Спосіб оцінки стійкості видів і гібридів рослин роду гіркокаштан (aesculus l.) до каштанової мінуючої молі (cameraria ohridella deschka et dimic)
Номер патенту: 59624
Опубліковано: 25.05.2011
Автори: Мельничук Максим Дмитрович, Григорюк Іван Панасович, Машковська Світлана Петрівна, Дубровін Валерій Олександрович, Серга Олеся Іванівна, Демчук Тетяна Леонідівна





Формула / Реферат
Спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et Dimic), що включає пряме кількісне визначення вмісту ендогенних фітогормонів, зокрема індоліл-3-оцтової кислоти (ІОК), зеатину, зеатинрибозиду й абсцизової кислоти (АБК) в листках рослин, який відрізняється тим, що як основний показник використовують показник інтегрального біохімічного фітогормонального балансу Вр, при цьому відбір свіжих листків нижнього, середнього і верхнього ярусів видів й гібридів рослин роду Гіркокаштан проводять до початку появи каштанової мінуючої молі (фаза початку цвітіння рослин), далі здійснюють екстракцію, очищення та визначають вміст ендогенних фітогормонів в листках рослин, а потім розраховують показник інтегрального біохімічного фітогормонального балансу Вр за формулою:
![]() ,
,
де Вр - інтегральний показник, який характеризує сумарне відношення вмісту ендогенних фітогормонів стимулюючої дії (ЮК, зеатину і зеатинрибозиду) до кількості інгібітора росту АБК; при цьому, якщо його значення становить більше 10,0, то види та гібриди рослин роду Гіркокаштан відносяться до стійких до каштанової мінуючої молі, якщо менше 10,0 - до нестійких.
Текст
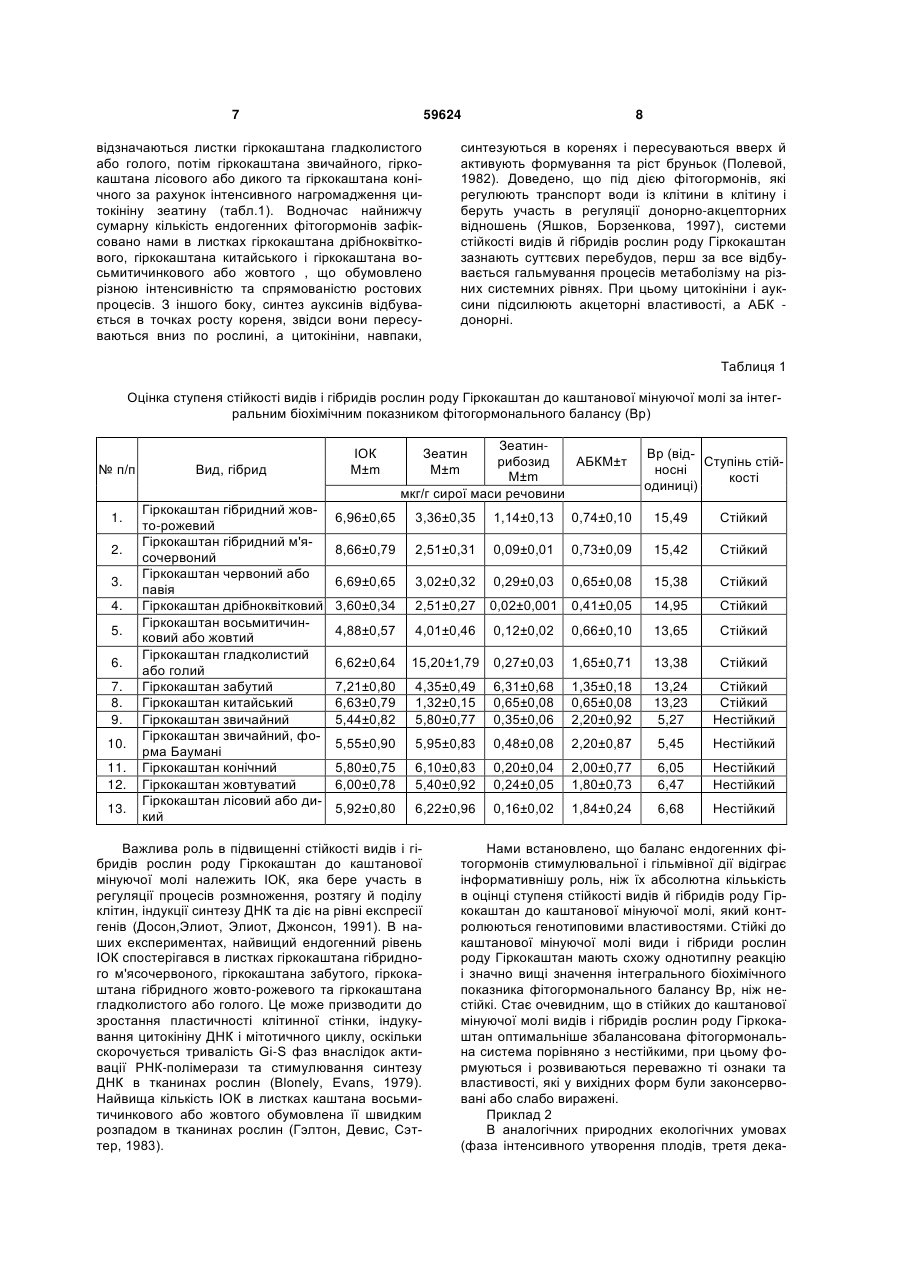
Спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et Dimic), що включає пряме кількісне визначення вмісту ендогенних фітогормонів, зокрема індоліл-3оцтової кислоти (ІОК), зеатину, зеатинрибозиду й абсцизової кислоти (АБК) в листках рослин, який відрізняється тим, що як основний показник ви U 2 (19) 1 3 умовах змін клімату. Спадкова мінливість геному ґрунтується на переміщенні рухливих контролюючих елементів, що відіграє важливу роль в мікро- і макроеволюції, зокрема, у швидкій адаптації організмів до несприятливих умов середовища. Доведено, що непостійність геному індукує перебудови диференціації клітин і систем регуляції активності генів, які спричиняють специфічні зміни їх структури та кількості в індивідуальному розвитку рослин (Хосин, 1984). Наявні чинники гальмують проведення ефективної селекції та застосування різноманіття генофонду рослин гіркокаштана для озеленення міських територій України. Для рослинного організму характерним є цілий спектр складних регуляторних механізмів, серед яких визначальне місце займає фітогормональна регуляція (Sembdner, Yness etol. 1980). Фітогормони (ауксини, цитокініни, гібереліни, абсцизова кислота, етилен) утворюються в органах рослин в -8 -5 надзвичайно низьких кількостях (10 -10 моль), які контролюють інтенсивність і спрямованість процесів обміну речовин й залежно від концентрації пригнічують чи стимулюють ріст та розвиток рослин (Біол. словник, 1986). Вони є медіаторами між екологічними факторами і рослинними організмами, причому містяться у тканинах у вільному й зв'язаному стані, різняться між собою за фізіологічними функціями (Иванов и др. 1999), впливають на всі етапи реалізації генетичної інформації від реплікації (подвоєння) ДНК до синтезу білка (Кефелі, Прусанова, 1985). Доведено, що універсальні перебудови в функціонуванні фітогормональної системи являють собою одну із ключових ланок в спектрі неспецифічних реакцій, які слугують основою для формування механізмів адаптації рослин до екстремальних факторів середовища (Пуревич, 1979). З огляду на це, роль фітогормонів у координації метаболічних і структурних змін за умов дії на рослини гіркокаштана каштанової мінуючої молі в літературі не з'ясована. Так, згідно наявних даних (Гэлстон, Дэвис, Сэттер, 1983), пропонується концепція гормонального балансу в рослинному організмі, яка грунтується на залежності між інгібітором росту АБК по відношенню до стимуляторів росту - гіберелінів і цитокінінів з метою оцінки сортової й видової реакції рослин на стресові фактори середовища. Проте, таке відношення недостатньо інформативне, оскільки в листках рослин не враховано сумарного вмісту ІОК. В іншій роботі (Кудоярова, Гюли-Заде, 1990), методом імуноферментного аналізу встановлено кореляцію між вмістом ендогенних фітогормонів і швидкістю ростових процесів в рослинах за дії гербіцидів. Однак, використання данної кореляції для оцінки ступеня стійкості рослин гіркокаштана до каштанової мінуючої молі є сумнівним у зв'язку з швидким гідролізом зв'язаних форм фітогормонів, індукованих токсичним впливом гербіцидів. Для оцінки ступеня проліферативної активності у кореневих меристемах етиольованих проростків рослин кукурудзи, які вирощували в темряві, використано співвідношення сумарної концентрації ендогенних цитокінінів до ІОК (Полевой, Полевой, 1992), в якому не враховано кількості АБК, що ме 59624 4 тодично є некоректним з точки зору постановки експерименту. З метою усунення наявних недоліків у запропонованих іншими авторами рішеннях запропоновано спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі за інтегральним біохімічним показником Вр. В основу корисної моделі покладене завдання розробки ефективного й інформативного способу оцінки стійкості видів і гібридів роду Гіркокаштан до каштанової мінуючої молі шляхом застосування ефекту прямого кількісного визначення вмісту ендогенних фітогормонів - індоліз-3-оцтової кислоти (ІОК), абсцизової кислоти (АБК), зеатину і зеатинрибозиду із розрахунком інтегрального біохімічного показника фітогормонального балансу Вр у комплексній взаємодії суттєвих ознак нового способу. Завдання вирішується завдяки тому, що в способі оцінки стійкості видів і гібридів рослин роду гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et dimic), що включає пряме кількісне визначення вмісту ендогенних фітогормонів, зокрема індоліл-3-оцтової кислоти (ІОК), зеатину, зеатинрибозиду й абсцизової кислоти (АБК) в листках рослин, в якості основного показника використовують показник інтегрального біохімічного фітогормонального балансу Вр, при цьому відбір свіжих листків нижнього, середнього і верхнього ярусів видів й гібридів рослин роду Гіркокаштан проводять до початку появи каштанової мінуючої молі (фаза початку цвітіння рослин), далі здійснюють екстракцію, очищення та визначають вміст ендогенних фітогормонів в листках рослин, а потім розраховують показник інтегрального біохімічного фітогормонального балансу Вр за формулою: ІОК Зеатин Зеатинтроб озид, ВР АБК Вр = ІОК+Зеатин+Зеатинрибозид, де Вр - інтегральний показник, який характеризує сумарне відношення вмісту ендогенних фітогормонів стимулюючої дії (ІОК, зеатину і зеатинрибозиду) до кількості інгібітора росту АБК; при цьому, якщо його значення становить більше 10, то види та гібриди рослин роду Гіркокаштан відносяться до стійких до каштанової мінуючої молі, якщо менше 10 - до нестійких. Нами встановлено наявність тісного взаємозв'язку між фітогормональним балансом і системами стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі. Ідея наукового пошуку полягала в тому, що гусениці першого, другого і третього віку, які проникають під кутикулу в шар клітин верхнього епідермісу, впродовж вегетаційного періоду порушують фізіологічні функції листкового апарату, причому живляться виключно лише клітинним соком (Зерова і ін., 2007; Трибель і ін., 2008), в якому в значних кількостях містяться фітогормони, вуглеводи, органічні кислоти, амінокислоти, алкалоїди, які разом з напівпроникністю протоплазми й пружністю клітинних стінок забезпечують належний тургор, який здійснює збереження органами рослин форми та структури в просторі, а також їх опір деформованій дії механічних 5 факторів (Роздорский, 1949). Надалі гусениці четвертого і п'ятого віку живляться тільки тканинами полісадної паренхіми листкової пластинки рослин гіркокаштана звичайного (Зерова і ін., 2007; Трибель і ін. 2008), які є джерелом значної кількості фітогормонів стимулювальної і гальмівної дії. Об'єктами досліджень слугували 13 видів і гібридів рослин роду Гіркокаштан віком 25-30 років, а саме: гіркокаштан звичайний (Aesculus hippocastanum L), гіркокаштан звичайний, форма Баумані (Aesculus hippocastanum, f.baumanii С.К. Schneid), гіркокаштан конічний (Aesculus turbinata Blume.), гіркокаштан лісовий або дикий (Aesculus silvatica Bartz.), гіркокаштан жовтуватий (Aesculus flava Ait), гіркокаштан китайський (Aesculus chinensis Bunge.), гіркокаштан забутий (Aesculus neglecta Lindl), гіркокаштан гладколистий або голий (Aesculus glabra Willd.), гіркокаштан восьмитичинковий або жовтий (Aesculus octandra March), гіркокаштан дрібноквітковий (Aesculus purviflora Walt.), гіркокаштан червоний або павія (Aesculus pavia L.), гіркокаштан гібридний (Aesculus hibrida D.C.), гіркокаштан гібридний м'ясочервоний {Aesculus cornea Hayne.), які інтродуковані в Національному ботанічному саду імені М.М. Гришка HAH України та частково зростають в Голосіївському лісі м. Києва. Запропонований нами спосіб реалізується наступним чином. До початку появи каштанової мінуючої молі (фаза початок цвітіння рослин) відбирають по 3 г матеріалу із свіжих листків нижнього, середнього і верхнього ярусів видів і гібридів рослин роду Гіркокаштан, які фіксують рідким азотом та екстрагують 96% етиловим спиртом в чотирьохкратній повторності. Кількісний вміст ендогенних фітогормонів в листках рослин визначають за допомогою спектроденситометричної тонкошарової хроматографії (Савинский, Кофман и др. 1987). Очищення і концентрування фітогормонів виконують на силікагельових пластинках (Silufol, UV254, Chemapol, Чехія) в суміші розчинників хлороформ, 12,5% водний аміак, етилацетатоцтова кислота. Для приготування стандартних розчинів використовують цис-транс-АБК фірми «Sigma», зеатин і зеатинрибозид фірми «Serva» та ІОК фірми «Fluka» A.G. Очищені екстракти ІОК і АБК розділяють на пластинках з оксидом кремнію (Merck, Art 5715, F254, ФРН) в суміші розчинників хлороформетилацетат-оцтова кислота у співвідношенні 100:100:2, а зеатину й зеатинрибозиду на пластинках з оксидом алюмінію (Merck, Art 5715, F254, ФРН) в суміші розчинників хлороформ-оцтова кислота у співвідношенні 19:1. Кількісне детектування ендогенних фітогормонів здійснюють за допомогою сканувального спектроденситометра Camag TLC Scanner II (Швейцарія). Баланс ендогенних фітогормонів в листках рослин гіркокаштана розраховують за запропонованою нами формулою: ІОК Зеатин Зеатинтроб озид, ВР АБК де Вр - інтегральний показник, який характеризує сумарне відношення вмісту ендогенних фітогормонів стимулювальної дії (ІОК, зеатину і зеа 59624 6 тинрибозиду) до кількості інгібітора росту АБК. Його використовують тільки за умов визначення кількості фітогормонів методом спектроденситометричної тонкошарової хроматографії, який відзначається повнотою їх виділення з однієї рослинної проби. Особливими результатами даного способу є: - підвищення ефективності, точності і інформативності методу спектроденситометричної тонкошарової хроматографії для оцінки ступеня стійкості видів й гібридів рослин роду Гіркокаштан до каштанової мінуючої молі за інтегральним біохімічним показником фітогормонального балансу Вр, який враховує також вміст ІОК в листках; - забезпечення повноти виділення ендогенних фітогормонів з листків рослин, що максимально підвищує достовірність отриманих результатів. Приклад 1 Проводили визначення вмісту ІОК, зеатину, зеатинрибозиду і АБК в листках видів й гібридів роду Гіркокаштан та величин інтегрального біохімічного показника фітогормонального балансу Вр з метою оцінки стійкості рослин до каштанової мінуючої молі (таблиця 1). Встановлено, що в листках видів і гібридів рослин роду Гіркокаштан до початку масової появи каштанової мінуючої молі в найбільшій кількості містяться ендогенні фітогормони ІОК та активне похідне цитокініну зеатин, що виявляється в цисформі в тРНК. В значно меншій кількості в листках рослин локалізоване похідне цитокініну зеатинрибозид, наявність якого в клітинах розглядають як спосіб регуляції рівня цитокінінової активності, який може знаходитись у вигляді транспортної або запасної форми в судинній системі ксилеми (Полевой, 1982). Дещо більші концентрації, ніж зеатинрибозиду, нами визначено в листках різних видів і гібридів гіркокаштана стресового гормону АБК, яка впливає на замикальні клітини продихів шляхом «викачування» з них пулів води з метою їх швидкого закривання. Роль АБК в рухах продихів полягає в тому, що вона «вимушує» іони калію покинути замикальні клітини, в результаті продихи закриваються, зменшуються втрати води і рослини гіркокаштана легше переносять стресовий період (Гэлстон и др., 1983). Із наведених даних видно, що найбільш високий вміст АБК характерний для листків гіркокаштана звичайного, форма Баумані, потім гіркокаштана звичайного конічного, гіркокаштана жовтуватого та гіркокаштана лісового або дикого. Згідно існуючих даних (Barlow, Pilet, 1984), в присутності АБК знижується матрична активність хроматину, синтез білків та РНК в кореневих меристемах рослин. Заодно в листках гіркокаштана забутого і гіркокаштана гібридного жовто-рожевого зафіксовано найбільш високу концентрацію зеатину, який переважно синтезується в кореневій системі, звідси з током води по ксилемі транспортується в надземну частину і гальмує процеси передчасного старіння рослин. Наявність значної кількості транспортних форм цитокінінів пояснюють інтенсивним надходженням фітогормонів із кореневої системи рослин (Reid, Wample, 1985). Аналіз результатів засвідчує, що найвищим сумарним фітогормональним пулом 7 59624 відзначаються листки гіркокаштана гладколистого або голого, потім гіркокаштана звичайного, гіркокаштана лісового або дикого та гіркокаштана конічного за рахунок інтенсивного нагромадження цитокініну зеатину (табл.1). Водночас найнижчу сумарну кількість ендогенних фітогормонів зафіксовано нами в листках гіркокаштана дрібноквіткового, гіркокаштана китайського і гіркокаштана восьмитичинкового або жовтого , що обумовлено різною інтенсивністю та спрямованістю ростових процесів. З іншого боку, синтез ауксинів відбувається в точках росту кореня, звідси вони пересуваються вниз по рослині, а цитокініни, навпаки, 8 синтезуються в коренях і пересуваються вверх й активують формування та ріст бруньок (Полевой, 1982). Доведено, що під дією фітогормонів, які регулюють транспорт води із клітини в клітину і беруть участь в регуляції донорно-акцепторних відношень (Яшков, Борзенкова, 1997), системи стійкості видів й гібридів рослин роду Гіркокаштан зазнають суттєвих перебудов, перш за все відбувається гальмування процесів метаболізму на різних системних рівнях. При цьому цитокініни і ауксини підсилюють акцеторні властивості, а АБК донорні. Таблиця 1 Оцінка ступеня стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі за інтегральним біохімічним показником фітогормонального балансу (Вр) № п/п 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. Вид, гібрид Гіркокаштан гібридний жовто-рожевий Гіркокаштан гібридний м'ясочервоний Гіркокаштан червоний або павія Гіркокаштан дрібноквітковий Гіркокаштан восьмитичинковий або жовтий Гіркокаштан гладколистий або голий Гіркокаштан забутий Гіркокаштан китайський Гіркокаштан звичайний Гіркокаштан звичайний, форма Баумані Гіркокаштан конічний Гіркокаштан жовтуватий Гіркокаштан лісовий або дикий ІОК М±m Зеатинрибозид АБКМ±т М±m мкг/г сирої маси речовини Зеатин М±m Вр (відСтупінь стійносні кості одиниці) 6,96±0,65 3,36±0,35 1,14±0,13 0,74±0,10 15,49 Стійкий 8,66±0,79 2,51±0,31 0,09±0,01 0,73±0,09 15,42 Стійкий 6,69±0,65 3,02±0,32 0,29±0,03 0,65±0,08 15,38 Стійкий 3,60±0,34 2,51±0,27 0,02±0,001 0,41±0,05 14,95 Стійкий 4,88±0,57 4,01±0,46 0,12±0,02 0,66±0,10 13,65 Стійкий 6,62±0,64 15,20±1,79 0,27±0,03 1,65±0,71 13,38 Стійкий 7,21±0,80 6,63±0,79 5,44±0,82 4,35±0,49 1,32±0,15 5,80±0,77 6,31±0,68 0,65±0,08 0,35±0,06 1,35±0,18 0,65±0,08 2,20±0,92 13,24 13,23 5,27 Стійкий Стійкий Нестійкий 5,55±0,90 5,95±0,83 0,48±0,08 2,20±0,87 5,45 Нестійкий 5,80±0,75 6,00±0,78 6,10±0,83 5,40±0,92 0,20±0,04 0,24±0,05 2,00±0,77 1,80±0,73 6,05 6,47 Нестійкий Нестійкий 5,92±0,80 6,22±0,96 0,16±0,02 1,84±0,24 6,68 Нестійкий Важлива роль в підвищенні стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі належить ІОК, яка бере участь в регуляції процесів розмноження, розтягу й поділу клітин, індукції синтезу ДНК та діє на рівні експресії генів (Досон,Элиот, Элиот, Джонсон, 1991). В наших експериментах, найвищий ендогенний рівень ІОК спостерігався в листках гіркокаштана гібридного м'ясочервоного, гіркокаштана забутого, гіркокаштана гібридного жовто-рожевого та гіркокаштана гладколистого або голого. Це може призводити до зростання пластичності клітинної стінки, індукування цитокініну ДНК і мітотичного циклу, оскільки скорочується тривалість Gi-S фаз внаслідок активації РНК-полімерази та стимулювання синтезу ДНК в тканинах рослин (Blonely, Evans, 1979). Найвища кількість ІОК в листках каштана восьмитичинкового або жовтого обумовлена її швидким розпадом в тканинах рослин (Гэлтон, Девис, Сэттер, 1983). Нами встановлено, що баланс ендогенних фітогормонів стимулювальної і гільмівної дії відіграє інформативнішу роль, ніж їх абсолютна кільькість в оцінці ступеня стійкості видів й гібридів роду Гіркокаштан до каштанової мінуючої молі, який контролюються генотиповими властивостями. Стійкі до каштанової мінуючої молі види і гібриди рослин роду Гіркокаштан мають схожу однотипну реакцію і значно вищі значення інтегрального біохімічного показника фітогормонального балансу Вр, ніж нестійкі. Стає очевидним, що в стійких до каштанової мінуючої молі видів і гібридів рослин роду Гіркокаштан оптимальніше збалансована фітогормональна система порівняно з нестійкими, при цьому формуються і розвиваються переважно ті ознаки та властивості, які у вихідних форм були законсервовані або слабо виражені. Приклад 2 В аналогічних природних екологічних умовах (фаза інтенсивного утворення плодів, третя дека 9 59624 да липня 2010 р.) шляхом маршрутного обстеження проводили візуальну оцінку ступеня пошкодження листків тих же видів і гібридів рослин роду Гіркокаштан каштановою мінуючою міллю, згідно запропонованої в літературі (Трибель і ін., 2008) універсальної дев'ятибальної шкали з деякими нашими уточненнями стосовно кількісних показників (таблиця 2). Виявлено, що пошкодження листків каштановою мінуючою міллю у гіркокаштана гібридного жовто-рожевого, гіркокаштана гібридного м'ясочервоного, гіркокаштана червоного або павія, гіркокаштана дрібноквіткового, гіркокаштана восьмитичинкового або жовтого, гіркокаштана гладколистого або голого і гіркокаштана забутого відсутнє (1 бал), що спричинено, очевидно, відсутністю або нестачею живильного субстрату, екологічними умовами зростання та генетичним надходженням. Водночас нами зафіксовано дуже сильне (8-9 балів, 85% охоплення мінами площі листкової поверхні) пошкодження каштановою мінуючою міллю листків гіркокаштана звичайного, форма Баумані й сильне (6-7 балів, 70% охоплення мінами площі листкової поверхні) у гіркокаштана конічного, гіркокаштана жовтуватого та гіркокаштана лісового або дикого (табл.2). Отримані 10 результати тісно корелюють із величиною інтегрального біохімічного показника фітогормонального балансу Вр, який у листках стійких до каштанової мінуючої молі видів і гібридів рослин роду Гіркокаштан коливався в межах від 13,23 до 15,49, а у нестійких - від 5,27 до 6,68. Таким чином, величина фітогормонального балансу Вр зв'язана з стійкістю видів і гібридів рослин роду Гіркокаштан прямо пропорційною залежністю. Чим менше величина показника Вр в листках рослин до і після дії каштанової мінуючої молі, тим стійкіший до неї генотип. Якщо значення величини інтегрального біохімічного показника фітогормонального балансу Вр становить більше 10,0, то види і гібриди рослин роду Гіркокаштан відноситься до стійких до каштанової мінуючої молі, якщо менше 10,0 - до нестійких. Запропонований нами спосіб дозволяє здійснювати достовірну і точну оцінку ступеня стійкості видів і гібридів рослин роду Гіркокаштан до каштанової мінуючої молі, який може бути використаний в селекційній роботі та для розробки заходів з озеленення міських територій. Таблиця 2 Оцінка ступеня пошкодження листків видів і гібридів рослин роду Гіркокаштан каштановою мінуючою міллю № п/п 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. Вид, гібрид Гіркокаштан гібридний жовто-рожевий Гіркокаштан гібридний м'ясочервоний Гіркокаштан червоний або павія Гіркокаштан дрібноквітковий Гіркокаштан восьмитичинковий або жовтий Гіркокаштан гладколистий або голий Гіркокаштан забутий Гіркокаштан китайський Гіркокаштан звичайний Гіркокаштан звичайний, форма Баумані Гіркокаштан конічний Гіркокаштан жовтуватий Гіркокаштан лісовий або дикий Джерела інформації: 1. Біологічний словник. За ред. академіка АН УРСР К.М. Ситника і члена-кореспондента АН УРСР В.О. Топачевського. Видання друге, перероблене і доповнене. - К.: Головна редакція ІРЕ, 1986. - 678 с. 2. Гэлстон А., Дэвис П., Сэттер Р. Жизнь зеленого растения. - М.: Мир, 1983. - 543 с. 3. Гуревич Л.С. Роль гормонального баланса ауксина и этилена в адаптационных реакциях высших растений // Ботан. журн. - 1979.-64, №11. - С. 1600-1608. 4. Досон Р., Эллиот Д., Джонс К. Справочник по биохимии. - М.: Мир, 1991. - 543 с. 5. Зерова М.Д., Никитенко Г.Н., Нарольський Н.Б. и др. Каштановая минирующая моль в Украине. - Киев: ТОВ «Вегес», 2007. - 87 с. Бал 1 1 1 1 1 1 1 1 8-9 8-9 6-7 6-7 6-7 Ступінь пошкоджен- Ступінь охоплення ня листків каштано- мінами площі листвою мінуючою міллю кової поверхні, % Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Відсутня 0 Дуже сильна 85 Дуже сильна 85 Сильна 70 Сильна 70 Сильна 70 6. Иванова А.Б., Анцичина Л.А., Ярин А.П. Современные аспекты и изучения фитогормонов // Цитология. - 1999. - 41, №10. - С. 835-847. 7. Кефели В.И., Прусакова Л.Д. Химические регуляторы растений. - М.: Знание, 1985. - 63 с. 8. Кудоярова Г.Р., Гюли-Заде В.З. Уровень фитогормонов в растениях: физиологическое значение и способы регуляции // Тез. докл. Второго съеpда Вш. об-ва физиологов растений (Минск, 24-29 сентября 1990 г.). - Москва, 1990. - С. 48. 9. Полевой В.В. Фитогормоны. - Л.: Изд-во Ленингр. у-та, 1982. - 249 с. 10. Полевой В.В., Полевой А.В. Эндогенные фитогормоны этиолированых проростков кукурузы // Физиология растений. - 1992. - 39, №6. - С. 11651174. 11 59624 11. Раздорский В.Ф. Анатомия растений. - М: Гос. изд-во «Советская наука», 1949. -524 с. 12. Савинский С.В., Кофман И.Ш., Кофанов В.И., Стасевская И.П. Методические подходы к определению фитогормонов с помощью спектроденситометрической тонкослойной хроматографии // Физиология и биохимия культ.растений. 1987. - 10, №2. - С. 195-200. 13. Трибель С.М., Гаманова О.М., Свєнтославскі Я. Каштанова мінуюча міль. - К.: Колобіг, 2008. - 72 с. 14. Хосин Р.Б. Непостоянство генома. - М.: Наука, 1984. - 472 с. 15. Яшков М.Ю., Борзенкова Р.А. Характеристика гормональной системы различных видов картофеля в связи с особенностями донорноакцепторных связей // Тез.докл. IV Междунар. конф. «Регуляторы роста и развития растений.» М.: Моск. с.-х. акад., 1997. - С.44-45. 16. Barlow P.W., Pilat P.-E. The effect of abscisis in the maize root meristem // Physiol.plant. - 1984. 62, №2. - p. 125-132. Комп’ютерна верстка А. Рябко 12 17. Blanery L.M., Evans T.A. Cell dynamics studies the pericycle of radish seedling roots // Plant Sci. Let. - 1979. - 14, №1. - P. 79-83. 18. Reid D.M., Wample R.Z. Water relations and plant hormones // Hormonal regulation of development. - Berlin etc.: Springer, 1985. - P.513578. 19. Savinski S.V., Tkachev V.J., Grigoryuk LA. Quantitive estimatien of phytohormonal balance with coefficient Bp (Abstr. 8th Congress of the Federation of European Soc. Of Plant Physiol., Aug. 23-28, 1992, Antwerpen, Belgium) // Physiol. Plant. - 1992. - 85 (3), №2. - P. 30. 20. Sembdner C., Grass D., Liebish H.W., Schneider G. Biosynthesis and metabolism of plant hormones // Encyclopedia of Plant Physiologic Berlin etc.: Springer. 1980. - 9, №5. - P. 281-444. 21. Wright S.T.C An increasing in the inhibitor "B" content of detached wheat leaves follwing a period of wilting // Planta. - 1969. - 86, №1. - P.10-18. Підписне Тираж 24 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for assessing resistance of species and hybrids of plants of horse chestnut genus (aesculus l.) to chestnut miner moth (cameraria ohridella deschka et dimic)
Автори англійськоюHryhoriuk Ivan Panasovych, Demchuk Tetiana Leonidivna, Melnychuk Maksym Dmytrovych, Serha Olesia Ivaniva, Dubrovin Valerii Oleksandrovych, Mashkovska Svitlana Petrivna
Назва патенту російськоюСпособ оценки стойкости видов и гибридов растений рода конский каштан (aesculus l.) к каштановой минирующей моли (cameraria ohridella deschka et dimic)
Автори російськоюГригорюк Иван Афанасьевич, Демчук Татьяна Леонидовна, Мельничук Максим Дмитриевич, Серга Олеся Ивановна, Дубровин Валерий Александрович, Машковская Светлана Петровна
МПК / Мітки
МПК: A01G 13/00
Мітки: мінуючої, спосіб, dimic, deschka, молі, гіркокаштан, стійкості, каштанової, оцінки, cameraria, рослин, гібридів, aesculus, видів, ohridella, роду
Код посилання
<a href="https://ua.patents.su/6-59624-sposib-ocinki-stijjkosti-vidiv-i-gibridiv-roslin-rodu-girkokashtan-aesculus-l-do-kashtanovo-minuyucho-moli-cameraria-ohridella-deschka-et-dimic.html" target="_blank" rel="follow" title="База патентів України">Спосіб оцінки стійкості видів і гібридів рослин роду гіркокаштан (aesculus l.) до каштанової мінуючої молі (cameraria ohridella deschka et dimic)</a>
Спосіб оцінки стійкості видів і гібридів рослин роду гіркокаштан (aesculus l.) до каштанової мінуючої молі (cameraria ohridella deschka et dimic)

Номер патенту: 59330
Опубліковано: 10.05.2011
Автори: Машковська Світлана Петрівна, Серга Олеся Іванівна, Григорюк Іван Панасович, Демчук Тетяна Леонідівна, Дубровін Валерій Олександрович, Мельничук Максим Дмитрович
МПК: A01G 13/00
Мітки: рослин, гібридів, cameraria, стійкості, deschka, видів, каштанової, роду, гіркокаштан, молі, мінуючої, dimic, ohridella, спосіб, aesculus, оцінки
Формула / Реферат:
Спосіб оцінки стійкості видів і гібридів рослин роду Гіркокаштан (Aesculus L.) до каштанової мінуючої молі (Cameraria ohridella deschka et dimic), що ґрунтується на кількісному визначенні вмісту АМФ, АДФ й АТФ в листках рослин, який відрізняється тим, що як інтегральний біоенергетичний показник енергозабезпечення використовують величину енергетичного заряду аденозинфосфатної системи [ЕЗ], при цьому відбір свіжих листків нижнього, середнього...
Спосіб оцінки рівня життєздатності самиць лабораторних популяцій видів роду trichogramma

Номер патенту: 42889
Опубліковано: 27.07.2009
Автори: Дрозда Валентин Федорович, Кочерга Марина Олександрівна
МПК: A01K 67/00
Мітки: спосіб, роду, життєздатності, видів, популяцій, trichogramma, самиць, рівня, оцінки, лабораторних
Формула / Реферат:
Спосіб оцінки рівня життєздатності самиць лабораторних популяцій видів роду Trichogramma, що включає вирощування їх в лабораторіях, з наступною процедурою оцінки за тестовими характеристиками, який відрізняється тим, що зразу після відродження самиць трихограми проводять прижиттєве операційне препарування гонад самиць, з наступним їх фарбуванням органічним фарбником у вигляді водного розчину, причому високожиттєздатними вважаються популяції...
Спосіб оптимізації умов діапаузи видів роду trichogramma

Номер патенту: 22063
Опубліковано: 10.04.2007
Автор: Дрозда Валентин Федорович
МПК: A01K 67/04
Мітки: роду, умов, оптимізації, trichogramma, спосіб, видів, діапаузи
Формула / Реферат:
Спосіб оптимізації умов діапаузи видів роду Trichogramma, що включає спрямовану дію на різні стадії розвитку паразитів, який відрізняється тим, що в період розвитку трихограми на стадії личинки та на стадії передлялечки витримують температуру повітря +8...+11°С, при цьому імаго паразита підживлюють водним розчином 0,005-0,020 % абсцизової кислоти.
Спосіб переводу видів роду trichogramma на нового хазяїна

Номер патенту: 25002
Опубліковано: 25.07.2007
Автор: Дрозда Валентин Федорович
МПК: A01K 67/00
Мітки: нового, спосіб, переводу, trichogramma, видів, хазяїна, роду
Формула / Реферат:
Спосіб переводу видів роду Trichogramma на нового хазяїна, що включає їх вирощування в яйцях лускокрилих комах, який відрізняється тим, що самицям трихограми відразу після спаровування пропонують яйця нового хазяїна і одноразово згодовують їжу, до складу якої входить 3-5 %-ний водний розчин меду у суміші з нативним попередником рибонуклеїнової кислоти (РНК) у концентрації 0,02-0,003 %.
Спосіб контролю шкідливості популяцій видів роду ostrinia (lepidoptera, pyraustidae)

Номер патенту: 28182
Опубліковано: 26.11.2007
Автор: Дрозда Валентин Федорович
МПК: A01G 13/00, A01N 63/00
Мітки: спосіб, роду, видів, pyraustidae, контролю, популяцій, шкідливості, ostrinia, lepidoptera
Формула / Реферат:
Спосіб контролю шкідливості популяції видів роду Ostrinia (Lepidoptera, Pyraustidae), що включає використання трихограми, який відрізняється тим, що перед розселенням в агроценози трихограму спаровують, причому після спаровування заражають впродовж 1-1,5 години яйцями комахи-хазяїна у біолабораторії, крім того, перед розселенням трихограми та ізоляції її від яєць лабораторного хазяїна, імаго згодовують дієту у складі водного розчину...
Попередній патент: Пристрій для моніторингу стану сільськогосподарських угідь
Наступний патент: Робоче обладнання землерийно-транспортної машини
Випадковий патент: Фільтрувальний елемент трубчастого типу