Експресія генів у модифікованому вірусі коров’ячої віспи ankara з використанням коров’ячого поквірусного промотору аті







Формула / Реферат
1. Рекомбінантний модифікований вірус коров'ячої віспи Ankara (MVA), який містить у вірусному геномі експресійну касету, яка містить коров'ячий поксвірусний ATI промотор або його похідну та кодуючу послідовність, де експресія кодуючої послідовності регулюється вказаним промотором або його похідною, та де похідна ATI промотору являє собою:
(і) послідовність, що має гомологію принаймні 60 % порівняно з послідовністю SEQ ID No:1,
(іі) послідовність, у якій не більше 6 нуклеотидів замінені, видалені та/або вставлені в послідовність SEQ ID No:1, та/або
(ііі) підпослідовність послідовності відповідно до SEQ ID No:1, де згадана підпослідовність має довжину принаймні 10 нуклеотидів послідовності SEQ ID No:1, де згадані послідовність та підпослідовність (і), (іі) та/або (ііі) зберігають активність як промотор.
2. Рекомбінантний MVA за п. 1, де послідовність та підпослідовність відповідно до (і), (іі) та/або (ііі) зберігають активність як пізній промотор вірусу коров'ячої віспи.
3. Рекомбінантний MVA за будь-яким з пп. 1-2, де підпослідовність (ііі) має довжину принаймні 20 нуклеотидів послідовності SEQ ID No:1.
4. Рекомбінантний MVA за будь-яким з пп. 1-3, де підпослідовність (ііі) має довжину принаймні 25 нуклеотидів послідовності SEQ ID No:1.
5. Рекомбінантний MVA за п. 1, де послідовність (і) має гомологію принаймні 80 %, порівняно з послідовністю SEQ ID No:1.
6. Рекомбінантний MVA за будь-яким з пп. 1-5, де послідовність (і), (іі) та/або послідовність (ііі) включає нуклеотиди 25-29 та/або 22-29 SEQ ID No:1.
7. Рекомбінантний MVA за п. 1, де ATI промотор має послідовність SEQ ID No:1.
8. Рекомбінантний MVA за будь-яким з пп. 1-7, де MVA вибирають зі штаму MVA-BN депонованого в European Collection of Cell Cultures (ECACC) під номером V00083008 або його похідної та штаму MVA 575, депонованого під номером V00120707 в ЕСАСС.
9. Рекомбінантний MVA за будь-яким з пп. 1-8, де експресійна касета вставлена в сайт делеції геному MVA, що зустрічається в природі, та відповідно у вірусі коров'ячої віспи Copenhagen або у міжгенному регіоні геному MVA.
10. Рекомбінантний MVA за будь-яким з пп. 1-9, де кодуюча послідовність кодує принаймні один антиген або антигенний епітоп та/або терапевтичну сполуку.
11. Рекомбінантний MVA за будь-яким з пп. 1-10 як вакцина або медикамент.
12. Вакцина або фармацевтична композиція, що включає рекомбінантний MVA згідно з будь-яким з пп. 1-10.
13. Використання рекомбінантного MVA за будь-яким з пп. 1-10 для виготовлення вакцини або медикаменту, для викликання імунної відповіді проти агента, з якого походить антиген/епітоп, кодований кодуючою послідовністю.
14. Використання за п. 13, де антиген/епітоп являє собою частину поліпептиду, пептиду або протеїну, який походить з вірусу, бактерій або грибів.
15. Використання за п. 14, де вірус вибрано з ВІЛ, HTLV (вірус Т-клітинного лейкозу людини), герпесвірусу, вірусу тропічної лихоманки, поліовірусу, вірусу кору, вірусу свинки, вірусу краснухи та вірусу гепатиту.
16. Вакцина або фармацевтична композиція за п. 12 або використання за будь-яким з пп. 13-15, де вакцина, фармацевтична композиція або медикамент містять принаймні 102 ТСID50 (доза, що інфікує тканинну культуру - tissue culture infectious dose) вірусу.
17. Вакцина або фармацевтична композиція за будь-яким з пп. 12 або 16 або використання за будь-яким з пп. 13-15, де вакцину, фармацевтичну композицію або медикамент слід уводити в терапевтично ефективних кількостях при першій інокуляції ("первинній інокуляції") та при другій інокуляції ("підсилюючій інокуляції").
18. Спосіб уведення кодуючої послідовності в клітини-мішені, що включає інфікування клітин-мішеней вірусом, згідно з будь-яким з пп. 1-10.
19. Спосіб одержання пептиду, протеїну та/або вірусу, який включає:
a) інфікування клітини-хазяїна вірусом згідно з будь-яким з пп. 1-10,
b) культивування інфікованої клітини-хазяїна за придатних умов, та
c) виділення та/або збагачення пептиду та/або протеїну, та/або вірусів, вироблених вказаною клітиною-хазяїном.
20. Клітина, що містить вірус відповідно до будь-якого з пп. 1-10.
21. Використання коров'ячого поксвірусного ATI промотору або його похідної, визначеної в будь-якому з пп. 1-7, для експресії кодуючих послідовностей у MVA.
22. Спосіб продукування рекомбінантного MVA за будь-яким з пп. 1-10, що включає етап введення експресійної касети в геном MVA.
23. Спосіб індукування імунологічної відповіді в живому тваринному організмі, що включає введення вірусу згідно з будь-яким з пп. 1-10 або композиції згідно з п. 12 тварині або людині, що лікується.
24. Спосіб за п. 23, що включає введення принаймні 102 ТСID50 (доза, що інфікує тканинну культуру - tissue culture infectious dose) вірусу.
25. Спосіб за будь-яким з пп. 23-24 або використання за п. 13, де вірус, композиція або вакцина застосовується в терапевтично ефективних кількостях, при першій інокуляції ("первинній інокуляції") та при другій інокуляції ("підсилюючій інокуляції").
Текст
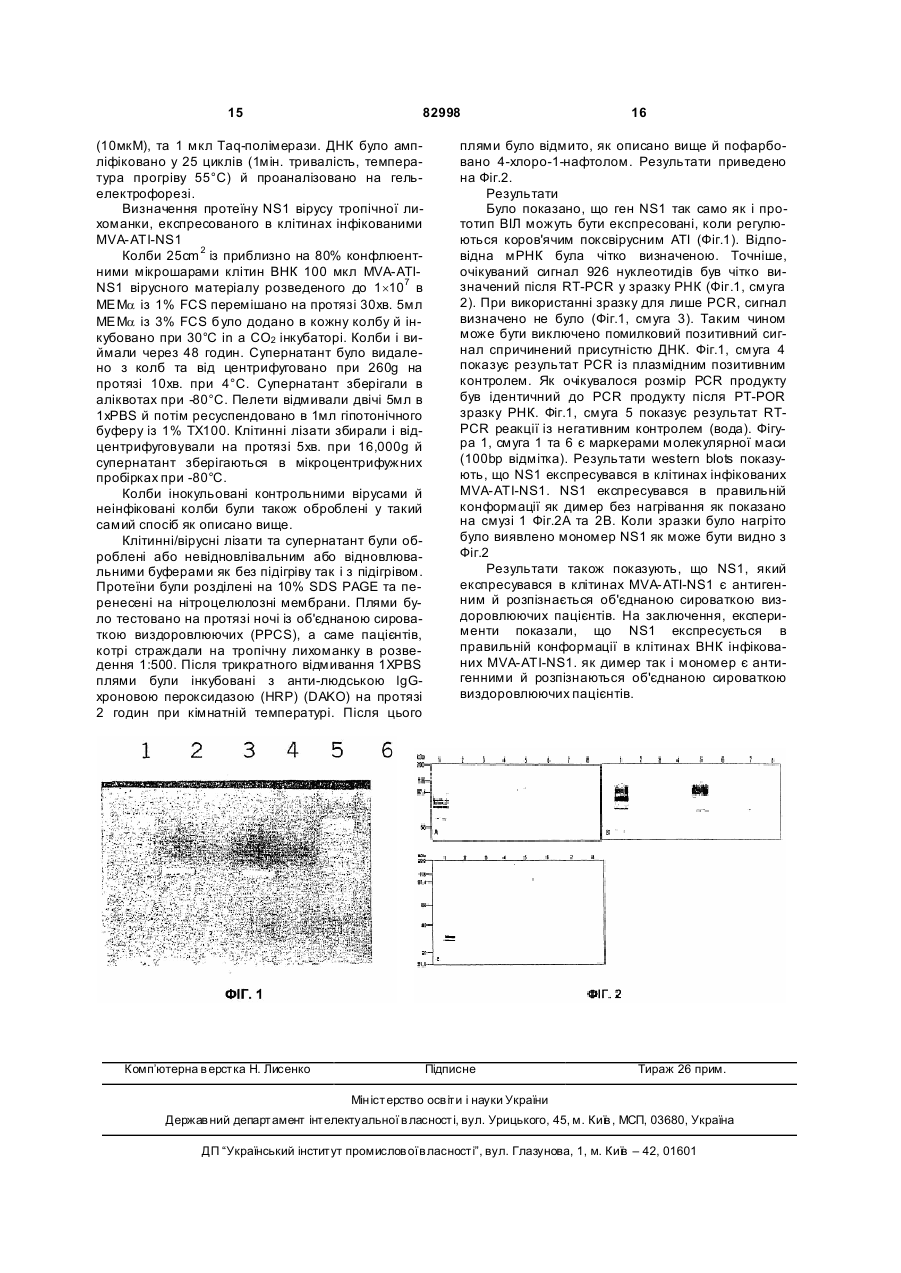
1. Рекомбінантний модифікований вірус коров'ячої віспи Ankara (MVA), який містить у вірусному геномі експресійну касету, яка містить коров'ячий поксвірусний ATI промотор або його похідну та кодуючу послідовність, де експресія кодуючої послідовності регулюється вказаним промотором або його похідною, та де похідна ATI промотору являє собою: (і) послідовність, що має гомологію принаймні 60 % порівняно з послідовністю SEQ ID NO:1, (іі) послідовність, у якій не більше 6 нуклеотидів замінені, видалені та/або вставлені в послідовність SEQ ID NO:1, та/або (ііі) підпослідовність послідовності відповідно до SEQ ID NO:1, де згадана підпослідовність має довжину принаймні 10 нуклеотидів послідовності SEQ ID NO:1, де згадані послідовність та підпослідовність (і), (іі) та/або (ііі) зберігають активність як промотор. 2. Рекомбінантний MVA за п.1, де послідовність та підпослідовність відповідно до (і), (іі) та/або (ііі) зберігають активність як пізній промотор вірусу коров'ячої віспи. 3. Рекомбінантний MVA за будь-яким з пп.1-2, де підпослідовність (ііі) має довжину принаймні 20 нуклеотидів послідовності SEQ ID NO:1. 4. Рекомбінантний MVA за будь-яким з пп.1-3, де підпослідовність (ііі) має довжину принаймні 25 нуклеотидів послідовності SEQ ID NO:1. 5. Рекомбінантний MVA за п.1, де послідовність (і) має гомологію принаймні 80 %, порівняно з послідовністю SEQ ID NO:1. 2 (19) 1 3 82998 4 композицію або медикамент слід уводити в терапевтично ефективних кількостях при першій інокуляції ("первинній інокуляції") та при другій інокуляції ("підсилюючій інокуляції"). 18. Спосіб уведення кодуючої послідовності в клітини-мішені, що включає інфікування клітинмішеней вірусом, згідно з будь-яким з пп.1-10. 19. Спосіб одержання пептиду, протеїну та/або вірусу, який включає: a) інфікування клітини-хазяїна вірусом згідно з будь-яким з пп.1-10, b) культивування інфікованої клітини-хазяїна за придатних умов, та c) виділення та/або збагачення пептиду та/або протеїну, та/або вір усів, вироблених вказаною клітиною-хазяїном. 20. Клітина, що містить вірус відповідно до будьякого з пп.1-10. 21. Використання коров'ячого поксвірусного ATI промотору або його похідної, визначеної в будь якому з пп.1-7, для експресії кодуючих послідовностей у MVA. 22. Спосіб продукування рекомбінантного MVA за будь-яким з пп.1-10, що включає етап введення експресійної касети в геном MVA. 23. Спосіб індукування імунологічної відповіді в живому тваринному організмі, що включає введення вірусу згідно з будь-яким з пп.1-10 або композиції згідно з п.12 тварині або людині, що лікується. 24. Спосіб за п.23, що включає введення принаймні 102 ТСID50 (доза, що інфікує тканинну культуру tissue culture infectious dose) вірусу. 25. Спосіб за будь-яким з пп.23-24 або використання за п.13, де вірус, композиція або вакцина застосовується в терапевтично ефективних кількостях, при першій інокуляції ("первинній інокуляції") та при другій інокуляції ("підсилюючій інокуляції"). Винахід стосується рекомбінантного модифікованого вірусу коров'ячої віспи Ankara, який містить у вірусному геномі експресійну касету, яка включає промотор ATI коров'ячої віспи або його похідну, та кодуючу послідовність, де експресія кодуючої послідовності регулюється вказаним промотором. Вірус може бути придатним для використання у якості вакцини, або як частина фармацевтичної композиції. Рекомбінантні поксвіруси широко використовуються для експресії чужорідних антигенів у інфікованих клітинах. Більше того, рекомбінантні поксвіруси на даний момент тестуються як надзвичайно перспективні вакцини для індукування імунної відповіді проти чужорідного антигену, експресованого з поксвірусного вектору. Найбільш популярними є, з одного боку віруси віспи птахів, а з іншого боку віруси курячої віспи. [В US 5,736,368 та US 6,051,410] розкривають рекомбінантний штам вірусу коров'ячої віспи Wyeth, який експресує антигени та протеїни ВІЛу. [US 5,747,324] розкриває рекомбінантний штам вірусу коров'ячої віспи NYCBH, що експресує антивірусні гени. [ЕР 0243029] розкриває рекомбінантний штам вірусу коров'ячої віспи Western Reserve, що експресує ретровірусні гени людини. Середньому спеціалісту в даній галузі техніки відомі різноманітні промотори для експресії гетерологічних генів у поксвірусах, наприклад такі, як 30К та 40К [дивись, наприклад, US 5,747,324], сильний синтетичний ранній/пізній промотор [дивись, наприклад, Sutter et al., Vaccine (1994) 12, 1032.40], Р7.5 промотор [дивись, наприклад, Endo et al., J. Gen. Virol. (1991) 72, 699.703] та промотор, одержаний з гену включення А-типу (ATI) вірусу коров'ячої віспи [Li et al., J. Gen. Virol. (1998) 79, 613]. Всі ці промотори були використані в рекомбінантних вірусах коров'ячої віспи для експресії гетерологічних генів та було показано, що можуть дуже ефективно експресувати вказані гени, що може призводити до відносно високих кількостей протеїну, який кодується гетерологічним геном. Для багатьох підходів імунізації надзвичайно бажаним є те, щоб антиген, проти якого індукують імунну відповідь експресувався у великих кількостях. Однак, так буває не завжди. Було описано, що різні типи цитотоксичних Т-клітин (CTL) індукуються імунною системою залежно від концентрації антигену. Низька авидність CTL індукується високими концентраціями антигену, тоді як висока авидність CTL індукується низькими концентраціями антигену. Було продемонстровано, що CTL з високою авидністю є більш ефективними при вивільненні заражуючого вірусу в тваринних тестсистемах ніж CTL з низькою авидністю. Більш того, було продемонстровано, що високі концентрації антигену можуть пригнічувати або навіть знищувати активність CTL. Підсумовуючи, можна сказати, що іноді більш бажано використовувати невеликі кількості антигену для індукування більших кількостей CTL високої авидності й, таким чином, індукувати ефективну імунну відповідь [Berzofsky et al., Immunological Reviews (1999) 170,151-172]. Об'єктом даного винаходу було створення вірусної системи, яка базується на вірусі коров'ячої віспи, та дозволяє експресувати гетерологічний ген, включений в геном вірусу коров'ячої віспи у відносно малих кількостях після його введення тваринам, включаючи людину, що може бути передумовою для індукування відносно великої кількості CTL з високою авидністю. Об'єкт було одержано введенням у вірусний геном рекомбінантного модифікованого вірусу коров'ячої віспи Ankara (MVA) експресійної касети, яка включає коров'ячий поксвірусний ATI промотора або його похідний та кодуючу послідовність, в якій експресія вказаної послідовності регулюється вказаним промотором. Неочікуваним було те, що ATI промотор має відносно низьку активність у MVA, 5 82998 хоча відомо, що даний промотор дуже активний в інших поксвірусних системах (e.g. Li et al., J. Gen. Virol. (1998) 79, 613). Також, в розділі прикладів даного опису було продемонстровано, що ATI промотор має від двох до чотирьох разів меншу активність в системах основаних на MVA, ніж у системах основаних на інших штамах вір усу коров'ячої віспи, таких як Western Reserve, Elstree або Copenhagen. Отже, ATI промотор в MVA є гарним промотором для експресії генів, які кодують протеїни, проти котрих індукуються високо авидні CTL. Модифікований вірус коров'ячої віспи Ankara (MVA) відноситься до вірусу коров'ячої віспи, члену роду Orthopoxvirus родини Poxviridae. MVA було створено 516-ю послідовними пасажами штаму вірусу коров'ячої віспи Ankara (CVA) на фібробластах ембріону курчати (for review see Mayr, A. , et al.[1975], Infection 3, 6-14). Наслідком цих довготривалих пасажів став MVA, вірус із видаленими 31 тис. основ геномної послідовності, та таким чином, він був охарактеризований, як дуже обмежений по відношенню до клітин-хазяїнів до пташиних клітин. (Me yer, H. et al., J. Gen. Virol. 72, 10311038 [1991]). На різноманітних тваринних моделях було продемонстровано, що одержаний MVA в значній мірі є авірулентним (Mayr, A. & Danner, K. [1978] Dev. Biol. Stand. 41: 225.34). Також, цей штам MVA було досліджено в 5 клінічних випробуваннях, як вакцину для імунізації проти віспи людини (Mayr et al., Zbl. Bakt. Hyg. I, Abt. Org. В 167, 375-390 [1987], Stickl et al., Dtsch. med. Wschr. 99, 2386-2392 [1974]). Згідно даного винаходу, може бути використаний будь-який штам MVA. прикладами штамів MVA, використаних відповідно до даного винаходу та депонованих у відповідності до вимог Будапештської угоди є штами MVA 572 та 575 депоновані в Європейській колекції тваринних клітинних культур (European Collection of Animal Cell Cultures) (ECACC), Salisbury (UK) під депозитними номерами ECACC V94012707 та ECACC V00120707, відповідно, та MVA-BN під депозитним номером ECACC V00083008. Найбільш переважним штамом MVA є MVA-BN або його похідні. Характерні ознаки MVA-BN, опис біологічних методів, котрі дозволяють чи є штам MVA MVA-BN або його похідним та методи, котрі дозволяють одержувати MVA-BN або його похідні розкрито в [WO02/42480]. Зміст цієї заявки включено в дану заявку шля хом посилання. Для розмноження MVA, е укаріотичні клітини були інфіковані цим вірусом. Еукаріотичні клітини є клітинами, котрі є придатними до інфікування відповідним поксвірусом та дозволяють реплікувати та продукувати інфекційний вірус. Для MVA прикладом такого типу клітин є фібробласти курячих ембріонів (CEF) та клітини ВНК [Drexler I., Heller K., Wahren В., Erfle V. and Sutter G. "Highly attenuated modified vaccinia Ankara replicates in baby hamster kidney cells, 30 a potential host for virus propagation, but not in various human transformed and primary cells" J. Gen. Virol. (1998), 79, 347-352]. Клітини CEF можуть культивуватися за умов відомих досвідченій персоні. Переважно, клітини CEF культи 6 вуються в безсироватковому середовищі у лабораторних колбах або ролерних флаконах. Інкубація зазвичай триває від 48 до 96 годин при 37°С±2°С. Для інфікування MVA, переважно, використовували при значенні інфікувальної кількості (МОЇ) від 0,05 до 1 ТСID50 й інкубація зазвичай проводилася від 48 до 72 години при 37°С±2°С. Послідовність гену промотору включеного протеїну коров'ячого поксвірусу тип у A (ATI промотор), є відомим досвідченій особі. В цьому контексті робиться посилання на об'єкт у Genebank із вхідним номером D00319. Переважна послідовність ATI промотора є такою як приведено на SEQ ID: No. 1 та є наступною: 5' GTTTT GAATA АААТТ TTTTT АТААТ АААТ 3'¢ Згідно даного винаходу, є можливим використовувати ATI промотор як зазначено в SEQ. ID.:No. 1 або використовувати похідний ATI промотора, котрі можуть бути підпослідовностями послідовності, згідно із SEQ. ID.:No. 1. Термін „підпослідовність послідовності, згідно із SEQ. ID.:No. 1" відповідає більш коротким фрагментам послідовності SEQ. ID.:No.1, котрі зберігають таку ж активність, що і промотор, зокрема пізній промотор вірусу коров'ячої віспи. Типовий фрагмент SEQ. ID.:No.1 має довжину, принаймні, 10 нуклеотидів, більш бажано, 15 нуклеотидів, ще більш бажано 20 нуклеотидів, та найбільш бажано 25 нуклеотидів послідовності SEQ. ID.:No.1. Підпослідовність може включати від 25 до 29 нуклеотиди SEQ. ID.:No.1, наприклад, послідовність 5'-ТАААТ-3', розташовану на 3' кінці SEQ. ID.:No.1. Підпослідовність також може включати від 22 до 29 нуклеотиди SEQ. ID.:No.1, наприклад, послідовність 5'-ТААТАААТ-3', розташовану на 3' кінці SEQ. ID.:No.1. Промотор може бути вставлений раніше кодуючою послідовності, таким чином, що н уклеотиди з 28 по 29 з SEQ. ID: 1 (підкреслені в послідовності вище) є частиною стартового кодону трансляції 5' ATG 3'. Також, промотор може бути відокремлений декількома нуклеотидами від стартового кодону трансляції. Спейсер між 3' кінцем промотору, згідно із SEQ ID.: No 1 та А в 5' ATG 3' стартового кодону, бажано менший за 100 нуклеотидів, більш бажано менший за 50 нуклеотидів, ще більш бажано менший за 25 нуклеотидів. Однак, спейсер може мати таку довжину, на скільки промотор здатний спрямовувати експресію кодуючою послідовності розташованої за промотором. Похідний ATI промотора також може бути послідовністю, яка має одну або більше нуклеотидних замін, делецій та/або вставок котрі відповідають послідовності SEQ ID.:No.1, де вказані похідні залишаються активними як промотор, зокрема пізній промотор, вірусу коров'ячої віспи. Послідовність яка має одну або більше нуклеотидних замін є послідовністю в котрій один або більше нуклеотид послідовності, згідно з SEQ ID.: No.1, замінені іншими нуклеотидами. Послідовність яка має одну або більше нуклеотидних вставок є послідовністю в котрій один або більше нуклеотид вставлені в одному або більше місць в послідовності, згідно з SEQ ID.: No.1. Послідовність, яка має одну або більше нуклеотидних делецій є послідовністю в 7 82998 котрій один або більше нуклеотид видалені в одному або більше місць в послідовності, згідно з SEQ ID.: No.1. В похідних SEQ ID.: No. 1. делеції, заміни та вста вки можуть комбінуватися в одній послідовності. Бажано, щоб похідні мали гомологію, принаймні, 40%, більш бажано, принаймні 60%, ще більш бажано, принаймні, 80%, найбільш бажано, принаймні, 90% порівняно із послідовністю SEQ ID.: No.1. Згідно із найбільш переважним втіленням, не більше 6 нуклеотидів, ще більш бажано, не більш як 3 нуклеотиди, можуть бути замінені, видалені, та/або вставлені в послідовність SEQ ID: No. 1. Зокрема, було б бажаним залишити нуклеотиди з 25 по 29 з SEQ. ID.:No. 1, тобто, послідовність 5'-ТАААТ-3' в промоторі для досягнення максимальної активності. Також було б бажаним залишити нуклеотиди з 22 по 29 з SEQ. ID.:No. 1, тобто, послідовність 5'-ТААТАААТ-3' в промоторі. Маса праць в даній галузі дозволяє досвідченій персоні передбачити, котрі з похідних SEQ ID.: No.1 будуть мати біологічну активність як промотор вірусу коров'ячої віспи, зокрема, як пізній промотор вірусу коров'ячої віспи. В цьому контексті можна посилатися на Chakrarbarti et al., Biotechniques (1997) 23, 1094-1097 and Davison and Moss, J. Мої. Biol. (1989) 210, 771-784. Більш того, чи фрагмент зберігає активність як промотор вірусу коров'ячої віспи, зокрема, як пізній промотор вірусу коров'ячої віспи можна бути з'ясовано будьякою досвідченою особою. Зокрема, похідна послідовність може бути клонована в сайти раніше репортерного гену плазмідного конструкту. Вказаний конструкт може бути трансфікований в еукаріотичну клітину або клітинну лінію, таку як клітини CEF або ВНК, котрі були інфіковані MVA. Експресія репортерного гену потім визначається і порівнюється із експресією репортерного контрольованого послідовністю згідно із SEQ ID.: No. 1. Експериментальні значення відповідають прикладам приведеним у вказаному описі. Похідні, згідно даного винаходу, мають промоторну активність у вказаних тест системах, принаймні 10%, бажано, принаймні 30%, більш бажано, принаймні 50%, ще більш бажано, принаймні 70%, найбільш бажано, принаймні 90%, порівняно із активністю промотора послідовності SEQ ID.: No.1. Також ті похідні SEQ ID.: No.1, які мають промоторну активність вищу за активність SEQ ID.: No. 1, також охоплюються даним винаходом. У більш загальних словах, даний винахід відноситься до використання коров'ячого поксвірусного промотора або його похідного, як вище зазначено для експресії кодуючою послідовності MVA. ATI промотор може використовуватися для експресії гену який вже є частиною геному MVA. такий ген може бути геном, який природно є частиною вірусного геному або чужорідним геном, котрий вставлено в геном MVA. в цих випадках ATI промотор вставляють раніше гену в геномі MVA, експресія котрого буде керуватися ATI промотором. 8 ATI промотор також може використовуватися для регуляції експресії гену, котрий не є частиною геному MVA. в цьому випадку, бажано створити експресійну касету, яка включала б ATI промотор та кодуючу послідовність, експресія котрої буде керуватися ATI промотором та вставити таку експресійну касету в геном MVA. Переважними сайтами вставки є вибрані з (і) сайтів делецій геному MVA, особливо геному вірусу коров'ячої віспи Copenhagen або (іі) міжгенні регіони MVA. Термін "міжгенний регіон", переважно, відноситься до тих частин вірусного геному, розташованих між двома сусідніми генами, в яких нема ні кодуючи послідовностей а ні регулюючих. Однак, сайти вставки не обмежуються цими переважними сайтами, оскільки даними винаходом передбачається, що експресійна касета може бути вставлена будь-де у вірусному геномі, в такій кількості скільки це потрібно для одержання рекомбінантів, котрі можуть бути ампліфіковані й розмножені, принаймні в одній клітинній культурі, такій як фібробласти курячого ембріону (CEF). Отже, касети вставки, також може бути вставлена, наприклад, в неважливі гени або гени, функція яких може бути доповнена клітинною системою, використовуваною для розведення MVA. Методи, необхідні для конструювання рекомбінантного MVA є відомими досвідченій особі. Як приклад, експресійна касета та/або ATI промотор або його похідний може бути вста влений в геном MVA гомологічною рекомбінацією. Для цього нуклеїнова кислота трансфікується в клітинну лінію таку, як клітини CEF або ВНК, де нуклеїнова кислота містить експресійну касету та/або ATI промотор або його похідний фланкований нуклеотидними проміжками, котрі є гомологічними до регіону в геномі MVA, в котрий вставлена експресійна касета та/або ATI промотор або його похідний. Клітини, інфіковані MVA та в інфікованих клітинах гомологічна рекомбінація відбувається між нуклеїновою кислотою та вірусним геномом. Також, можливо, спочатку інфікувати клітини MVA, а потім трансфікувати нуклеїнову кислоту в інфіковані клітини. Наступна рекомбінація відбувається в клітинах. Рекомбінантний MVA потім селектується загальновідомими на практиці методами. Конструктування MVA не обмежується цим конкретним методом. До того ж, для цієї мети може бути застосовано будь-який прийнятний метод відомий, досвідченій особі. ATI промотор в MVA може використовуватися для керування експресією будь-якої кодуючою послідовності. Кодуюча послідовність, переважно, може кодувати принаймні один антигенний епітоп або антиген. В цьому випадку рекомбінантний MVA може використовуватися для експресії вказаного антигену після інфекції клітин в організмі, наприклад, ссавця, включаючи людину. Презентація вказаного антигену/епітопу може викликати імунну відповідь в організмі, що може призводити до вакцинації організму проти агенту, з якого походить антиген/епітоп.Також, антиген/епітоп може бути частиною більшої амінокислотної послідовності, такої як поліепітоп, пептид або протеїн. Прикладами таких поліепітопів, пептидів або протеїнів мо 9 82998 жуть бути поліепітопів, пептидів або протеїнів одержані з (і) вірусів, таких як ВІЛ, HTLV (вірус Тклітинного лейкозу людини), герпесвірусу, вірусу тропічної лихоманки, поліовірусу, вірусу кору, вірусу свинки, вірусу гепатиту та таке інше, (іі) бактерій, (ііі) грибів. Також кодуюча послідовність може кодувати терапевтичну речовину, таку як інтерлейкини, інтерферони, рибозими або ензими. Рекомбінантний MVA може вводитися в організм тварини або людини згідно знанням досвідченої особи. Отже, рекомбінантний MVA, згідно даного винаходу може бути корисним як медикамент (тобто, фармацевтична композиція) або вакцина. Фармацевтична композиція або вакцина, взагалі, може включати один або більше прийнятних та/або дозволених носіїв, добавок, антибіотиків, консервантів, ад'ювантів, дилюентів та/або стабілізаторів на доданок до рекомбінантного MVA. Такими допоміжними речовинами можуть бути вода, сольовий розчин, гліцерил, етанол, зволожуючі або емульгуючи агенти, рН-буферні речовини, або подібні. Придатними носіями, зазвичай є великі, повільно розщеплювані молекули, такі як протеїни, полісахариди, полі молочні кислоти, полігліколеві кислоти, полімерні амінокислоти, амінокислотні сополімери, ліпідні агрегати або подібні до них. Для виготовлення фармацевтичних композицій або вакцин рекомбінантний MVA перетворюють у фізіологічно прийнятну форму. Це може бути проведено ґрунтуючись на досвіді виготовлення поксвірусних вакцин, які застосовували для вакцинації проти віспи людини (як описано у Stickl, H. et al. [1974] Dtsch. med. Wschr. 99, 2386-2392). Наприклад, очищений вірус зберігається при D-80°C при титрі 5x108 TCID50/ml у 10mM Tris, 140тМ NaCI рН 7.4. Для виготовлення ін'єкційних доз, наприклад брали 101-109 часток рекомбінантного вірусу, відповідно до даного винаходу й ліофілізували у фосфа тно-буферному сольовому (PBS) в 2% пептону та 1% альбуміну людини в ампулі, бажано в скляній ампулі. Також, вакцинні дози можуть бути виготовлені поетапною ліофільною сушкою вірусу у композиції. Композиція може містити додаткові домішки як манітол, декстран, цукор, гліцин, лактоза або полівінілпіролідон або інші допоміжники, такі як антиоксиданти або інертний газ, стабілізатори або рекомбінантні протеїни (наприклад, сироватковий альбумін людини) придатні для застосування in vivo. Типова вірусовмісна композиція придатна для ліофільного сушіння включає 10mМ Tris-буфер, 140mМ NaCI, 18.9г/л декстрану (MW 36000-40000), 45г/л сахарози, 0.108г/л Lглютамінової кислоти моногідрату мононатрієвої солі з рН 7.4. Скляна ампула потім запаюється й зберігається при температурі в межах від 4°С до кімнатної на протязі декількох місяців. Хоча, за відсутності потреби, майже необмежено можна зберігати при температурі нижче -20°С. Для вакцинації або терапії ліофілізат або ліофільно висушений може бути розведений у від 0.1 до 0.5мл водного розчину, переважно води, фізіологічного сольового розчину або Tris-буферу, та застосовується як системно так і місцево, напри 10 клад, парентерально, внутрішньом'язево, або іншим прийнятним шляхом введення відомим досвідченому професіоналу. Спосіб застосування, доза та кількість застосувань може бути оптимізовано відомим чином досвідченим спеціалістом. Отже, згідно даного втілення, винахід відноситься до способу впливу, переважно індукування імунологічної відповіді в живому організмі тварини включаючи людину, і включає введення вірусу, композиції або вакцини, згідно даного винаходу тварині або людині, що лікується. Зазвичай, вакцинні дози містять, принаймні, 102, переважно, принаймні, 104, більш переважно, принаймні, 106, ще більш переважно, принаймні, 108 TCID50 (доз, що інфікують тканинну культур у) вірусу. Однією з переваг рекомбінантного MVA, згідно даного винаходу, зокрема, рекомбінантного MVABN та його похідних, котрі можуть бути використані для первинного-підсилювального введення. Таким чином винахід також відноситься до методу, де вірус, композиція, або вакцина вводиться тварині, включаючи людину, за її потреби, в терапевтично ефективних кількостях при першій інокуляції ("первинній інокуляції") та другій інокуляції ("підсилюючій інокуляції"). Винахід, також, стосується методів введення кодуючою послідовності в клітини-мішені, включаючи інфікування клітин-мішеней вірусом, згідно даного винаходу. Клітини-мішені можуть бути клітинами, в котрих вірус здатен реплікуватися, такі як клітини CEF або ВНК або клітини, котрі можуть бути ін фіковані MVA, в котрих вірус, однак, не може реплікуватися, такі як усі типи клітин людини. Винахід, також відноситься до методу продукування пептиду, протеїну та/або вірусу, включаючи інфікування клітини-хазяїна рекомбінантним вірусом, згідно даного винаходу, наступним культивуванням інфікованої клітини-хазяїна з сприятливих умов, та подальшим виділенням та/або збагаченням пептиду та/або протеїну та/або вірусів, продукованих даною клітиною-хазяїном. Якщо це призначається для продукування, тобто ампліфікації вірусу, відповідно до даного винаходу, клітина має бути клітиною в котрій вірус здатний реплікуватися, такі як клітини CEF або ВНК. Якщо це призначається для продукування пептиду/протеїну, кодованого вірусом, переважно пептиду/протеїну, кодованого кодуючою послідовністю, експресія котрої керується ATI промотором або його похідним, клітини можуть бути будьякими клітинами, котрі можуть бути інфіковані рекомбінантним вірусом, та котрі дозволяють експресуватися протеїнам/пептидам, кодованим МВА. Винахід, також стосується клітин, інфікованих вірусом, згідно даного винаходу. Стислий опис фігур. Фіг.1: Результат PCR-реакції для виявлення присутності РНК експресованої ATI промотором в рекомбінантному MVA після інфікування клітин CEF (дивись Приклад 2). Фіг.2: Результат Western blot із протеїнами виділеними з клітин інфікованих різними рекомбінантними MVA. Фігура 2А: умови невідновлюючі, протеїни нетеплооброблені; Фіг.2В: умови відновлюючі, протеїни теплооброблені. Смуги 11 82998 1,3,4 є клітинними лізатами клітин інфікованих MVA-ATI-NS1, MVA-GFP, а також неінфікованих клітин, відповідно. Смуги 5, 7, 8 є супернатантами клітин інфікованих MVA- ATI-NS1, MVA-GFP та супернатанти клітин контролю, відповідно. Смуги 2 та 6 залишилися порожніми. Наступні приклади далі проілюструють даний винахід. Має бути добре зрозумілим досвідченою особою, що наведені приклади ні в якому разі не можуть бути інтерпретовані, як обмеження застосовності даного технології створеної даним винаходом до меж даних прикладів. Приклад 1: Активність коров'ячого поксвірусного ATI промотора в різних вірусу коров'ячої віспи Метою цього прикладу було проаналізувати силу ATI промотору в різних штамах вір усу коров'ячої віспи. Індукування Коров'ячий поксвірусний ATI промотор було злито із GUS (|3-Глюкоуронідаза Е. соїі), репортерним геном для експресійного аналізу. Клітини ВНК (нирки малого ховраха) були інфіковані різними вірусами коров'ячої віспи та трансфіковані плазмі дою, яка містила ATI промотор злитий із геном GUS. Досліджувані штами вірусу коров'ячої віспи включали OVA, Copenhagen, Elstree, IHD, Western reserve та MVA-BN. Якщо промотор був функціональним, GUS був би екс пресованим і міг би бути визначеним ферментативною реакцією. Матеріали і методи - клітини ВНК (ECACC No. 84100501) - усі віруси коров'ячої віспи використовувалися у титрі 7.5´107×ТСID50/мл - Плазміда pBNX73 (pBluescript+ATI промотор+GUS) - трансфекційний набір Effectene (Qiagen) - клітинне культуральне середовище DMEM (Gibco BRL) - FCS (Gibco BRL) - Лізисбуфер (PBS+0.1% Triton+1mM протеазного інгібітору) - субстрат GUS 1mM (р-Нітрофеніл-бета-(D)люкуронід; Sigma, Cat. No.N1627) - Стоп-розчин 2.5М (2-аміно-2-метил-1,3пропандіол; Sigma, Cat. No. A 9754) Метод: Посів клітин 5´105 клітин ВНК було посіяно при реакції трансфекції в лунки 6-линкової планшетки й утримувалося в DMEM/10%FCS на протязі ночі при 37°С та 5% СО2. Інфікування/трансфікування Клітини було інфіковано різними штамами вірусу коров'ячої віспи Клітини було інфіковано різними штамами вірусу коров'ячої віспи (МОI 0.1) в 0.5мл DMEM/10% FCS на лунку й інкубували на протязі 1г при кімнатній температурів шейкері. Трансфекція виконувалася як описано в прописі виготовника. 2мкг плазмід було розведено в буфері ЕВ (100мкл загального об'єму). Після додавання 3.2мкл енхансерного розчину розчин було перемішано й інкубовано на протязі 5хв. при кімнатній температурі. Потім було додано 10мкл реагенту Effectene, суспензію розмішали й інкубували а протязі 10хв. 12 при кімнатній температурі. З клітин було видалено вірусну суспензію й додано 1.6мл DMEM/10%FCS. До суміші ДНК Effectene було додано 0.6 мл DMEM/10%FCS й додано до клітин при обертанні культуральної платівки. Потім клітини інкубували на протязі 48 годин. Збирання клітин З клітин було видалене середовище й додано 0.5мл лізис-буферу. Після 15хв. встряхування при кімнатній температурі клітини було зіскребемо в лізис-буфер, поміщено в реакційну пробірку 1.5мл сильно перемішували. Лізовані клітини було центрифуговано на протязі 1хв. при 500rcf та 4°С, прозорий сурфактант було перенесено у свіжим посуд й зберігали при -20°С до використання. Визначення активності GUS 10мкл клітинного екстракту (= протеїновий вихід з 2´104 клітин) було додано до 1 мл попередньо підігрітого субстратного розчину (37°С) й інкубовано при 37°С до появи жовтого забарвлення. Зразки одразу були поміщені на лід й додано 0.4мл стоп-розчину. Екстинкцію було визначено при 415нм й порівняно із активністю GUS як значення між 0,05 та 2,0 в лінійній області. Субстрат ний розчин було використано як зразок, а клітинний екстракт неінфікованих клітин як негативний контроль. Результати: Підрахунок активності GUS експресованої конструктом ATI промотор - ген GUS в клітинах ВНК інфікованих різними штамами вірусу коров'ячої віспи дало наступні результати: Штами вірусу коров'ячої віспи викорис- Активність тані для інфікування клітин ВНК трансGUS фікованих PBNX73 OVA 1.30 Copenhagen 1.86 Elstree 2.07 IHD 1.30 Western Reserve 0.96 MVA-BN 0.48 Приклад 2: Експресія чужорідних генів вставлених в геном MVA й регульованих коров'ячим коксовим ATI промотором Метою цього прикладу було показати, що ATI промотор здатний регулювати й експресувати гени вставлені в геном MVA. Введення Коров'ячий поксвірусний промотор ATI було злито із неструктурним (NS) геном 1 вірусу тропічної лихоманки. Експресійна касета була вставлена в рекомбінантний вектор представлений послідовністю гомологічною до геному MVA. В одержаній рекомбінантній плазміді експресійна касета була франкована послідовностями гомологічними до послідовностей геному MVA котрим була вставлена експресійна касета. Клітини CEF було інфіковано MVA-BN та трансфіковано рекомбінантним вектором, який містив ATI промотор експресійної касети NS1. В клітинах відбувалася гомологічна рекомбінація між геномом MVA та рекомбінаційною плазмідою яка призводила до появи рекомбінантного геному MVA. Після декількох циклів очи 13 82998 щення було проаналізовано чи був протеїн NS1 в рекомбінантному MVA екс пресований з ATI промотору. В паралельних експериментах експресійна касета включала послідовність, яка кодувала прототип ВІЛ під контролем ATI промотору, була вставлена в геном MVA й знову проаналізована на активність ATI промотору в MVA й експресію прототипу ВІЛ. Матеріали й обладнання: - первинні клітини CEF - клітини нирки малого ховраха (ВНК; депоновані під 85011433 в European Collection of Animal Cell Cultures) - MVA-BN із ти тром 108 TCID50/ml. - плазміди pBN74 та pBN84: pBN74 включає послідовність, яка кодує прототип ВІЛ під контролем ATI промотору та pBN84 включає кодуючу послідовність протеїну вірусу тропічної лихоманки, під контролем ATI промотору. На додачу, експресійні касети, які містили кодуючу послідовність протеїну вірусу тропічної лихоманки та прототип ВІЛ, відповідно, обидві плазміди містили ген резистентності G418 та ген, що кодує зелений флуоресцентний протеїн як маркерні гени. Маркерні гени, так само як і експресійні касети NS1 вірусу тропічної лихоманки та прототипу ВІЛ, відповідно, були фланковані послідовностями MVA, гомологічними до регіону в геномі MVA в котрий вставлялися гетерологічні гени. - трансфекційний набір Effectene (Qiagen) - клітинне культуральне середовище VP-SFM (Gibco BRL) - набір для виділення РНК RNeasy (Qiagen) - вільні ДНК-аза та РНК-аза (Roche) - MML V реверсна транскриптаза (Promega) - ДНК-полімераза Taq (Roche) - інгібітор РНК-ази RNAsin (Promega) - у MWG (Німеччина), були придбані наступні олігонуклеотиди: праймер oBN465 ggtctgatttccatcccgtac (21 нуклеотид), використаний для реакції реверсної транскриптази; праймер oBN463 gaactgaagtgtggcagt (18 нуклеотидів), використаний для PCR; праймер oBN464 cggtggtaatgtgcaagatc (20 нуклеотидів), використаний для PCR. Методи: Інтеграція в геном MVA шляхом гомологічної рекомбінації Описані вище плазміди pBN74 або pBN84 були використані для інтеграції кодуючою послідовності прототипу ВІЛ та експресійної касети NS1 тропічної лихоманки, відповідно, в геном MVA гомологічною рекомбінацією між послідовностями MVA, які фланкують експресійні касети pBN74 або pBN84 на одному боці гомологічної цільової послідовності в геномі MVA з іншого боку. Це досягнуто трансфекцією лінеаризованої плазміди pBN74 або pBN84 в клітинах фібробластів ембріону курча (CEF) перед інфікуванням MVA при низькій множинності зараження. Точніше, клітини CEF були посіяні в 6-ти лункові платівки та утримувалися в VP-SFM на протязі ночі при 37°С та 5% СО2. клітини було інфіковано MVA-BN (МОІ 1.0) в 0.5 мл VPSFM на лунку й інкубовано на протязі 1г при кімнатній температурі в шейкері. Трансфекцію клітин як 14 pBN74 або pBN84 проводили як описано в інструкції виробника. Через 48 годин після інфікування або досягнення інфекцією конфлюентності було приготовано вірусний екстракт й поміщено в -20°С готовим до селекції та очищення клону бажаного рекомбінантного MVA (rMVA). Селекція рекомбінантного MVA (rMVA) та очищення клону Елімінація нерекомбінантного MVA (пустого вірусного вектору) та ампліфікація MVA досягалася інфікуванням злитих клітин фібробластів курячих ембріонів (CEF) при низькому МОЇ в присутності G418 (кількість G418 має бути оптимізовано для визначення найвищої дози, введення котрої на вбиває клітини CEF). Будь-який вірус, котрий не містить інтегрований ген NPT II не буде реплікуватися в присутності G418 в клітинному підтримуючому середовищі. G418 інгібує реплікацію ДНК, але оскільки клітини CEF будуть стабільно в нерепліковному стані, вони не зможуть піддаватися дії G418. Клітини CEF інфіковані rMVA можна відрізнити під флуоресцентним мікроскопом завдяки експресії посиленого зеленого флуоресцентного протеїну. Вірусні екстракти з етапу гомологічної рекомбінації послідовно розводять й використовують для інфікування свіжих клітин CEF в присутності G418. Інфіковані клітини покривають низькоплавкою агарозою. Через 2 дні після інфікування, платівки оглядають під флуоресцентним мікроскопом визначаючи одиничні локуси зведених інфікованих клітин. Ці клітини маркують й агарозу, яка містить очами інфікованих клітин переносять на 1.5мл центрифужні пробірки із стерильним стабілізуючим клітинним середовищем. Вірус вивільнюють з агарози триразовим заморожуваннямтаненням в пробірці при -20°С. Рекомбінантний MVA із вставленою експресійною касетою NS1 тропічної лихоманки, описаний в цьому винаході названо MVA-ATI-NS1 Визначення експреси РНК з ATI промотору в рекомбінантному MVA Для Визначення експресії РНК з ATI промотору в клітинах інфікованих рекомбінантним MVA, було проведено екстракцію РНК як описано в інструкції виробника (Rneasy Mini Protocol for the Isolation of Total RNA from Animal Cells). Реакція ДНК-зного розщеплення було виконано додаванням 3мкл вільних ДНК-аза та РНК-аза (=30U; Roche), 3мкл 10´буфер А (звичайно використовується для рестрикцій, Roche) до 5мкг РНК в доведеному водою до 30мкл об'ємі. Суміш інкубува ти на протязі 90хв. при 37°С. РНК було очищено із застосуванням колонок Rneasy згідно інструкції виробника. Для реверсної транскрипції 2мкг РНК було змішано із 1мкг OBN465 для реверсної транскрипції й загальний об'єм було доведено додаванням води до 10мкл. Це було інкубовано на протязі 5хв. при 70°С й пробірки було перенесено на лід. Потім 5мкл 5´буфер у, 5мкл dNTP Mix (10мкМ), 0.5мкл Rnasin, 2мкл M-MLV RT (200U) та 2.5 Н2О було додано і суміш було інкубовано на протязі 60хв. при 42°С. Для ампліфікації PCR 5мкл RT-суміші було змішано із 36мкл Н 2О, 5мкл 5´буферу, 1мкл dNTP, 2мкл кожного з праймерів OBN4G3 та OBN464 15 82998 (10мкМ), та 1 мкл Taq-полімерази. ДНК було ампліфіковано у 25 циклів (1мін. тривалість, температура прогріву 55°С) й проаналізовано на гельелектрофорезі. Визначення протеїну NS1 вірусу тропічної лихоманки, експресованого в клітинах інфікованими MVA-ATI-NS1 Колби 25cm 2 із приблизно на 80% конфлюентними мікрошарами клітин ВНК 100 мкл MVA-ATINS1 вірусного матеріалу розведеного до 1´107 в МЕМa із 1% FCS перемішано на протязі 30хв. 5мл МЕМa із 3% FCS було додано в кожну колбу й інкубовано при 30°С in а СО2 інкубаторі. Колби і виймали через 48 годин. Супернатант було видалено з колб та від центрифуговано при 260g на протязі 10хв. при 4°С. Супернатант зберігали в аліквотах при -80°С. Пелети відмивали двічі 5мл в 1xPBS й потім ресуспендовано в 1мл гіпотонічного буферу із 1% ТХ100. Клітинні лізати збирали і відцентрифуговували на протязі 5хв. при 16,000g й супернатант зберігаються в мікроцентрифужних пробірках при -80°С. Колби інокульовані контрольними вірусами й неінфіковані колби були також оброблені у такий самий спосіб як описано вище. Клітинні/вірусні лізати та супернатант були оброблені або невідновлівальним або відновлювальними буферами як без підігріву так і з підігрівом. Протеїни були розділені на 10% SDS PAGE та перенесені на нітроцелюлозні мембрани. Плями було тестовано на протязі ночі із об'єднаною сироваткою виздоровлюючих (PPCS), а саме пацієнтів, котрі страждали на тропічну лихоманку в розведення 1:500. Після трикратного відмивання 1XPBS плями були інкубовані з анти-людською lgGхроновою пероксидазою (HRP) (DAKO) на протязі 2 годин при кімнатній температурі. Після цього Комп’ютерна в ерстка Н. Лисенко 16 плями було відмито, як описано вище й пофарбовано 4-хлоро-1-нафтолом. Результати приведено на Фіг.2. Результати Було показано, що ген NS1 так само як і прототип ВІЛ можуть бути експресовані, коли регулюються коров'ячим поксвірусним ATI (Фіг.1). Відповідна мРНК була чітко визначеною. Точніше, очікуваний сигнал 926 нуклеотидів був чітко визначений після RT-PCR у зразку РНК (Фіг.1, смуга 2). При використанні зразку для лише PCR, сигнал визначено не було (Фіг.1, смуга 3). Таким чином може бути виключено помилковий позитивний сигнал спричинений присутністю ДНК. Фіг.1, смуга 4 показує результат PCR із плазмідним позитивним контролем. Як очікувалося розмір PCR продукту був ідентичний до PCR продукту після PT-POR зразку РНК. Фіг.1, смуга 5 показує результат RTPCR реакції із негативним контролем (вода). Фігура 1, смуга 1 та 6 є маркерами молекулярної маси (100bp відмітка). Результати western blots показують, що NS1 експресувався в клітинах інфікованих MVA-ATI-NS1. NS1 експресувався в правильній конформації як димер без нагрівання як показано на смузі 1 Фіг.2А та 2В. Коли зразки було нагріто було виявлено мономер NS1 як може бути видно з Фіг.2 Результати також показують, що NS1, який експресувався в клітинах MVA-ATI-NS1 є антигенним й розпізнається об'єднаною сироваткою виздоровлюючих пацієнтів. На заключення, експерименти показали, що NS1 експресується в правильній конформації в клітинах ВНК інфікованих MVA-ATI-NS1. як димер так і мономер є антигенними й розпізнаються об'єднаною сироваткою виздоровлюючих пацієнтів. Підписне Тираж 26 прим. Міністерство осв іт и і науки України Держав ний департамент інтелектуальної в ласності, вул. Урицького, 45, м. Київ , МСП, 03680, Україна ДП “Український інститут промислов ої в ласності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюExpression of genes in modified vaccinia virus ankara using bovine poxvirus promotor аті
Автори англійськоюLEYRER SONJA, Leyrer Sonja
Назва патенту російськоюЭкспрессия генов в модифицированном вирусе коровьей оспы ankara с использованием коровьего поксвирусного промотора аті
Автори російськоюХаули Пол, Лейрер Сонья
МПК / Мітки
МПК: C12N 15/86
Мітки: використанням, промотору, генів, коров'ячої, коров'ячого, поквірусного, аті, вірусі, віспи, експресія, ankara, модифікованому
Код посилання
<a href="https://ua.patents.su/8-82998-ekspresiya-geniv-u-modifikovanomu-virusi-korovyacho-vispi-ankara-z-vikoristannyam-korovyachogo-pokvirusnogo-promotoru-ati.html" target="_blank" rel="follow" title="База патентів України">Експресія генів у модифікованому вірусі коров’ячої віспи ankara з використанням коров’ячого поквірусного промотору аті</a>
Міжгенні регіони як сайти вставки в геном модифікованого вірусу коров’ячої віспи ankara (mva)

Номер патенту: 82479
Опубліковано: 25.04.2008
Автори: Лейрер Сонья, Хаулі Пол
МПК: C12N 15/863, A61K 31/70, C07K 14/07, C07K 14/16, C07K 14/18
Мітки: коров'ячої, вставки, регіони, міжгенні, віспи, вірусу, модифікованого, mva, геном, сайті, ankara
Формула / Реферат:
1. Рекомбінантний модифікований вірус коров'ячої віспи Ankara (MVA), який містить гетерологічну послідовність ДНК, вставлену в міжгенний регіон (IGR) вірусного геному.2. MVA, згідно з пунктом формули 1, де гетерологічна послідовність ДНК вставлена в IGR між двома відкритими рамками зчитування (ORF), вибраними з групи, яка включає: 007R-008L, 018L-019L, 044L-045L, 064L-065L, 136L-137L, 148R-149L.3. MVA, згідно з пунктами 1 або 2,...
Штам mva-bn модифікованого вірусу коров’ячої віспи ankara, фармацевтична композиція, вакцина, застосування mva-bn для приготування лікарського препарату та для приготування вакцини, спосіб введення гомологічної

Номер патенту: 76731
Опубліковано: 15.09.2006
Автори: Хаулі Пол, Мейсінгер Крістін, Чаплін Пол
МПК: C12N 15/39, A61K 48/00, C12N 5/10, A61P 37/04, A61K 39/39, A61K 39/285, C12N 15/863, C12P 21/02, C12N 7/02, C12N 15/09, C12N 7/04, A61K 39/275, C12N 7/01
Мітки: композиція, фармацевтична, введення, вакцина, спосіб, віспи, застосування, препарату, штам, гомологічної, mva-bn, вірусу, коров'ячої, вакцини, приготування, лікарського, модифікованого, ankara
Формула / Реферат:
1. Штам MVA-BN модифікованого вірусу коров'ячої віспи Ankara, депонований в Європейській колекції клітинних культур (ЕСАСС), Солсбері (UK), під номером V00083008, та його похідні, що мають принаймні одну з таких властивостей:(і) здатність до репродукції шляхом реплікації у фібробластах ембріонів курчат (CEF) і в лінії клітин нирок дитинчат хом'яка ВНК, але нездатність до репродукції шляхом реплікації у клітинних лініях...
Ізольований фрагмент днк промотору гена синтази ацетооксикислот (ahas) для експресії генів в рослинах, вектор для трансформації рослин, спосіб високорівневого експресування гетерологічного гена в рослині та в

Номер патенту: 48951
Опубліковано: 16.09.2002
Автори: Сміт Джейн, Пенг Джіань Їнг., Дітріх Габріель
МПК: C12N 9/88, C12N 15/60, C12N 15/29, A01H 5/00, C12N 5/10, C12N 15/82, C12N 15/09
Мітки: високорівневого, днк, синтази, гена, експресування, генів, ацетооксикислот, рослинах, рослин, фрагмент, ізольований, експресії, спосіб, гетерологічного, вектор, промотору, рослини, ahas, трансформації
Формула / Реферат:
1. Изолированный фрагмент ДНК промотора гена синтазы ацетооксикислот (AHAS) для экспрессии генов в растениях, выбранный из группы, состоящей из SEQ ID №1,SEQ ID № 2,SEQ ID № 3.2. Изолированный фрагмент ДНК по п. 1, отличающийся тем, что растение является однодольным.3. Изолированный фрагмент ДНК по п. 2, отличающийся тем, что растение представляет собой кукурузу.4. Вектор для...
Спосіб прогнозування розвитку тяжкого перебігу вітряної віспи

Номер патенту: 16824
Опубліковано: 15.08.2006
Автори: Лоскутова Ірина Володимирівна, Фролов Валерій Мітрофанович
МПК: G01N 33/48
Мітки: розвитку, спосіб, прогнозування, віспи, перебігу, вітряної, тяжкого
Формула / Реферат:
Спосіб прогнозування розвитку тяжкого перебігу вітряної віспи, що включає імунологічний аналіз крові з подальшою інтерпретацією отриманих результатів, який відрізняється тим, що вивчають концентрацію прозапального цитокіну - фактора некрозу пухлин (ФНПα) у сироватці крові і, при рівні ФНПα 356 пг/мл та більше з вірогідністю 95,56±2,4 %, прогнозують розвиток тяжкого перебігу вітряної віспи.
Спосіб лікування вітряної віспи

Номер патенту: 10456
Опубліковано: 15.11.2005
Автори: Савчук Антоніна Іванівна, Гайдей Віктор Романович
МПК: A61K 38/21
Мітки: лікування, вітряної, спосіб, віспи
Формула / Реферат:
Спосіб лікування вітряної віспи шляхом впливу на імунний статус пацієнтів, який відрізняється тим, що застосовують віферон-4 у свічках ректально, по 1 свічці 2 рази на добу з 12-годинним інтервалом протягом 4-6 діб.
Попередній патент: Бурильна труба
Наступний патент: Конструктивний утеплювач
Випадковий патент: Кормова суміш для годування м`ясних каченят