегувалися, гени резистентності в цих лініях є або алелями в локусі FSвідповідають співвідношенню 15:1 резистентностг4, або є дуже щільно зв'язаними. Оскільки ці попучутливості (Р=0,08), вказуючи на сегрегацію двох ляції не сегрегувалися в поколінні F2, групи F2:3 від незалежних, комплементарних генів (Фігура 2). цих схрещувань не досліджували. Популяція BCF1 відповідала співвідношенню 3:1 Усі схрещування за участю лінії 11А сегрегурезистентності:чутливості зі значенням Ρ Chiвалися в поколінні F2, вказуючи на присутність square 0,35, підтверджуючи результати для F2 (Фіунікального гена резистентності у 11А (Фігура 4). У гура 2). разі присутності двох незалежно сегрегованих Оскільки на основі даних для F2 було висловгенів резистентності в результаті схрещування лено припущення, що резистентність у лініях 1А, двох ліній, кожна з яких несе один ген резистент9А, 10А, 11A та 16А контролюється одним голоності, слід очікувати співвідношення 15:1 резистевним геном, групи F2:3 мають сегрегуватися і віднтності: чутливості в поколінні F2. У поколінні F2, повідати співвідношенню 1:2:1 груп гомозиготної резистентності: сегрегації: гомозиготної чутливосгібриди SWP965001´11A, 1А´11A, 10А´11А та ті. Оцінка груп F2:3 показала, що гібриди Teal´1А, 16А´11Α відповідають очікуваному співвідношенTeal´9Α, Teal´10Α, Teal´11A та Teal´16Α усі відню 15:1 резистентності:чутливості, вказуючи на незалежну сегрегацію двох головних генів резисповідають співвідношенню групи F2:3 1:2:1 резистентності (Фігура 4). Співвідношення у групі F2:3 від тентності: сегрегації: чутливості зі значеннями Ρ цих схрещувань також відповідали співвідношенChi-squares 0,64, 0,66, 0,52, 0,40 та 0,94, відповідням 7:8:1 резистентності: сегрегації: чутливості, но (Фігура 3). Ці результати підтверджують репідтверджуючи результати, отримані для поколінзультати даних для F2 та BCF1, за якими резистентність у лініях 1А, 10А, 9А, 11А та резистентність у ня F2 (Фігура 5). Гібрид 11А´9Α давав сегреговану 16А контролюється одним головним геном. Ця популяцію F2, але співвідношення не відповідало модель спадковості узгоджується з іншими данипоказникові сегрегації 15:1 через надлишок чутлими, які свідчили про генетичний контроль резистевих сегрегантів. Випробували різні інші дводенні нтності до інгібуючих AHAS гербіцидів. На даний гіпотези, але всі виявилися високо значущими (дачас майже всі мутації рослин, які забезпечують ні не показано). Однак, оцінка груп F2:3 від цього резистентність до імідазолінонів, демонструють, схрещування відповідала показникові сегрегації що єдиний, частково домінантний ген контролює 7:8:1, вказуючи на сегрегацію двох незалежних характеристику резистентності. У Triticun aestivum, генів (Фігура 5). Ці результати підтверджують, що Zea mays, Glycine max, Arabidopsis thaliana та 41 87261 42 ген резистентності у 11А є відмінним від генів у AHAS, яка складається з п'яти та шести членів, лініях SWP96001, 1A, 9A, 10А та 16А. відповідно [Rutledge et al., 1991 Mol Gen. Genet. 229:31-40; Grula et al., 1995 Plant Mol. Biol. 28:837Гібрид 11Α´15Α не давав сегрегованої попу846]. Вищий рівень резистентності до гербіцидів ляції F2. Оскільки 15А несе два гени резистентносспостерігали в поліплоїдних видах у разі присутті, один алель ний до FS-4, сегрегована популяція ності багатьох алелей резистентності. Swanson et F2 у гібриді 11А´15A має вказувати на присутність al. [1989 Theor. Appl. Gen. 78:525-530] комбінували трьох сегрегованих генів. Сегреговані покоління, дві унікальні алелі резистентності до імідазолінону одержані в результаті схрещування 15А´11A, було з двох гомозиготних ліній Brassica napus, в резульвипробувано на сегрегацію трьох незалежних лотаті чого одержували потомство з вищим рівнем кусів. Рослини F2 відповідали очікуваному співвідрезистентності, ніж у кожної з цих гомозиготних ношенню 63:1 резистентності:чутливості, вказуючи ліній окремо. Creason and Chaleff [1988 Theor. на сегрегацію трьох незалежних локусів (Фігура 4). Appl. Genet. 76:177-182] ідентифікували рослини Ці результати вказують на те, що друга мутація у Nicotiana tabacum, гомозиготні для двох мутацій, 15А не є алельною генові резистентності у 11A. які забезпечували резистентність до сульфонілсеГрупи F2:3 не піддавали відборові, оскільки довечовини. Рослини, гомозиготні для обох мутацій, лося б відбирати понад 330 рослин у кожній групі, були в п'ять разів більш резистентними до нанещоб забезпечити достатню потужність критерію сення на листя хлорсульфурону, ніж рослини, го[Hanson, 1959 Agron. J. 51:711-716]. мозиготні для кожної окремої мутації. Даний винаБуло ідентифіковано три незалежні локуси рехід пропонує вироблення вищого рівня зистентності, кожен з яких мав алель, яка забезперезистентності до імідазолінонового гербіциду у чує резистентність до імазамоксу. Опубліковано пшениці шляхом комбінування будь-яких двох або рекомендовані правила для генного локусу та сивсіх трьох алелей резистентності. мволізації алелей [Mclntosh et al., 1998 Catalogue Приклад 5 of Gene Symbols. Volume 5, Proceedings of the 9th Толерантність до імідазолінонових гербіцидів International Wheat Genetics Symposium. у ТеаІ11А, ТеаІ15А та гібриді Teal11A/15А Saskatoon, Saskatchewan]. Неалельні генні локуси Підвищена толерантність, яку демонструють ферменту, які каналізують одну реакцію, мають Teal11А та Teal15A до 20 грамів на гектар імазапозначатися однаковим символом, який відповідає моксу, підтверджувалася в наведених вище притрадиційній назві ферменту. Традиційною назвою кладах можливістю відрізнення толерантних від для AHAS є ALS. За відсутності даних для призначутливих батьківських та сегрегованих рослин у чення локусів для конкретних хромосом та геномів дослідженні успадкування. Було виявлено, що вони позначаються послідовною серією. ПознаTeal11А забезпечує такий самий рівень толерантчення фенотипу, який спостерігають у разі змін у ності до імідазолінонових гербіцидів, як і той, що гені, в результаті чого виникає нова алель, має забезпечується мутацією FS4 у Fidel у різних оравідображати цей фенотип. Таким чином, пропонунжерейних та польових порівняннях. Подібність ється, щоб алель резистентності до імідазолінону толерантності також відбивається в порівнянні in FS-4 позначалася як lmi1, а її локус - позначався vitro активності AHAS, видобутої з толерантних як Als1. Imi означає резистентність до імідазолінорослин. Це є можливим, оскільки толерантність у ну. Це позначення вказує, що ген є домінантною Teal11A, Teal15A та FS4 зумовлюється мутаціями характеристикою, і він є першою ідентифікованою у ферменті AHAS, що забезпечує резистентність алеллю. Дані для сегрегованих популяцій F2 та F2:3 до інгібування імідазоліноновими гербіцидами. вказують, що 15А та 11А несуть дві нові незалежні Фігура 6 вказує, що активність ферменту AHAS, алелі резистентності в різних локусах (Фігури 2 та видобутого з Teal11А та BW755, лінії, що містить 3). Позначеннями для цих алелей є Іmі2 для мутаFS4, змінює подібність зі збільшенням норми імації 11А в локусі Als2 і lmi3 для другої мутації 15А в замоксу, й обидва мають вищий відсоток активнолокусі Als3. го (резистентного) ферменту при найвищій концеАвторами було ідентифіковано три незалежно нтрації імазамоксу, ніж у контрольної рослини Teal сегреговані алелі, які забезпечують резистентність дикого типу. до імазамоксу, а саме, lmi1 (1А, 9А, 10А, 15А та Присутність двох ІМІ нуклеїнових кислот у 16А), Іmі2 (11A) та lmi3 (15А). Пропонується, щоб Teal15A забезпечує підвищену толерантність до кожна з трьох ідентифікованих алелей пов'язуваімідазолінонових гербіцидів порівняно з такою лілася зі своїм структурним геном, який кодує нечунією, як BW755, що має лише одну ІМІ нуклеїнову тливі до гербіциду форми AHAS. Оскільки пшеникмслоту. Ця підвищена толерантність відбиваєтьця є гексаплоїдною, слід очікувати багато локусів ся й у меншому ураженні при більш високих норAHAS. Було виявлено, що інші поліплоїдні види мах гербіциду, й у більшій неінгібованій активності мають більше однієї копії AHAS. У Nicotiana AHAS ферменту. Фігура 7 пояснює, що при 10´ tabacum, алотетраплоїді, було ідентифіковано і нормі імазамоксу (200г/га) усі оброблені одногенні охарактеризовано два гени AHAS [Mazur et at. рослини зазнавали ураження, тоді як жодна з дво1987]. Chaleff and Ray (1984) ідентифікували два генних рослин не була ураженою. При всіх конценнезалежно сегреговані алелі резистентності до траціях імазамоксу в in vitro аналізі активності сульфонілсечовини в Nicotiana tabacum, кожна з AHAS (Фігура 6), але особливо - при найвищих яких кодує змінену форму AHAS. Zea mays має концентраціях, рослина ТеаІ15А мала вищий віддва конститутивно експресовані ідентичні гени соток активного (резистентного) ферменту, ніж у AHAS [Fang et al., 1992 Plant Mol. Biol. 18:1185будь-якої з моногенних ліній, Теаl11А та BW755. 1187]. В алотетраплоїді Brassica napus та Gossypium hirsutum присутня багатогенна група 43 87261 44 Комбінування трьох неалельних генів, кожен з більше половини рослин не витримали уражень у яких забезпечує толерантність до імідазоліноноще сегрегованій змішаній популяції ТеаІ15А, вих гербіцидів, у результаті дає більшу толерантсхрещеній з Теаl11А, тоді як усі рослини гомозигоність, ніж у разі лише двох неалельних генів (Фігутної популяції Teal15А витримали ураження. ра 7). При нормі 30Х або 600г/га імазамоксу 45 87261 46 47 87261 48 49 87261 50 51 87261 52 53 87261 54 55 87261 56 57 87261 58 59 87261 60
ДивитисяДодаткова інформація
Назва патенту англійськоюWheat plants having increased resistance to imidazolinone herbicides
Автори англійськоюPOZNIAK CURTIS J, HUCL PIERRE
Назва патенту російськоюРастение пшеницы с повышенной резистентентностью к имидазолиновым гербицидам
Автори російськоюПозняк Кертис Дж., Хакл Пьер
МПК / Мітки
МПК: C12N 15/82, C12N 15/60, A01H 5/00, C12N 9/88, C12Q 1/68
Мітки: імідазолінонових, резистентністю, підвищеною, пшениці, рослина, гербіцидів
Код посилання
<a href="https://ua.patents.su/41-87261-roslina-pshenici-z-pidvishhenoyu-rezistentnistyu-do-imidazolinonovikh-gerbicidiv.html" target="_blank" rel="follow" title="База патентів України">Рослина пшениці з підвищеною резистентністю до імідазолінонових гербіцидів</a>
Спосіб визначення імідазолінонових гербіцидів у воді та грунті

Номер патенту: 24827
Опубліковано: 10.07.2007
Автори: Бардов Василь Гаврилович, Гіренко Дельбар Бекджанівна, Горбачевський Руслан Вікторович, Омельчук Сергій Тихонович, Коршун Марія Михайлівна
МПК: G01N 30/00
Мітки: грунті, гербіцидів, імідазолінонових, води, спосіб, визначення
Формула / Реферат:
Спосіб визначення імідазолінонових гербіцидів у воді та грунті, що включає підготовку проб води та ґрунту до екстракції, проведення екстракції, очищення екстрактів, реекстракції, концентрування та хроматографування, який відрізняється тим, що змінюють умови хроматографування для чіткого розходження піків речовин (імазапіру, імазетапіру, імазамоксу), що знаходяться в суміші проби, після отримання хроматографій вивчають піки кожної речовини,...
Спосіб передпосівної обробки насіння озимої пшениці
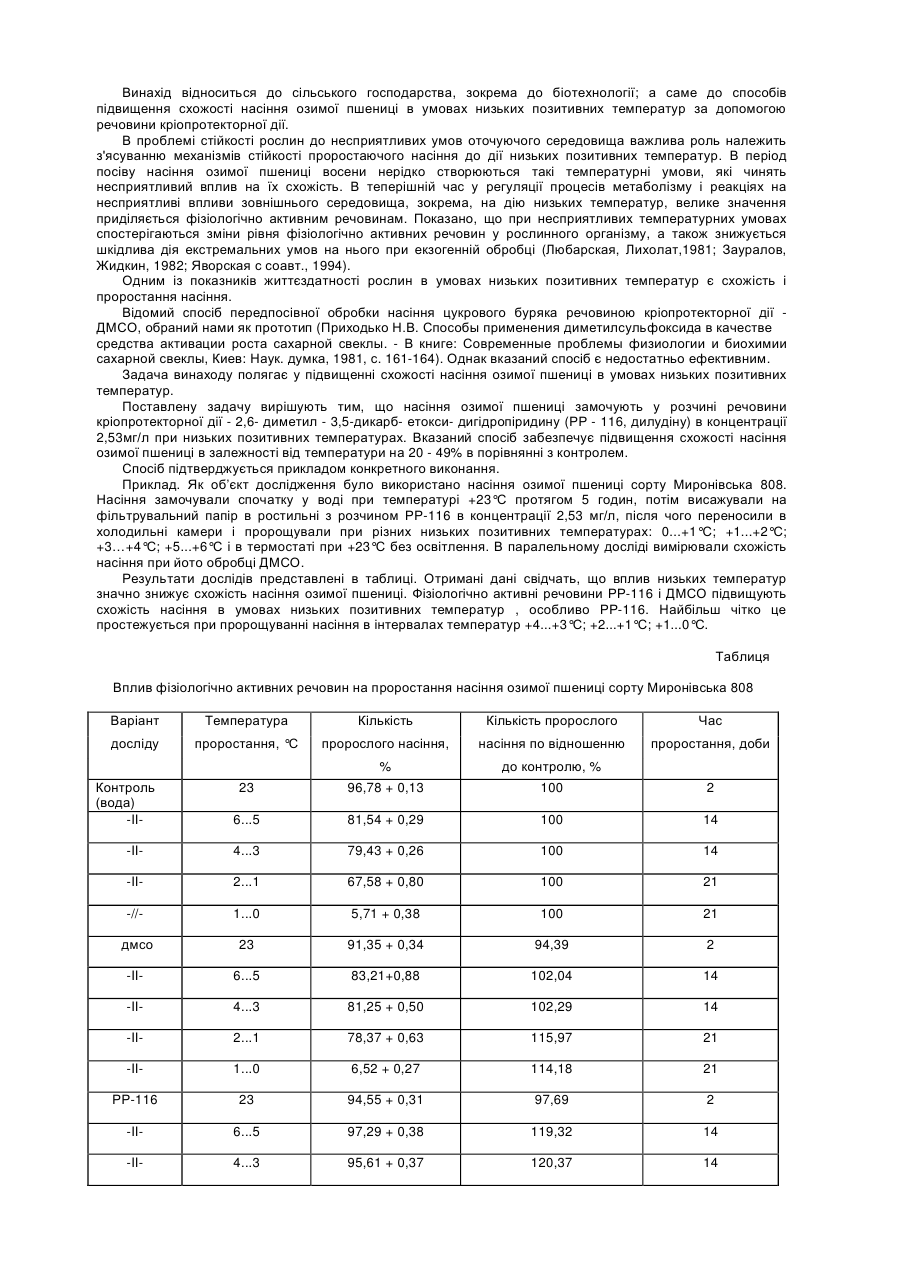
Номер патенту: 46260
Опубліковано: 15.05.2002
Автор: Хілько Тетяна Дмитрівна
МПК: A01C 1/00
Мітки: спосіб, озимої, пшениці, насіння, передпосівної, обробки
Формула / Реферат:
Спосіб передпосівної обробки насіння озимої пшениці, який включає замочування насіння озимої пшениці в розчині речовини кріопротекторної дії при низьких позитивних температурах, який відрізняється тим, що як речовину кріопротекторної дії використовують 2,6-диметил-3,5-дикарб-етокси-дигідропіридин в концентрації 2,53 мг/л.
Спосіб підвищення врожайності пшениці ярої
Номер патенту: 65661
Опубліковано: 15.04.2004
Автори: Шевченко Ярослав Олексійович, Шатурський Ярослав Павлович, Лісовал Анатолій Петрович
МПК: A01N 47/40
Мітки: ярої, спосіб, підвищення, врожайності, пшениці
Формула / Реферат:
Спосіб підвищення врожайності пшениці ярої шляхом передпосівної обробки насіння або вегетативної маси пшениці ярої водним розчином активної речовини, який відрізняється тим, що як активну речовину використовують гідройодид S-(2-метокси-2-феніл-етил)-ізотіосечовини формули:а її водний розчин готують у концентрації 0,05 мас.%.
Спосіб вирощування ярої пшениці на зрошуваних землях

Номер патенту: 36024
Опубліковано: 10.10.2008
Автор: Заєць Сергій Олександрович
МПК: A01B 79/02
Мітки: спосіб, вирощування, землях, ярої, зрошуваних, пшениці
Формула / Реферат:
Спосіб вирощування ярої пшениці на зрошуваних землях, що включає основний обробіток ґрунту, внесення добрив, поливи, посів, догляд за посівами та збирання врожаю, який відрізняється тим, що проводять плоскорізальний обробіток ґрунту на 18 см, під передпосівну культивацію вносять 60 кг/га діючої речовини карбаміду, насіння обробляють імуномодулятором Табод (100 г/т), а рослини обприскують у фазу кущіння і в налив зерна...
Спосіб підвищення урожаю і якості зерна озимої пшениці

Номер патенту: 29090
Опубліковано: 16.10.2000
Автори: Ткачук Катерина Семенівна, Богдан Тетяна Зіновіївна, Кузьменко Лідія Михайлівна
МПК: A01G 7/00
Мітки: якості, пшениці, урожаю, підвищення, озимої, спосіб, зерна
Формула / Реферат:
Спосіб підвищення урожаю і якості зерна озимої пшениці, що передбачає внесення під зяблеву оранку N60P60K60, який відрізняється тим, що додатково вносять сірчанокислий літій в дозі 2 кг/га.
Попередній патент: Підшипникова опора бурового долота
Наступний патент: Спосіб лікування первинного безсоння