Стійка до засухи рослина
Номер патенту: 113503
Опубліковано: 10.02.2017
Автори: Маліт Джон, Борел Ендрю Кенет, Клейн Патрисія, Джордан Девід Роберт






Формула / Реферат
1. Спосіб отримання генетично модифікованої рослини, яка використовує воду ефективніше, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує локус Stg1 або Stg2 з сорго, який асоційований з фенотипом підтримування зеленим, який включає перемикання використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури, або збільшення ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеженого доступу води.
2. Спосіб за п. 1, де локус Stg1 кодує білок суцвіття в формі шпильки (PIN), вибраний з SbPIN3, SbPIN4 або SbPIN10.
3. Спосіб за п. 1, де локус Stg2 кодує білок PIN, який являє собою SbPIN2.
4. Спосіб за п. 1, де локус Stg1 кодує білок PIN, який являє собою SbPIN4.
5. Спосіб за п. 1, де генетичний агент являє собою область геному рослини, вибрану так, щоб вона мала конкретний профіль експресії PIN.
6. Спосіб за п. 1, де генетично модифікована рослина являє собою рослину сорго.
7. Спосіб за п. 1, де генетично модифікована рослина вибрана з пшениці, вівса, маїсу, жита і рису.
8. Спосіб за п. 1, де генетично модифікована рослина вибрана з абаки, люцерни, мигдалю, яблука, спаржі, банана, квасолі, чорної смородини, кормових бобів, каноли, кеш'ю, маніоки, гороху баранячого, цитруса, кокоса, кави, кукурудзи, бавовни, фіги, льону, винограду, земляного горіха, конопель, кенафу, лаванди, манго, грибів, оливок, цибулі, гороху, арахісу, груші, проса, картоплі, рами, рапсу, плевелів, сої, полуниці, цукрового буряка, цукрової тростини, соняшника, солодкої картоплі, колоказії, чаю, тютюну, томата, тритикале, трюфеля і ямсу.
9. Спосіб за п. 1, де фенотип підтримування зеленим додатково включає фенотип, вибраний із збільшеної пластичності архітектури листяного покриву, зменшеного розміру листяного покриву, збільшеної біомаси на одиницю площі листка під час цвітіння, вищої ефективності транспірації, збільшеного використання води в ході наливання зерна, зниженої продукції біомаси до і після цвітіння і уповільненого старіння.
10. Спосіб за п. 1, де фенотип підтримування зеленим додатково включає більший розмір зерен.
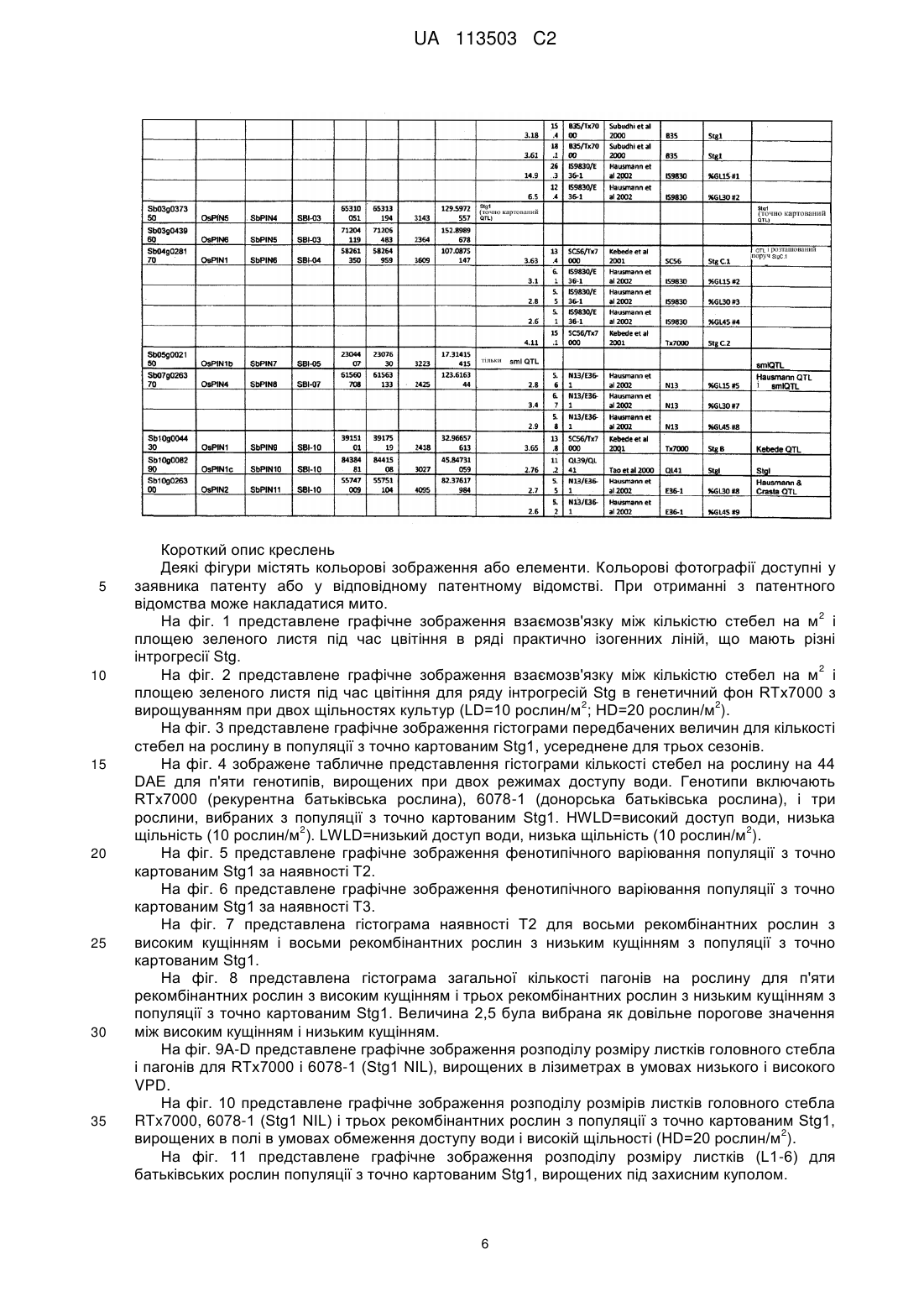
Текст
Реферат: Винахід належить до способу отримання генетично модифікованої рослини, яка використовує воду ефективніше, ніж не модифікована генетично рослина того ж виду. Спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує локус Stg1 або Stg2 з сорго, який асоційований з фенотипом підтримування зеленим, який включає перемикання використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури або збільшення ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеженого доступу води. UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 Дата подачі Дана заявка пов'язана з тимчасовою заявкою США № 61/413902, поданою 15 листопада 2010 року, повний зміст якої включений в даний опис як посилання. Галузь техніки У даному описі описано отримання стійких до засухи рослин. Даний винахід дозволяє маніпулювання фенотипічною ознакою, званою в даному описі "підтримування зеленим", для отримання стійких до засухи рослин рекомбінантними способами, способами мутагенезу і/або способами схрещування і селекції. Також в даному описі описані системи практики керування обробкою культур для підвищення виходу культури і ефективності збирання урожаю в умовах обмеженого доступу води. Рівень техніки Бібліографічні деталі публікацій, що указуються автором в даний заявці, зібрані в алфавітному порядку в кінці опису. Посилання на будь-який із документів рівня техніки в даному описі не є і не повинне тлумачитися як визнання або яка-небудь форма припущення, що цей документ рівня техніки формує частину загальної інформації в якій-небудь країні. Збільшення людської популяції вимагає збільшення врожайності сільськогосподарських культур. Це є головною метою для рослинників і генетиків рослин. Один з підходів підвищення врожайності сільськогосподарських культур залучає селекцію ознак рослин, які сприяють вищому виходу зерна і стійкості (Springer, Nature Genetics 42:475-476, 2010). Цей підхід був названий "зеленим революцією". Інші підходи включають розвиток архітектури рослин, який привів, наприклад, до ідентифікації локусу кількісних ознак (QTL), який кодує білок, подібний білку, зв'язуючому промотор лусочок 14 (OsSPL14) в рисі, і який сприяє збільшенню виходу рису (Jiao et al., Nature Genetics 42:541-544, 2010; Miura et al., Nature Genetics 42:545-549, 2010). Засуха являє собою одне з найважливіших обмежень продукції злаків по всьому світу. Сорго є сховищем механізмів стійкості до засухи, які включають C 4-фотосинтез, глибоке коріння і товстий віск листя, які забезпечують ріст в жарких і сухих умовах. Стійкість до засухи робить сорго особливо важливим в сухих регіонах, таких як країни Африки південніше Сахари, західноцентральна Індія, північно-східна Австралія і південні рівнини США. Враховуючи зростаючий тиск на доступність бідних водних ресурсів, ідентифікація ознак, асоційованих з виходом зерна в умовах засухи, стає більш важливою. Механізм адаптації до засухи, ідентифікований в сорго, який приводить до збереження листя зеленими протягом більш тривалих періодів часу в ході наливання зерна в умовах засухи, відомий як "підтримування зеленим". Підтримування зеленим асоційовано з високим виходом зерна при засусі після цвітіння в сорго (Borrell et al., Crop Sci. 40:1037-1048, 2000b; Kassahun et al., Euphytica 72:351-362, 2010), пшениці (Triticum aestivum L.) [Spano et al., J. Exp. Bot. 54:14151420, 2003; Christopher et al., Aust. J. Agric. Res. 59:354-364, 2008], рисі (Oryza sativa L.) [Kashiwagi et al., Plant Physiology and Biochemistry 44:152-157, 2006] і кукурудзі (Zea mays L.) [Zheng et al., Plant Breed 128:54-62, 2009]. Крім того, воно може прямо впливати на вихід зерна в умовах засухи шляхом підвищення стійкості до вугільної гнилі (Macrophomina phaseolina [Tassi] Goid.) (Tenkouano et al., Theor. Appl. Genet. 85:644-648, 1993; Garud et al., Int. Sorghum and Millets Newsl. 43:63-65, 2002). Це зменшує вилягання (Reddy et al., Euphytica 159:191-198, 2008), що дозволяє рослинникам використовувати позитивну асоціацію між висотою рослини і виходом зерна (Jordan et al., Theor. Appl. Genet. 106:559-567, 2003). Підтримування зеленим є важливим критерієм селекції для програм виведення сорго, націлених на адаптацію до засухи як в США (Rosenow et al., Agric. Water Manag. 7:207-222, 1983), так і в Австралії (Henzell et al., Australia Int. Sorghum and Millets Newsl. 38:1-9, 1997). На підтримку цієї ознаки отримані істотні докази (Borrell et al., Crop Sci. 40:1026-1037, 2000a; Borrell and Hammer, Crop Sci. 40:1295-1307, 2000; Harris et al., J. Exp. Bot. 58:327-338, 2007; Christopher et al., 2008, вище; Van Oosterom et al., Field Crops Res. 115:19-28, 2010a і Van Oosterom et al., Field Crops Res. 115:29-38, 2010b). Хоча цей механізм стійкості до засухи використовується фахівцями з розведення сорго в США і Австралії протягом 25 років, і широка фізіологічна основа цієї ознаки стає більш зрозумілою, причинні механізми і генетичні локуси, залучені до нього, до цього часу невідомі. В умовах обмеження води вихід зерна є функцією транспірації (Т), ефективності транспірації (TE) і індексу урожаю (HI) [Passioura, J. Aust. Inst. Agric. Sci. 43:117-120, 1977]. У цих рамках, вихід зерна пов'язаний з Т періоду після цвітіння (Turner, J. Exp. Bot. 55:2413-2425, 2004; Manschadi et al., Funct. Plant. Biol. 33:823-837, 2006), оскільки HI зростає із ростом частки загальної Т культури, що використовується після цвітіння (Passioura, 1977, вище; Sadras and Connor, Field Crops Res. 26:227-239, 1991; Hammer, Agric. Sci. 19:16-22, 2006). Збільшена Т 1 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 після цвітіння асоційована зі зниженням стресу, що викликається засухою, що може позитивно впливати на швидкість росту культури злаків під час цвітіння і, таким чином, число зерен (Andrade et al., Crop Sci. 42:1173-1179, 2002; Van Oosterom і Hammer, Field Crops Res. 108:259268, 2008). Якщо загальна кількість доступної води обмежена, Т після цвітіння може збільшуватися шляхом обмеження Т до цвітіння. Цього можна досягати шляхом обмеження розміру листяного покриву або генетично, або шляхом керування обробкою культур. Однак зменшення листяного покриву знизить загальну Т, тільки якщо TE не буде порушена. Для сорго були описані значні генотипічні відмінності в TE (Hammer et al., Aust. J. Agric. Res. 48:649-655, 1997; Henderson et al., Aust. J. Plant Physiol. 25:111-123, 1998; Mortlock and Hammer, J. Crop Prod. 2:265-286, 1999; Xin et al., Field Crops Res. 111:74-80, 2009). Альтернативно використання води після цвітіння може збільшуватися при збільшенні загальної кількості води, доступної для культури, або шляхом більш глибокого укорінення, або шляхом зниження нижньої межі витягання води (Manschadi et al., 2006, вище). Ознака підтримування зеленим впливає на ряд із вказаних вище процесів в сорго. Поперше, підтримування зеленим знижує споживання води в період до цвітіння шляхом обмеження розміру листяного покриву (внаслідок зменшення кущіння і зменшення розміру листя). По-друге, підтримування зеленим підвищує засвоюваність води шляхом збільшення співвідношення коріння:пагони. Існують деякі експериментальні дані про краще витягання води в лініях з ознакою підтримування зеленим, хоча потрібно більше досліджень. Ці кореневі відповіді також можуть бути пояснені посиленим транспортом ауксину (Wang et al., Molecular Plant 2(4):)(823-831, 2009). По-третє, підтримування зеленим збільшує зеленість листя під час цвітіння, ефективно збільшуючи здатність до фотосинтезу, і, таким чином, TE (за умови, що фотосинтез зростає пропорційно провідності). Збільшення зеленості листя є непрямим ефектом зниження маси листя, тобто в листках концентрується азот. Виробництво більшої кількості продуктів живлення при меншій кількості води є одним з основних завдань, які в цей час стоять перед людством. Існує реальна і невідкладна потреба як в країнах, що розвиваються, так і в розвинених країнах, в ідентифікації генів і генних мереж, контролюючих адаптацію до засухи в сільськогосподарських рослинах. Це забезпечує збільшення адаптації до засухи в широкому діапазоні видів рослин, які вирощуються в оточуючих умовах з обмеженим доступом води по всьому світу. Суть винаходу Даний винахід стосується локусів кількісних ознак (QTL), асоційованих з фенотипом підтримування зеленим і/або іншим чином сприяючих йому. QTL позначають в даному описі як "підтримування зеленим (Stg) X", де X являє собою число, що перевищує 1, яке відповідає області на хромосомі, асоційованій з фенотипом підтримування зеленим. Як описано в даному описі, QTL ідентифікують генетичні області на сорго, які несуть локуси, кодуючі від однієї до деякої кількості білків або регуляторних агентів, таких як мікроРНК, які сприяють фенотипу підтримування зеленим. Модулювання експресії одного або декількох цих локусів в сільськогосподарській рослині забезпечує архітектуру листяного покриву, який сприяє переключенню використання води рослиною на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури або збільшенню ефективності транспірації, тим самим, збільшенню індексу урожаю (HI) і виходу зерна в умовах обмеженого доступу води. У одному варіанті здійснення локуси кодують білок PIN, який асоційований з ауксином. Білки PIN являють собою ефлюксні переносники ауксину, які містять трансмембранні домени і, головним чином, локалізуються в плазматичних мембранах. Термін "PIN" походить від "суцвіття в формі шпильки". Термін "SbPINn" використовують для опису такого гена в сорго, де n являє собою число, що визначає компонент ефлюксного переносника ауксину, і n дорівнює від 1 до 11. Вказівка на "SbPINn" включає його гомологи і ортологи з інших рослин. Приклади локусів SbPINn в сорго включають локуси, наведені в таблиці 1A, такі як, але не обмежуючись ними, SbPIN4 і SbPIN2, або їх еквіваленти в інших рослинах. У даному описі описано, що модулювання експресії PIN або експресія PIN з конкретною поліморфною зміною сприяють вияву фенотипу підтримування зеленим. Даний винахід стосується PIN з інших рослин, таких як рис. Буквений префікс у PIN визначає його джерело (наприклад Sb - сорго; Os - рис і т. д.). Положення SbPIN в сорго визначається ID-номером гена (див. таблицю 1A). Як приклади, SbPIN4 відповідає OsPIN5, і SbPIN2 відповідає OsPIN3a. SbPIN4 і SbPIN2 є прикладами SbPIN, описаних в даному описі, відповідальних за ознаку підтримування зеленим в сорго, асоційовану з Stg1, і Stg2, відповідно, що приводить до ряду 2 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 фенотипів, які забезпечують адаптацію до засухи шляхом зниженого використання води до цвітіння (внаслідок зменшення кущіння і меншого розміру листя), збільшеної засвоюваності води (внаслідок збільшення співвідношення коріння:пагони), збільшеної ефективності транспірації при м'якому дефіциті води (внаслідок вищої концентрації азоту в листках), збільшеної біомаси на площу листка при термінальному дефіциті води (внаслідок збільшення транспірації на площу листка), і збільшений вихід зерна, розмір зерен і стійкість до вилягання. Інші приклади наведені в таблиці 1A і включають їх еквіваленти в інших рослинах. Даний винахід стосується способу отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає модулювання рівня експресії існуючого або внесеного локусу суцвіття в формі шпильки (PIN) у всіх або в окремій тканині рослини для сприяння фенотипу підтримування зеленим, який включає переключення використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури або збільшення ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеженого доступу води. У даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, що кодує білок PIN, вибраний із SbPIN1-11 або їх еквівалента з іншої рослини або їх функціонального гомолога або ортолога; або модулюючого експресію власного білка PIN; де рівень і місце експресії PIN сприяють фенотипу підтримування зеленим, який включає, серед інших, архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури, що приводить до збільшеного індексу урожаю і виходу зерна в умовах обмеження доступу води. Крім того, даний винахід стосується способу отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує білок PIN сорго, вибраний із SbPIN4 і SbPIN2 або їх еквівалента з іншої рослини, або який модулює експресію власного PIN. Вказівка на модулювання включає збільшення і зниження рівня експресії. Більше того, може бути вибраний PIN, що має бажаний профіль експресії у всіх або окремих тканинах в рослині. У даному описі передбачений спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує продукт, асоційований або сприяючий фенотипу підтримування зеленим, що включає переключення використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури або збільшення ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеженого доступу води, і де продукт вибраний із переліку, що складається з SbPIN1-11 або їх еквівалентів з іншої рослини, або який модулює експресію власного PIN в рослині. Також в даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду ефективніше, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує білок, вибраний із списку, що складається з SbPIN4 і SbPIN2, або їх еквівалентів із іншої рослини, або який модулює експресію власного PIN в рослині. Термін "введення" включає введення шляхом рекомбінантного втручання, а також за допомогою протоколу мутагенезу або схрещування з подальшою селекцією. У даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину і/або батьківську рослину генетичного агента, який кодує два або більше білків PIN або який модулює експресію двох або більше власних білків PIN в рослині. Приклади PIN в сорго включають SbPIN1-11, такі як SbPIN4 і SbPIN2, або їх еквіваленти з іншої рослини. Наприклад, SbPIN4 відповідає OsPIN5, і SbPIN2 відповідає OsPIN3a. SbPIN і OsPIN визначені в таблиці 1A. Генетично модифіковані рослини і їх нащадки, що виявляють ознаку підтримування зеленим, також передбачені в рамках даного винаходу, також як і насіння, плоди і квітки і інший репродуктивний матеріал або матеріал розмноження. У рамках даного винаходу передбачені як генетичний матеріал, який кодує білок PIN або його функціональний гомолог або ортолог, який асоційований або сприяє фенотипу 3 UA 113503 C2 5 10 15 20 25 30 35 підтримування зеленим, який включає архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури або збільшенню ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеження доступу води, так і агент, який модулює експресію PIN. У одному прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN4; і (ii) агента, який модулює рівні експресії SbPIN4 або його еквівалента з іншої рослини. У іншому прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN2; і (ii) агента, який модулює експресію SbPIN2 або його еквівалента з іншої рослини. Генетичний матеріал включає агент, який кодує білок SbPIN4 або SbPIN2 або їх функціональний гомолог або ортолог, або їх еквівалент із іншої рослини, який асоційований або сприяє фенотипу підтримування зеленим, що включає архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури, або збільшенню ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеження доступу води; і агент, який модулює експресію SbPIN4 або SbPIN2 або їх еквівалента з іншої рослини. У даному описі описана система керування рослинництвом для зниження залежності від води або іншого підвищення ефективності використання води і збільшення виходу зерна або продукту. Система керування рослинництвом включає отримання адаптованої до засухи сільськогосподарської культури, що включає злакові рослини, з використанням вибору і модулювання експресії локусу PIN або його функціонального еквівалента, як визначено в даному описі, окремо або в комбінації з введенням інших корисних ознак, таких як розмір зерна, розмір коріння, стійкість до солей, стійкість до гербіцидів, стійкість до шкідників і т. п. Альтернативно або додатково, система керування рослинництвом включає отримання адаптованих до засухи рослин і сільськогосподарські процедури, такі як зрошування, вимоги до поживних речовин, щільність і геометрія насадження культури, боротьба з бур'янами, боротьба з комахами, аерація ґрунту, зменшення кущіння, підняття грядок і т. п. Приклади локусу PIN включають SbPIN1-11 (таблиця 1A), такі як SbPIN4 і SbPIN2, і їх еквіваленти з інших рослин. Також в даному описі описана комерційна модель для підвищення економічної віддачі відносно виходу культури, причому модель включає отримання сільськогосподарських рослин, що мають архітектуру листяного покриву, який сприяє переключенню використання води рослиною на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури, або збільшення ефективності транспірації, тим самим, збільшуючи HI і вихід зерна в умовах обмеженого доступу води, отримання насіння отриманої сільськогосподарської рослини і розподіл насіння виробникам зерна для підвищення виходу і прибутку. У даному описі використовуються наступні скорочення: CI довірчий інтервал, CWU використання води культурою, DW маса сухої речовини, GLA площа зеленого листка, HD висока щільність, HI індекс урожаю, HT високе кущіння, HW високий доступ води, HWH високий доступ води, висока щільність (проміжний дефіцит води), D HWL високий доступ води, низька щільність (найменший дефіцит води), D LA площа листка, LD низька щільність, LT низьке кущіння, LW низький доступ води, LWH низький доступ води, висока щільність (найбільший дефіцит води), D LWL низький доступ води, низька щільність (проміжний дефіцит води), D NIL практично ізогенна лінія, OsPI PIN з рису, 4 UA 113503 C2 N PAB PAS біомаса після цвітіння, маса стебел після цвітіння, PIN PPB суцвіття в формі шпильки, співвідношення біомаси до:після цвітіння, QTL ROS RWC SbPI локус кількісних ознак, укриття від дощу, відносний вміст води, PIN з сорго, M R N SLW питома маса листя, SML навчання статистичної машини, Stg підтримування зеленим, Т транспірація, T2 пагін в пазусі листка 2, T3 пагін в пазусі листка 3, T4 пагін в пазусі листка 4, T5 пагін в пазусі листка 5, T6 пагін в пазусі листка 6, TE ефективність транспірації, TS термінальний стрес, VPD дефіцит тиску пари, WW що добре поливається. У таблиці 1A представлена інформація про PIN з сорго і рису. 5 UA 113503 C2 5 10 15 20 25 30 35 Короткий опис креслень Деякі фігури містять кольорові зображення або елементи. Кольорові фотографії доступні у заявника патенту або у відповідному патентному відомстві. При отриманні з патентного відомства може накладатися мито. 2 На фіг. 1 представлене графічне зображення взаємозв'язку між кількістю стебел на м і площею зеленого листя під час цвітіння в ряді практично ізогенних ліній, що мають різні інтрогресії Stg. 2 На фіг. 2 представлене графічне зображення взаємозв'язку між кількістю стебел на м і площею зеленого листя під час цвітіння для ряду інтрогресій Stg в генетичний фон RTx7000 з 2 2 вирощуванням при двох щільностях культур (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 3 представлене графічне зображення гістограми передбачених величин для кількості стебел на рослину в популяції з точно картованим Stg1, усереднене для трьох сезонів. На фіг. 4 зображене табличне представлення гістограми кількості стебел на рослину на 44 DAE для п'яти генотипів, вирощених при двох режимах доступу води. Генотипи включають RTx7000 (рекурентна батьківська рослина), 6078-1 (донорська батьківська рослина), і три рослини, вибраних з популяції з точно картованим Stg1. HWLD=високий доступ води, низька 2 2 щільність (10 рослин/м ). LWLD=низький доступ води, низька щільність (10 рослин/м ). На фіг. 5 представлене графічне зображення фенотипічного варіювання популяції з точно картованим Stg1 за наявності T2. На фіг. 6 представлене графічне зображення фенотипічного варіювання популяції з точно картованим Stg1 за наявності T3. На фіг. 7 представлена гістограма наявності T2 для восьми рекомбінантних рослин з високим кущінням і восьми рекомбінантних рослин з низьким кущінням з популяції з точно картованим Stg1. На фіг. 8 представлена гістограма загальної кількості пагонів на рослину для п'яти рекомбінантних рослин з високим кущінням і трьох рекомбінантних рослин з низьким кущінням з популяції з точно картованим Stg1. Величина 2,5 була вибрана як довільне порогове значення між високим кущінням і низьким кущінням. На фіг. 9A-D представлене графічне зображення розподілу розміру листків головного стебла і пагонів для RTx7000 і 6078-1 (Stg1 NIL), вирощених в лізиметрах в умовах низького і високого VPD. На фіг. 10 представлене графічне зображення розподілу розмірів листків головного стебла RTx7000, 6078-1 (Stg1 NIL) і трьох рекомбінантних рослин з популяції з точно картованим Stg1, 2 вирощених в полі в умовах обмеження доступу води і високій щільності (HD=20 рослин/м ). На фіг. 11 представлене графічне зображення розподілу розміру листків (L1-6) для батьківських рослин популяції з точно картованим Stg1, вирощених під захисним куполом. 6 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 На фіг. 12 представлене графічне зображення розподілу довжини листків (L1-6) для батьківських рослин популяції з точно картованим Stg1, вирощених під захисним куполом. Фігура 13 представлено графічне зображення розподілу ширини листків (L1-6) для батьківських рослин популяції з точно картованим Stg1, вирощених під захисним куполом. На фіг. 14 представлене графічне зображення розподілу розміру листків (l1-11) для батьківських рослин популяції з точно картованим Stg1, вирощених під захисним куполом. На фіг. 15 представлене графічне зображення розподілу довжини листків (L1-10) для батьківських рослин популяції з точно картованим Stg1, вирощених під захисним куполом. На фіг. 16 представлена гістограма фенотипічного варіювання довжини L10 в підгрупі рослин популяції з точно картованим Stg1, вирощених під захисним куполом. На фіг. 17 представлена діаграма, що показує, що збільшеної доступності води під час цвітіння досягають шляхом зниження використання води за допомогою двох механізмів (зниження кущіння і менший розмір листків) в рослинах, що містять область Stg1. На фіг. 18 показано, що розмір листяного покриву модулюється як конститутивною, так і адаптивною відповідями, що контролюються геном(ами) в області Stg1. На фіг. 19 представлене графічне зображення розподілу розміру листків головного стебла RTx7000, 6078-1 (Stg1 NIL) і трьох рекомбінантних рослин із популяції з точно картованим Stg1, 2 вирощених в полі в умовах обмеженого доступу води і високій щільності (HD=20 рослин/м ). На фіг. 20 представлене графічне зображення взаємозв'язку між площею листя 12 і загальною площею зеленого листя під час цвітіння для двох батьківських рослин (6078-1 і RTx7000) і трьох рекомбінантних рослин із популяції з точно картованим Stg1. На фіг. 21 представлене графічне зображення взаємозв'язку між загальною площею 2 2 зеленого листя (см /м ) і використанням води культурою (мм) під час цвітіння для двох батьківських рослин (6078-1 і RTx7000) і трьох рекомбінантних рослин із популяції з точно картованим Stg1. На фіг. 22 представлене графічне зображення взаємозв'язку між площею зеленого листя і використанням води (Т) в чотирьох рослинах з QTL Stg і рекурентній батьківській рослині (RTx7000) в лізиметричних дослідженнях при двох рівнях VPD. На фіг. 23 представлене графічне зображення гістограми фенотипічного варіювання "співвідношення коріння:пагони" для L6 в популяції з точно картованим Stg1, вирощеної під захисним куполом. На фіг. 24 представлене графічне зображення тимчасового профілю сукупного використання води культурою для RTx7000 і Stg1, вирощених в умовах низького доступу води і 2 низькій щільності (20 рослин/м ). Вертикальна лінія означає цвітіння. На фіг. 25 представлене графічне зображення взаємозв'язку між довжиною (мм) і зеленістю (SPAD) листя 10 в популяції з точно картованим Stg1, вирощеної під захисним куполом. На фіг. 26 представлене графічне зображення взаємозв'язку між зеленістю листя (SPAD) і фотосинтезом листя в підгрупі ліній з популяції з точно картованим Stg1, включаючи батьківські рослини. На фіг. 27 представлене графічне зображення взаємозв'язку між зеленістю листя (SPAD) і WUE (Licor) в підгрупі ліній із популяції з точно картованим Stg1, включаючи батьківські рослини. На фіг. 28 представлене графічне зображення взаємозв'язку між зеленістю листя (SPAD) і WUE (Licor) в чотирьох Stg Nil (Stg1, Stg2, Stg3 і Stg4) і рекурентній батьківській рослині (RTx7000). На фіг. 29 представлене графічне зображення взаємозв'язку між транспірацією на площу листя і ефективністю транспірації в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000) в лізиметричних дослідженнях при двох рівнях VPD. На фіг. 30 представлене графічне зображення взаємозв'язку між CWU (мм) до і після цвітіння в підгрупі ліній з популяції з точно картованим Stg1, включаючи батьківські рослини, вирощених в умовах високої щільності (HD) і низькій щільності (HD). На фіг. 31A і В представлено графічне зображення профілів сукупного використання води для Stg1 і RTx7000, вирощених в умовах LWHD. На фіг. 32 представлене графічне зображення взаємозв'язку між CWU (мм) до і після цвітіння в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах низького доступу води (LW) і низькій щільності (LD). На фіг. 33 представлене графічне зображення взаємозв'язку між PPBR і PAB в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). 7 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 На фіг. 34 представлене графічне зображення взаємозв'язку між GLAA і PPBR в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 35 представлене графічне зображення взаємозв'язку між GLAA і PASB в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 36 представлене графічне зображення взаємозв'язку між PPBR і PAB в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 37 представлене графічне зображення взаємозв'язку між PPBR і PASM в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 38 представлене графічне зображення взаємозв'язку між PPBR і виходом зерна в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого 2 2 доступу води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 39 представлене графічне зображення взаємозв'язку між RWC при середньому наливанні зерна (FL-2) і відносній швидкості старіння листя в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох 2 2 щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 40 представлене графічне зображення взаємозв'язку між відносною швидкістю старіння листя і площею зеленого листя в зрілому стані в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох 2 2 щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 41 представлене графічне зображення взаємозв'язку між відносним вмістом води (RWC) при середньому наливанні зерна (FL-2) і масою стебла в зрілому стані в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу 2 2 води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 42 представлене графічне зображення взаємозв'язку між масою стебла після цвітіння (PASM) і біомасою після цвітіння (PAB) в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях 2 2 культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 43 представлене графічне зображення взаємозв'язку між біомасою стебла після цвітіння і виходом зерна в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), 2 вирощених в умовах обмеженого доступу води при двох щільностях культури (LD=10 рослин/м ; 2 HD=20 рослин/м ). На фіг. 44 представлене графічне зображення взаємозв'язку між масою стебла після цвітіння (PASM) і біомасою після цвітіння (PAB) в чотирьох Stg1 QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях 2 2 культури, яка була вирощені в експерименті в 2005 році (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 45 представлене графічне зображення взаємозв'язку між масою стебла після цвітіння (PASM) і виходом зерна в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях культури (LD=10 2 2 рослин/м ; HD=20 рослин/м ), які були вирощені в експерименті в 2005 році. На фіг. 46 представлене графічне зображення взаємозв'язку між масою стебла після цвітіння (PASM) і біомасою після цвітіння (PAB) в різних лініях з точно картованим Stg1 і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при 2 2 двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 47 представлене графічне зображення взаємозв'язку між вмістом води (RWC) при середньому наливанні зерна (FL-2) і виходом зерна при різних комбінаціях Stg QTL і рекурентної батьківської рослини (RTx7000), вирощених в умовах обмеженого доступу води при 2 2 двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ) в експерименті з вирощування в 2004 році. На фіг. 48 представлене графічне зображення взаємозв'язку між водним потенціалом листя 2 (LWP) FL-2 при середньому наливанні зерна (бар) і виходом зерна (г/м ) в Stg1 QTL (6078-1) і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при 2 2 двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 49 представлене графічне зображення взаємозв'язку між PPBR і CWU в ході наливання зерна в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в 2 умовах обмеженого доступу води при двох щільностях культури (LD=10 рослин/м ; HD=20 2 рослин/м ). 8 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 На фіг. 50 представлене графічне зображення взаємозв'язку між CWU в ході наливання 2 зерна (мм) і виходом зерна (г/м ) в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях культури (LD=10 2 2 рослин/м ; HD=20 рослин/м ). На фіг. 51 представлене графічне зображення взаємозв'язку між PPBR і виходом зерна в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого 2 2 доступу води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 52 представлене графічне зображення взаємозв'язку між CWU в ході наливання зерна (мм) і розміром зерен (мг) в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях культури (LD=10 2 2 рослин/м ; HD=20 рослин/м ). На фіг. 53 представлене графічне зображення взаємозв'язку між PPBR і розміром зерен в чотирьох Stg QTL і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого 2 2 доступу води при двох щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 54 представлене графічне зображення взаємозв'язку між PPBR і CWU в ході наливання зерна в різних лініях з точно картованим Stg1 і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох щільностях культури (LD=10 2 2 рослин/м ; HD=20 рослин/м ). На фіг. 55 представлене графічне зображення взаємозв'язку між CWU в ході наливання 2 зерна (мм) і виходом зерна (г/м ) в різних лініях з точно картованим Stg1 і рекурентній батьківській рослині (RTx7000), вирощених в умовах обмеженого доступу води при двох 2 2 щільностях культури (LD=10 рослин/м ; HD=20 рослин/м ). На фіг. 56A-C представлене графічне зображення результатів дослідження імітаційної моделі культури сорго з використанням генетичного сорту Buster із звичайними 2 пагонами/рослиною (HT) проти Buster тільки з 1 пагоном/рослиною (LT) у віртуальних навколишніх умовах з хорошим доступом води (WW) і в умовах термінального стресу (TS). Для обох віртуальних навколишніх умов були вибрані наступні параметри: щільність посіву 5 2 рослин/м з відстанню між рядами 1 м; глибина ґрунту=1800 мм; PAWC ґрунту=324 мм; без обмежень N. На фіг. 57A представлене графічне зображення диференціальної експресії SbPIN4 (кандидат Stg1) в умовах хорошого доступу води. У умовах хорошого доступу води цей ген приглушується в молодих кореневих кінчиках Tx642 і Stg1 NIL в порівнянні з Tx7000. На фіг. 57B представлене графічне зображення диференціальної експресії SbPIN4 (кандидат за Stg1) в умовах дефіциту води. У умови дефіциту води цей ген активується в більшості тканин, але особливо в зростаючому листі Tx642 і Stg1 NIL в порівнянні з Tx7000. На фіг. 57C представлене графічне зображення диференціальної експресії SbPIN2 (кандидат за Stg2) в умовах хорошого доступу води. У умовах хорошого доступу води цей ген слабо активований в тканинах стебла і коріння Tx642 і Stg1 NIL в порівнянні з Tx7000. На фіг. 57D представлене графічне зображення диференціальної експресії SbPIN2 (кандидат за Stg2) в умовах дефіциту води. У умовах дефіциту води цей ген активується в більшості тканин Tx642 і Stg1 NIL в порівнянні з Tx7000. Докладний опис Протягом даного опису, якщо контекст не вказує на інше, слово "містять" або його варіанти, такі як "містить" або що "містить" потрібно розуміти як такі, що передбачають включення вказаного елемента або цілого числа або стадії способу або групи елементів або цілих чисел або стадій способів, але не виключення якого-небудь іншого елемента або цілого числа або стадії способу або групи елементів або цілих чисел або стадій способу. Як використовують в даній заявці, форма однини включає множинні аспекти, якщо контекст явно не вказує на інше. Таким чином, наприклад, вказівку на "локус" включать один локус, а також два або більше локусів; вказівка на "ауксин" включає один ауксин, а також два або більше ауксинів; вказівка на "опис" включає один або декілька аспектів, представлених в описі. Аспекти, описані в даному описі, охоплюються терміном "винахід". Всі аспекти, представлені, описані або заявлені в даному описі, входять в об'єм винаходу. У даному описі описані QTL, асоційовані і сприяючі фенотипу підтримування зеленим в культурі, включаючи злакові рослини. QTL позначають загалом як StgX, де X являє собою число від 1 і вище, відповідне генетичному локусу або області генетичних локусів на конкретній хромосомі в сільськогосподарській рослині. Підобласть позначають як StgXm, де m являє собою алфавітне позначення області в StgX. У даному описі описано, що модулювання експресії StgX у всіх або окремих тканинах рослини забезпечує фізіологічну і генетичну мережу, яка індукує або забезпечує переключення використання води сільськогосподарською рослиною на 9 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури або збільшення ефективності транспірації, тим самим, збільшуючи індекс урожаю (HI) і вихід зерна в умовах обмеженого доступу води. "Експресія" StgX включає активацію або пригнічення рівнів експресії (тобто модулювання експресії) локусу, а також вибір поліморфного варіанта, який експресується на вищому або нижчому рівні або який кодує більш або менш активний продукт у всіх або окремих тканинах в рослині. Фенотип може забезпечувати сам локус, або його функціональний еквівалент, такий як кДНК, що кодує той же білок, який кодується локусом. QTL ідентифікують локуси, що кодують білки PIN. Білки PIN являють собою ефлюксні переносники ауксину, які містять трансмембранні домени і, головним чином, локалізовані в плазматичних мембранах. Термін "PIN" походить від "суцвіття в формі шпильки". PIN для сорго являє собою "SbPIN". У даному описі описаний PIN із будь-якої рослини (наприклад, OsPIN із рису). Геномне положення SbPIN із сорго описано в таблиці 1A. У даному описі передбачений спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує білок PIN або його функціональну гомолог або ортолог; або який модулює експресію власного білка PIN; де рівень і місце експресії PIN забезпечує фенотип підтримування зеленим, який включає, серед інших, архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури, що приводить до збільшеного індексу урожаю і виходу зерен в умовах обмеження доступу води. Крім того, в даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує білок SbPIN сорго, вибраний із SbPIN1-SbPIN11, або еквівалент із іншої рослини, або який модулює експресію власного PIN. Приклади PIN включають SbPIN4 і SbPIN2 і інші SbPIN, наведені в таблиці 1A і їх еквіваленти в іншій рослині, а також PIN, що має особливо бажану поліморфну зміну, яка, наприклад, забезпечує змінений профіль експресії у вигляді підвищених рівнів білка PIN. Прикладами PIN в інших рослинах є OsPIN5, який відповідає SbPIN4, і OsPIN3a, який відповідає SbPIN2. Генетичний агент може являти собою локус або геномну область або їх функціональний еквівалент, такий як кДНК або фрагмент геномної ДНК. Альтернативно агент може модулювати експресію власного локусу PIN в конкретній рослині. Під "введенням" мають на увазі введення шляхом рекомбінантного втручання, шляхом мутагенезу або схрещування з подальшою селекцією. Без обмеження технології, описаної в даному описі, якою-небудь конкретною теорією або способом дії, модулювання експресії PIN окремо або в комбінації з генетичною або фізіологічною мережею змінює архітектуру рослини, посилюючи або іншим чином забезпечуючи ефективне використання води. У одному аспекті модифікована архітектура являє собою модифіковану архітектуру листяного покриву рослини. Термін "потомство" включає пряме потомство, а також віддалених родичів рослини, за умови, що вони стабільно експресують ознаку підтримування зеленим, внесений раніше в батьківську рослину. Вказівка на "сільськогосподарську рослину" включає злакову рослину. Сільськогосподарська культура, передбачена в рамках винаходу, включає сорго, пшеницю, жито, кукурудзу, ячмінь, рис, абаку, люцерну, мигдаль, яблуко, спаржу, банан, квасолю, чорну смородину, кормові боби, канолу, кеш'ю, маніоку, горох баранячий, цитрус, кокос, каву, кукурудзу, бавовну, інжир, льон, виноград, земляний горіх, коноплі, кенаф, лаванду, манго, гриби, оливки, цибулю, горох, арахіс, грушу, просо, картоплю, рами, рапс, плевел, сою, полуницю, цукровий буряк, цукрову тростину, соняшник, солодку картоплю, колоказію, чай, тютюн, томат, тритикале, трюфель і ямс. У одному прикладі, механізми сорго стійкості до засухи використовують для забезпечення стійкість до засухи в сорго, а також в інших сільськогосподарських рослинах. У одному прикладі генетично модифікована рослина використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду. Локуси PIN, що є в кожній з вказаних вище рослин називають "власними" PIN. У даному описі описана активація і пригнічення власного PIN або селекція PIN, що має конкретний профіль експресії. "Власний" PIN являє собою локус PIN в батьківській рослині до маніпулювання (рекомбінантні способи, мутагенез або схрещування). Термін "стійкість до засухи" включає витримування засухи, адаптацію до засухи, стійкість при засусі, знижену чутливість до умов засухи, збільшену ефективність використання води, а 10 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 також здатність змінювати використання води на використання води в період після цвітіння або збільшену засвоюваність води в ході росту культури, тим самим збільшуючи HI, і виходу зерна в умовах обмеженого доступу води. Рослини, що виявляють стійкість до засухи, описують як "адаптовані до засухи рослини" або "рослини, що виявляють знижену чутливість до умов обмеженого доступу води". У даному описі описано, що стійкість до засухи індукується, забезпечується або іншим способом асоційована з фенотипом підтримування зеленим. Термін "генетично модифікований", відносно рослини, включає спочатку отриману генетично модифіковану рослину, а також нащадків, прямих або віддалених, які стабільно експресують ознаку підтримування зеленим. Таким чином, в даному описі описані як класичні способи схрещування для введення генетичного агента, тобто QTL підтримування зеленим або його функціонального еквівалента, такого як кДНК або геномний фрагмент, або агента, який активує або пригнічує (тобто модулює) експресію QTL, або білка, який кодується ними, так і технологія генетичної інженерії. Остання охоплюється термінами "способи генетичної інженерії" і "рекомбінантні способи". Маркери, що визначають підтримку зеленим, також можна піддавати скринінгу в протоколах схрещування для моніторингу перенесення конкретних генетичних областей. Більш того, конкретну область підтримування зеленим генетично вбудовують рекомбінантними способами в клітину рослини або в калюс рослини і паросток регенерують. "Генетично модифікована" рослина включає батьківську рослину або будь-яких нащадків, а також будь-які продукти рослини, такі як зерно, насіння, матеріал розмноження, пилок і зародкові клітини. Крім того, локус PIN може експресуватися в одній конкретній тканині рослини, але не експресується або його експресія знижена в іншій тканині. Більш того, рослину можна піддавати мутагенезу, такому як генетичний, радіоактивний або хімічний мутагенез, і можна відбирати мутантні рослини з PIN, що мають бажаний фенотип. Вказівка на "фенотип підтримування зеленим" включає характеристики, вибрані із збільшеної пластичності архітектури листяного покриву, зменшеного розміру листяного покриву, збільшеної біомаси на одиничну площу листя під час цвітіння, вищої ефективності транспірації, збільшеного використання води в ході наливання зерна, збільшеного водного режиму рослини в ході наливання зерна, зниженого співвідношення біомаси до:після цвітіння, уповільненого старіння, збільшення виходу зерна, вищого розміру зерен і зниженого вилягання. У даному описі передбачений спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує продукт, асоційований або сприяючий фенотипу підтримування зеленим, який включає переключення використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури, або збільшення ефективності транспірації, що приводить до збільшення індексу урожаю і виходу зерна в умовах обмеженого доступу води, і де продукт вибраний з переліку, що складається з SbPIN1-11, включаючи SbPIN4 і SbPIN2, і інші SbPIN, наведені в таблиці 1A, або їх еквівалента з іншої рослини, або який модулює експресію власного PIN в рослині. Таким чином, в даному описі описане використання генетичного матеріалу, що кодує PIN, або генетичного матеріалу, який модулює рівні власного PIN для сприяння фенотипу підтримування зеленим. У даному описі передбачені генетично модифіковані рослини, які проявляють фенотип підтримування зеленим внаслідок генетичної модифікації, а також насіння, плоди, квітки і інший репродуктивний матеріал або інший матеріал розмноження. Також передбачені вихідний матеріал коріння або вихідний матеріал розмноження. Це основано на припущенні, що насіння, плоди, квітки, репродуктивний матеріал або матеріал розмноження проявляють або можуть переходити на фенотип підтримування зеленим, введений в кінцевого батька(ів). Вказівка на "агент, який модулює рівні експресії PIN," включає промотори, мікроРНК, гени і хімічні сполуки, які сприяють збільшеній або зниженій експресії гена у всіх або окремих тканинах або збільшеній або зниженій активності продукту гена, а також кДНК і геномні фрагменти. Агент також може являти собою інтрон геномного гена, який є частиною генетичної мережі, для сприяння модулюванню експресії. Білок PIN забезпечує градієнт ауксину в клітинах і містить трансмембранний домен і, головним чином, локалізований на плазматичній мембрані. Білки PIN є обмежуючими швидкість факторами транспорту ауксину і забезпечують векторний напрямок для потоків ауксину. У даному описі описано, що щонайменше один із Stg1 або Stg2 кодує білок PIN. Вважають, що введення Stg1 або Stg2 de novo в рослину або підвищення модулювання або експресії власних Stg1 або Stg2 сприяють вияву одного або декількох ознак або підознак, асоційованих з фенотипом підтримування зеленим. 11 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 Як указано вище, білки PIN являють собою ефлюксні переносники ауксину, які опосередковують полярний транспорт ауксину (PAT) з клітини в клітину в протилежність транспорту ауксину через ксилему (Rashotte et al., Plant Cell 13:1683-1697, 2000; Friml et al., Current Opinion in Plant Biology 6:7-12, 2003). Термін "PIN" походить від суцвіття в формі шпильки, яке розвивається в Arabidopsis, коли транспорт ауксину є дефектним. У даному описі передбачений SbPINn із сорго, де n являє собою число від 1 до 11, а також PIN із будь-якої іншої рослини. Також в даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує білок, вибраний із переліку, що складається з SbPIN1-11, такого як SbPIN4 і SbPIN2 або інші SbPIN, наведені в таблиці 1A, або їх еквівалента з іншої рослини, або яке модулює експресію власного PIN в рослині. У даному описі описано, що SbPIN4 і SbPIN2 сорго є основними генами адаптації до засухи, спільно з іншими SbPIN, а також їх еквівалентами з інших рослин. Відмінності в передачі сигналу ауксину пояснюють все з множини фенотипів, що спостерігаються в рослинах з профілем експресії PIN. Фенотипи, які проявляють рослини SbPIN4 або SbPIN2, наприклад, пояснюються змінами ефлюксу ауксину і включають зниження кущіння, листків менших розмірів (як в довжину, так і в ширину), зниження маси листя і збільшення співвідношення корінь:пагін. Фенотипи, які проявляють рослини SbPIN4 або SbPIN2, наприклад, також можуть бути пояснені непрямо (або як наслідок цих прямих ефектів) і включають збільшену доступність води під час цвітіння, вищу концентрацію N в листі під час цвітіння, збільшену транспірацію і біомасу на одиницю площі листя, вищу ефективність транспірації, збереження площі зеленого листя в ході наливання зерна, збільшений індекс урожаю, вищий вихід зерна, вищий розмір зерен і збільшену стійкість до вилягання. У даному описі передбачено, що еквіваленти SbPIN4 і SbPIN2 і інші SbPIN функціонують серед інших основних видів злаків і сільськогосподарських культур для підвищення адаптації до засухи по всьому світу, де доступність води обмежує ріст культури після цвітіння. Відповідно до вказівок в даному описі, модулювання експресії PIN, вибраного з SbPIN1-11, таких як SbPIN4 (Stg1) і SbPIN2 (Stg2), або їх еквівалента з іншої рослини у всіх або окремих тканинах забезпечує адаптацію до засухи прямо і непрямо, що в кінцевому результаті приводить до вищого виходу зерна, більшому розміру зерен, і стійкості до вилягання в умовах обмеженого доступу води. У одному прикладі фенотип підтримування зеленим залучає присутність декількох білків, таких як два або більше за SbPIN1-11, таких як SbPIN4 і SbPIN2. У даному описі описаний спосіб отримання генетично модифікованої рослини, яка використовує воду більш ефективно, ніж не модифікована генетично рослина того ж виду, причому спосіб включає введення в рослину або батьківську рослину генетичного агента, який кодує два або більше PIN або їх функціональну гомолог або ортолог, які асоційовані або сприяють фенотипу підтримування зеленим, який включає зміну використання води на використання води в період після цвітіння або збільшення засвоюваності води в процесі росту культури, або збільшення ефективності транспірації, що приводить до збільшення індексу врожаю і виходу зерна в умовах обмеження доступу води; або агента, який модулює рівні експресію двох або більше PIN. Під "двома або більше" мають на увазі 2 або 3, або 4, або 5, або 6, або 7, або 8, або 9, або 10, або 11. Одиничний або множинні локуси PIN також можуть експресуватися або бути інгібованими. У одному прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN4; і (ii) агента, який модулює рівні експресії SbPIN4 або його еквівалента в іншій рослині. У іншому прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN2; і (ii) агента, який модулює експресію SbPIN2 або його еквівалента в іншій рослині. Збільшена доступність води під час цвітіння в рослинах, що містять область Stg1, забезпечується зниженням використання води за допомогою двох механізмів (зниження кущіння і менший розмір листя). Виявилося, що обидва механізми окремо знижують розмір листяного покриву в середньому приблизно на 9%. Механізм "низького кущіння" переважає в умовах низької щільності, коли потенціал до кущіння є високим. Механізм "невеликого розміру листя" переважає в оточуючих умовах високої щільності, коли потенціал до кущіння є низьким. У сукупності, ці механізми забезпечують сільськогосподарські рослини із значною пластичністю відносно модифікації архітектури листяного покриву у відповідь на складність обмеження води. 12 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 Підтримування зеленим збільшує пластичність архітектури листяного покриву за допомогою конститутивних і адаптивних відповідей. Розмір листяного покриву у випадку Stg1 або Stg2 знижується приблизно на 5%, навіть коли доступ води не обмежений (конститутивна відповідь). Розмір листяного покриву далі знижується (адаптивна відповідь) при м'якій засусі (~10%) і більш важкій засусі (~15%). Низьке кущіння, головним чином, є конститутивною відповіддю. Невеликий розмір листя є як конститутивною, так і адаптивною відповіддю. Більше того, в даному описі описано, що область Stg1 або Stg2 забезпечує адаптацію до засухи шляхом зниження розміру листяного покриву (внаслідок зниження кущіння і меншого розміру листя) і зниження використання води культурою під час цвітіння. Це показано за допомогою високої кореляції (r2=0,9) між розміром листяного покриву і використанням води культурою в дослідженнях із штучною засухою (укриття від дощу [ROS]) і в лізиметричних дослідженнях. Збільшеної доступності води під час цвітіння також досягають за допомогою збільшення засвоюваності води (краще витягання води і більш глибоке і більш латеральне поширення). Підтримування зеленим збільшує біомасу на одиницю площі листя під час цвітіння. Передбачаючи, що маса коріння еквівалентна (або щонайменше трохи менше), ці відмінності могли б бути пояснені відмінностями в транспірації (Т) на одиничну площу листя [LA] (Т/LA) і/або ефективність транспірації (TE). Лізиметричні дослідження вказують на те, що збільшення Т/LA, а не TE, ініціюють спостережувані збільшення біомаси на площу листка. Потрібно зазначити, що збільшення Т/LA відбувалося, тільки коли дефіцит води був достатнім, щоб зменшити площу листя. Коли дефіцит води був менш важким (тобто недостатнім, щоб зменшити площу листя), тоді відбувалося зниження Т/LA, що приводило до вищої TE. Вищу TE в лініях StgX, таких як Stg1 або Stg2, також спостерігають, коли дефіцит води є менш складним. Вважають, що збільшення TE шляхом інтрогресії Stg1 або Stg2 є наслідком a) пропорціонально вищої здатності до фотосинтезу в порівнянні з продиховою провідністю, внаслідок меншого розміру, меншої товщини листя і більш зеленого листя, і/або b) зниження транспірації при збереженні біомаси. Лізиметричні дослідження вказують на те, що обидва з цих механізмів приводять до вищої TE в лініях Stg1, причому основним механізмом є зниження транспірації. У даному описі вважають, що зміна транспірації на одиничну площу листя є наслідком a) кількості продихів, b) розміру отвору продихів, с) зміни часу відкриття і закриття продихів відносно VPD, і/або d) кількості клітин основи волосків (яка впливає на пограничний шар і, таким чином, Т/LA). Інтрогресія Stg1, наприклад, в RTx7000 модифікувала анатомію листя шляхом збільшення кількості клітин обкладки судинних пучків, оточуючих судинний пучок. Відмінності в морфології листя є очевидними між RTx7000 і Stg1 або Stg2. У цьому випадку, в Stg1 або Stg2 обкладка судинних пучків, оточуюча судинний пучок, була більш представлена і була меншого розміру. Також було менше продихів і більше клітин основи волоска на одиничну площу листя (листя 7 і 10) в Stg1 в порівнянні з RTx7000. Збільшеного використання води в ході наливання зерна досягають шляхом (i) збільшеної доступності води під час цвітіння і (ii) збільшеної засвоюваності води (краще витягання води і більш глибоке або більш латеральне поширення) в ході наливання зерна. Використання води культурою (CWU) до цвітіння негативно корелювало з CWU після цвітіння в експерименті з штучною засухою (укриття від дощу [ROS]). Наприклад, в одному експерименті 25% збільшення використання води після цвітіння (80 проти 60 мм) приводило до 2 25% збільшення виходу зерна (400 проти 300 г/м ). Це відповідало 50 кг/га зерна на кожний додатковий мм доступної води. Stg1 або Stg2 проявляли збільшене використання води в період наливання зерна в умовах як низької, так і високої щільності в експерименті з укриттям від дощу (ROS). Це було наслідком, головним чином, (i) зниженого використання води під час цвітіння в умовах високої щільності, і (ii) збільшеної засвоюваності води в ході наливання зерна в умовах низької щільності. У даному описі описано, що StgX, такий як Stg1 або Stg2, забезпечує адаптацію до засухи тим, що він асоційований з продукцією біомаси до і після цвітіння. Область Stg1 або Stg2, наприклад, знижує співвідношення біомаси до:після цвітіння нижче критичного рівня, збільшуючи вихід зерна і стійкість до вилягання. Відповідно до вказівок даного опису, експресія StgX, такого як Stg1, Stg2, Stg3 і/або Stg4, сприяє одному або декільком їх наступних фенотипів: (i) уповільнене старіння листя (підтримування зеленим), вищий вихід зерна і стійкість до вилягання є наслідком вищого водного режиму рослин в ході наливання зерна (внаслідок збільшеного використання води в ході наливання зерна); 13 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 (ii) інтрогресія StgX, наприклад, в фон RTx7000 збільшує водний режим рослини при середньому наливанні зерна, як показано по a) вищому відносному вмісту води (RWC), і b) нижчому водному потенціалу листя (LWP); (iii) вищий вихід зерна і більший розмір зерен є наслідком збільшеної доступності води в ході наливання зерна; (iv) вищий вихід зерна, більший розмір зерен, і збільшена стійкість до вилягання не є взаємовиключними (тобто StgX виявляє всі три ознаки); (v) переваги виходу і розміру зерен є відносно вищими при важкій термінальній засусі, чому при м'якій термінальній засусі; (vi) користь генів підтримування зеленим, наприклад, в фоні RTx7000 (інбредні рослини) виникає при діапазоні виходу 1-3 т/га (12-22%), а потім з меншою, але ще істотною користю при діапазоні виходу 3-4 т/га (8-10%). Однак було невелике погіршення, асоційоване з цими областями (2-4%) при вищих виходах (5-8 т/га) внаслідок більш вологих умов. Потрібно зазначити, що ці діапазони виходів були б значно вищими в гібридах. Оскільки середній вихід зерна в сорго для гібридів в північній зерновій зоні становить приблизно 2,5 т/га, користь генів підтримування зеленим повинна бути значною. У гібридах не спостерігали зниження виходу зерна в більш вологих умовах (без обмеження води) завдяки підтримуванню зеленим; (vii) інтрогресія StgX, наприклад, в RTx7000 також збільшує розмір зерен на 11%, в середньому, при важкій термінальній засусі. Не було впливу StgX QTL на розмір зерен при м'якій термінальній засусі або без засухи; і (viii) кожний з ключових механізмів StgX картується в певній області, вказуючи на те, що дія одного гена має множину плейотропних ефектів. Крім того, в даному описі описана комерційна модель для підвищення економічного прибутку від продукції культури. Відповідно до цих ідей, передбачається комерційна модель для підвищення економічного прибутку від виходу культури, причому модель включає отримання сільськогосподарських рослин, які мають профіль експресії PIN, що приводить до сільськогосподарської рослини, що має переключення використання води рослиною на використання води в період після цвітіння або збільшенням засвоюваності води в ході росту культури, тим самим збільшуючи HI і вихід зерна в умовах обмеженого доступу води, отримання насіння від отриманої сільськогосподарської рослини і розподіл насіння виробникам зерна для досягнення збільшеного виходу і прибутку. Вказівка на PIN включає SbPIN1-11 такі як SbPIN4 і SbPIN2, а також їх еквіваленти з інших рослин. У даному описі описана система керування рослинництвом для зниження залежності культури від води або іншого підвищення ефективності використання води і збільшення виходу зерна або продукту. Система керування рослинництвом включає отримання адаптованої до засухи культури, яка включає злакові рослини, з використанням вибору і модульованої експресії локусу PIN або його функціонального еквівалента, як визначено в даному описі, окремо або в комбінації з введенням інших корисних ознак, таких як розмір зерен, розмір коренів, стійкість до солі, стійкість до гербіцидів, стійкість до шкідників і т. п. Альтернативно або додатково, система керування рослинництвом включає отримання адаптованих до засухи рослин і сільськогосподарським процедурам, таким як зрошування, вимоги до поживних речовин, щільності і геометрія насадження культури, боротьба з бур'янами, боротьба з комахами, аерація ґрунту, зменшення кущіння, підняття грядок і т. п. У даному описі описано, як індукувати або посилювати здатність до адаптації до засухи в рослині шляхом внесення de novo одного або декількох ознак фенотипу підтримування зеленим або збільшення, або зниження експресії існуючого одного або декількох локусів PIN в рослині і/або селекції поліморфного варіанта PIN із збільшеною або посиленою експресією або активністю продукту. Маніпулювання фенотипом підтримування зеленим можна провести окремо або як частина інтегрованої системи керування рослинництвом, яка може включати подальшу селекцію ознаки і/або удосконалення агрономічних технологій. Отримані культури використовують воду більш ефективно і мають вищий вихід зерна і збільшений розмір зерен. Ідеї даного винаходу включають комерційні моделі для збору насіння від адаптованих до засухи або посилених сільськогосподарських рослин для розподілу фахівцям з вирощування для збільшення в кінцевому результаті виходу зерна. У даному описі передбачається використання генетичного агента, кодую-чого білок PIN або модулюючого рівні експресії власного білка PIN, для виробництва адаптованої до засухи рослини. QTL ідентифіковані в рамках даного винаходу як несучі один або декілька локусів PIN, де селекцію рівня їх експресії проводять за допомогою протоколів схрещування або шляхом генетичної інженерії, для забезпечення фенотипу підтримування зеленим. 14 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 60 Також в даному описі описані генетично модифіковані рослини і їх нащадки, які виявляють ознаку підтримування зеленим, а також насіння, плоди, і квітки і інший репродуктивний матеріал або матеріал розмноження. У рамках даного винаходу передбачені як генетичний матеріал, який кодує білок PIN або його функціональну гомолог або ортолог, який асоційований або сприяє фенотипу підтримування зеленим, який включає архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури або збільшенню ефективності транспірації, які приводять до збільшення індексу урожаю і виходу зерна в умовах обмеження доступу води, так і агент, який модулює рівень PIN. У одному прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN, наведений в таблиці 1A; і (ii) агента, який модулює рівні експресії SbPIN, наведеного в таблиці 1A, або його еквівалента з іншої рослини. У одному прикладі генетичний матеріал вибраний із (i) агента, який кодує SbPIN4; і (ii) агента, який модулює рівні експресії SbPIN4 або його еквівалента з іншої рослини. У іншому прикладі генетичний матеріал вибраний з (i) агента, який кодує SbPIN2; і (ii) агента, який модулює експресію SbPIN2 або його еквівалента з іншої рослини. Генетичний матеріал включає агент, який кодує PIN, або його функціональний гомолог або ортолог, який асоційований або сприяє фенотипу підтримування зеленим, який включає архітектуру листяного покриву, який сприяє переключенню використання води на використання води в період після цвітіння або збільшенню засвоюваності води в процесі росту культури або збільшенню ефективності транспірації, які приводять до збільшення індексу урожаю і виходу зерна в умовах обмеження доступу води. Активний агент включає агент, який підвищує або знижує рівні PIN. Гени, які розглядаються також можна використовувати як маркери для перенесення областей геномів, які включають один або декілька з генів або їх еквівалентів з інших рослин, в конкретну рослину для включення в нього фенотипу підтримування зеленим. Приклади Аспекти, описані і передбачені в даному описі, далі описані за допомогою наступних необмежуючих прикладів. Приклад 1 Ідентифікація гена StgX Був ідентифікований локус кількісних ознак (QTL), який позначають в даному описі як Stg1, що є прикладом StgX, який підвищує або посилює ефективність використання води рослинами сорго. Stg1 кодує представник сімейства компонента 4 переносників вихідного ауксину, PIN4 (або SbPIN4) із сорго зернового. Цей основний ген адаптації до засухи був точно картований в геномі сорго. Зміни ефлюксу ауксину можуть пояснити всі з множини фенотипів, спостережуваних в рослинах, які містять SbPIN4. Ген-кандидат (і промоторну область) секвенують в двох батьківських рослинах популяції з точним картуванням (RTx7000 і Tx642) для ідентифікації однонуклеотидного поліморфізму. Також проводять визначення профілю експресії РНК в популяції з точно картованим Stg1 для підгруп ліній, моментів часу і органів. Фенотипи, що проявляються рослинами SbPIN4, які можуть бути пояснені безпосередньо змінами ефлюксу ауксину, включають знижене кущіння, менший розмір листя (як в довжину, так і в ширину), знижену масу листя і збільшене співвідношення коріння:пагони. Фенотипи, що проявляються рослинами SbPIN4, які можуть бути пояснені непрямо (або як наслідки цих прямих ефектів), включають збільшену доступність води під час цвітіння, вищу концентрацію N під час цвітіння, збільшену транспірацію і біомасу на одиницю площі листка, знижене співвідношення біомаси до:після цвітіння, вищу ефективність транспірації, збереження площі зеленого листя в ході наливання зерна, збільшений індекс урожаю, вищий вихід зерна, більший розмір зерен і збільшену стійкість до вилягання. Передбачається, що SbPIN4 функціонує серед інших основних злакових і сільськогосподарських видів, підвищуючи адаптацію до засухи по всьому світу, де доступ води обмежує ріст культури після цвітіння. Stg1 (SbPIN4) забезпечує адаптацію до засухи як прямо, так і непрямо, що в кінцевому результаті приводить до вищого виходу зерна, більшого розміру зерен і стійкості до вилягання в умовах обмеженого доступу води. Збільшена доступність води під час цвітіння в рослинах, які містять область Stg1, забезпечується зниженням використання води за допомогою двох механізмів (зниження кущіння і менший розмір листя). Виявилося, що обидва механізми окремо знижують розмір листяного покриву в середньому приблизно на 9%. Механізм "низького кущіння" переважає в умовах низької щільності, коли потенціал до кущіння є високим. Механізм "невеликого розміру листя" 15 UA 113503 C2 5 10 15 20 25 30 35 40 45 50 55 переважає в оточуючих умовах високої щільності, коли потенціал до кущіння є низьким. У сукупності, ці механізми забезпечують сільськогосподарські рослини із значною пластичністю відносно модифікації архітектури листяного покриву у відповідь на складність обмеження води. Підтримування зеленим збільшує пластичність архітектури листяного покриву за допомогою конститутивних і адаптивних відповідей. Розмір листяного покриву у випадку Stg1 або Stg2 знижується приблизно на 5%, навіть коли доступ води не обмежений (конститутивна відповідь). Розмір листяного покриву далі знижується (адаптивна відповідь) при м'якій засусі (~10%) і більш важкій засусі (~15%). Низьке кущіння, головним чином, є конститутивною відповіддю. Невеликий розмір листя є як конститутивною, так і адаптивною відповіддю. Існує зв'язок між зменшеним розміром листяного покриву (за допомогою зниженого кущіння і меншого розміру листя) і зниженим використанням води культури під час цвітіння. Існує висока кореляція (r2=0,9) між розміром листяного покриву і використанням води культурою в дослідженнях з ROS і лізиметричних дослідженнях. Збільшеної доступності води під час цвітіння також досягають за допомогою збільшення засвоюваності води (краще витягання води і більш глибоке і більш латеральне поширення). Підтримування зеленим збільшує біомасу на одиницю площі листя під час цвітіння. Передбачаючи, що маса коренів еквівалентна (або щонайменше трохи менша), ці відмінності могли б бути пояснені відмінностями в транспірації на одиничну площу листя (Т/LA) і/або ефективність транспірації (TE). Лізиметричні дослідження вказують на те, що збільшення Т/LA, а не TE, ініціює спостережувані збільшення біомаси на площу листка. Потрібно зазначити, що збільшення Т/LA відбувалося тільки в умовах низького VPD; Т/LA була насправді знижена в умовах високого VPD, приблизно як механізм утримання води. Вищу TE в лініях Stg1 також спостерігали в умовах вищого VPD. Збільшення TE шляхом інтрогресії Stg1 може бути наслідком a) пропорційно вищої здатності до фотосинтезу в порівнянні з продиховою провідністю, внаслідок меншого розміру, меншої товщини листя і більш зеленого листя, і/або b) зниження транспірації при збереженні біомаси. Лізиметричні дослідження вказують на те, що обидва з цих механізмів приводять до вищої TE в лініях Stg1, причому основним механізмом є зниження транспірації. Зміна транспірації на одиничну площу листка може бути наслідком a) кількості продихів, b) розміру отвору продихів, с) зміни часу відкриття і закриття продихів відносно VPD, і/або d) кількості клітин основи волосків (яке впливає на пограничний шар і, таким чином, Т/LA). Інтрогресія Stg1 в RTx7000 знижувала кількість продихів і збільшувала кількість клітин основи волосків на одиницю площі листка в листках 7 і 10; причому обидва механізми можуть зберігати воду шляхом зниження Т/LA. Інтрогресія Stg1, наприклад, в RTx7000 модифікувала анатомію листя шляхом збільшення кількості клітин обкладки судинних пучків, оточуючих судинний пучок. Збільшена кількість клітин в обкладці судинних пучків також може приводити до збільшеної фотосинетичної асиміляції і, таким чином, TE. Відмінності в морфології листя (наприклад, листя 7 і 10) є очевидними між RTx7000 і Stg1. У цьому випадку, в Stg1 обкладка судинних пучків, оточуюча судинний пучок, була більш представлена і була меншого розміру. Збільшена кількість клітин в обкладці судинних пучків також може приводити до збільшеної фотосинетичної асиміляції і, таким чином, TE. Збільшеного використання води в ході наливання зерна досягають шляхом (i) збільшеної доступності води під час цвітіння і (ii) збільшеної засвоюваності води (краще витягання води і більш глибоке або більше латеральне поширення) в ході наливання зерна. a) Збільшена доступність води під час цвітіння Використання води культурою (CWU) до цвітіння негативно корелювало з CWU після цвітіння в експерименті з ROS. Наприклад в одному експерименті 25% збільшення використання води після цвітіння (80 проти 60 мм) приводило до 25% збільшення виходу зерна 2 (400 проти 300 г/м ). Це відповідало 50 кг/га зерна на кожний додатковий мм доступної води. b) Збільшена засвоюваність води в процесі наливання зерна Рослини з Stg1 проявляли збільшене використання води в ході періоду наливання зерна в умовах як низької, так і високої щільності в експерименті з ROS. Це було наслідком, головним чином, (i) зниженого використання води під час цвітіння при високій щільності, і (ii) збільшеній засвоюваності води в ході наливання зерна при низькій щільності. Область Stg1 забезпечує адаптацію до засухи через зв'язок між продукцією біомаси до і після цвітіння. Область Stg1 знижує співвідношення біомаси до:після цвітіння нижче критичного рівня, збільшуючи вихід зерна і стійкість до вилягання. 16 UA 113503 C2 5 10 15 20 25 30 35 40 45 Наслідком вищого водного режиму рослини в ході наливання зерна (внаслідок збільшеного використання води в ході наливання зерна) є уповільнене старіння листя (підтримування зеленим), вищий вихід зерна і стійкість до вилягання. Інтрогресія Stg1 в фон RTx7000 збільшувала водний режим рослини при середньому наливанні зерна, як показано по a) вищому відносному вмісту води (RWC), і b) нижчому водному потенціалу листя (LWP). Вищий вихід зерна і більший розмір зерен є наслідком збільшеної доступності води в ході наливання зерна. Вищий вихід зерна, більший розмір зерен і збільшена стійкість до вилягання не є взаємовиключними (тобто Stg1 проявляє всі три ознаки). Переваги виходу і розміру зерен є відносно вищими при важкій термінальній засусі, ніж при м'якій термінальній засусі. Дослідження вказують на те, що найбільша користь генів підтримування зеленим, наприклад, в фоні RTx7000 (інбредні рослини) виникає при діапазоні виходу 1-3 т/га (12-22%), а потім з меншою, але ще істотною користю при діапазоні виходу 3-4 т/га (8-10%). Однак було невелике погіршення, асоційоване з цими областями (2-4%), при вищих виходах (5-8 т/га) внаслідок більш вологих умов. Потрібно зазначити, що ці діапазони виходів були б значно вищими в гібридах. Оскільки середній вихід зерна в сорго для гібридів в північній зерновій зоні становить приблизно 2,5 т/га, користь генів підтримування зеленим повинна бути значною. У гібридах не спостерігали зниження виходу зерна в більш вологих умовах (без обмеження води) завдяки підтримуванню зеленим. Інтрогресія Stg1 в RTx7000 також збільшує розмір зерна, в середньому, на 11% при важкій термінальній засусі. Не було впливу цього QTL на розмір зерен при м'якій термінальній засусі або без засухи. Кожний з ключових механізмів Stg1 картується в одній і тій же області, вказуючи на те, що дія одного гена має множину плейотропних ефектів. Приклад 2 Зниження кущіння (фізіологічні дослідження NIL в полі) Дані показують вплив Stg1 на кущіння в умовах як високого доступу води (HW), так і низького доступу води (LW). Відмінності в розвитку листяного покриву до цвітіння здебільшого 2 були наслідком варіювання кущіння серед ліній. Кількість стебел на м під час цвітіння була найкращим загальним показником ефекту кущіння на динаміку листяного покриву. Кількість 2 стебел на м була еквівалентною при обох водних режимах (12,89), вказуючи на те, що зниження кущіння є конститутивною ознакою. Генотипи варіювали значно (Р
ДивитисяДодаткова інформація
Назва патенту англійськоюDrought tolerant plants
Автори англійськоюBorrell, Andrew Kenneth, Jordan, David Robert, Mullet, John, Klein, Patricia
Автори російськоюБоррелл Эндрю Кеннет, Джордан Дэвид Роберт, Маллит Джон, Клейн Патрисия
МПК / Мітки
МПК: C12N 15/05, C12N 15/29, A01H 4/00, C12N 15/82
Мітки: рослина, стійка, засухи
Код посилання
<a href="https://ua.patents.su/67-113503-stijjka-do-zasukhi-roslina.html" target="_blank" rel="follow" title="База патентів України">Стійка до засухи рослина</a>
Рослина цукрового буряку, стійка до гліфосату

Номер патенту: 85364
Опубліковано: 26.01.2009
Автори: Стен Пер, Маннерлоф Марія, Теннінг Пауль Петер
МПК: C12N 15/52, C12N 9/10, A01H 5/10, C12N 15/82, A01H 5/00
Мітки: цукрового, стійка, гліфосату, рослина, буряку
Формула / Реферат:
1. Рослина цукрового буряку, в тому числі її нащадки, яка експресує фермент з 5-енолпірувілшикімат-3-фосфатсинтазною активністю, одержуваною з Agrobacteria sp.СР-4, яка відрізняється тим, що ПЛР-амліфікація з використанням як матриці її геномної ДНК приводить до ампліфікації фрагменту ДНК, розміром 739 пар основ, при використанні пари нуклеотидних праймерів з послідовностями, представленими в SEQ ID NO: 18 і SEQ ID NO:...
Рослина капусти качанної (brassica oleracea), стійка до albugo candida

Номер патенту: 100121
Опубліковано: 26.11.2012
Автори: Постма-Хаарсма Адріана Дорін, Схрейвер Альбертус Йоханесс Марія, Гес де Ян, Хоогланд Йоханесс Герардус Марія
МПК: A01H 5/00, C12N 15/82, C12Q 1/68
Мітки: капусти, candida, brassica, oleracea, рослина, albugo, качанної, стійка
Формула / Реферат:
1. Рослина Brassica oleracea, що містить домінантний ген стійкості до Albugo Candida, який викликає білу іржу хрестоцвітих, де ген стійкості одержують з рослини B. oleracea, насіння якої депоновано 1 березня 2006 р. до Американської колекції типових культур (ATCC) під номером PTA-7412, де наявність інтрогресії гену стійкості може бути показана за допомогою принаймні двох специфічних маркерів ДНК вибраних з...
Рослина капусти brassica oleracea, стійка до захворювання килою

Номер патенту: 84548
Опубліковано: 10.11.2008
Автори: Тжертес Петер, де Хаас Йоханнес Марія, Ліндерс Енріо Герардус Альбертус, Хуанг Ціа-Ченг
МПК: A01H 1/00, A01H 5/00, A01H 5/10, C12N 15/09, C12Q 1/68
Мітки: oleracea, капусти, рослина, захворювання, brassica, килою, стійка
Формула / Реферат:
1. Рослина B. oleracea, стійка до захворювання килою, де ознаку стійкості одержують зі стійких до кили рослин B. rapa і де ознака стійкості до кили є моногенною та домінантною. 2. Рослина за п. 1, де ураження захворюванням цієї рослини B. oleracea відповідає балу 2 або нижче при оцінці ураження, яке викликається захворюванням, за 1-9-бальною шкалою, або балу 1 або нижче при оцінці ураження, що викликається...
Нетрансгенна стійка до посухи рослина

Номер патенту: 110774
Опубліковано: 25.02.2016
Автори: Врізен Віллем Хендрік, Нітс Лісетт
МПК: C12N 15/55, C12N 15/82, A01H 5/08, A01H 5/00, A01H 1/06
Мітки: нетрансгенна, стійка, посухи, рослина
Формула / Реферат:
1. Нетрансгенна рослина, що містить алель Solanum lycopersicum PP2C1 (SIPP2C1) у своєму геномі, де алель SIPP2C1 - це алель, що кодує білок, який виявляє, щонайменше 90 % амінокислотної послідовності, ідентичної до послідовності SEQ ID NO: 2, яка відрізняється...
Стійка до гербіцидів соняшникова рослина

Номер патенту: 108062
Опубліковано: 25.03.2015
Автори: Сала Карлос, Булос Маріано
МПК: A01H 5/10, A01H 5/00, C12N 9/10, C12N 15/82, C12N 15/55
Мітки: стійка, рослина, соняшникова, гербіцидів
Формула / Реферат:
1. Соняшникова рослина, яка має стійкість до імідазолінонового гербіциду, сульфонілсечовинного гербіциду, триазолопіримідинового гербіциду, піримідинілоксибензоатного гербіциду, що містить мутантний ген AHAS, який кодує мутацію амінокислоти в положенні 574 або еквівалентному положенні, де амінокислота в положенні 574 або еквівалентному положенні є амінокислотою, відмінною від триптофану, де амінокислотою в положенні 574 або еквівалентному...
Попередній патент: Спосіб підвищення родючості піщаних ґрунтів
Наступний патент: Система і спосіб обробки насіння для підприємств роздрібного продажу
Випадковий патент: Вічний двигун