Комбінована вакцина проти кору та малярії
Номер патенту: 104180
Опубліковано: 10.01.2014
Автори: Глюек Реінхард, Гіанно Вівіана, Біллетер Мартін, Фазіо Агата







Формула / Реферат
1. Комбінована вакцина проти кору та малярії, яка включає рекомбінантну вірусну вакцину кору, що експресує антигени малярії, здатні викликати імунну відповідь та захист проти як кору, так і малярії.
2. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує один або різні антигени малярії.
3. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії.
4. Комбінована вакцина проти кору та малярії за п. 1, у якій антиген MSP1 малярії становить від 190 до 200 кДа (d190).
5. Комбінована вакцина проти кору та малярії за п. 1, у якій антигеном MSP1 малярії є d83-30-38.
6. Комбінована вакцина проти кору та малярії за п. 1, у якій антигеном MSP1 малярії є d42.
7. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах.
8. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах штаму 3D7 та MAD 20, краще, штаму FCB1.
9. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах штаму FCB1.
10. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує Diversity Covering (DiCo) антигену малярії ΑΜΑ1.
11. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-1 антигену малярії АМА1.
12. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-2 антигену малярії АМА1.
13. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-3 антигену малярії АМА1.
14. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-1, DiCo-2 та DiCo-3 антигену малярії АМА1.
15. Комбінована вакцина проти кору та малярії за будь-яким з пп. 10-14, у якій рекомбінантна вірусна вакцина кору експресує Diversity Covering (DiCo) антигену малярії АМА1 у трансмембранній та секретованій формах.
16. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує антиген малярії CS.
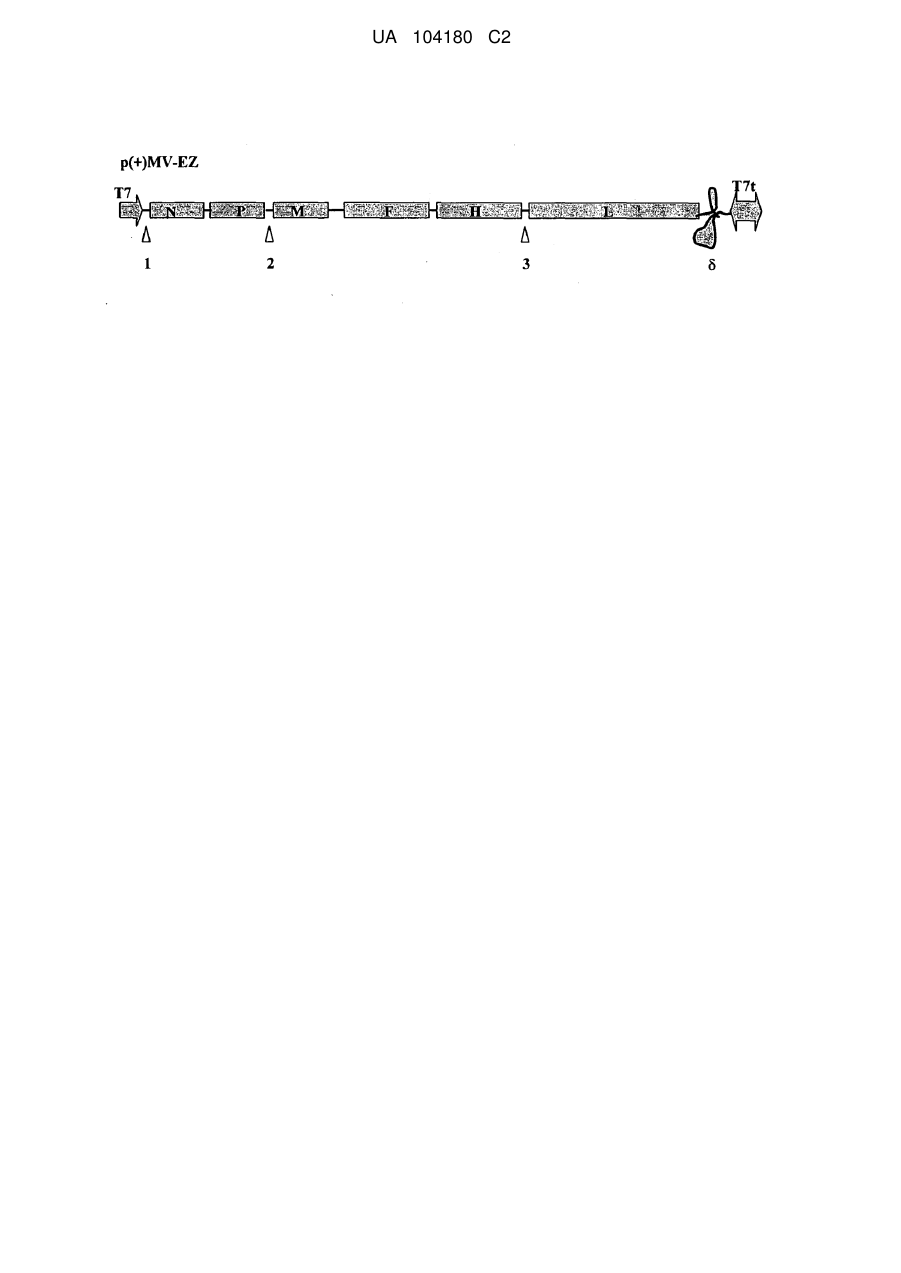
17. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій антиген малярії клонований між білками Ρ та Μ або Η та L рекомбінантної вірусної вакцини кору.
18. Вектор вірусної вакцини кору, що містить нуклеотидну послідовність антигену малярії з будь-якого з пунктів 1-17 формули винаходу.
19. Вектор за п. 18, у якому нуклеотидну послідовність вибирають з наведених SEQ ID NO 3-12.
20. Вектор за будь-яким з пп. 18 та 19, який додатково включає нуклеотидну послідовність, вибрану з наведених SEQ ID NO 1-2.
21. Вектор за п. 18, у якому нуклеотидна послідовність кодує антигени малярії, вибрані з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS.
22. Хазяїн, який включає вектор за п. 18.
23. Хазяїн за п. 22, вибраний з Е. coli або клітинної лінії ссавця.
24. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантний вірус кору походить від штаму вакцин, одержаного з Edmonston Zagreb.
25. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантну вірусну вакцину кору, що експресує щонайменше один антиген малярії, вибирають з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS.
26. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантну вірусну вакцину кору, що експресує два чи більше антигенів малярії, вибирають з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS або їх комбінації.
27. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору включає послідовності SEQ ID NO 3-12.
28. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору кодує, на додаток до антигенів малярії, білок з ад'ювантними властивостями.
29. Вакцина за будь-яким з попередніх пунктів формули, яка включає рекомбінантні віруси кору, що кодують, на додаток до антигенів малярії, інтерлейкін, краще інтерлейкін 2.
30. Вакцина за будь-яким з попередніх пунктів формули, що включає один з описаних рекомбінантних вірусів кору-малярії або суміш від двох до декількох таких вірусів.
31. Вакцина за будь-яким з попередніх пунктів формули, у якій описані рекомбінантні віруси кору-малярії або суміш від двох до декількох таких вірусів позбавлені дефектних інтерферуючих частинок (DI).
32. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють методом очищення бляшок.
33. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють методом кінцевого розведення.
34. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють фізичними методами, такими як диференційне центрифугування.
35. Вакцина за будь-яким з попередніх пунктів формули, яка є компонентом комбінованої вакцини, де іншими компонентами є краснуха, епідемічний паротит, вітряна віспа або інша жива атенуйована вірусна вакцина, природно атенуйована або рекомбінантна, сама або в комбінації.
36. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор, такий як желатин та/або альбумін сироватки людини і сорбіт як основні компоненти, ідеально, для парентерального застосування.
37. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для інтраназального застосування.
38. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для застосування шляхом інгаляції.
39. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для перорального застосування.
40. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для трансдермального застосування.
41. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для будь-якої композиції супозиторія.
42. Склад комбінованої вакцини проти кору-малярії, як заявлено в будь-якому попередньому пункті, що містить стабілізатор та/або ад'ювант.
Текст
Реферат: Винахід стосується комбінованої вакцини проти кору та малярії, яка включає різні атенуйовані рекомбінантні вектори кору-малярії, що містять гетерологічну нуклеїнову кислоту, яка кодує декілька антигенів Plasmodium falciparum; вірусного вектора, який включає нуклеїнову кислоту, що кодує циркумспорозоїтний (CS) білок P. falciparum, мерозоїтний поверхневий білок 1 (MSP1) P. falciparum, або його похідні (р-42; р-83-30-38) в глікозилованій та секретованій формах, або антиген апікальної мембрани 1 (АМА1) P. falciparum у закріпленій або секретованій формі. UA 104180 C2 (12) UA 104180 C2 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Область техніки Даний винахід стосується комбінованої вакцини проти кору та малярії, яка містить різні атенуйовані рекомбінантні вектори кору-малярії, що включають гетерологічну нуклеїнову кислоту, яка кодує декілька антигенів Plasmodium falciparum. Краще, він стосується вірусних векторів, які включають нуклеїнові кислоти, що кодують циркумспорозоїтний (circumsporozoite) (CS) білок P. falciparum, мерозоїтний (merozoite) поверхневий білок 1 (MSP-1) P. falciparum, та його похідні (p-42; p-83-30-38) у глікозильованій та секретованій формах, та антиген апікальної мембрани 1 (AMA1) P. falciparum, у його закріпленій (anchored) або секретованій формі. Вірусний вектор походить з атенуйованого вірусу кору, на основі штаму, що використовується як вакцина та є ефективним для доставки бажаного гена, і зв’язується з та ефективно інфікує релевантні імунні клітини. У кращому варіанті втілення, білки CS, MSP1 та AMA1 одержують з вірусу так, щоб вони викликали сильну імунну відповідь у ссавців, краще, у людини; експресія білків підвищується завдяки оптимізації кодонів для людини. Крім того, винахід стосується використання рекомбінантної вакцини у профілактичному лікуванні малярії. Відомий рівень техніки Вірус кору Винахід стосується вакцини, що містить рекомбінантні атенуйовані віруси кору, які експресують антигени Plasmodium falciparum (Pf), та їх використання для виготовлення рекомбінантної вакцини проти кору та малярії, яка буде створювати імунітет проти антигенів як кору, так і малярії. Вірус кору (MV) є членом порядку Мононегавірусів (Mononegavirales), тобто, вірусів з несегментованим РНК геномом з негативним ланцюгом. Несегментований геном MV має антисмислову (antimessage) полярність; отже, геномна РНК не транслюється in vivo або in vitro. Крім цього, вона є біологічно активною тільки коли вона дуже специфічно асоційована з трьома вірусними білками у формі рибонуклеопротеїнового (RNP) комплексу (див. нижче). Транскрипція та реплікація несегментованих РНК вірусів з (-)-ланцюгом та їх складання у вірусні частинки було детально описано (1). Транскрипція та реплікація вірусу кору не зачіпає ядро інфікованих клітин, а відбувається у цитоплазмі інфікованих клітин. Геном вірусу кору включає гени, які кодують шість великих структурних білків з шести генів (позначених N, P, M, F, H та L), і додатково два неструктурних білка, що походять з P-гена - C та V - які беруть участь в протидії конститутивним імунним відповідям та в регуляції транскрипції/реплікації. Гени розташовані у порядку 3' N, P (включаючи C та V), M, F, H та L 5'. Крім того, з 3'-кінцевої ділянки транскрибується коротка лідерна РНК довжиною приблизно 50 нуклеотидів. Названі гени відповідно кодують білки рибонуклеокапсиду (RNP) вірусу, тобто, нуклеопротеїн (N), фосфопротеїн (P) та великий білок полімерази/реплікази (L), який дуже щільно асоційований з геномною РНК, утворюючи RNP. Інші гени кодують білки вірусної оболонки, включаючи гемаглютинін (H), злитий (F) та матричний (M) білки. Транскрипція генів MV відбувається з низхідним градієнтом: при взаємодії полімерази з геномною матрицею вона синтезує більше РНК з генів, розташованих ближче до початку ланцюга, ніж з далі розташованих генів. В режимі такої періодичної транскрипції відбувається кепіювання та поліаденілювання мРНК. Навпаки, в режимі реплікації, L-білок продукує повнорозмірну антигеномну та геномну РНК, які негайно покривають N, P та L білки з утворенням RNP інфекційного потомства. Вірус кору був виділений в 1954 р.: Enders and Peebles інокулювали первинні клітини нирки людини кров’ю, що належала David Edmonston (Edmoston) - дитині, ураженій кором, і одержаний штам Edmonston MV (2) був згодом адаптований для вирощування у різноманітних клітинних лініях. Адаптація до курячих ембріонів, фібробластів курячих ембріонів (CEF) та/або клітин нирки собаки та людських диплоїдних клітин дозволила одержати атенуйований засівний матеріал Edmonston A та B (3), Zagreb (EZ) та AIK-C. Edmonston B був ліцензований у 1963 р. як перша вакцина MV. Подальші пасажи Edmonston A та B на CEF дали більш атенуйовані віруси Schwarz and Moraten (3), ідентичність послідовностей яких було недавно продемонстровано (4; 5). Оскільки вакцина Edmonston B була реактогенною, її використання припинили у 1975 р. та замінили на вакцину Schwarz/Moraten. Використовуються також декілька інших штамів вакцин: AIK-C, Schwarz F88, CAM70, TD97 у Японії, Leningrad-16 у Росії, та Edmonston Zagreb. Китайські штами CAM70 та TD97 були одержані не з Edmonston. Вакцини Schwarz/Moraten та AIK-C продукують на CEF. Вакцину Zagreb продукують на людських диплоїдних клітинах (WI-38). Зараз звичайно застосовуються вакцини Schwarz/Moraten, AIK-C та EZ (6), але у принципі було продемонстровано, що будь-який з цих атенуйованих штамів вакцин, які усі належать до одного унікального серотипу MV, є безпечними та індукує тривалі імунні відповіді, можуть бути використані в цілях винаходу. 1 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Вакцини MV індукують довічний імунітет після однієї або двох ін’єкцій малими дозами. Захист проти кору медіюється як антитілами, так і T-клітинами CD4 та CD8. Була продемонстрована персистенція MV-специфічних антитіл та клітин CD8 на протязі 25 років після вакцинації (7). Вакцина MV є простою у виробництві у великих масштабах в більшості країн та може розповсюджуватися з низькими витратами. Завдяки тому, що атенуація геному MV спричинена вдалою комбінацією численних мутацій, вакцина є дуже стабільною і повернення до патогенності ніколи не спостерігалося (6). Що стосується безпеки, то MV реплікується виключно у цитоплазмі, виключаючи можливість інтеграції в ДНК хозяїна. Такі характеристики роблять живу атенуйовану вакцину MV приваблювим кандидатом для використання як вектор для багатовалентної вакцинації. Така вакцина може виявитися ефективною для створення тривалого імунного захисту проти інших патогенних агентів, так само як і проти власне вірусного вектора. Martin Billeter зі співробітниками клонували кДНК, що відповідає антигеному Edmonston MV, та розробили оригінальну та ефективну процедуру зворотної генетики для рятування (rescue) вірусу (8), яка описана у міжнародній патентній заявці WO 97/06270. Рекомбінантний вірус кору виділяють з лінії клітин-помічників 293-3-46, стабільно трансфекованих та експресуючих білки N та P MV, а також РНК-полімеразу бактеріофага T7. Для рятування будь-якого варіанта або рекомбінантного MV, лінію клітин-помічників потім транзієнтно трансфекують експресійною плазмідою, яка кодує L-білок та, найважливіше, будь-якою антигеномною плазмідою, належно сконструйованою для створення будь-якої мутованої або рекомбінантної антигеномної РНК, сумісною для утворення потомства MV. Стадія транзієнтної трансфекції приводить спочатку до транскрипції, краще, резидентною РНК-полімеразою T7. Одержана антигеномна РНК негайно (in statu nascendi) покривається вірусними білками N, P та L з утворенням антигеномного RNP, з якого продукується геномний RNP. По-друге, геномний RNP транскрибується приєднаним L, з утворенням усіх вірусних мРНК та відповідних білків. Зрештою, як геномний, так і антигеномний RNP ампліфікують реплікацією. У невеликому варіанті цієї процедури, замість використання стабільно трансфекованих 2933-46 клітин-помічників, транзієнтно трансфекували комерційно доступні клітини 293T, використовуючи одночасно усі 5 плазмід, описані у вихідному описі патенту, що кодують N, P та T7 полімеразу (раніше використані для створення лінію клітин-помічників), а також плазміду, яка кодує L, та антигеномну плазміду. Відзначимо, що у процедурі "повністю транзієнтної трансфекції" можна також використовувати варіант експресійних плазмід та цілком уникнути використання РНК полімерази T7, застосовуючи замість неї резидентну РНК полімеразу II для експресії також L-білка та антигеному (9). Для рятування індивідуальних рекомбінантних MV використовувані антигеномні плазміди включали кДНК, яка кодує повнорозмірну антигеномну (+)РНК вірусу кору, рекомбіновану з нуклеотидними послідовностями, що кодують бажаний гетерологічний антиген (гетерологічну нуклеотидну послідовність), фланкований MV-специфічними послідовностями початку та термінації транскрипції, з утворенням при цьому додаткових одиниць транскрипції (ATU). Цей вектор штаму MV Edmonston був розроблений винахідниками, які винайшли рятування MV для експресії чужорідних генів (10), і продемонстрував свою велику придатність для інсерцій (аж до 5 тпн) та високу стабільність при експресії трансгенів (11; 12), таких як поверхневий антиген вірусу гепатиту B, віруси імунодефіциту мавп або людини (SIV або HIV), вірус епідемічного паротиту, та IL-12 людини. Зокрема, на ранніх етапах були продуковані рекомбінантні віруси кору, що експресують поверхневі та корові антигени вірусу гепатиту B індивідуально або в комбінації, і було продемонстровано, що вони індукують гуморальні імунні відповіді у генетично модифікованих мишей. На основі спостережень властивостей вірусу кору, і особливо його здатності викликати високі титри нейтралізуючих антитіл in vivo та його здатності бути сильним індуктором тривалої клітинної імунної відповіді, автори винаходу припустили, що він може бути гарним кандидатом для продукування рекомбінантних вірусів, що експресують антигени P. falciparum, для індукування нейтралізуючих антитіл проти зазначеного малярійного паразита, які краще можуть бути придатними для досягнення щонайменше певного ступеня захисту у хазяїв-тварин і, ще краще, у людини. Так, штами MV і, зокрема, штами вакцин були вибрані в даному винаході як векторикандидати для індукування імунітету проти як вірусу кору, так і паразита P. falciparum, компонент якого експресується в сконструйованому рекомбінантному MV, в експонованих дитячих популяціях, тому що вони не мають імунітету до MV. 2 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 Однак, дорослі популяції, навіть вже MV-імунізовані особи, також можуть одержати користь від імунізації рекомбінантним MV, тому що повторне введення вірусу MV в рекомбінантній формі за даним винаходом спричинює бустинг анти-MV антитіл (13). Винахід стосується, зокрема, одержання рекомбінантних вірусів кору, які несуть гетерологічні гени паразитів P. falciparum. Кращі імунологічні властивості рекомбінантних вірусів кору відповідно до винаходу можуть бути продемонстровані на тваринній моделі, яку вибирають з тварин, сприйнятливих до вірусів кору, та у якій визначають гуморальну та/або клітинну імунну відповідь проти гетерологічного антигену та/або проти вірусу кору. З таких тварин, придатних для використання як модель для характеризації імунної відповіді, кваліфікований фахівець може, зокрема, використовувати трансгенних мишей, що експресують CD46, один із специфічних рецепторів MV. Найбільш перспективні рекомбінантні продукти можуть згодом бути випробувані на мавпах. Нуклеотидна послідовність рекомбінантного вірусу кору повинна включати повне число нуклеотидів, яке є кратним шести. Вимога дотримання цього так званого "правила шести" є абсолютно необхідною не лише для MV, але для усіх вірусів, що належать до підродини Paramyxovirinae. Очевидно, молекули N-білка, кожна з яких контактує з шістьма нуклеотидами, мають точно покривати геномну та антигеномну РНК від 5'- до 3'-кінця. Слід відзначити, що положення ATU по довжині антигеномної кДНК може змінюватися. Таким чином, використовуючи природний градієнт експресії мРНК MV, згаданий вище, можна змінювати рівень експресії вставлених ATU до потрібних рівнів. Кращими положеннями ATU є зліва (upstream) від L-гена, зліва від M-гена та зліва від N-гена, які приводять до низької, середньої та сильної експресії, відповідно, гетерологічних білків. Малярійний паразит Малярія зараз є однією з найбільш розповсюджених інфекційних хвороб у світі, особливо, в тропічних та субтропічних регіонах. Щорічно, малярійні інфекції спричинюють тяжкі захворювання у сотень мільйонів людей в усьому світі, вбиваючи від 1 до 3 мільйонів, переважно, дітей молодшого віку у молодих країнах та країнах, що розвиваються. Поширеність та підвищена захворюваність на малярію є наслідком розповсюдженої заборони застосування ДДТ та зростання кількості резистентних до лікарських засобів паразитів, а також резистентних до інсектицидів переносників паразитів. Інші фактори включають екологічні та кліматичні зміни, громадянські заворушення та підвищену мобільність популяцій. Збудниками малярії є гематопротозойні паразити, що переносяться москітами, які належать до роду Plasmodium типу Apicomplexa. Чотири види роду Plasmodium інфікують людину: P. malariae, який викликає чотириденну малярію, P. vivax та P. ovale, які обидва викликають триденну малярію, і P. falciparum, патоген тропічної малярії, що відповідає за майже усі смертельні інфекції. Багато інших видів викликають хвороби тварин, такі як P. yoelii та P. berghei у мишей. Малярійні паразити мають життєвий цикл, що складається з декількох стадій. Кожна стадія є здатною індукувати специфічні імунні відповіді, спрямовані проти відповідних присутніх стадіяспецифічних антигенів. Малярійні паразити переносяться до людини самками декількох видів москітів Anopheles. Інфіковані москіти вносять "спорозоїтну" форму малярійного паразита в кровотік ссавця. Спорозоїти протягом декількох хвилин залишаються в циркуляції, після чого заражають гепатоцити. На цій стадії, паразит знаходиться в позаклітинному середовищі та зазнає атак антитіл, переважно націлених на "циркумспорозоїтний" (CS) білок, основний компонент поверхні спорозоїтів. Потрапивши до печінки, паразити реплікуються та розвиваються в так звані "шизонти". Ці шизонти є присутніми в кількості до 20000 на інфіковану клітину. На цій внутрішньоклітинній стадії паразита, головними гравцями імунної відповіді хозяїна є T-лімфоцити, особливо CD8+ T-лімфоцити. Після приблизно одного тижня інфекції печінки, тисячи так званих "мерозоїтів" вивільняються в кровотік. Антиген апікальної мембрани 1 (AMA1) та мерозоїтний поверхневий білок 1 (MSP1) обидва присутні на мерозоїтах, які вивільняються з інфікованих клітин печінки: вони є суттєвими компонентами безстатевих мерозоїтів стадії кровотоку, які відповідають за інвазію еритроцитів. Після потрапляння в червоні кров’яні клітини вони робляться мішенями антитіло-медійованої імунної відповіді та секретованих T-клітинами цитокінів. Після інвазії еритроцитів, мерозоїти проходять декілька стадій реплікації, утворюючи так звані "трофозоїти" та шизонти і мерозоїти, які можуть інфікувати нові червоні кров’яні клітини. Обмежена кількість трофозоїтів може розвиватися в "гаметоцити", які є статевою стадією розвитку паразита. При потраплянні еритроцитів до сприйнятливих москітів, гаметоцити вивільняються з еритроцитів, утворюючи декілька чоловічих гаметоцитів та один жиночий гаметоцит. Запліднення цих гамет приводить до утворення зиготи та подальшого перетворення на оокінети, потім на ооцисти, і зрештою на спорозоїти слинної 3 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 залози. Націлювання антитіл на специфічні для гаметоцитної стадії поверхневі антигени може блокувати цей цикл у середній кишці москіта. Такі антитіла не захищають ссавця-хазяїна, але будуть зменшувати перенесення малярії шляхом зменшення кількості інфікованих москітів та їх паразитарного навантаження. MSP-1 синтезується у вигляді 190-200 кДа (d-190) прекурсора, який протеолітично процесується на фрагменти 83, 30, 38 та 42 кДа (d-42) під час шизогонії (14). В момент еритроцитарної інвазії 42-кДа далі розщеплюється з утворенням 33 кДа фрагмента, який відкидається з рештою комплексу, та 19 кДа фрагмента, який містить два епідермальний фактор росту (EGF)-подібних домени, що залишаються асоційованими з мерозоїтною мембраною під час інвазії. Це вторинне розщеплення є передумовою успішної інвазії еритроцитів (15). MSP-1 є по суті диморфним білком з високим ступенем консерватизму у диморфних алелях, що характеризуються прототипами K1 та MAD20. AMA-1 (16) є структурно консервативним інтегральним мембранним білком типу I, який у P. falciparum містить 622 aa (амінокислот) (PfAMA-1), організовані в цитозольну ділянку (50 aa), трансмембранну ділянку та ектодомен, який укладається в N-кінцеву пропослідовність та три домени (DI, DII, DIII). Експресія білка є максимальною у пізній шизогонії: прекурсор AMA-1 (83 кДа) протеолітично процесується з відщепленням пропослідовності, перетворюючи білок на 66 кДа форму, яка дозволяє релокалізацію мерозоїтів. Антитіла розпізнають переважно Dl та DII і, певно, однаково гарно реагують з декількома алельними варіантами. Реакція антитіл на DIII є звичайно слабкою, причому у дорослих рівні зростають (17, 18). PfAMA-1 містить 64 поліморфні положення (9 у пропослідовності, 52 у ектодомені, 3 у цитозольній ділянці), більшість з них є диморфними, які є важливими епітопами для імунних відповідей. Для розробки вакцин на основі PfAMA-1 важливо охопити (to cover) поліморфізми: Diversity Covering (DiCo 1, 2 та 3) PfAMA-1 є штучними послідовностями, які представляють, в максимально можливому ступені, природний поліморфізм ектодомену PfAMA1. Було продемонстровано, що вони індукують імунні відповіді, які є функціональними проти ряду паразитів, що несуть різні алелі PfAMA1. Цей підхід може забезпечити засоби, за допомогою яких можуть продукуватися вакцини, націлені на PfAMA1, так щоб міг бути індукований сильний та функціональний захист від широкого спектра природних алелей PfAMA1 (19). CS-білок (CSP) містить приблизно 420 aa та має молекулярну вагу 58 кДа. Він є основним поверхневим білком спорозоїтів: його функція є фундаментальною для визрівання спорозоїтів з ооцист та для інвазії гепатоцитів, яка медіюється консервативним мотивом позитивно заряджених амінокислот. CSP організований в дві неповторювані ділянки на 5'- та 3'-кінцях, та варіабельну вид-специфічну центральну ділянку, що складається з численних повторів мотивів довжиною в чотири залишки, які утворюють головний епітоп CSP. Оскільки CSP залишається детектованим протягом щонайменше перших 3 днів шизогонії, він вважається привабливою мішенню для вакцини як для антитіло-медійованої імунної відповіді, спрямованої проти позаклітинних спорозоїтів, так і для клітинно-медійованих імунних відповідей, спрямованих проти шизонтів (20). Сучасні підходи до розробки вакцин проти малярії можуть бути класифіковані відповідно до різних стадій, в яких може існувати паразит, як описано вище. Можна виділити три типи можливих вакцин: i) до-еритроцитарні вакцини, які спрямовані проти спорозоїт- та/або шизонт-інфікованих клітин. Ці типи вакцини основані переважно на CS та повинні, ідеально, створювати стерильний імунітет, медійований гуморальною та клітинною імунними відповідями, запобігаючи малярійній інфекції; ii) вакцини безстатевої кров’яної стадії, які спрямовані проти мерозоїт-інфікованих клітин: MSP1 та AMA1 є головними кандидатами малярійних вакцин, розроблених для мінімізації клінічної тяжкості. Ці вакцини мають зменшувати захворюваність та смертність і повинні перешкоджати потраплянню паразитів до та/або їх розвитку в еритроцитах; iii) вакцини, що блокують перенесення, які призначені для перешкоджання розвитку паразитів в москіті-хазяїні. Цей тип вакцини повинен сприяти зменшенню зараженості популяції малярією. Крім цих вакцин, можливість розробки малярійних вакцин, націлених на численні стадії життєвого циклу паразита, використовується у так званих багатокомпонентних та/або багатостадійних вакцинах. На сьогодні глобальний портфель малярійних вакцин виглядає обнадійливо з 47 новими вакцинами-кандидатами, з яких 31 знаходяться на стадії доклінічної розробки, і лише 16 - на стадії клінічних випробувань. Одна з них, вакцина RTS,S, яку розробляють GSK Biologicals та PATH-MVI, має перейти на кінцеву стадію III клінічних випробувань у 2008 р. (21). Іншими цікавими вакцинами-кандидатами є такі, що основані на живих рекомбінантних вірусах, використовуваних як вектор, таких як модифікований вірус коров’ячої віспи Ankara (MVA), 4 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 описаний у міжнародній патентній заявці US2006127413, поксвірус (US6214353, AU7060294, AU1668197, WO9428930 та US5756101), аденовірус (US2007071726, US2005265974, US2007088156 та CA2507915), холодоадаптований атенуйований вірус грипу, або основані на дріжджах, таких як Pichia pastoris та Saccharomyces spp., або на бактеріальних системах експресії, таких як Salmonella spp. (US5112749) та Escherichia coli (EBO 191748) (22). Зараз немає комерційно доступної вакцини проти малярії, хоча розробка вакцин проти малярії була розпочата більш ніж 30 років назад. Багато факторів роблять розробку малярійної вакцини складною та важкою. По-перше, розмір та генетична складність паразита означає, що кожна інфекція презентує імунній системі людини тисячі антигенів. Важко зрозуміти, які з них можуть бути корисною мішенню для розробки вакцини, і на сьогодні було ідентифіковано щонайменше 40 різних перспективних антигенів. По-друге, зміни паразита на протязі декількох стадій життя, навіть у людині-хазяїні, презентують імунній системі, на кожній стадії життєвого циклу, різні підгрупи молекул. По-третє, паразит виробив ряд стратегій, які дозволяють йому заплутувати, ховати та спрямовувати на невірний шлях імунну систему людини. Зрештою, можна одночасно мати множинні малярійні інфекції не лише різних видів, але також і різних штамів. Отже, даний винахід задовольняє давню потребу відомого рівня техніки шляхом забезпечення комбінованої вакцини проти кору та малярії, яка містить різні атенуйовані рекомбінантні вектори кору-малярії, що включають гетерологічну нуклеїнову кислоту, яка кодує декілька антигенів Plasmodium falciparum. Суть винаходу В одному варіанті втілення даний винахід передбачає комбіновану вакцину проти кору та малярії, яка включає рекомбінантну вірусну вакцину кору, що експресує антигени малярії, здатні викликати імунну відповідь і захист проти кору та малярії. В іншому варіанті втілення, даний винахід передбачає рекомбінантну вірусну вакцину кору, що має нуклеотидну послідовність, яка експресує антиген MSP1 малярії. В кращому варіанті втілення, рекомбінантна вірусна вакцина кору має нуклеотидну послідовність, яка експресує малярійний антиген d190 або d83-30-38 або d42 як у закріпленій, так і в секретованій формах, зі штаму 3D7 та штаму FCB1. У ще одному варіанті втілення, даний винахід передбачає рекомбінантну вірусну вакцину кору, що має нуклеотидну послідовність, яка експресує малярійний антиген Diversity Covering (DiCo) AMA1. У ще одному варіанті втілення, даний винахід передбачає рекомбінантну вірусну вакцину кору, що має нуклеотидну послідовність, яка експресує малярійний антиген CS. Опис фігур Фіг. 1: Схематичне зображення антигеномної ДНК p(+)MV-EZ вірусу кору. p(+)MV-EZ є плазмідою, одержаною з pBluescript, що містить повну послідовність вірусу кору (Edmonston Zagreb), під контролем промотора РНК полімерази T7 (T7), який включає три ATU, відповідно, в положенні 1 (перед геном N вірусу кору), 2 (між генами P та M вірусу кору) та 3 (між генами H та L вірусу кору), і точно закінчується дельта-рибозимом гепатиту та термінатором РНК полімерази T7 (5T7t). Розмір плазміди 18941 пн. Фіг. 2: Зображення синтетичного гена MSP-1 (d-190) штаму 3D7. Показані кодуючі нуклеотиди на фланкуючих ділянках фрагментів гена d-190 (d-83-30-38 та d-42) відповідні амінокислоти. Унікальні сайти рестрикції, додані для процедур клонування, виділені кольором; SP: сигнальний пептид; GPI: послідовність глікозил-фосфатидил-інозит, яка кодує мембранноякорну ділянку. Фіг. 3: Зображення синтетичного гена MSP-1 (d-190) штаму FCB1. Показані кодуючі нуклеотиди на фланкуючих ділянках фрагментів гена d-190 (d-83-30-38 та d-42) відповідні амінокислоти. Унікальні сайти рестрикції, додані для процедур клонування, виділені кольором; SP: сигнальний пептид; GPI: послідовність глікозил-фосфатидил-інозит, яка кодує мембранноякорну ділянку. ділянки SP та GPI належать штаму 3D7. Фіг. 4. Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d190-3D7. Це плазміда, одержана з p(+)MV-EZ, яка містить ген d-190 малярії (штам 3D7), 5253 пн, що кодує GPI-закріплену форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 24323 пн. Фіг. 5: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d190*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-190* малярії (штам 3D7), 5160 пн, який кодує секретовану форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 24227 пн. 5 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Фіг. 6: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-d190-3D7 або p(+)MV3-EZ-d190*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-190 малярії (штам 3D7), 5253 пн, який кодує GPI-закріплену форму білка, або ген d-190* малярії (штам 3D7), 5160 пн, який кодує секретовану форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAI-BssHII. Рекомбінантна плазміда p(+)MV3-EZ-d190 має розмір 24323 пн, а p(+)MV3-EZ-d190* - 24227 пн Фіг. 7: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d83-30-83D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-83-30-38 малярії (штам 3D7), 4122 пн, який кодує GPI-закріплену форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 23195 пн. Фіг. 8: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d83-3038*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-83-30-38* малярії (штам 3D7), 4029 пн, який кодує секретовану форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 23105 пн. Фіг. 9: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-d83-30-383D7 або p(+)MV3-EZ-d83-30-38*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-8330-38 малярії (штам 3D7), 4122 пн, який кодує GPI-закріплену форму білка, або ген d-83-30-38* (штам 3D7), 4029 пн, який кодує секретовану форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAI-BssHII. Рекомбінантна плазміда p(+)MV3-EZ-d83-30-38 має розмір 23195 пн, p(+)MV3-EZ-d83-30-38* - 23105 пн. Фіг. 10: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d42-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-42 малярії (штам 3D7), 1347 пн, який кодує GPI-закріплену форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 20417 пн. Фіг. 11: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d42*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-42* малярії (штам 3D7), 1254 пн, який кодує секретовану форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 20345 пн. Фіг. 12: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-d42-3D7 або p(+)MV3-EZ-d42*-3D7. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-42 малярії (штам 3D7), 1347 пн, який кодує GPI-закріплену форму білка, або ген d-42* малярії (штам 3D7), 1254 пн, який кодує секретовану форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAI-BssHII. Рекомбінантна p(+)MV3-EZ-d42 має розмір 20417 пн, p(+)MV3-EZ-d42* має розмір 20345 пн. Фіг. 13: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-d190FCB1. Це плазміда, одержана з p(+)MV-EZ, містить ген d-190 малярії (FCB1 штам), 5013 пн, який кодує GPI-закріплену форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 24083 пн. Фіг. 14: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-d190FCB1. Це плазміда, одержана з p(+)MV-EZ, що містить ген d-190 малярії (FCB1 штам), 5013 пн, який кодує GPI-закріплену форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAI-BssHII. Рекомбінантна плазміда p(+)MV3-EZ-d190 має розмір 24083 пн. Фіг. 15: Зображення синтетичного гена CS. Зображені кодуючі нуклеотиди на фланкуючих ділянках гена CS та відповідні амінокислоти. Унікальні сайти рестрикції, додані для процедур клонування, виділені кольором. Фіг. 16: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-CS. Це плазміда, одержана з p(+)MV-EZ, що містить ген CS, 1119 пн, клонований в положенні два геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 20219 пн. Фіг. 17: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-CS. Це плазміда, одержана з p(+)MV-EZ, що містить ген CS, 1119 пн, клонований в положенні три геному кору шляхом гідролізу SgrAI-BssHII. Розмір рекомбінантної плазміди становить 20219 пн. Фіг. 18: Зображення повного синтетичного гена DiCo-1. Показані кодуючі нуклеотиди на фланкуючих ділянках доменів повного гена DiCo1 (екто та транс-цито) та відповідні амінокислоти. Унікальні сайти рестрикції, додані для процедур клонування, виділені кольором; SP: сигнальний пептид, оптимізований для людини за складом кодонів. Фіг. 19: Зображення екто синтетичного гена DiCo-1. Показані кодуючі нуклеотиди на фланкуючих ділянках екто домену DiCo1 та відповідні амінокислоти. Унікальні сайти рестрикції, додані для процедур клонування, виділені кольором; SP: сигнальний пептид (оптимізований для людини за складом кодонів). 6 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Фіг. 20: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-DiCo1повної. Це плазміда, одержана з p(+)MV-EZ, що містить повний ген DiCo1, 1689 пн, який кодує трансмембранну форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAIBssHII. Розмір рекомбінантної плазміди становить 20753 пн. Фіг. 21: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-DiCo1повної. Це плазміда, одержана з p(+)MV-EZ, що містить повний ген DiCo1, 1689 пн, який кодує трансмембранну форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAIBssHII. Розмір рекомбінантної плазміди становить 20753 пн. Фіг. 22: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV2-EZ-DiCo1екто. Це плазміда, одержана з p(+)MV-EZ, що містить ген DiCo1 екто, 1458 пн, який кодує секретовану форму білка, клонований в положенні два геному кору шляхом гідролізу SgrAIBssHII. Розмір рекомбінантної плазміди становить 20525 пн. Фіг. 23: Схематичне зображення рекомбінантної плазміди кору-малярії p(+)MV3-EZ-DiCo1екто. Це плазміда, одержана з p(+)MV-EZ, що містить ген DiCo1 екто, 1458 пн, який кодує секретовану форму білка, клонований в положенні три геному кору шляхом гідролізу SgrAIBssHII. Розмір рекомбінантної плазміди становить 20525 пн. Фіг. 24: Повна нуклеотидна послідовність p(+)MV2EZ-GFP. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 592-608 промотор T7 - 609-17354 антигеном MV Edmonston Zagreb - 4049-4054 сайт рестрикції MluI - 4060-4067 сайт рестрикції SgrAI - 4079-4084 сайт рестрикції BssHII - 4085-4801 ORF (відкрита рамка зчитування) зеленого флуоресцентного білка (GFP) - 4805-4810 сайт рестрикції BssHII - 4817-4822 сайт рестрикції AatII - 17355-17580 рибозим HDV та термінатор T7 Фіг. 25: Повна нуклеотидна послідовність p(+)MV3EZ-GFP. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 592-608 промотор T7 - 609-17359 антигеном MV Edmonston Zagreb - 9851-9856 сайт рестрикції MluI - 9862-9869 сайт рестрикції SgrAI - 9886-9891 сайт рестрикції BssHII - 9892-10608 ORF (відкрита рамка зчитування) зеленого флуоресцентного білка (GFP) - 10612-10617 сайт рестрикції BssHII - 10624-10629 сайт рестрикції AatII - 17360-17585 рибозим HDV та термінатор T7 Фіг. 26: AN101TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-190 3D7, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-1903D7 - 100-105 сайт рестрикції BamHl - 4014-4020 сайт рестрикції BstEII - 5152-5157 сайт рестрикції AclI - 5158-5250 послідовність GPI - 5251-5253 стоп-кодон Фіг. 27: AN102TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-190* 3D7, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-190* 3D7 - 100-105 сайт рестрикції BamHl - 4014-4020 сайт рестрикції BstEII - 5152-5157 сайт рестрикції AclI - 5158-5160 стоп-кодон 7 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 Фіг. 28: AN103TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-83-30-38 3D7, клонованоїавторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-83-30-38 3D7 - 100-105 сайт рестрикції BamHl - 4014-4020 сайт рестрикції BstEII - 4021-4026 сайт рестрикції AclI - 4027-4119 послідовність GPI - 4120-4122 стоп-кодон Фіг. 29: AN104TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-83-30-38* 3D7, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-83-30-38* 3D7 - 100-105 сайт рестрикції BamHl - 4014-4020 сайт рестрикції BstEII - 4027-4029 стоп-кодон Фіг. 30: AN105TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-42 3D7, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-42 3D7 - 100-105 сайт рестрикції BamHl - 108-114 сайт рестрикції BstEII - 1246-1251 сайти рестрикції AclI - 1252-1344 послідовність GPI - 1345-1347 стоп-кодон Фіг. 31: AN106TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-42* 3D7, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-42* 3D7 - 100-105 сайт рестрикції BamHl - 108-114 сайт рестрикції BstEII - 1246-1251 сайти рестрикції AclI - 1252-1254 стоп-кодон Фіг. 32: AN107TE: це відкрита рамка зчитування (ORF) послідовності MSP1 d-190 FCB1, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 сигнальний пептид d-190 FCB1 - 100-105 сайт рестрикції BamHl - 146-151 сайт рестрикції HindIII - 3825-3831 сайт рестрикції BstEII - 4912-4917 сайти рестрикції AclI - 4918-5010 послідовність GPI - 5011-5013 стоп-кодон Фіг. 33: AN108TE: це відкрита рамка зчитування (ORF) послідовності CS, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-1116 послідовність CS - 1117-1119 стоп-кодон Фіг. 34: AN109TE: це відкрита рамка зчитування (ORF) повної послідовності DiCo 1, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 повни сигнальний пептид DiCo1 - 100-105 сайт рестрикції BamHl 8 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 - 106-1686 відкрита рамка зчитування (ORF) повної послідовності DiCo 1 - 1687-1689 стоп-кодон Фіг. 35: AN110TE: це відкрита рамка зчитування (ORF) екто послідовності DiCo 1, клонованої авторами винаходу. Послідовність може бути описана із зазначенням положень нуклеотидів у такий спосіб: - 1-3 стартовий кодон - 4-99 екто сигнальний пептид DiCo1 - 100-105 сайт рестрикції BamHl - 106-1455 відкрита рамка зчитування (ORF) екто послідовності DiCo 1 - 1456-1458 стоп-кодон Фіг. 36: Порівнянні цитопатичні ефекти, створювані у клітинах Vero після інфекції рекомбінантною вірусною вакциною на основі вірусу MV p-42 кору-малярії. Фіг. 37: Експресія трансгена d-42 3D7, вставленого в положення три вектора кору (MV3EZ-d42 SgrAI). Клітинні лізати з пасажу 1, 5 та 10 аналізують методом вестерн-блотингу у порівнянні з пустим вектором кору (MVEZ) та негативним контролем (MV3L1, рекомбінантний вірус MVпапіломи). Фіг. 38: Експресія трансгена d-42 3D7, вставленого в положення три вектора кору (MV3EZ-d42 SgrAI) за результатами аналізу методом імунофілуоресценції, у порівнянні з пустим вектором кору (MVEZ) та негативним контролем (MV2EZL1, рекомбінантним вірусом MV-папіломи). Стрілки показують одні й ті самі синцитії при спостереженні під оптичним мікроскопом до та після імунофарбування. Фіг. 39: Крива кінетики росту рекомбінантного вірусу p-42 кору-малярії у порівнянні з MVвірусною вакциною. Фіг. 40: .Експресія трансгена d-190 FCB1, вставленого в положення два та три вектора кору (MV2-3EZ-d-190 SgrAI FCB1). Клітинні лізати аналізують методом вестерн-блотингу порівняно з пустим вектором кору (MVEZ) та негативним контролем (MV2EZL1, рекомбінантним вірусом MVпапіломи). Фіг. 41: Крива кінетики росту рекомбінантного вірусу p-190-FCB1 кору-малярії у порівнянні з MV-вірусною вакциною. Детальний опис винаходу Метою винаходу є продукування комбінованої вакцини проти кору та малярії з рекомбінантних векторів кору, здатних містити стабільно інтегровані ДНК послідовності, що кодують CS, MSP-1 або його часткові фрагменти та AMA-1 або часткові фрагменти, у секретованих або поверхнево закріплених формах, які належать P. falciparum. Винахід повинен також включати рятування рекомбінантних вірусів MV-малярії, які є здатними інфікувати, реплікуватися та експресувати антигени PfCS, PfMSP-1 та PfAΜΑ-1 у сприйнятливих трансгенних хазяях-мишах, мавпах та людях. Крім цього, передбачається, що винахід включає конструювання багатовалентних рекомбінантних векторів кору-малярії, у яких два різні антигени одночасно клонуються та експресуються в одному векторі, створюючи імунітет проти них обох. Крім того, винахід стосується комбінації трьох різних рекомбінантних вірусів кору-малярії, кожен з яких несе різні гени та експресує різні антигени, у такий спосіб, щоб викликати імунну відповідь у хазяїна, спрямовану проти різних стадій життєвого циклу паразита. На додаток, винахід включає спосіб виробництва рекомбінантних вірусів кору-малярії, які не містять дефектних інтерферуючих частинок (DI). Відомо, що DI в значному ступені інгібують ріст вірусу в будь-якій системі продукування та успішно пригнічують імунну відповідь у людей. Крім того, винахід включає спосіб виробництва вакцини, що містить такі рекомбінантні віруси. Наведені нижче приклади описують кращий спосіб здійснення винаходу. Слід розуміти, що ці приклади наведені з метою ілюстрації та не повинні тлумачитися як такі, що обмежують у будь-який спосіб обсяг винаходу. Приклад 1: Конструювання рекомбінантних плазмід MV-PfMSP-1 Усі процедури клонування здійснюють згідно з методиками, описаними у Sambrook et al. (1989). Усі рестрикційні ферменти були одержані від New England Bio Labs; олігонуклеотиди ПЛРпраймерів та ДНК-полілінкери були одержані від Invitrogen. PfMSP1 та його фрагменти (d-83-30-38 та d-42) у секретованій та закріпленій формі, були хімічно синтезовані та оптимізовані до кодонів людини. Їх клонують у проміжний вектор pZE21MV та трохи модифікують, додаючи сайт клонування SgrAI на 5'-кінці, за яким йде 9 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 оптимізована послідовність Козака (Kozak) (TCATCA). Ці модифікації перевіряють шляхом секвенування у MWG Biotech. Перелік рекомбінантних плазмід MV-PfMSP-1, GPI-закріплених та секретованих (*) форм, одержаних зі штаму 3D7, що належать до прототипу MAD20, та одержаних зі штаму FCB1, що належать до прототипу K1: p(+)MV2EZ-d-190-SgrAI (3D7) p(+)MV3EZ-d-190-SgrAI (3D7) p(+)MV2EZ-d-83-30-38-SgrAI (3D7) p(+)MV3EZ-d-83-30-38-SgrAI (3D7) p(+)MV2EZ-d-42-SgrAI (3D7) p(+)MV3EZ-d-42-SgrAI (3D7) p(+)MV2EZ-d-190*-SgrAI (3D7) p(+)MV3EZ-d-190*-SgrAI (3D7) p(+)MV2EZ-d-83-30-38*-SgrAI (3D7) p(+)MV3EZ-d-83-30-38*-SgrAI (3D7) p(+)MV2EZ-d-42*-SgrAI (3D7) p(+)MV3EZ-d-42*-SgrAI (3D7) p(+)MV2EZ-d-190-SgrAI (FCB1) p(+)MV3EZ-d-190-SgrAI (FCB1) 1a) Конструювання p(+)MV2EZ-d-190-SgrAI (3D7, 24323 пн) та p(+)MV3EZ-d-190-SgrAI (3D7, 24323 пн). 1 мкг ДНК плазміди MV, що містить зелений флуоресцентний білок (GFP) (p(+)MV2-3EZ-GFP, штам Berna, 19774 пн: фігури 24 та 25) гідролізують однією одиницею кожного з рестрикційних ферментів SgrAI та BssHII, протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель та подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (19048 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Видобувають 1 мкг гена d-190, вставленого в проміжну плазміду (pZE21MV-d-190 SgrAI, 7564 пн) шляхом гідролізу за допомогою SgrAI-BssHII (по одній одиниці кожного ферменту), протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (5275 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Таким чином, вектор (ДНК MV: фігура 1) та вставку (ДНК d-190: фігура 2) лігують в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4 та його власний реакційний буфер в кінцевому об’ємі 100 мкл. Потім хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування, відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-190-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 9335) представлений на фігурі 4, а його відкрита рамка зчитування (ORF) наведена на фігурі 26. Ген d-190-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 15137), представлений на фігурі 6. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 21884) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних вірусів MV2-3-d-190-3D7. 1b) Конструювання p(+)MV2EZ-d-83-30-38-SgrAI (3D7, 23195 пн) та p(+)MV3EZ-d-83-30-38SgrAI (3D7, 23195 пн). Вектори кору були одержані, як детально описано у прикладі 3a. Гідролізують pZE21MV-d-190 SgrAI за допомогою BstEII-AclI для того, щоб вирізати фрагмент d-42; лігують полілінкер з липкими кінцями BstEII та AclI для одержання проміжної плазміди pZE21MV-d-83-30-38-SgrAI (6436 пн). Послідовність полілінкера: 5'-GTCACCAGCGGCCGCAA-3'. 10 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 Гідролізують 1 мкг pZE21MV-d-83-30-38 SgrAI за допомогою SgrAI-BssHII (по одній одиниці кожного ферменту), протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (4147 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Таким чином, вектор (ДНК MV: фігура 1) та вставку (ДНК d-83-30-38: фігура 2) лігують в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4 та його власний реакційний буфер в кінцевому об’ємі 100 мкл. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності потім вирівнюють з гаданими за допомогою прикладної програми DNA Strider. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-83-30-38-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 8207), представлений на фігурі 7, а його відкрита рамка зчитування (ORF) наведена на фігурі 28. Ген d-83-30-38-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 14006), представлений на фігурі 9. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 20756) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних вірусів MV2-3-d-83-30-38-3D7. 1c) Конструювання p(+)MV2EZ-d-42-SgrAI (3D7, 20417 пн) та p(+)MV3EZ-d-42-SgrAI (3D7, 20417 пн). Вектори кору були одержані, як детально описано у прикладі 3a. Видобувають 1 мкг гена d-42, вставленого в проміжну плазміду (pZE21MV-d-42 SgrAI, 3658 пн), шляхом гідролізу SgrAI-BssHII (по одній одиниці кожного ферменту), протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (1369 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Таким чином, вектор (ДНК MV: фігура 1) та вставку (ДНК d-42: фігура 2) лігують в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4 та його власний реакційний буфер в кінцевому об’ємі 10 мкл. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-42-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 5429), представлений на фігурі 10, а його відкрита рамка зчитування (ORF) наведена на фігурі 30. Ген d-42-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 11231), представлений на фігурі 12. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 17978) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних MV2-3-d-423D7 віруси. Рекомбінантні віруси p-42 кору-малярії та вакцина MV індукували подібні цитопатичні ефекти (фігура 36). Трансген досить стабільно експресується: його експресія повністю зберігалася в усіх проаналізованих клонах потомства, одержаних з окремих вихідних врятованих клонів після десяти послідовних пасажів вірусу у диплоїдних клітинах людини MRC5 (фігура 37-38). Криві росту рекомбінантного вірусу MV-малярії та вакцини MV продемонстрували однакову кінетику (фігура 39). 11 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 1d) Конструювання p(+)MV2EZ-d-190*-SgrAI (3D7, 24227 пн) та p(+)MV3EZ-d-190*-SgrM (3D7, 24227 пн). Вектори кору були одержані, як детально описано у прикладі 3a. Використовуючи проміжний вектор pZE21MVd-190-SgrAI як матрицю, проводили ПЛРреакцію для видалення якорної ділянки GPI, розташованої між сайтами AclI (поз. 5434) та СlaI (поз. 5536). ПЛР-ампліфікації проводили з використанням коригувальної (proofreading) ДНК-полімерази Pfu (Stratagene). ДНК-послідовності синтетичних олігонуклеотидних праймерів наведені малими літерами для нуклеотидів MV та великими літерами для не-MV нуклеотидів; послідовності відповідних сайтів розпізнавання ендонуклеаз рестрикції підкреслені. Були використані такі олігонуклеотиди праймери: For-ClaI, 5'CCAATAAACGTTTAATAGatcgattacgcgcgctctagc-3', та Rev-ΑνrII, 5'-gcctttgagtgagctgatacc-3'. For-ClaI є гомологічним матриці на рівні сайтів ClaI та BssHII та містить виступний кінець (великими літерами) з двома стоп-кодонами (TAATAG), сайт AclI (AACGTT), та захисний сайт AclI довжиною 6 пн (CCAATA). У так званому ПЛР-GPI та в кінцевому конструкті d-190*, AclI буде розташований поряд з ClaI. Rev-Avrll є гомологічним матриці (від поз. 5704 до 5724). Продукт ПЛР має довжину 207 пн: його гідроліз за допомогою AclI+ΑνrII та лігування з попередньо гідролізованим AclI+ΑνrII проміжним вектором pZE21MVd-190-SgrAI привів до одержання pZE21MVd-190*-SgrAI. Детальніше, гідроліз вектора за допомогою AclI+ΑνrII дав дві смуги 7318 пн та 246 пн (що містить ділянку GPI, яку треба видалити): фрагмент 7,3 тпн очищають з агарозного геля за допомогою набору для очищення QIAEX II (Qiagen) та лігують з гідролізованим AclI+ΑνrII ПЛР (вставка), одержуючи pZE21MVd-190*-SgrAI. Для скринінгу позитивних клонів, проводять гідроліз за допомогою NcoI, одержуючи одну смугу 7 тпн з проміжного вектора d-190*, та дві смуги 1,3 та 5,7 тпн з вихідного конструкту GPIякір. Для конструювання кінцевих рекомбінантних p(+)MeV2EZ-d190* та p(+)MeV3EZ-d190* (фігура 5 та фігура 6), відповідно до "правила шести", вектори MeV та проміжну плазміду гідролізують за допомогою SgrAI+BssHII і потім лігують один з одним. Детальніше, pZE21MVd-190*-SgrAI, гідролізований SgrAI+BssHII, дає три смуги, 5,2 тпн + 1,3 тпн + 900 пн. Послідовність D-190* знаходиться у фрагменті 5,2 тпн, який вирізають, очищають та лігують з векторами MeV2EZ та MeV3EZ, гідролізованими SgrAI+BssHII (довжиною 19 тпн), в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al., 1989), висівають та проводять селекцію на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-190*-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 9239), представлений на фігурі 5, а його відкрита рамка зчитування (ORF) наведена на фігурі 27. Ген d-190*-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 15041), представлений на фігурі 6. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 21788) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних вірусів MV2-3-d-190*-3D7. 1e) Конструювання p(+)MV2EZ-d-83-30-38*-SgrAI (3D7, 23105 пн) та p(+)MV3EZ-d-83-30-38*SgrAI (3D7, 23105 пн). Вектори кору були одержані, як детально описано у прикладі 3a. Проміжний вектор pZE21MVd-190-SgrAI гідролізують BstEII-ClaI для того, щоб вирізати фрагмент d-42 та ділянку GPI, розташовану між сайтами AclI (поз. 5434) та ClaI (поз. 5536); полілінкер з липкими BstEII та ClaI кінцями лігують для одержання проміжної плазміди pZE21MV-d-83-30-38*-SgrAI (6346 пн). Послідовність полілінкера: 5'-GTCACCGGGGAATAATAGCGCAT-3'. 12 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 ДНК-послідовність синтетичного олігонуклеотидного полілінкера представлена великими літерами для не-MV нуклеотидів; послідовності відповідних сайтів розпізнавання ендонуклеаз рестрикції підкреслені. Полілінкер включає липкі кінці BstEII (GTCACC) та ClaI (AT), два стоп-кодони (TAATAG), та триплет (GCG) для забезпечення дотримання правила шести. 1 мкг pZE21MV-d-83-30-38* SgrAI гідролізують за допомогою SgrAI-BssHII (по одній одиниці кожного ферменту), протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (4057 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Таким чином, вектор (MV, ДНК: Фіг. 1) та вставку (d-83-30-38*, ДНК: Фіг. 2) лігують в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4 та його власний реакційний буфер у кінцевому об’єм 10 мкл. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності потім вирівнюють з гаданими за допомогою прикладної програми DNA Strider. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-83-30-38*-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 8117), представлений на фігурі 8, а його відкрита рамка зчитування (ORF) наведена на фігурі 29. Ген d-83-30-38*-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 13919), представлений на фігурі 9. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 20666) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних вірусів MV2-3-d-83-30-38*-3D7. 1f) Конструювання p(+)MV2EZ-d-42*-SgrAI (3D7, 20345 пн) та p(+)MV3EZ-d-42*-SgrAI (3D7, 20345 пн). Вектори кору були одержані, як детально описано у прикладі 3a. Використовуючи проміжний вектор pZE21MVd-42-SgrAI (3658 пн) як матрицю, проводять ПЛР-реакці для видалення якорної ділянки GPI, розташованої між сайтами AclI (поз. 1528) та ClaI (поз. 1630). ПЛР-ампліфікації проводять з використанням коригувальн ДНК-полімерази Pfu (Stratagene). ДНК-послідовності синтетичних олігонуклеотидних праймерів наведені малими літерами для нуклеотидів MV та великими літерами для не-MV нуклеотидів; послідовності відповідних сайтів розпізнавання ендонуклеаз рестрикції підкреслені. Були використані такі олігонуклеотиди праймери: For-ClaI, 5'CCAATAAACGTTTAATAGategattacgcgcgctctagc-3', та Rev-AvrII, 5'-gcctttgagtgagctgatacc-3'. For-ClaI є гомологічним матриці на рівні сайтів ClaI (поз. 1630) та BssHII (поз. 1639) та містить виступний кінець (великими літерами) з двома стоп-кодонами (TAATAG), сайт AclI (AACGTT), та захисний сайт AclI довжиною 6 пн (CCAATA). У так званому ПЛР-GPI та в кінцевому конструкті d-42*, AclI буде розташований поряд з ClaI. Rev-AvrII є гомологічним матриці (від поз. 1798 до 1818). Продукт ПЛР мав довжину 207 пн: його гідроліз за допомогою AclI+AvrII та лігування з попередньо гідролізовани AclI+AvrII проміжним вектором pZE21MVd-42-SgrAI давало pZE21MVd-42*-SgrAI. Детальніше, гідроліз вектора за допомогою AclI+AvrII давав дві смуги 3412 пн та 246 пн (що містить ділянку GPI, яку треба видалити): фрагмент 3,4 тпн очищають з агарозного геля за допомогою набору для очищення QIAEX II (Qiagen) та лігують з гідролізованим AclI+AvrII продуктом PCR (вставка), одержуючи pZE21MVd-42*-SgrAI. Для скринінгу позитивних клонів, має бути проведений гідроліз NcoI, з одержанням однієї смуги 3,4 тпн з проміжного вектора d-42*, та двох смуг 1,3 та 2,3 тпн з вихідного конструкту GPIякір. 13 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Для конструювання кінцевих рекомбінантних p(+)MeV2EZ-d42* та p(+)MeV3EZ-d42*, відповідно до "правила шести", вектори MeV та проміжну плазміду гідролізують за допомогою SgrAI+BssHII і потім лігують один з одним. Детальніше, pZE21MVd-42*-SgrAI, гідролізована за допомогою SgrAI+BssHII+Spel, давала чотири смуги, 1,3 тпн + 936 пн + 800 пн + 400 пн. Послідовність d-42* знаходиться у фрагменті 1,3 тпн, який вирізають, очищають та лігують з векторами MeV2EZ та MeV3EZ, гідролізованими SgrAI+BssHII (довжиною 19 тпн), в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНК-лігази T4. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та проводять селекцію на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-42*-3D7, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 5357), представлений на фігурі 11, а його відкрита рамка зчитування (ORF) наведена на фігурі 31. Ген d-42*-3D7, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 11159), представлений на фігурі 12. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 17906) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних MV2-3-d42*-3D7 віруси. 1g) Конструювання p(+)MV2EZ-d-190-SgrAI (FCB1, 24083 пн) та p(+)MV3EZ-d-190-SgrAI (FCB1, 24083 пн). По-перше, проводять клонування синтетичного гена MSP-1 штаму FCB1 в проміжну плазміду pZE21MV-SgrAI, зі збереженням сигнального пептиду та GPI-якірної ділянки MSP-1 штаму 3D7. Ген d-190 (FCB1) одержують у декілька стадій з проміжного вектора, названого pZE23f-GX-190H, у такий спосіб: i) гідролізують 1 мкг плазміди pZE21MV-d-190-SgrAI (3D7) за допомогою рестрикційних ферментів HindIII + AclI, протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (2558 пн), що відповідає вектору, вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм. ii) проводять ПЛР-реакцію, використовуючи pZE23f-GX-190H як матрицю, для ампліфікації та виділення ділянки d-42 MSP-1/FCB1. ПЛР-ампліфікацію проводять використанням коригувальної ДНК полімерази Pfu (Stratagene). ДНК-послідовності синтетичних олігонуклеотидних праймерів наведені малими літерами для нуклеотидів MV та великими літерами для не-MV нуклеотидів; послідовності відповідних сайтів розпізнавання ендонуклеаз рестрикції підкреслені. Були використані такі олігонуклеотиди праймери, вставлені в послідовність pZE23f-GX-190H: For-1 FCB1, 5'-CCCAAGCTTccaggtggtcaccggAgagctgtcactcc-3', та Rev-1 FCB1, 5'GCCTGCaacgttGCTagagctggagcaGaaGatcccgtcg-3'. For-1 FCB1 є гомологічним матриці від поз. 4509 до поз. 4538, включаючи сайт BstEII (ggtcacc). A (великою літерою) відповідала t у матриці, і була модифікована для усунення сайта SgrAI. Він має виступний кінець (великими літерами) із сайтом HindIII (AAGCTT), розташованим після його захисним сайтом довжиною 3 пн (CCC). Rev-1 FCB1 містить сайт AclI (aacgtt), перед яким розташований захисний сайт з 6 пн (GCCTGC). До нього був введений триплет GCT, що кодує серин, для забезпечення дотримання правила шести; два були модифіковані у G, щоб уникнути полі(A)-сайта. Одержаний PCR-HindHI-AclI (1,1 тпн) гідролізують за допомогою HindIII+AclI та лігують протягом ночі при 16 °C в еквімолярному співвідношенні, з попередньо гідролізованим за допомогою HindIII+AclI pZE21MV-d-190-SgrAI (стадія i), одержуючи pZE21MV-d-42-SgrAI-FCB1 (3657 пн). Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та проводять селекцію на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та проведення гідролізу рестрикційними ферментами з використанням HindIII+AclI (очікувані фрагменти 2558 пн + 1099 пн). 14 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 iii) pZE21MV-d-42-SgrAI-FCB1, одержану, як описано на стадії ii, гідролізують HindIII + BstEII (HindIII, поз. 428, та BstEII, поз. 440), і потрібну смугу (3645 пн), що відповідає відкритому вектору, завантажують на 1% агарозний гель, вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм. iv) pZE23f-GX-190H гідролізують HindIII + BstEII і потрібну смугу 3679 пн (вставка), що відповідає фрагменту d-83-30-38/FCB1, очищають з гелю, як описано вище. v) гідролізований за допомогою HindIII + BstEII фрагмент 3657 пн (вектор), одержаний з pZE21MV-d-42-SgrAI-FCB1, лігують з гідролізованим HindIII + BstEII фрагментом 3679 пн (вставка), який містить d-83-30-38/FCB1 та був одержаний гідролізом з pZE23f-GX-190H. Лігування здійснюють в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНК-лігази T4, одержуючи pZE21MV-d-190-SgrAI-FCB1 (7324 пн). Після цього, хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та проводять селекцію на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Вектори кору для конструювання p(+)MV2EZ-d-190-SgrAI-FCB1 та p(+)MV3EZ-d-190-SgrAIFCB1 були одержані, як детально описано у прикладі 3a. 1 мкг гена d-190/FCB1, вставленого в проміжну плазміду (pZE21MV-d-190 SgrAI-FCB1, 7324 пн), виділяють шляхом гідролізу SgrAI-BssHII (по одній одиниці кожного ферменту) протягом двох годин при їх оптимальній температурі, у кінцевому об’ємі 50 мкл. Всю гідролізовану ДНК завантажують на 1% агарозний гель, подають напругу 80 вольт протягом приблизно 2 годин. Потім потрібну смугу (5035 пн) вирізають з гелю, очищають за допомогою набору QIAEX для очищення гелю і концентрацію ДНК обчислюють за поглинанням при 260 нм та доводять до 1 мкг/мл. Таким чином, вектор (ДНК MV: фігура 1) та вставку (ДНК d-190/FCB1: фігура 3) лігують в еквімолярному співвідношенні протягом ночі при 16 °C, використовуючи одну одиницю ДНКлігази T4 та його власний реакційний буфер в кінцевому об’ємі 10 мкл. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді-, та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Ген d-190-FCB1, вставлений в положення 2 вектора MV (SgrAI, поз. 4060, та BssHII, поз. 9095), представлений на фігурі 13, а його відкрита рамка зчитування (ORF) наведена на фігурі 32. Ген d-190-FCB1, вставлений в положення 3 вектора MV (SgrAI, поз. 9862, та BssHII, поз. 14897), представлений на фігурі 14. Довжина геному (починаючи від ACC, поз. 609, до GGT, поз. 21884) рекомбінантних плазмід кору-малярії була кратною шести, дозволяючи проводити рятування рекомбінантних вірусів MV2-3-d-190-FCB1. Трансген досить стабільно експресується: його експресія повністю зберігалася в усіх проаналізованих клонах потомства, одержаних з окремих вихідних врятованих клонів після десяти серійних пасажів вірусу у диплоїдних клітинах людини MRC5 (фігура 40). Криві росту рекомбінантного вірусу MV-малярія та вакцини MV демонстрували однакову кінетику (фігура 41). Приклад 2: Конструювання послідовності нуклеїнової кислоти DiCo1. Починаючи з амінокислотної послідовності DiCo1 (екто, транс та цитоплазматичні домени: aa 97-622) та використовуючи прикладну програму DNA Strider, була сконструйована послідовність нуклеїнової кислоти шляхом порівняння виродженої послідовності ДНК DiCo 1 з вибраним геном PfAMA1 (номер доступу AAG141.1), який є найбільш подібною до DiCo1 послідовністю після вирівнювання за допомогою BLAST. На 5'-кінці були додані придатні унікальні сайти рестрикції (MluI та SgrAI) як сайти клонування, за якими йде оптимальна послідовність KOZAC та людський оптимізований сигнальний пептид (SP). З 3'-кінця були додані два стоп-кодони та сайт клонування BssHII. Відповідно до цієї схеми, ми сконструювали дві нуклеотидні послідовності (дотримуючись "правила шести" для подальшої експресії у вектор кору), що кодують закріплену та секретовану форми білка DiCo1: перший ген включає ектоплазматичний, трансмембранний та 15 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 цитоплазматичний домени (Фігура 18), у той час як другий відповідає самому лише ектодомену (Фігура 19). Дві послідовності були оптимізовані до кодонів людини за допомогою GENEART, для зменшення % вмісту AT, для усунення полі(A) послідовності та мотив нестабільності РНК. ORF повного DiCo1 та ORF ектодомену DiCo1 наведені, відповідно, на фігурах 34 та 35. Приклад 3: Конструювання рекомбінантних плазмід MV-PfAMA-1 Усі процедури клонування здійснювалися відповідно до методик, описаних Sambrook et al. (1989). PfAMA1, та зокрема послідовності Diversity Covering 1 (DiCo1), в секретованій або закріпленій формі, були хімічно синтезовані та оптимізовані до кодонів людини. Секретовану та закріплену форми DiCo1 з оптимізованими кодонами гідролізують SgrAI+BssHII та лігують протягом ночі при 16 °C в еквімолярному співвідношенні, з попередньо гідролізованими MeV2EZ та MeV3EZ векторами (довжиною 19 тпн), використовуючи одну одиницю ДНК-лігази T4, одержуючи такі рекомбінантні плазміди MV-PfAMA-1: p(+)MV2EZ-DiCo1повна (Фігура 20), p(+)MV3EZ-DiCo1-повна (Фігура 21), p(+)MV2EZ-DiCo1-екто (Фігура 22), та p(+)MV3EZ-DiCo1-екто (Фігура 23). Приклад 4: Конструювання рекомбінантних плазмід MV-PfCS Конструювання p(+)MV2EZ-CS-SqrAI (20219 пн) та p(+)MV3EZ-CS-SqrAI (20219 пн) Усі процедури клонування були в основному такими, як описано у Sambrook et al. (1989). PfCS1, клонований в проміжний вектор pAdApt35Bsu.CS.Pfalc.aa-sub.gcc, ампліфікують методом ПЛР, та безпосередньо клонують в кінцеві вектори MV, одержуючи дві рекомбінантні плазміди MV-PfCS: p(+)MV2EZ-CS та p(+)MV3EZ-CS. Детальніше, проводять ПЛР-реакцію, використовуючи pAdApt35Bsu.CS.Pfalc.aa-sub.gcc як матрицю, для ампліфікації та виділення гена CS (фігура 15). ПЛР-ампліфікацію проводять з використанням коригувальної ДНК полімерази Pfu (Stratagene). ДНК послідовності синтетичних олігонуклеотидних праймерів наведені малими літерами для нуклеотидів MV та великими літерами для не-MV нуклеотидів; послідовності відповідних сайтів розпізнавання ендонуклеаз рестрикції підкреслені. Були використані такі олігонуклеотиди праймери, розроблені для послідовності pAdApt35Bsu.CS.Pfalc.aa-sub.gcc: For-SgrAI, 5'-ACTTCTCACCGGTGTggaagcttgccaccatgat-3', та Rev-BssHII-CS 5'- TAGCGCGCtctagaggatccttatcagc-3'. For-SgrAI є гомологічним матриці від поз. 1356 до поз. 1375, включаючи сайт HindIII (aagctt). Він містить виступний кінець (великими літерами) із сайтом рестрикції SgrAI (CACCGGTG), розташований після захисного сайта довжиною 6 пн (ACTTCT). Rev-BssHII-CS містить виступний кінець (великими літерами) із сайтом рестрикції BssHII (GCGCGC), який буде знаходитися поряд з Xbal (tctaga) у ПЛР-CS (1187 пн). Одержаний ПЛР-CS гідролізують SgrAI + BssHII та лігують протягом ночі при 16 °C в еквімолярному співвідношенні, з векторами MeV2EZ та MeV3EZ, попередньо гідролізованими SgrAI+BssHII (довжиною 19 тпн), використовуючи одну одиницю ДНК-лігази T4, і одержуючи, відповідно, p(+)MV2EZ-CS-SgrAI (20219 пн, фігура 16) та p(+)MV3EZ- CS-SgrAI (20219 пн, фігура 17). ORF CS наведена на фігурі 33. Хімічно компетентні клітини XL10 Gold трансформують з використанням усього об’єму для лігування відповідно до стандартного протоколу трансформації (Sambrook et al. 1989), висівають та піддають селекції на чашках з агаром LB на резистентність до ампіциліну. Проводять скринінг колоній шляхом приготування ДНК-плазмід (QIAGEN, міні-, міді- та максинабори) та гідролізу рестрикційними ферментами. Правильні клони надсилають в MWG для секвенування: послідовності, вирівняні з гаданими за допомогою прикладної програми DNA Strider, продемонстрували 100% ідентичність. Приклад 5: Клітини та віруси Клітини підтримують як моношари у середовищі Ігла, модифікованому за Дульбекко (DMEM), з добавками 5% сироватки плоду корови (FCS) для клітин Vera (нирки африканської зеленої мавпи) та з 10% FCS і 1% пеніциліну/стрептоміцину (P/S) для клітин 293T (нирки ембріона людини); DMEM з добавкою Glutamax (F12) та 10% FCS для MRC-5 (фібробласти плоду людини); DMEM з добавкою 10% FCS та 1,2 мг/мл G 418 для 293-3-46. 7 Для вирощування культур вірусу MV з титрами близько 10 буо/мл (бляшкоутворювальних одиниць/мл), рекомбінантні віруси та штам вакцини Edmonston Zagreb розмножують у клітинах MRC-5: очищення бляшок здійснюють шляхом перенесення синцитію на 35 мм клітинну культуру MRC-5, яку розширяють спочатку до 10 см чашки, а потім до колби 175 см. Вірусні 2 культури одержують з культур 175 см , коли утворення синцитію виражене приблизно на 90%. Збирають середовище, що відповідає так званій "безклітинній (free-cell) вірусній фракції", заморожують та відтаюють три рази та центрифугують для усунення клітинних уламків. 16 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 Середовище потім зберігають при -80 °C. Клітини, що відповідають так званій "клітинноасоційованій вірусній фракції", зішкрябають в 3 мл OPTIMEM (Gibco BRL) з подальшими трьома циклами заморожування та відтаювання, центрифугують і очищений супернатант (surnatant) зберігають при -80 °C. Приклад 6: Трансфекція плазмід та рятування вірусів MV Клітини 293T висівають у 35 мм лунки до досягнення ~50-70% злиття, при якому проводять трансфекцію. За 4 год. до трансфекції, середовище заміняють на 3 мл DMEM, що містить 10% FCS. Усі рекомбінантні плазміди були одержані з використанням набору для приготування 2+ плазмід QIAGEN. Для Ca -фосфатної копреципітації ДНК використовували набір від Invitrogen. Клітини піддають ко-трансфекції плазмідами у таких кінцевих концентраціях: pCA-L 0,5 мкг, pCA-N 0,5 мкг, pCA-P 0,1 мкг, pCA T7 1 мкг, і рекомбінантна плазміда кору-малярії - 4 мкг. Усі п’ять плазмід, розведені H2O, поміщають в пробірку Еппендорфа, що містить 2M CaCl2, суміш додають в іншу пробірку Еппендорфа, що містить буфер HEPES, при струшуванні, та інкубують протягом 30 хв. при кімнатній температурі (RT). Таким чином, ко-преципітати додають по краплях до культури і трансфекцію проводять при 37 °C та 5% CO2 протягом приблизно 18 год. Потім середовище для трансфекції заміняють на 3 мл DMEM, що містить 10% FCS. Інший спосіб одержання рекомбінантних вірусів вакцини проти кору та малярії описаний далі, з використанням клітини-помічників 293-3-46 (клітини нирки ембріона людини), які стабільно експресують білки N та P кору, а також РНК полімерази T7. Вірусна РНК-полімераза (великий білок, L) експресують шляхом ко-трансфекції клітини 15 нг плазміди peMCLa. Для підвищення ефективності трансфекції додають 300 нг pSC6-T7 Neo. Для проведення трансфекції використовують кальцій-фосфатний метод. Перші синцитії з’являються через 3-4 дні після трансфекції, коли клітини ще не досягають стадії слиття. Для полегшення утворення синцитій, майже злитий шар моношар клітин з кожної 35 мм лунки потім переносять на 10 см чашку. Кожен синцитій розводять у 300 мкл середовища для трансфекції та поміщають в стерильну пробірку Еппендорфа, що містить 700 мкл OPTIMEM, заморожують та відтаюють три рази, і зберігають при -80 °C. Приклад 7: Титрування вірусу методом аналізу бляшкоутворювання Проводять серійні 10-кратні розведення вірусу з використанням OPTIMEM до кінцевого об’єму 0,5 мл. Кожне розведення додають до 35 мм клітинних культур Vero. Через 1 год. адсорбції вірусу, інокулят видаляють і інфіковані клітини заливають 2 мл DMEM, що містить 5% FCS та 1% легкоплавкої агарози (LMP агароза). Після 5 днів інкубації при 37 °C та 5% CO2, культури фіксують 1 мл 10% TCA протягом 1 год., потім зшивають УФ протягом 30 хв. Після видалення верхнього шару агарози, моношари клітин забарвлюють кристалічним фіолетовим, розчиненим в 4% етанолі, промивають водою і бляшки підраховують під інвертованим мікроскопом. Приклад 8: Серійні пасажі рекомбінантних вірусів на MRC-5 Проводять серійні пасажі 10 разів врятованих вірусів на клітинах MRC5, висіяних на чашки діаметром 10 см, які були інфіковані стандартним та рекомбінантними вірусами MV при MOI (множинність зараження) 0,01 буо/клітину. Після того, як моношар був повністю інфікованим, 1% супернатанта (surnatant) кожної культури використовують для інфікування наступного моношару клітин MRC5. Для випробування експресії та стабільності трансгенів, віруси з пасажів 1, 5 та 10 використовувалися для додаткової характеризації експресії методами вестерн-блотингу та імунофілуоресценції. Приклад 9: Вестерн-блотинг, імунофілуоресценція Для аналізу експресії MV та малярії проводили вестерн-блотинг та імунофілуоресценцію. 5 Для вестерн-блотингу, клітини Vero, висіяні на 35 мм чашки (1-5×10 ) перевіряють наступного дня на досягнення 90% злиття та інфікують очищеною вірусною суспензією з клітинно-асоційованої вірусної фракції, використовуючи 0,1 MOI (множинність інфекції), включаючи MVEZ як контроль. Коли спостерігалося утворення приблизно 80% синцитій, клітини спочатку промивають PBS (фосфатно-сольовий буфер) і потім зішкрябають в 1 мл PBS та збирають в пробірку Еппердорфа, і центрифугують при 2000 об./хв. протягом 4 хв. Клітини потім лізують 5 хв./RT (кімнатна температура) за допомогою 70 мкл буфера для лізису (1% NP-40, 50 мМ Tris pH 8, 150 мМ NaCl) з добавкою коктейлю інгібітора протеази (Complete Mini, Roche, 1 836 153). Супернатанти очіщають центрифугуванням при 13000 об./хв. протягом 5 хв., та переносять в нову пробірку: додають 30 мкл 4× буфера для завантаження (Invitrogen); зразки перемішують та кип’ятять при 95 °C протягом 2 хв., центрифугують та зберігають при -20 °C. Проводять міграцію на SDS-PAGE, використовуючи NuPAGE 12% біс-акриламідний гель у відновних умовах, використовуючи 1× рухомий буфер, протягом 50 хв. при 200 В (на початку 100-125 мА, у кінці 60-80 мА). 17 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 Потм використовують напівсухий метод для перенесення виділених клітинних білків на нітроцелюлозну мембрану, при 14 В протягом 1 год. 30 хв. Як перші антитіла використовують кролячі поліклональні антитіла проти MSP1-p-83, розведені у PBST щонайменше до 1:30000, і проти MSP1-p-42, розведені щонайменше 1:50000. Другим антитілом було свиняче анти-кроляче антитіло, спряжене з пероксидазою хрону, що дозволяє проводити візуалізацію смуг за допомогою набору для посиленої хемілюмінесценції (ECL™, Amersham LifeScience). Для імунофілуоресценції, клітини Vero висівають на накривних стеклах 24 мм × 24 мм в 35 мм лунках, культивують протягом ночі та інфікують врятованим рекомбінантним вірусом. Через 3 дні після інфекції клітини на накривних стеклах фіксують 3,7% параформальдегідом у PBS, та пермеабілізують за допомогою 0,1% TX-100, промивають блокувальним розчином (PBS, що містить 1% BSA) протягом 1 год., та забарвлюють специфічними антитілами. Використовують mAb 5.2 із супернатанта мишачої гібридоми, що розпізнає EGF-подібний домен у p-19 ділянці p42, у розведенні 1:100, а потім FITCH-кон’юговану козячу анти-мишачу сироватку, розведену 1:250. Приклад 10: Крива кінетики росту 5 Клітини MRC5, висіяні на 35 мм чашку (1-5×10 ), контролюють до досягнення 90% злиття та інфікують очищеною вірусную суспензією з клітинно-асоційованої вірусної фракції, використовуючи 0,1 MOI, включаючи MVEZ як контроль. Зразки, що відповідають так званій "безклітинній (free-cell) вірусній фракції" та так званій "клітинно-асоційованій вірусній фракції", збирають щоденно протягом одного тижня та титрують. Приклад 11: Імунізація мишей Імуногенна активність описаних врятованих рекомбінантних вірусів MV-малярії перевіряють шляхом тестів імунізації, що проводяться на трансгенних мишах IFNAR/CD46, сприйнятливих до інфекції MV. Тварин утримують в оптимальних гігієнічних умовах та імунізують у віці 6-8 тижнів. Нижче наведений приклад імунізації мишей двома рекомбінантними вірусами кору-малярії: MeV2EZ-d-p42-SgrAI (GPI закріплена форма) та MeV2EZ-d-p42* (секретована форма). 5 Імунізацію виконують внутрішньом’язово з використанням 10 буо (PFU) кожного рекомбінантного вірусу MV-малярії трьома ін’єкціями в тижні 0, 4 та 8. Як негативний контроль використовували мишей, імунізованих рекомбінантним-пустим вірусом кору (клонований rMVEZ13-пустий). УФ-інактивований rMV використовують як контроль для визначення впливу реплікації вірусу на активацію імунної відповіді. Тестують імунну відповідь векторного антигену MV порівняно з очищеним білком d-42 (0,5 мг/мл): мишей імунізували підшкірно 20 мкг білка у неповному ад’юванті Фрейнда. Присутність MV-специфічних антитіл у сироватці імунізованих IFNAR/CD46 мишей (6 на експериментальну групу та 3 у контрольній групі) визначають методом ELISA з використанням 96-лункових мікропланшетів, покритих об’ємною фазою (bulk) вірусу кору EIA (ATCC VR-24), для детектування IgG антитіла. Білок розводять до 0,6 мкг/мл 0,05M карбонатним буфером (pH 9,4), та додають в 96-лункові мікротитрувальні планашети по 100 мкл на лунку. Планшети інкубують протягом ночі при 4 °C, промивають PBS/0,05% Tween 20 (PT) (pH 7.4), інкубують з PT (0,1 мл/лунку)-10% BSA протягом 60 хв. при 37 °C, та знов промивають PT. Додають серійні 2-кратні розведення тестованої сироватки (100 мкл/лунку), та планшети інкубують протягом 60 хв. при 37 °C. Планшети промивають PT та інкубують з 100 мкл козячого анти-мишачого IgG HRP, розведеного 1:2000 у PT протягом 30 хв. при 37 °C. Планшети промивають PT та інкубують з 100 мкл OPD (о-фенілендіамін, Fluka 78411). Реакцію зупиняють через 3-4 хв. Планшети зчитують за допомогою пристрою MicroElisa Reader на довжині хвилі 490 нм. Результати, які більш ніж утричі перевищують негативний контроль, вважають позитивною реакцією. Присутність MV-малярія-специфічних антитіл у сироватці імунізованих CD46 мишей (щонайменше 10 на експериментальну групу) визначають за допомогою аналізу методом ELISA. Стисло, 96-лункові мікропланшети покривають 50 нг/лунку штамами MSP-1-d42 3D7, розведеними карбонатним буфером, pH 9,4. Планшети інкубують протягом ночі при 4 °C, промивають PBS/0,05% Tween 20 (PT). Після цього, неспецифічні взаємодії блокують 10% знежиреного молока, розчиненого у PT, протягом 1 год. при 37 °C, і лунки знов промивають PT. Планшети послідовно інкубують з різними розведеннями мишачої сироватки (починаючи від 1:200, з подальшими серійними двократними розведеннями), пероксидаза-кон’югованим козячим анти-мишачим IgG та із субстратом OPD. Вимірюють величини оптичної густини при 490 нм. Значення вище рівня відсічення фону (середнє значення для сироватки MV імунізованих мишей, помножена на коефіцієнт 2,1) вважають позитивними. Титри зображені як обернені величини кінцевих розведень. 18 UA 104180 C2 Гуморальні імунн відповіді проти кору показані на фігурі A. Гуморальні імунні відповіді проти малярії p42 показані на фігурі B. Фігура A: Гуморальна відповідь проти кору Регулярно відбирають кров та визначають титри IgG кору. 5 Фігура B: Гуморальна відповідь проти малярії 19 UA 104180 C2 5 10 15 Регулярно відбирають кров та визначають титри IgG малярії. Приклад 12: Очищення рекомбінантного вірусу кору, що експресує антигени малярії, від дефектних інтерферуючих частинок (DI) методом очищення бляшок З літератури відомо, що після певного числа пасажів параміксовірусів, і зокрема, вірусів кору, відбувається накопичення дефектних інтерферуючих частинок (DI) (23, 24). Як було описано, ці DI призводять до розвитку різних дефектів: негативний вплив на безпечність вакцини, негативний вплив на вихід вірусу при продукуванні, нестабільність геному та пригнічення імунної реакції після вакцинації. Для того, щоб уникнути таких DI з нашими новими рекомбінантними вірусами, ми застосували метод очищення бляшок, як описано у прикладі 6, за тим винятком, що ми використовували клітини MRC5 замість клітин 293T. Після утворення прозорих, гарно визначених синцитій ми відсмоктували під мікроскопом мікропіпеткою такий матеріал для подальшого пасажу на свіжу тканинну культуру MRC5. Приклад 13: Очищення рекомбінантного вірусу кору, що експресує антигени малярії, від дефектних інтерферуючих частинок (DI) методом кінцевого розведення Методика кінцевого розведення була використана на мікропланшетах: в усіх лунках був щойно сформований свіжий моношар клітин MRC5. Вірусну суспензію, яка містить 20 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 рекомбінантні віруси кору-малярії, готують двократними розведеннями. З лунки останнього (latest) моношару, у якій були виявлені синцитії, супернатант відсмоктують піпеткою. Супернатант змішують з суспензією, що містить клітини MRC5. Цю суміш інкубують при 4 °C протягом 1 години. Зрештою, ї переносять у маленьку колбу Costar та інкубують при 35 °C/5% CO2 і через десять днів збирають рекомбінантні віруси кору-малярії для очищення. Приклад 14: Продукування комбінованої вакцини проти кору та малярії Робочий посівний матеріал описаного рекомбінантного вірусу кору-малярії інкубують на 2 моношарі клітин MRC5 у ролер-флаконах на 1750 см при 35 °С протягом десяти днів. Клітини кожного дня перевіряють на стан здоров’я та злиття. В день десять при найвищому рівні створення синцитій, супернатант відкачують у сталевий циліндр для зберігання в рідкому азоті. Цю ж саму процедуру повторюють через два дні. Після проведення усіх тестів (титр вірусу, стабільність геному, безпечність вірусу, безпечність клітин, хімічний аналіз, стерильність та нші), зібраний матеріал відтаюють та змішують із стабілізатором, що містить желатин, сорбіт, 5 амінокислоти та інші цукри, до кінцевого розведення 10 . За допомогою автоматичної машини для наповнення інокулюють дві маленькі пляшки для ліофілізації (F3) по 0,5 мл кожну. Використовують спеціально розраховану програму ліофілізації для гарантії максимального виживання продукту в процесі сушіння виморожуванням. Бібліографія 1. Fields Virology, fifth edition (2007), eds.-in-chief Knipe, D.M. &. Howley, P.M. Lippincott Williams & Wilkins, Philadelphia PA 19106, USA. 2. Enders, J.F., and Peebles, T.C. (1954). Propagation in tissue cultures of cytopathogenic agents from patients with measles. Proc. Soc. Exp. Biol. Med., 86: 277-286. 3. Griffin, D. (2007) Measles virus. In: Fields Virology, fifth edition, eds.-in-chief Knipe, D.M. &. Howley, P.M. Lippincott Williams & Wilkins, Philadelphia PA 19106, USA. 4. Parks, C.L., Lerch, R.A., Walpita, P., Wang, H.P., Sidhu, M.S., and Udem, S.A. (2001). Analysis of the noncoding regions of measles virus strains in the Edmonston vaccine lineage. J. Virol., 75: 921933. 5. Parks, C.L., Lerch, R.A., Walpita, P., Wang, H.P., Sidhu, M.S., and Udem, S.A. (2001). Comparison of predicted amino acid sequences of measles virus strains in the Edmonston vaccine lineage. J. Virol., 75: 910-920. 6. Hilleman, M.R. (2002). Current overview of the pathogenesis and prophylaxis of measles with focus on practical implications. Vaccine, 20: 651-665. 7. Ovsyannikova I.G., Reid, K.C., Jacobson, R.M., Oberg, A.L., Klee, G.G., Poland, G.A. (2003). Cytokine production patterns and antibody response to measles vaccine. Vaccine, 21(25-26): 394653. 8. Radecke, F., P. Spielhofer, H. Schneider, K. Kaelin, M. Huber, K. Dotsch, G. Christiansen, and M. Billeter. (1995). Rescue of measles viruses from cloned DNA. EMBO Journal., 14: 5773-5784. 9. Martin, A., Staeheli, P. and Schneider, U. (2006). RNA polymerase H-controlled expression of antigenomic RNA enhances the rescue efficacies of two different members of the Mononegavirales independently of the site of viral genome replication. J. Virol., 80:5708-5715. 10. Radecke, F., and M. Billeter. (1997). Reverse genetics meets the nonsegmented negativestrand RNA viruses. Rev. Med. Virol., 7: 49-63. 11. Singh M.R., Cattaneo, R., Billeter, M.A. (1999). A recombinant measles virus expressing hepatitis B virus surface antigen induces humoral immune responses in genetically modified mice. J. Virol., 73: 4823-4828. 12. Wang, Z.L., Hangartner, L., Cornu, T.I., Martin, L. R., Zuniga, A., and Billeter, M. (2001). Recombinant measles viruses expressing Deterologous antigens of mumps and simian immunodeficiency viruses. Vaccine, 19: 2329-2336. 13. Dilraj, Α., Cutts, F.T., de Castro, J.F., Wheeler, J.G., Brown, D., Roth, C, Coovadia, H.M., Bennett, J.V. (2000). Response to different measles vaccine strains given by aerosol and subcutaneous routes to schoolchildren: a randomised trial. Lancet, 355(9206): 798-803. 14. Holder A.A. and Freeman, R.R. (1984). The three major antigens on the surface of Plasmodium falciparum merozoites are derived from a single high molecular weight precursor. J. Exp. Med, 160(2): 624-9. 15. Blackman M.J., Whittle H., and Holder A.A. (1991). Processing of the Plasmodium falciparum major merozoite surface protein-1: identification of a 33-kilodalton secondary processing product which is shed prior to erythrocyte invasion. MoI. Biochem. Parasitol., 49(1): 35-44. 16. Remarque, E.J., Faber, B.W., Kocken, CH.M., and Thomas, A.W. (2007). Apical membrane antigen 1: a malaria vaccine candidate in review. Trends Parasitol, 24: 74-83. 21 UA 104180 C2 5 10 15 20 17. Polley, S.D., Mwangi, T., Kocken, C.H., Thomas, A.W., Dutta, S., Lanar, D.E., Remarque, E., Ross, A., Williams, T.N., Mwambingu, G., Lowe, B., Conway, D.J., and Marsh, K. (2004). Human antibodies to recombinant protein constructs of Plasmodium falciparum apical membrane antigen 1 (AMA1) and their association with protection from malaria. Vaccine, 23: 718-728. 18. Cortes, A., Mellombo, M., Masciantonio, R., Murphy, V.J., Reeder, J.C., and Anders, R.F. (2005). Allele specificity of naturally acquired antibody responses against Plasmodium falciparum apical membrane antigen 1. Infect. Immun., 73: 422-430. 19. Remarque, E.J., Faber, B.W., Kocken, C.H.M., and Thomas, A.W. (2008). A diversity-covering approach to immunisation with Plasmodium falciparum AMA1 induces broader allelic recognition and growth inhibition responses in rabbits. Infect. Immun. 20. Garcia, J.E., Puentes, A., and Patarroyo, M.E. (2006). Developmental biology of sporozoitehost interactions in Plasmodium falciparum malaria: implications for vaccine design. Clin. Microbiol. Rev., 19(4): 686-707. 21. Ballou, W.R., and Cahill, CP. (2007). Two Decades of Commitment to Malaria Vaccine Development: GlaxoSmithKline Biologicals. Am. J. Trop. Med. Hyg., 77(6_Suppl): 289-295. 22. Girard, M.P, Reed, Z.H., Friede, M., and Kieny, M.P. (2007). A review of human vaccine research and development: Malaria. Vaccine, 25: 1567-1580. 23. Roux, L., Simon, A.E., Holland, J.J. (1991). Effects of Defective Interfering Viruses on virus replication and pathogenesis in vitro and in vivo. Adv. Virus Res., 40: 181-221. 24. Calain, P., and Roux, L. (1988). Generation of measles virus defective interfering particles and their presence in a preparation of attenuated live-virus vaccine. J. Virol., 62 (8):2859-2866. ФОРМУЛА ВИНАХОДУ 25 30 35 40 45 50 55 1. Комбінована вакцина проти кору та малярії, яка включає рекомбінантну вірусну вакцину кору, що експресує антигени малярії, здатні викликати імунну відповідь та захист проти як кору, так і малярії. 2. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує один або різні антигени малярії. 3. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії. 4. Комбінована вакцина проти кору та малярії за п. 1, у якій антиген MSP1 малярії становить від 190 до 200 кДа (d190). 5. Комбінована вакцина проти кору та малярії за п. 1, у якій антигеном MSP1 малярії є d83-3038. 6. Комбінована вакцина проти кору та малярії за п. 1, у якій антигеном MSP1 малярії є d42. 7. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах. 8. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах штаму 3D7 та MAD 20, краще, штаму FCB1. 9. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору експресує антиген MSP1 малярії як у закріпленій, так і в секретованій формах штаму FCB1. 10. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує Diversity Covering (DiCo) антигену малярії ΑΜΑ1. 11. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-1 антигену малярії АМА1. 12. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-2 антигену малярії АМА1. 13. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-3 антигену малярії АМА1. 14. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує DiCo-1, DiCo-2 та DiCo-3 антигену малярії АМА1. 15. Комбінована вакцина проти кору та малярії за будь-яким з пп. 10-14, у якій рекомбінантна вірусна вакцина кору експресує Diversity Covering (DiCo) антигену малярії АМА1 у трансмембранній та секретованій формах. 22 UA 104180 C2 5 10 15 20 25 30 35 40 45 50 55 60 16. Комбінована вакцина проти кору та малярії за п. 1, у якій рекомбінантна вірусна вакцина кору експресує антиген малярії CS. 17. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій антиген малярії клонований між білками Ρ та Μ або Η та L рекомбінантної вірусної вакцини кору. 18. Вектор вірусної вакцини кору, що містить нуклеотидну послідовність антигену малярії з будьякого з пунктів 1-17 формули винаходу. 19. Вектор за п. 18, у якому нуклеотидну послідовність вибирають з наведених SEQ ID NO: 3-12. 20. Вектор за будь-яким з пп. 18 та 19, який додатково включає нуклеотидну послідовність, вибрану з наведених SEQ ID NO: 1-2. 21. Вектор за п. 18, у якому нуклеотидна послідовність кодує антигени малярії, вибрані з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS. 22. Хазяїн, який включає вектор за п. 18. 23. Хазяїн за п. 22, вибраний з Е. coli або клітинної лінії ссавця. 24. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантний вірус кору походить від штаму вакцин, одержаного з Edmonston Zagreb. 25. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантну вірусну вакцину кору, що експресує щонайменше один антиген малярії, вибирають з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS. 26. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантну вірусну вакцину кору, що експресує два чи більше антигенів малярії, вибирають з фрагментів d83-30-38 та d42 та d190 MSP1 або Diversity Covering (DiCo) AMA1 або білка CS або їх комбінації. 27. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору включає послідовності SEQ ID NO: 3-12. 28. Комбінована вакцина проти кору та малярії за будь-яким з попередніх пунктів формули, у якій рекомбінантна вірусна вакцина кору кодує, на додаток до антигенів малярії, білок з ад'ювантними властивостями. 29. Вакцина за будь-яким з попередніх пунктів формули, яка включає рекомбінантні віруси кору, що кодують, на додаток до антигенів малярії, інтерлейкін, краще інтерлейкін 2. 30. Вакцина за будь-яким з попередніх пунктів формули, що включає один з описаних рекомбінантних вірусів кору-малярії або суміш від двох до декількох таких вірусів. 31. Вакцина за будь-яким з попередніх пунктів формули, у якій описані рекомбінантні віруси кору-малярії або суміш від двох до декількох таких вірусів позбавлені дефектних інтерферуючих частинок (DI). 32. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють методом очищення бляшок. 33. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють методом кінцевого розведення. 34. Вакцина за будь-яким з попередніх пунктів формули, у якій випадково утворювані DI частинки видаляють фізичними методами, такими як диференційне центрифугування. 35. Вакцина за будь-яким з попередніх пунктів формули, яка є компонентом комбінованої вакцини, де іншими компонентами є краснуха, епідемічний паротит, вітряна віспа або інша жива атенуйована вірусна вакцина, природно атенуйована або рекомбінантна, сама або в комбінації. 36. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор, такий як желатин та/або альбумін сироватки людини і сорбіт як основні компоненти, ідеально, для парентерального застосування. 37. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для інтраназального застосування. 38. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для застосування шляхом інгаляції. 39. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для перорального застосування. 40. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для трансдермального застосування. 41. Вакцина за будь-яким з попередніх пунктів формули, яка включає придатний стабілізатор та/або ад'ювант, ідеально, для будь-якої композиції супозиторія. 42. Склад комбінованої вакцини проти кору-малярії, як заявлено в будь-якому попередньому пункті, що містить стабілізатор та/або ад'ювант. 23 UA 104180 C2 24 UA 104180 C2 25 UA 104180 C2 26 UA 104180 C2 27 UA 104180 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюCombined measles-malaria vaccine
Автори англійськоюGlueck, Reinhard, Fazio, Agata, Gianino, Viviana, Billeter, Martin
Автори російськоюГлюек Реинхард, Фазио Агата, Гианно Вивиана, Биллетер Мартин
МПК / Мітки
МПК: A61K 39/165, A61K 39/295, A61K 39/015, A61P 33/06
Мітки: кору, комбінована, вакцина, малярії
Код посилання
<a href="https://ua.patents.su/63-104180-kombinovana-vakcina-proti-koru-ta-malyari.html" target="_blank" rel="follow" title="База патентів України">Комбінована вакцина проти кору та малярії</a>
Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, вакцина проти багатосистемного синдрому виснаження свиней (бсвс) та набір для вакцинації

Номер патенту: 72891
Опубліковано: 16.05.2005
Автори: Мак-Нейлі Френсіс, Краковка Джордж Стівен, Елліс Джон Алберт, Одонне Жан-Крістоф Франсіс, Аллан Гордон Мур, Міхен Брайан Мартін
МПК: A61P 31/12, A61P 37/04, C07K 14/01, A61P 31/22, A61P 43/00, C12N 7/00, A61K 39/295
Мітки: цирковірусу, препарат, бсвс, парвовірусу, викликає, синдрому, імуногенний, вакцинації, багатосистемного, вакцина, відповідь, свиней, імунну, виснаження, набір
Формула / Реферат:
1. Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, що включає щонайменше один антиген цирковірусу свиней та щонайменше один антиген парвовірусу свиней у розчиннику або наповнювачі, прийнятному з ветеринарної точки зору.2. Імуногенний препарат за п.1, що включає антиген цирковірусу свиней типу II.3. Імуногенний препарат за п.2, у якому антиген цирковірусу свиней типу II вибраний з...
Вакцина абацилярна антракол проти сибірки тварин

Номер патенту: 52796
Опубліковано: 15.01.2003
Автори: Завірюха Анатолій Іванович, Завірюха Ганна Анатолієвна
МПК: A61K 39/07, A61K 39/00, A61K 39/02
Мітки: абацилярна, антракол, сибірки, вакцина, тварин
Формула / Реферат:
Вакцина абацилярна проти сибірки тварин, яка містить продукти культивування вакцинного штаму збудника сибірки Вас. anthracis K-79Z, яка відрізняється тим, що титр сибіркового білка у продуктах культивування вакцинного штаму збудника сибірки складає 1:2-1:512, причому вакцина додатково містить продукти культивування вірулентного штаму Е.coli при такому співвідношенні компонентів, %:культуральна рідина після культивування Вас. anthracis...
Композиція вакцини проти малярії та спосіб лікування або попередження захворювання малярією з використанням цієї композиції

Номер патенту: 68336
Опубліковано: 16.08.2004
Автор: Кохен Джозеф
МПК: A61K 39/29, A61K 39/015, A61K 39/39, A61K 39/002
Мітки: спосіб, композиції, лікування, попередження, використанням, малярією, малярії, вакцини, цієї, композиція, захворювання
Формула / Реферат:
1. Композиція протималярійної вакцини, що містить принаймні два специфічні для малярії антигени, в комбінації з ад¢ювантом, що є вибірковим стимулятором відповіді ТН1 клітин, і де принаймні два малярійних антигени вибрані з групи, яка включає:а) гібридний протеїн, що включає всю С-термінальну ділянку циркумспорозоїтного (CS) протеїну, чотири або більше подвійних повторів імунодомінантної ділянки та поверхневий антиген вірусу...
Комбінована педіатрична вакцина, спосіб одночасної комбінованої вакцинації

Номер патенту: 27754
Опубліковано: 16.10.2000
Автори: Хекелл Джіл Дж., Хогерман Дебора А., Парадізо Пітер Р., Мадоре Дейс Віцепс
МПК: A61K 39/02, A61K 39/116
Мітки: комбінована, спосіб, комбінованої, одночасної, педіатрична, вакцина, вакцинації
Текст:
...композиция комбинированной вакцины настоящего изобретения является гомогенной белой суспензией. Кроме того, вакцинные компоненты стабильны и совместимы, и не проявляют взаимной подавляющей активности между компонентами. Также были найдены способы для одновременно комбинированной вакцинации младенцев и для повышения иммуногенности полисахарида Haemophilus influenzae типа b дифтерийного антигена, столбнячного антигена и коклюшного антигена,...
Асоційована вакцина проти інфекційного ринотрахеїту та вірусної діареї

Номер патенту: 7152
Опубліковано: 15.06.2005
Автори: Кассіч Володимир Юрійович, Герус Галина Борисівна, Волосянко Олена Вікторівна, Вовк Сергій Іванович, Чечоткіна Наталія Павлівна, Стегній Борис Тимофійович, Данілова Ірина Сергіївна
МПК: A61K 39/00
Мітки: інфекційного, асоційована, діареї, вакцина, вірусної, ринотрахеїту
Формула / Реферат:
Асоційована вакцина проти інфекційного ринотрахеїту та вірусної діареї, що містить віруси ІРТ та ВД, інактиватор та ад'ювант, яка відрізняється тим, що використовують суміш суспензій інактивованих вірусів штам "IPT-LG" та штам "ВД-М", а як ад'ювант використовують масло вакцинне синтетичне (типу Маrсоl 52 або МИ), як інактиватор - уротропін та ще додають емульгатор 139, при наступному співвідношенні компонентів, мас. %:...
Попередній патент: Пристрій для визначення відхилень прямовисних ліній
Наступний патент: Завантажувальна установка шахтної печі з приводним механізмом для розподільного лотка та шахтна піч, що її містить
Випадковий патент: Плоский металодіелектричний хвилевід