Білкова конструкція, яка включає модифікований поліпептид casb7439
Номер патенту: 99801
Опубліковано: 25.09.2012
Автори: Ріоу Клемен, Блейс Норманд, Гарві Мартін, Пілорджет Ентоні







Формула / Реферат
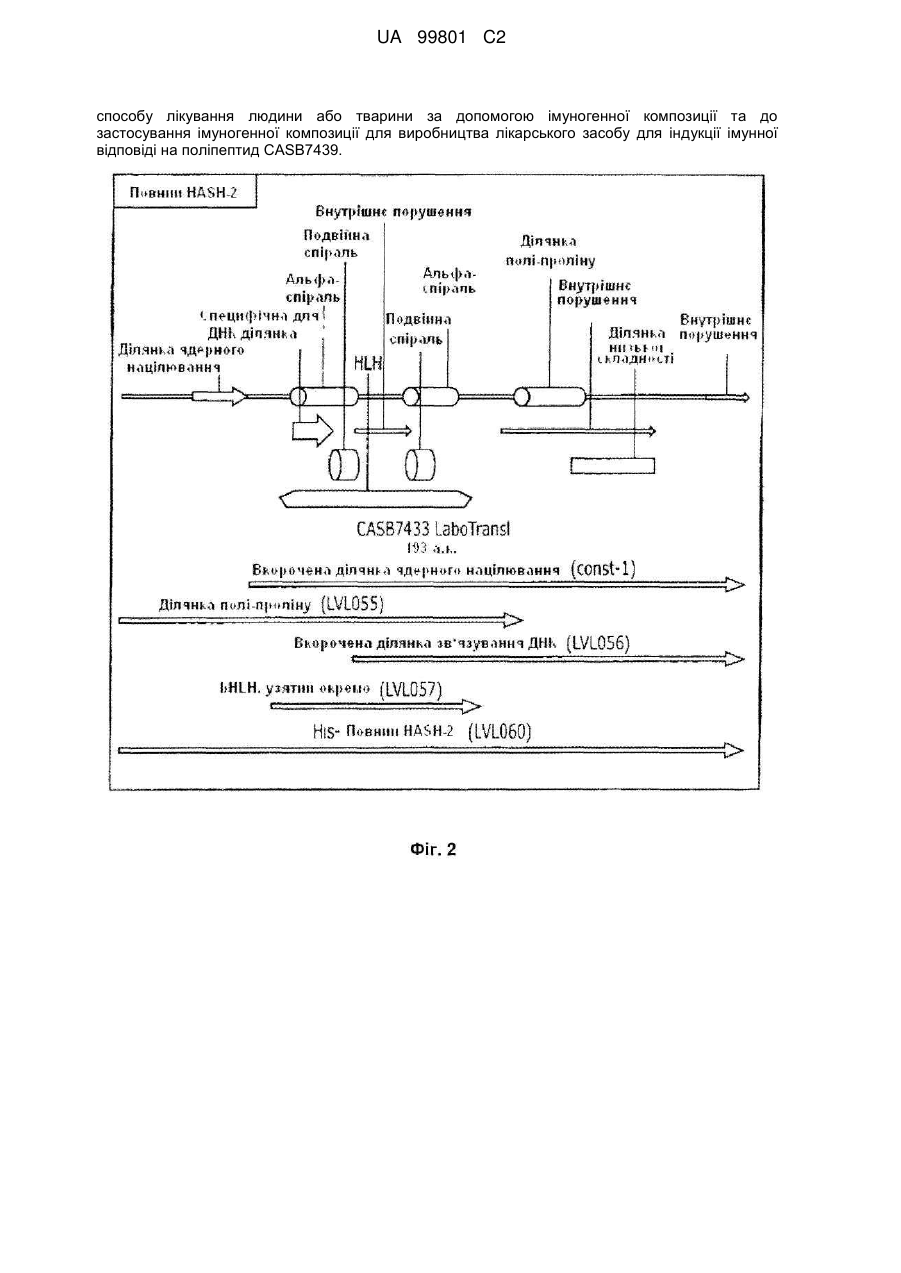
1. Білкова конструкція, що включає модифікований CASB7439 поліпептид, який є вибраним з:
(a) LVL055 (SEQ ID NO: 1);
(b) LVL111 (SEQ ID NO: 3);
(c) LVL137(SEQ ID NO: 5);
(d) LVL141 (SEQ ID NO: 7);
(e) LVL144 (SEQ ID NO: 9); та
(f) LVL168 (SEQ ID NO: 11).
2. Імуногенна композиція, що включає білкову конструкцію за пунктом 1 та фармацевтично прийнятний носій або наповнювач, де носій або наповнювач може необов'язково включати буфер.
3. Імуногенна композиція за пунктом 2, що додатково включає ад'ювант.
4. Імуногенна композиція за пунктом 3, де ад'ювант включає принаймні один з наступних: TLR-4 агоніст, імунологічно активну фракцію сапоніну, TLR-9 агоніст.
5. Імуногенна композиція за пунктом 4, де вказаний TLR-4 агоніст являє собою 3D-MPL та/або вказаний TLR-9 агоніст являє собою CpG олігонуклеотид, та/або імунологічно активна фракція сапоніну являє собою QS21.
6. Імуногенна композиція за пунктом 5, що включає 3D-MPL та/або CpG, та/або CpG, та/або QS21.
7. Імуногенна композиція за пунктом 6, що додатково включає холестерин.
8. Молекула нуклеїнової кислоти, що включає полінуклеотидну послідовність, яка кодує білкову конструкцію за пунктом 1.
9. Спосіб індукції імунної відповіді на CASB7439 у людини або у тварини, відмінної від людини, що включає введення людині або тварині, відмінній від людини, ефективної кількості композиції, що включає ад'ювант та білок, який включає поліпептидну послідовність, вибрану із групи, що складається з:
(a) поліпептидної послідовності, представленої у SEQ ID NO: 9; та
(b) поліпептидної послідовності, представленої у SEQ ID NO: 11.
10. Спосіб лікування людини або тварини, відмінної від людини, що включає етапи:
(a) вибору людини або тварини, відмінної від людини, що має ракові клітини, які експресують CASB7439; та
(b) введення людині або тварині, відмінній від людини, ефективної кількості композиції, що включає ад'ювант та білок, який включає поліпептид, вибраний із групи, яка складається з:
і) конструкції, представленої у SEQ ID NO: 9; та
іі) конструкції, представленої у SEQ ID NO: 11.
11. Імуногенна композиція, як визначено у пунктах 2-7, для застосування у способі індукції імунної відповіді на CASB7439 у людини або у тварини, відмінної від людини, де вказаний спосіб включає введення ефективної кількості вказаної імуногенної композиції.
12. Імуногенна композиція за пунктом 11, яка включає білок, який включає поліпептид, вибраний із групи, яка складається з:
(a) конструкції, представленої у SEQ ID NO: 9; та
(b) конструкції, представленої у SEQ ID NO: 11.
13. Застосування імуногенної композиції, як визначено у пунктах 2-7, у виробництві лікарського засобу для застосування у способі індукції імунної відповіді на CASB7439 у людини або у тварини, відмінної від людини, де вказаний спосіб включає введення ефективної кількості вказаної імуногенної композиції.
Текст
Реферат: Винахід належить до білкової конструкції, що включає модифікований поліпептид CASB7439. Також винахід належить до імуногенної композиції, що містить білкову конструкцію, молекули нуклеїнової кислоти, яка її кодує, способу індукції імунної відповіді на поліпептид CASB7439, UA 99801 C2 (12) UA 99801 C2 способу лікування людини або тварини за допомогою імуногенної композиції та до застосування імуногенної композиції для виробництва лікарського засобу для індукції імунної відповіді на поліпептид CASB7439. UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 Передумови створення винаходу Гомологи Achaete-Scute ссавців являють собою консервативних родичів комплексу AchaeteScute дрозофіли у ссавців. Вони є членами родини основного гена спіраль-петля-спіраль (bHLH або HLH), що функціонують як специфічні для лінії фактори транскрипції, які є суттєвими для розвитку. Один мишачий член родини, MASH2, є суттєвим для розвитку плаценти, але не є важливим для розвитку самого ембріона. HASH2, людський ортолог гена MASH2, було клоновано у 1997 Alders та ін. (позначається авторами як "ASCL2"). Alders та ін. (1997) Hum. Molec. Genet. 6: 859-867. Як описується у W001/62778, дослідження експресії виявили, що транскрипт HASH2 (називається у даній заявці як CASB7439) понадекспресується у колоректальних пухлинах у порівнянні із сусідніми нормальними тканинами кишечнику та іншими дослідженими нормальними тканинами. Цей ген понадекспресується у пацієнтів, які мають стадію I - IV аденокарциноми. Таким чином, цей білок може вважатися раковим антигеном, корисним в імунотерапевтичному підході, що використовується для полегшення раку у суб‗єкта, наприклад, при використанні рекомбінантного CASB7439 як антигенспецифічного імунотерапевтичного засобу для лікування раку (ASCI). Короткий виклад суті винаходу В даній заявці забезпечуються сполуки та способи для підвищення рекомбінантної продукції CASB7439 поліпептидів. У деяких втіленнях забезпечуються модифіковані CASB7439 поліпептиди, а також білкові конструкції, що включають такі модифіковані CASB7439 поліпептиди. Вказані модифіковані CASB7439 поліпептиди включають принаймні одну модифікацію для поліпшеної продукції CASB7439. Заявники також розкрили молекули нуклеїнової кислоти, що включають полінуклеотидну послідовність, яка кодує модифіковані CASB7439 поліпептиди та білкові конструкції, що включають такі модифіковані CASB7439 поліпептиди, як описується в даній заявці. У деяких аспектах винахідники розкрили білкові конструкції, що мають амінокислотні (aa) послідовності та нуклеотидну послідовність (ДНК), що їх кодує, як описано нижче: - LVL055 [SEQ ID N0:1 (aa); SEQ ID NО:2 (ДНК)]; - LVL111 [SEQ ID N0:3 (aa); SEQ ID NО:4 (ДНК)]; - LVL137 [SEQ ID N0:5 (aa); SEQ ID NО:6 (ДНК)]; - LVL141 [SEQ ID N0:7 (aa); SEQ ID NО:8 (ДНК)]; - LVL144 [SEQ ID N0:9 (aa); SEQ ID NО:10 (ДНК)]; та - LVL168 [SEQ ID N0:11 (aa); SEQ ID NО:12 (ДНК)]. Також розкриваються способи та процеси, що використовують молекули нуклеїнової кислоти, описані в даній заявці, для продукції білкових конструкцій. Заявники також розкривають імуногенні композиції, при цьому ці композиції включають один або більше модифікованих CASB7439 поліпептидів або конструкцій, описаних в даній заявці, та фармацевтично прийнятний носій або наповнювач, де носій або наповнювач може необов‗язково включати буфер. У деяких втіленнях розкривається застосування модифікованих CASB7439 поліпептидів або білкових конструкцій, описаних в даній заявці — або молекул нуклеїнової кислоти, що їх кодують — у приготуванні лікарського засобу для лікування колоректального раку. У деяких втіленнях розкриваються білкові конструкції, що описані в даній заявці, для застосування у терапії, зокрема терапії колоректального раку. У деяких втіленнях розкриваються способи для стимуляції імунної відповіді проти CASB7439 у суб‗єкта з колоректальним раком, при цьому такий спосіб включає: (a) відбір суб‗єкта з колоректальним раком; та (b) введення такому суб‗єктові ефективної кількості імуногенної композиції, що включає модифіковані CASB7439 поліпептиди або конструкції, описані в даній заявці, та фармацевтично прийнятний носій або наповнювач, де носій або наповнювач може необов‗язково включати буфер. Короткий опис фігур Фігура 1/35. Ця фігура представляє схематичну діаграму, що відображає різноманітні ділянки CASB7439 поліпептиду, зокрема: специфічний для ДНК домен (домен зв‗язування ДНК), bHLH домен та ділянку, збагачену на пролін. Див. Приклад 2 для деталей. Схематичне зображення є також представленим для наступних ділянок: ядерного націлювання (сіра стрілка); альфа-спіраль (сірі циліндри); подвійна спіраль (чорні циліндри); внутрішнє порушення (чорні стрілки); та ділянка низької складності(сірий прямокутник). Фігура 2/35. Ця фігура представляє схематичну діаграму, що відображує різноманітні вкорочені варіанти CASB7439 поліпептиду (чорні стрілочки), вирівняні з повним варіантом CASB7439 поліпептиду. Див. Приклад 4 для детальної інформації. 1 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 Фігура 3/35. Ця фігура представляє схематичну діаграму CASB7439, демонструючи приблизне розміщення збагаченої на пролін ділянки, а також ідентифікованих та спрогнозованих епітопів. Різні втілення включають CASB7439 поліпептид, модифікований шляхом видалення деяких або усіх амінокислот у межах ділянки, збагаченої на пролін. Такі модифіковані CASB7439 поліпептиди зберігають максимальну частину епітопів у межах CASB7439 поліпептиду. Фігура 4/35. Показане вирівнювання між послідовністю немодифікованого CASB7439 поліпептиду (SEQ ID NO:13) та послідовністю поліпептиду, що походить від двох можливих модифікацій CASB7439 поліпептиду: перша модифікація є такою, в якій 21 суміжних амінокислотних залишків (133-153, включно) є делетованими; друга модифікація є такою, в якій 21 суміжних амінокислотних залишків (131-151, включно) є делетованими. Крім того, може бути представлена третя модифікація CASB7439 поліпептиду, в якій 21 суміжних амінокислотних залишків (132-152, включно) є делетованими. Як представлено на цій фігурі одержана модифікована послідовність CASB7439 поліпептиду (вставка, обведена кружечком) у ділянці делеції є однаковою для усіх трьох модифікацій. Причиною цього є повторювані RG амінокислотні залишки у положеннях 131-132 та 152-153 SEQ ID NO:13. Фігура 5/35. Ця фігура представляє вирівнювання амінокислотної послідовності між білковими конструкціями Прикладу 6. CASB7439 = амінокислотна послідовність HASH2 (SEQ ID NO:13); LVL088 (SEQ ID NO:27); LVL111 (SEQ ID NO:3); LVL137 (SEQ ID NO:5); LVL138 (SEQ ID NO:33); LVL168 (SEQ ID NO:11). Фігура 6/35. Ця фігура представляє вирівнювання між амінокислотною послідовністю HASH2 (CASB7439) (SEQ ID NO: 13) та наступними білковими конструкціями: LVL168 (SEQ ID NO:11); pD1/3 (SEQ ID NO:39) та LVL144 (SEQ ID NO:9). Фігура 7/35. Кількісний ПЛР аналіз у реальному часі експресії іРНК CASB7439 у нормальних тканинах (12) та тканинах колоректального раку (83). Див. Приклад 9. Фігура 8/35. Відповідність між експресією іРНК CASB7439 та білка у різноманітних зразках колоректального раку людини (CRC). • (чорні кружечки) представляють експресію іРНК CASB7439 у зразках колоректального раку, у той час як O (незаштриховані кружечки) представляють експресію іРНК CASB7439 у нормальних прилеглих тканинах. Відповідний рівень CASB7439 білка, виявлений за допомогою імунофлуоресценції позначається зверху кожного зразка (- відсутність експресії, +/- дуже низька експресія, + низька експресія, ++ сильна експресія та +++ дуже сильна експресія). CRC: первинні пухлини, METS: метастази, NAT: нормальні прилеглі тканини. Усі зразки являють собою аденокарциноми товстого кишечнику за винятком зразків 22440 та 24762, які являють собою слизоутворювальні карциноми. Див. Приклад 9. Фігура 9/35: Банк пептидів, що охоплюють білкову послідовність CASB7439, яка використовується для повторної стимуляції спленоцитів після імунізації за допомогою різних CASB7439 конструкцій, є представленим у цій таблиці. Див. Приклад 10. Фігура 10/35. Представлений матрикс CASB7439 пептиду, що використовується для дослідження імуногенності T-клітин. Див. Приклад 10. Фігура 11/35. CD4 відповідь, виражена як процент подвійно позитивних (IFNγ/TNFα) CD4 Tклітин у чотирьох лініях інбредних мишей (C57BL6, BALBC, CB6F1 та C3H), імунізованих при використанні LVL111 + AS01B з наступною повторною стимуляцією за допомогою CASB7439 пептидів, що перекриваються (пули, матричний підхід). Див. Приклад 10, експеримент з порівняння декількох ліній інбредних мишей. Фігура 12/35. CD4 відповідь, виражена як процент подвійно позитивних (IFNγ/TNFα) CD4 Tклітин у чотирьох лініях інбредних мишей, імунізованих при використанні LVL111 + AS15 з наступною повторною стимуляцією за допомогою CASB7439 пептидів, що перекриваються (пули, матрикчний підхід). Див. Приклад 10, експеримент з порівняння декількох ліній інбредних мишей. Фігура 13/35. Ідентифікація CD4 імуногенних CASB7439 пептидів у чотирьох лініях інбредних мишей, що обговорюються у попередньому абзаці. Світло-сірий = 0,2-0,4 % подвійно позитивних (IFNγ/TNFα) CD4 T-клітин; темно-сірий > 0,5% подвійно позитивних (IFNγ/TNFα) CD4 T-клітин. Див. Приклад 10, експеримент з порівняння декількох ліній інбредних мишей. Фігура 14/35. Ідентифікація CD4 імуногенних CASB7439 пептидів у аутбредних CD1 мишей (AS01B верхня частина; AS15 нижня частина). Світло-сірий = 0,2-0,4 % подвійно позитивних (IFNγ/TNFα) CD4 T-клітин; темно-сірий > 0,5% подвійно позитивних (IFNγ/TNFα) CD4 T-клітин. Див. Приклад 10, імуногенність CASB7439 у мишей: T-клітинна імуногенність CASB7439 у аутбредних мишей. 2 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 Фігура 15/35. CD4 та CD8 T-клітинні відповіді, виражені як процент подвійної позитивності (IFNγ/TNFα) в індивідуальних CD1 аутбредних мишей, імунізованих при використанні LVL111, LVL168 або LVL144, рецептованих з AS15, наступна повторна стимуляція при використанні CASB7439 пептидів, що перекриваються (пули імунодомінантних пептидів). Див. Приклад 10, дослідження LVL111, LVL168 та LVL144 у аутбредних мишей. Фігура 16/35. CD4 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα)) в об‗єднаних зразках лейкоцитів периферичної крові (PBL), ізольованих з HLA A2.1/DR-1 трансгенних мишей, імунізованих при використанні або LVL168 (SEQ ID NO:11), або LVL144 (SEQ ID NO:9), рецептованих з AS15, наступна повторна стимуляція при використанні CASB7439 пептидів, що перекриваються (7 пулів 7 пептидів/пул + пептид 24 (SEQ ID NO:76)). Фігура 17/35. CD8 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα)) в об‗єднаних PBL, ізольованих з HLA A2.1/DR-1 трансгенних мишей, імунізованих при використанні або LVL168 (SEQ ID NO:11), або LVL144 (SEQ ID NO:9), рецептованих з AS15, наступна повторна стимуляція при використанні CASB7439 пептидів, що перекриваються (7 пулів 7 пептидів/пул + пептид 24 (SEQ ID NO:76)). Фігура 18/35. CD4 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα)) в об‗єднаних PBL, ізольованих з HLA A2.1/DR-1 трансгенних мишей, імунізованих при використанні або LVL168 (SEQ ID NO:11), або LVL144 (SEQ ID NO:9), рецептованих з AS15, наступна повторна стимуляція при використанні CASB7439 пептидів, що перекриваються (7 пулів 7 пептидів/пул + пептид 24 (SEQ ID NO:76)). Фігура 19/35. CD8 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα)) у спленоцитах, ізольованих з HLA A2.1/DR-1 трансгенних мишей, імунізованих при використанні або LVL168 (SEQ ID NO:11), або LVL144 (SEQ ID NO:9), рецептованих з AS15, наступна повторна стимуляція при використанні CASB7439 пептидів, що перекриваються (7 пулів 7 пептидів/пул + пептид 24 (SEQ ID NO:76)). Фігура 20/35. Аналіз залежності від часу утворення антитіл, специфічних до CASB7439 (lgG1 та lgG2a), що запускається при LVL111 + AS01B або LVL111 + AS15 імунізаціях CB6f1 інбредних мишей. Див. Приклад 10, імуногенність CASB7439 у мишей: опосередкована CASB7439 гуморальна відповідь. Фігура 21/35. Специфічна для CASB7439 гуморальна відповідь у інбредних CB6f1 мишей (lgG1 та lgG2a) при імунізаціях з використанням LVL111, LVL168 або LVL144 рецептованих з AS15 ад‗ювантом або з AS15, узятим окремо. Див. Приклад 10, імуногенність CASB7439 у мишей: опосередкована CASB7439 гуморальна відповідь. Фігура 22/35. Специфічна для CASB7439 гуморальна імунна відповідь (титр IgG) у CD1 аутбредних мишей, імунізованих при використанні LVL111, LVL168 або LVL144, рецептованих з AS15 ад‗ювантом, або при використанні AS15 ад‗юванту, узятого окремо; інтактних мишей використовували як контроль. Див. Приклад 10, дослідження CD1 мишей. Фігура 23/35. TC1/CASB7439 #14 пухлинний трансплантат у CB6f1 мишей, імунізованих при використанні LVL111, рецептованого з AS01B (верхня частина) або AS15 (нижня частина). Див. Приклад 11, дослідження #1.1. У верхній частині порядок кривих на графіку від верхньої частини до нижньої частини є наступним 'буфер', LVL111 (10 мкг), LVL111 (10 мкг) + AS01B та LVL111 (1 мкг) + AS01B. У нижній частині верхня крива являє собою LVL111 (10 мкг) + AS15. Фігура 24/35. Криві виживання з Прикладу 11, дослідження #1.2 є представленими у верхній, середній та нижній частині. TF: вільний від пухлини. (Верхня частина) Крива виживання CB6f1 мишей, імунізованих при використанні буфера на основі фізіологічного розчину, LVL111 без ад‗юванта або тільки ад‗юванта (AS01 або AS15) та сенсибілізованих при використанні TC1/CASB7439 #14 клітин. Остання миша, що залишилася живою, з групи мишей, імунізованих при використанні LVL111, померла першою, після неї померла остання миша, що залишилася живою, з групи AS01B, після неї померла остання миша, що залишилася живою, з групи AS15 та остання миша, що залишилася живою, з групи буфера. (Середня частина) Крива виживання CB6f1 мишей, імунізованих при використанні AS15, узятого окремо, або зазначеними дозами LVL111, рецептованими з AS15, та сенсибілізованих при використанні TC1/CASB7439 #14 клітин. Остання миша, що залишилася живою, з групи AS15, померла першою. З інших груп група 30 мкг мала найменше значення особин, що вижили, далі йшла група 1 мкг, потім група 10 мкг, яка мала найвищу кількість особин, що вижили. (Нижня частина) Крива виживання CB6f1 мишей, імунізованих при використанні AS01B, узятого окремо, або зазначеними дозами LVL111, рецептованими з AS01B, та сенсибілізованих за допомогою TC1/CASB7439 #14 клітин. У цій частині останні миші з груп 10 мкг та AS01B гинули в один й той самий час. Групи 1 мкг та 30 мкг мали однаковий процент виживання у заключний день. 3 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Фігура 25/35. Представлені криві виживання з Прикладу 11, дослідження #1.3. TF: вільні від пухлини. (Верхня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні LVL111, LVL144 або LVL168, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14 клітин. (Нижня частина) Крива виживання CB6f1 мишей, імунізованих при використанні LVL111, LVL144 або LVL168, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою MC38/CASB7439#35 клітин. Фігура 26/35. Представлені криві виживання з Прикладу 11, дослідження #1.4. TF: вільні від пухлини. (Верхня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні LVL144 або LVL168, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14 клітин. Група мишей, імунізованих при використанні LVL144, мала трохи більше значення проценту виживання. (Середня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні LVL144 або LVL168, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою клональних TC1/CASB7439 #14-2 клітин. Миші, яких інокулювали при використанні LVL144, мали трохи більше значення проценту виживання. (Нижня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні LVL144 або LVL168, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою MC38/CASB7439 #35 клітин. Миші, яких інокулювали LVL144, не мали особин, що вижили. Фігура 27/35. Представлені криві виживання з Прикладу 11, дослідження #2.1. TF: вільні від пухлини. (Верхня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 6,25 мкг LVL168 та 10 мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, імунізованих при використанні LVL168, мала трохи більше значення проценту виживання. (Верхня частина середини) Крива виживання для CB6f1 мишей, імунізованих при використанні 3,1 мкг LVL168 та 5 мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, інокульованих при використанні LVL144, мала трохи більше значення проценту виживання. (Нижня частина середини) Крива виживання для CB6f1 мишей, імунізованих при використанні 1,55 мкг LVL168 та 2,5 мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, інокульованих за допомогою LVL144, мала трохи більше значення проценту виживання. (Нижня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 0,77 мкг LVL168 та 1,25 мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, інокульованих за допомогою LVL168, мала трохи вище значення проценту виживання. Фігура 28/35. Представлені криві виживання з Прикладу 11, дослідження #2.2. TF: вільні від пухлини. (Верхня частина) Крива виживання для C57BI/6 мишей, імунізованих при використанні 6,25 мкг LVL168 або 10 мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. (Верхня частина середини) Крива виживання для C57BI/6 мишей, імунізованих при використанні 3,1 мкг LVL168 або мкг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. (Нижня частина середини) Крива виживання для C57BI/6 мишей, імунізованих при використанні 1,55 нг LVL168 або 2,5 нг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. (Нижня частина) Крива виживання для C57BI/6 мишей, імунізованих при використанні 0,77 нг LVL168 або 1,25 нг LVL144, рецептованих з AS15, або при використанні AS15, узятого окремо, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Фігура 29/35. Представлені криві виживання з Прикладу 11, Дослідження #2.3. TF: вільні від пухлини. (Верхня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 6,25 мкг LVL168 або 10 мкг LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання, після неї йшла група, яка одержувала LVL144. Не було особин, що вижили, у групі, яка одержувала тільки AS15. (Середня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 3,1 мкг LVL168 та 5 мкг LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання, після неї йшла група, яка одержувала LVL144. Не було особин, що вижили, у групі, яка одержувала тільки AS15. (Нижня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 1,55 мкг LVL168 та 2,5 мкг 4 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання, після неї йшла група, яка одержувала LVL144. Не було особин, що вижили, у групі, яка одержувала тільки AS15. Буфер на основі фізіологічного розчину або AS15, узятий окремо, використовували як контролі. Фігура 30/35. Криві виживання з Прикладу 11, дослідження #2.3. TF: вільні від пухлин. (Верхня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 0,77 мкг LVL168 або 1,25 мкг LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання, після неї йшла група, яка одержувала LVL144. Не було особин, що вижили, у групі, яка одержувала тільки AS15. (Середня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 0,19 нг LVL168 або 0,31 нг LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання. Остання миша, що залишилася живою, з групи, яка одержувала LVL144, переживала останню особину, що вижила, як з групи AS15, так і з групи буфера. (Нижня частина) Крива виживання для CB6f1 мишей, імунізованих при використанні 0,048 мкг LVL168 або 0,078 нг LVL144, рецептованих з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. Група мишей, що одержувала LVL168, мала найвище значення проценту виживання. Остання миша, що залишилася живою, з групи, яка одержувала LVL144, переживала останню особину, що вижила, як з групи AS15, так і з групи буфера. Буфер на основі фізіологічного розчину або AS15, узятий окремо, використовували як контролі. Фігура 31/35. TC1/CASB7439 #14-2 пухлинні трансплантати у самців та самок CB6f1 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15; AS15, узятий окремо, використовували як контроль (верхня частина). TC1/CASB7439 #14-2 пухлинні трансплантати у C57BI/6 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15; AS15, узятий окремо, використовували як контроль (нижня частина). Фігура 32/35. Верхня частина: Крива виживання для самців або самок CB6f1 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. AS15, узятий окремо, використовували як контроль як у самців, так і у самок CB6f1 мишей. Серед CB6f1 мишей, імунізованих при використанні LVL168 плюс AS15, були 7 вільних від пухлин (TF) самців та 3 TF самки. Не було TF серед мишей, які одержували AS15, узятий окремо. Нижня частина: Крива виживання для C57BI/6 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15, та сенсибілізованих за допомогою TC1/CASB7439 #14-2 клітин. AS15, узятий окремо, використовували як контроль як у самців, так і у самок C57BI/6 мишей. Серед C57BI/6 мишей, імунізованих при використанні LVL168 плюс AS15, були 2 TF самці та 1 TF самка. Серед мишей, які одержували AS15, узятий окремо, були 5 TF самців та 1 TF самка. Фігура 33/35. Специфічна для CASB7439 гуморальна імунна відповідь (титр IgG) у самців (чорні кружечки) та самок (сірі кружечки) CB6F1 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15, або при використанні AS15, узятого окремо (зліва). Специфічна для CASB7439 гуморальна імунна відповідь (титр IgG) у самців (чорні кружечки) та самок (сірі кружечки) C57/BL6 мишей, імунізованих при використанні 1 мкг LVL168, рецептованого з AS15, або при використанні AS15, узятого окремо (справа). Фігура 34/35. CD4 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα) в об‗єднаних PBL, ізольованих з CB6F1 мишей, імунізованих при використанні або 1 мкг LVL168 (SEQ ID NO:11), рецептованого з AS15, або при використанні AS15, узятого окремо, наступна повторна стимуляція при використанні банку 46 пептидів (див. Фігуру 9), що охоплюють повну білкову послідовність CASB7439 (верхня частина). CD4 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα) в об‘єднаних PBL, ізольованих з C57/BL6 мишей, імунізованих при використанні або 1 мкг LVL168 (SEQ ID NO:11), рецептованого з AS15, або при використанні AS15, узятого окремо, наступна повторна стимуляція при використанні CASB7439 об‗єднаного банку 46 пептидів (див. Фігуру 9), що охоплюють повну білкову послідовність CASB7439 (нижня частина зліва); CASB7439 пептиду 39 (SEQ ID NO:91) (нижня частина посередині); або RPMI (нижня частина справа). Фігура 35/35. CD8 T-клітинні відповіді (виражені як процент подвійної позитивності (IFNγ/TNFα) в об‗єднаних PBL, ізольованих від CB6F1 мишей, імунізованих при використанні або 1 мкг LVL168 (SEQ ID NO:11), рецептованого з AS15, або при використанні AS15, узятого окремо, наступна повторна стимуляція при використанні банку 46 пептидів (див. Фігуру 9), що охоплюють повну білкову послідовність CASB7439 (верхня частина). CD8 T-клітинні відповіді 5 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 (виражені як процент подвійної позитивності (IFNγ/TNFα) в об‗єднаних PBL, ізольованих з C57/BL6 мишей, імунізованих при використанні або 1 мкг LVL168 (SEQ ID NO:11), рецептованого з AS15, або при використанні AS15, узятого окремо, наступна повторна стимуляція при використанні CASB7439 об‗єднаного банку 46 пептидів (див. Фігуру 9), що охоплюють повну білкову послідовність CASB7439 (нижня частина зліва); CASB7439 пептиду 39 (SEQ ID NO:91) (нижня частина посередині); або RPMI (нижня частина справа). Детальний опис винаходу Однією з перешкод для одержання комерційно прийнятного імунотерапевтичного агента є продукція комерційно прийнятних кількостей рекомбінантного ракового антигену. У загальному випадку та без обмеження, рівень комерційно прийнятної продукції для рекомбінантного поліпептиду приблизно складає 5% від кількості загального білка, який виробляється хазяйськими клітинами, як оцінюється на забарвленому Кумасі блакитним SDS-ПАГЕ гелі. Існує багато причин, чому нативна поліпептидна послідовність може забезпечувати тільки невеликі кількості, коли їх одержують за допомогою рекомбінантних способів. Підходи до підвищення продукції поліпептидів включають варіацію хазяйських систем експресії або оптимізацію кодонів. Проте такі стандартні підходи не завжди приводять до прийнятної поліпептидної продукції, тому продукція деяких поліпептидів залишається комерційно недосяжною. Як описано більш докладно нижче, заявники дослідили, чи буде модифікований CASB7439 поліпептид продукувати прийнятні рівні білка при використанні рекомбінантних способів. У деяких втіленнях забезпечуються модифіковані CASB7439 поліпептиди, в яких модифікація полягає у або включає C-термінальне вкорочення CASB7439 поліпептиду такого, що уся або частина ділянки, збагаченої на пролін, видаляється. У деяких втіленнях модифікація CASB7439 поліпептиду полягає у або включає делецію усієї або частини ділянки, збагаченої на пролін. У деяких втіленнях забезпечуються модифіковані CASB7439 поліпептиди, які зберігають принаймні амінокислотний залишок 193 SEQ ID NO: 13, проте де уся або частина ділянки, збагаченої на пролін, є делетованою. В інших втіленнях забезпечуються білкові конструкції, що включають модифікований CASB7439 поліпептид, як описано в даній заявці. У деяких втіленнях такі білкові конструкції додатково включають один або більше гетерологічних поліпептидів. У деяких втіленнях гетерологічний поліпептид являє собою партнера злиття, як описується більш детально у будьякому місці даної заявки. В особливо прийнятних втіленнях такі білкові конструкції включають гетерологічний поліпептид, розміщений N-термінально від модифікованого CASB7439 поліпептиду. У додаткових прийнятних втіленнях забезпечуються білкові конструкції, в яких гетерологічний поліпептид являє собою фрагмент білка D H. influenzae (як описується більш детально нижче), розміщений N-термінально від модифікованого CASB7439 поліпептиду. У деяких втіленнях забезпечуються білкові конструкції, в яких гетерологічний поліпептид включає полігістидинову ділянку. У додаткових втіленнях полігістидинова послідовність є розміщеною у C-термінальній частині білкової конструкції. В інших втіленнях полігістидинова послідовність є розміщеною поблизу C-термінальної частини білкової конструкції. У додаткових втіленнях полігістидинова послідовність є розміщеною у N-термінальній частині білкової конструкції. В інших втіленнях полігістидинова послідовність є розміщеною поблизу Nтермінальної частини білкової конструкції. У прийнятних втіленнях полігістидинова ділянка включає десять або більше послідовних залишків гістидину. В особливо прийнятних втіленнях полігістидинова ділянка включає десять або більше послідовних залишків гістидину, розміщених на N-термінальному кінці модифікованого CASB7439 поліпептиду. У деяких втіленнях більше однієї полігістидинової ділянки може включатися в одному або більше положеннях, описаних в даній заявці. У деяких втіленнях забезпечуються білкові конструкції, в яких гетерологічний поліпептид являє собою білок носія, хімічно кон‗югований з модифікованим CASB7439 поліпептидом. У деяких втіленнях забезпечуються білкові конструкції, що включають гетерологічний поліпептид, що являє собою полігістидинову ділянку, та гетерологічний поліпептид, що є білком носія. У деяких втіленнях забезпечуються білкові конструкції, що включають гетерологічний поліпептид, що являє собою полігістидинову ділянку, та гетерологічний поліпептид, що є партнером злиття. У деяких втіленнях забезпечуються білкові конструкції, що включають гетерологічний поліпептид, який являє собою полігістидинову ділянку, гетерологічний поліпептид, що є партнером злиття, та гетерологічний поліпептид, який є білком носія. Заявники також розкривають втілення кожної конструкції, описаної у цих абзацах даної заявки, в яких деякі або усі з будь-яких гетерологічних поліпептидів, таких, як полігістидинова ділянка, були видалені. Композиції та способи для видалення гетерологічних поліпептидів є 6 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 відомими у галузі техніки, включаючи, але без обмеження, перетравлювання за допомогою ендо- або eкзопептидаз, хімічної модифікації або розщеплення зв‗язку між гетерологічним поліпептидом та іншою частиною молекули, та подібні до них Заявники також розкривають у даній заявці імуногенні композиції; такі композиції включають будь-який модифікований CASB7439 поліпептид або білкову конструкцію, що включає модифікований CASB7439 поліпептид, як описано в даній заявці, та фармацевтично прийнятий носій або наповнювач, де носій або наповнювач може необов‗язково включати буфер. У деяких аспектах такі композиції додатково включають ад‗ювант. У деяких аспектах такі композиції включають ад‗ювант, що викликає принаймні TH1 імунну відповідь. У деяких аспектах ад‗ювант, описаний в даній заявці, включає принаймні один з: 3D-MPL, QS21 та CpG. У деяких втіленнях композиція включає 3D-MPL. В інших втіленнях композиція включає CpG. У деяких втіленнях композиція включає QS21. У деяких втіленнях композиція включає QS21 та холестерин. У деяких втіленнях композиція включає холестерин, 3D-MPL та QS21 у ліпосомній композиції. Заявники також розкривають в даній заявці молекули нуклеїнової кислоти, що включають полінуклеотидні послідовності, які кодують модифіковані CASB7439 поліпептиди та конструкції, як описано в даній заявці. У деяких втіленнях розкривається вектор, що включає такі молекули нуклеїнової кислоти, як описано в даній заявці. У деяких втіленнях вектор включає прокаріотичний експресійний вектор. У деяких втіленнях вектор включає еукаріотичний експресійний вектор. У деяких втіленнях полінуклеотидна послідовність, що кодує конструкцію, є оптимізованою за кодонами для експресії у хазяйській клітині. Заявники також розкривають хазяйські клітини, що включають (i) молекулу нуклеїнової кислоти, як описано в даній заявці, або (ii) вектор, що включає молекулу нуклеїнової кислоти, як описано в даній заявці. У деяких втіленнях хазяйська клітина є вибраною з групи: бактеріальних клітин, клітин комах та клітин ссавців. Деякі втілення забезпечують імуногенну композицію, що включає (i) молекулу нуклеїнової кислоти, описану в даній заявці або (ii) вектор, що включає молекулу нуклеїнової кислоти, як описано в даній заявці, та фармацевтично прийнятний носій або наповнювач. У деяких втіленнях забезпечується застосування модифікованих CASB7439 поліпептидів або білкових конструкцій, описаних в даній заявці (або молекул нуклеїнової кислоти, що їх кодують), в одержанні лікарського засобу для лікування колоректального раку. У деяких втіленнях розкриваються модифіковані CASB7439 поліпептиди або білкові конструкції, описані в даній заявці (або молекули нуклеїнової кислоти, що їх кодують) для застосування у терапії, зокрема, терапії колоректального раку. У деяких втіленнях забезпечуються способи для стимуляції імунної відповіді проти CASB7439 у суб‗єкта з колоректальним раком, при цьому такий спосіб включає: (i) відбір суб‗єкта з колоректальним раком; та (ii) введення суб‗єктові ефективної кількості імуногенної композиції, що включає модифікований CASB7439 поліпептид або білкову конструкцію, як описано в даній заявці, та фармацевтично прийнятний носій або наповнювач, де носій або наповнювач може необов‗язково включати буфер. У деяких втіленнях способів, розкритих в даній заявці, імуногенна композиція додатково включає ад‗ювант. У деяких втіленнях способу, описаного в даній заявці, ад‗ювант викликає принаймні TH1 імунну відповідь. У деяких втіленнях o способу, описаного в даній заявці, ад‗ювант включає принаймні один з: 3D-MPL, QS21 та CpG. У деяких втіленнях способу, описаного в даній заявці, суб‗єкт являє собою людину. Білкові конструкції CASB7439 поліпептиди CASB7439, як він називається в даній заявці, є також відомим як HASH2 або ASCL2. HASH2, являє собою поліпептид, що походить від людини та складається з 193 амінокислотних залишків (SEQ ID NO: 13). Див., наприклад, номер доступу AAB86993. В даній заявці посилання на характеристики CASB7439 буде зроблене стосовно поліпептидної послідовності, що забезпечується у SEQ ID NO: 13. Збагачена на пролін ділянка Ділянка з високою повторюваністю проліну може бути знайдена у послідовності SEQ ID NO: 13 у ділянці амінокислот 127-158. Заявники називають ділянку послідовності SEQ ID NO:13 від амінокислоти 133 дo амінокислоти 153, включно, "збагаченою на пролін," "поліпроліновою," "подібною до поліпроліну," або "поліпро" ділянкою або доменом. Згідно з цим, як використовується в даній заявці, "збагачена на пролін," "поліпролінова," "подібна до поліпроліну," або "поліпро" відноситься до цієї ділянки (або домену) CASB7439 білка. Модифікації 7 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Розкриття забезпечує білкові конструкції, що включають CASB7439 поліпептид, що включає принаймні одну модифікацію, що поліпшує експресію. У деяких втіленнях частина CASB7439 поліпептиду видаляється, при цьому вказана частина включає усю або частину ділянки, збагаченої на пролін. У деяких втіленнях ціла C-термінальна частина CASB7439 поліпептиду видаляється, що приводить до одержання вкороченого CASB7439 поліпептиду, який включає амінокислотні залишки 1-117 послідовності SEQ ID NO:13. У деяких втіленнях гетерологічний поліпептид розміщується N-термінально стосовно CASB7439 поліпептиду. Як використовується в даній заявці, "поліпшення експресії" або "поліпшена експресія" у контексті модифікованого CASB7439 поліпептиду відноситься до модифікації, призначеної для підвищення рівнів експресії білкових конструкцій, що включають вказаний модифікований CASB7439 поліпептид у порівнянні із експресією іншої еквівалентної білкової конструкції, що включає немодифікований CASB7439 поліпептид послідовності SEQ ID NO:13, тобто, такий самий вектор, хазяйські клітини, тощо. Будь-який зі способів, що є відомими у галузі техніки, для визначення кількості білка, є прийнятними для визначення, чи буде дана конструкція, яка включає модифікований CASB7439 поліпептид, мати підвищений рівень експресії у порівнянні із іншою еквівалентною білковою конструкцією, що включає немодифікований CASB7439 поліпептид. У деяких втіленнях продукція може бути оцінена при використанні забарвленого за Кумассі SDS-ПАГЕ. Видалення ділянки, збагаченої на пролін У деяких втіленнях модифікація CASB7439 поліпептиду включає делецію частини ділянки, збагаченої на пролін. У деяких втіленнях модифікація CASB7439 поліпептиду включає делецію всієї ділянки, збагаченої на пролін. У деяких втіленнях модифікація CASB7439 поліпептиду включає делецію амінокислот за межами ділянки, збагаченої на пролін, на доповнення до делеції ділянки, збагаченої на пролін. У деяких втіленнях модифікацію здійснюють шляхом вкорочення C-термінальної частини SEQ ID NO: 13. Tаким чином, у деяких втіленнях одержують білкову конструкцію, в якій Cтермінальний амінокислотний залишок модифікованого CASB7439 поліпептиду відповідає залишку, розміщеному на N-термінальному кінці ділянки, збагаченої на пролін послідовності SEQ ID NO: 13. У деяких втіленнях одержують білкову конструкцію, в якій C-термінальний амінокислотний залишок модифікованого CASB7439 поліпептиду відповідає залишку, розміщеному у межах ділянки, збагаченої на пролін послідовності SEQ ID NO:13. У деяких втіленнях C-термінальний амінокислотний залишок модифікованого CASB7439 поліпептиду відповідає амінокислотному залишку 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151 або 152 послідовності SEQ ID NO:13. У деяких втіленнях модифікація включає делецію частини або всієї ділянки, збагаченої на пролін. У деяких втіленнях послідовність модифікованого CASB7439 поліпептиду відповідає послідовності SEQ ID NO:13, з якої 21 або більше суміжних амінокислотних залишків були делетовані. Ці 21 або більше суміжних залишків відповідають суміжній амінокислотній послідовності SEQ ID NO:13, починаючи з будь-якого із залишків 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151 або 152, включно. У додаткових втіленнях модифікована поліпептидна послідовність CASB7439 відповідає SEQ ID NO:13, з якої 21 суміжних амінокислотних залишків були делетовані, при цьому вказані 21 суміжних залишків починаються з будь-якого із залишків 131, 132 та 133. У деяких втіленнях модифікована поліпептидна послідовність CASB7439 відповідає SEQ ID NO: 13, з якої один або більше залишків 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153 були делетовані. У деяких втіленнях модифікована поліпептидна послідовність CASB7439 відповідає SEQ ID NO: 13, з якої два або більше суміжних амінокислотних залишків були делетовані у ділянці амінокислотних залишків 133-153, включно. У деяких втіленнях модифікована поліпептидна послідовність CASB7439 відповідає SEQ ID NO:13, з якої залишки від 133 дo 153 включно послідовності SEQ ID NO: 13 були делетовані. Приклади необмежуючих втілень включають наступні: - LVL055, SEQ ID NO:1 (aa); - LVL111, SEQ ID NO:3 (aa); - LVL137, SEQ ID NO:5 (aa); - LVL141, SEQ ID NO:7 (aa); - LVL144, SEQ ID NO:9 (aa); та - LVL168, SEQ ID NO:11 (aa). 8 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Гетерологічні послідовності У деяких втіленнях модифікований CASB7439 поліпептид поєднують з гетерологічним поліпептидом. Термін "гетерологічний" стосовно молекули нуклеїнової кислоти, поліпептиду або іншого клітинного компоненту вказує на те, що компонент існує там, де його в нормі не виявляють у природі, та/або те, що він має походження від відмінного джерела або видів, ніж згадана молекула. Як використовується в даній заявці, "гетерологічні" молекули включають без обмеження гібридні білки, білки носія та мітки для очищення. У деяких втіленнях гетерологічний поліпептид може бути хімічно кон‗югований з модифікованим CASB7439 поліпептидом, тобто з білком носія. В інших втіленнях гетерологічний поліпептид та модифікований CASB7439 поліпептид може бути експресований як один рекомбінантний гібридний білок. В інших втіленнях модифікований CASB7439 поліпептид та гетерологічний поліпептид експресуються у вигляді одного рекомбінантного гібридного білка та хімічного кон‗югованого з іншим гетерологічним поліпептидом. Гетерологічний поліпептид може допомагати у забезпеченні T-хелперних епітопів, включаючи T-хелперні епітопи, що впізнаются людиною. У випадку білкової конструкції, що включає CASB7439 поліпептид та гетерологічний поліпептид, що є гібридним білком, гетерологічний поліпептид (партнер злиття) можуть допомагати у забезпеченні таких епітопів або допомагати в експресії білка (енхансер експресії) з більш високим виходом, ніж нативного рекомбінантного білка. Партнер злиття може являти собою як імунологічний партнер злиття, так і партнер, що покращує експресію. Білки носія Білок носія є хімічно кон‗югованим з іншим поліпептидом, що представляє інтерес. Хімічне злиття поліпептиду з білком носія може бути здійснене будь-яким із способів, відомих у галузі техніки. Таким чином, для безпосереднього ковалентного зв‗язування є можливим застосовувати карбодіімід, глутаральдегід або N-[γ-малеїмідобутирилокси]сукцинімідний естер, використовуючи традиційні комерційно доступні гетеробіфункціональні лінкери, такі, як CDAP та SPDP (при застосуванні інструкцій виробника). Після реакції злиття кон‗югат поліпептид-білок носія може бути легко ізольований та очищений за допомогою способу діалізу, методу гельфільтрації, способу фракціонування, тощо. Типи білків носія, що використовуються у деяких втіленнях, будуть відомими кваліфікованому спеціалісту у даній галузі техніки. Функція деяких білків носія полягає у забезпеченні цитокінової допомоги для того, щоб допомогти індукувати імунну відповідь проти злитого поліпептиду. Наприклад, невичерпний список білків носія, які можуть використовуватися у даному винаході, включає: гемоціанін лімфи равлика (KLH), сироваткові альбуміни, такі, як альбумін бичачої сироватки (BSA), інактивовані бактеріальні токсини, такі, як токсини правця та дифтерії (TT та DT), або їх рекомбінантні фрагменти (наприклад, домен 1 фрагменту C TT, або домен транслокації DT), або очищені похідні білка туберкуліну (PPD). Партнери злиття Незважаючи на доступність численних систем для злиття генів, експресія рекомбінантного білка в Escherichia coli залишається складною. Формування партнера злиття, що найкращим чином поліпшує експресію складних для експресії білків, залишається емпіричним. Звичайні партнери злиття включають маркерні послідовності C-термінального кінця білка для зв‗язування мальтози (MBP), глутатіон S-трансферази (GST), тіоредоксину (TRX), NUS A, убіквітину (Ub) та подібного до убіквітину модифікатора невеликого розміру (SUMO), кожна із цих послідовностей є описаною у літературі. Див., наприклад, Marblestone (2006) Prot. Sci. 15:182-7; Hunt (2005) Protein Exp. & Purif 40:1-22; Hammarstom та ін. (2002) Protein Sci. 11:313321. Імуногенні партнери злиття включають білок D з Haemophilus influenza B та неструктурний білjк з вірусу грипу, NS1 (гемаглютинін). Інший імунологічний партнер злиття являє собою білок, відомий як LYTA. При цьому може використовуватися C-термінальна частина такої молекули. LYTA походить від Streptococcus pneumoniae, що синтезує N-ацетил-L-аланін амідазу, амідазу LYTA, (кодується lytA геном, Garcia та ін. (1986) Gene 43:265-272). Є можливим використовувати повторювану частину молекули Lyta, що виявлена у C-термінальному кінці, починаючи від залишка 178, наприклад, залишки 188-305. Інший прийнятний партнер злиття являє собою білок аденілат циклази (CyaA) або його фрагмент, де вказаний фрагмент CyaA зберігає властивість вказаного білка аденілат циклази стосовно клітин, що презентують антиген. Див. W02005089792. В одному втіленні винаходу гетерологічний поліпептид являє собою білок D з Haemophilus influenzae (EP 0 594 610 B1), або його фрагмент. Білок D представляє собою IgD-зв‗язувальний білок з Haemophilus influenzae (WO 91/18926, виданий EP 0 594 610 B1). У деяких випадках, 9 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 наприклад, в експресійних системах рекомбінантного імуногену може бути бажаним використовувати N-термінальні фрагменти білка D, наприклад, такі, що включають від приблизно 100 до приблизно 110 N-термінальних амінокислот білка D (GB 9717953.5) (pD1/3, SEQ ID NO:39). Білок D H. influenzae синтезується як попередник з 18 амінокислотною сигнальною послідовністю (SEQ ID NO:41, див також номер доступу AAA24998). Коли сигнальна послідовність піддається обробці під час секреції, залишок цистеїну у положенні 19 у молекулі попередника стає амінотермінальним залишком. В одному втіленні гетерологічна поліпептидна послідовність включає першу частину, що містить приблизно одну третину перетвореної молекули білка D H. influenzae або N- термінальні 100-110 амінокислот перетвореного білка D. В одному втіленні гетерологічний білок включає амінокислотні залишки Met-Asp-Pro, приєднані до 109 N-термінальних амінокислот перетвореного білка D. В іншому втіленні гетерологічний білок включає амінокислотні залишки Met-Asp-Pro, приєднані до амінокислотних залишків 2-109 перетвореного білка D (SEQ ID NO:39). В одному втіленні, модифікований CASB7439 поліпептид або білкова конструкція є хімічно злитою з CyaA білком або його фрагментом. Див. W02005054851. В іншому втіленні модифікований CASB7439 поліпептид або білкова конструкція є хімічно приєднаною до B субодиниці токсину Шига або його імунологічно функціонального еквіваленту. Див. патент США №6,613,882; W002060937; W02005112991. Полігістидинові мітки, чужорідні амінокислоти Часто є бажаним включати гетерологічний поліпептид, що містить секреторну або лідерну послідовності, про-послідовності, послідовності, які сприяють очищенню, такі, як гістидинова мітка, наприклад, полігістидинова послідовність (включає численні залишки гістидину), або додаткові послідовності для стабільності під час рекомбінантного процесу одержання. Вектори гістидинової мітки та набори є комерційно доступними. Наприклад, вектори для додання шести гістидинових міток до поліпептиду є доступними від Roche. Набори для одержання та застосування мічених гістидином поліпептидів є доступними від Qiagen, Sigma, Thermo Scientific, GE Healthcare та інших. Після гістидинової мітки може також йти прийнятна амінокислотна послідовність, що сприяє видаленню полігістидинової послідовності при використанні ендопептидаз. Екзопептидаза, наприклад, Qiagen TAGZyme, може використовуватися для видалення термінальної полігістидинової послідовності без додаткової послідовності. У деяких втіленнях полігістидинова послідовність є розміщеною у N-термінальній ділянці білкової конструкції. У додаткових втіленнях полігістидинова послідовність є розміщеною на Nтермінальному кінці білкової конструкції. У деяких втіленнях полігістидинова послідовність включає 6, 7, 8, 9, 10, 11, 12, 13, 14, 15 або більше послідовних гістидинів. У додаткових втіленнях полігістидинова послідовність включає десять послідовних гістидинів. У деяких втіленнях гетерологічний поліпептид включає полігістидинову послідовність та одну або більше чужорідних амінокислот. Під "чужорідними амінокислотами" розуміють одну або більше амінокислот, що не є частиною полігістидинової послідовності або гібридного білка, та не є природною частиною CASB7439 поліпептидної послідовності. Наприклад, чужорідні амінокислоти можуть бути такими, що включаються для забезпечення сайту пептидази, вони можуть просто бути сторонньою послідовністю, такою, як артефакт клонування, або подібною до неї. Молекули нуклеїнової кислоти, що кодують casb7439 конструкції Інші втілення цього розкриття стосуються рекомбінантних нуклеїнових кислот, які кодують модифіковані CASB7439 поліпептиди та білкові конструкції, як описано в даній заявці. У деяких втіленнях рекомбінантні нуклеїнові кислоти є оптимізованими за кодонами для експресії у вибраних прокаріотичних або еаукаріотичних хазяйських клітинах. Для сприяння реплікації та експресії нуклеїнові кислоти можуть бути введені у вектор, такий, як прокаріотичний або еукаріотичний експресійний вектор. Хазяйські клітини, що включають рекомбінантні модифіковані CASB7439 поліпептиди або білкові конструкції, які кодуються нуклеїновими кислотами, також представляють собою ознаки даного розкриття. Бажані хазяйські клітини включають прокаріотичні (тобто, бактеріальні) хазяйські клітини, такі, як E. coli, а також численні еукаріотичні хазяйські клітини, включаючи грибкові (наприклад, дріжджові) клітини, клітини комах та клітини ссавців (такі, як HEK293, CHO та VERO клітини). Експресійні вектори Для сприяння реплікації та експресії нуклеїнові кислоти можуть бути введені у вектор, такий, як прокаріотичний або еукаріотичний експресійний вектор. Незважаючи на те, що нуклеїнові 10 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 кислоти, розкриті в даній заявці, можуть бути включені у будь-який один з різноманітних векторів (включаючи, наприклад, бактеріальні плазміди; фагову ДНК; бакуловірус; дріжджові плазміди; вектори, що походять від комбінацій плазмід та фагової ДНК, вірусної ДНК, такої, як осповакцини, аденовірусу, вірусу віспи птахів, псевдосказу, аденовірусу, аденоасоційованого вірусу, ретровірусу та багатьох інших), у найбільш загальному випадку вектор буде являти собою експресійний вектор, прийнятний для одержання поліпептидних продуктів експресії. В експресійному векторі нуклеїнова кислота, що кодує білкову конструкцію, типово розміщується поблизу та в орієнтації до прийнятної послідовності контролю транскрипції (промотор та необов‗язково, один або більше енхансерів) для направлення синтезу іРНК. Тобто, полінуклеотидна послідовність, що представляє інтерес, є оперативно зв‗язаною з прийнятною послідовністю контролю транскрипції. Приклади таких промоторів включають: передранній промотор CMV, LTR або SV40 промотор, поліедровий промотор бакуловірусу, промотор lac або trp E. coli, промотор PL фага T7 та лямбда та інші промотори, відомі для контролю експресії генів у прокаріотичних та еукаріотичних клітинах або їх вірусів. Експресійний вектор типово також містить сайт зв‘язування рибосоми для ініціації трансляції та термінатор транскрипції. Вектор необов‗язково включає прийнятні послідовності для ампліфікації експресії. Крім того, експресійні вектори необов‗язково включають один або більше генів селективних маркерів для забезпечення фенотипічної ознаки для селекції трансформованих хазяйських клітин, такі, як наявність дигідрофолатредуктази або стійкість до неоміцину для культури еукаріотичних клітин, або такі, як стійкість до тетраціикліну або ампіцилліну в E. coli. Типові процедури, достатні для того, щоб забезпечити керівництво середньому спеціалістові у галузі техніки стосовно продукції рекомбінантних нуклеїнових кислот CASB7439, можуть бути знайдені у Sambrook та ін., Molecular Cloning: A Laboratory Manual, 2-е вид., Cold Spring Harbor Laboratory Press, 1989; Sambrook та ін., Molecular Cloning: A Laboratory Manual, 3-є вид., Cold Spring Harbor Press, 2001; Ausubel та ін., Current Protocols in Molecular Biology, Greene Publishing Associates, 1992 (та Доповнення до 2003); та Ausubel та ін., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, 4-е вид., Wiley & Sons, 1999. Типові молекули нуклеїнової кислоти, що кодують білкові конструкції, є представленими послідовностями SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO: 10 та SEQ ID NO: 12. Додаткові варіанти послідовності, які мають ідентичність послідовності з типовою молекулою нуклеїнової кислоти, можуть бути одержані кваліфікованим спеціалістом у даній галузі техніки. Типово, варіанти нуклеїнової кислота будуть кодувати поліпептиди, які відрізняються не більше, ніж на 1% або 2%, або 5%, або 10%, або 15%, або 20% амінокислотних залишків. Тобто, кодовані поліпептиди мають принаймні 80% або 85%, головним чином, принаймні приблизно 90% або більше, наприклад, 95%, 98%, 99%,або 99,5% ідентичності послідовності. Кваліфікованому спеціалістові у даній галузі техніки буде зрозумілим, що полінуклеотидні послідовності, які кодують білкові конструкції, як такі можуть мати меншу ідентичність послідовності завдяки виродженості генетичного коду. У деяких випадках модифікований CASB7439 поліпептид або білкова конструкція має одну або більше амінокислотних модифікацій у порівнянні з амінокислотною послідовністю типової конструкції, що представлена в даній заявці (наприклад, на доповнення до згаданих вище модифікацій). Такі відмінності можуть бути доповненням, делецією або заміщенням одного або більше нуклеотидів або амінокислот, відповідно. Варіант типово відрізняється не більше, ніж приблизно на 1% або 2%, або 5%, або 10%, або 15%, або 20%, або 25%, або 30% нуклеотидних залишків. Наприклад, варіант CASB7439 поліпептиду або білкової конструкції, що кодується нуклеїновою кислотою, може включати 1 або 2, або аж до 5, або аж до приблизно 10, або аж до приблизно 15, або аж до приблизно 50, або аж до приблизно 100 нуклеотидних відмінностей у порівнянні із типовим CASB7439 поліпептидом або білковою конструкцією, що кодується нуклеїновими кислотами послідовностей SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID N0:10 та SEQ ID NO:12. Таким чином, варіант у контексті CASB7439 поліпептиду або білкової конструкції, що кодується нуклеїновою кислотою, типово має принаймні 70% або 75%, або 80%, або 85%, або 90%, або 95%, або 98%, або 99%, або 99,5% ідентичності послідовності із референтною послідовністю, наприклад, референтною послідовністю, проілюстрованою послідовностями SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID N0:10 та SEQ ID NO:12, або будь-яким іншим типовим CASB7439 поліпептидом, білковою конструкцією, що кодуються нуклеїновими кислотами, розкритими в даній заявці. Додаткові варіанти можуть виникати внаслідок випадкового поширення генетичних мутацій у популяції, або можуть бути одержані синтетичним способом при використанні сайт направленого або випадкового мутагенезу, або шляхом рекомбінації двох або більше попередньо існуючих варіантів. Такі 11 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 додаткові варіанти є також прийнятними у контексті CASB7439 поліпептиду або білкових конструкцій, розкритих в даній заявці. На доповнення до раніше описаного варіанту нуклеїнової кислоти, нуклеїнові кислоти, які можуть гібридизуватися з однією або більше типових нуклеїнових кислот, представлених послідовностями SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID N0:10 та SEQ ID NO:12, можуть також використовуватися для кодування CASB7439 поліпептидів або білкових конструкцій, розкритих в даній заявці. Кваліфікований спеціаліст у даній галузі техніки, зможе оцінити, що на доповнення до показника проценту (%) ідентичності послідовності, що обговорюється у даній заявці, інший показник подібності послідовностей між двома нуклеїновими кислотами є показником здатності до гібридизації. Більш подібними є послідовності двох нуклеїнових кислот, якщо більш жорсткими є умови, за яких вони будуть гібридизуватися. Умови жорсткості гібридизації є залежними від послідовності та є різними при різних параметрах навколишнього середовища. Tаким чином, умови гібридизації, що приводять до конкретних ступенів жорсткості, будуть варіювати у залежності від природи вибраного способу гібридизації, композиції та довжини послідовності нуклеїнової кислоти, що гібридизується. + В загальному випадку, температура гібридизації та іонна сила (зокрема, концентрація Na ++ та/або Mg ) гібридизаційного буфера будуть визначати жорсткість гібридизації, незважаючи на те, що кратність промивання також впливає на жорсткість. В загальному випадку жорсткі умови вибирають як такі, що є приблизно на 5°C - 20°C нижчими за температурну точку плавлення (Tm) для специфічної послідовності при визначеній іонній силі та pH. T m являє собою температуру (при визначеній іонній силі та pH), при якій 50% цільової послідовності гібридизується з ідеально підібраним зондом. Умови для гібридизації нуклеїнової кислоти та підрахунок жорсткості можуть бути знайдені, наприклад, у Sambrook та ін., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 2001; Tijssen, Hybridization With Nucleic Acid Probes, Part I: Theory and Nucleic Acid Preparation, Laboratory Techniques in Biochemistry and Molecular Biology, Elsevier Science Ltd., NY, NY, 1993 та Ausubel -е та ін. Short Protocols in Molecular Biology, 4 вид., John Wiley & Sons, Inc., 1999. Для цілей даного розкриття "жорсткі умови" охоплюють умови, за яких буде відбуватися гібридизація, якщо існує менше, ніж 25% невідповідності між гібридизаційними молекулами та цільовою послідовністю. "Жорсткі умови" можуть бути розділені на декілька рівнів жорсткості для більш точного визначення. Таким чином, як використовується в даній заявці, умови "помірної жорсткості" є такими, за яких молекули, які мають більше 25% невідповідності послідовності, не будуть гібридизуватися; умови "середньої жорсткості " є такими, за яких молекули, які мають більше 15% невідповідності, не будуть гібридизуватися, та умови "високої жорсткості" є такими, за яких послідовності, які мають більше 10% невідповідності, не будуть гібридизуватися. Умови "дуже високої жорсткості" " є такими, за яких молекули, які мають більше 6% невідповідності, не будуть гібридизуватися. На противагу до цього, нуклеїнові кислоти, які гібридизуються за "умов низької жорсткості " включають такі з набагато меншою ідентичністю послідовності, або з ідентичністю послідовності тільки на короткій субпослідовності нуклеїнової кислоти. Таким чином, буде зрозумілим, що різноманітні варіанти нуклеїнових кислот, що охоплюються цим розкриттям, є здатними гібридизуватися при жорстких умовах принаймні з однією із послідовностей SEQ ID NO:2, SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID N0:10 та SEQ ID NO:12 суттєво по усій довжині. Продукція білкових конструкцій Білкові конструкції, розкриті в даній заявці, можуть бути одержані при використанні добре відомих процедур для експресії та очистки рекомбінантних білків. Процедури, достатні для того, щоб забезпечити керівництво для середнього спеціаліста у галузі техніки можуть бути знайдені у: Sambrook та ін., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, -е Cold Spring Harbor, NY, 200; та Ausubel та ін. Short Protocols in Molecular Biology, 4 вид., John Wiley & Sons, Inc., 999. Додаткові та специфічні деталі забезпечуються в даній заявці нижче. Рекомбінантні нуклеїнові кислоти, які кодують білкові конструкції, можуть бути введені у хазяйські клітини за допомогою будь-якої з різноманітності відомих процедур, таких, як електропорація, опосередкована ліпосомами трансфекція (наприклад, при використанні комерційного доступного реагенту для ліпосомальної трансфекції, такого, як LIPOFECTAMINE™2000 або TRANSFECTIN™), преципітація при використанні фосфату кальцію, інфекція, трансфекція та подібні до них, у залежності від вибору векторів та хазяйських клітин. Хазяйські клітини можуть культивуватися у традиційному поживному середовищі, модифікованому, як є прийнятним для активації промоторів, селекції трансформантів або 12 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 ампліфікації вбудованих полінуклеотидних послідовностей. Умови культивування, такі, як температура, pH та подібні до них, є типово такими, які раніше використовувалися для хазяйських клітин, вибраних для експресії, та які будуть зрозумілими кваліфікованому спеціалістові у даній галузі техніки та які згадуються у посиланнях, приведених у даній заявці, включаючи, наприклад, Freshney (1994) Culture of Animal Cells, a Manual of Basic Technique, третє видання, Wiley-Liss, New York та посиланнях, які там наводяться. У деяких втіленнях прийнятна температура для культивування складає 16°C; в інших випадках 37°C. Продукти експресії, що відповідають нуклеїновим кислотам згідно з винаходом, можуть також бути одержані у клітинах нетваринного походження, таких, як рослини, дріжджі, гриби, бактерії та подібні до них. На доповнення до Sambrook, Berger та Ausubel, деталі стосовно культури тканин можуть бути знайдені у Payne та ін. (1992) Plant Cells and Tissue Culture in Liquid Systems John Wiley & Sons, Inc. New York, NY; Gamborg та Phillips (ред.) (1995) Plant Cells, Tissue and Organ Culture; Fundamental Methods Springer Lab Manual, Springer-Verlag (Berlin Heidelberg New York) та Atlas та Parks (ред.) The Handbook of Microbiological Media (1993) CRC Press, Boca Raton, FL. Хазяйські клітини Хазяйські клітини, що включають нуклеїнові кислоти, які кодують рекомбінантну білкову конструкцію, також являють собою, ознаку даного розкриття. Бажані хазяйські клітини включають прокаріотичні (тобто, бактеріальні) хазяйські клітини, такі, як E. coli, а також численні еукаріотичні хазяйські клітини, включаючи клітини грибів (наприклад, дріжджі, такі, як Saccharomyces cerevisiae та Picchia pastoris), клітини комах, рослинні клітини та клітини ссавців (такі, як CHO клітини). Нуклеїнові кислоти рекомбінантної білкової конструкції вводяться (наприклад, шляхом трансдукції, трансформації або трансфекції) у хазяйські клітини, наприклад, за допомогою вектора, такого, як експресійний вектор. Як описано в даній заявці, вектор найбільш типово являє собою плазміду, проте такі вектори можуть також представляти собою, наприклад, вірусні частинки, фаг, тощо. Приклади прийнятних хазяйських організмів для експресії включають: бактеріальні клітини, такі, як E. coli, Streptomyces та Salmonella typhimurium; клітини грибів, такі, як Saccharomyces cerevisiae, Pichia pastoris та Neurospora crassa; клітини комах, такі, як Drosophila та Spodoptera frugiperda; клітини ссавців, такі, як 3T3, COS, CHO, BHK, HEK293 або Bowes melanoma; рослинні клітини, включаючи клітини водоростей, тощо. Хазяйські клітини є необов‗язково вибраними за їх здатністю модулювати експресію вбудованої послідовності або піддавати процесингу експресований білок бажаним чином. Такі модифікації білка включають, але без обмеження, глікозилювання, (а також, наприклад, ацетилування, карбоксилування, фосфорилювання, ліпідизацію та ацилування). Посттрансляційний процесинг, наприклад, такий, що розщеплює форму попередника з одержанням форми зрілого білка (наприклад, за допомогою фуринпротеази) необов‗язково здійснюють у контексті хазяйських клітин. Різноманітні хазяйські клітини, такі, як 3T3, COS, CHO, HeLa, BHK, MDCK, HEK293, WI38, тощо, мають специфічний клітинний механізм та характерні механізми для таких пост-трансляційних активностей та можуть бути вибрані для забезпечення правильної модифікації та процесингу вбудованого чужорідного білка. Для тривалої продукції з високим виходом рекомбінантних білкових конструкцій, розкритих в даній заявці, типово використовуються стабільні експресійні системи. Наприклад, молекули нуклеїнової кислоти, що стабільно експресують білкову конструкцію, вводяться у хазяйські клітин при використанні експресійних векторів, що містять вірусну точку початку реплікації або eндогенні експресійні елементи та ген селективного маркера. Після введення вектора клітини залишають для вирощування протягом 1-2 днів у збагаченому середовищі перед перенесенням їх на селективне середовище. Мета селективного маркера полягає у наданні резистентності до селекції, присутність селективного маркера дозволяє здійснювати ріст та відновлення клітин, які успішно експресують введені послідовності. Наприклад, стійкі групи або колонії стабільно трансформованих клітин можуть розмножуватися при використанні методик тканинної культури, прийнятних для типу клітин. Хазяйські клітини, трансформовані за допомогою нуклеїнової кислоти, що кодує білкову конструкцію, необов‗язково культивують за умов, прийнятних для експресії та вилучення кодованого білка з клітинної культури. Eкспресовані білкові конструкції можуть бути вилучені та очищені з культур рекомбінантних клітин за допомогою будь-якого із численних способів, що є добре відомими у галузі техніки, включаючи осадження сульфатом амонію або етанолом, кислотну екстракцію, фільтрування, ультрафільтрацію, центрифугування, аніонообмінну або катіонообмінну хроматографію, хроматографію на основі фосфоцелюлози, хроматографію на основі гідрофобної взаємодії, афінну хроматографію (наприклад, при використанні будь-якої із систем мічення, що зазначені у даній заявці), хроматографію на основі гідроксіапатиту та хроматографію на основі лектину. 13 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Етапи рефолдінгу білка можуть використовуватися, як це є бажаним, у завершенні формування конфігурації зрілого білка. Крім того, високоефективна рідинна хроматографія (ВЕРХ) може використовуватися на заключних етапах очищення. На доповнення до посилань, зазначених у даній заявці, різноманітні способи очистки є добре відомими у галузі техніки, включаючи, наприклад, ті, що описані у Sandana (1997) Bioseparation of Proteins, Academic Press, Inc.; та Bollag та ін. (1996) Protein Methods, 2-е вид., Wiley-Liss, NY; Walker (1996) The Protein Protocols Handbook Humana Press, NJ, Harris та Angal (1990) Protein Purification Applications: A Practical Approach IRL Press at Oxford, Oxford, U.K.; Scopes (1993) Protein Purification: Principles and Practice 3-є вид. Springer Verlag, NY; Janson та Ryden (1998) Protein Purification: Principles, High Resolution Methods and Applications, 2-е вид. Wiley-VCH, NY; та Walker (1998) Protein Protocols on CD-ROM Humana Press, NJ. У деяких прикладах нуклеїнові кислоти вводяться у клітини за допомогою векторів, прийнятних для введення та експресії у прокаріотичних клітинах, наприклад, клітинах E. coli. Наприклад, нуклеїнова кислота, що включає полінуклеотидну послідовність, яка кодує білкову конструкцію, може бути введена у будь-який з різноманітних комерційно доступних або індивідуально одержаних векторів, таких, як pET серії експресійних векторів (наприклад, pET9b та pET2d). Експресія кодуючої послідовності індукується за допомогою ізопропіл п-D-1тіогалактопіранозиду (IPTG), що приводить до високих рівнів білкової експресії. Полінуклеотидна послідовність, що кодує CASB7439 білкову конструкцію транскрибується під контролем промотора фага T7. Aльтернативні вектори, такі, як pURV22, що включає промотор pL фага лямба, що індукується нагріванням, також є прийнятними. Експресійний вектор вводять (наприклад, шляхом електропорації) у прийнятних бактеріальних хазяїв. Численні прийнятні штами E. coli є доступними та можуть бути відібрані спеціалістом у даній галузі техніки (наприклад, BLR DE3, BL21 DE3 та Rosetta DE3 штами були схвалені як сприятливі для експресії рекомбінантних векторів, що містять полінуклеотидні послідовності, які кодують білкові конструкції). Необов‗язково, полінуклеотиди, які кодують білкові конструкції, вбудовують в експресійні вектори, які є прийнятними для введення та експресії в еукаріотичних організмах (наприклад, клітинах комах та ссавців). Є бажаним, коли такі нуклеїнові кислоти є оптимізованими за кодонами для експресії у вибраних векторах/хазяйських клітинах. Відібрані клітини можуть клонально розмножуватися та характеризуватися для експресії бажаної білкової конструкції. Методики для оптимізації кодонів є відомими у галузі техніки. Крім того, постачальники комерційних послуг пропонують оптимізацію кодонів, серед інших звичайних технічних послуг. Прокаріотичні організми У бактеріальних системах може бути вибраний ряд експресійних векторів у залежності від призначеного застосування для експресованого продукту. Наприклад, коли є необхідними великі кількості поліпептиду або його фрагментів для продукції антитіл, бажаним чином використовуються вектори, що керують високим рівнем експресії гібридних білків, які піддаються легкій очистці. Такі вектори включають, але без обмеження, мультифункціональні клонуючі та експресійні вектори E. coli, такі, як BLUESCRIPT (Stratagene), в яких кодуюча послідовність, що представляє інтерес, наприклад, полінуклеотиди згідно з винаходом, як описано в даній заявці, можуть лігуватися у вектор у рамці з послідовностями для амінотермінальної трансляції, ініціюючи метіонін та наступні 7 залишків бета-галактозидази, з продукцією каталітично активного гібридного білка бета-галактозидази; pIN вектори (Van Heeke & Schuster (1989) J Biol Chem 264:5503-5509); pET вектори (Novagen, Madison Wl), в яких амінотермінальний метіонін лігують у рамці з гістидиновою міткою; та подібні до них. У деяких втіленнях прийнятний вектор являє собою pET19; в інших - pET24; в інших - pET26. Еукаріотичні організми Подібно до цього, у дріжджах таких, як Saccharomyces cerevisiae, ряд векторів, що містять конститутивні або індуцибельні промотори, такі, як альфа фактор, алкоголь оксидаза та PGH, можуть використовуватися для продукції бажаних продуктів експресії. Для огляду, див. Berger, Ausubel, та, наприклад, Grant та ін. (1987; Methods in Enzymology 153:516-544). У хазяйських клітинах ссавців може використовуватися ряд експресійних систем, включаючи як плазміди, так і системи на основі вірусів. ІМУНОГЕННІ КОМПОЗИЦІЇ Фармацевтично прийнятні носії або наповнювачі Фармацевтично прийнятні носії або наповнювачі є добре відомими та можуть бути вибрані кваліфікованим спеціалістом у даній галузі техніки. Наприклад, носій або наповнювач може бажаним чином включати буфер. Необов‗язково, носій або наповнювач також містить принаймні один компонент, що стабілізує розчинність та/або стабільність. Приклади 14 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 солюбілізувальних/стабілізувальних агентів включають детергенти, наприклад, лаурилсаркозин та/або Твін. Альтернативні солюбілізувальні/стабілізувальні агенти включають аргінін, та склоутворювальні поліоли (такі, як сахароза, трегалоза та подібні до них). Численні фармацевтично прийнятні носії та/або фармацевтично прийнятні наповнювачі є відомими у галузі техніки та є розкритими, наприклад, у Remington's Pharmaceutical Sciences, by E. W. Martin, Mack Publishing Co., Easton, PA, 5-е вид. (975). Згідно з цим прийнятні наповнювачі та носії можуть бути вибрані спеціалістом у даній галузі техніки для одержання композиції, прийнятної для доставки суб‗єкту вибраним шляхом введення. Прийнятні наповнювачі включають, але без обмеження, гліцерин, поліетиленгліколь (ПЕГ), сорбіт, трегалозу, сіль N-лауроїлсаркозину натрію, L-пролін, сульфобетаїн, який не має 2+ 2+ поверхневої активності, гуанідин гідрохлорид, сечовину, триметиламін оксид, KCI, Ca , Mg , 2+ 2+ Mn , Zn та інші споріднені солі дивалентних катіонів, дитіотреїтол, та β-меркаптоетанол. Інші наповнювачі можуть являти собою детергенти, включаючи: Твін 80, Твін 20, Тритон X-00, NP40, Емпіген BB, Oктилглікозид, лауроїлмальтозид, Zwittergent 3-08, Zwittergent 3-0, Zwittergent 32, Zwittergent 3-4, Zwittergent 3-6, CHAPS, дезокисхолат натрію, додецилсульфат натрію, цетилтриметиламоній бромід). Ад‗юванти Необов‗язково, імуногенні композиції також включають ад‗ювант. У контексті імуногенної композиції прийнятні для введення суб‗єкту, такому, як суб‗єкт, який являє собою людину, для цілей стимуляції iмунної відповіді проти CASB7439, ад‗ювант є вибраним для стимуляції імунної відповіді Th1 типу. Ад‗ювант є типово вибраним для поліпшення імунної відповіді Th1 типу у суб‗єкта або у популяції суб‗єктів, яким вводиться композиція. Th1 імунна відповідь Імунна відповідь "Th1" типу характеризується індукцією CD4+ T-хелперних клітин, що продукують IL-2 та IFN-γ. На противагу до цього імунна відповідь "Th2" типу характеризується індукцією CD4+ T-хелперних клітин, що продукують IL-4, IL-5 та IL-13. TLR ефектори, включаючи без обмеження 3D-MPL Один прийнятний ад‗ювант для застосування у комбінації з модифікованими CASB7439 поліпептидами являє собою TLR-4-модулятор. Oдин приклад являє собою нетоксичну похідну ліпіду A, таку, як монофосфорил ліпід A або, зокрема, 3-деацилований монофосфорил ліпід A (3D-MPL). 3D-MPL продається під назвою MPL GlaxoSmithKline Biologicals S.A., та називається у цьому документі MPL або 3D-MPL. Див., наприклад, патенти США №№4,436,727; 4,877,611; 4,866,034 та 4,912,094. 3D-MPL перш за все стимулює CD4+ T-клітинні відповіді з IFN-γ (Th1) фенотипом. 3D-MPL може бути одержаний у відповідності зі способами, розкритими в GB2220211 A. З хімічної точки зору він являє собою суміш 3-деацилованого монофосфорил ліпіду A з 3, 4, 5 або 6 ацилованими ланцюгами. У композиції згідно з даним винаходом може використовуватися 3D-MPL у вигляді невеликих частинок. 3D-MPL, що складається з невеликих частинок, має такий розмір частинок, що вони стерильно фільтруються через фільтр 0,22 мкм. Такі препарати є описаними у WО94/21292. В інших втіленнях ліпополісахарид може являти собою Р(1-6) глюкозамін дисахарид, який описується у патенті США № 6,005,099 та патенті ЕР 0 729 473 B1. Спеціаліст у даній галузі техніки зможе легко одержати різноманітні ліпополісахариди, такі, як 3D-MPL, на основі методик вказаних посилань. На доповнення до згаданих вище імуностимуляторів (що є подібними за структурою до LPS або MPL або 3D-MPL), ациловані похідні моносахариду та дисахариду, що є субчастиною структури MPL ,є також прийнятними як ад‗юванти. В інших втіленнях ад‗ювант являє собою синтетичну похідну ліпіду A, деякі з яких були описані як TLR-4 aгоністи та включають, але без обмеження: OM174 (2-дезокси-6-o-[2-дезокси-2-[(R)-3-додеканоїлoкситетрадеканоїламіно]-4-o-фосфоно-P-D-глюкопіранозил]-2-[(R)-3-гідрокситетрадеканоїламіно]-α-Dглюкопіранозилдигідроген фосфат) (WO 95/14026); OM 294 DP (3S, 9R)-3-[(R)додеканоїлокситетрадеконоїлaміно]-4-oксо-5-aзa-9(R)-[(R)-3-гідрокситетра деканоїлaміно]декан1,10-діол, 1,10-біс(дигідрогенфосфат) (WO 99/64301 та WO 00/0462 ); та OM 197 MP-Ac DP (3S,9R)-3-[(R)-додеканоїлокситетрадеканоїламіно]-4-oксo-5-aзa-9-[(R) -3гідрокситетрадеканоїламіно] декан-1,10-діол,1-дигідрогенфосфат 10-(6-аміногексаноат) (WO 01/46127). Інші TLR-4 ліганди, які можуть використовуватися, являють собою алкілглюкозамінід фосфати (AGP) такі, як ті, що розкриті у WO 98/50399 або патенті США № 6,303,347 (способи для одержання AGP є також розкритими), прийнятним чином RC527 або RC529, або фармацевтично прийнятні солі AGP, як розкрито у патенті США № 6,764,840. Деякі AGP 15 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 являють собою TLR-4 агоністи, а деякі являють собою TLR-4 антагоністи. Обидва передбачаються корисними як ад‗юванти. Інші прийнятні TLR-4 ліганди, що є здатними викликати сигнальну відповідь при посередництві TLR-4 (Sabroe т, Jl 2003 p1630-5), являють собою, наприклад, ліпополісахарид з грам-негативних бактерій та їх похідних, або їх фрагменти, зокрема, нетоксичну похідну LPS (таку, як 3D-MPL). Інші прийнятні TLR агоністи являють собою: білок теплового шоку (HSP) 10, 60, 65, 70, 75 або 90; поверхнево-активний білок A, гіалуронанові олігосахариди, фрагменти гепаранан сульфату, фрагменти фібронектину, пептиди фібриногену, b-дефенсин-2 та дипептид мурамілу (MDP). В одному втіленні TLR агоніст являє собою HSP 60, 70 або 90. Інші прийнятні TLR-4 ліганди являють собою такі, як описано у WO 2003/011223 та у WO 2003/099195, такі, як сполука I, сполука II та сполука III, розкриті на сторінках 4-5 у WО2003/011223, або на сторінках 3-4 WО2003/099195 та, зокрема, такі сполуки, як розкрито у WО2003/011223 під назвою ER803022, ER803058, ER803732, ER804053, ER804057, ER804058, ER804059, ER804442, ER804680 та ER804764. Наприклад, один прийнятний TLR-4 ліганд являє собою ER804057. Сапонінові ад‗юванти Ініш ад‗юванти, що можуть використовуватися в імуногенних композиціях з CASB7439, наприклад, як такі або у комбінації з 3D-MPL або іншим ад‗ювантом, описаним в даній заявці, являють собою сапоніни, такі, як QS21. Сапоніни є розкритими у: Lacaille-Dubois, M та Wagner H. (1996. A review of the biological and pharmacological activities of saponins. Phytomedicine том 2, стор. 363- 386). Сапоніни являють собою стероїдні або тритерпенові глікозиди, що є широко поширеними у царствах рослин та морських тварин. Сапоніни є відомими як такі, що утворюють колоїдні розчини у воді, які піняться при струшуванні, та є призначеними для осадження холестерину. Коли сапоніни перебувають поблизу мембран клітин, вони утворюють пороподібні структури у мембрані, які спричинюють прориви мембрани. Гемоліз еритроцитів являє собою приклад цього явища, що є притаманним деяким, але не усім, сапонінам. Сапоніни, зокрема, імунологічно активна фракція санонінів, є відомими як ад‗юванти у вакцинах для системного введення. Ад‗ювантна та гемолітична активність індивідуальних сапонінів широко вивчалася у галузі техніки (Lacaille-Dubois та Wagner, див. вище). Наприклад, Quil A (є одержаним з кори північно-американського дерева Quillaja Saponaria Molina) та його фракції є описаними у US 5,057,540 та "Сапоніни як вакцинні ад‗юванти", Kensil, C. R., Crit Rev Ther Drug Carrier Syst, 1996, 12 (1-2): 1-55; та EP 0 362 279 B1. Структури, що складаються з частинок, які називають імуностимулювальними комплексами (ISCOMS) та включають фракції Quil A, є гемолітиками та використовються у виробництві вакцин (Morein, B., EP 0 109 942 B1; WO 96/11711; WO 96/33739). Гемолітичні сапоніни QS21 та QS17 (очищені за допомогою ВЕРХ фракції Quil A) були описані як потужні системні ад‗юванти, а спосіб їх одержання є розкритим у патенті США № 5,057,540 та EP 0 362 279 B1. Інші сапоніни, що використовувалися у дослідженнях системної вакцинації, включають ті, що мають походження від видів рослин, таких, як Gypsophila та Saponaria (Bomford та ін., Vaccine, 10(9):572-577, 1992). Такі композиції що включають QS21 та холестерин, були продемонстровані як успішні ад‗юванти, що стимулюють Th1, коли рецептуються разом з антигеном. Таким чином, наприклад, CASB7439 може сприятливим чином використовуватися в імуногенних композиціях з ад‗ювантом, що включає комбінацію QS21 та холестерину. Імуностимуляторні олігoнуклеїнові кислоти Один прийнятний ад‗ювант для застосування у комбінації з CASB7439 являє собою TLR агоніст бактеріальної ДНК, здатний викликати сигнальну відповідь за допомогою TLR-9, тобто, TLR-9 агоніст, зокрема, ДНК, що містить неметильовані CpG нуклеотиди, особливо послідовність, яка є відомою як CpG мотив. Олігонуклеотиди, що містять CpG, індукують головним чином Th1 відповідь. Такі олігонуклеотиди є добре відомими та описані, наприклад, у WO 96/02555, WO 99/33488 та патентах США №№ 6,008,200 та 5,856,462. Прийнятним чином CpG нуклеотиди являють собою CpG олігонуклеотиди. Прийнятні олігонуклеотиди для застосування в імуногенних композиціях згідно з даним винаходом являють собою олігонуклеотиди, що містять CpG, зокрема, необов‗язково містять два або більше динуклеотидних CpG мотивів, відокремлених принаймні трьома, бажано принаймні шістьома або більше нуклеотидами. CpG мотив являє собою цитозиновий нуклеотид, за яким йде гуаніновий нуклеотид. CpG олігонуклеотиди згідно з даним винаходом типово являють собою дезоксинуклеотиди. У специфічному втіленні інтернуклеотид в олігoнуклеотиді являє собою фосфородитіоат, або бажано фосфородитіоатний зв‗язок, незважаючи на те, що фосфодiефірні та інші інтернуклеотидні зв‗язки знаходяться у межах об‗єму правової охорони даного винаходу. 16 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 У деяких втіленнях забезпечуються олігонуклеотиди зі змішаними інтернуклеотидними зв‗язками. Способи одержання фосфоротіоатних олігонуклеотидів або фосфородитіоату є описаними у патентах США №№ 5,666,153, 5,278,302 та WO 95/26204. В альтернативному втіленні використовується TLR агоніст, що є здатним викликати сигнальну відповідь за допомогою TLR-9. В одному втіленні TLR агоніст, що є здатним викликати сигнальну відповідь за допомогою TLR-9, являє собою HSP90. Комбінації різних ад‗ювантів, такі, як ті, що згадані в даній заявці, можуть також використовуватися у композиції з CASB7439. Наприклад, як вже було зазначено, QS21 може рецептуватися разом з 3D-MPL. Співвідношення QS21 : 3D-MPL буде типово складати від 1 : 10 дo 10 : 1; напркилад, від 1:5 дo 5 : 1, та частіше суттєво 1 : 1 . Типово, співвідношення знаходиться у межах від 2.5 : 1 дo 1 : 1 3D-MPL: QS21. Інша комбінація ад‗юванту являє собою 3DMPL плюс QS21 у ліпосомній композиції, а також у комбінації з CpG. Суб‗єкти з колоректальним раком Вибір суб‗єкта з колоректальним раком для участі у способах, описаних в даній заявці, може здійснюватися за допомогою клінічного діагнозу, молекулярного діагнозу або їх комбінації. У деяких втіленнях суб‗єкта піддають аналізу для визначення, чи експресують його ракові клітини CASB7439 (HASH2). Клінічні способи для діагностики колоректального раку є відомими у галузі техніки. Молекулярні методики та способи для визначення, чи експресує ракова клітина або тканина CASB7439 (HASH2), розкриваються у W001/62778. Експериментальні приклади Приклад 1 Дві молекули нуклеїнової кислоти, що кодує білкові конструкції, що включають CASB7439 поліпептид, одержували у pET19b векторі. His-мітку приєднували до N-термінального кінця CASB7439 послідовності. Ці клони називали LVL007 [SEQ ID NО:15(aa); SEQ ID N0:16 (ДНК)] та LVL010 [SEQ ID NО:17 (aa); SEQ ID NО:18 (ДНК)], відповідно). Спостерігали, що непередбачена мутація виникала у LVL007 клоні, що приводило до однієї амінокислотної заміни проліну на лейцин (у положенні амінокислотного залишку 17 з посиланням на SEQ ID NO: 13). Ці молекули нуклеїнової кислоти використовувалися з трьома різними хазяйськими типами клітин, зокрема, штамами E. coli BLR DE3, BL21 DE3, та Rosetta DE3. Індукцію проводили при двох температурах під час інкубації (16°C або 37°C) та при використанні 1 мM IPTG. Див. розділ Матеріали та Способи, що приведений нижче, для деталей пропису. Збирали білки для кожної індукції та піддавали аналізу за допомогою Вестерн-блоттінгу та SDS-ПАГЕ при використанні забарвлювання Кумассі блакитним. У цих експериментах визначення білка візуалізували тільки при використанні Вестерн-блоттінгу. Приклад 2 Проводили біоінформаційні аналізи CASB7439 гена, виявляючи при цьому наступні структурні характеристики: - шпильковий РНК структурний елемент, розміщений між нуклеотидом 2 та 27 послідовності SEQ ID NO:14 - вміст GC у нуклеотидній послідовності складає 62% - нуклеотидна послідовність є збагаченою на кодони аргініну (26/193 кодованих амінокислот) - амінокислотна послідовність має високий рівень основних амінокислотних залишків - амінокислотна послідовність має високу ізоелектричну точку (11.18) - ділянка низької складності знаходиться у межах амінокислотної послідовності (залишки 727 послідовності SEQ ID NO:13 - домен спіраль-петля-спіраль (HLH) (IPR000014) знаходиться у межах амінокислотної послідовності (залишки 56-108 послідовності SEQ ID NO: 13) - перша ділянка внутрішнього порушення знаходиться у межах амінокислотної послідовності (залишки 118-164 послідовності SEQ ID NO:13) - друга ділянка внутрішнього порушення знаходиться у межах амінокислотної послідовності (залишки 118-164 послідовності SEQ ID NO:13) Заявники також ідентифікували ділянку з високою періодичністю проліну від амінокислоти 127 дo 158. Заявники сконструювали ділянку від амінокислоти 133 до 153, включно, послідовності SEQ ID NO: 13 (aa) ділянка, збагачена на пролін. Збагачена на пролін ділянка, а також деякі інші характеристики, згадані у цьому прикладі, схематично зображені на Фігурі 1/35 разом з ділянками, які згадуються в інших місцях даної заявки. Приклад 3 Немодифіковану CASB7439 ДНК нуклеотидну послідовність (SEQ ID NO: 14) піддавали оптимізації за кодонами за допомогою провайдера послуг синтезу ДНК, Geneart (специфічний ідентифікатор 0606597, Geneart AG, BioPark, Josef-Engert-Str. 11D-93053 Regensburg, Germany) 17 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 та клонували.. Нуклеотидну послідовність CASB7439 одержували з клону за допомогою ПЛР ампліфікації та використовували для одержання двох конструкцій. У першій конструкції, LVL016 [SEQ ID NO:29 (aa), SEQ ID NО:30 (ДНК)] послідовність CASB7439 вбудовували у pET21b (+) вектор при злитті з pD1/3 білком (з Haemophilus influenzae) на N-термінальному кінці CASB7439 послідовності. У другій конструкції, LVL018 [SEQ ID NO:31 (aa); SEQ ID NO:32 (ДНК)] CASB7439 вбудовували у pET21b (+) вектор (без будь-якого компоненту білка D). Ці конструкції піддавали аналізу на продукцію у BLR DE3 E. coli при температурах 16°C та 37°C з 1 мM IPTG. Продукцію визначали за допомогою Вестерн-блоттінгу як нерозчинний білок, що в основному продукується при 16°C. Чотири домени CASB7439 були вибрані для подальшого дослідження на поліпшення продукції білка: домен націлювання на ядро, ділянка, збагачена на пролін, домен зв‗язування ДНК та HLH домен. Приклад 4 Домен націлювання на ядро, домен зв‗язування ДНК, bHLH домен та ділянку, збагачену на пролін CASB7439 вибирали для оцінки продукції білка з конструкції, що включає нуклеотидні послідовності, які кодують CASB7439 поліпептиди. Різні фрагменти нуклеотидної послідовності CASB7439 піддавали ПЛР ампліфікації з оптимізованих за кодонами клонів Geneart, описаних в даній заявці та вбудованих у pET19b (+) вектор, що приводить до одержання п‗яти конструкцій, які мають відмінні послідовності CASB7439 (вкорочені певним чином або повні); кожна конструкція мала полігістидиновий хвіст на N-термінальному кінці від CASB7439 послідовності. Див. Фігуру 2/35. Ці конструкції, показані на Фігурі 2/35, піддавали оцінці на експресію білка у штамі BLR DE3 E.coli при температурах 37°C та 16°C з 1 мM IPTG. Як показано на Фігурі 2/35, ці конструкції включають CASB7439 повної довжини [LVL060, SEQ ID NO:19 (aa), SEQ ID N0:20 (ДНК)]; CASB7439 вкорочений для видалення домену націлювання на ядро [Const-1, SEQ ID NO:21 (aa), SEQ ID NO:22 (ДНК)]; CASB7439 вкорочений для видалення ділянки, збагаченої на пролін [LVL055, SEQ ID NO:1 (aa), SEQ ID NO:2 (ДНК)]; CASB7439, вкорочений для видалення домену зв‗язування ДНК [LVL056, SEQ ID NO:23 (aa), SEQ ID NO:24 (ДНК)]; CASB7439, вкорочений для видалення основного домену спіраль-петля-спіраль (bHLH) [LVL057, SEQ ID NO:25 (aa), SEQ ID NO:26 (ДНК)]. Завдяки помилці експерименту домен Const-1 не мав стоп-кодону, що приводило до трансляції частини векторної нуклеотидної послідовності (яку одержували з додатковими 21 амінокислотами у C-термінальній ділянці, починаючи з LED...). Продукцію LVL055 після інкубації при 16°C (має вкорочення на C-термінальному кінці) чітко визначали на забарвленому за Кумассі SDS-ПАГЕ гелі. Приклад 5 Базуючись на результатах, одержаних у Прикладі 4, конструювали нові конструкції для (i) видалення ділянки, збагаченої на пролін (ii) при збереженні інших ділянок C-термінальної частини поліпептиду. Див. опис Фігур 3/35 та 4/35, що обговорює (i) різноманітні ідентифіковані або спрогнозовані епітопи у межах поліпептиду та (ii) різноманітні альтернативні підходи до видалення ділянки, збагаченої на пролін. Оптимізовану за кодонами послідовність нуклеїнової кислоти гена, що кодує модифікований CASB7439 поліпептид, потім замовляли у Geneart (специфічний ідентифікатор 0706840), перетравлювали та вбудовували паралельно у два різні вектори для одержання наступної конструкції: (і) LVL088, що включає ген CASB7439 з делетованою ділянкою, збагаченою на пролін, яка складається з 21 амінокислоти, злитий з полігістидиновим хвостом на C-термінальному кінці, у pET26b (+) векторі (SEQ ID NO:27 (aa), SEQ ID NO:28 (ДНК)) та (іі) LVL111, що включає ген CASB7439 з делетованою ділянкою, збагаченою на пролін, яка складається з 21 амінокислоти, та з полігістидиновим хвостом як на N-термінальному, так і на C-термінальному кінц, у pET19b (+) векторі (SEQ ID NO:3 (aa), SEQ ID NO:4 (ДНК)). LVL111 конструкцію (див. SEQ ID NO:3) клонували у відповідності із тією самою стратегією, що й використовували для LVL055, тобто, при використанні pET19 (+) вектора, що включає полігістидиновий хвіст, який складається з 10 гістидинів, після яких йде сайт ентерокінази. Це приводило до одержання партнера злиття, який складається з 23 амінокислот (SEQ ID NO:42) на N-термінальному кінці. Інші конструкції одержували з синтетичним геном, оптимізованим за кодонами, що обговорюється у Прикладі 3 (специфічний ідентифікатор 0606597), включаючи наступні: - LVL090 (білкова конструкція, що включає (від N-термінального кінця) білок D, повнорозмірний оптимізований CASB7439 та 6-His у pET21 векторі) (SEQ ID NO:43 (aa); SEQ ID NO:44 (ДНК)) 18 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 - LVL112 (білкова конструкція, що включає (від N-термінального кінця) SUMO білок, повнорозмірний оптимізований CASB7439 у pSUMO векторі) (SEQ ID NO:45 (aa); SEQ ID NO:46 (ДНК)) - LVL113 (білкова конструкція, що включає (від N-термінального кінця) SUMO білок, повнорозмірний оптимізований CASB7439 у pSUMO векторі) (SEQ ID NO:47 (aa); SEQ ID NO:48 (ДНК)) - LVL114 (білкова конструкція, що включає (від N-термінального кінця) SUMO білок, CASB7439, вкорочений при амінокислоті 117 у pSUMO векторі) (SEQ ID NO:49 (aa); SEQ ID NО:50 (ДНК)) - LVL115 (білкова конструкція, що включає (від N-термінального кінця) SUMO білок, модифікований CASB7439 з ділянкою, збагаченою на пролін, видаленою у pSUMO векторі) (SEQ ID NO:51 (aa); SEQ ID NO:52 (ДНК)) Не спостерігали ніякого поліпшення продукції, незважаючи на те, що здійснювали тільки один повтор (дані не представлені). Продукцію LVL088 аналізували після індукції протягом ночі при використанні 1 мM IPTG при 37°C та при 16°C. При електрофоретичному аналізі продукцію LVL088 візуалізували при використанні Вестерн-блоттінгу. Одна серія очистки з чотирьох колб для культури клітин на 800 мл приводила до одержання 0,4 мг білка. З іншого боку, LVL111 легко визначали після забарвлення Кумассі блакитним. Рекомбінантний білок LVL111 одержували та очищали у 8 партіях. Декілька партій LVL111 (кожна з чотирьох колб для культури тканин на 800 мл) були оцінені як такі, що продукують білок в інтервалі від 3,3 мг/партія до 10 мг/партія очищеного білка. Таким чином, продукція LVL088 була нижча за ту, яку спостерігали для LVL111. Очищений білок потім піддавали аналізу на його реактивність з декількома анти-CASB7439 моноклональними антитілами та отримували позитивну реактивність у всіх випадках. Приклад 6 Досліджували видалення C-термінального полігістидинового хвоста з LVL111 конструкції. Молекулу нуклеїнової кислоти, що кодує поліпро (-) CASB7439 поліпептид без частини, що кодує C-термінальний полігістидин, піддавали ампліфікації за допомогою ПЛР з LVL111 конструкції та клонували у pET19b вектор. Цю конструкцію позначали як LVL137. LVL137 конструкція суттєво не містила LVL111 нуклеотидної послідовності, що кодує C-термінальний полігістидиновий хвіст. SEQ ID NO:5 (aa); SEQ ID NO:6 (ДНК). Видалення C-термінального полігістидинового хвоста не виявляло видимого впливу на рівень продукції. Як було проаналізовано при використанні SDS-ПАГЕ геля, LVL137 виробляється на тому самому рівні білка, що й LVL111. Молекулу нуклеїнової кислоти, що кодує поліпро (-) CASB7439 поліпептид потім піддавали ампліфікації при використанні ПЛР від Geneart (0706840) синтетичної CASB7439 послідовності та клонували у pET26 вектор із шістьома полігістидиновими мітками на N-термінальному кінці (LVL138). SEQ ID NO:33 (aa); SEQ ID NO:34 (ДНК). Ця конструкція демонстрували значно знижений рівень продукції білка та складності при визначенні за допомогою Вестерн-блоттінгу. Одержували молекулу нуклеїнової кислоти, що кодує шість полігістидинових міток на Nтермінальному кінці поліпро (-) CASB7439 поліпептиду у pET24 резистентному до канаміцину векторі. Нову лінкерну конструкцію використовували для видалення структури шпильки, створеної між послідовністю полігістидинової мітки та 5' кінцем CASB7439 гена. Зокрема, кодони для перших восьми амінокислотних залишків білкової конструкції змінювали до одержання наступної нуклеотидної послідовності: 5' ATG CAT CAT CAT CAT CAT CAT GAC... 3' (SEQ ID NO:35). (Вихідні кодони були наступними 5'ATG CAC CAT CAC CAT CAC CAT GAT... 3' (SEQ ID NO:36)). Цю конструкцію позначали як LVL160. SEQ ID NO:37 (aa); SEQ ID NO:38 (ДНК). При використанні аналізу у гелі та Вестерн-блоттінг аналізу було виявлено, що продукція була такою ж слабкою, як і та, що була одержана при використанні LVL138. Одержували молекулу нуклеїнової кислоти, що кодує десять полігістидинових міток на Nтермінальному кінці поліпро (-) CASB7439 поліпептиду у pET24 резистентному до канаміцину векторі. Цю конструкцію ідентифікували як LVL168. SEQ ID NO:11 (aa); SEQ ID NO: 12 (ДНК). Рівень продукції білка був порівнюваним з таким, що спостерігали для LVL111. Вирівнювання деяких з конструкцій, що обговорюються у цьому прикладі, може бути знайдене на Фігурі 5/35. Результати для конструкцій, що обговорюються у цьому прикладі та Прикладі 1, є підсумованими у Таблиці 1, що представлена нижче. № LVL LVL007 LVL010 Опис Вектор pET19b(+) pET19b(+) Довга His-CASB7439 Довга His-CASB7439 19 Продукція гель-/блот.+ гель-/блот.+ UA 99801 C2 LVL016 LVL111 LVL137 LVL138 LVL160 LVL168 5 10 15 20 25 30 35 40 45 50 pD1/3-CASB7439-OPT-6His Довга His-Дел.-полі-Pro-домен-His-OPT Довга His-Дел.-полі-Pro-домен-OPT 6His-Дел.-полі-Pro-домен-OPT His модиф.-Дел.-полі-Рro-домен-OPT 10His-Дел.-полі-Рro-домен-OPT pET21b(+) pET19b(+) pET19b(+) pET26b(+) pET24b(+) pET24b(+) гель-/блот.+ гель+++/ блот.+++ гель+++/блот.+++ гель-/ блот.+ гель-/блот.+ гель+++/ блот.+++ Taблиця 1. Рівні експресії для конструкцій, що обговорюються у Прикладах 1, 3 та 6. Конструкції, що приводять до "гель-/блот.+" продукції звичайно візуалізуються тільки за допомогою Вестерн-блоттінгу, коли лізати, що містять цільні клітини, піддають SDS-ПАГЕ. Конструкції, що приводять до "гель+++/блот.+++" продукції чітко візуалізуються за допомогою забарвлювання за Кумассі, коли лізати, що містять цільні клітини, піддають SDS-ПАГЕ. 6His: Гістидинова мітка = 6 залишків гістидину Довга His = 23aa (MGHHHHHHHHHHSSGHIDDDDKH) (SEQ ID NO:42) Делет.-полі-Pro-домен: CASB7439, модифікований шляхом делеції ділянки, збагаченої на пролін, OPT: оптимізований за кодонами His модиф.: Гістидинова мітка, модифікована для селекції на кодони (див. в даній заявці, Приклад 6), 1/3pD: Конструкція білка D, описана в даній заявці 10His: Гістидинова мітка з 10 залишками гістидину Приклад 7 Іншу CASB7439 конструкцію одержували шляхом злиття з фрагментом білка D з Haemophilus influenzae. Як показано у SEQ ID NO:39 (aa) (SEQ ID N0:40 (ДНК)), амінокислоти Met-Asp-Pro зливали з 108 N-термінальними амінокислотами процесованого білка D (амінокислотні залишки 2-109 процесованого білка D). SEQ ID NO:39 називається в даній заявці як 1/3pD або 1/3 білок D. Синтетичну нуклеотидну послідовність CASB7439, одержану від Geneart (0706840) (делетована ділянка, збагачена на пролін) субклонували у pET26-1/3pD, стійку до канаміцину плазміду, що містить 1/3pD, для одержання LVL141. Послідовність LVL141 є представленою у SEQ ID NO:7 (aa) та SEQ ID NO:8 (ДНК). Продукція є порівняною із такою для LVL111. Цей клон потім піддавали мутації за допомогою ПЛР для видалення лінкера, що складається з чотирьох амінокислот (Ala-Ala-Ala-His) між 1/3pD та CASB7439 компонентами. Послідовність одержаної конструкції, яка позначена як LVL144, є представленою у SEQ ID NO:9 (aa) та SEQ ID NO: 10 (ДНК). Продукція є порівняною із такою для LVL111. Фігура 6/35 показує вирівнювання CASB7439 (SEQ ID NO:13); LVL168 (SEQ ID NO:12), PD1/3 (SEQ ID NO:39) та LVL144 (SEQ ID NO:9). Приклад 8 Аналізи солюбілізації LVL168 В загальному випадку очищені білкові конструкції, що включають CASB7439, піддавали солюбілізації у буфері на основі 6M гуанідину, переводили у буфер на основі 8M сечовини та зберігали у 4M сечовині, 20 мM HEPES, 1 мM TCEP, 150 мM NaCI, pH7,0. Попередні результати, які проводили у малих об'ємах, дають змогу запропонувати, що LVL168 може також бути солюбілізований у фосфатному буфері без сечовини або без будь-якого відновлювального агенту, такого, як ЕДТА або TCEP, та з або без NaCI. Не спостерігали ніякої преципітації після діалізу у цих буферах. Аналізи солюбілізації проводили для характеристики LVL168 білка у шістнадцяти буферах, приведених у Taблиці 2. Сто мікролітрів очищених зразків LVL168 білка у буфері на основі 8M сечовини, 20 мM HEPES, 150 мM NaCI, 1 мM TCEP, pH7,0, поміщали у блок для проведення мінідіалізу 10000 MWCO (Biolynx), попередньо кондиціонований при використанні двох промивань буфером. Діаліз проводили протягом ночі у 100 мкл кожного із шістнадцяти буферів, приведених у Таблиці 2, при перемішуванні при 4°C. Потім зразки виймали з блоків, поміщали у пробірки Епендорфа та центрифугували протягом 5 хвилин при 20,000 g. Розчинні білки залишалися у суспензії, а нерозчинні білки осаджувалися та одержували осади після центрифугування.. Буфери, в яких LVL168 білок був розчинним, є приведеними у Таблиці 2. Супернатанти аналізували за допомогою SDS-ПАГЕ, та концентрації розчинних білків, що залишилися, вимірювали за допомогою RC DC способу при використанні інструкцій виробника. Набір для здійснення RC DC методу на основі модифікованого способу Лоурі є доступним від BioRad Laboratories (1000 Alfred Nobel Drive, Hercules, CA 94547). Зразки зберігали при 4°C для аналізу стабільності. 20 UA 99801 C2 5 Аналізи стабільності LVL168 LVL168 білки з аналізу стабільності витримували протягом 7 днів при температурі 4°C. Двадцять мікролітрів кожного із зразків 2, 3, 5, 6, 9 - 16 переносили в інші пробірки та піддавали аналізу на їх стабільність при 4°C, 37°C або при кімнатній температурі протягом 2 додаткових днів. Зразки центрифугували протягом 5 хвилин при 20,000 g. Супернатанти аналізували при використанні SDS-PAGE, та профілі піддавали аналізу на деградацію, агрегацію та втрату білка шляхом порівняння із вихідним очищеним білком. Білок був стабільним у більшості проаналізованих буферів. № зразка 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 Буфер Розчинність PBS 1X pH7,3 2M Сечовина, 20 мМ Трис, 150 мМ NaCI + 1 мМ TCEP, pH 8,0 4M Сечовина, 20 мМ Трис, 150 мМ NaCI + 1 мМ TCEP, pH 7,0 PBS 1X, 1 мM TCEP, 1 мМ ЕДТА, pH7,3 10 мМ NaAc 1 мМ ЕДТА. 1 мM, TCEP, 5 мМ NaCI, pH 5,01 20 мМ імідазол, 150 мМ, NaCI, 1 мМ ЕДТА, 1 мМ TCEP, pH 6,0 20 мМ біцину, 138 мМ NaCI, 1 мМ ЕДТА, 1 мМ TCEP, pH 8,7 50 мМ Трис, 250 мМ NaCI, 27 мМ сахарози, 0,1% Твін 80, 1 1 мM TCEP, pH 8,5 50 мМ Трис, 250 мМ NaCI, 270 мМ сахарози, 0,3 M L-аргініну, 1 мM TCEP, pH 8,5 20 мМ Na2HPО4, 150 мМ NaCI, 1 мМ ЕДТА + 1 мМ TCEP, pH 6,0 20 мМ Na2HPО4, 5 мМ NaCI, 1 мМ ЕДТА + 1 мМ TCEP, pH 6,0 20 мМ Na2HPО4, 5 мМ NaCI, 1 мМ ЕДТА pH 6,0 20 мМ Na2HPО4, 5 мМ NaCI + 1 мМ TCEP + pH 6,0 20 мМ Na2HPО4, 5 мМ NaCI pH 6,0 20 мМ Na2HPО4, 5 мМ NaCI pH 6,5 20 мМ Na2HPО4, 150 мМ NaCI pH 6,0 Нерозчинний Розчинний Розчинний Нерозчинний Розчинний Розчинний Нерозчинний Нерозчинний Розчинний Розчинний Розчинний Розчинний Розчинний Розчинний Розчинний Розчинний 10 Для зручності читачів підсумовані конструкції, згадані у даній заявці, та їх відносні рівні продукції забезпечуються у Таблиці 3 Таблиця 2 Аналізи розчинності та стабільності LVL168 № LVL Опис LVL007 Довга His-CASB7439 LVL010 Довга His-CASB7439 LVL016 pD1/3-CASB7439-OPT-6His LVL018 CASB7439-OPT-6His Const-1 Довга His-CASB7439-OPT-Const.-1 (націлююча на ядро вкороч.) LVL055 Довга His-полі-Pro-домен-вкороч.-OPT LVL056 Довга His-ДНК зв‗яз. вкороч.-OPT LVL057 Довга His-bHLH, узята окремо-OPT LVL060 Довга His-C7439-OPT LVL088 Делет.-полі-Pro-домен-OPT-6His LVL111 Довга His-делет.-полі-Pro-домен-His-OPT LVL137 LVL138 LVL141 LVL 144 LVL160 LVL168 Довга His-Делет-полі-Pro-домен-OPT 6His-Делет.-полі-Pro-домен-OPT 1/3pD-Делет.-полі-Pro-домен-OPT-6His 1/3pD-делет.полі-Pro-домен-OPT His мут. лінкер His модиф.-Делет.-полі-Рro-домен-OPT 10His-Делет.-полі-Рro-домен-OPT 21 Вектор Продукція pET19b(+) гель-/блот.+ pET19b(+) гель-:блот.+ pET21b(+) гель-/блот. + pET21b(+) гель-/блот. + pET19b(+) гель-/ блот. + pET19b(+) гель+++/блот. +++ pET19b(+) гель-/блот. pET19b(+) гель-/блот. pET19b(+) гель-/блот. + pET26b(+) гель-/блот. + pET19b(+) гель+++/ блот. +++ pET19b(+) гель+++/блот. +++ pET26b(+) гель-/ блот. + pET26b(+) гель+++/блот. +++ pET26b(+) гель+++/ блот. +++ pET24b(+) гель-/блот. + pET24b(+) гель+++/ блот. UA 99801 C2 +++ 5 10 15 20 25 30 35 40 45 50 55 Таблиця 3. Див. скорочення у Таблиці 1. Наступні додаткові скорочення використовуються у Таблиці 3: вкороч.: вкорочена Const.: конструкція мут.: послідовність, мутована для видалення неспоріднених амінокислот між pD1/3 та модифікованим CASB7439 поліпептидом лінкер: неспоріднені амінокислоти між двома частинами молекули ДНК зв‘яз.: домен зв‗язування ДНК bHLH узятий окремо: основний домен спіраль-петля-спіраль Матеріали та Способи Експресія - індукція у малих об‗ємах: Штами E. coli BLR (DE3) трансформували при використанні конструкцій, згаданих у даній заявці, поміщали в агарові планшети з бульйоном Лурія (LB), що містять прийнятний антибіотик (40 мкг/мл канаміцину або 100 мкг/мл карбеніциліну) та інкубували протягом ночі при 37°C. Одержаний бактеріальний газон використовували для інокуляції 20 мл середовища LB з альтернативним джерелом білка (APS) з прийнятним антибіотиком (40 мкг/мл канаміцину або 100 мкг/мл карбеніциліну) та поміщали при 37°C до досягнення OD600 від 0,5 дo 1,0. Індукцію проводили шляхом додання 1 мМ IPTG, або при температурі 16°C, або при 37°C. Для проведення індукції при 16°C культури, що ростуть, охолоджували на льоду за 1 годину до індукції експресії рекомбінантного білка. Культури потім поміщали назад для вирощування при температурі 16°C та підтримували за цих умов протягом 16 годин. Для проведення індукції при 37°C експресію рекомбінантних білків безпосередньо індукували шляхом додання IPTG до культур, що ростуть. Індукцію підтримували при 37°C протягом 3 годин. Невеликі аліквоти, що складають 1 мл, відбирали до та після індукції. Потім аліквоти центрифугували (10 хвилин, 4°C, 20000 x g) та бактеріальні осади витримували при -20°C перед аналізом. Аналіз проводили у відповідності із білковою екстракцією BugBuster згідно з рекомендаціями виробника. BugBuster, що виробляється Novagen, є комерційно доступним від VWR. Рівень експресії визначали за допомогою забарвлювання за Кумассі SDS-ПАГЕ гелів та при використанні Вестерн-блоттів. Розчинність оцінювали після визначення білка в осадах або у супернатантах за допомогою BugBuster eкстракції. Експресія - індукція у великих об‗ємах: Штами E. coli BLR (DE3) трансформували при використанні конструкцій, як описано у даній заявці, поміщали на планшети з LB агаром, що містять прийнятний антибіотик та інкубували протягом ночі при 37°C. Одержаний бактеріальний газон використовували для інокуляції бажаної кількості колб з 800 мл LB APS середовища, що містить прийнятний антибіотик (40 мкг/мл канаміцину або 100 мкг/мл карбеніциліну) та поміщали при 37°C до досягнення OD600 від 0,5 дo 1,0. Колби на 2,5 л, які містять 800 мл вирощеної культури, потім охолоджували на льоду за 1 годину перед індукцією експресії рекомбінантного білка шляхом додання 1 мM IPTG. Для проведення індукції при 16°C культури, що ростуть, охолоджували на льоду за 1 годину перед індукцією експресії рекомбінантного білка. Культури потім поміщали назад для вирощування при температурі 16°C та підтримували за цих умов протягом 16 годин. Культури центрифугували (15 хвилин, 4°C, 6000 x g) та бактеріальні осади витримували при -80 °C до проведення процесу очистки. Невеликі аліквоти, що складають 1 мл, відбирали до та після індукції BugBuster та аналізували за допомогою SDS-ПАГЕ і Вестерн-блоттінгу перед очищенням. Очистка Бактеріальні осади ресуспендували у літичному буфері (20 мМ Трис буфер (pH 8,0), що містить 500 мМ NaCI, 10 мМ TCEP) та суміші інгібіторів протеази (повна, вільна від ЕДТА). Бактерії піддавали лізису або за допомогою обробки ультразвуком при використанні 13 мМ зонда на ультразвуковому пристрої Vibra для клітин, при використанні Emulsiflex C3 (Avestin), або за допомогою системи для руйнування клітин Constant (Constant System). Розчинний (cупернатант) та нерозчинний (осад) компоненти розділяли шляхом центрифугування при 20 000 g протягом 20 хвилин при 4°C. Оскільки CASB7439 рекомбінантні білки, як очікуються, містяться в осадах, супернатанти відкидали. Звичайно нерозчинні компоненти (осади) повторно розчиняли у 50 мМ біциновому буфері pH 8,0, що містить 6M гуанідину HCI, 500 мМ NaCI, 10 мМ TCEP, та потім центрифугували (20 000 x g протягом 20 хвилин). Супернатант завантажували на 5 мл His Trap колонку (GE 22 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Healthcare), що вже була врівноважена у попередньому буфері, проте з концентрацією TCEP, зниженою до 1 мM. Після завантаження колонку промивали один раз при використанні того самого буфера. 6M GnHCI хаотропного агенту замінювали на 8M сечовину (8M сечовини, 20 мМ HEPES, 500 мМ NaCI, 1 мМ TCEP, pH 8,0). Елюювання проводили при використанні 20 мМ HEPES буфера (pH 80), що містить 8M сечовини, 500 мМ NaCI, 1 мМ TCEP та 250 мМ імідазолу. Зразки з фракцій елюювання піддавали аналізу за допомогою SDS-ПАГE, та фракції, які містять білок, що представляє інтерес, об‗єднували, концентрували на центрифузі, у разі необхідності, та використовували для другого етапу очистки. Проводили один або два етапи SEC (ексклюзійна хроматографія за розмірами) (Superdex 200, GE Healthcare), у залежності від необхідної чистоти очистки, у 8M сечовини, 20 мМ HEPES, 500 мМ NaCI, 1 мМ TCEP, pH 8,0, перед останнім етапом знесолювання. Знесолення проводили на G25 колонці для знесолювання, попередньо врівноваженій за допомогою заключного 20 мМ HEPES буфера (pH 70), що містить 4M сечовини, 150 мМ NaCI та 1 мМ TCEP. Зразки білка розділяли на аліквоти по 1 мл у пробірках та зберігали при - 80°C. Аналіз білка — визначення концентрації Концентрації білка визначали при використанні RC DC білкового аналізу (модифікований BioRad метод Лоурі). Цей аналіз використовували з причини несумісності деяких компонентів заключного буфера з класичними аналізами Лоурі або BCA (високий вміст сечовини та редукційних агентів). Відомі кількості заключного препарату білка також піддавали аналізу за допомогою SDS-ПАГЕ, Вестерн-блоттінгу та на вміст LAL для оцінки агрегації або деградації білка E. coli та рівня контамінації LPS Приклад 9 Підтвердждення експресії CASB7439 у нормальних тканинах та тканинах колоректального раку (CRC) Заявники проводили кількісний ПЛР аналіз у реальному часі експресії іРНК CASB7439 у 83 CRC зразках та 12 зразках нормальної тканини товстого кишечнику, одержаних від Corixa Corporation. іРНК CASB7439 понадекспресувалася принаймні 10-кратно у CRC у порівнянні із нормальним товстим кишечником у 63% CRC зразків, перевірених у порівнянні із нормальними тканинами (дані представлені як середнє значення співвідношення експресії CASB7439/β-актин) (Фігура 7/35). Крім того, заявники визначали CASB7439 білок шляхом імунофлуоресценції (IF) у CRC тканинах людини при використанні кролячого aнти-CASB7439 моноклонального антитіла (40-12) (антитіло обговорюється в іншому місці заявки); не виявляли ніякого CASB7439 білка у зрізах нормальних сусідніх тканин. Додаткові дані стосовно експресії для CASB7439 були одержані за допомогою GeneLogic (Gene Microarray) аналізу при використанні стандартних методик. Було виявлено, що іРНК CASB7439 експресується на високому рівні у CRC зразках на усіх стадіях хвороби (дані не показані). Заявники проводили кількісний ПЛР аналіз у реальному часі експресії іРНК CASB7439 у 83 CRC зразках та 12 зразках нормальної тканини товстого кишечнику, одержаних від Corixa Corporation. іРНК CASB7439 понадекспресувалася принаймні 10-кратно у CRC у порівнянні із нормальним товстим кишечником у 63% CRC зразків, перевірених у порівнянні із нормальними тканинами (дані представлені як середнє значення співвідношення експресії CASB7439/β-актин) (Фігура 7/35). Незалежні літературні джерела описують визначення CASB7439, тобто іРНК HASH-2, за допомогою in situ гібридизації (ISH) на дуже низькому рівні у нижній частині крипт кишечнику, але не в інших нормальних тканинах, за винятком плаценти. Jubb та ін. (2006) Oncogene 25: 3445-3457. У тканинах колоректального раку (CRC) іРНК CASB7439 була показана як така, що 15-кратно експресується у 70% проаналізованих тканинах людини. Експресія білка CASB7439 у CRC та нормальних сусідніх тканинах кишечнику Для того, щоб проаналізувати білкову експресію CASB7439 шляхом імунофлуоресценції у нормальних тканинах та у CRC тканинах, розробляли специфічне кроляче анти-CASB7439 моноклональне антитіло (40-12). Кролячі гібридоми проти CASB7439 одержували від комерційного постачальника Epitomics Inc., вони піддавалися скринінгу заявниками. Моноклональні антитіла, які виробляється гібридомами, спочатку піддавали скринінгу шляхом ELISA при використанні рекомбінантного білка LVL111 (SEQ ID NO:3). Специфічність моноклональних антитіл перевіряли за допомогою Вестерн-блоттінгу при використанні LVL111 та клітинного лізату, одержаного з рекомбінантної лінії клітин, сконструйованої для експресії CASB7439 (див. нижче, TC1/CASB7439). Експресію білка CASB7439 у TC1/CASB7439 пухлинах (вирощені у мишах), SW-620 та SW-480 клітинах (позитивна за CASB7439 клітинна лінія колоректального раку людини) та у HCT-116 клітинах (негативна за CASB7439 клітинна лінія 23 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 колоректального раку людини) оцінювали за допомогою імунофлуоресценції з моноклональними антитілами. Моноклональне антитіло 40-12 відбирали для підтвердження експресії білка CASB7439. Заявники одержували від NDRI (National Disease Research Interchange, 8 Penn Center, 8-ий поверх Floor, 1628 JFK Boulevard, Philadelphia, PA 19103) заморожені зразки тканин, заключені у OCT, як для CRC, так нормальної сусідньої тканини від 17 різних індивідуумів; для 5 із цих індивідуумів також одержували зразки метастатичних тканин. Заявники піддавали скринінгу 17 первинних пухлин, 17 нормальних прилеглих тканин та 5 метастазів. З 17 проаналізованих первинних CRC пухлин 13 були позитивними за експресією білка CASB7439 (76,5%). Усі ці позитивні за CASB7439 зразки являли собою аденокарциноми. Жодна із прилеглих нормальних тканин не експресувала здатних до визначення рівнів CASB7439 білка. Серед метастатичних тканин 3 з 5 були позитивними (60%) на експресію білка CASB7439. Кореляцію між експресією білка CASB7439 та іРНК вивчали у 16 CRC тканинах за допомогою кількісного ПЛР аналізу у реальному часі, який проводили при використанні звичайних методик як на ракових тканинах, так і на тканинах нормальних прилеглих тканинах кишечнику. Ki67 забарвлювання використовували як маркер проліферації. Кореляція між експресією білка CASB7439 та іРНК є представленою на Фігурі 8/35. Зразки тканин, які демонстрували білок CASB7439 при дослідженні із використання мунофлуоресценції, також показали найвищі рівні іРНК CASB7439. Приклад 10 Імуногенність CASB7439 у мишей: Імуногенність CASB7439 для T-клітин у інбредних мишей Для вивчення T-клітинної імуногенності CASB7439 у мишей одержували банк пептидів за допомогою звичайних способів синтезу для охоплення цільної послідовності CASB7439 білка (Фігура 9/35). Кожний пептид являв собою п‗ятнадцятимер, перекриваючи наступний пептид 11 амінокислотами. Для дослідження імунодомінантних ділянок 14 пулів пептидів піддавали класифікації у відповідності із матрицею так, що будь-які два пули пептидів мали тільки один спільний пептид (Фігура 10/35). Для дослідження T-клітинної імуногенності у мишей аж до 15 інбредних мишей та аж до 10 аутбредних мишей на групу одержували чотири внутрішньом‗язові імунізації кожні два тижні при використанні різних доз (від 0 дo 30 мкг) різних рекомбінантних CASB7439 білків, рецептованих з AS01B або AS15, обидва з яких являють собою власні ад‗юванти GSK. При рецептуванні з антигеном AS01B складався з MPL (100 мкг/мл) та QS21 (100 мкг/мл) у ліпосомах. При рецептуванні з антигеном AS15 складався з MPL (100 мкг/мл), QS21 (100 мкг/мл) та CpG7909 (840 мкг/мл) у ліпосомах. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у концентрації 2X. Таким чином, кожна миша у групі AS01B одержувала 5 мкг MPL та 5 мкг QS21. Кожна миша у групі AS15 одержувала 5 мкг MPL, 5 мкг QS21 та 5 мкг CpG7909. Після останньої імунізації одержували селезінки або PBL від мишей. Для проведення аналізу імуногенності CASB7439 у інбредних мишей селезінки або PBL аналізували індивідуального або об‗єднували їх. Для проведення аналізу імуногенності CASB7439 у CD1 аутбредних мишей, кожну селезінку піддавали обробці індивідуально. 6 PBL або ізольовані спленоцити ресуспендували при заключній концентрації 10x10 клітин на мл у RPMI 1640 культуральному середовищі з наступними добавками: пеніцилін-стрептоміцин (1X), неесенціальні амінокислоти (1X), 2-меркаптоетанол (55 мM), піруват натрію (100 мM), Lглутамін (200 мM). Клітини стимулювали при використанні пептидів з CASB7439 послідовності (використовувалися при заключній концентрації 1 мкг/мл/пептид у RPMI 1640 + добавки + 5% інактивований прогрівання FBS). Внутрішньоклітинне забарвлювання (ICS) для CD4, CD8, IL-2, IL-5, IFNγ та TNFα проводили у відповідності з класичними процедурами. Після стимуляції клітини обробляли за допомогою брефелдіну A та фіксували. Забарвлення CD4 та CD8поверхні клітин проводили за допомогою специфічних для CD4 та CD8 моноклональних антитіл. Потім клітин пермеабілізували та забарвлювали при використанні моноклональних антитіл, що утворилися проти IL-2, IL-5, IFNγ та TNFα білків. Клітини піддавали аналізу за допомогою проточної цитометрії при використанні BD FACSCanto II. Дані виражали у процентах подвійно позитивних IFNγ+ та TNFα+ CD4 або CD8 T-клітин. Експеримент з порівняння Т-клітинної імуногенності у декількох лініях інбредних мишей Заявники порівнювали імуногенність CASB7439 білка, рецептованого або з AS01B, або з AS15, у C57BI/6, Balb/C, CB6f1 (потомство першого покоління C57BI/6 мишей, схрещених з Balb/C) та C3H інбредних мишей, яких одержували від Charles River, Senneville, QC, Canada. П‗ять мишей на групу імунізували при використанні 10 мкг рекомбінантного CASB7439 білка (LVL111), рецептованого або з AS01B, або з AS15. Збирали селезінки та об‗єднували зразки (один пул з п‗яти селезінок на групу). CD4 T-клітинна відповідь (подвійно позитивні IFNγ+ та 24 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 TNFα+) на LVL111 для кожної лінії є представленою на Фігурі 11/35 для AS01B композиції та на Фігурі 12/35 для AS15 композиції. Ніякої CD8 T-клітинної відповіді (подвійно позитивні IFNγ+ та TNFα+) не спостерігали у жодної з чотирьох інбредних ліній мишей. CASB7439 імунодомінантні пептиди, ідентифіковані у інбредних мишей, імунізованих при використанні CASB7439+AS15, є представленими на Фігурі 13/35. Імунодомінантні пептиди з відповідними ділянками послідовності SEQ ID NO: 13 являють собою: - C57BI/6: пептид 39 (SEQ ID NO:91) (aa 153 - 167 послідовності SEQ ID NO:13) - Balb/c: пептид 24 (aa 93 - 107 послідовності SEQ ID NO:13) - CB6f1: пептиди 7-8 (aa 25 - 43 послідовності SEQ ID NO: 13) та 23-24 (aa 89 - 107 послідовності SEQ ID NO:13) - C3H: пептиди 9-11 (aa 33 - 55 послідовності SEQ ID NO: 13), 16-19 (aa 61 - 87 послідовності SEQ ID NO:13) та 41-43 (aa 161-183 послідовності SEQ ID NO:13). Дослідження T-клітинної імуногенності LVL111, LVL168 та LVL144 у інбредних мишей Імуногенність LVL111, LVL168, та LVL144 вивчали у інбредних CB6f1 мишей. У цьому експерименті п‗ять мишей на групу (Charles River, Senneville, QC, Canada) імунізували чотири рази кожні два тижні за допомогою рекомбінантних CASB7439 білків (LVL111, LVL168 та LVL144), рецептованих з AS15. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у 2X концентрації, як описано будь-де у даній заявці. CD4 та CD8 T-клітинні відповіді (подвійно позитивні IFNγ та TNFα) вимірювали шляхом внутрішньоклітинного забарвлювання та аналізу проточної цитометрії у індивідуальних CB6f1 мишей після стимуляції спленоцитів при використанні CASB7439 пептидної матриці. (Дані не показані) Імунізації при використанні усіх трьох конструкцій запускали CD4 T-клітинну імунну відповідь проти тих самих імуногенних ділянок (aa 25 - 43 послідовності SEQ ID NO:13 та aa 89 107 послідовності SEQ ID NO:13) у CB6f1 інбредних мишей. Імуногенність CASB7439 у мишей: T-клітинна імуногенність CASB7439 у аутбредних мишей CD1 аутбредних мишей (Charles River, Senneville, QC, Canada) імунізували чотири рази кожні два тижні за допомогою рекомбінантного CASB7439 білка (LVL111), рецептованого або з AS01B, або з AS15. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у 2X концентрації, як описано будь-де у даній заявкці. CD4 та CD8 T-клітинні відповіді (подвійно позитивні IFNγ та TNFα) вимірювали шляхом внутрішньоклітинного забарвлювання та аналізу проточної цитометрії у індивідуальних CD1 мишей після повторної стимуляції спленоцитів при використанні CASB7439 пептидної матриці. Таблиці CASB7439 CD4-імуногенних пептидів, ідентифікованих у аутбредних мишей, імунізованих при використанні або CASB7439+AS01B, або CASB7439+AS15, є представленими на Фігурі 14/35 верхня та нижня частини, відповідно. CD4 T-клітинні імуногенні ділянки, ідентифіковані у CD1 аутбредних мишей, перекривають наступні ділянки CASB7439 (експресовані у межах амінокислотних положень SEQ ID NO:13): пептиди 8-10 (aa 29 - 51 послідовності SEQ ID NO:13), пептиди 16-18 (aa 61 - 83 послідовності SEQ ID NO: 13) та пептиди 24-26 (aa 93 - 115 послідовності SEQ ID NO:13). CD8 T-клітинні імунодомінантні ділянки, ідентифіковані у CD1 аутбредних мишей, перекривають наступні ділянки CASB7439 (SEQ ID NO:13): пептиди 16-18 (aa 61 - 83 послідовності SEQ ID NO: 13), пептиди 24-26 (aa 93 115 послідовності SEQ ID NO: 13) та пептид 32 (aa 125 - 139 послідовності SEQ ID NO:13). (Дані не показані). Дослідження T-клітинної імуногенності LVL111. LVL168 та LVL144 у аутбредних мишей Опосередковані CASB7439+AS15 CD4 та CD8-T-клітинні відповіді, індуковані за допомогою LVL111, LVL168 та LVL144, аналізували у індивідуальних аутбредних CD1 мишей, яких закуповували у Charles River, Senneville, QC, Canada. П‗ять мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації, що складалися з 10 мкг CASB7439 рекомбінантного білка (LVL111, LVL168 або LVL144), рецептованого з AS15. (Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у концентрації 2X, як описано будь-де у даній заявці). П‗ять мишей у контрольній групі імунізували при використанні буфера на основі фізіологічного розчину. Через два тижні після четвертої імунізації мишей забивали. CD4 та CD8 T-клітинні відповіді (подвійно позитивні IFNγ та TNFα) вимірювали шляхом внутрішньоклітинного забарвлювання та аналізу проточної цитометрії після повторної стимуляції спленоцитів за допомогою чотирьох пулів пептидів (пул 1: пептиди 8-9-10, пул 2: пептиди 16-17-18, пул 3: пептиди 24-25-26 та пул 4: пептиди 30-31-32). Дані є представленими на Фігурі 15/35. Такі самі CD4 T-клітинні імуногенні ділянки послідовності SEQ ID NO:13, зокрема, пептиди 16-18 (aa 61 - 83 послідовності SEQ ID NO: 13) та пептиди 24-26 (aa 93 - 115 послідовності SEQ ID NO: 13) активувалися після імунізацій за допомогою кожної з трьох CASB7439 конструкцій, рецептованих з AS15. 25 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 Дослідження T-клітинної імуногенності LVL144 та LVL168 у HLA A2.1/DR1-трансгенних мишей Опосередковані CASB7439+AS15 CD4 та CD8-T-клітинні відповіді, індуковані за допомогою двох конструкцій (LVL168 або LVL144), аналізували в об‗єднаних HLA A2.1/DR1-трансгенних мишей (3 пулів з 3 мишей), яких закуповували в Інституті Пастера, Париж, Франція. 9 мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації, що складалися з 6,25 мкг LVL168 або 10 мкг LVL144, рецептованих з AS15. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у концентрації 2X, як описано будь-де у даній заявці. Двох мишей у контрольній групі імунізували при використанні ад‗юванта, узятого окремо. Через два тижні після четвертої імунізації мишей забивали. CD4 та CD8 T-клітинні відповіді (подвійно позитивні IFNγ та TNFα) вимірювали шляхом внутрішньоклітинного забарвлювання та аналізу проточної цитометрії після повторної стимуляції PBL та спленоцитів з чотирма пулами пептидів при використанні 7 пулів пептидів, що охоплюють повну CASB7439 послідовність [пул 1: пептиди 1-7 (SEQ ID NOS:53-59), пул 2: пептиди 8-14 (SEQ ID NOS:60-66), пул 3: пептиди 1521 (SEQ ID NOS:67-73), пул 4: пептиди 22-28 (SEQ ID NOS:74-80), пул 5: пептиди 29-35 (SEQ ID NOS81-87), пул 6: пептиди 36-42 (SEQ ID NOS:88-94) та пул 7: пептиди 43-46 (SEQ ID NOS:9598)]. Крім того, людський HLA DR імунодомінантний пептид 24 (SEQ ID NO:76) аналізували індивідуально. Результати, представлені на Фігурах 16-19, показують, що та сама імуногенна ділянка CD4 та CD8 T-клітин (пептиди 22-28 + пептид 24) активувалася після імунізацій за допомогою кожної з двох CASB7439 конструкцій, рецептованих з AS15. Таким чином, пептид 24 також являє собою HLA A2.1 імунодомінантний пептид. Ці результати підтверджувалися у другому експерименті, в якому такі самі CASB7439 пептиди використовували для повторної стимуляції спленоцитів, одержаних від 5 індивідуальних мишей на групу, імунізованих при використанні або LVL168, або LVL144, рецептованих з AS15. (Дані не представлені.) Імуногенність CASB7439 у мишей: опосередкована CASB7439 гуморальна відповідь у дослідженні CB6f1 мишей Опосередкована CASB7439 гуморальна відповідь на 10 нг CASB7439 (LVL111), рецептованого або з AS01B, або з AS15, що використовується при 1/10 людської дози, оцінювали у інбредних CB6f1 мишей (Charles River, Senneville, QC, Canada). Дев‗ятнадцять мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта (2X концентрація). У мишей брали кров безпосередньо перед кожною імунізацією та через два тижні після четвертої імунізації. В іншому експерименті опосередковану CASB7439 гуморальну відповідь на LVL111, LVL168 або LVL144 оцінювали у інбредних CB6f1 мишей (Charles River, Senneville, QC, Canada). П‗ятдесят мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації при використанні 10 мкг CASB7439 рекомбінантного білка (LVL111, LVL168 або LVL144), рецептованого з AS15, або при використанні AS15, узятого окремо для контрольної групи. Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта у концентрації 2X, як описано будь-де в даній заявці. У мишей брали кров через два тижні після четвертої імунізації. Титри анти-CASB7439 lgG1 та lgG2a сироваткового антитіла визначали за допомогою ELISA згідно зі стандартними процедурами. Стисло, CASB7439 рекомбінантний білок у забуференому фосфатом фізіологічному розчині, вносили у планшет, що складається з 96 комірок, (PBS). Після блокування за допомогою 1% бичачого сироваткового альбуміну у PBS-Твін (01%), сироватку, одержану від iмунізованих мишей, інкубували у планшетах протягом однієї години при 37°C. Після промивання планшету додавали специфічне мишаче мічене антитіло проти козячого lgG1 або lgG2a-HRP l (Southern Biotech Associates, Birmingham, AL, USA) та інкубували протягом 1 години при 37°C. Після промивання до планшету додавали TMB субстратний реагент (BD Pharmingen, Mississauga, ON, Canada) на 15 хвилин. Зупиняли колориметричну реакцію шляхом додання 1M сірчаної кислоти. OD вимірювали при 450 нм за допомогою Spectramax спектрофотометра (Molecular Device, Sunnyvale, CA, USA). Титр антитіла визначали за допомогою стандартної кривої, при використанні специфічного загального захоплення IgG, lgG1 або lgG2a антитіл та внутрішньої позитивної за CASB7439 мишачої сироватки. Фігура 20/35 відображає аналіз у часі титрів lgG1 та lgG2a антитіл у сироватці мишей, імунізованих при використанні CASB7439 (LVL111), рецептованого або з AS01B, або при використанні AS15. Ці дані демонструють, що AS15 запускає більш високу Th1 відповідь (приблизно у десять разів вищу за співвідношення lgG2a/lgG1 після 4 iмунізацій у порівнянні із AS01B (lgG2a являє собою сурогатний маркер імунної відповіді типу 1). Як показано на Фігурі 21/35, CASB7439 білок, рецептований з AS15, запускає дуже сильний lgG1 та lgG2a 26 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 60 антитілогенез у інбредних CB6f1 мишей, незалежно від використовуваної конструкції (LVL111, LVL168 або LVL144). Імуногенність CASB7439 у мишей: опосередкована CASB7439 гуморальна відповідь CD1 у дослідженні мишей Оцінювали опосередковану CASB7439 гуморальну відповідь при імунізаціях CASB7439 (LVL111, LVL168 або LVL144) у аутбредних CD1 мишей (Charles River, Senneville, QC, Canada). Десять мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації при використанні 10 мкг CASB7439 рекомбінантного білка (LVL111, LVL168 або LVL144), рецептованого з AS15 (або тільки при використанні AS15, узятого окремо, як у контрольній групі). Кожна імунізація містила 25 мкл рекомбінантного CASB7439 білка, змішаного з 25 мкл ад‗юванта (2X концентрація), як описується будь-де у даній заявці. У мишей брали кров через два тижні після четвертої імунізації та визначали титри загального IgG антитіла проти CASB7439 у крові за допомогою ELISA, як вже було описано. CD1 миші, імунізовані при використанні різних CASB7439 конструкцій (LVL111, LVL168 або LVL144), розвивали дуже сильну специфічну для CASB7439 гуморальну імунну відповідь (Фігура 22/35). Ці дані вказують на те, що індукувалися у 8 та у 10 разів більш високі титри загального IgG антитіла проти CASB7439 при використанні LVL144 у порівнянні із LVL168 та LVL111, відповідно, у CD1 аутбредних мишей. Приклад 11 Протипухлинна ефективність CASB7439 ASCI у мишей: розробка моделі мишачої пухлини Мишачу модель пухлини, що експресує CASB7439 білок при використанні TC-1 мишачих пухлинних клітин або MC38 мишачих клітин колоректального раку, розробляли для дослідження опосередкованого CASB7439 in vivo захисту від пухлин у мишей. TC-1 пухлинні клітини були люб‗язно надані T. C. Wu (Johns Hopkins University, Baltimore, MD). Вони були одержані з первинних клітин легень C57BI/6 мишей шляхом послідовного переносу HPV16 E6 та E7 генів та активованого ras окогену так, як було описано раніше (Lin та ін. (1996) Cancer Res. Jan 1;56(1):21-6). MC38 клітини, клітинну лінію мишачої колоректальної аденокарциноми, закуповували у ATCC (Manassas, VA). Ці клітини стабільно трансфікували за допомогою конструкції, що кодує CASB7439, при використанні неліпосомального трансфектантного реагенту Fugene-6 (Roche, Mississauga, ON, Canada), у відповідності із рекомендаціями виробника. Стисло, плазміду (pcДНК3.1/Zeocine), що кодує цільну послідовність CASB7439 білка та містить ген резистентності до зеоцину, змішували з Fugene-6 та вносили до TC-1 або MC38 пухлинних клітин у середовищі, вільному від сироватки на ніч. Після трансфекції клітини залишали для відновлення на 24 годин у культуральному середовищі + 10% інакативованого прогріванням FBS. Клітини потім збирали та розподіляли у планшети на 96 комірок (50 мкл клітинної суспензії на комірку) для проведення граничних розведень. Після висівання клітин середовище замінювали на культуральне середовище + 10% інакативованого прогріванням FBS, що містить 100 мкг/мл зеоцину (Invitrogen, Burlington, ON, Canada). Після проведення відбору кожний клон збирали, залишали для вирощування у планшетах на 24 комірки та заморожували. Експресію іРНК CASB7439 у TC1/CASB7439 та MC38/CASB7439 клітинах підтверджували за допомогою кількісної ПЛР у реальному часі при використанні специфічних для CASB7439 TaqMan праймерів та зонду. Кількісні ПЛР реакції у реальному часі проводили за допомогою TaqMan реакційного набору при використанні TaqMan 7900 пристрою (Applied Biosystem, Foster City, CA, USA). Експресію CASB7439 білка оцінювали шляхом Вестерн-блоттінгу та імунофлуоресценції при використанні специфічного анти-CASB7439 моноклонального антитіла (40-12). TC-1/CASB7439#14 популяція клітин не була повністю клональною, проте складалася принаймні з двох різних субклонів. Tаким чином, інший етап граничного розведення проводили для того, щоб одержати повністю клональну популяцію TC-1/CASB7439 клітин: TC- 1 /CASB7439#14-2 клітини. Для визначення оптимальної дози TC-1 або MC38 клітин для зараження їх пухлинами 5 6 мишей, різні дози клітин (від 1x10 дo 2 x 10 клітин на мишу) вводили CB6f1 інбредним мишам. 5 Оптимальну дозу визначали як таку, що складає 2,5 x 10 клітин на мишу, для зараження TC-1 5 /CASB7439#14 та TC1/CASB7439#14-2; доза, що складає 1 x 10 клітин на мишу, була виявлена як оптимальна для зараження MC38/CASB7439#35 клітинами. Протипухлинна ефективність у мишей: експерименти з імунізації та зараження Серію, що складалася із чотирьох досліджень (1.1-1.4), проводили для оцінки опосередкованого CASB7439 in vivo захисту від пухлини. В загальному випадку у кожному із цих чотирьох досліджень групи CB6f1 мишей одержували чотири внутрішньом‗язові імунізації з двотижневими інтервалами, з використанням різних доз (0, 1, 10 або 30 мкг) специфічного 27 UA 99801 C2 5 10 15 20 25 30 35 40 45 50 55 рекомбінантного CASB7439 білка, вибраного для дослідження, рецептованого або з AS01B, або з AS15 при заключній концентрації, так, як описано скрізь у даній заявці, буфера на основі фізіологічного розчину, CASB7439 білка, узятого окремо, або ад‗ювантів, узятих окремо. Через два тижні після четвертої імунізації CB6f1 мишей заражали шляхом підшкірної ін‗єкції 250000 TC-1/CASB7439 #14 клітинами, 100000 клітинами MC38/CASB7439#35 або 250000 пухлинними клітинами TC-1/CASB7439 #14-2. Після зараження розмір пухлин вимірювали тричі на тиждень 2 у кожної миші. Згідно з прописом мишей з пухлинами, більшими за 200 мм , забивали. Потім будували криві виживання. Також здійснювали три додаткові дослідження (2.1-2.4) для оцінки доз, відмінних від 0, 1, 10 або 30 мкг рекомбінантного CASB7439 білка. Дослідження 2.4 оцінювало кореляцію між імуногенністю та захистом від пухлини за допомогою CASB7439 у самців та самок C57BI/6 мишей; CB6f1 мишей піддавали оцінці паралельно. Дослідження 1.1-2.4 є описаними більш детально нижче. Дослідження #1.1 Заявники досліджували захист від пухлин, що забезпечується шляхом імунізації за допомогою LVL111 (1 або 10 нг), рецептованого або з AS01B, або з AS1, проти зараження TC1/CASB7439 #14 у CB6f1 мишей. Десять мишей на групу кожні два тижні одержували чотири внутрішньом‗язові імунізації за допомогою CASB7439 білка, рецептованого або з AS01B, або з AS15, при заключній концентрації, так, як описано будь-де у даній заявці. Дві контрольні групи, імунізовані при використанні або буфера на основі фізіологічного розчину, або при використанні CASB7439 білка, узятого самостійно, також досліджували. Через два тижні після четвертої імунізації, кожну мишу підшкірно заражали за допомогою 250000 клітин TC-1/CASB7439#14. Ріст пухлин продовжувався протягом 42 днів, проте не будували ніяких кривих виживання, оскільки тільки декілька мишей забивали на 42 день. Але криві росту пухлин будували для кожної групи. Фігура 23/35, верхня та нижня частини, представляють криві росту пухлин, одержані при використанні композицій з AS01B та AS15, відповідно. (Примітка: AS15 є помилково ідентифікованим на цій фігурі як "AS015".). Криві демонструють, що у CB6f1 мишей, які одержували CASB7439 білок, рецептований або з AS01B, або з AS15, пухлинний ріст TC1/CASB7439#14 знижувався. Дослідження #1.2 Групи з чотирнадцяти мишей одержували різні дозування LVL111 (1,10 та 30 мкг) або контроль. Мишей внутрішньовенно імунізували чотири рази з інтервалами у два тижні різними дозами LVL111, рецептованого або з AS01B, або з AS15 при заключній концентрації так, як описано будь-де у даній заявці. Чотири контрольні групи також були імунізовані при використанні тільки AS01B, тільки AS15, тільки буфера на основі фізіологічного розчину, або 30 мкг тільки LVL111. Через два тижні після четвертої імунізації кожну мишу підшкірно заражали 250000 клітинами TC-1 /CASB7439#14. Після зараження розмір пухлин вимірювали тричі на 2 тиждень у кожної миші. Згідно з прописом мишей з пухлинами, більшими за 200 мм забивали. Потім будували криві виживання. Ці результати свідчать про те, що миші, імунізовані при використанні LVL111, рецептованого з AS15, були краще захищені від пухлинного зараження TC1/CASB7439#14, ніж миші імунізовані при використанні LVL111, рецептованого з AS01B. Оптимальні результати для CB6f1 мишей були одержані при використанні 10 мкг LVL111, рецептованого з AS15. Див. Фігуру 24/35. Дослідження #1.3 Тридцять вісім CB6f1 мишей на групу внутрішньом‗язово імунізували чотири рази з інтервалами у два тижні при використанні 10 нг LVL111, LVL168 або LVL144, рецептованих з AS15 при заключній концентрації так, як описано будь-де у даній заявці. Також досліджували контрольну групу, імунізовану при використанні AS15, узятого окремо. Через два тижні після четвертої імунізації двадцять вісім мишей підшкірно заражали 250000 клітинами TC-1 /CASB7439#14 та десять мишей, що залишилися, підшкірно заражали 100000 клітин MC38/CASB7439#35. Після зараження розмір пухлин вимірювали тричі на тиждень у 2 кожної миші. Згідно з прописом мишей з пухлинами, більшими за 200 мм , забивали. Ці результати свідчать про те, що LVL111, LVL168 та LVL144, рецептовані з AS15, усі є протективними щодо зараження TC1/CASB7439#14 або MC38/CASB7439#35. Див. Фігуру 25/35. Дослідження #1.4 Шістдесят шість CB6f1 мишей на групу внутрішньом‗язово імунізували чотири рази з інтервалами у два тижні при використанні 10 мкг LVL168 або LVL144, рецептованого з AS15 при заключній концентрації так, як описано будь-де у даній заявці. Також досліджували контрольну групу, імунізовану при використанні AS15, узятого окремо. 28
ДивитисяДодаткова інформація
Назва патенту англійськоюProtein construct comprising a modified casb7439 polypeptide
Автори англійськоюBlais, Normand, Harvey, Matrine, Pilorget, Anthony, Rioux, Clement
Назва патенту російськоюБелковая конструкция, которая включает модифицированный полипептид casb7439
Автори російськоюБлейс Норманд, Гарви Мартин, Пилорджет Энтони, Риоу Клемен
МПК / Мітки
МПК: A61K 38/17, C07K 14/47, A61P 37/04, A61P 35/00
Мітки: конструкція, поліпептид, включає, білкова, casb7439, модифікований, яка
Код посилання
<a href="https://ua.patents.su/95-99801-bilkova-konstrukciya-yaka-vklyuchaeh-modifikovanijj-polipeptid-casb7439.html" target="_blank" rel="follow" title="База патентів України">Білкова конструкція, яка включає модифікований поліпептид casb7439</a>
Послідовність днк, що кодує поліпептид, який зв’язується з mort-1 (варіанти), вектор експресії, що включає послідовність днк, клітина- хазяїн, виділений поліпептид та фармацевтична композиція, що містить поліп

Номер патенту: 72423
Опубліковано: 15.03.2005
Автори: Гончаров Таня, Воллах Девід, Гольцев Юрій, Болдін Марк
МПК: C07K 14/705, C07K 14/47, A61P 35/00, A61K 45/00, A61P 31/12, C12N 9/64, C07K 16/28, C12N 15/09, A61K 48/00, C07K 14/715, C12N 5/10
Мітки: поліп, містить, варіанти, послідовність, експресії, вектор, композиція, клітина, поліпептид, виділений, зв'язується, днк, включає, хазяїн, mort-1, кодує, фармацевтична
Формула / Реферат:
1. Послідовність ДНК, що кодує поліпептид, який зв'язується з MORT-1, де MORT-1 є білком, який зв'язується з внутрішньоклітинним доменом FAS-R, і який зв'язується з білком TRADD, який зв'язується з внутрішньоклітинним доменом p55-TNF-R, де зазначений поліпептид має:a) послідовність, що містить залишки 1-182 послідовності SEQ ID №5;b) послідовність, що містить фрагмент а), де фрагмент зв'язується з MORT-1;c) послідовність,...
Рекомбінантний модифікований генно-інженерний поліпептид hccore-25, що імітує структурний білок нуклеокапсиду вірусу гепатиту с

Номер патенту: 60424
Опубліковано: 25.06.2011
Автори: Липова Надія Миколаївна, Ніколаєнко Ігор Васильович, Раєвська Галина Євгенівна
МПК: C12N 15/70, C12N 15/51
Мітки: нуклеокапсиду, поліпептид, структурний, вірусу, рекомбінантний, імітує, гепатиту, модифікований, білок, hccore-25, генно-інженерний
Формула / Реферат:
1. Рекомбінантний білок НСсоrе-25, продукований бактеріями Е.соlі, що імітує структурний білок нуклеокапсиду вірусу гепатиту С генотипу 1b, який містить послідовності олігогістидинового пептиду (His)6, складається зі 167 амінокислотних залишків (а.з.), має молекулярну масу 18,7 кДа, ізоелектричну точку РІ 12,04:MGSSHHHHHH SSGLVPRGSH MASMTGGQQM GRGSMSTNPK PQRKTKRNTN RRPQDVKFPG GGQIVGGVYL LPRRGPRLGV RATRKTSERS QPRGRRQPIP KARHPEGRTW...
Рекомбінантний очищений модифікований генно-інженерний поліпептид, що імітує основні імунодомінантні епітопи корового білка gag вірусу імунодефіциту людини

Номер патенту: 17186
Опубліковано: 15.09.2006
Автори: Кобозев Юрій Олексійович, Грабченко Наталія Іванівна, Іванська Наіля Валєєвна, Шевчук Олександр Анатолійович, Трояновський Борис Моісейович, Семиноженко Володимир Петрович, Пилипенко Віталій Григорович, Трохимчук Тетяна Юріївна, Горлов Юрій Іванович
МПК: C08H 5/00
Мітки: генно-інженерний, основні, білка, очищений, імунодефіциту, вірусу, модифікований, рекомбінантний, поліпептид, корового, імунодомінантні, людини, епітопи, імітує
Формула / Реферат:
Рекомбінантний очищений модифікований генно-інженерний поліпептид, що імітує основні імунодомінантні епітопи корового білка Gag вірусу імунодефіциту людини, який містить основні імунодомінантні епітопи, притаманні нативному коровому білку Gag ВІЛ-1, з молекулярною масою 45,4кД, має ізоелектричну точку 5,9, відповідає загальній формулі: N-(HIS)6-Gag-COOH і складається з 432 амінокислотних залишків:
Модифікований хімеричний поліпептид з покращеними фармакокінетичними властивостями

Номер патенту: 74146
Опубліковано: 15.11.2005
Автори: Янкопулос Джордж Д., Девіс Самуел, Пападопулос Ніколас Дж.
МПК: A61K 38/22, A61P 17/02, A61P 29/00, A61K 48/00, A61P 13/12, A61P 35/00, C12N 1/21, A61P 9/00, C07K 19/00, A61P 31/00, A61P 27/02, C12N 15/12, A61P 17/00, A61P 19/02, C12N 1/19, C12N 5/10, C12N 15/09, C07K 14/71, A61P 43/00, A61P 11/06, C07K 16/00, C12P 21/02
Мітки: властивостями, фармакокінетичними, поліпептид, покращеними, хімеричний, модифікований
Формула / Реферат:
1. Молекула нуклеїнової кислоти, що кодує злитий поліпептид, що має здатність до зв'язування VEGF поліпептиду, яка містить:(a) нуклеотидну послідовність, яка кодує компонент першого рецептора фактора росту ендотеліальних клітин судин (VEGF), що переважно містить імуноглобуліновий (Ig) домен 2 першого VEGF рецептора, оперативно зв'язану з(b) нуклеотидною послідовністю, що кодує компонент другого VEGF рецептора, що переважно...
Трансгенна рослинна клітина, що включає поліпептид з активністю гліфосат-n-ацетилтрансферази

Номер патенту: 94688
Опубліковано: 25.05.2011
Автори: Гайвер Лорайн Дж., Паттен Філліп А., Дак Ніколас Б., Сайхл Ден, Айві Крістіна, МакКатчен Біллі Ф., Кембл Роджер, Касл Лінда А., Чен Янг Хонг, Міншулл Джеремі
МПК: C12N 9/10, A01H 5/00, C12N 15/82, C12N 15/54, C12N 5/14
Мітки: поліпептид, гліфосат-n-ацетилтрансферази, включає, рослинна, активністю, клітина, трансгенна
Формула / Реферат:
1. Трансгенна рослинна клітина, що включає гетерологiчний поліпептид з активністю глiфосат-N-ацетилтрансферази, де вказана рослинна клітина включає метаболічний продукт гліфосату, який являє собою N-ацетилгліфосат після обробки гліфосатом.2. Трансгенна рослинна клітина за п. 1, де рослинна клітина виявляє підвищену стійкість до глiфосату порівняно з клітиною дикої рослини того ж виду, лінією або культурним сортом.3. Трансгенна...
Попередній патент: Спосіб оптимізації технологічного навантаження на родючий ґрунт
Наступний патент: Установка для доїння овець
Випадковий патент: Вузол з'єднання труб