Спосіб оцінки стійкості видів і гібридів рослин роду гіркокаштан (aesculus l.) до каштанової мінуючої молі (cameraria ohridella deschka et dimic)
Номер патенту: 59330
Опубліковано: 10.05.2011
Автори: Демчук Тетяна Леонідівна, Мельничук Максим Дмитрович, Дубровін Валерій Олександрович, Машковська Світлана Петрівна, Григорюк Іван Панасович, Серга Олеся Іванівна





Формула / Реферат
Спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et dimic), що ґрунтується на кількісному визначенні вмісту АМФ, АДФ й АТФ в листках рослин, який відрізняється тим, що як інтегральний біоенергетичний показник енергозабезпечення використовують величину енергетичного заряду аденозинфосфатної системи [ЕЗ], при цьому відбір свіжих листків нижнього, середнього і верхнього ярусів видів й гібридів рослин роду Гіркокаштан проводять до початку появи каштанової мінуючої молі (фаза початку цвітіння рослин), далі здійснюють екстракцію і очищення вільних аденозинфосфатів (АМФ, АДФ, АТФ) й визначають їх вміст у відібраних листках рослин, розраховують величину енергетичного заряду аденозинфосфатної системи за формулою:
ЕЗ = (АТФ+1/2АДФ)/(АМФ+АДФ+АТФ),
де АМФ - це величина вмісту у листках рослин аденозин-5-монофосфатів, АДФ - аденозин-5-дифосфатів, а АТФ - аденозин-5-трифосфатів; а вже за величиною енергетичного заряду аденозинфосфатної системи ЕЗ оцінюють стійкість видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі таким чином, що коли величина ЕЗ становить більше, ніж 0,5, то види й гібриди рослин роду Гіркокаштан належать до стійких до каштанової мінуючої молі, а якщо менше 0,5 - до нестійких.
Текст
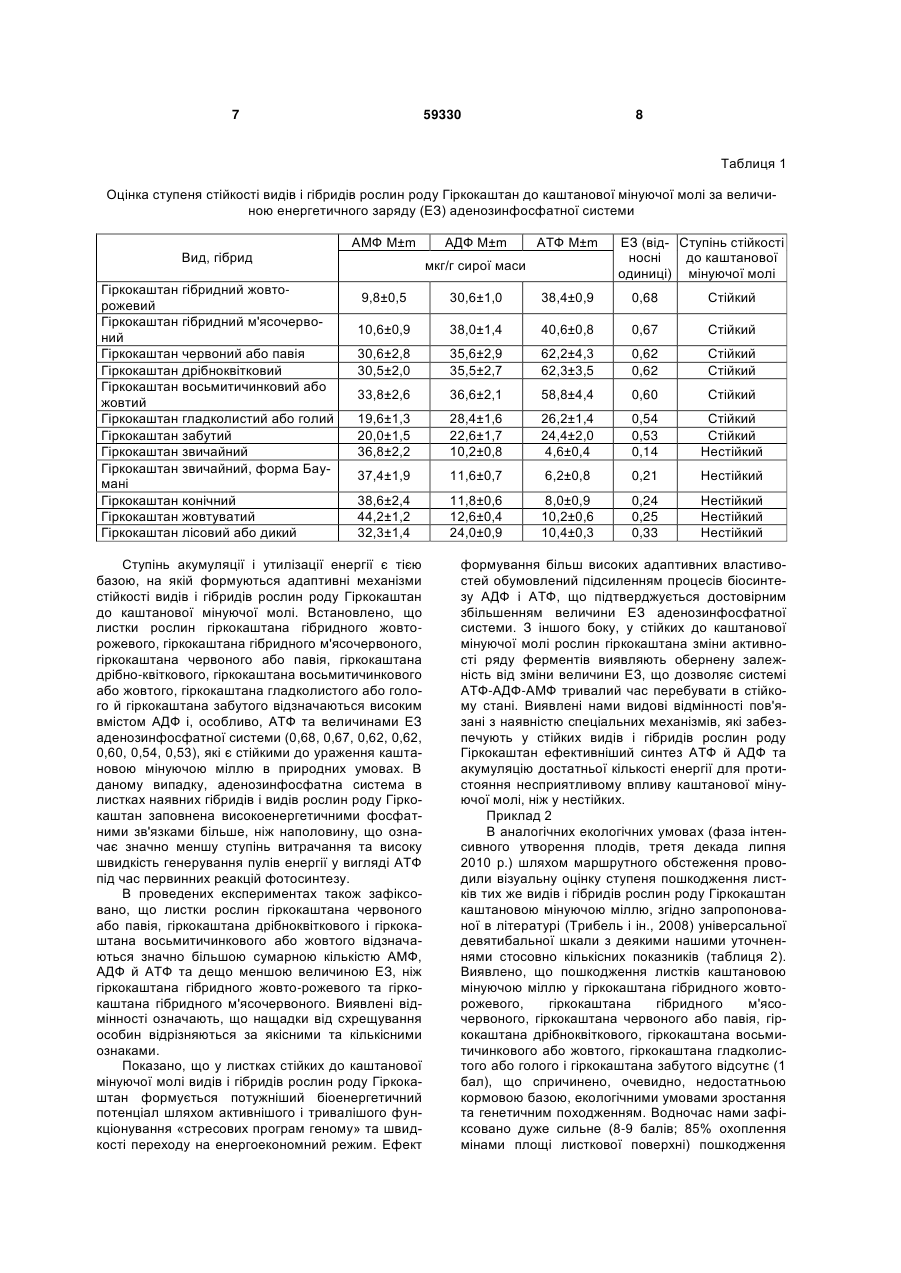
Спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et dimic), що ґрунтується на кількісному визначенні вмісту АМФ, АДФ й АТФ в листках рослин, який відрізняється тим, що як інтегральний біоенергетичний показник енергозабезпечення використовують величину енергетичного заряду аденозинфосфат U 2 (13) 1 3 6,9 т кисню [7]. Однак, деякі види рослин роду Гіркокаштан поступово вичерпують свій адаптивний потенціал і нерідко гинуть від промислового забруднення полютантами, посухи, високих температур, хвороб та шкідників. Нині, один із найагресивніших шкідників, що значною мірою уражує листки рослин гіркокаштана звичайного в Європі і Україні, є каштанова мінуюча міль (Сатегагіа ohridella Deschka et Dimic) - вид метеликів, який відноситься до ряду лускокрилих (Lepidoptera) та родини молей-строкаток {Glacillariidae) [4, 8]. Це - типовий ксерофіл, найнебезпечніший, адвентивний і особливо поширений інвазійний чужорідний вид, для якого характерна наявність достатньої кормової бази, відсутність природних ворогів й висока швидкість розселення ареалу, що спричиняє інфекційне усихання листків та передчасну загибель рослин гіркокаштана в природних умовах [4, 8]. За нашими оцінками, деякі види гіркокаштана, які зростають в Україні, в літній період не витримують пошкоджувального впливу каштанової мінуючої молі, в результаті відбувається розхитування спадковості, що пов'язано із значним витрачанням запасних енергетичних ресурсів та нестабільністю геному в умовах розбалансованих глобальних змін клімату на Земній кулі. Фізіолого-біохімічні і генетичні зміни, які виникають в результаті масового пошкодження листків рослин гіркокаштана різного походження каштановою мінуючою міллю, залишаються до теперішнього часу не з'ясованими. Такий стан проблеми стримує проведення селекційногенетичних експериментів та застосування різноманіття генофонду рослин роду Гіркокаштан для озеленення міських територій України. Відомий спосіб [1] визначення посухо-стійкості деревних рослин шляхом вимірювання максимальних значень швидкості потоку пасоки і водного потенціалу (ВП) в листках за допомогою датчиків впродовж 3-4 діб. Проте, даний спосіб недосконалий і недостатньо інформативний, який потребує тривалого часу, при цьому відбуваються значні втрати листками вільної води за рахунок збільшення інтенсивності транспірації, що індуковано перегрівом рослин. Найбільш близьким до пропонованого нами є спосіб оцінки посухостійкості сортів озимої м’якої пшениці [3], що ґрунтується на кількісному визначенні величини енергетичного заряду аденозинфосфатної системи [ЕЗ] тільки в прапорцевих листках рослин за умов недостатнього та оптимального водозабезпечення. Суттєвим недоліком цього способу є лише одностороння і часткова оцінка сумарного енергозабезпечення та відносної посухостійкості рослин у штучно створених вегетаційних умовах. Водночас, не оцінено вклад в енергетичний баланс листків рослин пшениці нижнього та середнього ярусів як головних постачальників асимілятів для фотосинтетичного апарату. В основу корисної моделі покладене завдання розробки ефективного й інформативного способу оцінки стійкості видів і гібридів роду Гіркокаштан до каштанової мінуючої молі шляхом застосування оригінального методу визначення вмісту аденозин-5-моно-(АМФ), ди-(АДФ) і трифосфатів (АТФ) у 59330 4 листках рослин та наступної оцінки стійкості рослин за величиною енергетичного заряду аденозинфосфатної системи (ЕЗ) у комплексній взаємодії суттєвих ознак нового способу. Завдання вирішується завдяки тому, що в способі оцінки стійкості видів і гібридів рослин роду гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et dimic), що ґрунтується на кількісному визначенні вмісту АМФ, АДФ й АТФ в листках рослин, в якості інтегрального біоенергетичного показника енергозабезпечення використовують величину енергетичного заряду аденозинфосфатної системи [ЕЗ], при цьому відбір свіжих листків нижнього, середнього і верхнього ярусів видів й гібридів рослин роду Гіркокаштан проводять до початку появи каштанової мінуючої молі (фаза початку цвітіння рослин), далі здійснюють екстракцію і очищення вільних аденозинфосфатів (АМФ, АДФ, АТФ) й визначають їх вміст у відібраних листках рослин, розраховують величину енергетичного заряду аденозинфосфатної системи за формулою: ЕЗ=(АТФ+1/2АДФ)/(АМФ+АДФ+АТФ), де АМФ - це величина вмісту у листках рослин аденозин-5-монофосфатів, АДФ - аденозин-5дифосфатів, а АТФ - аденозин-5-трифосфатів; а вже за величиною енергетичного заряду аденозинфосфатної системи ЕЗ оцінюють стійкість видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі таким чином, що коли величина ЕЗ становить більше, ніж 0,5, то види й гібриди рослин роду Гіркокаштан відносяться до стійких до каштанової мінуючої молі, а якщо менше 0,5 - до нестійких. В основу корисної моделі вперше покладено ефект визначення сумарного ступеня акумуляції багатих енергією сполук аденілату в листках нижнього, середнього і верхнього ярусів, який оцінюють за величиною ЕЗ аденозинфосфатної системи. Згідно літературних даних [10, 12], ЕЗ - це «міра» заповнення аденозинфосфатної системи (суми АТФ, АДФ, АМФ) високоенергетичними фосфатними групами, який свідчить про ступінь нагромадження, утилізації та відновлення енергії в рослинному організмі [5]. Якщо аденозинфосфати в клітині містяться у формі АТФ, то система енергетично заповнена до межі і її ЕЗ дорівнює 1,0. Якщо вони знаходяться у вигляді АМФ, то система не містить високоенергетичних зв'язків і її ЕЗ становить 0,0. У випадку, коли АМФ, АДФ і АТФ виявляються у формі АДФ або в суміші АТФ й АМФ, то система заповнена високоенергетичними зв'язками лише наполовину і її ЕЗ дорівнює 0,5 [5]. Об'єктами досліджень слугували 12 видів і гібридів рослин роду Гіркокаштан, а саме: гіркокаштан гібридний жовто-рожевий (Aesculus hibrida D.C.), гіркокаштан гібридний м'ясочервоний (Aesculus carnea Hayne.), гіркокаштан червоний або павія (Aesculus pavia L), гіркокаштан дрібноквітковий (Aesculus purviflora Walt.), гіркокаштан восьмитичинковий або жовтий (Aesculus octandra March.), гіркокаштан гладколистий або голий (Aesculus glabra Willd.), гіркокаштан забутий (Aesculus neglecta Lindl), гіркокаштан звичайний (Aesculus hippocastanum L), гіркокаштан звичай 5 ний, форма Баумані {Aesculus hippocastanum, f.baumanii C.K. Schneid), гіркокаштан конічний (Aesculus turbinata Blume.), гіркокаштан жовтуватий (Aesculus flava Ait) і гіркокаштан лісовий або дикий (Aesculus silvatica Bartz.), які інтрудуковані в Національному ботанічному саду імені М.М. Гришка HAH України та частково зростають в Голосіївському лісі м. Києва. Запропонований нами спосіб реалізується наступним чином. До початку появи каштанової мінуючої молі (фаза початок цвітіння рослин) відбирають по 3 г матеріалу із свіжих листків нижнього, середнього і верхнього ярусів видів й гібридів рослин роду Гіркокаштан, фіксують рідким азотом в чотирьохкратній повторності та розтирають в фосфоровій ступці. Екстракцію і очищення вільних аденозинфосфатів (АМФ, АДФ, АТФ) здійснюють за методикою [6], а пряме визначення вмісту в листках рослин за допомогою тонкошарової хроматографії, що дозволяє їх чітко розділяти безпосередньо в шарі сорбенту. Метод ґрунтується на екстракції аденозинфосфатів з листків рослин і подальшому їх очищенні шляхом багаторазової хроматографії в некоригуючих системах розчинників. Ідентифікацію і кількісне детектування аденозинфосфатів проводять із тонкого шару сорбенту із застосуванням сканувального спектроденситометра «Camag TLC Scanner II» (Швейцарія), що до мінімуму зменшує їх втрати та розпад під час аналізу. ЕЗ аденозинфосфатної системи в листках видів і гібридів рослин роду Гіркокаштан розраховують за формулою (АТФ+1/2АДФ)/(АМФ+АДФ+АТФ) [5, 11], величина якого є переважно стабільною величиною, що є передумовою метаболічного гомеостазу [13]. Доведено, що ЕЗ може бути індикатором стресового стану організму, визначати можливості синтезу рослинами нових сполук, контролювати значну кількість елементів азотного і вуглецевого метаболізму, наприклад через фітохромну систему [12, 14, 15]. Нами висловлено припущення щодо наявності тісного взаємозв'язку між рівнем енергозабезпечення і стійкістю рослин гіркокаштана до каштанової мінуючої молі. З'ясовано, що гусениці першого, другого і третього віку проникають під кутикулу в шар клітин верхнього епідермісу листка й живляться лише соком, в якому містяться запасні енергетичні сполуки, переважно вуглеводи, четвертого та п'ятого - тільки тканинами палісадної паренхіми листкової пластинки рослин гіркокаштана [4, 8]. За таких стресових умов відбувається гальмування процесів фотосинтезу, енергозабезпечення, біосинтезу хлорофілу, фітогормонів, ліпідів, надходження елементів мінерального живлення з підземної частини в надземну і підвищення 59330 6 інтенсивності транспірації, що прискорює старіння та зниження продуктивності рослин гіркокаштана [2]. Відмінними від прототипу ознак у корисній моделі, що з'являється, є: - підвищення ефективності, точності і надійності методу визначення ступеня оцінки стійкості видів та гібридів рослин роду Гіркокаштан до каштанової мінуючої молі за величиною ЕЗ аденозинфосфатної системи; - забезпечення зменшення втрат і розпаду АМФ, АДФ та АТФ під час аналізу; можливість отримувати достовірніші і точніші енергетичні характеристики з метою ранжування видів та гібридів рослин роду Гіркокаштан за ступенем стійкості до каштанової мінуючої молі. Приклад 1 Проводили порівняльну оцінку вмісту АМФ, АДФ і АТФ й величин ЕЗ аденозинфосфатної системи в листках видів та гібридів рослин роду Гіркокаштан з метою визначення їх стійкості до каштанової мінуючої молі (таблиця 1). В листках видів гіркокаштана звичайного, гіркокаштана звичайного, форма Баумані, гіркокаштана конічного, гіркокаштана жовтуватого і гіркокаштана лісового або дикого нами визначено найбільш високі концентрації АМФ й найменш низькі - АТФ та величини ЕЗ аденозинфосфатної системи, що засвідчує високий рівень витрачання запасних енергетичних ресурсів на процеси адаптації рослин до каштанової мінуючої молі. Зменшення значень величини ЕЗ аденозинфосфатної системи в листках рослин активує біосинтез аденозину із АМФ, антистресова дія якого спричинена 2+ блокадою входження іонів Са до клітин. Нами показано, що у листках рослин нестійких до каштанової мінуючої молі відбувається розбалансування енергетичних процесів і пригнічення генерування сумарного пулу енергії у вигляді АТФ, при гідролізі фосфатного зв'язку якої вивільнюється вільна енергія, яка витрачається на функціонування адаптивних процесів. Із наведених даних видно, що аденозинфосфатна система в листках гіркокаштана звичайного, гіркокаштана звичайного, форма Баумані, гіркокаштана конічного, гіркокаштана жовтуватого і гіркокаштана лісового або дикого заповнена високоенергетичними фосфатними зв'язками менше половини і її ЕЗ дорівнює 0,14, 0,21, 024, 0,25 та 0,33 відповідно, що дає підставу віднести їх до нестійких рослин до каштанової мінуючої молі (таблиця 1). Головним чинником, який спричиняє низьку стійкість даних видів гіркокаштана до каштанової мінуючої молі є швидкий розпад АТФ до АДФ й АМФ, що призводить до зменшення величини ЕЗ в листках рослин. 7 59330 8 Таблиця 1 Оцінка ступеня стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі за величиною енергетичного заряду (EЗ) аденозинфосфатної системи АМФ М±m Вид, гібрид Гіркокаштан гібридний жовторожевий Гіркокаштан гібридний м'ясочервоний Гіркокаштан червоний або павія Гіркокаштан дрібноквітковий Гіркокаштан восьмитичинковий або жовтий Гіркокаштан гладколистий або голий Гіркокаштан забутий Гіркокаштан звичайний Гіркокаштан звичайний, форма Баумані Гіркокаштан конічний Гіркокаштан жовтуватий Гіркокаштан лісовий або дикий АДФ М±m АТФ М±m мкг/г сирої маси ЕЗ (від- Ступінь стійкості носні до каштанової одиниці) мінуючої молі 9,8±0,5 30,6±1,0 38,4±0,9 0,68 Стійкий 10,6±0,9 38,0±1,4 40,6±0,8 0,67 Стійкий 30,6±2,8 30,5±2,0 35,6±2,9 35,5±2,7 62,2±4,3 62,3±3,5 0,62 0,62 Стійкий Стійкий 33,8±2,6 36,6±2,1 58,8±4,4 0,60 Стійкий 19,6±1,3 20,0±1,5 36,8±2,2 28,4±1,6 22,6±1,7 10,2±0,8 26,2±1,4 24,4±2,0 4,6±0,4 0,54 0,53 0,14 Стійкий Стійкий Нестійкий 37,4±1,9 11,6±0,7 6,2±0,8 0,21 Нестійкий 38,6±2,4 44,2±1,2 32,3±1,4 11,8±0,6 12,6±0,4 24,0±0,9 8,0±0,9 10,2±0,6 10,4±0,3 0,24 0,25 0,33 Нестійкий Нестійкий Нестійкий Ступінь акумуляції і утилізації енергії є тією базою, на якій формуються адаптивні механізми стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі. Встановлено, що листки рослин гіркокаштана гібридного жовторожевого, гіркокаштана гібридного м'ясочервоного, гіркокаштана червоного або павія, гіркокаштана дрібно-квіткового, гіркокаштана восьмитичинкового або жовтого, гіркокаштана гладколистого або голого й гіркокаштана забутого відзначаються високим вмістом АДФ і, особливо, АТФ та величинами ЕЗ аденозинфосфатної системи (0,68, 0,67, 0,62, 0,62, 0,60, 0,54, 0,53), які є стійкими до ураження каштановою мінуючою міллю в природних умовах. В даному випадку, аденозинфосфатна система в листках наявних гібридів і видів рослин роду Гіркокаштан заповнена високоенергетичними фосфатними зв'язками більше, ніж наполовину, що означає значно меншу ступінь витрачання та високу швидкість генерування пулів енергії у вигляді АТФ під час первинних реакцій фотосинтезу. В проведених експериментах також зафіксовано, що листки рослин гіркокаштана червоного або павія, гіркокаштана дрібноквіткового і гіркокаштана восьмитичинкового або жовтого відзначаються значно більшою сумарною кількістю АМФ, АДФ й АТФ та дещо меншою величиною ЕЗ, ніж гіркокаштана гібридного жовто-рожевого та гіркокаштана гібридного м'ясочервоного. Виявлені відмінності означають, що нащадки від схрещування особин відрізняються за якісними та кількісними ознаками. Показано, що у листках стійких до каштанової мінуючої молі видів і гібридів рослин роду Гіркокаштан формується потужніший біоенергетичний потенціал шляхом активнішого і тривалішого функціонування «стресових програм геному» та швидкості переходу на енергоекономний режим. Ефект формування більш високих адаптивних властивостей обумовлений підсиленням процесів біосинтезу АДФ і АТФ, що підтверджується достовірним збільшенням величини ЕЗ аденозинфосфатної системи. З іншого боку, у стійких до каштанової мінуючої молі рослин гіркокаштана зміни активності ряду ферментів виявляють обернену залежність від зміни величини ЕЗ, що дозволяє системі АТФ-АДФ-АМФ тривалий час перебувати в стійкому стані. Виявлені нами видові відмінності пов'язані з наявністю спеціальних механізмів, які забезпечують у стійких видів і гібридів рослин роду Гіркокаштан ефективніший синтез АТФ й АДФ та акумуляцію достатньої кількості енергії для протистояння несприятливому впливу каштанової мінуючої молі, ніж у нестійких. Приклад 2 В аналогічних екологічних умовах (фаза інтенсивного утворення плодів, третя декада липня 2010 р.) шляхом маршрутного обстеження проводили візуальну оцінку ступеня пошкодження листків тих же видів і гібридів рослин роду Гіркокаштан каштановою мінуючою міллю, згідно запропонованої в літературі (Трибель і ін., 2008) універсальної девятибальної шкали з деякими нашими уточненнями стосовно кількісних показників (таблиця 2). Виявлено, що пошкодження листків каштановою мінуючою міллю у гіркокаштана гібридного жовторожевого, гіркокаштана гібридного м'ясочервоного, гіркокаштана червоного або павія, гіркокаштана дрібноквіткового, гіркокаштана восьмитичинкового або жовтого, гіркокаштана гладколистого або голого і гіркокаштана забутого відсутнє (1 бал), що спричинено, очевидно, недостатньою кормовою базою, екологічними умовами зростання та генетичним походженням. Водночас нами зафіксовано дуже сильне (8-9 балів; 85% охоплення мінами площі листкової поверхні) пошкодження 9 59330 каштановою мінуючою міллю листків гіркокаштана звичайного, форма Баумані й сильне (6-7 балів, 70% охоплення мінами площі листкової поверхні) у гіркокаштана конічного, гіркокаштана жовтуватого та гіркокаштана лісового або дикого (таблиця 2). Отримані результати тісно корелюють із змінами величин інтегрального біоенергетичного показника аденозинфосфатної системи ЕЗ, який у листках стійких до каштанової мінуючої молі видів і гібридів рослин роду Гіркокаштан коливався в межах від 0,53 до 0,68, а у нестійких - від 0,14 до 0,33. Таким чином, види і гібриди роду Гіркокаштан акумулюють різну кількість енергії у вигляді АТФ, 10 яка інтенсивніше витрачається в процесах метаболізму нестійкими до каштанової мінуючої молі рослинами, ніж стійкими. Якщо величина ЕЗ аденозинфосфатної системи становить більше, ніж 0,50, то види і гібриди рослин роду Гіркокаштан відносяться до стійких, якщо менше 0,50 - до нестійких. Запропонований нами спосіб дозволяє здійснювати достовірну оцінку ступеня стійкості рослин видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі й може бути використаний в селекції та для розробки заходів з озеленення міських територій. Таблиця 2 Оцінка ступеня пошкодження листків видів і гібридів рослин роду Гіркокаштан каштановою мінуючою міллю № п/п 1 2 3 4 5 6 7 8 9 10 11 12 Вид, гібрид Гіркокаштан гібридний жовто-рожевий Гіркокаштан гібридний м'ясочервоний Гіркокаштан червоний або павія Гіркокаштан дрібноквітковий Гіркокаштан восьмитичинковий або жовтий Гіркокаштан гладколистий або голий Гіркокаштан забутий Гіркокаштан звичайний Гіркокаштан звичайний, форма Баумані Гіркокаштан конічний Гіркокаштан жовтуватий Гіркокаштан лісовий або дикий Джерела інформації: 4 1. А.с. №1496704 ССР, МКИ А01С 7/00, А01Н 1/04. Способ определения засухоустойчивости древесных растений /Н. Е. Надеждина, Л. К. Кайбияйнен. - Опубл. 30.07.1989, Бюл. №28. 2. Григорюк І.П., Машковська С.П., Яворовський П.П., Колесніченко О.В. Біологія каштанів. - К.: Логос, 2004. - 380 с. 3. Декл. патент на винахід №45880 А Україна, МКВ7 A01G/00. Спосіб оцінки посухостійкості сортів озимої пшениці / І.П. Григорюк, В.І. Ткачов, О.І. Серга, М.Ф. Михальський. - Опубл. 15.04.2002, Бюл. №4. 4. Зерова М.Д., Никитенко Г.Н., Нарольський Н.Б. и др. Каштановая минирующая моль в Украине. - Киев: ТОВ «Вегес», 2007. - 87 с. 5. Ленинджер А. Биохимия. Молекулярные основы структуры и функции клетки. - М.: Мир, 1976. - 957 с. 6. Никулина Г.Н. Методы определения нуклеотидов в растениях. - Л.: Наука, 1989. - 88 с. 7. Соколов В.Б. Каштан. - М.: Лесн. пром-ть, 1984. - 80 с. 8. Трибель С.М., Гаманова О.М., Свєнтославскі Я. Каштанова мінуюча міль. - К.: Колобіг, 2008. 72 с. 9. Atkinson D.E., Walton G.M. Adenosine triphosphate conservation in metabolic regulation. Rat Бал 1 1 1 1 1 1 1 8-9 8-9 6-7 6-7 6-7 Ступінь пошкоджен- Ступінь охоплення ня листків каштано- мінами площі листвою мінуючою міллю кової поверхні, % Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Дуже сильна 85 Дуже сильна 85 Сильна 70 Сильна 70 Сильна 70 liver citrate glavage enzyme // J. Biol. Chem. - 1967. 242, №13. - P. 3239-3241. 10. Atkinson D.E. The energy charge of the adenilate pool as a regulatory parameter. Interaction with feedback modifiers // Biochem. - 1968. - Vol.7. P. 4030-4034. 11. Hoffmann P. Photosynthesis and biomass production in higher plants // Photosynthesis solar energy conversion and storages: Proceed. 18 th Conf. SEV-Problem (Warszawa 1-18.02.1980). Warszawa: Univ.Press, 1983. - P. 239. 12. Hoffmann P., Leopold D. Energiekonvertierung in pflanzenlichen System // Wiss. Z. Humboldt - Univ.Berlin.Math. - Naturwiss.R. 1984. - 33, №4. - S. 275-279. 13. Miginiac - Maslow M, Hoarou A. The adenine nucleotide levels and the adenylate energy charge values of different Triticum and Aegilops species // Z. Pflanzenphysiol. - 1979. - 93, №5. - P. 387-394. 14. Quebedeaux B. Adenylate and nicotianamide nucleotides in developing soybean seeds duringseed-till // Plant Physiol. - 1981. - 68, №1. - P. 23-27. 15. Stitt M.R., Lilley Me, Heldt H.W. Adenine nucleotide levels in the cytosol chloroplasts and mitochondria of wheat leaf protoplasts // Plant Physiol. - 1982. - 70, №4. - P. 971-977. 11 Комп’ютерна верстка А. Рябко 59330 Підписне 12 Тираж 23 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod to assess resistance of species and hybrids of plants of horse chestnut genus (aesculus l.) to horse-chestnut leafminer (cameraria ohridella deschka et dimic)
Автори англійськоюHryhoriuk Ivan Panasovych, Demchuk Tetiana Leonidivna, Melnychuk Maksym Dmytrovych, Serha Olesia Ivaniva, Dubrovin Valerii Oleksandrovych, Mashkovska Svitlana Petrivna
Назва патенту російськоюСпособ оценки стойкости видов и гибридов растений рода каштан конский (aesculus l.) к каштановой минирующей моли (cameraria ohridella deschka et dimic)
Автори російськоюГригорюк Иван Афанасьевич, Демчук Татьяна Леонидовна, Мельничук Максим Дмитриевич, Серга Олеся Ивановна, Дубровин Валерий Александрович, Машковская Светлана Петровна
МПК / Мітки
МПК: A01G 13/00
Мітки: ohridella, гібридів, молі, cameraria, aesculus, стійкості, роду, каштанової, мінуючої, рослин, видів, оцінки, dimic, deschka, гіркокаштан, спосіб
Код посилання
<a href="https://ua.patents.su/6-59330-sposib-ocinki-stijjkosti-vidiv-i-gibridiv-roslin-rodu-girkokashtan-aesculus-l-do-kashtanovo-minuyucho-moli-cameraria-ohridella-deschka-et-dimic.html" target="_blank" rel="follow" title="База патентів України">Спосіб оцінки стійкості видів і гібридів рослин роду гіркокаштан (aesculus l.) до каштанової мінуючої молі (cameraria ohridella deschka et dimic)</a>
Спосіб оцінки рівня життєздатності самиць лабораторних популяцій видів роду trichogramma

Номер патенту: 42889
Опубліковано: 27.07.2009
Автори: Дрозда Валентин Федорович, Кочерга Марина Олександрівна
МПК: A01K 67/00
Мітки: лабораторних, популяцій, рівня, trichogramma, оцінки, життєздатності, спосіб, роду, видів, самиць
Формула / Реферат:
Спосіб оцінки рівня життєздатності самиць лабораторних популяцій видів роду Trichogramma, що включає вирощування їх в лабораторіях, з наступною процедурою оцінки за тестовими характеристиками, який відрізняється тим, що зразу після відродження самиць трихограми проводять прижиттєве операційне препарування гонад самиць, з наступним їх фарбуванням органічним фарбником у вигляді водного розчину, причому високожиттєздатними вважаються популяції...
Спосіб переводу видів роду trichogramma на нового хазяїна

Номер патенту: 25002
Опубліковано: 25.07.2007
Автор: Дрозда Валентин Федорович
МПК: A01K 67/00
Мітки: видів, trichogramma, хазяїна, переводу, спосіб, нового, роду
Формула / Реферат:
Спосіб переводу видів роду Trichogramma на нового хазяїна, що включає їх вирощування в яйцях лускокрилих комах, який відрізняється тим, що самицям трихограми відразу після спаровування пропонують яйця нового хазяїна і одноразово згодовують їжу, до складу якої входить 3-5 %-ний водний розчин меду у суміші з нативним попередником рибонуклеїнової кислоти (РНК) у концентрації 0,02-0,003 %.
Спосіб контролю шкідливості популяцій видів роду ostrinia (lepidoptera, pyraustidae)

Номер патенту: 28182
Опубліковано: 26.11.2007
Автор: Дрозда Валентин Федорович
МПК: A01G 13/00, A01N 63/00
Мітки: pyraustidae, шкідливості, видів, популяцій, lepidoptera, ostrinia, контролю, спосіб, роду
Формула / Реферат:
Спосіб контролю шкідливості популяції видів роду Ostrinia (Lepidoptera, Pyraustidae), що включає використання трихограми, який відрізняється тим, що перед розселенням в агроценози трихограму спаровують, причому після спаровування заражають впродовж 1-1,5 години яйцями комахи-хазяїна у біолабораторії, крім того, перед розселенням трихограми та ізоляції її від яєць лабораторного хазяїна, імаго згодовують дієту у складі водного розчину...
Спосіб визначення стійкості сортів та гібридів картоплі до вірусного раку

Номер патенту: 10063
Опубліковано: 15.11.2005
Автори: Голик Іван Васильович, Мовчан Олексій Миколайович, Хома Інга Павлівна, Мілашевська Жанна Іванівна, Хом'як Віра Василівна, Мельник Павло Олексійович
МПК: A01G 7/00, G01N 33/00
Мітки: картоплі, сортів, спосіб, визначення, стійкості, гібридів, раку, вірусного
Формула / Реферат:
Спосіб визначення стійкості сортів та гібридів картоплі до вірусного раку, що включає застосування метеорологічних факторів для активізації патологічних процесів у заражених рослин, який відрізняється тим, що дослідні сортозразки спочатку заражають однаковими дозами інфекції вірусу, а потім заражені рослини піддають дії сприятливих для розвитку патогену метеофакторів, під впливом яких патоген протягом 4-5 діб викликає максимальне ураження...
Спосіб оптимізації вирощування видів роду trichogramma (hymenoptera, trichogrammatidae)

Номер патенту: 35292
Опубліковано: 10.09.2008
Автор: Дрозда Валентин Федорович
МПК: A01K 67/00
Мітки: роду, trichogrammatidae, оптимізації, спосіб, видів, trichogramma, hymenoptera, вирощування
Формула / Реферат:
Спосіб оптимізації вирощування видів роду Trichogramma (Hymenoptera, Trichogrammatidae), що включає лабораторний режим вирощування в яйцях комах-живителів, який відрізняється тим, що лабораторні популяції трихограми формують шляхом щорічного збору паразита із яєць природних популяцій лускокрилих видів комах з незайманих природних угідь, крім того в лабораторних умовах проводять 2-3-разовий пасаж через яйця совок, а перед розселенням...
Попередній патент: Спосіб визначення типу імунної реакції організму хворих на інтраабдомінальну інфекцію (перитоніт та абдомінальний сепсис)
Наступний патент: Спосіб одержання лектину бруслини європейської (euonymus europaea l.)
Випадковий патент: Спосіб ароматизації виробу