Пентаспецифічне антитіло
Номер патенту: 103157
Опубліковано: 25.09.2013
Автори: Клегг Стефані Джейн, Льюїс Алан П., Йонак Зденка Людмила, Уайт Джон Р., Гермашевскі Фолкер, Елліс Джонатан Г., Добрзинські Ерік, Годіллот Алексіс Поль





















































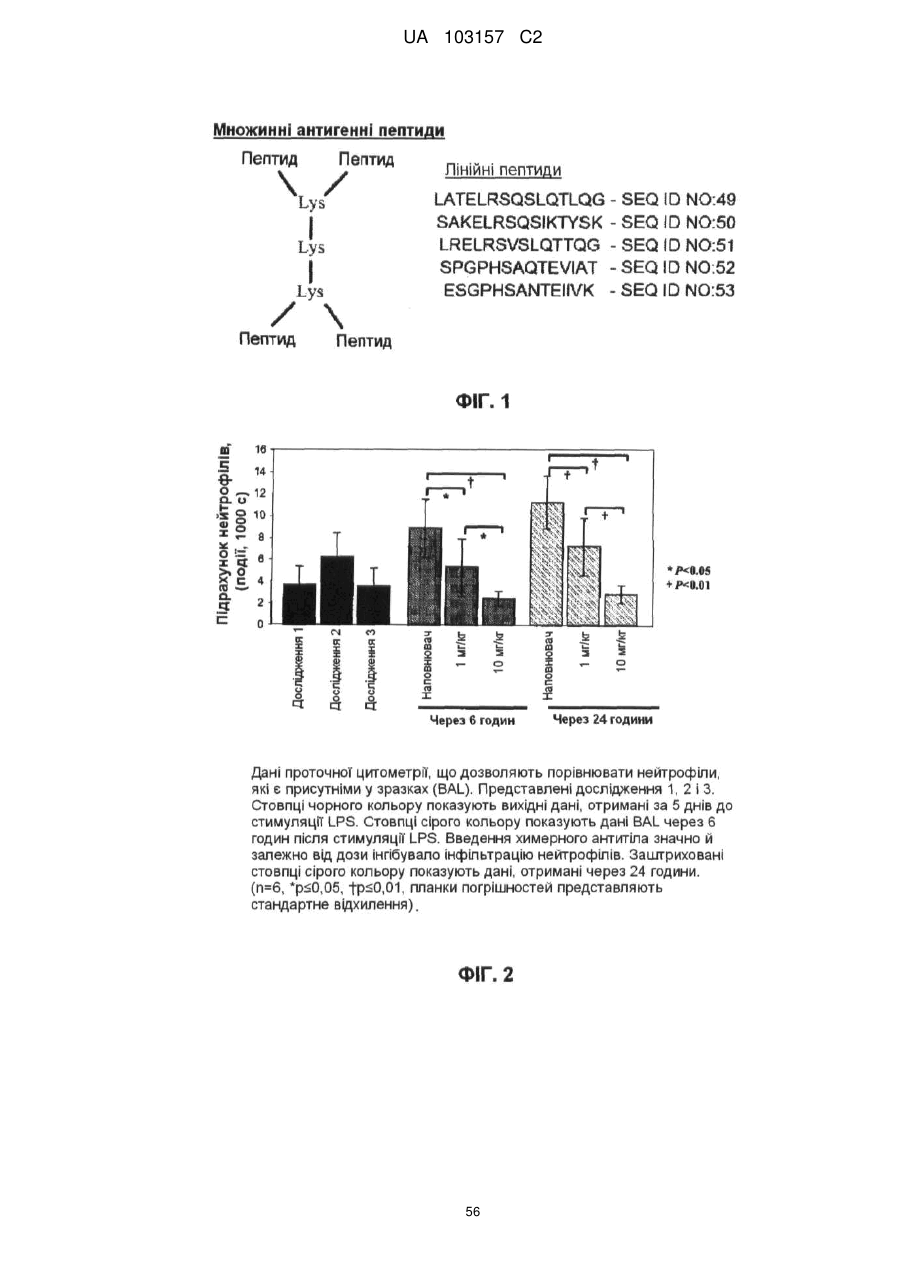
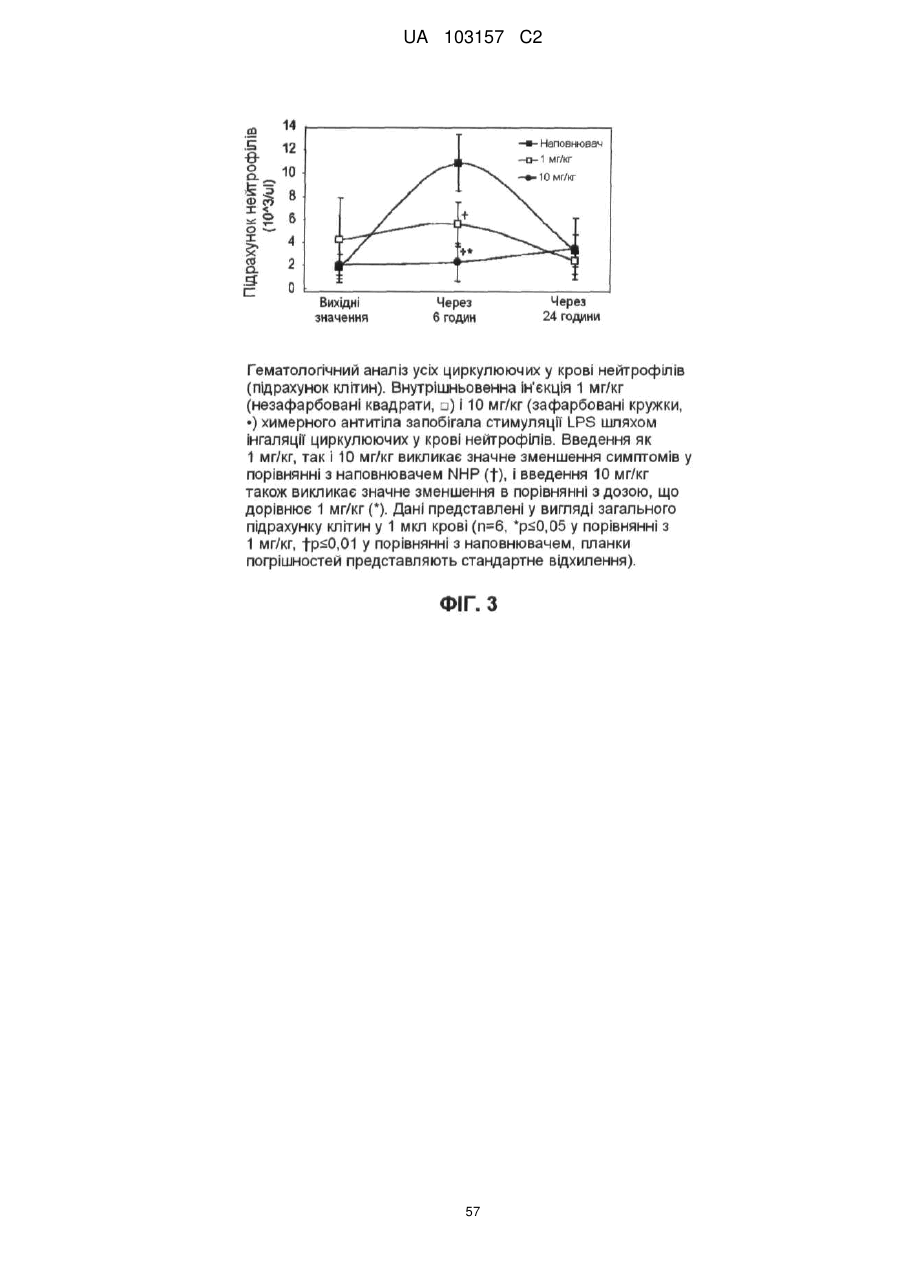
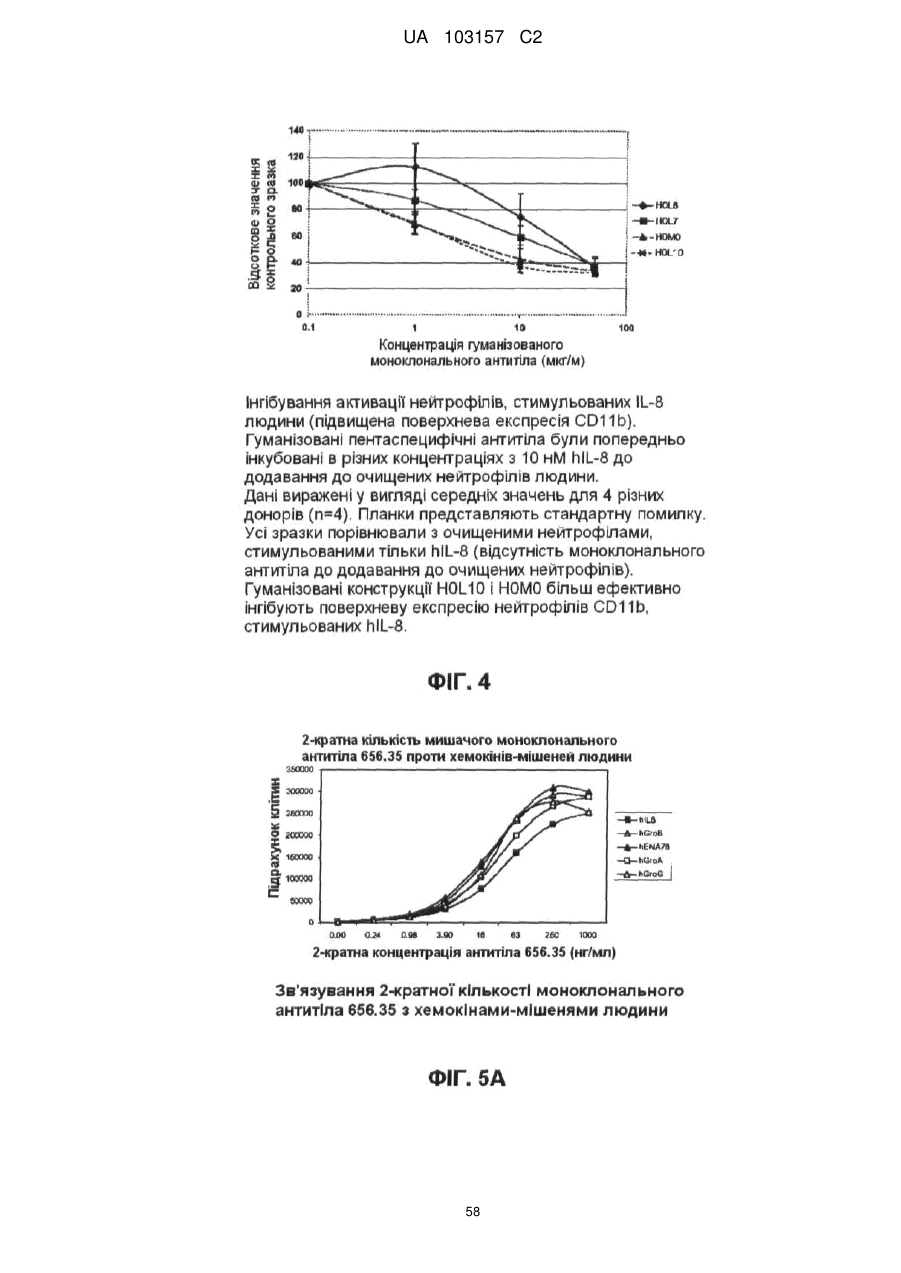

Формула / Реферат
1. Ізольована рекомбінантна еукаріотична або прокаріотична клітина, яка продукує антитіло, що містить варіабельні ділянки важкого та легкого ланцюгів, що містять амінокислотні послідовності CDR SEQ ID NOs: 13, 14 та 15; та SEQ ID NOs: 16, 17 та 18, відповідно.
2. Ізольоване антитіло, що містить варіабельні ділянки важкого та легкого ланцюгів, які містять амінокислотні послідовності CDR SEQ ID NOs:13, 14 та 15; та SEQ ID NOs:16, 17 та 18, відповідно.
3. Фармацевтична композиція, що містить антитіло за п. 2 та фармацевтично прийнятний носій.
4. Спосіб лікування або попередження COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, або псоріазу, відторгнення трансплантата, подагри, раку, гострого ураження легенів, гострого захворювання легенів, сепсису, ARDS, захворювання периферійних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми та COPD, муковісцидозу, дифузного панбронхіоліту, ураження, викликаного реперфузією, та/або ендометріозу, що включає введення ефективної кількості антитіла за п. 2.
5. Ізольоване антитіло за п. 2, що містить важкий та легкий ланцюги, кодовані нуклеотидними послідовностями, що включають послідовності SEQ ID NO:11 та SEQ ID NO:61, відповідно.
6. Ізольоване антитіло за п. 2, яке містить важкий та легкий ланцюги, що містять амінокислотні послідовності SEQ ID NO: 46 та SEQ ID NO:60, 62 або 64, відповідно.
7. Ізольоване антитіло, що містить важкий та легкий ланцюги, що містять амінокислотні послідовності SEO ID NO:46 та SEQ ID NO:62, відповідно.
8. Ізольована рекомбінантна еукаріотична або прокаріотична клітина-хазяїн, яка продукує антитіло, що має важкий або легкий ланцюг, що містить амінокислотну послідовність SEQ ID NO:46 та SEQ ID NO:62, відповідно.
9. Антитіло за п. 2, яке є гуманізованим.
Текст
Реферат: Винахід стосується антитіла, що має множинну специфічність до (або перехресно взаємодіє з) IL-8, Gro-альфа, Gro-бета, Gro-гамма і ENA-78 людини, та містить варіабельні ділянки важкого та легкого ланцюгів, що містять амінокислотні послідовності CDR SEQ ID NOs:13, 14 та 15; та SEQ ID NOs: 16, 17 та 18, відповідно. UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 Перехресне посилання на родинну заявку За даною заявкою заявляється пріоритет тимчасової заявки на патент США 60/912229, поданої 17 квітня 2007 р., і 61/044132, поданої 11 квітня 2008 р., що цілком включені в даний опис винаходу шляхом посилання. Галузь техніки, до якої належить винахід Дійсний винахід відноситься до антитіла, що володіє широкою специфічністю. Зокрема, антитіло за дійсним винаходом зв'язується (перехресно взаємодіє) з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини. Дійсний винахід відноситься також до способів лікування зазначеним антитілом захворювань або порушень, які характеризуються підвищеним або незбалансованим рівнем одного або декількох хемокінів, що включають IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, а саме, хронічного обструктивного захворювання легень (COPD), остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, респіраторного дистрес-синдрому в дорослих (ARDS), захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. Рівень техніки Опубліковані дані та звіти свідчать про те, що в цілому ряді захворювань мають місце підвищені рівні членів надсімейства ELRCXC хемокінів CXCL. Сімейство CXCL нараховує в цілому 16 членів. Як повідомлялося в науковій літературі, рівні хемокінів підвищені в цілому ряді запальних захворювань, включаючи COPD, у якому CXCL 1-3, 5 і 8 відомі також як Gro-α, -β, -γ (Haskill, S., et al. Proc. Natl. Acad. Sci., 1990; 87, 7732-7736), ENA-78 (Wang, D. and Richmond, A., Cytokine Reference. Oppenheim, J.J. and Feldman, M. ed., Academic Press, London, 1023-1027, Powerm C.A. et al. Gene., 1994: 151, 333-334) і IL-8 (Iizasa, H. and Matsushima, K., Cytokine Reference. Oppenheim, J.J. and Feldman, M. ed., Academic Press, London, 1061-1067, Matsushima, K. et el., J. Exp. Med. 1988: 167, 1883-1893) (Am. J. Respir. Crit. Саrе Med., 163: 349-355, 2001, Am. J. Respir. Crit. Care Med., 168:968-975, 2003, Thorax, 57: 590-595, 2002). Теоретично обґрунтовано, що тривала і підвищена експресія зазначених хемокінів може викликати розвиток таких захворювань, як COPD, остеоартрит, ревматоїдний артрит, ерозійний артрит, астма, атеросклероз, запальне захворювання кишечнику, псоріаз, відторгнення трансплантату, подагра, рак, гостре ураження легень, гостре захворювання легень, сепсис, ARDS, захворювання периферичних артерій, системний склероз, респіраторний дистрес-синдром у немовлят, загострення астми і COPD, муковісцидоз, дифузійний панбронхіоліт, ураження, викликане реперфузією, або ендометріоз. Зазначені хемокіни СХС, як відомо, стимулюють хемотаксис нейтрофілів у результаті залучення й активації рецепторів CXCR1 і/або СХС2. Таким чином, інгібування зазначених хемокінів може перешкоджати інфільтрації клітин запального інфільтрату в тканину легень і, отже, запобігати ураженню тканини. Метою дійсного винаходу є інгібування активації рецепторів CXCR1 і CXCR2 за допомогою антитіла, здатного зв'язуватися з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, тобто пентаспецифічного антитіла. Суть винаходу Дійсний винахід відноситься до антитіла (імуноглобуліну), яке володіє широкою специфічністю, що міститься в одному імуноглобуліні. Зокрема, антитіло за дійсним винаходом зв'язується (перехресно взаємодіє) з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини. Дійсний винахід відноситься також до способів лікування зазначеним антитілом захворювань або порушень, які характеризуються підвищеним або незбалансованим рівнем одного або декількох хемокінів, що включають IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, а саме, хронічного обструктивного захворювання легень (COPD), остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, респіраторного дистрес-синдрому в дорослих (ARDS), захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. Одним об'єктом дійсного винаходу є виділене антитіло, щоволодіє широкою специфічністю (або яке перехресно взаємодіє) до IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, тому антитіло за дійсним винаходом було визначене як пентаспецифічне (або пан-ELR) антитіло. Зазначене визначення антитіла включає антиген-зв'язувальну частину (або фрагмент) антитіла, що зв'язується з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини. Пентаспецифічне 1 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 антитіло за дійсним винаходом переважно є мишачим моноклональним, химерним, людським або гуманізованим антитілом. Щоб уникнути неясності слід зазначити, що пентаспецифічне антитіло за дійсним винаходом не обов'язково зв'язується тільки з антигенами IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, воно може також зв'язуватися з іншими родинними білками (такими як GCP-2 людини, або може навіть зв'язуватися з відмінними від людських ортологами IL-8, Gro-альфа, Gro-бета, Gro-гама, ENA-78 і GCP-2); іншими словами, пентаспецифічне антитіло за дійсним винаходом як мінімум зв'язується з IL-8, Gro-альфа, Groбета, Gro-гама і ENA-78 людини. Пентаспецифічне антитіло за дійсним винаходом переважно зв'язується з кожним з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини при значеннях константи рівноваги KD менше -7 -8 -9 10 М, більш переважно менше 10 М і ще більш переважно менше 10 М, що визначаються шляхом вимірювання поверхневого плазмонного резонансу. Вимірювання поверхневого плазмонного резонансу звичайно виконують відповідно до приведеного нижче опису. Один варіант здійснення дійсного винаходу відноситься до способу зменшення хемотаксису нейтрофілів шляхом інгібування активації рецепторів CXCR1 і CXCR2 у результаті нейтралізації IL-8, Gro-альфа, Gro-бета, Gro-гама, GCP-2 і ENA-78 людини пентаспецифічним антитілом за дійсним винаходом. Один варіант здійснення дійсного винаходу відноситься до способу зменшення хемотаксису нейтрофілів у пацієнта шляхом введення пентаспецифічного антитіла за дійсним винаходом. В одному варіанті здійснення винаходу пентаспецифічне антитіло зв'язується з епітопом KELRCQCIKTYSKP (SEQ ID NO:54) у IL-8 людини. В одному варіанті здійснення винаходу пентаспецифічне антитіло за дійсним винаходом одержують способом за протоколом RIMM (Kilpatrick, K.E., et al. Hybridoma. 1997: 16, 381), використовуючи суміш (коктейль) IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини разом з набором з п'яти множинних антигенних пептидів (МАР), у яких кожна ланка містить одну окрему послідовність з поліпептидів з ID NO:49-53. LATELRSQSLQTLQG SEQ ID NO:49 SAKELRSQSIKTYSK SEQ ID NO:50 LRELRSVSLQTTQG SEQ ID NO:51 SPGPHSAQTEVIAT SEQ ID NO:52 ESGPHSANTEIIVK SEQ ID NO:53 Не обмежуючись якою-небудь теорією, слід зазначити, що МАР виконують дві функції в протоколі імунізації. По-перше, МАР забезпечують селективне множинне представлення відомої амінокислотної послідовності-мішені імунній системі хазяїна. По-друге, викликають збільшення маси завдяки численним копіям даної послідовності, пов'язаним ядром, таким як лізин, але не обмежуючись ним, що підвищує імуногенність послідовності у порівнянні з окремими пептидами (Francis, J.P., et al., Immunology, 1991: 73; 249, Schott, M.E., et al., Cell. Immuno. 1996: 174: 199209, Tam, J.P. Proc. Natl. Acad. Sci. 1988: 85; 5409-5413). МАР, використовувані в дійсному винаході, складаються з численних копій консервативних послідовностей-мішеней (наприклад, SEQ ID NO:49-53), виявлених в ділянках ELRCXC і GPHCA і поруч із зазначеними ділянками хемокінів-мішеней. Типовий набір МАР показаний на фігурі 1. В одному варіанті здійснення винаходу пентаспецифічне антитіло за дійсним винаходом одержують способом, що включає наступні стадії: а. ін'єкування миші суміші IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини в повному ад'юванті Фрейнда (cFA); b. ін'єкування миші суміші IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини в неповному ад'юванті Фрейнда (iFA); с. ін'єкування миші суміші IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини і набору з п'яти множинних антигенних пептидів (МАР), у яких кожна ланка містить одну окрему послідовність з поліпептидів з ID NO:49-53, у неповному ад'юванті Фрейнда; d. виділення у миші В-клітин; е. злиття В-клітин з мієломними клітинами з утворенням іморталізованих клітин гібридоми, що секретують необхідне пентаспецифічне антитіло; і f. виділення пентаспецифічного антитіла із супернатанту культури гібридоми. За бажанням, між стадіями с і d можна необов'язково ін'єкувати миші суміш IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини і набір МАР, що включають амінокислотні послідовності SEQ ID NO:49-53, у фізіологічному розчині з фосфатним буфером (PBS). В іншому варіанті здійснення винаходу пентаспецифічне антитіло за дійсним винаходом одержують способом, що включає наступні стадії: 2 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 а. ін'єкування миші набору з п'яти множинних антигенних пептидів (МАР), у яких кожна ланка містить одну окрему послідовність з поліпептидів з ID NO:49-53 (далі визначається також як набір МАР) у повному ад'юванті Фрейнда; b. ін'єкування миші набору МАР у неповному ад'юванті Фрейнда; с. ін'єкування миші суміші всіх IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини і набору МАР у неповному ад'юванті Фрейнда; d. виділення у миші В-клітин; е. злиття В-клітин з мієломними клітинами з утворенням іморталізованих клітин гібридоми, що секретують необхідне пентаспецифічне антитіло; і f. виділення пентаспецифічного антитіла із супернатанту культури гібридоми. За бажанням, між стадіями с і d можна необов'язково ін'єкувати миші суміш IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини і набір МАР, що включають SEQ ID NO:49-53, у фізіологічному розчині з фосфатним буфером (PBS). Інший варіант здійснення дійсного винаходу відноситься до пентаспецифічного антитіла, отриманого вищевказаним способом. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що кодуються нуклеотидними послідовностями, які включають, відповідно, послідовності SEQ ID NO:1 і SEQ ID NO:3 або консервативні модифікації зазначених послідовностей. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, шо кодуються нуклеотидними послідовностями, які включають, відповідно, послідовності SEQ ID NO:5 і SEQ ID NO:7 або консервативні модифікації зазначених послідовностей. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельну ділянку важкого ланцюга, що кодується нуклеотидною послідовністю, яка включає послідовність SEQ ID NO:9 або консервативні модифікації зазначеної послідовності. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають амінокислотні послідовності SEQ ID NO:2 і SEQ ID NO:4 або консервативні модифікації зазначених послідовностей. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають поліпептиди, які щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:2 і SEQ ID NO:4. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають, відповідно, амінокислотні послідовності SEQ ID NO:6 і SEQ ID NO:8 або консервативні модифікації зазначених послідовностей. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають поліпептиди, які щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:6 і SEQ ID NO:8. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають, відповідно, амінокислотні послідовності SEQ ID NO:10 і 12 або консервативні модифікації зазначених послідовностей. В одному варіанті здійснення винаходу пентаспецифічне антитіло має варіабельні ділянки важкого і легкого ланцюгів, що включають поліпептидні послідовності, які щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:10 і 12. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше одну варіабельну ділянку, обрану з (і) амінокислоти SEQ ID NO:2, 4, 6, 8, 10 або 12; або (ii) амінокислотної послідовності, яка щонайменше на 90 %, 95 %, 98 % або 99 % ідентична до однієї з амінокислотних послідовностей, зазначених у пункті (і). В одному варіанті здійснення винаходу пентаспецифічне антитіло включає послідовності CDR SEQ ID NO:13, 14, 15, 16, 17 і 18; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей SEQ ID NO:13, 14, 15, 16, 17 і 18. Один варіант здійснення дійсного винаходу відноситься до гібридоми або трансфектоми, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:13, 14, 15, 16, 17 і 18. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:13, 14, 15, 16, 17 і 18. 3 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше одну послідовність CDR, обрану з (і) SEQ ID NO:13, 14, 15, 16, 17 або 18; або (ii) консервативної модифікації послідовностей, зазначених у пункті (і). В одному варіанті здійснення винаходу пентаспецифічне антитіло включає поліпептид SEQ ID NO:15. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше чотири послідовності CDR, обрані з групи, що включає SEQ ID NO:13, 14, 15, 16, 17 і 18; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей SEQ ID NO:13, 14, 15, 16, 17 і 18. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає варіабельні ділянки важкого і легкого ланцюгів, що містять, відповідно, амінокислотні послідовності CDR SEQ ID NO:13, 14 і 15 і SEQ ID NO:16, 17 і 18. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає послідовності CDR SEQ ID NO:19, 20, 21, 22, 23 і 24; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей, зазначених у SEQ ID NO:19, 20, 21, 22, 23 і 24. Один варіант здійснення дійсного винаходу відноситься до гібридоми або трансфектоми, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:19, 20, 21, 22, 23 і 24. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:19, 20, 21, 22, 23 і 24. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше одну послідовність CDR, обрану з (і) SEQ ID NO:19, 20, 21, 22, 23 або 24; або (ii) консервативної модифікації послідовностей, зазначених у пункті (і). В одному варіанті здійснення винаходу пентаспецифічне антитіло включає поліпептид SEQ ID NO:21. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше чотири послідовності CDR, обрані з групи, що включає SEQ ID NO:19, 20, 21, 22, 23 і 24; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей, зазначених у SEQ ID NO:19, 20, 21, 22, 23 і 24. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає варіабельні ділянки важкого і легкого ланцюгів, що містять, відповідно, амінокислотні послідовності CDR SEQ ID NO:19, 20 і 21 і SEQ ID NO:22, 23 і 24. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає послідовності CDR SEQ ID NO:25, 26, 27, 28, 29 і 30; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей, зазначених у SEQ ID NO: 25, 26, 27, 28, 29 і 30. Один варіант здійснення дійсного винаходу відноситься до гібридоми або трансфектоми, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:25, 26, 27, 28, 29 і 30. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини, що продукує пентаспецифічне антитіло, яке включає послідовності CDR SEQ ID NO:25, 26, 27, 28, 29 і 30. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше одну послідовність CDR, обрану з (і) SEQ ID NO:25, 26, 27, 28, 29 або 30; або (ii) консервативної модифікації послідовностей, зазначених у пункті (і). В одному варіанті здійснення винаходу пентаспецифічне антитіло включає поліпептид SEQ ID NO:27. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає щонайменше чотири послідовності CDR, обрані з групи, що включає SEQ ID NO:25, 26, 27, 28, 29 і 30; або одна або кілька послідовностей CDR можуть бути консервативними модифікаціями послідовностей, зазначених у SEQ ID NO:25, 26, 27, 28, 29 і 30. В одному варіанті здійснення винаходу пентаспецифічне антитіло включає варіабельні ділянки важкого і легкого ланцюгів, які містять, відповідно, амінокислотні послідовності CDR SEQ ID NO:25, 26 і 27 і SEQ ID NO:28, 29 і 30. Один варіант здійснення дійсного винаходу відноситься до гібридоми, що продукує моноклональне антитіло, яке має варіабельну ділянку важкого або легкого ланцюга, що кодується нуклеотидними послідовностями, які включають, відповідно, нуклеотидні послідовності SEQ ID NO:1, 5 або 9, або SEQ ID NO:3 або 7. 4 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 Один варіант здійснення дійсного винаходу відноситься до гібридоми, що продукує моноклональне антитіло, яке має варіабельну ділянку важкого або легкого ланцюга, що включає, відповідно, амінокислотні послідовності SEQ ID NO:2, 6 або 10, або SEQ ID NO:4, 8 або 12 і консервативні модифікації зазначених послідовностей. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини-хазяїна, що продукує пентаспецифічне антитіло, яке має варіабельну ділянку важкого або легкого ланцюга, що включає, відповідно, амінокислотні послідовності SEQ ID NO:2, 6 або 10, або SEQ ID NO:4, 8 або 12 і консервативні модифікації зазначених послідовностей. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидні послідовності, які кодують варіабельну ділянку важкого або легкого ланцюга пентаспецифічного антитіла, яка містить, відповідно, послідовності CDR SEQ ID NO:13, 14 і 15 або SEQ ID NO:16, 17 і 18. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидну послідовність, яка кодує послідовність CDR пентаспецифічного антитіла, обрану з SEQ ID NO:13, 14, 15, 16, 17 або 18. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидні послідовності, які кодують щонайменше чотири послідовності CDR пентаспецифічного антитіла, обрані з групи, що включає SEQ ID NO:13, 14, 15, 16, 17 і 18. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності SEQ ID NO:31, 32 або 33. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності SEQ ID NO:34, 35 і 36. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає щонайменше чотири полінуклеотидні послідовності, обрані з групи, що включає SEQ ID NO:31, 32, 33, 34, 35 і 36. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидні послідовності, які кодують варіабельну ділянку важкого або легкого ланцюга пентаспецифічного антитіла, що містить, відповідно, послідовності CDR SEQ ID NO:19, 20 і 21 або 22, 23 і 24. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидну послідовність, яка кодує послідовність CDR пентаспецифічного антитіла, обрану з SEQ ID NO:19, 20, 21, 22, 23 або 24. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидні послідовності, які кодують щонайменше чотири послідовності CDR пентаспецифічного антитіла, обрані з групи, що включає SEQ ID NO:19, 20, 21, 22, 23 і 24. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності SEQ ID NO:37, 38 і 39. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності SEQ ID NO:40, 41 і 42. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає щонайменше чотири полінуклеотидні послідовності, обрані з групи, що включає SEQ ID NO:37, 38, 39, 40, 41 і 42. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидні послідовності, які кодують варіабельну ділянку важкого ланцюга пентаспецифічного антитіла, що містить послідовності CDR SEQ ID NO:25, 26 і 27. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає нуклеотидну послідовність, яка кодує послідовність CDR пентаспецифічного антитіла, обрану з SEQ ID NO:25, 26 або 27. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності SEQ ID NO:SEQ ID NO:43, 44 і 45. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які кодуються нуклеотидними послідовностями, що включають, відповідно, послідовності SEQ ID NO:11 і SEQ ID NO:47, 59, 61 або 63. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які кодуються нуклеотидними послідовностями, що включають послідовності, які щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до послідовностей SEQ ID NO:11 і SEQ ID NO:47, 59, 61 або 63. 5 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:48. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:60. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:62. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:64. В одному варіанті здійснення винаходу пентаспецифічне антитіло має важкі і легкі ланцюги, які включають поліпептиди, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини-хазяїна, що продукує пентаспецифічне антитіло, яке має важкий або легкий ланцюг, який включає, відповідно, амінокислотну послідовність SEQ ID NO:46 або SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини-хазяїна, що продукує пентаспецифічне антитіло, яке має важкі і легкі ланцюги, які включають, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини-хазяїна, що продукує пентаспецифічне антитіло, яке має важкий або легкий ланцюг, який включає амінокислотну послідовність, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентична, відповідно, до амінокислотної послідовності SEQ ID NO:46 або SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до рекомбінантної еукаріотичної або прокаріотичної клітини-хазяїна, що продукує пентаспецифічне антитіло, яке має важкі і легкі ланцюги, які включають амінокислотні послідовності, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності, які містять, відповідно, послідовності SEQ ID NO:11 і SEQ ID NO:47, 59, 61 або 63. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності, які містять послідовності, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до послідовностей SEQ ID NO:11 і SEQ ID NO:47, 59, 61 або 63. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидну послідовність, яка містить послідовність SEQ ID NO:11, 47, 59, 61 або 63. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидну послідовність, яка містить послідовність, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентична, відповідно, до послідовності SEQ ID NO:11, 47, 59, 61 або 63. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності, які кодують пентаспецифічне антитіло, яке включає важкі і легкі ланцюги, що містять, відповідно, амінокислотні послідовності SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидні послідовності, які кодують пентаспецифічне антитіло, що включає важкі і легкі ланцюги, які містять амінокислотні послідовності, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентичні, відповідно, до амінокислотних послідовностей SEQ ID NO:46 і SEQ ID NO:48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидну послідовність, яка кодує поліпептид, що містить амінокислотну послідовність SEQ ID NO:46, 48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до експресуючого вектора, що включає полінуклеотидну послідовність, яка кодує поліпептид, що містить амінокислотну 6 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовність, що щонайменше на 90 %, 95 %, 98 % або 99 % ідентична до амінокислотних послідовностей SEQ ID NO:46, 48, 60, 62 або 64. Один варіант здійснення дійсного винаходу відноситься до способу продукування пентаспецифічного антитіла (імуноглобуліну) в одиничній клітині-хазяїні, що включає наступні стадії: (і) трансформацію зазначеної одиничної клітини-хазяїна першою послідовністю ДНК, що кодує важкий ланцюг, який включає поліпептид SEQ ID NO:46; і другою послідовністю ДНК, що кодує легкий ланцюг, який включає поліпептид SEQ ID NO:48, 60, 62 або 64; і (ii) експресію зазначеної першої послідовності ДНК і зазначеної другої послідовності ДНК, у результаті якої зазначені важкі і легкі ланцюги імуноглобуліну одержують у вигляді окремих молекул у зазначеній трансформованій одиничній клітині-хазяїні; причому даний спосіб може бути виконаний таким чином, що зазначені перша і друга послідовності ДНК присутні в різних векторах або зазначені перша і друга послідовності ДНК присутні в одному векторі. Один варіант здійснення дійсного винаходу відноситься до способу продукування пентаспецифічного антитіла (імуноглобуліну) в одиничній клітині-хазяїні, що включає наступні стадії: (і) трансформацію зазначеної одиничної клітини-хазяїна першою послідовністю ДНК, що кодує щонайменше варіабельну ділянку важкого ланцюга імуноглобуліну, що включає ділянки CDR SEQ ID NO:13, 14 і 15; і другою послідовністю ДНК, що кодує щонайменше варіабельну ділянку легкого ланцюга імуноглобуліну, що включає ділянки CDR SEQ ID NO:16, 17 і 18; і (ii) експресію зазначеної першої послідовності ДНК і зазначеної другої послідовності ДНК, у результаті якої зазначені важкі і легкі ланцюги імуноглобуліну одержують у вигляді окремих молекул в зазначеній трансформованій одиничній клітині-хазяїні; причому даний спосіб може бути виконаний таким чином, що зазначені перша і друга послідовності ДНК присутні в різних векторах або зазначені перша і друга послідовності ДНК присутні в одному векторі. Один варіант здійснення дійсного винаходу відноситься до способу продукування пентаспецифічного антитіла (імуноглобуліну) в одиничній клітині-хазяїні, що включає наступні стадії: (і) трансформацію зазначеної одиничної клітини-хазяїна першою послідовністю ДНК, що кодує щонайменше варіабельну ділянку важкого ланцюга імуноглобуліну, що включає ділянки CDR SEQ ID NO:19, 20 і 21; і другою послідовністю ДНК, що кодує щонайменше варіабельну ділянку легкого ланцюга імуноглобуліну, що включає ділянки CDR SEQ ID NO:22, 23 і 24; і (ii) експресію зазначеної першої послідовності ДНК і зазначеної другої послідовності ДНК, у результаті якої зазначені важкі і легкі ланцюги імуноглобуліну одержують у вигляді окремих молекул в зазначеній трансформованій одиничній клітині-хазяїні; причому даний спосіб може бути виконаний таким чином, що зазначені перша і друга послідовності ДНК присутні в різних векторах або зазначені перша і друга послідовності ДНК присутні в одному векторі. Один варіант здійснення дійсного винаходу відноситься до антитіла, яке повністю або частково блокує зв'язування будь-якого з вищевказаних пентаспецифічних антитіл з IL-8, Groальфа, Gro-бета, Gro-гама, ENA-78 і GCP-2 людини при виконанні імуноаналізу, такого як аналіз ELISA. В одному варіанті здійснення винаходу часткове блокування відбувається тоді, коли антитіло блокує зв'язування пентаспецифічного антитіла більш ніж на 10 %, 20 %, 40 % або 50 %. Один варіант здійснення дійсного винаходу відноситься до антитіла, яке конкурує за зв'язування з будь-яким з вищевказаних пентаспецифічних антитіл з IL-8, Gro-альфа, Gro-бета, Gro-гама, ENA-78 і GCP-2 людини. Один варіант здійснення дійсного винаходу відноситься до антитіла, яке повністю або частково блокує зв'язування будь-якого з вищевказаних пентаспецифічних антитіл з епітопом KELRCQCIKTYSKP (SEQ ID NO:54) у IL-8 людини при виконанні імуноаналізу, такого як аналіз ELISA. В одному варіанті здійснення винаходу часткове блокування відбувається тоді, коли антитіло блокує зв'язування пентаспецифічного антитіла більш ніж на 10 %, 20 %, 40 % або 50 %. Один варіант здійснення дійсного винаходу відноситься до антитіла, яке конкурує з будьяким з вищевказаних пентаспецифічних антитіл за зв'язування з епітопом KELRCQCIKTYSKP (SEQ ID NO:54) у IL-8 людини. Один варіант здійснення дійсного винаходу відноситься до композиції, що включає вищевказане пентаспецифічне антитіло і фармацевтично прийнятний носій. 7 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 Один варіант здійснення дійсного винаходу відноситься до способу лікування або запобігання у ссавця COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, ARDS, захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу, що включає введення зазначеному ссавцю ефективної кількості вищевказаного пентаспецифічного антитіла. Один варіант здійснення дійсного винаходу відноситься до вищевказаного пентаспецифічного антитіла, призначеного для використання при лікуванні захворювань або порушень, що характеризуються підвищеним або незбалансованим рівнем одного або декількох хемокінів, що включають IL-8, Gro-альфа, Gro-бета, Gro-гама, GCP-2 і ENA-78 людини, а саме, COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, ARDS, захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, або ендометріозу. Одним об'єктом дійсного винаходу є вищевказане пентаспецифічне антитіло, призначене для запобігання і/або лікування у ссавця COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, ARDS, захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. Одним об'єктом дійсного винаходу є застосування вищевказаного пентаспецифічного антитіла для одержання лікарського засобу, використовуваного для запобігання і/або лікування у ссавця COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, ARDS, захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. Одним об'єктом дійсного винаходу є застосування вищевказаного пентаспецифічного антитіла для одержання лікарського засобу, призначеного для запобігання і/або лікування у ссавця COPD, остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, ARDS, захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. В одному варіанті здійснення винаходу вищевказаним ссавцем є людина. Опис креслень На фіг. 1 показаний типовий набір МАР, використовуваний для створення пентаспецифічного антитіла. Щоб уникнути неясності зображені п'ять пептидних ланок МАР. Кожна ланка містить одну ідентичну амінокислотну послідовність, обрану з лінійних пептидів SEQ ID NO:49-53. На фіг. 2 приведені дані проточної цитометрії, що дозволяють порівнювати нейтрофіли, які є присутніми у зразках бронхоальвеолярного лаважа (BAL). Представлені дослідження 1, 2 і 3. Стовпчики чорного кольору показують вихідні дані, отримані за 5 днів до стимуляції LPS. Стовпчики сірого кольору показують дані BAL, отримані через 6 годин після стимуляції LPS. Введення химерного антитіла (HcLc) значно й залежно від дози інгібувало інфільтрацію нейтрофілів. Заштриховані стовпчики сірого кольору показують дані, отримані через 24 години. (n=6, *р≤0,05, †р≤0,01, планки погрішностей представляють стандартне відхилення). На фіг. 3 показаний гематологічний аналіз усіх циркулюючих у крові нейтрофілів (підрахунок клітин). Внутрішньовенна ін'єкція 1 мг/кг (незафарбовані квадрати, □) і 10 мг/кг (зафарбовані кружки, ●) химерного антитіла (HcLc) запобігала стимуляції LPS циркулюючих у крові нейтрофілів шляхом інгаляції. Введення як 1 мг/кг, так і 10 мг/кг викликає значне зменшення симптомів у порівнянні з наповнювачем NHP (†), і введення 10 мг/кг також викликає значне зменшення в порівнянні з дозою, що дорівнює 1 мг/кг (*). Дані представлені у вигляді загального 8 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 підрахунку клітин у 1 мкл крові (n=6, *р≤0,05 у порівнянні з 1 мг/кг, †р≤0,01 у порівнянні з наповнювачем, планки погрішностей представляють стандартне відхилення). На фіг. 4 показане інгібування активації нейтрофілів, стимульованих IL-8 людини (підвищена поверхнева експресія CD11b). Гуманізовані пентаспецифічні антитіла попередньо інкубували в різних концентраціях з 10 нм hIL-8 до додавання до очищених нейтрофілів людини. Дані виражені у вигляді середніх значень для 4 різних донорів (n=4). Планки представляють стандартну помилку. Усі зразки порівнювали з очищеними нейтрофілами, стимульованими тільки hIL-8 (відсутність моноклонального антитіла до додавання до очищених нейтрофілів). Гуманізовані конструкції H0L10 і H0M0 більш ефективно інгібують поверхневу експресію CD11b, стимульовану hIL-8. На фіг. 5А показане зв'язування моноклонального антитіла 656.35 з хемокінами-мішенями людини. На фіг. 5В показане зв'язування химерного моноклонального антитіла (HcLc) з хемокінамимішенями людини. На фіг. 5С показане зв'язування гуманізованих моноклональних антитіл з хемокінамимішенями людини. Докладний опис винаходу Термін "виділене пентаспецифічне антитіло" або просте "пентаспецифічне антитіло" у значенні, використаному в дійсному описі винаходу, означає антитіло, що зв'язується і, отже, перехресно взаємодіє з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, власне кажучи не містить інших антитіл, що володіють іншою антигенною специфічністю, і має моносклад. Щоб уникнути неясності слід зазначити, що пентаспецифічне антитіло за дійсним винаходом не обов'язково зв'язується тільки з антигенами IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, воно може також зв'язуватися з іншими родинними білками, такими як GCP-2 людини, або навіть може зв'язуватися з відмінними від людських ортологами IL-8, Gro-альфа, Gro-бета, Gro-гама, ENA-78 і GCP-2; іншими словами, пентаспецифічне антитіло за дійсним винаходом як мінімум зв'язується з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини. Крім того, виділене пентаспецифічне антитіло власне кажучи не містить інших клітинних і/або хімічних речовин. Пентаспецифічне антитіло в значенні, використаному в даному контексті, включає також антиген-зв'язувальні фрагменти і/або похідні, що володіють характеристиками зв'язування "батьківського" пентаспецифічного антитіла. Пентаспецифічне антитіло за дійсним винаходом може включати два ідентичні важкі ланцюги і два ідентичні легкі ланцюги, які утворюють типову білатеральну симетричну молекулу імуноглобуліну, що складається з двох гетеродимерів, кожний з яких включає важкий ланцюг і легкий ланцюг. Таким чином, пентаспецифічне антитіло за дійсним винаходом може включати дві копії одного антиген-зв'язувального домену, утвореного в результаті асоціації одного важкого ланцюга з одним легким ланцюгом. Пентаспецифічне антитіло за дійсним винаходом, незважаючи на наявність зв'язувального домену тільки одного типу, може зв'язуватися і, отже, взаємодіяти з IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини. У використовуваному в описі значенні термін "антитіло" означає також "імуноглобулін". У використовуваному в описі значенні термін "антитіло, що перехресно взаємодіє з" означає, що зазначене антитіло зв'язується не тільки з одним антигеном, але і також з іншими антигенами. Термін "нейтралізація" у використовуваному в описі значенні означає часткове або повне інгібування щонайменше однієї біологічної активності IL-8, Gro-альфа, Gro-бета, Gro-гама, GCP2 або ENA-78 людини. Наприклад, однієї з біологічних активностей IL-8, Gro-альфа, Gro-бета, Gro-гама, GCP-2 або ENA-78 людини є їхня здатність індукувати хемотаксис нейтрофілів. Одним способом вимірювання кінетики зв'язування антитіла є вимірювання поверхневого плазмонного резонансу. Термін "поверхневий плазмонний резонанс" у використовуваному в описі значенні означає оптичне явище, яке дозволяє аналізувати біоспецифічні взаємодії в реальному часі шляхом виявлення змін у концентрації білків у біосенсорній матриці, наприклад, за допомогою системи BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, N.J.). Для ознайомлення з більш докладним описом див. публікації Jonsson, U., et al. (1993) Ann. Biol. Clin. 51:19-26; Jonsson, U., et al. (1991) Biotechniques 11:620-627; Johnsson, B., et al. (1995) J. Mol. Recognit. 8:125-131; і Johnnson, B., et al. (1991) Anal. Biochem. 198:268-277. Термін "епітоп" означає детермінанту білка, здатну специфічно зв'язуватися з антитілом. Епітопи звичайно складаються з хімічно активних поверхневих груп молекул, таких як бічні ланцюги амінокислот або цукрів, і мають специфічну тривимірну структуру, а також характерні заряди. 9 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 Термін "моноклональне антитіло" або mAb (на відміну від поліклонального антитіла) у використовуваному в описі значенні означає молекули антитіл, що мають однакову молекулярну сполуку. Наприклад, моноклональне антитіло, виділене у миші (мишаче моноклональне антитіло), може бути отримане методом утворення гібридом, таким як стандартна методика утворення гібридом Колера і Мілстейна (Kohler and Milstein). Антитіло-продукуючі клітини можуть бути отримані у суб'єкта і використані для одержання моноклональних антитіл стандартними методами, такими як метод гібридом, спочатку описаний Колером і Мілстейном (1975, Nature 256:495-497) (див. також публікації Brown et al. (1981) J. Immunol. 127:539-46; Brown et al. (1980) J. Biol. Сhем. 255:4980-83; Yeh et al. (1976) PNAS 76:2927-31; і Yeh et al. (1982) Int. J. Cancer 29:269-75). Методика одержання гібридом моноклонального антитіла добре відома (див. публікації R.H. Kenneth, in Monoclonal Antibodies: A New Dimension In Biological Analyses, Plenum Publishing Corp., New York, N.Y. (1980); E.A. Lerner (1981) Yale J. Biol. Med., 54:387-402; M.L. Gefter et al. (1977) Somatic Cell Genet., 3:231-36). Термін "трансфектома" у використовуваному в описі значенні означає рекомбінантну еукаріотичну клітину-хазяїна, що експресує антитіло, таку як клітини СНО, клітини NS/0, клітини НЕК293, рослинні клітини або гриби, включаючи дріжджові клітини. У використовуваному в описі значенні термін "специфічне" зв'язування означає зв'язування антитіла із заздалегідь визначеним антигеном. Антитіло звичайно зв'язується при значеннях -7 константи рівноваги KD, рівних приблизно 110 М або менше, і зв'язується із заздалегідь визначеним антигеном зі спорідненістю, що відповідає значенню KD, яке щонайменше на два порядки величини менше спорідненості зв'язування з неспецифічним антигеном (наприклад, BSA, казеїн), відмінним від заздалегідь визначеного антигену або близько родинного антигену. Фрази "антитіло, що впізнає антиген" і "антитіло, специфічне до антигену" мають взаємозамінні значення з терміном "антитіло, що специфічно зв'язується з антигеном". -1 Термін "kd" (сек ) у використовуваному в описі значенні означає константу швидкості дисоціації визначеної взаємодії антитіло-антиген. -1 Термін "ka" (Мсек ) у використовуваному в описі значенні означає константу швидкості асоціації визначеної взаємодії антитіло-антиген. Термін "KD" (M) у використовуваному в описі значенні означає константу рівноваги визначеної взаємодії антитіло-антиген, що одержують у результаті розподілу kd на ka. Термін "консервативні модифікації послідовностей" для модифікацій нуклеотидних і амінокислотних послідовностей означає зміни, які не чинять істотного впливу або не змінюють характеристики зв'язування антитіла, що кодується даною нуклеотидною послідовністю або містить таку амінокислотну послідовність. Такі консервативні модифікації послідовностей включають заміни, додавання і делеції нуклеотидів і амінокислот. Модифікації можуть бути введені в послідовності стандартними методами, відомими в даній галузі, такими як сайтнаправлений мутагенез, ПЦР-опосередкований мутагенез. Консервативні заміни амінокислот включають такі заміни, у яких амінокислотний залишок заміняється іншим амінокислотним залишком, що має подібний бічний ланцюг. У даній галузі відомі сімейства амінокислотних залишків, що мають подібні бічні ланцюги. Зазначені сімейства включають амінокислоти з основними бічними ланцюгами (наприклад, лізин, аргінін, гістидин), кислотними бічними ланцюгами (наприклад, аспарагінова кислота, глутамінова кислота), незарядженими полярними бічними ланцюгами (наприклад, гліцин, аспарагін, глутамін, серин, треонін, тирозин, цистеїн, триптофан), неполярними бічними ланцюгами (наприклад, аланін, валін, лейцин, ізолейцин, пролін, фенілаланін, метіонін), бета-розгалуженими бічними ланцюгами (наприклад, треонін, валін, ізолейцин) і ароматичними бічними ланцюгами (наприклад, тирозин, фенілаланін, триптофан, гістидин). Таким чином, прогнозований замінний амінокислотний залишок в антитілі, що має розглянуту послідовність, переважно заміняють іншим амінокислотним залишком із сімейства з таким самим бічним ланцюгом. Таким чином, одним об'єктом дійсного винаходу є пентаспецифічне антитіло за дійсним винаходом, що включає всі консервативні модифікації розглянутих амінокислотних послідовностей. В обсяг дійсного винаходу входять також "похідні" описаних амінокислотних послідовностей, у яких один або кілька амінокислотних залишків є похідними залишками, отриманими, наприклад, шляхом ацилювання або глікозилювання, що не чинять істотного впливу і не змінюють характеристики зв'язування антитіла, що містить зазначені амінокислотні послідовності. Термін "власне кажучи ідентичні" стосовно нуклеїнових кислот означає, що дві нуклеїнові кислоти або їх послідовності є ідентичними при оптимальному сполученні і в порівнянні з відповідними інсерціями або делеціями нуклеотидів щонайменше приблизно в 80 % нуклеотидів, звичайно щонайменше приблизно в 90 %-95 % і більш переважно щонайменше 10 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 приблизно в 98 %-99,5 % нуклеотидів. Альтернативно, власне кажучи ідентичними нуклеїнові кислоти є тоді, коли сегменти гібридизують в умовах селективної гібридизації з комплементом ланцюга. Термін "ідентичний" стосовно до нуклеотидних і амінокислотних послідовностей означає ступінь ідентичності двох нуклеотидних або амінокислотних послідовностей при оптимальному сполученні і порівнянні з відповідними інсерціями або делеціями. Альтернативно, власне кажучи ідентичними послідовності є тоді, коли сегменти ДНК гібридизують в умовах селективної гібридизації з комплементом ланцюга. Процентне значення ідентичності двох послідовностей є функцією числа ідентичних положень у зазначених послідовностях (тобто % ідентичності = число ідентичних положень/загальне число положень, помножене на 100) з урахуванням числа розривів і довжини кожного розриву, що повинні бути введені для оптимального порівняння двох послідовностей. Порівняння послідовностей і визначення процентного значення ідентичності двох послідовностей може бути виконане за допомогою математичного алгоритму, описаного в приведених нижче необмежуючих прикладах. Процентне значення ідентичності двох нуклеотидних або поліпептидних послідовностей може бути визначене за допомогою програми GAP у пакеті програмного забезпечення GCG (представленого на сайті http://www.gcg.com) з використанням матриці NWSgapdna.CMP, вагового коефіцієнта розриву, рівного 40, 50, 60, 70 або 80, і вагового коефіцієнта довжини розривів, рівного 1, 2, 3, 4, 5 або 6. Процентне значення ідентичності двох нуклеотидних або амінокислотних послідовностей можна визначити, використовуючи алгоритм Е. Мейерса і В. Міллера (E. Meyers and W. Miller, Comput. Appl. Biosci., 4:11-17 (1988)) який включений у програму ALIGN (версія 2.0), таблицю вагових коефіцієнтів залишків РАМ120, штраф за довжину розриву, що дорівнює 12, і штраф за розрив, що дорівнює 4. Крім того, процентне значення ідентичності двох амінокислотних послідовностей можна визначити, використовуючи алгоритм Нідлмана і Ванша (Needleman and Wunsch, J. Mol. Biol. 48:444-453 (1970)), що включений у програму GAP пакета програмного забезпечення GCG (представлений на сайті http://www.gcg.com), матрицю Blossum 62 або матрицю РАМ250, ваговий коефіцієнт розриву, що дорівнює 16, 14, 12, 10, 8, 6 або 4, і ваговий коефіцієнт довжини розриву, що дорівнює 1, 2, 3, 4, 5 або 6. Нуклеїнова кислота є "функціонально зв'язаною" при перебуванні у функціональній взаємодії з послідовністю іншої нуклеїнової кислоти. Наприклад, промотор або енхансер функціонально зв'язаний з кодуючою послідовністю, якщо він впливає на транскрипцію даної послідовності. Що стосується транскрипції регуляторних послідовностей, то термін "функціонально зв'язаний" означає, що зв'язані послідовності ДНК є суміжними і, за необхідності, поєднують кодуючі ділянки двох білків, що є суміжними і знаходяться в рамці зчитування. Термін "функціонально зв'язані" стосовно перемикаючих послідовностей означає, що зазначені послідовності можуть здійснювати рекомбінацію на етапі переключення. Термін "вектор" у використовуваному в описі значенні означає молекулу нуклеїнової кислоти, здатну транспортувати зв'язану з ним іншу нуклеїнову кислоту. Одним типом вектора є "плазміда", що являє собою кільцеву петлю дволанцюгової ДНК, у яку можуть бути ліговані додаткові сегменти ДНК. Іншим типом вектора є вірусний вектор, у якому додаткові сегменти ДНК можуть бути ліговані у вірусний геном. Деякі вектори можуть автономно реплікувати у клітині-хазяїні, у яку вони введені (наприклад, бактеріальні вектори, що мають бактеріальну природу реплікації, і епісомні вектори ссавців). Інші вектори (наприклад, неепісомні вектори ссавців) можуть бути вбудовані в геном клітини-хазяїна при введенні в клітину-хазяїна, після чого зазначені вектори реплікують разом з геномом хазяїна. Крім того, деякі вектори можуть направляти експресію генів, з якими вони функціонально зв'язані. Такі вектори визначаються в дійсному описі винаходу як "рекомбінантні експресуючі вектори" (або просто "експресуючі вектори"). Як правило, експресуючі вектори, використовувані в методах рекомбінантних ДНК, часто мають форму плазмід. У дійсному описі винаходу терміни "плазміда" і "вектор" можуть мати взаємозамінні значення, тому що плазміда є найбільш часто використовуваною формою вектора. Однак в обсяг дійсного винаходу входять також інші форми експресуючих векторів, такі як вірусні вектори (наприклад, ретровіруси з дефектом реплікації, аденовіруси й аденоасоційовані віруси), що виконують такі ж функції. Термін "рекомбінантна клітина-хазяїн" (або просто "клітина-хазяїн") у використовуваному в описі значенні означає клітину, у яку введений рекомбінантний експресуючий вектор. Очевидно, що такі терміни відносяться не тільки до визначеної клітини, але і до потомства такої клітини. Тому що в наступних поколіннях можуть з'явитися визначені модифікації внаслідок мутації або впливу навколишніх умов, таке потомство фактично не може бути ідентично материнській 11 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 клітині, але все-таки входить у визначення терміну "клітина-хазяїн" у значенні, використаному в дійсному описі винаходу. Рекомбінантні клітини-хазяї включають, наприклад, трансфектоми, такі як клітини СНО, клітини NS/0 і лімфоцити. У використовуваному в описі значенні термін "суб'єкт" означає людину або тварину, відмінну від людини. Термін "тварина, відмінна від людини" означає всіх хребетних, наприклад, ссавців і нессавців, таких як примати крім людини, вівці, собаки, корови, кури, амфібії, рептилії і т.д. 1. Структури антитіл Інтактні антитіла Інтактні антитіла звичайно є гетеробагатомірними глікопротеїнами, що мають щонайменше два важкі і два легкі ланцюги. Крім IgM інтактні антитіла є гетеротетрамірними глікопротеїнами з довжиною ланцюга близько 150 кДа, що складаються з двох ідентичних легких (L) ланцюгів і двох ідентичних важких (Н) ланцюгів. Кожен легкий ланцюг звичайно зв'язаний з важким ланцюгом одним ковалентним дисульфідним зв'язком, причому число дисульфідних зв'язків між важкими ланцюгами різних ізотипів імуноглобуліну може бути різним. Усі важкі і легкі ланцюги мають також дисульфідні містки всередині ланцюга. З одного кінця кожного важкого ланцюга знаходиться варіабельна ділянка (VH), за якою наступає ряд константних ділянок. Кожен легкий ланцюг має варіабельну ділянку (VL) і константну ділянку з іншого кінця; константна ділянка легкого ланцюга сполучається з першою константною ділянкою важкого ланцюга, і варіабельна ділянка легкого ланцюга сполучається з варіабельною ділянкою важкого ланцюга. Легкі ланцюги антитіл у більшості хребетних можуть бути віднесені до одного з двох типів, іменованих капа і лямбда на підставі амінокислотної послідовності константної ділянки. Залежно від амінокислотної послідовності константної ділянки важких ланцюгів людські антитіла можуть бути віднесені до п'яти різних класів IgA, IgD, IgE, IgG і IgM. IgG і IgA можуть бути далі підрозділені на підкласи IgG1, IgG2, IgG3 і IgG4; і IgA1 і IgA2. Варіанти існують у мишей і щурів, що мають щонайменше IgG2a, IgG2b. Варіабельна ділянка повідомляє антитілу специфічність зв'язування з визначеними ділянками, що характеризуються конкретною варіабельністю, які іменуються ділянками, що визначають комплементарність (CDR). Більш консервативні частини варіабельної ділянки іменуються каркасними ділянками (FR). Варіабельні ділянки важких і легких ланцюгів інтактних антитіл включають чотири каркасні ділянки, з'єднані трьома CDR. CDR у кожнім ланцюзі утримуються в безпосередній близькості один від одного каркасними ділянками і разом з CDR з іншого ланцюга сприяють утворенню антиген-зв'язувального центра антитіл. Константні ділянки безпосередньо не беруть участь у зв'язуванні антитіла з антигеном, але виконують різні ефекторні функції, наприклад, опосередковують антитілозалежну клітиноопосередковану цитотоксичність (ADCC), беруть участь у фагоцитозі завдяки зв'язуванню з рецептором Fcγ, визначають період напіввиведення/кліренс за допомогою неонатального рецептора Fc (FcRn) і комплементзалежну цитотоксичність за допомогою компонента С1q шляху активації комплементу. Константна ділянка IgG2 людини не може активувати комплемент за класичним шляхом активації комплементу або опосередкувати антитілозалежну клітиноопосередковану цитотоксичність. Константна ділянка IgG4 не може активувати комплемент за класичним шляхом активації комплементу і може лише слабко опосередковувати антитілозалежну клітино-опосередковану цитотоксичність. Антитіла, у яких власне кажучи відсутні зазначені ефекторні функції, можуть іменуватися "нелітичними" антитілами. Людські антитіла Людські антитіла можуть бути отримані рядом методів, відомих фахівцям у даній галузі. Людські антитіла можуть бути отримані методом гібридом з використанням ліній мієломних клітин людини або гетеромієломних клітин миші-людини; див. публікації Kozbor, J. Immunol. 133, 3001, (1984) і Brodeur, Monoclonal Antibody Production Techniques and Applications, pp.51-63 (Marcel Dekker Inc, 1987). Альтернативні методи включають використання фагових бібліотек або трансгенних мишей на основі наборів V-ділянок людини (див. публікації Winter G. (1994), Annu. Rev. Immunol. 12, 433-455, Green L.L. (1999), J. Immunol. methods 231, 11-23). В даний час існує кілька ліній трансгенних мишей, у яких мишачі локуси імуноглобуліну замінені сегментами генів імуноглобуліну людини (див. публікації Tomizuka K. (2000) PNAS 97, 722-727; Fishwild D.M. (1996) Nature Biotechnol. 14, 845-851, Mendez M.J., 1997, Nature Genetics, 15, 146-156). У результаті антигенної стимуляції такі миші можуть продукувати набір людських антитіл, з яких можуть бути обрані антитіла, що представляють інтерес. Особливої уваги заслуговує система Trimera (див. публікацію Eren R. et al. (1998) Immunology 93:154-161), відповідно до якої лімфоцити людини трансплантують опроміненим мишам, а саме, система відібраних лімфоцитних антитіл (SLAM, див. публікацію Babcook et al., PNAS (1996) 93:7843-7848), у якій лімфоцити людини (або інші різновиди) ефективно піддають процедурі масового утворення антитіл in vitro з наступним виконанням процедури 12 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 неруйнуючого, обмеженого розведення і відбору, і система Xenomouse II (Abgenix Inc). Альтернативним підходом є метод Morphodoma компанії Morphotek Inc. Для одержання людських антитіл (і їх фрагментів) може бути використаний метод відображення на фазі, див. публікації McCafferty; Nature, 348, 552-553 (1990) і Griffiths A.D. et al. (1994) EMBO 13:3245-3260. Відповідно до зазначеного методу гени V-ділянки антитіла клонують у рамці зчитування в головному або мінорному гені оболонки білка нитковидного бактеріофагу, такого як М13 або fd, і відображають (звичайно за допомогою фагу-хелпера) у вигляді функціональних фрагментів антитіла на поверхні фагової частинки. На підставі функціональних властивостей антитіла відбирають ген, що кодує антитіло, яке володіє зазначеними властивостями. Метод відображення на фазі може бути використаний для відбору антигенспецифічних антитіл з бібліотек, отриманих з В-клітин людини, узятих у суб'єктів, що страждають вищевказаним захворюванням або порушенням, або, альтернативно, у неімунізованих людей-донорів (див. публікацію Marks; J. Mol. Bio. 222, 581-597, 1991). Якщо бажано одержати інтактне людське антитіло, що включає Fc-ділянку, необхідно реклонувати фрагмент, виділений з антитіла, відображеного на фазі, в експресуючих векторах ссавця, що включають необхідні константні ділянки, і одержати лінії клітин, стійко експресуючих дане антитіло. Метод дозрівання афінності (Marks; Bio/technol. 10, 779-783 (1992)) може бути використаний для поліпшення спорідненості зв'язування, при цьому спорідненість первинного людського антитіла поліпшується в результаті послідовної заміни V-ділянок Н- і L-ланцюгів природними варіантами і відбору, виконуваного на підставі поліпшеної спорідненості зв'язування. В даний час відомі також варіанти розглянутого методу, такі як "імпринтінг епітопу"; див. WO 93/06213. Див. також публікацію Waterhouse; Nucl. Acids Res. 21, 2265-2266 (1993). Химерні та гуманізовані антитіла Використання інтактних антитіл, відмінних від людських, при лікуванні захворювань або порушень у людини створює добре відомі проблеми можливої імуногенності, особливо при повторному введенні антитіла, що полягають у тому, що імунна система пацієнта може впізнати чужорідне інтактне антитіло, відмінне від людського, як невласне і викликати нейтралізуючу реакцію. Крім створення цілком людських антитіл (див. вище), протягом останніх років були розроблені різні методи, які дозволяють перебороти вищевказані проблеми, які звичайно припускають зменшення вмісту амінокислотних послідовностей, відмінних від людських, в інтактному терапевтичному антитілі при збереженні відносної простоти одержання антитіл, відмінних від людських, в імунізованих тварин, наприклад, миші, щура або кролика. Для досягнення зазначеної мети широко використовуються два методи. Перший метод включає створення химерних антитіл, які звичайно містять варіабельну ділянку, відмінну від людської (наприклад, гризуна, такого як миша), злиту з константною ділянкою людини. Оскільки антигензв'язувальний центр антитіла локалізований у варіабельних ділянках, химерне антитіло зберігає спорідненість зв'язування з антигеном, але здобуває ефекторні функції константної ділянки людини і тому може виконувати вищеописані ефекторні функції. Химерні антитіла звичайно одержують методами рекомбінантних ДНК. ДНК, що кодує антитіла, (наприклад, кДНК) виділяють і секвенують стандартними методами (наприклад, використовуючи олігонуклеотидні зонди, здатні специфічно зв'язуватися з генами, що кодують варіабельні ділянки H- і L-ланцюгів антитіла за дійсним винаходом. Клітини гібридоми є типовим джерелом такої ДНК. Виділену ДНК вводять у експресуючі вектори, які трансфікують у клітини-хазяї, такі як E.coli, клітини COS, клітини СНО або мієломні клітини, які у противному випадку не продукують білок імуноглобуліну, викликаючи синтез зазначеного антитіла. ДНК може бути модифікована шляхом заміни L- і H-ланцюгів людини кодуючою послідовністю для відповідних константних ділянок Н- і L-ланцюгів, відмінних від людських (наприклад, миші); див., наприклад, публікацію Morrison; PNAS 81, 6851 (1984). Таким чином, інший варіант здійснення винаходу відноситься до химерного антитіла, що включає VН-ділянку, що має послідовність SEQ ID NO:2, 6 або 10, і VLділянку, що має послідовність SEQ ID NO:4, 8 або 12, злиті з константною ділянкою людини (яка може бути ізотипом IgG, наприклад, IgG1). Другий метод включає створення гуманізованих антитіл, у яких вміст частини, відмінної від людської, зменшений шляхом гуманізації варіабельних ділянок. Широко застосовуються два методи гуманізації. Перший метод включає гуманізацію шляхом трансплантації CDR. CDR формують петлі поруч з N-кінцем антитіла, де вони утворюють поверхню в каркасних ділянках. Антиген-зв'язувальна специфічність антитіла визначається, головним чином, топографією і хімічними характеристиками поверхні CDR. Зазначені властивості, у свою чергу, визначаються конформацією окремих CDR, відносним розташуванням CDR, характером і розташуванням бічних ланцюгів залишків, що утворюють CDR. Значне зменшення імуногенності може бути 13 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 досягнуте в результаті трансплантації тільки CDR антитіл, відмінних від людських, (наприклад, миші) ("донорські" антитіла) у підходящу каркасну ділянку людини ("каркасна ділянка акцепторного антитіла") і константні ділянки (див. публікації Jones et al. (1986) Nature 321, 522525 і Verhoeyen M. et al. (1988) Science 239, 1534-1536). Однак сама по собі трансплантація CDR не може гарантувати повного збереження антиген-зв'язувальних властивостей, тому часто виявляється, що необхідно зберегти деякі залишки каркасної ділянки донорського антитіла (іноді визначені як "зворотні мутації") в гуманізованій молекулі для досягнення значної антигензв'язувальної спорідненості (див. публікації Queen C. et al. (1989) PNAS 86, 10029-10033, Co, M. et al. (1991) Nature 351, 501-502). У даному випадку V-ділянки людини, які характеризуються найбільшою гомологією послідовностей (звичайно 60 % або більше) з донорським антитілом, відмінним від людського, можуть бути обрані з бази даних для створення каркасної ділянки людини (FR). FR людини можуть бути обрані з консенсусних або індивідуальних антитіл людини. За необхідності основні залишки з донорського антитіла вводять у каркасну ділянку акцепторного антитіла людини для збереження конформацій CDR. Для ідентифікації таких структурно важливих залишків може бути використане комп'ютерне моделювання антитіла; див. WO99/48523. Альтернативно, гуманізація може бути виконана методом "маскування". Статистичний аналіз унікальних варіабельних ділянок важких і легких ланцюгів імуноглобуліну людини і миші показав, що точні паттерни залишків різні в антитілах людини і миші й у більшості випадків надають перевагу положенням з невеликим числом різних залишків (див. публікації Padlan E.A. et al.; (1991) Mol. Immunol. 28, 489-498 і Pedersen J.T. et al. (1994) J. Mol. Biol. 235; 959-973). Тому імуногенність Fv-фрагмента, відмінного від людського, можна зменшити, заміняючи в каркасних ділянках залишки, що відрізняються від залишків, які виявляються звичайно в антитілах людини. Оскільки антигенність білка можна корелювати з доступністю поверхні, заміна поверхневих залишків може бути достатньою для того, щоб зробити варіабельну ділянку миші "невидимою" для імунної системи людини (див. також публікацію Mark G.E. et al. (1994) in Handbook of Experimental Pharmacology vol. 113: The pharmacology of monoclonal Anthibodies, Springer-Verlag, pp.105-134). Даний процес гуманізації визначається як "маскування", оскільки змінюється тільки поверхня антитіла, при цьому несучі залишки залишаються без зміни. Інший альтернативний метод описаний у WO04/006955. Інші альтернативні підходи включають методи, розглянуті в WO04/006955, і метод HumaneeringTM (Kalobios), який завдяки використанню бактеріальних експресуючих систем дозволяє одержати антитіла, у яких послідовність близька до зародкової лінії людини (Alfenito-M Advancing Protein Therapeutics, January 2007, San Diego, California). Інший метод гуманізації включає відбір каркасних ділянок акцепторного антитіла людини на підставі структурної подібності ділянок CDR людини з ділянками CDR донорського антитіла миші, а не на гомології інших ділянок антитіла, таких як каркасні ділянки. Зазначений метод також відомий як супергуманізація ((Evogenix Inc.; Hwang et al. (2005) Methods 36:35-42). Фахівцям у даній галузі повинно бути зрозуміло, що термін "виділений" означає не тільки джерело фізичного походження матеріалу, але також служить для визначення матеріалу, який ідентичний у структурному відношенні до даного матеріалу, але отриманий з іншого джерела. Таким чином, "залишки, виявлені в донорському антитілі" необов'язково повинні бути очищені від донорського антитіла. Фрагменти антитіла Деякі варіанти здійснення винаходу відносяться до терапевтичного антитіла, яке є антигензв'язувальним фрагментом. Такі фрагменти можуть бути функціональними антигензв'язувальними фрагментами інтактних, гуманізованих і/або химерних антитіл, такими як Fab, Fd, Fab", F(ab")2, Fv, ScFv фрагменти вищеописаних антитіл. Фрагменти, у яких відсутня константна ділянка, втрачають здатність активувати комплемент за класичним шляхом або опосередкувати антитілозалежну клітино-опосередковану цитотоксичність. Такі фрагменти традиційно одержують у результаті протеолітичного розщеплення інтактних антитіл, наприклад, папаїном (див., наприклад, WO 94/29348), але фрагменти можуть бути отримані безпосередньо з рекомбінантно трансформованих клітин-хазяїнів. Для одержання ScFv див. публікацію Bird et al. (1988) Science, 242, 423-426. Крім того, фрагменти антитіла можуть бути отримані різними методами генної інженерії, описаними нижче. Fv-фрагменти, очевидно, характеризуються більш низькою енергією взаємодії двох ланцюгів, ніж Fab-фрагменти. Щоб стабілізувати асоціацію VН- і VL-ділянок, зазначені ділянки зв'язують пептидами (Bird et al., (1988) Science 242, 423-426, Huston et al., PNAS, 85, 5879-5883), дисульфідними містками (Glockshuber et al. (1990) Biochemistry, 29, 1362-1367) і мутаціями "виступ в отворі" (Zhu et al. (1997), Protein Sci., 6, 781-788). ScFv-фрагменти можуть бути 14 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 отримані методами, добре відомими фахівцям у даній галузі; див. публікації Whitlow et al. (1991) Methods companion Methods Enzymol., 2, 97-105 і Huston et al. (1993) Int.Rev.Immunol. 10, 195217. ScFv-фрагменти можуть бути отримані в бактеріальних клітинах, таких як E.Coli, але звичайно зазначені фрагменти одержують в еукаріотичних клітинах. Одним недоліком ScFvфрагментів є одновалентність продукту, що унеможливлює підвищену авідність через багатовалентне зв'язування і визначає короткий період напіввиведення. Спроби перебороти зазначені проблеми привели до використання двовалентного (ScFv") 2- фрагмента, отриманого з ScFv-фрагмента, що містить додатковий С-кінцевий цистеїн, шляхом хімічного зв'язування (Adams et al. (1993) Can.Res. 53, 4026-4034 і McCartney et al. (1995) Protein Eng. 8, 301-314) або спонтанної сайт-спеціфічної димерізації ScFv-фрагмента, що містить неспарений С-кінцевий залишок цистеїну (див. публікацію Kipriyanov et al. (1995) Cell. Biophys 26, 187-204). Альтернативно, з ScFv-фрагмента можуть бути отримані багатоміри в результаті укорочування пептидного лінкера між залишками 3-12 з утворенням "діатіл"; див. публікацію Holliger et al., PNAS (1993), 90, 6444-6448. Зменшення лінкера може далі привести до одержання тримерів ScFv ("триатіла"; див. публікацію Kortt et al. (1997) Protein Eng., 10, 423-433) і тетрамерів ("тетратіла"; див. публікацію Le Gall et al. (1999) FEBS Lett., 453, 164-168). Двовалентні молекули ScFv можуть бути також створені шляхом генетичного злиття з фрагментами, що димеризують білок, з утворенням "мініантитіл" (див. публікацію Pack et al. (1992) Biochemistry 31, 1579-1584) і "мінітіл" (див. публікацію Hu et al. (1996) Cancer Res. 56, 3055-3061). Можуть бути також отримані тандеми ScFv-Sc-Fv ((ScFv)2) шляхом зв'язування двох ланок ScFv третім пептидним лінкером; див. публікацію Kurucz et al. (1995) J. Immol. 154, 4576-4582. Біспецифічні діатіла можуть бути отримані шляхом нековалентного зв'язування двох одноланцюгових продуктів злиття, що складаються з VН-ділянки одного антитіла, з'єднаного коротким лінкером з VL-ділянкою іншого антитіла; див. публікацію Kipriyanov et al. (1998), Int. J. Can. 77, 763-772. Стійкість таких біспецифічних діатіл може бути збільшена шляхом введення дисульфідних містків або мутацій "виступ в отворі", описаних вище, або утворення одноланцюгових діатіл (ScDb), у яких два гібридних ScFv-фрагменти з'єднані пептидним лінкером; див. публікацію Kontermann et al. (1999) J. Immunol. Methods 226, 179-188. Чотиривалентні біспецифічні молекули можуть бути отримані, наприклад, шляхом злиття ScFv-фрагмента з СН3-ділянкою молекули IgG або з Fab-фрагментом за допомогою шарнірної ділянки; див. публікацію Coloma et al. (1997) Nature Biotechnol. 15, 159-163. Альтернативно, чотиривалентні біспецифічні молекули були створені шляхом злиття біспецифічних одноланцюгових діатіл (див. публікацію Alt et al., (1999) FEBS Lett 454, 90-94). Більш дрібні чотиривалентні біспецифічні молекули можуть бути також утворені шляхом димеризації тандемів ScFv-ScFv за допомогою лінкера, що містить фрагмент спіраль-петля-спіраль (DiBi-мініантитіла; див. публікацію Muller et al. (1998) FEBS Lett 432, 45-49), або одноланцюгової молекули, що містить чотири варіабельні ділянки антитіла (V H і VL) в орієнтації, що перешкоджає внутрішньомолекулярному спарюванню (тандемне антитіло; див. публікацію Kipriyanov et al.,(1999) J. Mol. Biol. 293, 41-56). Біспецифічні F(ab")2-фрагменти можуть бути створені шляхом хімічного зв'язування Fab'-фрагментів або гетеродимерізації за допомогою лейцинових зіперів (див. публікації Shalaby et al. (1992) J. Exp. Med. 175, 217-225 і Kostelny et al. (1992), J. Immunol. 148, 1547-1553). Крім того, можуть бути отримані так звані антитіла-домени, створені на основі виділених VН- або VL-ділянок (Domantis Ltd.); див. патенти США №№ 6248516, 6291158, 6172197. Гетерокон'югатні антитіла Гетерокон'югатні антитіла є похідними антитілами, що також відносяться до одного варіанту здійснення дійсного винаходу. Гетерокон'югатні антитіла складаються з двох ковалентно зв'язаних антитіл, утворених за допомогою будь-яких відомих методів перехресного зв'язування. Див. патент США № 4676980. Інші модифікації Антитіла за дійсним винаходом можуть також включати будь-які інші модифікації в константних ділянках. Наприклад, відомо, що глікозілювання антитіл у консервативних положеннях константних ділянок впливає на функцію антитіл, зокрема, на вищеописані ефекторні функції; див., наприклад, публікацію Boyd et al. (1996), Mol. Immunol. 32, 1311-1318. Варіанти глікозілювання терапевтичних антитіл за дійсним винаходом включають додавання, заміну, делецію або модифікацію однієї або декількох вуглеводних частин. Введення фрагмента аспарагін-Х-серин або аспарагін-Х-треонін створює потенційний сайт для ферментативного приєднання вуглеводних частин і тому може бути використане для керування глікозілюванням антитіла. У публікації Raju et al. (2001) Biochemistry 40, 8868-8876 зазначено, що сіалілування кінця імуноадгезину TNFR-IgG було збільшено в результаті регалактозилювання і/або ресіалілування з використанням бета-1,4-галактозилтрансферази і/або альфа-2,3 15 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 сіалілтрансферази. Вважається, що підвищене сіалілування кінця збільшує період напіввиведення імуноглобуліну. Антитіла, поряд з більшістю глікопротеїнів, звичайно утворюються в природі у вигляді суміші глікоформ. Зазначена суміш є особливо очевидною при одержанні антитіл в еукаріотичних клітинах, зокрема, у клітинах ссавців. Були розроблені різні методи одержання визначених глікоформ; див. публікації Zhang et al., Science (2004), 303, 371, Sears et al., Science, (2001) 291, 2344, Wacker et al. (2002) Science, 298, 1790, Davis et al. (2002) Chem. Rev. 102, 579, Hang et al. (2001) Acc. Chem. Res. 34, 727. Таким чином, дійсний винахід відноситься до безлічі терапевтичних антитіл (які можуть бути ізотипом IgG, наприклад, IgG1), що включає визначене число (наприклад, 7 або менше, зокрема, 5 або менше, а саме, дві або одну) глікоформ зазначених антитіл. Похідні за дійсним винаходом включають також терапевтичні антитіла за дійсним винаходом, зв'язані з небілковим полімером, таким як поліетиленгліколь (РEG), поліпропіленгліколь або поліоксіалкілен. Кон'югація білків з PEG є відомим методом збільшення періоду напіввиведення білків, а також зменшення антигенності та імуногенності білків. Метод обробки поліетиленгліколем з різними молекулярними масами і різними типами (лінійні або розгалужені) був досліджений з використанням інтактних антитіл, а також Fab'-фрагментів; див. публікацію Koumenis I.L. et al. (2000) Int. J. Pharmaceut. 198:83-95. Конкретний варіант здійснення винаходу відноситься до антиген-зв'язувального фрагмента за дійсним винаходом без ефекторних функцій а) активації комплементу за класичним шляхом і b) опосередкування антитілозалежної клітино-опосередкованої цитотоксичності; (такого як Fab-фрагмент або scFv), зв'язаного з PEG. Вважається, що взаємодія Fc-ділянки антитіла з різними Fc-рецепторами (FсγR) опосередковує ефекторні функції антитіла, що включають антитілозалежну клітиноопосередковану цитотоксичність (ADCC), фіксацію комплементу, фагоцитоз і період напіввиведення/кліренс антитіла. Fc-ділянка антитіл за дійсним винаходом може бути піддана різним модифікаціям залежно від необхідної ефекторної функції. Зокрема, константні ділянки людини, у яких відсутні функції а) активації комплементу за класичним шляхом і b) опосередкування антитілозалежної клітино-опосередкованої цитотоксичності, включають константну ділянку IgG4, константну ділянку IgG2 і константну ділянку IgG1, що містять специфічні мутації, наприклад, мутації в положеннях 234, 235, 236, 237, 297, 318, 320 і/або 322, описані у EP0307434 (WO8807089), EP0629240 (WO9317105) і WO 2004/014953. Крім того, було відзначено, що мутації в положенні залишків 235 або 237 СН2-домена константної ділянки важкого ланцюга (нумерація Кабата; європейська система індексації) зменшують зв'язування з FcγRI, FcγRII та FcγRIII і, таким чином, зменшують антитілозалежну клітино-опосередковану цитотоксичність (ADCC) (Duncan et al. Nature 1988, 332; 563-564; Lund et al. J. Immunol. 1991, 147; 2657-2662; Chappel et al. PNAS, 1991, 88; 9036-9040; Burton and Wооf, Adv. Immunol. 1992, 51; 1-84; Morgan et al., Immunology 1995, 86; 319-324; Hеzаrеh et al., J. Virol. 2001, 75 (24); 1216112168). У деяких наукових звітах була описана участь деяких із зазначених залишків у рекрутинзі або опосередкуванні комплементзалежної цитотоксичності (СDС) (Morgan et al., 1995; Xu et al., Cell. Immunol. 2000; 200:16-26; Hezareh et al., J. Virol. 2001, 75(24); 12161-12168). Тому залишки 235 і 237 були мутовані з введенням залишків аланіну (Brett et al., Immunology 1997, 91; 346-353; Bartholomew et al., Immunology 1995, 85; 41-48; і WO9958679 для зменшення комплемент-опосередкованих і FcγR-опосередкованих впливів. Антитіла, які включають зазначені константні ділянки, можуть іменуватися "нелітичними" антитілами. В антитіло можна ввести епітоп зв'язування з рецептором реутилізації для збільшення періоду напіввиведення з кров'яного русла; див. патент США № 5739277. В даний час відомі п'ять пізнаваних рецепторів Fcγ, а саме, FcγR(I), FcγRIIa, FcγRIIb, FcγRIIIa і неонатальний рецептор FcRn. У публікації Shields et al., (2001) J. Biol. Chem. 276, 6591-6604 було продемонстровано, що загальний набір залишків IgG1 бере участь у зв'язуванні всіх рецепторів FcγR, у той час як FcγRII і FcγRIII використовують інші сайти, які не входять у зазначений загальний набір. Одна група залишків IgG1 зменшувала зв'язування з усіма рецепторами FcγR при заміні на аланін: Pro-238, Asp-265, Asp-270, Asn-297 і Pro-239. Усі зазначені залишки знаходяться в СН2-домені IgG і утворюють кластер поруч із шарнірною сполукою СН1 і СН2. У той час як FcγRI використовує тільки загальний набір залишків IgG1 для зв'язування, FcγRII і FcγRIII взаємодіють також з іншими залишками крім загального набору. Заміна деяких залишків зменшувала зв'язування тільки з FcγRII (наприклад, Arg-292) або FcγRIII (наприклад, Glu-293). Деякі варіанти характеризувалися кращим зв'язуванням з FcγRII або FcγRIII, але не впливали на зв'язування з іншим рецептором (наприклад, заміна Ser-267 аланіном (Ala) поліпшувала зв'язування з FcγRII, не впливаючи на зв'язування з FcγRIII). Інші варіанти характеризувалися кращим зв'язуванням з FcγRII або FcγRIII при зменшенні 16 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 зв'язування з іншим рецептором (наприклад, заміна Ser-298 аланіном (Ala) поліпшувала зв'язування з FcγRIII і зменшувала зв'язування з FcγRII). Варіанти IgG1, які забезпечують краще зв'язування з FcγRIIIa, характеризувалися загальним введенням аланіну в положеннях Ser-298, Glu-333 і Lys-334. Вважається, що неонатальний рецептор FcRn захищає молекули IgG від руйнування і, таким чином, збільшує період напіввиведення з кров'яного русла і трансцитоз у тканинах (див. публікації Junghans R.P. (1997) Immunol. Res. 16, 29-57 і Ghetie et al. (2000) Annu. Rev. Immunol. 18, 739-766). Встановлено, що залишки IgG1 людини, які безпосередньо взаємодіють з FcRn людини, включають Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 і His435. Терапевтичне антитіло за дійсним винаходом може включати будь-які вищевказані модифікації константних ділянок. 2. Способи одержання Антитіла за дійсним винаходом можуть бути продуковані в трансгенних організмах, таких як кози (див. публікацію Pollock et al. (1999), J. Immunol. Methods 231:147-157), курчата (див. публікацію Morrow KJJ (2000) Genet. Eng. News 20:1-55), миші (див. вищевказану публікацію Pollock) або рослини (див. публікації Doran P.M. (2000) Curr. Opinion Biotechnol. 11, 199-204, Ma JK-C (1998), Nat. Med. 4; 601-606, Baez J. et al., BioPharm (2000) 13:50-54, Stoger E. et al. (2000) Plant Mol. Biol. 42:583-590). Антитіла можуть бути також отримані методами хімічного синтезу. Однак антитіла за дійсним винаходом звичайно одержують методом культивування рекомбінантних клітин, добре знайомим фахівцям у даній галузі. Полінуклеотид, який кодує антитіло, виділяють і вводять у вектор, що реплікується, такий як плазміда, для наступного клонування (ампліфікації) або експресії в клітині-хазяїні. Однією прийнятною експресуючою системою є система на основі глутаматсинтетази (компанії Lonza Biologics), особливо при використанні у якості клітини-хазяїна СНО або NS0 (див. нижче). Полінуклеотид, що кодує антитіло, легко виділяють і секвенують стандартними методами (наприклад, за допомогою олігонуклеотидних зондів). Застосовувані вектори включають плазміду, вірус, фаг, транспозони, мініхромосоми, з яких найбільш типовими є плазміди. Такі вектори звичайно додатково включають сигнальну послідовність, оріджин реплікації, один або кілька генів-маркерів, енхансер, промотор і послідовності термінації транскрипції, функціонально зв'язані з полінуклеотидом легкого і/або важкого ланцюга для полегшення експресії. Полінуклеотид, що кодує легкі і важкі ланцюги, може бути вставлений в окремі вектори й одночасно або послідовно введений (наприклад, шляхом трансформації, трансфекції, електропорації або трансдукції) у ту ж клітину-хазяїна, або, при бажанні, важкий і легкий ланцюги можуть бути вставлені в той самий вектор до введення в клітину-хазяїна. Фахівцям у даній галузі повинно бути очевидно, що внаслідок надмірності генетичного коду, крім вищевказаних полінуклеотидів, також існують інші полінуклеотиди, що кодують поліпептиди за дійсним винаходом. Сигнальні послідовності Антитіла за дійсним винаходом можуть бути отримані у вигляді злитого білка з гетерологічною сигнальною послідовністю, що має специфічний сайт розщеплення біля N-кінця зрілого білка. Сигнальна послідовність повинна бути впізнана і процесована клітиною-хазяїном. Для прокаріотичних клітин-хазяїнів сигнальною послідовністю може бути лужна фосфатаза, пеніциліназа або лідерні послідовності теплостійкого ентеротоксину II. Для секреції в дріжджах сигнальними послідовностями можуть бути лідерна послідовність інвертази дріжджів, лідерна послідовність α-фактора або лідерні послідовності кислої фосфатази; див., наприклад, WO90/13646. У системах клітин ссавців можуть бути використані секретуючі лідерні послідовності вірусів, такі як сигнальна послідовність вірусу простого герпеса gD і сигнальна послідовність природного імуноглобуліну (наприклад, важкий ланцюг Ig людини). Сигнальна послідовність звичайно лігована в рамці зчитування з полінуклеотидом, що кодує антитіло за дійсним винаходом. Оріджин реплікації Оріджини реплікації добре відомі в даній галузі, при цьому pBR322 придатний для більшості грамнегативних бактерій, 2µ плазміда придатна для більшості дріжджів, і різні віруси, такі як SV40, вірус поліоми, аденовірус, VSV або BPV, придатні для більшості клітин ссавців. Оріджин реплікації звичайно не потрібний для інтегрованих експресуючих векторів ссавців, за винятком введення вектора в E.Coli. Однак оріджин реплікації SV40 може бути використаний, оскільки даний вірус містить ранній промотор. Селектуючий маркер Типові селектуючі гени кодують білки, які (а) повідомляють стійкість до антибіотиків або інших токсинів, такимх як ампіцилін, неоміцин, метотрексат або тетрациклін, або (b) заповнюють недолік ауксотрофів або надають поживні речовини, відсутні в комплексному середовищі, або 17 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 (с) виконують обидві функції. Схема селекції може включати уповільнення росту клітин-хазяїнів, що не містять вектора або векторів. Клітини, які були успішно трансформовані генами, що кодують терапевтичне антитіло за дійсним винаходом, виживають, наприклад, завдяки лікарській стійкості, повідомленої доставленим селектуючим маркером. Одним прикладом є DHFR-селектуюча система, у якій трансформанти утворюються в DHFR-негативних лініях хазяїнів (див., наприклад, публікацію Page and Sydenham, 1991, Biotechnology 9:64-68). У зазначеній системі ген DHFR доставляють разом з полінуклеотидними послідовностями антитіла за дійсним винаходом і потім відбирають DHFR-позитивні клітини шляхом видалення нуклеозиду. За необхідності, для відбору трансформантів з ампліфікацією гена DHFR також використовують інгібітор DHFR метотрексат. У результаті функціонального зв'язування гена DHFR з кодуючими послідовностями антитіла за дійсним винаходом або функціональними похідними зазначених послідовностей ампліфікація гену DHFR викликає супутню ампліфікацію послідовностей необхідного антитіла, що представляють інтерес. Клітини СНО являють собою особливо придатну лінію клітин для відбору за допомогою DHFR/метотрексату, і в даній галузі добре відомі методи ампліфікації і відбору клітин-хазяїнів з використанням системи DHFR; див. публікацію Kaufman R.J. et al., J. Mol. Biol. (1982) 159, 601-621; для огляду див. публікацію Werner R.G., Noe W., Kopp K., Schluter M., "Appropriate mammalian expression systems for biopharmaceuticals", Arzneimittel-Forschung, 48(8):870-80, 1988 Aug. Іншим прикладом є експресуюча система на основі глутаматсинтетази (Lonza Biologics). Підходящим селектуючим геном, призначеним для використання в дріжджах, є ген trp1; див. публікацію Stinchcomb et al., Nature, 282, 38, 1979. Промотори Підходящі промотори для експресії антитіл за дійсним винаходом функціонально зв'язані з ДНК/полінуклеотидом, що кодує дане антитіло. Промотори для прокаріотичних хазяїнів включають промотор phoA, промоторні системи на основі бета-лактамази і лактози, лужну фосфатазу, триптофан і гібридні промотори, такі як Tac. Промотори, придатні для експресії в дріжджових клітинах, включають 3-фосфогліцераткіназу або інші гліколітичні ферменти, наприклад, енолазу, гліцеральдегід-3-фосфатдегідрогеназу, гексокіназу, піруватдекаробоксилазу, фосфофруктокіназу, глюкоза-6-фосфатізомеразу, 3фосфогліцератмутазу, глюкокіназу та інші. Індуцибельні дріжджові промотори включають алкогольдегідрогеназу-2, ізоцитохром С, кислу фосфатазу, металотіонеїн, ферменти, що відповідають за азотистий обмін або утилізацію мальтози/галактози, та інші. Промотори, призначені для експресії в клітинних системах ссавців, включають РНКполімеразу II, у тому числі, вірусні промотори, такі як вірус поліоми, пташиний поксвірус і аденовіруси (наприклад, аденовірус 2), вірус папіломи великої рогатої худоби, пташиний вірус саркоми, цитомегаловірус (зокрема, промотор передраннього гену), ретровірус, вірус гепатиту В, актин, вірус саркоми Рауса (RSV) і ранній або пізній вакуолізуючий мавпячий вірус 40, і невірусні промотори, такі як EF-1-альфа (Mizushima and Nagata, Nucleic Acids Res., 1990, 18(17):5322), та інші. Вибір промотору може бути заснований на підходящій сумісності з клітиною-хазяїном, використовуваною для експресії. Енхансер У разі потреби, наприклад, для експресії у вищих еукаріотичних клітинах, можуть бути використані додаткові енхансерні елементи замість промоторів або енхансерні елементи, виявлені в описаних вище промоторах. Підходящі енхансерні послідовності ссавців включають енхансерні елементи з глобіну, еластази, альбуміну, фетопротеїну, металотіоніну й інсуліну. Альтернативно можна використовувати енхансерний елемент із вірусу еукаріотичної клітини, такий як енхансер SV40, енхансер раннього промотору цитомегаловірусу, енхансер вірусу поліоми, енхансер бакуловірусу або локус IgG2a миші (див. WO04/009823). Незважаючи на те, що такі енхансери звичайно знаходяться у векторі в сайті зверху від промотору, вони можуть бути розташовані в будь-якому місці, наприклад, у нетрансльованій ділянці або внизу від сигналу поліаденілювання. Енхансер і його місце розташування можуть бути обрані з урахуванням підходящої сумісності з клітиною-хазяїном, використовуваною для експресії. Поліаденілювання/термінація В еукаріотичних системах сигнали поліаденілювання функціонально зв'язані з полінуклеотидом, що кодує антитіло за дійсним винаходом. Такі сигнали звичайно знаходяться біля 3'-кінця відкритої рамки зчитування. У системах ссавців необмежуючі приклади сигналів включають послідовності, виділені з гормонів росту, альфа-фактора елонгації 1 і вірусних (наприклад, SV40) генів або довгих кінцевих повторів ретровірусів. У дріжджових системах необмежуючі приклади сигналів поліаденілювання/термінації включають послідовності, виділені з генів фосфогліцераткінази (PGK) і алкогольдегідрогенази 1 (ADH). В прокаріотичній системі 18 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 сигнали поліаденілювання звичайно не потрібні, і замість них звичайно використовують більш короткі і більш визначені термінаторні послідовності. Вибір послідовностей поліаденілювання/термінації може бути заснований на підходящій сумісності з клітиноюхазяїном, використовуваною для експресії. Інші методи/елементи для підвищення виходу Крім вищевикладеного, інші ознаки, які дозволяють збільшити вихід, включають елементи ремоделювання хроматину, інтрони і модифікацію конкретного кодону клітини-хазяїна. Кодон, використовуваний в антитілі за дійсним винаходом, може бути модифікований для відповідності до зсуву кодону клітини-хазяїна, наприклад, для збільшення виходу транскрипту і/або продукту (див., наприклад, публікацію Hoekema A. et al., Mol. Cell Biol., 1987, 7(8):2914-24). Вибір кодонів може бути заснований на підходящій сумісності з клітиною-хазяїном, використовуваною для експресії. Клітини-хазяї Підходящі клітини-хазяї для клонування або експресії векторів, що кодують антитіла за дійсним винаходом, є, наприклад, прокаріотичними, дріжджовими або вищими еукаріотичними клітинами. Підходящі прокаріотичні клітини включають еубактерії, наприклад, ентеробактерії, такі як Escherichia, наприклад, E.Coli (наприклад, АТСС 31446, 31537, 27325), Enterobacter, Erwinia, Klebsiella Proteus, Salmonella, наприклад, Salmonella typhimurium, Serratia, наприклад, Serratia marcescans, і Shigella, а також Bacilli, такі як B.subtilis і B.licheniformis (див. DD 266710), Pseudomonas, такі як P.aeruginosa і Streptomyces. З дріжджових клітин-хазяїнів можуть бути також використані Saccharomyces cerevisiae, schizosaccharomyces pombe, Kluyveromyces (наприклад, АТСС 16045, 12424, 24178, 56500), yarrowia (EP402226), Pichia Pastoris (EP183070, див. також публікацію Peng et al., J. Biotechnol. 108 (2004) 185-192), Candida, Trichoderma reesia (EP244234), Penicillin, Tolypocladium і Aspergillus, такі як A.nidulans і A.niger. Незважаючи на те, що в обсяг дійсного винаходу входять прокаріотичні та дріжджові клітини-хазяї, звичайно клітинами-хазяїнами за дійсним винаходом є клітини хребетних. Підходящі клітини-хазяї хребетних включають клітини ссавців, такі як COS-1 (АТСС № CRL 1650), COS-7 (АТСС CRL 1651), лінія клітин нирок ембріону людини 293, PerC6 (Crucell), клітини нирок дитинчати хом'ячка (ВНК) (АТСС CRL 1632), ВНК570 (АТСС № CRL 10314), 293 (АТСС № CRL 1573), клітини яєчника китайського хом'ячка СНО (наприклад, СНО-К1, АТСС № CCL 61), лінія клітин DHFR-CHO, така як DG44 (див. приведену вище публікацію Urlaub et al., (1986)), зокрема, лінії клітин СНО, адаптовані для суспензійної культури, клітини Сертолі миші, клітини нирки мавпи, клітини нирки африканської зеленої мавпи (АТСС CRL 1587), клітини HELA, клітини нирки собаки (АТСС CCL 34), клітини легень людини (АТСС CCL 75), клітини Hep G2 і мієломні або лімфомні клітини, наприклад, NS0 (див. патент США № 5807715), Sp2/0, Y0. Таким чином, один варіант здійснення винаходу відноситься до стійко трансформованої клітини-хазяїна, що містить вектор, який кодує важкий ланцюг і/або легкий ланцюг терапевтичного антитіла за дійсним винаходом. Такі клітини-хазяї звичайно містять перший вектор, що кодує легкий ланцюг, і другий вектор, що кодує важкий ланцюг. Такі клітини-хазяї можуть бути також створені або адаптовані для зміни якості, функції і/або виходу антитіла за дійсним винаходом. Необмежуючі приклади включають експресію специфічних модифікуючих (наприклад, глікозилюючих) ферментів і чаперонів, що виробляють укладання білку. Методи культивування клітин Клітини-хазяї, трансформовані векторами, які кодують терапевтичні антитіла за дійсним винаходом, можна культивувати будь-яким методом, відомим фахівцям у даній галузі. Клітинихазяї можна культивувати в обертових колбах, струшуваних колбах, ролер-флаконах або системах порожніх волокон, але для широкомасштабного виробництва кращі реактори з мішалками або реактори з кошиками (див., наприклад, Wave Biotech. Somerset, New Jersey, USA) особливо для суспензійних культур. Баки з мішалкою звичайно пристосовують для аерації за допомогою барботерів, дефлекторів або імпелерів зі слабким зрушувальним зусиллям. Для барботажних колон і реакторів з перемішуванням потоку повітря може бути використана пряма аерація пухирцями повітря або кисню. При культивуванні клітин-хазяїнів у безсироватковому культуральному середовищі бажано ввести в середовище агент, що захищає клітини, такий як плюронік F-68, для запобігання руйнуванню клітин у результаті процесу аерації. Залежно від характеристик клітини-хазяїна як субстрати росту для якірної підкладки можуть бути використані мікроносії, або клітини можуть бути адаптовані для суспензійної культури (яка є типовою). При культивуванні клітин-хазяїв, зокрема, клітини-хазяїв хребетних, можуть бути використані різні методи, такі як періодичне культивування, культивування з підживленням, повторюване періодичне культивування (див. публікацію Drapeau et al. (1994) cytotechnology 15:103-109), 19 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 тривале періодичне культивування або перфузійне культивування. Хоча рекомбінантно трансформовані клітини-хазяї ссавців можна культивувати в сироватковому середовищі, що включає фетальну телячу сироватку (FCS), клітини-хазяї переважно культивують у синтетичному безсироватковому середовищі, аналогічному до описаного в публікації Keen et al. (1995) Cytotechnology 17:153-163, або в комерційно доступному середовищі, такому як ProCHOCDM UltraCHO (Cambrex NJ, USA), що містить при необхідності джерело енергії, таке як глюкоза, і синтетичні фактори росту, такі як рекомбінантний інсулін. Культивування клітин-хазяїв у безсироватковому середовищі може зажадати адаптації зазначених клітин для росту в безсироваткових умовах. Одним методом адаптації є культивування таких клітин-хазяїв у сироватковому середовищі і послідовній заміні 80 % культурального середовища безсироватковим середовищем, у результаті чого клітини-хазяї будуть адаптуватися до безсироваткових умов (див., наприклад, Scharfenberg K. et al. (1995) in Animal Cell technology: st Developments towards the 21 cеntury (Beuvery E.C. et al., eds), pp.619-623, Kluwer Academic publishers). Антитіла за дійсним винаходом, які секретуються в середовищі, можуть бути виділені й очищені від середовища різними методами з досягненням ступеня чистоти, що підходить для передбачуваного застосування. Наприклад, застосування терапевтичних антитіл за дійсним винаходом для лікування людей вимагає щонайменше 95 % чистоти, визначеної методом SDSPAGE, звичайно 98 % або 99 % чистоти в порівнянні з культуральним середовищем, що містить терапевтичні антитіла. У першому випадку клітинний дебріс з культурального середовища звичайно видаляють центрифугуванням з наступним виконанням стадії очищення супернатанту за допомогою мікрофільтрації, ультрафільтрації і/або глибинної фільтрації. Альтернативно антитіло може бути отримане мікрофільтрацією, ультрафільтрацією або глибинною фільтрацією без попереднього центрифугування. Існує цілий ряд інших методів, таких як діаліз і гельелектрофорез, і хроматографічних методів, таких як хроматографія на колонці з гідроксіапатитом (НА), афінна хроматографія (яка необов'язково включає афінну систему мічення, таку як полігістидин) і/або хроматографія гідрофобної взаємодії (HIC, див. патент США № 5429746). В одному варіанті здійснення антитіла за дійсним винаходом одержують після виконання різних стадій очищення за допомогою афінної хроматографії білків А або G з наступним виконанням іонообмінної хроматографії і/або хроматографії на колонці з гідроксіапатитом, аніонообмінної або катіонообмінної хроматографії, гель-хроматографії й осадження сульфатом амонію. Крім того, можуть бути використані різні стадії видалення вірусу (наприклад, нанофільтрація з використанням, наприклад, фільтра DV-20). У результаті виконання вищевказаних стадій одержують очищений (звичайно моноклональний) препарат, що містить щонайменше 10 мг/мл або більше, наприклад, 100 мг/мл або більше антитіла за дійсним винаходом, що являє собою ще один варіант здійснення винаходу. Концентрація до 100 мг/мл або більше може бути досягнута ультрацентрифугуванням. Такі препарати власне кажучи не містять агреговані форми антитіл за дійсним винаходом. Бактеріальні системи особливо придатні для експресії фрагментів антитіла. Такі фрагменти локалізовані всередині клітини або в периплазмі. Нерозчинні периплазматичні білки можуть бути екстраговані і піддані повторному укладанню з утворенням активних білків відповідно до методів, відомих фахівцям у даній галузі; див. публікації Sanchez et al. (1999) J. Biotechnol. 72, 13-20 і Cupit P.M. et al. (1999) Lett. Appl. Microbiol., 29, 273-277. 3. Фармацевтичні композиції і спосіб введення Очищені препарати антитіл за дійсним винаходом, описані вище, можуть бути введені у фармацевтичні композиції для застосування при лікуванні вищевказаних захворювань і порушень у людини. Такі композиції звичайно додатково включають фармацевтично прийнятний (тобто інертний) носій, використовуваний у загальноприйнятій фармацевтичній практиці; див., наприклад, Remingtons Pharmaceutical Sciences, 16th ed, (1980), Mack Publishing Co. Приклади таких носіїв включають стерилізовані носії, такі як фізіологічний розчин, розчин Рінгера або розчин декстрози, що містять підходящі буфери для досягнення значення рН, що дорівнює 5-8. Фармацевтичні композиції для ін'єкцій (наприклад, внутрішньовенних, внутрішньочеревних, внутрішньошкірних, підшкірних, внутрішньом'язових або внутрішньопортальних) або вливань не містять видимих частинок і можуть включати від 0,1 мг до 10 г терапевтичного антитіла, звичайно від 5 мг до 25 мг антитіла. Методи одержання таких фармацевтичних композицій добре відомі фахівцям у даній галузі. В одному варіанті здійснення винаходу фармацевтичні композиції містять від 0,1 мг до 10 г терапевтичних антитіл за дійсним винаходом в стандартній лікарській формі з прикладеною інструкцією із застосування. Фармацевтичні композиції за дійсним винаходом можуть бути ліофілізовані (висушені виморожуванням) для відновлення перед введенням способами, добре відомими фахівцям у 20 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 даній галузі. Коли варіанти здійснення винаходу відносяться до антитіл за дійсним винаходом з ізотипом IgG1, у фармацевтичну композицію може бути доданий хелатор міді, такий як цитрат (наприклад, цитрат натрію), EDTA або гістидин, для зменшення ступеня опосередковуваного міддю руйнування антитіл даного ізотипу; див. EP0612251. Ефективні дози і схеми введення антитіла за дійсним винаходом звичайно визначаються емпірично і залежать від таких факторів, як вік, маса тіла і стан здоров'я пацієнта, захворювання, що підлягає лікуванню, або порушення. Такі фактори повинні враховуватися лікуючим лікарем. Посібник з вибору відповідних доз приведений, наприклад, у публікації Smith et al. (1977) Antibodies in human diagnosis and therapy, Raven Press, New York, але прийнятна доза звичайно знаходиться в межах від 0,1 мг до 1 г. В одному варіанті здійснення винаходу схема введення при лікуванні людини складає від 0,1 мг до 10 г терапевтичного антитіла за дійсним винаходом, що вводиться підшкірно один раз на тиждень або через два тижні, або у вигляді внутрішньовенного вливання через 1 або 2 місяці. Композиції за дійсним винаходом можуть бути також використані в профілактичних цілях. 4. Клінічні застосування Дійсний винахід відноситься до антитіла, що зв'язується з IL-8, Gro-альфа, Gro-бета, Groгама і ENA-78 людини. Дійсний винахід відноситься також до способів лікування зазначеним антитілом захворювань або порушень, що характеризуються підвищеним або незбалансованим рівнем одного або декількох хемокінів, що включають IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини, а саме, хронічного обструктивного захворювання легень (COPD), остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, респіраторного дистрес-синдрому в дорослих (ARDS), захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу, до фармацевтичних композицій, що містять зазначене антитіло, і способів приготування таких композицій. Дійсний винахід відноситься також до застосування антитіла для приготування лікарського засобу для лікування захворювань або порушень, що характеризуються підвищеним або незбалансованим рівнем одного або декількох хемокінів, що включають IL-8, Gro-альфа, Groбета, Gro-гама, GCP-2 і ENA-78 людини, а саме, хронічного обструктивного захворювання легень (COPD), остеоартриту, ревматоїдного артриту, ерозійного артриту, астми, атеросклерозу, запального захворювання кишечнику, псоріазу, відторгнення трансплантату, подагри, раку, гострого ураження легень, гострого захворювання легень, сепсису, респіраторного дистрес-синдрому в дорослих (ARDS), захворювання периферичних артерій, системного склерозу, респіраторного дистрес-синдрому в немовлят, загострення астми і COPD, муковісцидозу, дифузійного панбронхіоліту, ураження, викликаного реперфузією, і/або ендометріозу. Незважаючи на те, що дійсніий винахід описаний, головним чином, стосовно до лікування захворювань або порушень у людини, дійсний винахід може бути також використаний для лікування аналогічних захворювань або порушень у ссавців, відмінних від людини. Конкретні варіанти здійснення винаходу Приклад 1 Одержання мишачого моноклонального антитіла 656.35, 197.2 і 81.1 Одержання пентаспецифічного моноклонального антитіла проти хемокінів-мішеней (IL-8, Gro-альфа, Gro-бета, Gro-гама і ENA-78 людини) Були використані різні методи і схеми імунізації мишей з метою створення панспецифічних моноклональних антитіл (mAb). Панспецифічні моноклональні антитіла були отримані з використанням різних сумішей множинних антигенних пептидів (МАР) і/або інтактних хемокінівмішеней (IL-8, Gro-α, -β, -γ і ENA-78), змішаних у повному або неповному ад'юванті Фрейнда (cFA або iFA), відповідно до зміненого протоколу повторної імунізації декількох сайтів (RIMMS) у лінії мишей SJL/JOrlCrl. МАР або множинні антигенні пептиди виконують дві функції в протоколі імунізації. По-перше, МАР забезпечують селективне множинне представлення відомої амінокислотної послідовностімішені. По-друге, викликають збільшення маси завдяки численним копіям послідовності, зв'язаним, наприклад, за допомогою лізинового ядра, що також підвищує імуногенність послідовності в порівнянні з окремими пептидами (Francis, J.P. et al., Immunology, 1991: 73; 249, Schott, M.E. et al., Cell. Immuno. 1996: 174:199-209, Tam, J.P. Proc. Natl. Acad. Sci. 1988: 85; 54095413). На фіг.1 представлене схематичне зображення набору МАР, що мають амінокислотні послідовності SEQ ID NO:49-53. Лінкер у МАР може бути будь-яким лінкером, крім лізинів. Загальна часова послідовність імунізації 21 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 55 60 Панспецифічні моноклональні антитіла були отримані відповідно до двох різних протоколів імунізації в зазначеній тимчасовій послідовності. 1. Первинна імунізація (0-й день) складалася з декількох підшкірних ін'єкцій (кінцівки, спина і шия) усіх хемокінів-мішеней, змішаних з cFA (по 10 мкг кожного). Нижченаведені 4 бустер-дози складалися із суміші всіх хемокінів-мішеней, змішаних в iFA (по 10 мкг кожного). П'ята і всі наступні бустер-дози складалися з коктейлю всіх хемокінів-мішеней і всіх 5 лінійних МАР (по 10 мкг кожного) в iFA. Кінцева бустер-доза, що вводиться за допомогою внутрішньочеревної (IP) ін'єкції за 3 дні до умертвіння і злиття, складалася з усіх хемокінів-мішеней і лінійних МАР в PBS. 2. Первинна імунізація (0-й день) складалася з декількох підшкірних ін'єкцій (кінцівки, спина і шия) усіх п'яти лінійних МАР, змішаних в cFA (по 10 мкг кожного). Наступні 4 бустер-дози складалися із суміші всіх п'яти лінійних МАР в iFA (по 10 мкг кожного). П'ята і всі наступні бустер-дози складалися з коктейлю всіх 5 лінійних МАР і всіх хемокінів-мішеней (по 10 мкг кожного) в iFA. Остання бустер-доза, що вводиться за допомогою внутрішньочеревної (IP) ін'єкції за 3 дні до умертвіння і злиття, складалася з усіх 5 лінійних МАР і всіх хемокінів-мішеней (по 10 мкг кожного) в PBS. Приклад 2 Панзв'язування моноклонального антитіла з хемокінами-мішенями (IL-8, Gro-альфа, Groбета, Gro-гама, GCP-2 і ENA-78 людини) було підтверджено за допомогою імунофлуоресцентного аналізу з флуоресцентним посиленням і роздільною здатністю за часом Кожний антиген (IL-8, Gro-альфа, Gro-бета, Gro-гама, GCP-2, ENA-78 людини і hIL-18, використовуваний у якості негативного контрольного зразка) окремо наносили на 96-лунковий імунофлуоресцентний планшет. Потім планшети промивали і блокували комерційно доступним блокуючим розчином. Блокуючий розчин видаляли, у кожну лунку додавали зразки, що містять моноклональне антитіло, та інкубували протягом 40-60 хвилин (протягом зазначеного часу моноклональне антитіло зв'язувалося з хемокінами-мішенями). Планшети знову промивали й у кожну лунку додавали детектуюче антитіло. У якості детектуючих реагентів була використана відповідно 2-кратна кількість антитіла 656.35, химерні (HcLc) і гуманізовані моноклональні антитіла: біотинільований IgG проти миші, біотинільований Fab2 IgG проти людини і IgG проти людини, оброблений європієм. Детектуюче антитіло було кон'юговане з європієм або біотином. Планшет знову промивали, щоб видалити незв'язане детектуюче антитіло, і в аналізовані зразки з біотином, що виявляється, додавали розчин стрептавідину-європію. Суміш стрептавідину-європію зв'язується з біотином, дозволяючи знайти зазначений реагент. Після виконання останнього промивання для видалення незв'язаного стрептавідину-європію або у випадку аналізу гуманізованого моноклонального антитіла після наступного промивання кон'югату з європієм усі лунки заповнювали розчином хелатуючого детергенту для активації флуоресценції, що продукується європієм. Більш сильна флуоресценція пропорційна кількості детектуючого антитіла, присутньої у лунці, яка пропорційна кількості хемокін-зв'язувального моноклонального антитіла. На фіг. 5А показане зв'язування мишачого моноклонального антитіла 656.35 з хемокінамимішенями людини. На фіг. 5В показане зв'язування химерного антитіла (HcLc) з хемокінами-мішенями людини. На фіг. 5С показане зв'язування гуманізованих моноклональних антитіл з хемокінамимішенями людини. Приклад 3 Функціональне панінгібування мишачим моноклональним антитілом було підтверджено 2+ різними методами: CXCR2-опосередкована мобілізація Са , хемотаксис нейтрофілів людини й активація нейтрофілів людини (поверхнева експресія CD11b) Аналіз мобілізації кальцію на титраційному мікропланшеті FLIPR (спектрофотометр для прочитування планшетів із флуорометричною візуалізацією, Molecular Devices, Sunnyvale, CA, [Schroeder, 1996] виконували для функціонального дослідження нейтралізуючого впливу антитіл 2+ на [Ca ]i-мобілізацію, індуковану ELR + хемокіном, у клітинах СНО-К1, трансфікованих hCXCR2 і Gα16 і стійко експресуючих зазначені речовини. За один день до аналізу клітини висівали на 96-лункові планшети з чорними стінками і світлою основою (Packard View) у кількості 40000 клітин на лунку. Через 18-24 години з лунок відсмоктували середовище і заміняли 100 мкл робочого середовища, що містить мінімальне есенціальне середовище Ігла (ЕМЕМ) із солями Ерла і L-глутаміном, 0,1 % BSA (Serologicals Corporation), 4 мкМ флуоресцентні індикаторні барвники на основі флуо-4-ацетоксиметилового ефіру (флуо-4 АМ) і 2,5 мМ пробенециду. Клітини інкубували в зазначеному середовищі, що містить барвник, протягом 1 години при 37С. Середовище, що містить барвник, відсмоктували і 22 UA 103157 C2 5 10 15 20 заміняли таким самим середовищем без флуо-4 АМ, що містить 0,1 % желатину (BSA видаляли) і 2,5 мМ пробенециду. Клітини інкубували протягом 10 хвилин при 37С і потім тричі промивали буфером для аналізу KRH [буфер Кребса–Рінгера-Хенселейта (120 мМ NaCl, 4,6 мМ KCl, 1,03 мМ КН2РО4, 25 мМ NaHCO3, 1 мМ CaCl2, 1,1 мМ MgCl2, 11 мМ глюкози, 20 мМ HEPES (рН 7,4)), що містить 0,1 % желатину і 2,5 мМ пробенециду]. Після останнього промивання буфером до клітин додавали 100 мкл буфера для аналізу KRH, що містить 0,1 % желатину і 2,5 мМ пробенециду, нагрівали до 37С протягом 10 хвилин і вводили в FLIPR, де клітини, пофарбовані барвником, піддавали впливові збудливого випромінювання (488 нм), 2+ створюваного аргонним лазером потужністю 6 Вт. [Ca ]i-мобілізацію вимірювали у вигляді 2+ збільшення інтенсивності випромінювання флуо-4, зв'язаного із Са , при 516 нм. Зміна 2+ інтенсивності випромінювання безпосередньо пов'язана з рівнями цитозольного кальцію [Ca ]i. Після вимірювання вихідного значення протягом 10 секунд у планшет додавали 50 мкл 3кратної кількості суміші ERL + хемокін, яку попередньо інкубували з антитілом у різних концентраціях, і реєстрували дані щосекунди протягом 1 хвилини, потім ще протягом півхвилини з інтервалами в 3 секунди. Максимальну клітинну реакцію, що була вище вихідного значення, використовували для побудови графіку в програмі GraphPad Prism (версія 4.03). Значення IC50 визначали у вигляді концентрації антитіла, необхідної під час попередньої обробки 3-кратною кількістю хемокіну в концентрації ЕС 80 для нейтралізації CXCR2опосередкованої стимулюючої дії суміші ELR + хемокін у концентрації ЕС 80 на 50 %. Вторинну клітинну реакцію на 25 мкм АТР вимірювали для дослідження життєздатності клітин [Sarau, 1999]. Інгібування індукованого сумішшю ERL + хемокін кальцієвого потоку в людини (геометрично усереднене значення IC50, мкг/мл) трьома мишачими моноклональними антитілами. (n=3-6) (n.d.=визначення не здійснювалося, N/A=дані відсутні) 3 нМ hIL-8 10 нМ hGROα Мишаче моноIC50 IC50 клональ95 % C.I. 95 % C.I. (мкг/мл) (мкг/мл) не антитіло 656.35 0,18 0,14-0,25 1,43 0,60-3,42 81.1 67,65 18,80-243,50 0,94 0,37-2,38 197.2 2,19 0,61-7,88 2,24 0,39-12,98 25 30 35 40 45 6 нМ hGROβ 30 нМ hENA-78 IC50 IC50 (мкг/ 95 % C.I. (мкг/ мл) мл) 1,06 n.d. n.d. 95 % C.I. 0,78-1,42 2,09 0,71-6,14 N/A 1,84 0,69-4,93 N/A 13,86 1,71-112,49 100 нМ hGCP-2 IC50 (мкг/ мл) 95 % C.I. 9,58 9,11 >157 1,17-78,57 1,44-57,61 N/A Schroeder KS, Neagle, BD. FLIPR: a new instrument for accurate, high throughput optical screening. J. Biomol. Screen. 1996:1-75. Sarau HM, Ames RS, Chambers J, Ellis C, Elshourbagy N, Foley JJ et al. Identification, molecular cloning, expression, and characterization of a cysteinyl leukotrien receptor. Mol Pharmacol. 1999;56:657-663. Інгібування стимульованого IL-8 хемотаксису нейтрофілів людини було також продемонстроване при використанні мікрокамери Бойдена. Мікрокамера Бойдена складається з двох невеликих камер, розташованих вище і нижче пористої мембрани товщиною 3 мікрони. У нижню камеру поміщають агент, що викликає хемотаксис, (тобто IL-8) або суміш хемокіну з пентаспецифічним моноклональним антитілом. У верхній камері знаходяться очищені неактивовані нейтрофіли людини. Після зборки камери виникає градієнт концентрацій з нижньої частини у верхню, стимулюючий рух нейтрофілів через мембрану до хемотаксину. Результати аналізу виражені у вигляді CI (хемотаксичного індексу), що являє собою відношення числа клітин, підданих хемотаксису під впливом стимулятора, до числа нестимульованих клітин. У процесі попередньої інкубації хемокіну (тобто IL-8) з пентаспецифічним моноклональним антитілом у нижній камері зазначене антитіло інгібувало залежно від дози стимульований IL-8 хемотаксис нейтрофілів. IL-8 у кількості 10 нм забезпечував досягнення CI у межах 3,6±0,8. Попередня інкубація IL-8 з пентаспецифічним моноклональним антитілом (656.35) у зростаючих концентраціях викликала інгібування залежно від дози хемотаксису нейтрофілів людини, при цьому статистично значиме значення було досягнуте при CI, що дорівнювало 2,2±0,6, при дозі 1 мкг/мл. Інші хемокіни (Gro-α, -β, -γ і ENA-78) не можна було досліджувати через використовувані кількості та чутливість аналізу. Інгібування активації очищених нейтрофілів людини шляхом вимірювання поверхневої експресії CD11b. CD11b або Мас-1 опосередковують адгезію до субстратів, агрегацію і хемотаксис, і, як відомо, їхній рівень підвищується на поверхні активованих нейтрофілів (Molad 23 UA 103157 C2 5 10 15 20 25 Y.J. et al., Clin. Immunol. Immunopathol. 1994: 71; 281-286). Неактивовані нейтрофіли людини очищають і стимулюють ex vivo хемокінами-мішенями (тобто IL-8) або хемокінами, попередньо інкубованими з пентаспецифічним моноклональним антитілом проти хемокінів. Дані виражені у вигляді процентного значення активації при максимальній поверхневій експресії CD11b у результаті стимуляції IL-8. Попередня інкубація IL-8 з моноклональним антитілом 656.35 дозволяла інгібувати залежно від дози підвищені рівні поверхневої експресії CD11b, що свідчить про інгібування активації нейтрофілів (79,9 %±3,7, 48,5 %±7,2, 28,7 %±3,2 і 31,6 %±3,4 відповідно в дозі 0,01, 0,1, 1, 10 і 50 мкг/мл). Приклад 4 Значення асоціації/дисоціації пентаспецифічних моноклональних антитіл для кожного хемокіна-мішені (IL-8, Gro-бета і ENA-78 людини) Методи аналізу Biacore Для виконання експерименту, іменованого KL, IgG-Fc кролика проти миші (Biacore BR-100514) імобілізують на чипі СМ5 шляхом зв'язування з первинним аміном відповідно до інструкцій виробників. Для виконання експерименту, іменованого ВЕ, очищені антитіла імобілізують на чипі шляхом зв'язування з аміном. Супернатант або очищене антитіло, виділене з батьківського мишачого моноклонального антитіла, уловлюють на поверхні IgG-Fc проти миші. Кожну аналізовану речовину (IL-8, Groбета, ENA-78) у визначених концентраціях пропускають над поверхнею імобілізованого або уловленого антитіла, при цьому для кожного упорскування аналізованої речовини використовують знову уловлене антитіло. Після кожного упорскування аналізованої речовини поверхню відновлюють, упорскуючи слабкий кислотний розчин, що видаляє уловлене антитіло, але не чинить значного впливу на здатність IgG-Fc проти миші виконувати інше уловлювання і не впливає на імобілізовані антитіла. На поверхню уловленого антитіла також упорскують буфер для подвійного контролю. Дані аналізують за допомогою програмного забезпечення для аналізу, призначеного для даного комп'ютера, використовуючи модель зв'язування у відношенні 1:1. IL-8 KL BE KL BE KL BE Моноклональне антитіло 656.35 81.1 197.2 Gro-B ENA-78 kd ka KD kd ka KD kd ka KD 1,48E-03 6,87E-04 9,32E-03 1,08E-01 3,69E-03 4,99E-04 1,53E+06 8,19E+06 1,95E+06 1,95E+06 6,27E+05 7,23E+05 9,64E-10 8,39E-11 4,80E-09 5,55E-08 5,89E-09 6,90E-10 2,01E-03 1,46E-03 3,12E-04 4,89E-04 9,93E-04 4,80E-08 3,82E+06 1,32E+07 2,76E+06 5,44E+06 1,55E+05 1,04E+06 5,26E-10 1,11E-10 1,13E-10 8,98E-11 6,39E-10 4,62E-14 1,87E-03 7,96E-04 3,76E-04 3,94E-06 2,69E-03 3,02E-03 1,07E+06 2,94E+06 6,11E+05 1,65E+05 2,99E+05 3,01E+05 1,74E-09 2,71E-10 6,16E-10 2,39E-11 9,00E-09 1,01E-08 30 35 40 45 50 Приклад 5 Картування епітопів Було зроблено картування епітопів моноклонального антитіла 656.35, при цьому було встановлено, що зазначене антитіло зв'язується з епітопом KELRCQCIKTYSKP (SEQ ID NO:54) у IL-8 людини. Таким чином, інший варіант здійснення дійсного винаходу відноситься до пентаспецифічного антитіла, що зв'язується з епітопом SEQ ID NO:54 IL-8 людини. Приклад 6 Дослідження ефективності химерного антитіла, отриманого з антитіла 656.35, що має послідовність важкого ланцюга SEQ ID NO:56 і послідовність легкого ланцюга SEQ ID NO:58. (Зазначене антитіло буде іменуватися химерним антитілом) Дослідження in vivo Для дослідження здатності пентаспецифічного антитіла інгібувати інфільтрацію нейтрофілів в легені приматів крім людини (NHP – циномолгус) була використана модель гострого запального захворювання легень, викликаного інгаляцією LPS. У приматів (NНР) обстежували стан здоров'я і сприйнятливість до екзогенно доданого IL-8 циномолгусу (циноIL-8). У відібраних приматів брали зразки для одержання вихідних даних (кров і бронхоальвеолярний лаваж – BAL) за п'ять днів до першої стимуляції LPS. У процесі стимуляції LPS приматів транквілізували за допомогою однієї внутрішньом'язової ін'єкції гідрохлориду кетаміну (~10 мг/кг). Після досягнення седативного ефекту приматів анестезували шляхом внутрішньовенного вливання пропофолу (~0,2 мг/кг/хв, за потреби). Анестезованих тварин поміщали на грілку-ковдру з циркулюючою теплою водою і/або обертали ковдрою з циркулюючим теплим повітрям і в кожне око вводили 24 UA 103157 C2 5 10 15 20 25 30 35 40 45 50 очну мазь. Потім тварин інтубували і штучно вентилювали легені для досягнення стимуляції. Для стимуляції використовували дихальний апарат з позитивним тиском і регульованим обсягом (дихальний апарат для котів/кроликів компанії Stoelting, www.stoeltingco.com № за каталогом 5019510). Стимуляцію ліпополісахаридом (LPS) виконували шляхом інгаляції за допомогою ультразвукового розпилювача Ultraneb-99 компанії DeVilbiss. Аерозоль LPS вводили протягом 5 хвилин у кількості 100 мкг/мл. Під час стимуляції контролювали і реєстрували частоту серцевих скорочень, температуру тіла і частоту подиху. Первинна стимуляція підтвердила, що примати реагують на LPS (підвищена інфільтрація нейтрофілів у легені та підвищені рівні циноIL-8). Індивідуальні реакції приматів були підтверджені зразками, отриманими через 6 і 24 години після стимуляції. Приматів (NHP), підданих первинній стимуляції LPS, довільно розподіляли в дві групи. Після відновлення протягом 4 тижнів у всіх приматів брали зразки крові і BAL. Через чотири дні після вимірювання вихідних показників піддослідним тваринам внутрішньовенно ін'єкували наповнювач або 1 мг/кг химерного антитіла. Наступного дня приматів знову стимулювали інгальованим LPS і через 6 і 24 години після стимуляції брали зразки для аналізу. Після додаткового періоду відновлення в приматів, яким вводили наповнювач, брали вихідні зразки і через 4 дні зазначеним приматам робили одну внутрішньовенну ін'єкцію ударної дози химерного антитіла в кількості 10 мг/кг. Наступного дня тварин стимулювали LPS і через 6 і 24 години брали зразки для аналізу. Інгаляція LPS являє собою модель гострого запального захворювання, що стимулює підвищення хемотаксичних хемокінів, таких як IL-8, що викликають підвищену інфільтрацію нейтрофілів у легені. Первинним параметром ефективності химерного антитіла є інгібування інфільтрації нейтрофілів у легені приматів, стимульованих LPS. Інфільтрація нейтрофілів у зазначеній моделі гострого запального захворювання відбувається протягом перших 24 годин після стимуляції LPS, після чого більш вираженими стають інші запальні процеси. Попереднє введення химерного антитіла значно й залежно від дози інгібує інфільтрацію нейтрофілів у легені приматів, стимульованих LPS. Введення химерного антитіла також запобігає підвищенню рівнів циркулюючих нейтрофілів, не впливаючи на дійсну функцію нейтрофілів (тобто здатність клітин до фагоцитозу). Введення химерного антитіла також не чинило сильного впливу на клітини інших типів, такі як макрофаги/моноцити, у легенях або кровотоці. Див. фіг. 2. Приклад 7 Подальші дослідження гуманізованих моноклональних антитіл Було отримано кілька конструкцій на основі гуманізованого пентаспецифічного антитіла, чотири з яких, як було показано, міцно зв'язуються з усіма хемокінами-мішенями та інгібують зазначені хемокіни. (Для ознайомлення з кожною послідовністю див. приведену нижче інформацію про послідовності). Зазначений аналіз складався з вимірювань спорідненості (BiaCore), аналізів мобілізації кальцію (функціональний аналіз in vitro, FLIPR) і аналізу стимуляції нейтрофілів CD11b людини і приматів (функціональний аналіз ex vivo, проточна цитометрія). BiaCore Кінетику гуманізованих і химерних конструкцій визначали методом уловлювання білка А, виконуваним у такий спосіб. Білок А імобілізують на чипі СМ5 шляхом зв'язування з первинним аміном відповідно до інструкцій виробника. Очищене гуманізоване або химерне антитіло уловлюють на поверхні білка А. Кожну аналізовану речовину (IL-8, Gro-β і ENA-78) у визначених концентраціях пропускають над поверхнею уловленого антитіла, виконуючи окреме уловлювання для кожного упорскування аналізованої речовини. Після кожного упорскування аналізованої речовини поверхню регенерують, упорскуючи слабкий кислотний розчин, що видаляє уловлене антитіло, але не чинить значного впливу на здатність білка А виконувати наступне уловлювання. На поверхню уловленого антитіла також упорскують буфер для подвійного контролю. Дані аналізують за допомогою програмного забезпечення для аналізу, призначеного для даного комп'ютера, використовуючи модель зв'язування у відношенні 1:1. 25 UA 103157 C2 Вимірювання з використанням Gro-бета Конструкція HcLc HcLc HcLc HcLc HcLc HcLc HcLc HcLc HcLc HcLc HcLc HcLc ka(1/Ms) 7,39E+06 2,11E+07 1,59E+07 1,19E+07 1,43E+07 1,33E+07 1,51E+07 1,49E+07 2,54E+07 1,81E+07 2,04E+07 1,82E+07 kd(1/s) 8,16E-04 0,0011 8,88E-04 6,16E-04 5,43E-04 6,76E-04 6,73E-04 6,44E-04 0,001163 0,001058 8,54E-04 5,06E-04 KD(M) 1,11E-10 5,21E-11 5,57E-11 5,17E-11 3,79E-11 5,09E-11 4,45E-11 4,33E-11 4,57E-11 5,85E-11 4,18E-11 2,78E-11 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 6,99E+06 1,76E+07 1,63E+07 1,14E+07 1,20E+07 1,42E+07 1,76E+07 1,63E+07 1,12E+07 2,63E+07 2,10E+07 9,66E-04 0,001305 0,001039 0,005616 0,004398 9,57E-04 0,00118 0,001389 0,008212 0,001546 9,28E-04 1,38E-10 7,40E-11 6,39E-11 4,93E-10 3,66E-10 6,73E-11 6,72E-11 8,52E-11 7,32E-10 5,88E-11 4,41E-11 H0L8 H0L8 H0L8 H0L8 H0L8 H0L8 1,28E+07 1,99E+07 1,19E+07 1,18E+07 1,61E+07 1,85E+07 0,005462 0,01065 0,007727 0,007799 0,006903 0,004586 4,27E-10 5,36E-10 6,52E-10 6,60E-10 4,29E-10 2,48E-10 H0L10 H0L10 H0L10 H0L10 H0L10 H0L10 1,23E+07 1,45E+07 1,44E+07 1,28E+07 1,98E+07 1,94E+07 0,001395 0,001261 0,001477 0,001394 0,00144 0,00109 1,13E-10 8,68E-11 1,03E-10 1,09E-10 7,28E-11 5,62E-11 H0M0 H0M0 H0M0 H0M0 H0M0 H0M0 H0M0 1,20E+07 1,13E+07 1,58E+07 1,43E+07 1,03E+07 1,85E+07 1,81E+07 0,001394 0,007071 0,001708 0,001829 0,009588 0,001728 0,001252 1,17E-10 6,24E-10 1,08E-10 1,28E-10 9,34E-10 9,35E-11 6,91E-11 26 UA 103157 C2 Вимірювання з використанням IL-8 Конструкція HcLc HcLc HcLc HcLc HcLc HcLc ka(1/Ms) 3,77E+06 1,03E+07 8,20E+06 1,04E+07 9,44E+06 9,97E+06 kd(1/s) 2,34E-04 2,87E-04 2,76E-04 2,68E-04 2,53E-04 2,30E-04 KD(M) 6,22E-11 2,78E-11 3,37E-11 2,58E-11 2,68E-11 2,31E-11 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 2,74E+06 8,99E+06 7,64E+06 5,98E+06 8,74E+06 7,51E+06 9,10E+06 2,84E-04 4,00E-04 2,84E-04 0,001167 3,52E-04 3,63E-04 3,89E-04 1,04E-10 4,45E-11 3,71E-11 1,95E-10 4,03E-11 4,83E-11 4,28E-11 H0L8 H0L8 H0L8 H0L8 5,90E+06 7,89E+06 6,80E+06 7,30E+06 0,00111 0,001126 0,001193 0,001112 1,88E-10 1,43E-10 1,75E-10 1,52E-10 H0L10 H0L10 H0L10 H0L10 6,86E+06 7,20E+06 7,21E+06 7,43E+06 4,57E-04 3,67E-04 3,96E-04 3,73E-04 6,66E-11 5,10E-11 5,48E-11 5,02E-11 H0M0 H0M0 H0M0 H0M0 H0M0 7,02E+06 6,40E+06 7,96E+06 7,97E+06 7,50E+06 3,98E-04 0,001467 4,09E-04 4,60E-04 4,18E-04 5,67E-11 2,29E-10 5,14E-11 5,77E-11 5,57E-11 Вимірювання з використанням ENA-78 Конструкція HcLc HcLc HcLc HcLc HcLc HcLc ka(1/Ms) 3,67E+06 5,81E+06 5,11E+06 5,28E+06 4,87E+06 4,78E+06 kd(1/s) 1,70E-04 1,71E-04 1,84E-04 1,54E-04 1,56E-04 1,23E-04 KD(M) 4,63E-11 2,94E-11 3,60E-11 2,91E-11 3,20E-11 2,58E-11 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 H0L7 2,95E+06 5,28E+06 4,59E+06 3,03E+06 5,11E+06 4,76E+06 4,91E+06 2,04E-04 2,97E-04 2,17E-04 3,33E-04 2,58E-04 3,07E-04 2,77E-04 6,93E-11 5,63E-11 4,74E-11 1,10E-10 5,04E-11 6,45E-11 5,63E-11 H0L8 H0L8 H0L8 H0L8 2,88E+06 2,82E+06 2,95E+06 3,29E+06 2,88E-04 2,66E-04 2,69E-04 2,48E-04 1,00E-10 9,41E-11 9,12E-11 7,55E-11 27 UA 103157 C2 H0L10 H0L10 H0L10 H0L10 4,07E+06 4,72E+06 4,20E+06 4,38E+06 3,66E-04 3,40E-04 3,12E-04 2,68E-04 8,99E-11 7,22E-11 7,42E-11 6,13E-11 H0M0 H0M0 H0M0 H0M0 H0M0 3,91E+06 2,97E+06 4,39E+06 4,18E+06 4,52E+06 3,09E-04 3,49E-04 3,16E-04 3,34E-04 2,94E-04 7,89E-11 1,17E-10 7,20E-11 7,99E-11 6,51E-11 Спорідненість (KD) визначають, досліджуючи швидкість асоціації (ka) і швидкість дисоціації (kd) білків. 5 10 15 20 25 30 35 HcLc означає химерне антитіло, отримане з антитіла 656.35, що має послідовність важкого ланцюга SEQ ID NO:56 і послідовність легкого ланцюга SEQ ID NO:58. Функціональний аналіз: мобілізація кальцію (FLIPR) з використанням CXCR2 (з Gα16) у клітинах СНО-К1 При виконанні аналізу мобілізації кальцію на титраційному мікропланшеті FLIPR (спектрофотометр для прочитування планшетів із флуорометричною візуалізацією, Molecular Devices, Sunnyvale, CA, [Schroeder, 1996]) був використаний для функціонального дослідження 2+ нейтралізуючого впливу антитіл на [Ca ]i-мобілізацію, індуковану ELR + хемокіном, у клітинах СНО-К1, трансфікованих hCXCR2 і Gα16 та стійко експресуючих зазначені речовини. За один день до аналізу клітини висівали на 96-лункові планшети з чорними стінками і світлою основою (Packard View) у кількості 40000 клітин на лунку. Через 18-24 години з лунок відсмоктували середовище і заміняли 100 мкл робочого середовища, що містить мінімальне есенціальне середовище Ігла (ЕМЕМ) із солями Ерла і L-глутаміном, 0,1 % BSA (Serologicals Corporation), 4 мкМ флуоресцентні індикаторні барвники на основі флуо-4-ацетоксиметилового ефіру (флуо-4 АМ) і 2,5 мМ пробенециду. Клітини інкубували в зазначеному середовищі, що містить барвник, протягом 1 години при 37С. Середовище, яке містить барвник, відсмоктували і заміняли таким самим середовищем без флуо-4 АМ, що містить 0,1 % желатину (BSA видаляли) і 2,5 мМ пробенециду. Клітини інкубували протягом 10 хвилин при 37С і потім тричі промивали буфером для аналізу KRH [буфер Кребса-Рінгера-Хенселейта (120 мМ NaCl, 4,6 мМ KCl, 1,03 мМ KН2РО4, 25 мМ NaHCO3, 1,0 мМ CaCl2, 1,1 мМ MgCl2, 11 мМ глюкози, 20 мМ HEPES (рН 7,4)), що містить 0,1 % желатину і 2,5 мМ пробенециду]. Після останнього промивання буфером до клітин додавали 100 мкл буферу для аналізу KRH, що містить 0,1 % желатину і 2,5 мМ пробенециду, нагрівали до 37С протягом 10 хвилин і вводили в FLIPR, де клітини, пофарбовані барвником, піддавали впливові збудливого випромінювання (488 нм), 2+ створюваного аргонним лазером потужністю 6 Вт. [Ca ]i-мобілізацію вимірювали у вигляді 2+ збільшення інтенсивності випромінювання флуо-4, зв'язаного із Са , при 516 нм. Зміна 2+ інтенсивності випромінювання безпосередньо пов'язана з рівнями цитозольного кальцію [Ca ]i. Після вимірювання вихідного значення протягом 10 секунд у планшет додавали 50 мкл 3кратної кількості суміші ERL + хемокін, яку попередньо інкубували з антитілом у різних концентраціях, і реєстрували дані щосекунди протягом 1 хвилини, потім ще протягом півхвилини з інтервалами в 3 секунди. Максимальну клітинну реакцію, що була вище вихідного значення, використовували для побудови графіку в програмі GraphPad Prism (версія 4.03). Значення IC50 визначали у вигляді концентрації антитіла, необхідної під час попередньої обробки 3-кратною кількістю хемокіну в концентрації ЕС 80 для нейтралізації CXCR2опосередкованої стимулюючої дії суміші ELR + хемокін у концентрації ЕС80 на 50 %. Вторинну клітинну реакцію на 25 мкМ АТР вимірювали для дослідження життєздатності клітин [Sarau, 1999]. 28
ДивитисяДодаткова інформація
Назва патенту англійськоюNovel compounds
Автори російськоюClegg, Stephanie, Jane, Dobrzynski, Eric, Ellis, Jonathan, H., Germaschewski, Volker, Godillot, Alexis, Paul, Jonar, Zdenka, Ludmila, Lewis, Alan, P., White, John, R.
МПК / Мітки
МПК: C07K 16/24, A61K 39/395, A61P 29/00
Мітки: антитіло, пентаспецифічне
Код посилання
<a href="https://ua.patents.su/61-103157-pentaspecifichne-antitilo.html" target="_blank" rel="follow" title="База патентів України">Пентаспецифічне антитіло</a>
Моноклональне антитіло, яке спричиняє апоптоз кров’яних клітин, що містять ядро та інтегрин-пов’язаний протеїн людини (іпп), фрагмент моноклонального антитіла, гібридома, яка продукує моноклональне антитіло (ва

Номер патенту: 72192
Опубліковано: 15.02.2005
Автори: Уно Шінсуке, Фукушіма Наоші
МПК: C12N 5/20, C12P 21/08, C07K 16/28
Мітки: апоптоз, містять, антитіло, гібридома, яке, ядро, іпп, моноклональне, клітин, спричиняє, моноклонального, фрагмент, ва, яка, антитіла, протеїн, інтегрин-пов'язаний, людини, продукує, кров'яних
Формула / Реферат:
1. Моноклональне антитіло, яке зв’язується із інтегрин-пов’язаним протеїном людини (ІПП) і спричиняє апоптоз кров'яних клітин, що містять ядро та ІПП.2. Моноклональне антитіло за п.1, яке відрізняється тим, що антитіло отримують способом, що включає:а) імунізацію тварин з використанням ІПП людини як сенсибілізуючого антигена,б) одержання гібридом;в) відбір антитіл, що розпізнають ІПП людини та спричиняють апоптоз...
Химерне антитіло проти cd40 людини, молекула нуклеїнової кислоти, вектор експресії, фармацевтична композиція для лікування захворювань, опосередкованих т-клітинами, яка містить химерне антитіло

Номер патенту: 71909
Опубліковано: 17.01.2005
Автори: Сайдек Ентоні В., Хьюз Уільям Д., Аруффо Аледжандро А., Уоткінс Джеффрі Д., Херріс Лінда Дж., Ву Херрен, Торн Барбара А., Беррі Карен К., Холленбау Даєн, Бейорет Юрген
МПК: A61P 43/00, A61P 19/02, C07K 14/725, C07K 16/28, A61P 37/02, C12N 1/21, C07K 14/46, A61K 39/395, C12N 15/09, A61P 37/06, A61P 29/00, C12N 15/13
Мітки: лікування, кислоти, молекула, нуклеїнової, людини, опосередкованих, фармацевтична, вектор, яка, експресії, композиція, захворювань, т-клітинами, химерне, містить, антитіло
Формула / Реферат:
1. Варіабельна ділянка легкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:1 (Фіг. 4а).2. Варіабельна ділянка важкого ланцюга химерного антитіла, що зв’язується з CD40 людини, яка містить амінокислотну послідовність SEQ ID NО:2 (Фіг. 4b).3. Химерне антитіло, що зв'язується з CD40 людини, яке містить легкий і важкий ланцюг, причому зазначений легкий ланцюг містить ...
Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент до рецептора ліпопротеїнів низької щільності (лпнщ) людини, спосіб їх одержання

Номер патенту: 78489
Опубліковано: 10.04.2007
Автори: Антонетті Франческо, Суісса Дані, Йонах Начум, Бельзер Ілана, Смоларскі Моше, Дреано Мішель
МПК: G01N 33/68, C07K 16/28, G01N 33/577, C12P 21/08, C12N 5/20
Мітки: ліпопротеїнів, спосіб, одержання, лпнщ, антитіло, моноклональне, щільності, фрагмент, людини, низької, антитіла, рецептора, антиідіотипічного, химерне, гуманізоване, повністю
Формула / Реферат:
1. Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент, що специфічно розпізнає і зв'язує розчинний рецептор ліпопротеїнів низької щільності (ЛПНЩ)людини і його фрагменти, а також здатне інгібувати реплікацію вірусу гепатиту С, і яке одержують шляхом імунізації тварин формою +291 розчинного рецептора ЛПНЩ людини, тобто формою розчинного рецептора людини, що містить...
Цитокіноподібний поліпептид-10 ссавців, полінуклеотид, що його кодує, антитіло та антиідіотипічне антитіло

Номер патенту: 73275
Опубліковано: 15.07.2005
Автори: Гросман Анжеліка, Хелдемен Бетті А., Конклін Деррел К.
МПК: A61P 35/00, A61P 11/00, C12N 15/09, A61K 48/00, C12N 15/19, C07K 16/42, C07K 16/24, A61P 37/02, A61P 43/00, A61K 38/00, A61P 17/00, A61P 1/02, A01K 67/027, C07K 14/52
Мітки: поліпептид-10, полінуклеотид, кодує, антитіло, антиідіотипічне, цитокіноподібний, ссавців
Формула / Реферат:
1. Виділений полінуклеотид, який кодує поліпептид, цей поліпептид щонайменше на 80% ідентичний одному чи більше з поліпептидів, які вибрані з групи, яка складається з ПОСЛІДОВН. №2, ПОСЛІДОВН. №4, ПОСЛІДОВН. №12, ПОСЛІДОВН. №13, ПОСЛІДОВН. №19, ПОСЛІДОВН. №20, ПОСЛІДОВН. №25, ПОСЛІДОВН. №26, ПОСЛІДОВН. №34 та ПОСЛІДОВН. №35, або виділений полінуклеотид, який кодує поліпептид, який вибраний з групи, яка складається з ПОСЛІДОВН. №14, ПОСЛІДОВН....
Антитіло зі специфічною афінністю до характерної антигенної детермінанти ed-b домену фібронектину, його застосування для лікування ангіогенезу, кон’югати та діагностичний набір, до складу якого входить вказане

Номер патенту: 74134
Опубліковано: 15.11.2005
Автори: Бірхлер Манфред, Віті Франческа, Тарлі Лоренцо, Нері Даріо
МПК: A61K 47/48, A61K 49/00, C12N 15/09, C07K 16/18
Мітки: характерної, антигенної, афінністю, застосування, вказане, специфічною, антитіло, входить, складу, кон'югати, домену, детермінанти, лікування, діагностичний, фібронектину, якого, набір, ангіогенезу
Формула / Реферат:
1. Антитіло зі специфічною афінністю до характерної антигенної детермінанти ED-B домену фібронектину, де згадане антитіло має афінність до згаданої ED-B антигенної детермінанти у субнаномолярному діапазоні.2. Антитіло за п. 1, яке розпізнає ED-B(+) фібронектин.3. Антитіло за п. 1, яке має scFv формат.4. Антитіло за п. 3, яке є рекомбінантним.5. Антитіло за п. 3, де згадана афінність поліпшена шляхом індукування...
Попередній патент: Вдосконалена дерев’яна бочка
Наступний патент: Кон’югат білок-полімер та спосіб лікування за його допомогою
Випадковий патент: Акустична система