Спосіб здійснення експресії амінопептидазного антигену н110д
Номер патенту: 32437
Опубліковано: 15.12.2000
Автори: Манн Едвард Альберт, Грехем Маргарет, Ролф Тімоті Пітер, Сміт Тревор Стенлі, Ньютон С'юзен Елізабет

















Текст
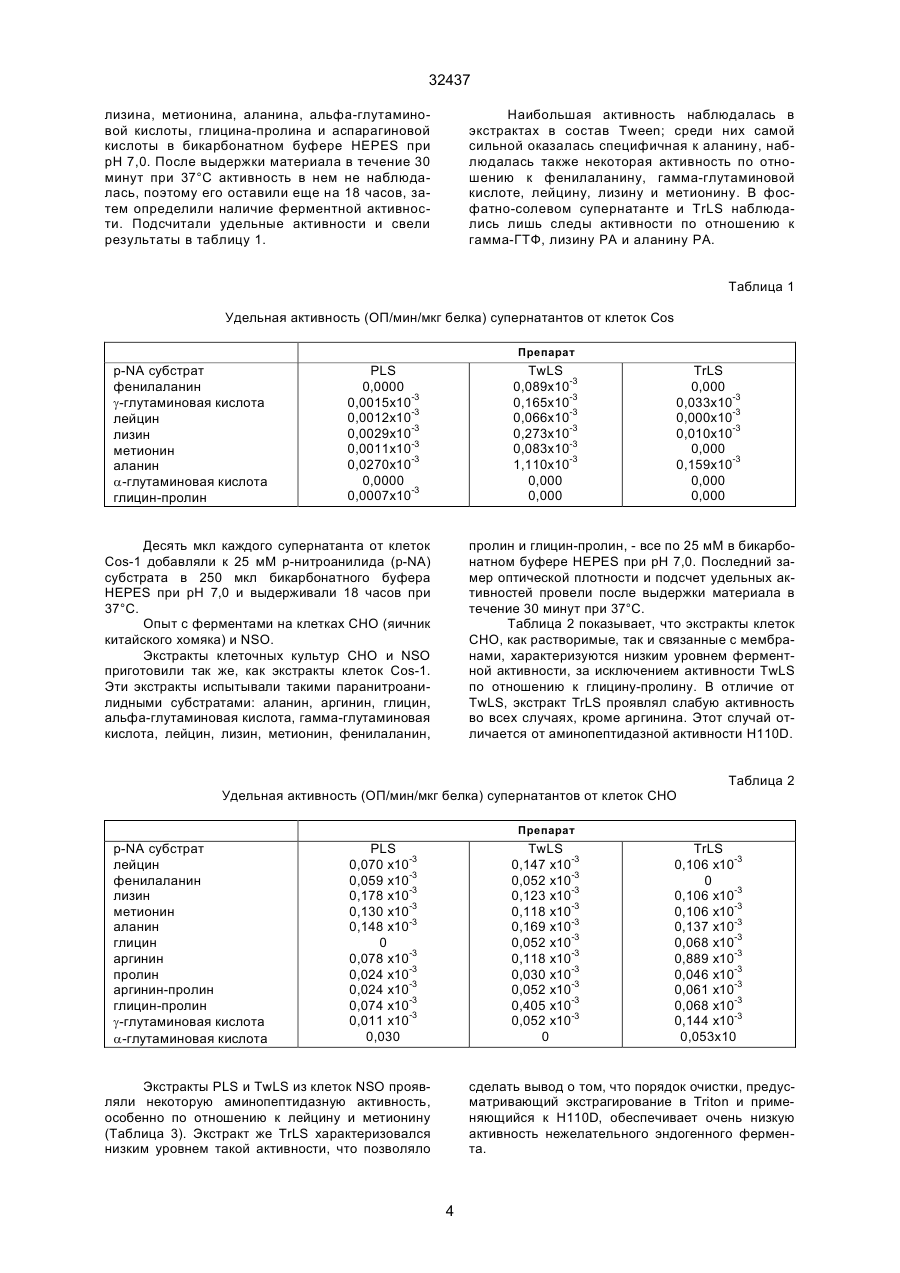
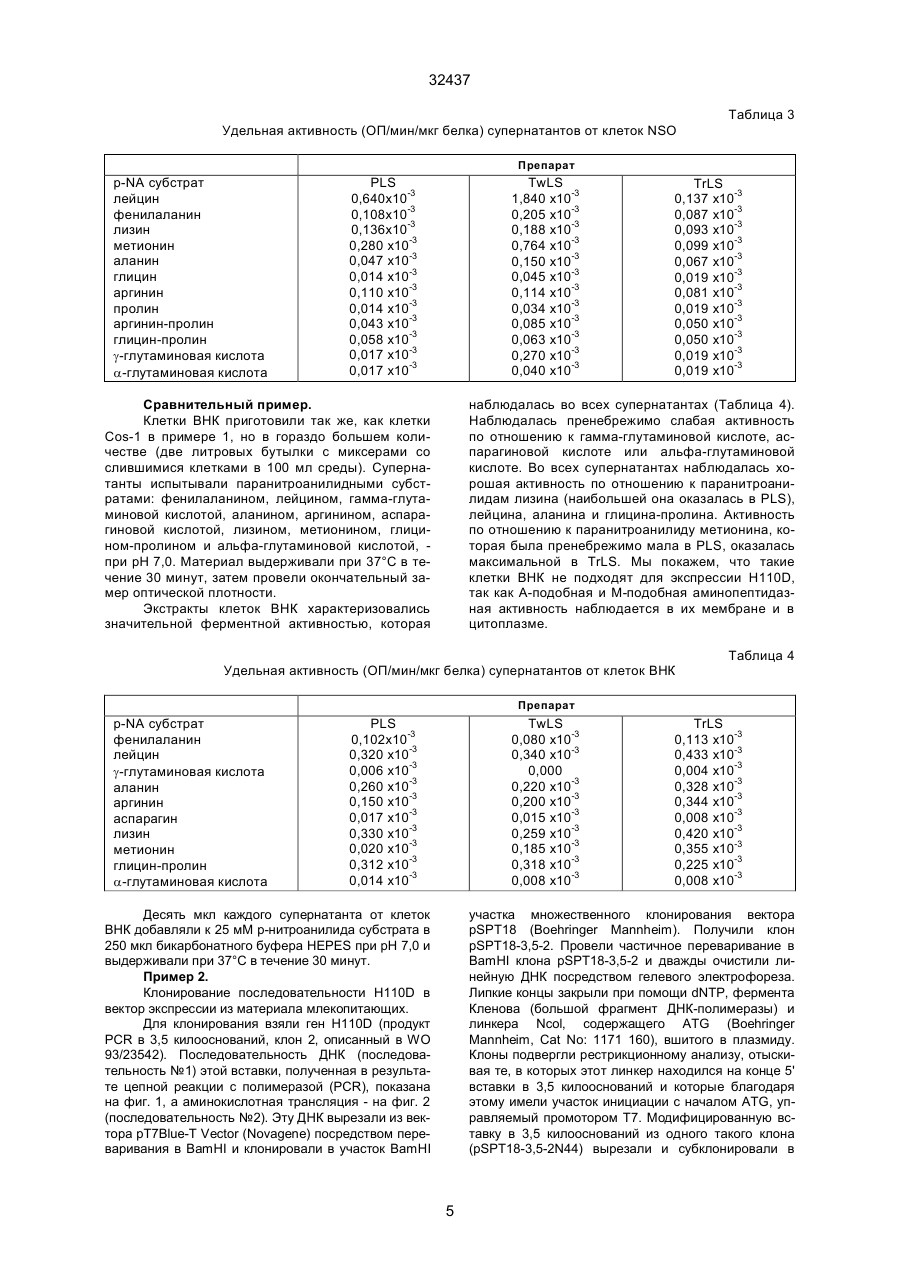
1. Способ осуществления экспрессии аминопептидазного антигена H110Д или его фрагмента, обладающего сходной энзиматической и/или анти C2 (54) СПОСІБ ЗДІЙСНЕННЯ ЕКСПРЕСІЇ АМІНОПЕПТИДАЗНОГО АНТИГЕНУ Н110Д 32437 тепсин. Они присутствуют в кишках многих паразитов, например, гельминтов, таких как различные виды семейств Haemonchus, Ostertagia, Trichostrongylus, Nematodirus, Dictуocaulus, Cooperia, Ascaris, Dirofilaria, Trichuris, Strongylus и Fasciola, членистоногих, особенно представителей классов паукоообразных и насекомых, в частности, эктопаразитов, таких как кровососущие насекомые (например, члены отделов насекомых с полным и неполным превращением), мухи, такие как мясная муха (Lucilia), мухи myiasis и сосущие, вши, клещи, блохи, кровососки и клопы. В документе W093/23542, на который мы ссылались выше, описано изготовление фрагментов антигена Н110D методом рекомбинации ДНК. Согласно описанной технологии, материал H110D экспрессировали в Е. coli при помощи вектора pGEX и вводили овцам, после чего у них образовывались антитела к Н110D. В ходе дальнейшей работы, используя бакуловирус в клетке-хозяине (клетки Sf9), мы успешно экспрессировали клон гена H110D длиной в 3,5 килооснований. Соответствующая ему последовательность нуклеотидов показана на фиг. 1 (последовательность №1). Соответствующая трансляция показана на фиг. 2 (последовательность №2). Однако, для приготовления вакцин, вводимых животным, предпочтительно экспрессировать антиген в клетках млекопитающих. Это обеспечивает хорошее воспроизведение нативной формы и защитного эпитопа антигена, так как эукариотическая система экспрессии дает более сходные схемы гликозилирования, дисульфидного связывания и других посттрансляционных преобразований, чем Е. coli, которые образуют нерастворимый белок, требующий другой укладки и плохо воспроизводящий нативную форму. Кроме того, гликозилирование в клетках млекопитающих с меньшей вероятностью вызывает иммунную реакцию, которая могла бы помешать защитной противобелковой реакции, как это случается с материалом, полученным из клеточной линии насекомых, где гликозилирование происходит совсем по-другому. Итак, для защиты человека и домашних животных предпочтительно использовать клеточные линии человеческих или животных фибробластов или миелом, такие как HeLa - человеческая клеточная линия; ВНК - клетки почек детенышей хомяка; VERO и COS - обезьянья почечная клеточная линия; FR3T3 -фибробласты крыс Фишера, NIH3T3 - линия мышиных фибробластов; С127І клеточная линия опухоли мышиной молочной железы; CV-1 - фибробласты почек африканской зеленой мартышки; ЗТ6 - фибробласты мышиных зародышей; L-клетки - мышиная клеточная линия; СНО - клеточная линия яичников китайского хомяка; NSO NSI, SP2 и другие клеточные линии мышиных миелом, а также клеточные линии крысиных миелом, например, YB2/0 и Y3. Поскольку скрытые антигены, которые необходимо получить, играют ведущую роль в переваривании паразитом пищи, ферменты или другие функциональные белки, обладающие теми же свойствами, присутствуют в различных эукариотических клеточных линиях и потому не только мешают отбору клонов, продуцирующих нужный антиген, но и затрудняют выделение этого антигена из продуктов жизнедеятельности клетки. Кроме того, избранная клеточная линияхозяин неизбежно окажется генетически близкой - если говорить о последовательности полипептидов - животному, которое необходимо иммунизировать, так что заражение искомого чужого скрытого антигена таким собственным антигеном с большой вероятностью вызовет нежелательную аутоиммунную реакцию со стороны хозяина. И наконец, контроль качества экспрессированного скрытого антигена будет затруднен. Настоящее изобретение исходит из того, что экспрессию рекомбинантной ДНК искомого чужого скрытого ферментного антигена следует проводить в преобразованной линии клеток млекопитающего-хозяина, которая, когда оба фермента связаны с клеточной мембраной или оба находятся в цитоплазме и физико-химическое разъединение их исключается, практически не содержит эндогенных антигенов, обладающих теми же ферментными свойствами, что и чужой скрытый антиген. Поэтому, в соответствии с настоящим изобретением, мы предлагаем способ экспрессии антигена-фермента, который в природе является ферментом, связанным с мембранами кишечника паразитов, или его фрагментом, обладающим теми же ферментными и/или антигенными свойствами; этот способ предусматривает трансфекцию клеток-хозяев, взятых у млекопитающих, вектором, подготовленным к экспрессии упомянутого антигена-фермента или его фрагмента, характеризующийся тем, что перед трансформацией клетки-хозяева практически не содержат эндогенных ферментов, которые а) имеют ту же функцию и б) связаны или не связаны с мембраной так же, как и упомянутый паразитный антиген-фермент или его фрагмент. Таким образом, если эндогенный фермент, такой как аминопептидаза, содержится в цитоплазме клетки-хозяина, а чужой фермент экспрессируется трансмембранной последовательностью, которая остается в мембране клетки-хозяина, то разъединить эндогенный и чужой фермент нетрудно: нужно отобрать из клеток мембраны, например, посредством центрифугирования. С другой стороны, чужой антиген можно модифицировать - удалить участок связывания с мембраной, и если клетка-хозяин содержит в мембране эндогенный фермент, обладающий теми же свойствами, что и чужой, то разъединить их также можно путем отбора клеточной мембраны; так, последовательность, кодирующую трансмембранный участок паразитного фермента, можно заместить - в гене, который нужно экспрессировать, - сигнальной последовательностью, вызывающей секрецию. Особый интерес представляют глистные антигены-аминопептидазы, такие как H110D, упомянутый выше, и его фрагменты. Их можно экспрессировать в различных клетках млекопитающих при помощи подходящих векторов. Для антигена к H.contortus H110D, который проявляет преимущественно А-подобную и М-подоб 2 32437 ную аминопептидазную активность и потому расщепляет преимущественно пептидные связи метионина и лейцина, мы подобрали клетки COS-1: А-подобная и М-подобная аминопептидазная активность в них незначительна, и их можно использовать в соответствии с настоящим изобретением. Выяснилось, что в них содержится фермент-аминопептидаза, расщепляющий пептидные связи аланина и слабо связанный с мембраной, поэтому эндогенный фермент нетрудно отделить от экспрессируемого H110D, содержащегося в мембране. Оказалось, что протеолитические ферменты, такие как трипсин, отщепляют паразитный антиген Н110D от мембраны, в результате чего получается растворимый Н110D (Н11S). Таким образом, эти ферменты можно использовать для дифференциального отделения чужого скрытого антигена от эндогенного фермента со сходной активностью. Клеточные линии для использования в соответствии с настоящим изобретением можно выбрать из существующих штаммов путем поиска профилей подходящей активности и/или обнаружения нужного фермента в мембране или цитоплазме. В последнем случае связь с мембраной можно установить путем экстрагирования клеток после лизиса сначала детергентом, таким как Tween, который не вымывает мембранные ферменты, а затем детергентом, таким как Triton, который высвобождает эти ферменты. Можно также вырастить линию клеток млекопитающего с пониженным содержанием какого-либо фермента. Один из способов сделать это таков: культивируют антитела к ферменту млекопитающего, который нужно удалить. Клеточные линии модифицируют облучением или химическими средствами, вызывая точечные мутации. Клетки культивируют в специальной среде, компенсирующей действие утраченного фермента. Затем клетки приводят во взаимодействие с антителами, меченными флуоресцентным веществом и пропускают через прибор для сортировки клеток по флуоресценции Fluorescence Activated Cell Sorter (FACS). Клетки с отрицательной флуоресценцией клонируют и снова подвергают селекции, отслеживая стабильность потери ферментных свойств. Клетки также можно модифицировать путем удаления гена или рациональной (направленной) мутации, удаляющей или изменяющей ген эндогенного фермента. Векторы, подходящие к различным классам клеточных линий из клеток млекопитающих, хорошо известны специалистам. Как правило, они состоят из промотора и/или энхансера (гена-усилителя), оперативно соединенного с геном, экспрессирующим антиген-фермент или его фрагмент. Так, в частности, ген фрагмента Н110 в 3,5 килооснований можно соединить в рамке с подходящим промотором. К подходящим промоторам относятся ранний или поздний промотор SV40, например, вектор PSVL, цитомегаловирус, мышиный металлотионеин I и длинный концевой повтор вируса опухоли мышиной молочной железы. Вектор предпочти тельно включает подходящий маркер, например, ген дигидрофолат-редуктазы или глутаминсинтетазы. Векторы этих типов описаны в документах W086/05807, W087/04462, W089/01036 и W089/10404. Трансфекцию клеток-хозяев можно осуществить стандартными методами, например, с использованием фосфата кальция, декстрана диэтиламиноэтила, полибрена, слияния протопластов, липосом, при помощи непосредственной микроинъекции, генной пушки или электропорации. Предпочтительным является последний метод. Способы трансфекции клеток млекопитающих путем электропорации описаны y Andreason G.L. и Evans G.A., Introduction and expression of DNA molecules in eukaryotic cells by electroporation, Biotechniques 6, 650, 1980. В общем случае линейная ДНК вводится легче, чем кольцевая. Антиген Н110D проявляет лейцинаминопептидазные (М-подобные) и метионинаминопептидазные (А-подобные) свойства и в общем случае предпочтительными являются условия, когда клетка-хозяин лишена хотя бы аминопептидазных свойств этих типов. Приведенные ниже примеры служат исключительно иллюстрацией к настоящему изобретению. В этих примерах на фигурах изображено следующее: Фигура 1 - изображение последовательности ДНК в 3,5 килооснований (продукт цепной реакции с полимеразой (PCR), клон 2) - последовательности №1. Фигура 2 - изображение аминокислотной трансляции продукта цепной реакции с полимеразой в 3,5 килооснований, клона 2, (последовательность №2). Пример 1. Опыт с ферментами на клетках Cos-1. Клетки Cos-1 были получены из университета графства Суррей в 5 мл среды Игла, модифицированной по способу Дульбекко (DMEM). Их рассадили в две 200-миллилитровые чашки, содержащие по 11 мл среды, и выращивали, пока они не слились. Клетки Cos-1 цепкие; их отбирали из чашек путем аспирации при помощи стеклянной пастеровской пипетки. Затем их помещали в 10 мл забуференного фосфатом физиологического раствора (ЗФР). Когда все клетки оказались в суспензии, их переместили в универсальную пробирку и заморозили при минус 20°С. Несколько раз их замораживали-размораживали в жидком азоте, чтобы они лопнули, затем центрифугировали, чтобы отделить фосфатно-солевой супернатант (PLS). Последующие экстрагирования в 500 мкл сначала ЗФР 0,1% Tween, затем ЗФР 2% Triton дали супернатанты TwLS и TrLS. Все супернатанты сконцентрировали до 200 мкл при помощи микроконцентраторов Millipore. Сам по себе ЗФР экстрагирует ферменты, свободно содержащиеся в цитоплазме, Tween экстрагирует ферменты, которые могут иметь слабую связь с клеточной мембраной, a Triton экстрагирует мембранные белки. Супернатанты испытывали порциями по 25 мМ р-нитроанилидов субстратов: фенилаланина, гамма-глутаминовой кислоты, лейцина, 3 32437 лизина, метионина, аланина, альфа-глутаминовой кислоты, глицина-пролина и аспарагиновой кислоты в бикарбонатном буфере HEPES при рН 7,0. После выдержки материала в течение 30 минут при 37°С активность в нем не наблюдалась, поэтому его оставили еще на 18 часов, затем определили наличие ферментной активности. Подсчитали удельные активности и свели результаты в таблицу 1. Наибольшая активность наблюдалась в экстрактах в состав Tween; среди них самой сильной оказалась специфичная к аланину, наблюдалась также некоторая активность по отношению к фенилаланину, гамма-глутаминовой кислоте, лейцину, лизину и метионину. В фосфатно-солевом супернатанте и TrLS наблюдались лишь следы активности по отношению к гамма-ГТФ, лизину РА и аланину РА. Таблица 1 Удельная активность (ОП/мин/мкг белка) супернатантов от клеток Cos Препарат р-NA субстрат фенилаланин g-глутаминовая кислота лейцин лизин метионин аланин a-глутаминовая кислота глицин-пролин PLS 0,0000 0,0015x10-3 0,0012x10-3 0,0029x10-3 0,0011x10-3 0,0270x10-3 0,0000 0,0007x10-3 TwLS 0,089x10-3 0,165x10-3 0,066x10-3 0,273x10-3 0,083x10-3 1,110x10-3 0,000 0,000 Десять мкл каждого супернатанта от клеток Cos-1 добавляли к 25 мМ р-нитроанилида (р-NA) субстрата в 250 мкл бикарбонатного буфера HEPES при рН 7,0 и выдерживали 18 часов при 37°С. Опыт с ферментами на клетках СНО (яичник китайского хомяка) и NSO. Экстракты клеточных культур СНО и NSO приготовили так же, как экстракты клеток Cos-1. Эти экстракты испытывали такими паранитроанилидными субстратами: аланин, аргинин, глицин, альфа-глутаминовая кислота, гамма-глутаминовая кислота, лейцин, лизин, метионин, фенилаланин, TrLS 0,000 0,033x10-3 0,000x10-3 0,010x10-3 0,000 0,159x10-3 0,000 0,000 пролин и глицин-пролин, - все по 25 мМ в бикарбонатном буфере HEPES при рН 7,0. Последний замер оптической плотности и подсчет удельных активностей провели после выдержки материала в течение 30 минут при 37°С. Таблица 2 показывает, что экстракты клеток СНО, как растворимые, так и связанные с мембранами, характеризуются низким уровнем ферментной активности, за исключением активности TwLS по отношению к глицину-пролину. В отличие от TwLS, экстракт TrLS проявлял слабую активность во всех случаях, кроме аргинина. Этот случай отличается от аминопептидазной активности H110D. Таблица 2 Удельная активность (ОП/мин/мкг белка) супернатантов от клеток СНО Препарат p-NA субстрат лейцин фенилаланин лизин метионин аланин глицин аргинин пролин аргинин-пролин глицин-пролин g-глутаминовая кислота a-глутаминовая кислота PLS 0,070 x10-3 0,059 x10-3 0,178 x10-3 0,130 x10-3 0,148 x10-3 0 0,078 x10-3 0,024 x10-3 0,024 x10-3 0,074 x10-3 0,011 x10-3 0,030 TwLS 0,147 x10-3 0,052 x10-3 0,123 x10-3 0,118 x10-3 0,169 x10-3 0,052 x10-3 0,118 x10-3 0,030 x10-3 0,052 x10-3 0,405 x10-3 0,052 x10-3 0 Экстракты PLS и TwLS из клеток NSO проявляли некоторую аминопептидазную активность, особенно по отношению к лейцину и метионину (Таблица 3). Экстракт же TrLS характеризовался низким уровнем такой активности, что позволяло TrLS 0,106 x10-3 0 0,106 x10-3 0,106 x10-3 0,137 x10-3 0,068 x10-3 0,889 x10-3 0,046 x10-3 0,061 x10-3 0,068 x10-3 0,144 x10-3 0,053x10 сделать вывод о том, что порядок очистки, предусматривающий экстрагирование в Triton и применяющийся к H110D, обеспечивает очень низкую активность нежелательного эндогенного фермента. 4 32437 Таблица 3 Удельная активность (ОП/мин/мкг белка) супернатантов от клеток NSO Препарат p-NA субстрат лейцин фенилаланин лизин метионин аланин глицин аргинин пролин аргинин-пролин глицин-пролин g-глутаминовая кислота a-глутаминовая кислота PLS 0,640x10-3 0,108x10-3 0,136x10-3 0,280 x10-3 0,047 x10-3 0,014 x10-3 0,110 x10-3 0,014 x10-3 0,043 x10-3 0,058 x10-3 0,017 x10-3 0,017 x10-3 TwLS 1,840 x10-3 0,205 x10-3 0,188 x10-3 0,764 x10-3 0,150 x10-3 0,045 x10-3 0,114 x10-3 0,034 x10-3 0,085 x10-3 0,063 x10-3 0,270 x10-3 0,040 x10-3 Сравнительный пример. Клетки ВНК приготовили так же, как клетки Cos-1 в примере 1, но в гораздо большем количестве (две литровых бутылки с миксерами со слившимися клетками в 100 мл среды). Супернатанты испытывали паранитроанилидными субстратами: фенилаланином, лейцином, гамма-глутаминовой кислотой, аланином, аргинином, аспарагиновой кислотой, лизином, метионином, глицином-пролином и альфа-глутаминовой кислотой, при рН 7,0. Материал выдерживали при 37°С в течение 30 минут, затем провели окончательный замер оптической плотности. Экстракты клеток ВНК характеризовались значительной ферментной активностью, которая TrLS 0,137 x10-3 0,087 x10-3 0,093 x10-3 0,099 x10-3 0,067 x10-3 0,019 x10-3 0,081 x10-3 0,019 x10-3 0,050 x10-3 0,050 x10-3 0,019 x10-3 0,019 x10-3 наблюдалась во всех супернатантах (Таблица 4). Наблюдалась пренебрежимо слабая активность по отношению к гамма-глутаминовой кислоте, аспарагиновой кислоте или альфа-глутаминовой кислоте. Во всех супернатантах наблюдалась хорошая активность по отношению к паранитроанилидам лизина (наибольшей она оказалась в PLS), лейцина, аланина и глицина-пролина. Активность по отношению к паранитроанилиду метионина, которая была пренебрежимо мала в PLS, оказалась максимальной в TrLS. Мы покажем, что такие клетки ВНК не подходят для экспрессии Н110D, так как А-подобная и М-подобная аминопептидазная активность наблюдается в их мембране и в цитоплазме. Таблица 4 Удельная активность (ОП/мин/мкг белка) супернатантов от клеток ВНК Препарат p-NA субстрат фенилаланин лейцин g-глутаминовая кислота аланин аргинин аспарагин лизин метионин глицин-пролин a-глутаминовая кислота PLS 0,102x10-3 0,320 x10-3 0,006 x10-3 0,260 x10-3 0,150 x10-3 0,017 x10-3 0,330 x10-3 0,020 x10-3 0,312 x10-3 0,014 x10-3 TwLS 0,080 x10-3 0,340 x10-3 0,000 0,220 x10-3 0,200 x10-3 0,015 x10-3 0,259 x10-3 0,185 x10-3 0,318 x10-3 0,008 x10-3 TrLS 0,113 x10-3 0,433 x10-3 0,004 x10-3 0,328 x10-3 0,344 x10-3 0,008 x10-3 0,420 x10-3 0,355 x10-3 0,225 x10-3 0,008 x10-3 участка множественного клонирования вектора pSPT18 (Boehringer Mannheim). Получили клон pSPT18-3,5-2. Провели частичное переваривание в BamHI клона pSPT18-3,5-2 и дважды очистили линейную ДНК посредством гелевого электрофореза. Липкие концы закрыли при помощи dNTP, фермента Кленова (большой фрагмент ДНК-полимеразы) и линкера Ncol, содержащего ATG (Boehringer Mannheim, Cat No: 1171 160), вшитого в плазмиду. Клоны подвергли рестрикционному анализу, отыскивая те, в которых этот линкер находился на конце 5' вставки в 3,5 килооснований и которые благодаря этому имели участок инициации с началом ATG, управляемый промотором Т7. Модифицированную вставку в 3,5 килооснований из одного такого клона (pSPT18-3,5-2N44) вырезали и субклонировали в Десять мкл каждого супернатанта от клеток ВНК добавляли к 25 мМ р-нитроанилида субстрата в 250 мкл бикарбонатного буфера HEPES при рН 7,0 и выдерживали при 37°С в течение 30 минут. Пример 2. Клонирование последовательности H110D в вектор экспрессии из материала млекопитающих. Для клонирования взяли ген H110D (продукт PCR в 3,5 килооснований, клон 2, описанный в WO 93/23542). Последовательность ДНК (последовательность №1) этой вставки, полученная в результате цепной реакции с полимеразой (PCR), показана на фиг. 1, а аминокислотная трансляция - на фиг. 2 (последовательность №2). Эту ДНК вырезали из вектора pT7Blue-T Vector (Novagene) посредством переваривания в BamHI и клонировали в участок BamHI 5 32437 вектор экспрессии для клетки млекопитающего pRC/CMV - следующим образом. 1. ДНК клона pSPT18(T7)-3,5-2N44) переварили рестрикционным ферментом Smal. 2. В этот модифицированный участок Smal вшили линкер NotI для получения клонов с участком NotI на конце 5' вставки, предшествующим линкеру Ncol, содержащему начальный участок ATG. 3. Очищенную ДНК из подходящего клона переварили ферментами NotI и Xbal, чтобы высвободить вставку в 3,5 килооснований. К материалу добавили фермент Pvul, чтобы разрезать ДНК вектора pSPT18 пополам. Дело в том, что одинаковые размеры вектора и вставки затрудняют очистку вставки. 4. Вставку в 3,5 килооснований очистили на 0,6 - 0,7% агарозном геле. 5. Вектор для клеток млекопитающих pRC/CMV переварили ферментами NotI и Xbal, a полосу линейной плазмиды очистили на агарозном геле. 6. Вставку в 3,5 килооснований сшили с вектором, преобразованном в линейную форму. 7. Отобрали подходящие клоны - те, в которых после переваривания NotI и Xbal имелась вставка в 3,5 килооснований. Их назвали pRC/CMV-3,5-2. Трансфекция клеток млекопитающих Cos-1. ДНК вектора экспрессии для клеток млекопитающих с клонами вставок Н110D - клонами pRC/CMV-3,5-2 - тщательно очищается путем центрифугирования в градиенте хлорида цезия (Сэмбрук, Фрич, Маниатис - Sambrook J., Fritsch E. F., Maniatis T. Molecular Cloning, A Laboratory Manual, Second edition. Cold Spring Harbor Press, 1989). Преходящую экспрессию H110D можно получить, используя эту очищенную ДНК клона pRC/CMV-3,5-2 для трансфекции клеток Cos-1 (которые можно получить в Европейской коллекции культивируемых клеток животных, в Портоне). Трансфекция осуществляется с использованием диэтиламиноэтилового декстрана (Cullen B.R., Use of Eukaryotic expression technology in the functional analysis of cloned genes, Methods in Enzymology: Guide to molecular cloning techniques, Eds S.L. Berger and A.R. Kimmal, Academic Press, 1987, pp684704). Клетки выращивают в среде Игла, модифицированной по Дульбекко (DMEM, Gibco BRL) и 10% фетальной телячьей сыворотке (FCS, Gibco BRL). Экспрессию анализируют через 48-72 часа выдержки. Трансфекция клеток яичника китайского хомяка (СНО). Вектор ДНК вводится в клетки СНО (которые можно получить в Европейской коллекции культивируемых клеток животных, в Портоне) с использованием фосфата кальция, как описано у Каллен (Cullen B.R., Use of Eukaryotic expression technology in the functional analysis of cloned genes, Methods in Enzymology: Guide to molecular cloning techniques, Eds S.L. Berger and A.R. Kimmal, Academic Press, 1987, pp684-704). Трансформированные клетки выращивают в среде Игла, модифицированной по Дульбекко, плюс 10% FCS (фирма Gibco BRL). Генетецин (G418) - это среда, где хорошо растут только трансформированные клеточные линии, а нетрансформированные - плохо, его можно использовать в количестве до 800 мкг/мл. Тогда трансформированные клетки клонируют в микротитровальных чашках способом предельного разведения. Исследование трансфециpованных клеток млекопитающих. Трансформированные и нетрансформированные клетки СНО можно перенести для роста на покровные стекла, с которыми проводится иммунофлуоресцентный анализ. Клеткам дают разрастись в небольшие колонии, фиксируют метанолом и испытывают овечьей антисывороткой к H110D. Затем их покрывают флуоресцентным составом (напр. FTTC), конъюгированным с овечьей иммуноглобулиновой антисывороткой. Для отыскания положительных колоний используется флуоресцентный микроскоп. Трансформированные клеточные линии разделяют на клетки в буфере RIPA (150 мМ натрия хлорида, 1% Nonidet P40, 0,5% деоксихолата, 0,1% натрия додецилсульфата, 50 мМ трис-HCI, рН 8,0): среду отделяют от клеток, а клетки осторожно перемешивают в буфере RIPA в течение 5 минут. Затем клетки помещают в пробирку микроцентрифуги и прокручивают на полной скорости в течение 15 минут. Получают прозрачный лизат, который помещают в чистую пробирку. Порции этого лизата, эквивалентные 2 х 105 клеток, подвергают электрофорезу на полиакриламидном геле с додецилсульфатом натрия, а белки в геле переносят на нитроцеллюлозную мембрану посредством вестерн-блоттинга. Мембрану обрабатывают, возможно, с использованием периодата, а затем исследуют антисыворотками к различным формам антигена H110D. Обработка периодатом отделяет углеводные эпитопы; углеводные эпитопы млекопитающих могут значительно отличаться от нативных углеводов гельминта. Вестерн-блоттинги трансформированных клеток показывают присутствие белка, распознанного антисывороткой, специфической к H110D. Экстракты трансформированных и нетрансформированных клеток, приготовленные, как описано в примере 1, и лизаты, полученные с использованием состава RIPA, испытываются на ферментную активность в точности так, как описано в примере 1. Клетки, трансформированные при помощи H110D, характеризуются более высокими уровнями аминопептидазной активности, чем нетрансформированные. Присутствие трансфицированной векторной ДНК, содержащей продукт PCR в 3,5 килооснований, клон 2, в невосприимчивых к генетицину (Geneticin) клетках СНО определяется сазерн-анализом препаратов ДНК из этих клеток. ДНК извлекают из клеток по способу Сэмбрука, Фрича, Маниатиса (Sambrook J., Fritsch E. F., Maniatis T. Molecular Cloning, A Laboratory Manual, Second Ed. Cold Spring Harbor Press, 1989). Десять - двадцать микрограммов ДНК переваривают рестрикционными ферментами, затем подвергают электрофорезу на агарозном геле, затем ДНК переносят на мембрану посредством сазернблоттинга (Southern, E. Detection of specific sequences among DNA fragments separated by gel electrophoreses. J Mol. Biol. 98 p503, 1975). Эту мембрану гибридизуют с зондом, кодирующим продукт PCR в 3,5 килооснований, клон 2, промывают в условиях высокой жесткости и авторадиографируют. В трансформированных клетках имеются полосы, специфические для продукта PCR в 3,5 килооснований, клона 2. Экспрессия продукта PCR в 3,5 килооснований,клона 2, на уровне РНК в трансформированных 6 32437 клетках млекопитающих определяется при помощи назерн-анализа РНК, выделенной из этих клеток. РНК извлекается из клеток с использованием состава RNAzol (Cinna/Biotecx, Техас) в соответствии с инструкциями изготовителя. До 20 мкг подвергается электрофорезу на агарозном геле и переносится на мембрану посредством назерн-блоттинга (Сэмбрук, Фрич, Маниатис - Sambrook J., Fritsch E. F., Maniatis Т., Molecular Cloning, A Laboratory Manual, Second Ed. Cold Spring Harbor Press, 1989). Эту мембрану затем гибридизуют с зондом, кодирующим продукт PCR в 3,5 килооснований, клон 2, промывают в условиях высокой жесткости и радиографируют. Трансформированные клетки, экспрессирующие продукт PCR в 3,5 килооснований, клон 2, дают специфическую полосу гибридизации. (2) СВЕДЕНИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ № 1 (I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ: (A) ДЛИНА: 3303 ПАР ОСНОВАНИЙ (B) ТИП: НУКЛЕИНОВАЯ КИСЛОТА (C) КОЛИЧЕСТВО ЦЕПОЧЕК: ОДНА (D) ТОПОЛОГИЯ:ЛИНЕЙНАЯ (II) ТИП МОЛЕКУЛЫ: кДНК (XI) СВЕДЕНИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ №1 7 32437 8 32437 9 32437 10 32437 11 32437 (2) СВЕДЕНИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ № 2 (I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ: (A) ДЛИНА: 962 АМИНОКИСЛОТ (B) ТИП: АМИНОКИСЛОТА (C) КОЛИЧЕСТВО ЦЕПОЧЕК: ОДНА (D) ТОПОЛОГИЯ: ЛИНЕЙНАЯ (II) ТИП МОЛЕКУЛЫ: пептид (ХІ) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ № 2 12 32437 13 32437 14 32437 15 32437 16 32437 Фиг. 1 17 32437 Фиг. 1 (продолжение) 18 32437 Фиг. 2 Тираж 50 екз. Відкрите акціонерне товариство «Патент» Україна, 88000, м. Ужгород, вул. Гагаріна, 101 (03122) 3 – 72 – 89 (03122) 2 – 57 – 03 19
ДивитисяДодаткова інформація
Назва патенту англійськоюA method for carrying out expression of aminopeptidase antigen n110d
Автори англійськоюMann Edvard Albert, Graham Margaret, Smith Travor Stanley, Rolf Timothy Peter, Newton Suzanne Elizabeth
Назва патенту російськоюСпособ осуществления экспрессии аминопептидазного антигена н110д
Автори російськоюМанн Эдвард Альберт, Грехем Маргарет, Смит Тревор Стенли, Ролф Тимоти Питер, Ньютон Сьюзен Элизабет
МПК / Мітки
МПК: C12N 9/64, C12N 15/09, C12N 15/12, C12N 5/10, C12N 9/48
Мітки: амінопептидазного, спосіб, здійснення, антигену, експресії, н110д
Код посилання
<a href="https://ua.patents.su/19-32437-sposib-zdijjsnennya-ekspresi-aminopeptidaznogo-antigenu-n110d.html" target="_blank" rel="follow" title="База патентів України">Спосіб здійснення експресії амінопептидазного антигену н110д</a>
Saf-поліпептид, фрагмент днк, що кодує saf-поліпептид, рекомбінантна плазмідна днк для експресії safполіпептиду (варіанти), штам грибів streptomyces lividans, спосіб експресії saf-поліпептиду

Номер патенту: 27696
Опубліковано: 16.10.2000
Автори: Жіль Хосе Антоніо, Гарсіа Томас Вігаль, Ортега Антоніо Даза, Мартін Хуан Франціско
МПК: C12N 15/31, C07K 14/00, C07K 14/37, C12P 21/02, C07K 14/36, C12N 1/21, C12N 15/09, C07K 14/41
Мітки: streptomyces, рекомбінантна, saf-поліпептиду, saf-поліпептид, варіанти, грибів, кодує, спосіб, safполіпептиду, експресії, фрагмент, плазмідна, lividans, штам, днк
Текст:
...от штамма АТСС 10137 S. griseus. 7.2 кб Bgl II фрагмент от pULADI и 1 кб Bgl II фрагмент от pULAD3 субклониовали раздельно в обеих ориентациях в ВамНI-переваренной pUC 19. Все эти плазмидные конструкции проверялись в клетках Е. coli M103 и затем использовались для трансформирования Е. coli E15 (Sartiy и др. J. Bacteriol, 145:288-292,1981) и Е. coli A 1046, в В-ХР не было обнаружено никаких синих колоний, позволяющих предположить, что...
Спосіб одержання трепонемного антигену для діагностики сифілісу

Номер патенту: 23229
Опубліковано: 19.05.1998
Автор: Тацька Лариса Семенівна
МПК: G01N 33/571
Мітки: трепонемного, спосіб, одержання, антигену, діагностики, сифілісу
Формула / Реферат:
Спосіб одержання трепонемного антигену для діагностики сифілісу, який відрізняється тим, що створюють захист трепонемному антигену шляхом висушування стабілізатора, стабілізатор заморожують на протязі 10-20 хвилин до температури-20 —79°С, створюють вакуум, высушують на протязі 20 - 30 годин з наступним доданням до нього нативного трепонемного антигену, потім высушують вдруге у тому ж режимі.
Спосіб експресії мітогенного білка ендотеліальних клітин людини

Номер патенту: 13321
Опубліковано: 28.02.1997
Автори: Майкл Джей, Вільям Дрохан, Вільсон Берджесс, Томас Мейсіаг
МПК: C12N 15/24, C12N 15/74
Мітки: спосіб, експресії, мітогенного, ендотеліальних, клітин, людини, білка
Формула / Реферат:
(57) Способ экспресии митогенного белка эндотелиальных клеток человека в Escherichla coll, заключающийся в том, что выделяют мРНК из вытяжки ствола мозга человека, получают из мРНК поли(А+)мРНК, синтезируют кДНК с помощью указанной мРНК, колонируют кДНК в вектор лямбда gt10 или лямбда gt11, отбирают клоны кДН К, кодирующие митогенный белок эндотелиальных клеток путем гибридизации указанных клонов с олигонуклеотидной пробой, имеющей следующую...
Спосіб отримання ротавірусного антигену

Номер патенту: 14568
Опубліковано: 20.01.1997
Автори: Онуфріев Владислав Петрович, Міськевич Степан Володимирович, Скибіцький Володимир Гурійович, Постой Віктор Петрович
МПК: A61K 39/225, C12R 1/91
Мітки: антигену, ротавірусного, отримання, спосіб
Формула / Реферат:
Спосіб отримання ротавірусного антигену шляхом зараження клітин, руйнування їх триразовим заморожуванням і розморожуванням та видалення клітинного детриту центрифугуванням з подальшим виділенням віруса з інфікованих клітин, який відрізняється тим, шо із суспензії після обробки її поліетіленгліколем вірус сорбують на тканину Петрянова, сорбційний шар якої з осадом розчиняють в хлороформі і додають фізіологічний розчин при струшуванні,...
Спосіб приготування ротавірусного антигену з польових матеріалів

Номер патенту: 14592
Опубліковано: 20.01.1997
Автори: Постой Віктор Петрович, Міськевич Степан Володимирович, Герстман Микола Володимирович, Онуфріев Владислав Петрович, Скибіцький Володимир Гурійович
МПК: A61K 39/225, C12N 7/06
Мітки: польових, приготування, матеріалів, антигену, спосіб, ротавірусного
Формула / Реферат:
Спосіб приготування ротавірусного антигену з польових матеріалів шляхом дворазового послідовного центрифугування суспензії, який відрізняється тим, що суспензію очищають хлороформом, концентрують поліетіленгліколем, фільтрують через тканину Петрянова, сорбційний шар якої з осадом розчиняють в хлороформі і додають фізіологічний розчин при струшуванні, відділяючи потім надосад, в якому концентрується вірус, центрифугуванням.
Попередній патент: Спосіб випробування матеріалів на тріщиностійкість
Наступний патент: Спосіб виробництва корму тваринного походження
Випадковий патент: Рідина для глушіння та капітального ремонту свердловин