Атенуйований штам вірусу захворювання свиней на репродуктивний та респіраторний синдромом (prrs), вакцина для захисту свиней від prrs, спосіб одержання активної вакцини проти prrs
Номер патенту: 68326
Опубліковано: 16.08.2004
Автори: Рієра Пуджадас Пере, Саубі Роса Нарцис, Еспуна Масо Енрік, Алемані Бурх Рейна




Формула / Реферат
1. Атенуйований штам вірусу захворювання свиней на репродуктивний та респіраторний синдром (PRRS), депонований 23.11.95 у CNCM Інституту Пастера під реєстраційним номером I-1642.
2. Вакцина для захисту свиней від захворювання на репродуктивний та респіраторний синдром (PRRS), яка відрізняється тим, що містить штам вірусу за п. 1 та/або принаймні один вірусний антиген, одержаний з даного штаму вірусу.
3. Вакцина за п. 2, яка відрізняється тим, що вірусний антиген являє собою інактивований вірус.
4. Вакцина за п. 2, яка відрізняється тим, що містить від 102 до 106 ТСID50 атенуйованого штаму вірусу за п. 1.
5. Вакцина за будь-яким з пп. 2 або 3, яка відрізняється тим, що вакцинна доза містить кількість вірусного антигену, еквівалентну кількості, що відповідає 102-106 ТСID50 атенуйованого штаму вірусу.
6. Вакцина за будь-яким з пп. 2-5, яка відрізняється тим, що додатково містить імуностимулюючі ад'юванти та/або емульгатори, та/або стабілізатори.
7. Вакцина за будь-яким з пп. 2-6, яка відрізняється тим, що додатково містить як живі, так і інактивовані віруси свиней, як окремо, так і в комбінації.
8. Вакцина за будь-яким з пп. 2-6, яка відрізняється тим, що додатково містить живі або інактивовані бактерії.
9. Спосіб одержання активної вакцини проти PRRS за п. 2, який відрізняється тим, що має такі ознаки:
- атенуювання вірулентного штаму PRRS шляхом його 15-200 пасажів у пермісивному клітинному клоні для одержання вакцини за п. 2,
- розмноження зазначеного штаму вірусу PRRS у пермісивному клітинному клоні до високих титрів, що становлять 104-107 ТСID/мл,
- пасаж зазначеного штаму PRRS у клітинному субстраті, що має період реплікації 20-175 годин при температурі 30-39°С.
10. Спосіб за п. 9, який відрізняється тим, що включає переведення одержаного атенуйованого штаму вірусу в форму водної дисперсії, масляної емульсії, ліпосомної композиції або ліофілізацію як у присутності, так і за відсутності ад'ювантів за п. 6.
11. Спосіб за будь-яким з пп. 9 і 10, який відрізняється тим, що здійснюють інактивацію, термальну або хімічну, одержаного атенуйованого штаму вірусу до його повної інактивації.
Текст
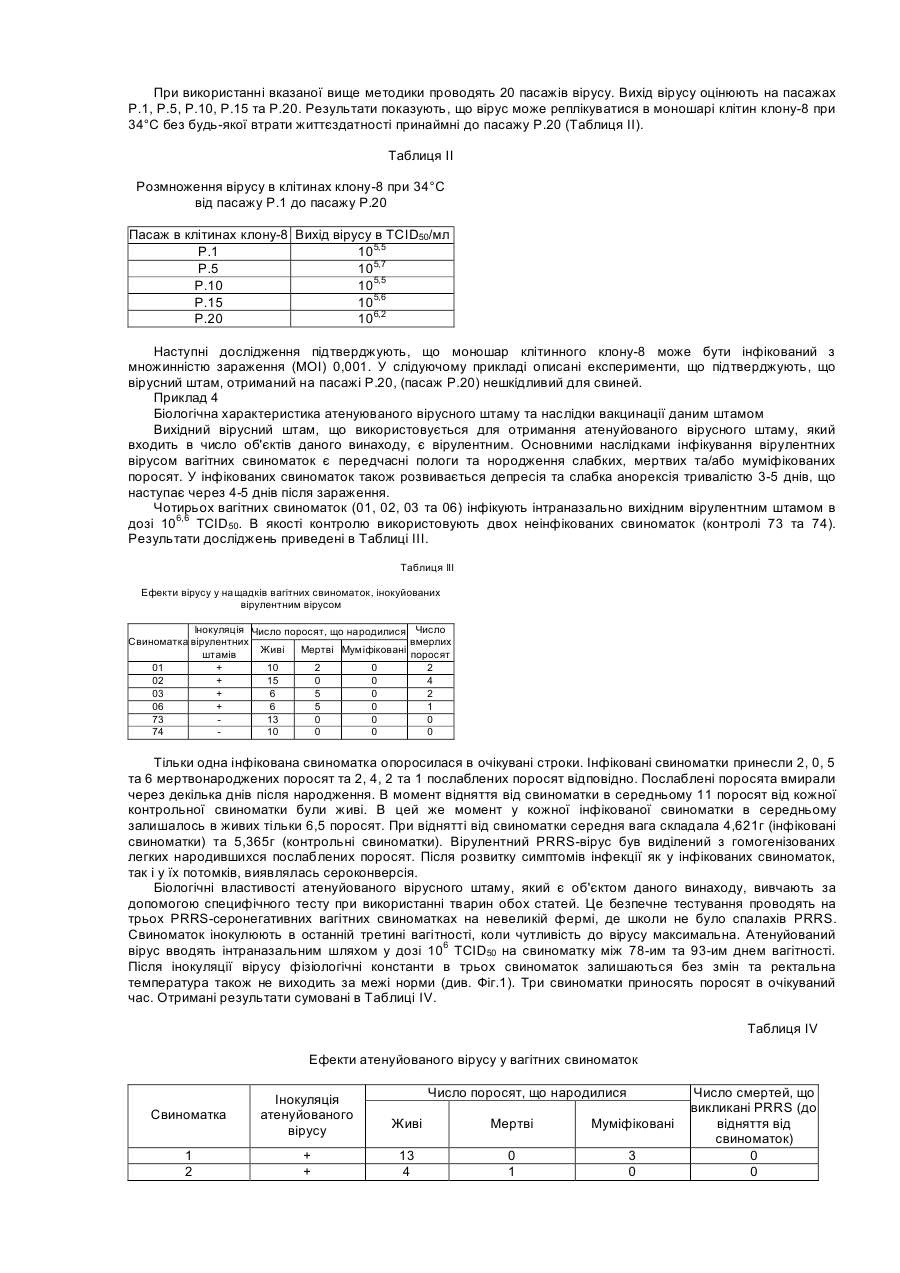
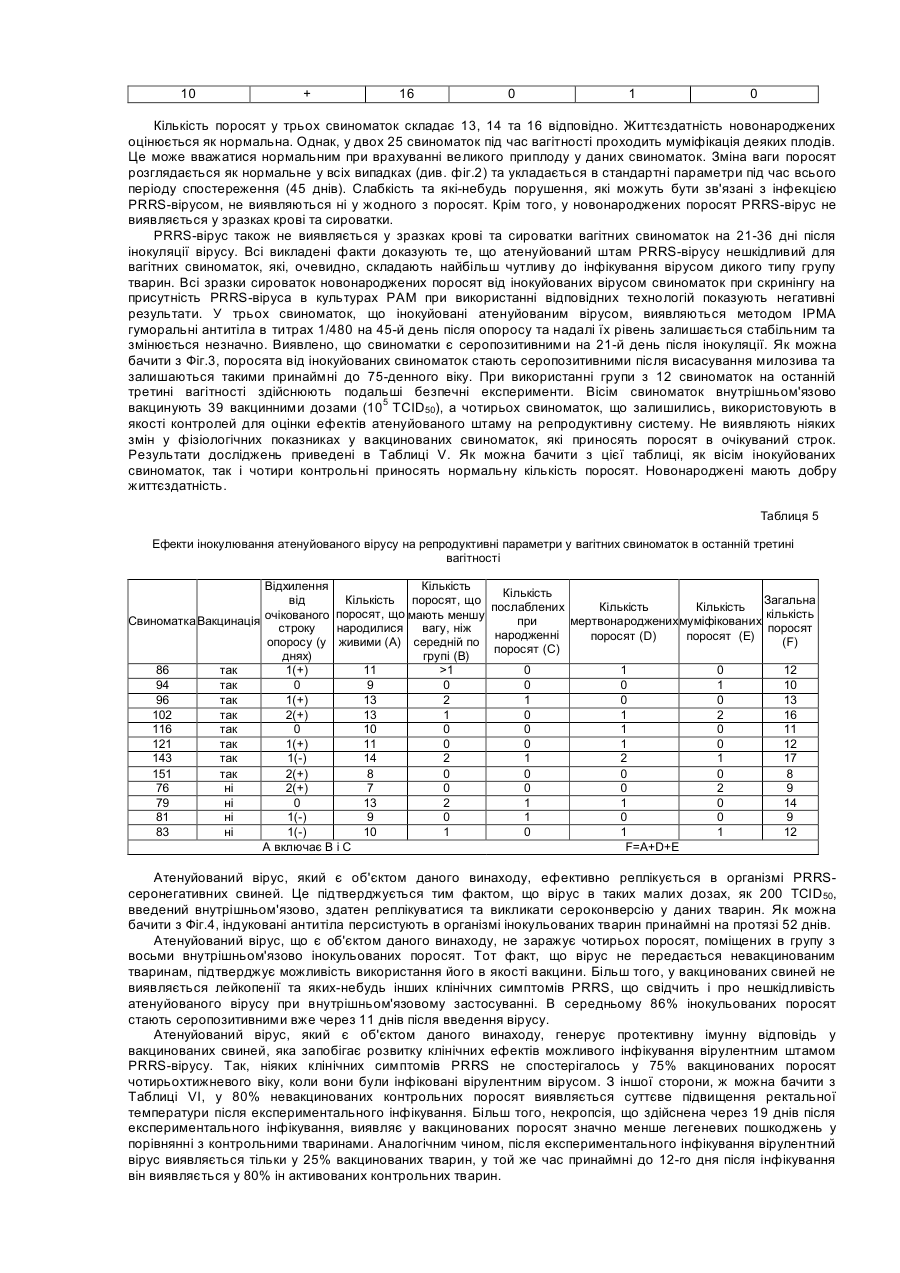
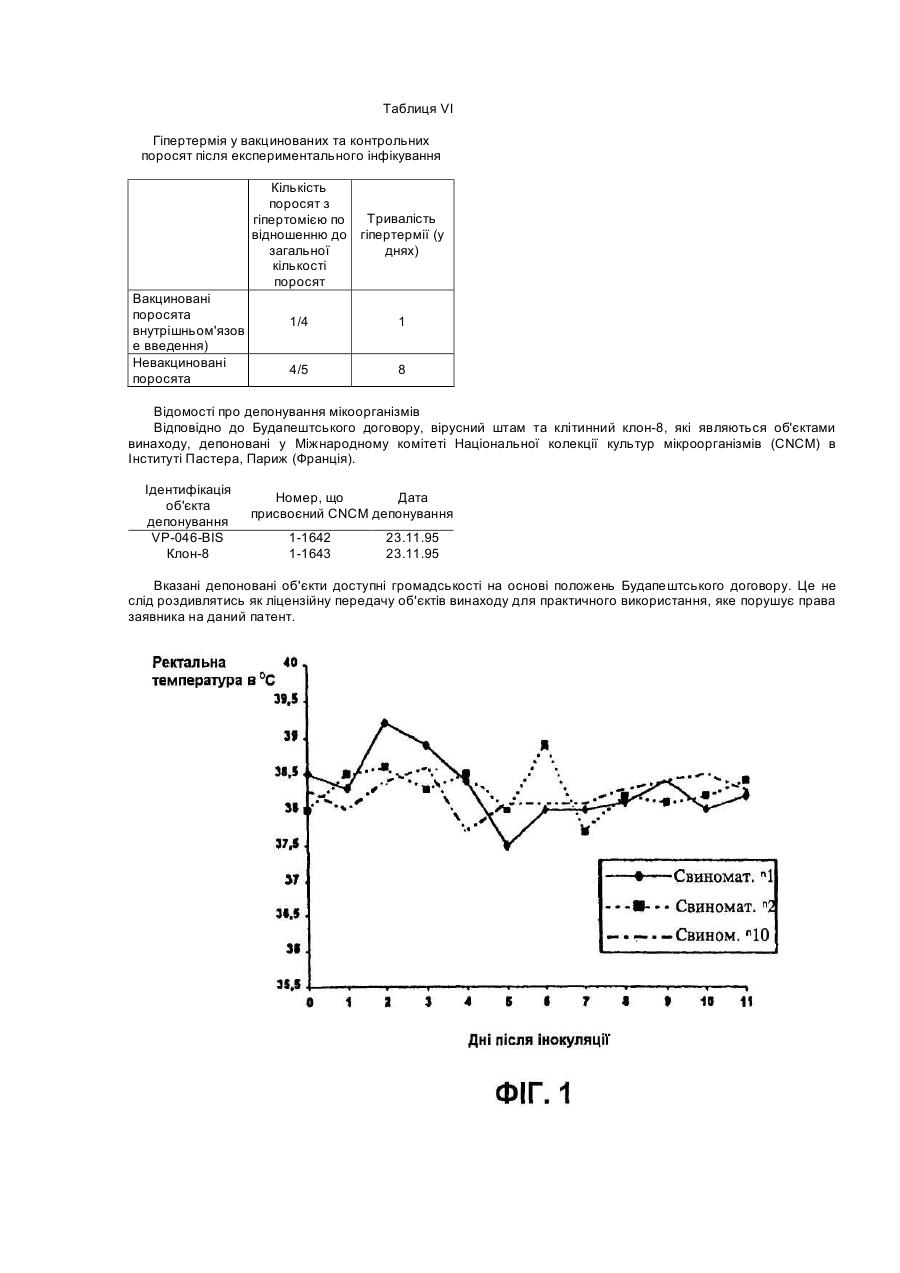
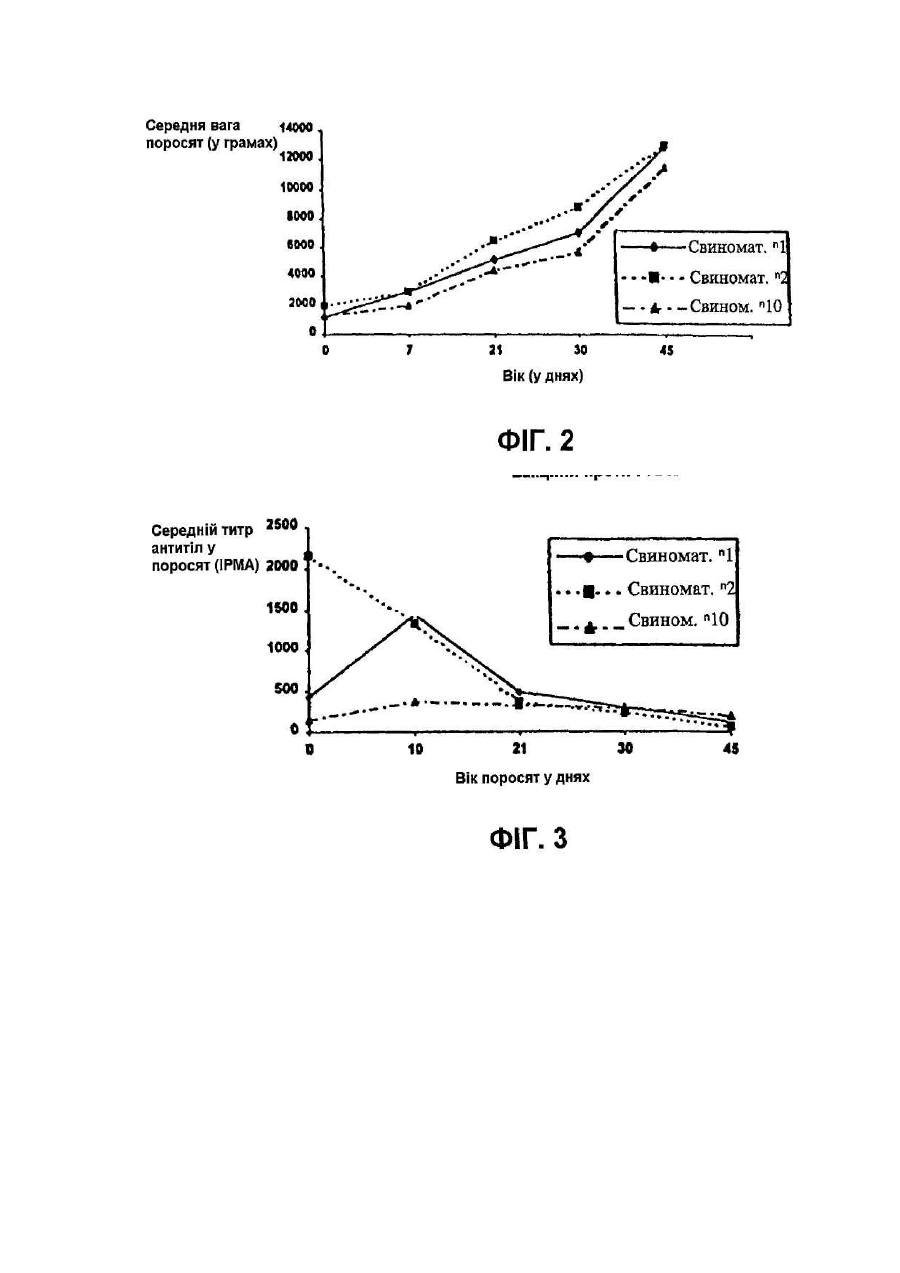
Винахід відноситься до нового атенуйованого штаму вірусу, що викликає хворобу свиней, яка відома як репродуктивний та респіраторний синдром свиней (PRRS). Спосіб атенуації та реплікації вірулентного штаму при використанні нового клітинного клону, отриманого з нирки мавпи, забезпечує створення вакцин та діагностичних наборів, за допомогою яких можливі рання діагностика PRRS та ефективне профілактичне лікування захворювання. У 1987 у Північній Африці була вперше виявлена хвороба свиней, яку назвали Таємною хворобою свиней, або ТХС, а пізніше - Синдромом, що пов'язаний з безпліддям та респіраторними порушеннями, або SIRS. Дуже схожий синдром був вперше виявлений у Центральній Європі у 1990p., та пізніше захворювання поширилось й в інші країни, включаючи Іспанію. У перший час у Європі хворобу називали Епідемічним абортивним та респіраторним синдромом, або PEARS, а у подальшому - репродуктивним та респіраторним синдромом свиней або PRRS. Ця назва вказаного захворювання отримала поширення у всьому світі. У даний час відомо, що етіологічним агентом PRRS є невеликий вірус, що містить інкапсульовану РНК, який вперше був виділений у Нідерландах та названий вірусом Лелістаду. Вважали, що даний вірус належить до групи Arterviridae. Він описаний у заявці РСТ WO 92/21375 та у Європейському патенті ЕР-В-058 7780 (Stickting Centraal Diegeneeskundig Instituut). Остання заявка надана на основі першої заявки РСТ. Для цілей вказаних заявок ізолят вірусу, що охарактеризований вище, депонований в Інституті Пастера у Парижі під номером 1-1102. Північноамериканський тип даного вірусу був ізольований практично одночасно з Європейським, як це описано у заявці РСТ WO 93/03760 (Collins et al.) та заявці на Європейський патент ЕР-А-052 9584 (Boehringer Ing.). Для цілей вказаних заявок ізолят вказаного вище вірусу депонований в Американській колекції типових культур (АТСС) під номером VR-2332. Європейський та Північноамериканський типи вірусів явним чином розрізняються не тільки по серологічній реактивності, але й по ступеню гомології нуклеотидних послідовностей істотних ДНК-фрагментів. На перших двох сторінках заявки на Європейський патент ЕР-А-0676467 (Akzo) приведений подрібний опис вказаних різниць з багаточисельними посиланнями. У заявці на Європейський патент зроблено заключения, що Європейський та Американський типи вірусів дивергували тривалий час тому. У відповідності з цим можна очікувати, що дійсно ефективні вакцини проти одного з вказаних типів вірусу будуть малоефективні або взагалі неефективні проти вірусу іншого типу. Різні штами були виділені з Європейського та Північноамериканського типів вірусу. Кожний штам має свої власні специфічні властивості, та деякі штами стали предметом заявок на видачу патенту. Наприклад, у заявці РСТ WO 93/07898 (Akzo) описується Європейський штам, депонований в CNCM (Інститут Пастера) під номером 1-1140, та отримані на його основі вакцини. У заявці РСТ WO 93/14196 та заявці на Європейський патент ЕР-А-541418 (Phone-Merieux), що мають однаковий пріоритет по даті подачі першої заявки, описується новий штам, який виділений у Франції та депонований в CNCM (Інститут Пастера) під номером 1-1153. У заявці на Європейський патент ЕР-А-0595436 (Solvay) охарактеризований новий штам Американського типу вірусу, більш вірулентний у порівнянні з будь-яким вище описаним, та отримані на його основі вакцини. Вказаний штам депоновано в АТСС, однак регістраційний номер депонованого об’єкту у заявці не вказано. Та, нарешті, в Іспанській заявці на патент ES-A-2074950 (Cyanamid lberica) описується так званий "Іспанський штам", який відрізняється від інших Європейських та Американських штамів. Вказаний "Іспанський штам" депонований у Європейській колекції культур клітин тварин (ЕАССС) під номером V93070108. У заключения відмітимо, що оскільки виявлені багаточисленні варіанти етіологічного агенту PRRS та існує необхідність підвищення ефективності боротьби з захворюванням, наполегливо треба розробляти вакцини, що активні по відношенню до різних типів вірусу, на основі конкретного вірусного штаму, який інфікує тварин. У свиноматок хвороба характеризується втратою апетиту, анорексією, репродуктивними порушеннями (викидні, передчасні пологи, народження мертвих або послаблених поросят, загибель плоду, у тому числі з його муміфікацією). Іноді інфіковані свиноматки гинуть. Більш рідкою ознакою захворювання є тимчасове посиніння вухів, живота або вульви. По цій причині хворобу у перший час називали у Нідерландах "Абортус блау", а у Великобританії - "Хворобою синього вуха". У поросят виявлення вказаних симптомів залежить від віку. У новонароджених можуть спостерігатися віддишка та тремор м'язів, у той час коли у більш старших поросят частіше зустрічаються параліч та атаксія задніх кінцівок. Під час спалаху захворювання смертність новонароджених перших днів життя обмежена, але досягає 80% у поросят десятиденного віку. На протязі деякого часу інфіковані свині, що відкормлюються, менше їдять та мають респіраторні порушення. Інкубаційний період захворювання у значному ступеню варіює, складаючи від 5 до 37 днів (I.B.Robertson Eurp.Comm.Seminar on PRRS, 11:4-5, Brussels, 1991). Іноді хвороба поширюється дуже повільно, але якщо заражена вся свиноферма, тоді захворювання може персистувати на протязі декількох місяців (B.Thacker, Int.Symp. on SIRS, St.Paul/Minnesota, 1992). За допомогою імунопероксидазної технололгії (імунопероксидазного дослідження моношару, що далі позначається ІРМА) на 6-й день після інфекції спостерігаються антивірусні антитіла, як описано Wensvoort G., et al. (The Vet.Quart. 13:121-130, 1991). Після 5 днів титри антитіл досягають 1/20000 та звичайно зберігаються такими на протязі 12 місяців та більше. Однак, як було повідомлено, V.Ohlinger et al. (Meredith, M.De.Pig Dis.Info. Center Cambridge, Dec. 1992) деякі тварини стають серонегативними через 4-5 місяців. Вказані автори змогли ізолювати вірус після інфікування з різних органів, причому титр вірусу досягав 104 ТСІD50 (50%-на доза інфікування тканинних культур) через 6 тижнів після початку інфекції у легенях, сироватці, плазмі та гомогенатах клітин крові. Це свідчить про те, що вірус та антитіла можуть сумісно персистувати на протязі декількох тижнів. Більш того, добре відомо, що свині, які виживають, при спалаху даного захворювання, здатні служити джерелом інфекції для сприйнятливих тварин. Віремія може бути виявлена, починаючи з першого дня після інфекції, та може продовжуватися до 56 днів, однак звичайно цей період більш короткий. При поширенні вірусу через кров він здатний інфікувати плаценту вагітних свиноматок. Показано, що вірус може проходити через плаценту та викликати загибель плоду. Максимальна сприйнятливість плоду до інфікування вірусом спостерігається під час останнього триместру вагітності. Окрім того, вірус здатний реплікуватися у клітинах плоду, не викликаючи його загибелі. Проте, вірус ніколи не був виділений з муміфікованих або з ембріонів, що підлягали аутолізису. У поросят хвороба починає розвиватися, коли рівень набутих з молозивом материнських антитіл зменшується. Серед поросят, які народилися живими у інфікованих у останньому триместрі вагітності свиноматок, спостерігаються тварини, які мають антитіла до вірусу ще до початку годування молозивом. Звичайно, при народженні у таких поросят також виявляється віремія (C.Terpstra et al., Vet.Q. 13:131-136. 1991). Не дивлячись на руйнування великої кількості макрофагів, імуносупресорна активність PRRS вірусу, що викликається, явним чином не показана. Однак, часто розвиваються асоційовані з PRRS вторинні інфекції, наносячи значні економічні втрати свинофермам. У даний час PRRS-вірус представляє собою одне з найважливіших захворювань свиней, що призводить до економічних втрат, як за рахунок безпосередньої, так й опосередкованої (вторинні інфекції, що пов’язані з PRRS) дії вірусу. Інактивовані вакцини, що отримані за допомогою культур альвеолярних макрофагів свині (РАМ), дають придатні результати на лабораторному рівні, однак їх ефективність у польових умовах частково залежить від дії зовнішнього середовища та вмісту вакцинованих тварин. Однією з причин, що заважають отриманню імунологічних продуктів, які активні по відношенню до PRRSвірусу, є обмежений доступ стабільних субстратів для реплікації вірусу. До недавнього часу PRRS-вірус міг бути ампліфікований тільки у культурах альвеолярних макрофагів свині (РАМ) (Wensvoort G. et al., in The Vet.Quart. 13:121-130, 1991). Необхідність використання здорових свиней певного віку для отримання вказаних макрофагальних культур має свої проблеми. Більш того, сприйнятливість до вірусної інфекції культурами РАМ не гарантована, оскільки клітинні субстрати, що отримані від різних тварин, завжди неоднорідні. Це створює серйозну перешкоду для отримання партій антигенів з постійними та гомологічним властивостями, та необхідно оцінювати показники кожної партії. По всім вищевказаним причинам низька доступність стабільних та постійних клітин-хазяїв серйозним чином ускладнює розробку стратегій отримання вірусних мутантів, що основана на реплікації PRRS-вірусу у клітинних субстратах та селекції атенуйованих варіантів. Одна з основних проблем, яку необхідно вирішити, це нейтралізація вірулентних вірусів, що уникнули реплікації у культурах РАМ, оскільки реплікація призводить до деструкції вірусів. У відповідності з вказаним, адаптація вірусу до стабільного субстрату, що походить з трансформованої клітинної лінії, зможе забезпечити належний підхід до отримання атенуйованих мутантів та інактивованих вакцин, таким чином знімаючи залежність від РАМ та неоднорідності даних культур. У заявці РСТ WO 94/18311 (Miles) описується розмноження деяких PRRS-вірусних штамів в унікальному клітинному клоні, що позначений як клон 900 9В, який був отриманий з клітинної лінії нирки африканської зеленої мавпи, що має дане зяавником позначення МА-104 (М). Вказана лінія у свою чергу отримана з комерційної клітинної лінії МА-104. У даній патентній заявці не приведені дані про депонування вказаного унікального клону та тому здійснення та відтворення відповідного винаходу ускладнено. Винахід відноситься до нового атенуйованого штаму PRRS-вірусу, за допомогою якого стабільним та відтвореним чином можуть бути отримані нешкідливі для свиней вакцини, що попереджують розвиток PRRS з високим ступенем ефективності. Іншим об'єктом даного винаходу є новий клітинний клон, що отриманий на основі стабільної клітинної ліній нирки мавпи МА-104, який здатний підтримувати розмноження PRRS-вірусу у високих титрах та забезпечувати постійний вихід вірусу відібраного атенуйованого штаму, та спосіб отримання вказаного клітинного клону. Винахід також відноситься до ефективних вакцин проти PRRS, які можуть бути розроблені на основі нового атенуйованого штаму та його мутантів, та способу отримання вказаних вакцин. Ще одним об'єктом винаходу є набори для діагностики PRRS на основі нового атенуйованого штаму та його мутантів, та до способу отримання вказаних наборів. Атенуйований штам PRRS-вірусу, який є об'єктом даного винаходу, отриманий в результаті атенуації вірулентного штаму, що виділений з інфікованих свиней на іспанській фермі. У відповідності з вимогами до опису винаходу, вказаний атенуйований штам депонований у Національній колекції культур мікроорганізмів (CNCM) в Інституті Пастера під регістраційним номером І-1642. Дата депонування - 23.11.95. Атенуація та реплікація атенуйованого вірулентного штаму здійснюється шляхом серійних пасажів у клітинах певного клону, який також є об'єктом даного винаходу, та позначений авторами як клон-8. Вказаний клон отриманий з комерційної лінії нирки мавпи, яка відома під позначенням МА-104. Також у відповідності з вимогами до опису винаходу, вироблено депонування вказаного клону-8 у CNCM в Інституті Пастера під регістраційним номером І-1643. Дата депонування аналогічна вказаної вище. В першу чергу комерційну лінію МА-104 клонують для селекції та виділення клону-8. Клонування забезпечується суспендуванням клітин у ростовому середовищі, що підходить (наприклад, мінімального необхідного середовища Ігла (MEM Ігла)) з додаванням ембріональної бичачої сироватки (FBS), висіванням клітинної суспензії на чашки у різних концентраціях, відбором та трипсинізацією клонів та розмноженням їх у культуральних флаконах. Отримані таким чином клони успішно субклонують, як описано вище, до тих пір, поки не будуть виділені добре диференційовані клони. Після видалення клонів, які виявляють нестійкі властивості або важко ампліфікуються, відібрані клони тестують на сприйнятливість до інфекції вихідним вірулентним штамом PRRS-вірусу. Клон-8 відбирають на основі його високої чутливості до реплікації вірусного штаму (високе значення ТСІD50), репродуктивності та однорідності отриманого вірусного матеріалу. Вірулентний штам атенуюють послідовною реплікацією у культурах клону-8, переважно при 34°С. Вміст інфекційних вірусних частинок досліджують для того, щоб оцінити реплікаційну активність, та тестують цитопатичну дію (СРЕ) вірусу з метою визначення ступеню адаптації. У відповідності з результатами досліджень, вірус здатний реплікуватися у клітинах 8 без втрати життєздатності принаймні на протязі 20 пасажів, та така атенуація забезпечує отримання практично нешкідливого вірусу, що зберігає антигенність. У відповідності з вказаним, атенуйований штам PRRS-вірусу, який є об'єктом даного винаходу, отримують стабільним та промислово відтворюваним чином, що забезпечує його використання для приготування як PRRS-вакцин, так й діагностичних наборів. Порівняльні дослідження між двома групами свиней, інфікованих вірулентним або атенуйованим штамом, явним чином показують, що атенуйований штам нешкідливий для тварин. З іншої сторони, атенуйований штам, який є об'єктом даного винаходу,демонструє високу ефективність реплікації у серонегативних свиней, та здатний викликати сероконверсію у тварин, інокульованих внутрішньом'язовим шляхом у таких низьких дозах, як 200 TCID50. Утворені відповідним чином антитіла персистують у крові принаймні 80 днів. Таким чином, атенуйований штам, що є об'єктом даного винаходу, служить високоефективною базою для отримання вакцин, що забезпечують профілактичний захист свиней від PRRS. Такі вакцини можуть бути отримані за допомогою якого-небудь добре відомого з рівня техніки методу у різних формах, таких як водні дисперсійні розчини, масляні емульсії, ліпосомні композиції, ліофілізовані препарати та т.п. Вакцинні композиції можуть бути доповнені різними ад'ювантами, наприклад, імуностимуляторами, емульгаторами, стабілізаторами та т.п. Вакцини можуть бути введені внутрішньом'язовим, підшкірним, інтраназальним, інтратрахеальним, шкірним, черезшкірним або внутрішньошкірним шляхом. Ефективні вакцинні дози можуть варіювати, однак переважно містять від 102 до 106ТСІD50 атенуйованого штаму, який є об'єктом даного винаходу. Заявлені вакцини можуть бути представлені полівалентними вакцинами, що містять інший живий або інактивований вірус свиней, або живі або інактивовані бактерії. Як очевидно спеціалісту у даній області досліджень, можуть також бути створені вакцини, що містять вірусні антигени, отримані з вірусного штаму, який є об'єктом даного винаходу. Прикладом таких вакцин можуть служити вакцини, що містять вказаний штам, повністю інактивований яким-небудь підходящим способом, може термічним або хімічним шляхом, вакцини, що містять капсулярні структури або фрагменти ДНК вказаного штаму, та т.п. Атенуйований штам, який є об'єктом даного винаходу, може бути також використаний для отримання (за допомогою відповідних методик) підходящих діагностичних наборів, що містять антигенні структури, які здатні виявляти антитіла у серопозитивних тварин. Наприклад, методика ІРМА для виявлення PRRS-антитіл може включати наступні стадії: а) адаптацію атенуйованого вірусу, який є об'єктом винаходу, до стабільної клітинної культури, переважно клону-8, у мікропланшетах для культивування таким чином, що кожна лунка інфікована приблизно 20-40 інфекційними частинками; б) закріплення інфікованих клітин на твердій фазі за допомогою відомих фіксаторів; в) виявлення антитіл у сироватці крові свиней шляхом інкубування їх у мікропланшетах з послідуючим забарвленням вказаних антитіл у мікропланшетах, відповідно методиці ІРМА. Перелік фігур креслень: До даного винаходу додаються дві сторінки з чотирма кресленнями, які є частиною опису. Креслення представлені з ілюстративною метою, не обмежують винахід та відображають слідуюче. Фіг.1. Приведений графік, що побудований у двомірній системі координат, який відображає хід ректальної температури вагітних свиноматок, яким інтраназально інокулювали атенуйований вірусний штам, що є об'єктом даного винаходу. Фіг.2. Приведений графік, що побудований у двомірній системі координат, який відображає зміну ваги поросят, що народились у свиноматок, яким інокулювали атенуйований вірусним штам, що є об'єктом даного винаходу. Фіг.3. Приведений графік, що побудований у двомірній системі координат, який відображає кінетику антитіл молозива, як це визначено за допомогою ІРМА, у нащадків свиноматок, які вакциновані атенуйованим вірусним штамом, що є об'єктом даного винаходу. Знижені титри у нащадків свиноматок 1 та 10 при народженні зумовлені тією обставиною, що деякі поросята ще не встигли відсмоктувати молозиво до моменту забору крові. Фіг.4. Приведена трьохмірна діаграма, що демонструє розвиток гуморальної антитільної відповіді у поросят, внутрішньом'язово інокулюваних 200, 2000 та 20000 ТСІD50 атенуйованого вірусного штаму, який є об'єктом даного винаходу. Приклади Нижче наведені приклади з метою більш детальної ілюстрації опису до даного винаходу. Вказані приклади не слід розглядати, як ті, що обмежують об'єм винаходу. Приклад 1. Отримання клітинних клонів на основі клітинної лінії нирки мавпи МА-104 Здійснюють шість пасажів отриманого у результаті селекції вірулентного штаму PRRS-вірусу у стабільній клітинній лінії нирки мавпи МА-104, яка зберігається у Європейській колекції культур клітин тварин (ЕАССС) під регістраційним номером 85102918. При цьому використовують статичні клітинні культури з утвореним моношаром, що вирощені у культуральних флаконах у середовищі MEM Ігла з додаванням 10% ембріональної бичачої сироватки (FBS) при 37°С та без внесення СО2. Вірус збирають на 6-7 день після інокуляції на кожному пасажі. Для визначення виходу вірусу клітинну лінію інокулюють вірулентним штамом з різною багаточисельністю зараження (далі - МОІ). Вихід вірусу навіть після чотирьох адаптаційних пасажів виявляється досить низьким (між 102 та 104 ТСІD50/мл) та недостатнім для отримання вакцини. У вказаних експериментах виявляється, що тільки частина інфікованих клітин чутлива до вірулентного вірусу, у той час як клітини, що залишились, зберігають стікість до інфікування. По цій причині вказану клітинну лінію клонують з метою відбору таких клітинних популяцій, які у цілому мають загальну сприйнятливість до вірулентного штаму. Клональну селекцію здійснюють слідуючим чином. Суспензію клітин лінії розводять в MEM Ігла з 20% FBS та різні розведення поміщають у 96-лункові мікропланшети (NUNC). Планшети інкубують на протязі 8 днів при 37°С та 5% СО2. На третій день відбирають під мікроскопом лунки, що містять тільки одну клітину, та вміст таких лунок на 8-й день оброблюють трипсином. Таким чином отримують 44 клона. Потім вказані клони розмножують у культуральних флаконах до отримання суспензії, що містить 5x107 клітин/мл. Клітини послідовно клонують за допомогою аналогічної процедури другий або третій раз. Після третього клонування 44 вказаних клона розмножують до отримання 25мл клітинної суспензії кожного клону, що містить 6x106 клітин/мл. Середовище, яке використовують при здійсненні даних процедур, містить 20% FBS. Отримані клони далі відбирають у відповідності з їх ростовими властивостями та показниками життєздатності. Приклад 2 Селекція клітинного клону-8 Клони, що отримані як це описано у попередньому прикладі, оцінюють на здатність до ефективного росту. Відкидають клони, які важко ампліфікувати та підтримувати при 37°С, та клітини, які мають порушену морфологію. У цілому, після попередньої селекції відкидають 35 клонів та відбирають 9 клонів, що залишились. Відібрані клітинні клони інфікують суспензією вірулентного штаму PRRS-вірусу на восьмому пасажі (Р-8), отриманого у культурах РАМ (як це описано Bloomberg, m. et al., Vet.Micribio. 42, 361-371, 1994). Процес адаптації до клітинних клонів проходить слідуючим чином: вірус реплікується при 37°С на протязі 3 пасажів при підтриманні 80-90% злитого клітинного моношару, який контактує з вірусною суспензією на протязі 6 днів, потім вірус заморожують при -80°С та відтаюють через 24 години. В якості середовища інфікування використовують MEM Ігла з додаванням 10% FBS та гентаміцину (0,4мг/мл). Не застосовують антигрибкових та антидріжджевих сполук. Кожний збір вірусу, отриманий вказаним шляхом, титрують при використанні відповідного клітинного клону. Після аналізу результатів роблять заключення про те, що 9 клонів сприйнятливі до вірусної інфекції, причому титр вірусу складає 104,2 ТСІD50/мл або перевищує цю величину. Результати досліджень приведені в Таблиці 1. Таблиця 1 Чутливість отриманих клітинних клонів до відібраного вірулентного штаму Клон або клітинна лінія Неклонована клітинна лінія Клон 1 Клон 2 Клон 3 Клон 4 Клон 8 Клон 29 Клон 30 Клон 41 Клон 44 Титр як величина ТСІD50/мл 103 105,75 104,5 105,2 105,3 106 105,75 105,38 105,38 104,2 Як можна бачити з даних Таблиці 1, неклонована клітинна лінія виявляє дуже низьку чутливість до вірусу, та таким чином, застосування її в якості антигену при отриманні ефективних вакцин виявляється неможливим. Дані Таблиці 1 й показують, що деякі клони, особливо клон-8, більш чутливий до вірусу, ніж неклонована клітинна лінія. Клон-8 особливо виділяється серед інших. При його використанні вірус може бути отриманий в титрах, що досягають 106 ТСІD50/мл. У відповідності з результатами, що приведені вище, відбирають клон-8. Проводять певні дослідження, щоб бути певним в надійності властивостей вказаного клону. В різних дослідженнях титри зібраного вірусу відтворюються з високою точністю та варіюють від 105 до 107 ТСІD50/мл. Приклад 3 Отримання атенуйованого вірусного штаму Атенуйований вірусний штам, який являється об'єктом даного винаходу, отримують шляхом реплікації зворотнього вірулентного штаму в клітинах клону-8 при 34°С. Клітини клону-8 у стані моношару, що інфіковані вірусом, підтримують при 34°С до проявлення СРЕ. Цитопатичний ефект завжди виявляється між 24 та 48 годинами після інфікування, пізніше, ніж в культурах, які підтримуються при 37°С. Однак, ніяких суттєвих різниць в СРЕ при обох вказаних температурах не відмічається. Зібраний вірус титрують при використанні клітинного моношару клону 8. Крім того, вірус ідентифікують за допомогою ІРМА. Вірусом інокулюють клітини клону-8, які практично досягли стану моношару (75см2), та залишають для адсорбції на 2 години при 34°С. Потім до моношару додають середовище для інфікування, яка являє собою мінімальне необхідне середовище, модифіковану Іглом, з додаванням 10% FBS та попередньо прогріту при 34°С. Флакони з площею клітинного моношару розмішують в термостат при 34°С та кожен день продивляються до чіткого прояву СРЕ. Коли СРЕ зачіпає 80-95% клітинного моношару, що звичайно проходить між п'ятим та сьомим днем після інфікування, збирають вірус. Вірусну фракцію центрифугують при 2000 rpm та потім титрують супернатант для визначення титру вірусу (ТСІD50/мл). При використанні вказаної вище методики проводять 20 пасажів вірусу. Вихід вірусу оцінюють на пасажах Р.1, Р.5, Р.10, Р.15 та Р.20. Результати показують, що вірус може реплікуватися в моношарі клітин клону-8 при 34°С без будь-якої втрати життєздатності принаймні до пасажу Р.20 (Таблиця II). Таблиця II Розмноження вірусу в клітинах клону-8 при 34°С від пасажу Р.1 до пасажу Р.20 Пасаж в клітинах клону-8 Вихід вірусу в ТСІD50/мл Р.1 105,5 Р.5 105,7 Р.10 105,5 Р.15 105,6 Р.20 106,2 Наступні дослідження підтверджують, що моношар клітинного клону-8 може бути інфікований з множинністю зараження (МОI) 0,001. У слідуючому прикладі описані експерименти, що підтверджують, що вірусний штам, отриманий на пасажі Р.20, (пасаж Р.20) нешкідливий для свиней. Приклад 4 Біологічна характеристика атенуюваного вірусного штаму та наслідки вакцинації даним штамом Вихідний вірусний штам, що використовується для отримання атенуйованого вірусного штаму, який входить в число об'єктів даного винаходу, є вірулентним. Основними наслідками інфікування вірулентних вірусом вагітних свиноматок є передчасні пологи та нородження слабких, мертвих та/або муміфікованих поросят. У інфікованих свиноматок також розвивається депресія та слабка анорексія тривалістю 3-5 днів, що наступає через 4-5 днів після зараження. Чотирьох вагітних свиноматок (01, 02, 03 та 06) інфікують інтраназально вихідним вірулентним штамом в дозі 106,6 ТСІD50. В якості контролю використовують двох неінфікованих свиноматок (контролі 73 та 74). Результати досліджень приведені в Таблиці III. Таблиця III Ефекти вірусу у нащадків вагітних свиноматок, інокуйованих вірулентним вірусом Інокуляція Число поросят, що народилися Число Свиноматка вірулентних вмерлих Живі Мертві Муміфіковані поросят штамів 01 + 10 2 0 2 02 + 15 0 0 4 03 + 6 5 0 2 06 + 6 5 0 1 73 13 0 0 0 74 10 0 0 0 Тільки одна інфікована свиноматка опоросилася в очікувані строки. Інфіковані свиноматки принесли 2, 0, 5 та 6 мертвонароджених поросят та 2, 4, 2 та 1 послаблених поросят відповідно. Послаблені поросята вмирали через декілька днів після народження. В момент відняття від свиноматки в середньому 11 поросят від кожної контрольної свиноматки були живі. В цей же момент у кожної інфікованої свиноматки в середньому залишалось в живих тільки 6,5 поросят. При віднятті від свиноматки середня вага складала 4,621г (інфіковані свиноматки) та 5,365г (контрольні свиноматки). Вірулентний PRRS-вірус був виділений з гомогенізованих легких народившихся послаблених поросят. Після розвитку симптомів інфекції як у інфікованих свиноматок, так і у їх потомків, виявлялась сероконверсія. Біологічні властивості атенуйованого вірусного штаму, який є об'єктом даного винаходу, вивчають за допомогою специфічного тесту при використанні тварин обох статей. Це безпечне тестування проводять на трьох PRRS-серонегативних вагітних свиноматках на невеликій фермі, де школи не було спалахів PRRS. Свиноматок інокулюють в останній третині вагітності, коли чутливість до вірусу максимальна. Атенуйований вірус вводять інтраназальним шляхом у дозі 106 ТСІD50 на свиноматку між 78-им та 93-им днем вагітності. Після інокуляції вірусу фізіологічні константи в трьох свиноматок залишаються без змін та ректальна температура також не виходить за межі норми (див. Фіг.1). Три свиноматки приносять поросят в очікуваний час. Отримані результати сумовані в Таблиці IV. Таблиця IV Ефекти атенуйованого вірусу у вагітних свиноматок Свиноматка Інокуляція атенуйованого вірусу 1 2 + + Число поросят, що народилися Живі Мертві Муміфіковані 13 4 0 1 3 0 Число смертей, що викликані PRRS (до відняття від свиноматок) 0 0 10 + 16 0 1 0 Кількість поросят у трьох свиноматок складає 13, 14 та 16 відповідно. Життєздатність новонароджених оцінюється як нормальна. Однак, у двох 25 свиноматок під час вагітності проходить муміфікація деяких плодів. Це може вважатися нормальним при врахуванні великого приплоду у даних свиноматок. Зміна ваги поросят розглядається як нормальне у всіх випадках (див. фіг.2) та укладається в стандартні параметри під час всього періоду спостереження (45 днів). Слабкість та які-небудь порушення, які можуть бути зв'язані з інфекцією PRRS-вірусом, не виявляються ні у жодного з поросят. Крім того, у новонароджених поросят PRRS-вірус не виявляється у зразках крові та сироватки. PRRS-вірус також не виявляється у зразках крові та сироватки вагітних свиноматок на 21-36 дні після інокуляції вірусу. Всі викладені факти доказують те, що атенуйований штам PRRS-вірусу нешкідливий для вагітних свиноматок, які, очевидно, складають найбільш чутливу до інфікування вірусом дикого типу групу тварин. Всі зразки сироваток новонароджених поросят від інокуйованих вірусом свиноматок при скринінгу на присутність PRRS-віруса в культурах РАМ при використанні відповідних технологій показують негативні результати. У трьох свиноматок, що інокуйовані атенуйованим вірусом, виявляються методом ІРМА гуморальні антитіла в титрах 1/480 на 45-й день після опоросу та надалі їх рівень залишається стабільним та змінюється незначно. Виявлено, що свиноматки є серопозитивними на 21-й день після інокуляції. Як можна бачити з Фіг.3, поросята від інокуйованих свиноматок стають серопозитивними після висасування милозива та залишаються такими принаймні до 75-денного віку. При використанні групи з 12 свиноматок на останній третині вагітності здійснюють подальші безпечні експерименти. Вісім свиноматок внутрішньом'язово вакцинують 39 вакцинними дозами (105 ТСІD50), а чотирьох свиноматок, що залишились, використовують в якості контролей для оцінки ефектів атенуйованого штаму на репродуктивну систему. Не виявляють ніяких змін у фізіологічних показниках у вакцинованих свиноматок, які приносять поросят в очікуваний строк. Результати досліджень приведені в Таблиці V. Як можна бачити з цієї таблиці, як вісім інокуйованих свиноматок, так і чотири контрольні приносять нормальну кількість поросят. Новонароджені мають добру життєздатність. Таблиця 5 Ефекти інокулювання атенуйованого вірусу на репродуктивні параметри у вагітних свиноматок в останній третині вагітності Відхилення Кількість Кількість Кількість поросят, що від Загальна послаблених Кількість Кількість кількість очікованого поросят, що мають меншу Свиноматка Вакцинація при мертвонароджених муміфікованих строку народилися вагу, ніж поросят народженні поросят (D) поросят (Е) опоросу (у живими (А) середній по (F) поросят (С) днях) групі (В) 86 так 1(+) 11 >1 0 1 0 12 94 так 0 9 0 0 0 1 10 96 так 1(+) 13 2 1 0 0 13 102 так 2(+) 13 1 0 1 2 16 116 так 0 10 0 0 1 0 11 121 так 1(+) 11 0 0 1 0 12 143 так 1(-) 14 2 1 2 1 17 151 так 2(+) 8 0 0 0 0 8 76 ні 2(+) 7 0 0 0 2 9 79 ні 0 13 2 1 1 0 14 81 ні 1(-) 9 0 1 0 0 9 83 ні 1(-) 10 1 0 1 1 12 А включає В і С F=A+D+E Атенуйований вірус, який є об'єктом даного винаходу, ефективно реплікується в організмі PRRSсеронегативних свиней. Це підтверджується тим фактом, що вірус в таких малих дозах, як 200 ТСІD50, введений внутрішньом'язово, здатен реплікуватися та викликати сероконверсію у даних тварин. Як можна бачити з Фіг.4, індуковані антитіла персистують в організмі інокульованих тварин принаймні на протязі 52 днів. Атенуйований вірус, що є об'єктом даного винаходу, не заражує чотирьох поросят, поміщених в групу з восьми внутрішньом'язово інокульованих поросят. Тот факт, що вірус не передається невакцинованим тваринам, підтверджує можливість використання його в якості вакцини. Більш того, у вакцинованих свиней не виявляється лейкопенії та яких-небудь інших клінічних симптомів PRRS, що свідчить і про нешкідливість атенуйованого вірусу при внутрішньом'язовому застосуванні. В середньому 86% інокульованих поросят стають серопозитивними вже через 11 днів після введення вірусу. Атенуйований вірус, який є об'єктом даного винаходу, генерує протективну імунну відповідь у вакцинованих свиней, яка запобігає розвитку клінічних ефектів можливого інфікування вірулентним штамом PRRS-вірусу. Так, ніяких клінічних симптомів PRRS не спостерігалось у 75% вакцинованих поросят чотирьохтижневого віку, коли вони були інфіковані вірулентним вірусом. З іншої сторони, ж можна бачити з Таблиці VI, у 80% невакцинованих контрольних поросят виявляється суттєве підвищення ректальної температури після експериментального інфікування. Більш того, некропсія, що здійснена через 19 днів після експериментального інфікування, виявляє у вакцинованих поросят значно менше легеневих пошкоджень у порівнянні з контрольними тваринами. Аналогічним чином, після експериментального інфікування вірулентний вірус виявляється тільки у 25% вакцинованих тварин, у той же час принаймні до 12-го дня після інфікування він виявляється у 80% ін активованих контрольних тварин. Таблиця VI Гіпертермія у вакцинованих та контрольних поросят після експериментального інфікування Кількість поросят з Тривалість гіпертомією по відношенню до гіпертермії (у загальної днях) кількості поросят Вакциновані поросята внутрішньом'язов е введення) Невакциновані поросята 1/4 1 4/5 8 Відомості про депонування мікоорганізмів Відповідно до Будапештського договору, вірусний штам та клітинний клон-8, які являються об'єктами винаходу, депоновані у Міжнародному комітеті Національної колекції культур мікроорганізмів (CNCM) в Інституті Пастера, Париж (Франція). Ідентифікація об'єкта депонування VP-046-BIS Клон-8 Дата Номер, що присвоєний CNCM депонування 1-1642 1-1643 23.11.95 23.11.95 Вказані депоновані об'єкти доступні громадськості на основі положень Будапештського договору. Це не слід роздивлятись як ліцензійну передачу об'єктів винаходу для практичного використання, яке порушує права заявника на даний патент.
ДивитисяДодаткова інформація
Назва патенту англійськоюAn attenuated strain of the virus causing the porcine reproductive and respiratory syndrom (prrs), a vaccine for porcine protection from prrs, a method for preparation of active vaccine against prrs
Назва патенту російськоюОслабленный штамм вируса заболевания свиней репродуктивным и респираторным синдромом (prrs), вакцина для защиты свиней от prrs, способ получения активной вакцины против prrs
МПК / Мітки
МПК: A61K 39/295, C12R 1/93, A61P 31/14, C12N 7/08, A61K 39/12
Мітки: вакцина, репродуктивний, свиней, штам, респіраторний, захворювання, prrs, активної, спосіб, вакцини, захисту, синдромом, атенуйований, одержання, вірусу
Код посилання
<a href="https://ua.patents.su/9-68326-atenujjovanijj-shtam-virusu-zakhvoryuvannya-svinejj-na-reproduktivnijj-ta-respiratornijj-sindromom-prrs-vakcina-dlya-zakhistu-svinejj-vid-prrs-sposib-oderzhannya-aktivno-vakcini-pr.html" target="_blank" rel="follow" title="База патентів України">Атенуйований штам вірусу захворювання свиней на репродуктивний та респіраторний синдромом (prrs), вакцина для захисту свиней від prrs, спосіб одержання активної вакцини проти prrs</a>
Вакцини проти живого вірусу prrs з низькою патогенністю і способи їх одержання

Номер патенту: 47433
Опубліковано: 15.07.2002
Автор: Дзоо Хан Соо
МПК: A61K 39/12, C12N 7/04, A61P 31/12, C12N 7/00, C12N 15/38, A61K 39/155
Мітки: вакцини, живого, способи, одержання, патогенністю, вірусу, низькою
Формула / Реферат:
1. Вакцина, яка вміщує вірус Північно-американського свинячого репродуктивного та респіраторного синдрому і який має позначення АТСС VR2509.2. Вакцина згідно з п. 1, яка відрізняється тим, що вірус є живий.3. Вакцина згідно з п. 1, яка відрізняється тим, що доза вакцини містить від 104 до 108 БУО.4. Спосіб імунізації свині проти Північно-американського свинячого репродуктивного та респіраторного синдрому, який включає...
Штам вірусу герпесу курей sbg другого серотипу як компонент для виготовлення бівалентної вакцини проти хвороби марека

Номер патенту: 53969
Опубліковано: 17.02.2003
Автори: Соловйов Сергій Тихонович, Білокінь Віктор Степанович, Берус Павел Тихонович, Фісенко Светлана Анатоліївна, Герман В'ячеслав Валентинович, Стегній Борис Тимофійович
МПК: A61K 39/295, A61K 39/255, C12N 7/00, C12R 1/93, A61K 39/245
Мітки: марека, компонент, хвороби, курей, вакцини, штам, виготовлення, герпесу, серотипу, бівалентної, вірусу, другого
Формула / Реферат:
Штам вірусу герпесу курей SBG другого серотипу як компонент для виготовлення бівалентної вакцини проти хвороби Марека, який депонований та зберігається за номером 7 у колекції вірусів лабораторії біотехнології та біохімії Інституту експериментальної та клінічної ветеринарної медицини УААН, м. Харків, Україна.
Штам вірусу інфекційного ринотрахеїту ipt-lg /іеквм/ для виготовлення вакцини

Номер патенту: 30385
Опубліковано: 15.11.2000
Автори: Волосянко Олена Вікторівна, Морозова Олена Михайлівна, Фукс Поліна Павлівна
МПК: A61K 39/265, C12N 7/00, C12R 1/93
Мітки: ipt-lg, штам, виготовлення, ринотрахеїту, вірусу, інфекційного, іеквм, вакцини
Формула / Реферат:
Штам вірусу інфекційного ринотрахеїту ІРТ-LG (ІЕКВМ) для виготовлення вакцини проти інфекційного ринотрахеїту великої рогатої худоби.
Штам вірусу синдрому зниження несучості-1976 для виготовлення вакцини

Номер патенту: 45400
Опубліковано: 15.04.2002
Автори: Герман В'ячеслав Валентинович, Герман Євгеній В'ячеславович, Бабкін Валерій Федорович, Герман Ірина В'ячеславна, Ольховик Лідія Андрійовна
МПК: C12N 5/06, C12R 1/93, C12R 1/91, C12N 7/00
Мітки: синдрому, вакцини, вірусу, штам, виготовлення, зниження, несучості-1976
Формула / Реферат:
Штам вірусу синдрому зниження несучості-197б ЄГ-96 №3 (колекція ІЕКВМ) сімейство Adenoviridae роду Aviadenovirus для виготовлення вакцини.
Вакцинна композиція проти репродуктивного та респіраторного дистрес-синдрому свиней, спосіб імунізації свиней проти репродуктивного та респіраторного дистрес-синдрому свиней, спосіб одержання вакцини проти реп

Номер патенту: 39991
Опубліковано: 16.07.2001
Автори: Кладек Денні В., Коркіка Дейвід Е., Харріс Луі Л.
МПК: A61K 39/12, A61K 47/42, C12N 7/00, C12N 7/08, C12N 7/02, A61P 31/12, A61K 35/76
Мітки: вакцинна, вакцини, спосіб, дистрес-синдрому, свиней, реп, репродуктивного, одержання, респіраторного, композиція, імунізації
Формула / Реферат:
1. Вакцинна композиція проти репродуктивного та респіраторного дистрес-синдрому свиней (PRRS), що містить у собі авірулентний штам вірусу репродуктивного та респіраторного дистрес-синдрому свиней (PRRS) АТСС-VR2332 і фармакологічно прийнятний агент-носій.2. Композиція за п. 1, де зазначений агент-носій містить у собі сахарозо-желатиновий агент-стабілізатор.3. Композиція за п. 1, де зазначений штам вірусу був пасований 75 разів і...