Рослина з підвищеними показниками врожайності та спосіб її одержання




































































































































































































































































































































































Формула / Реферат
1. Спосіб підвищення показників врожайності рослин, який включає збільшення експресії у рослині: (i) послідовності нуклеїнової кислоти, яка кодує поліпептид Фактора Регулювання Росту (GRF); та (ii) послідовності нуклеїнової кислоти, яка кодує поліпептид транслокації синовіальної саркоми (SYT), де вказані показники врожайності є підвищеними відносно до рослин з підвищеною експресією однієї з: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, або (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид,
причому вказаний GRF поліпептид містить:
(i) домен, амінокислотна послідовність якого на щонайменше 50 % ідентична до QLQ домену, представленого SEQ ID NO:115; та
(ii) домен, амінокислотна послідовність якого на щонайменше 50 % ідентична до WRC домену, представленого SEQ ID NO:116, і
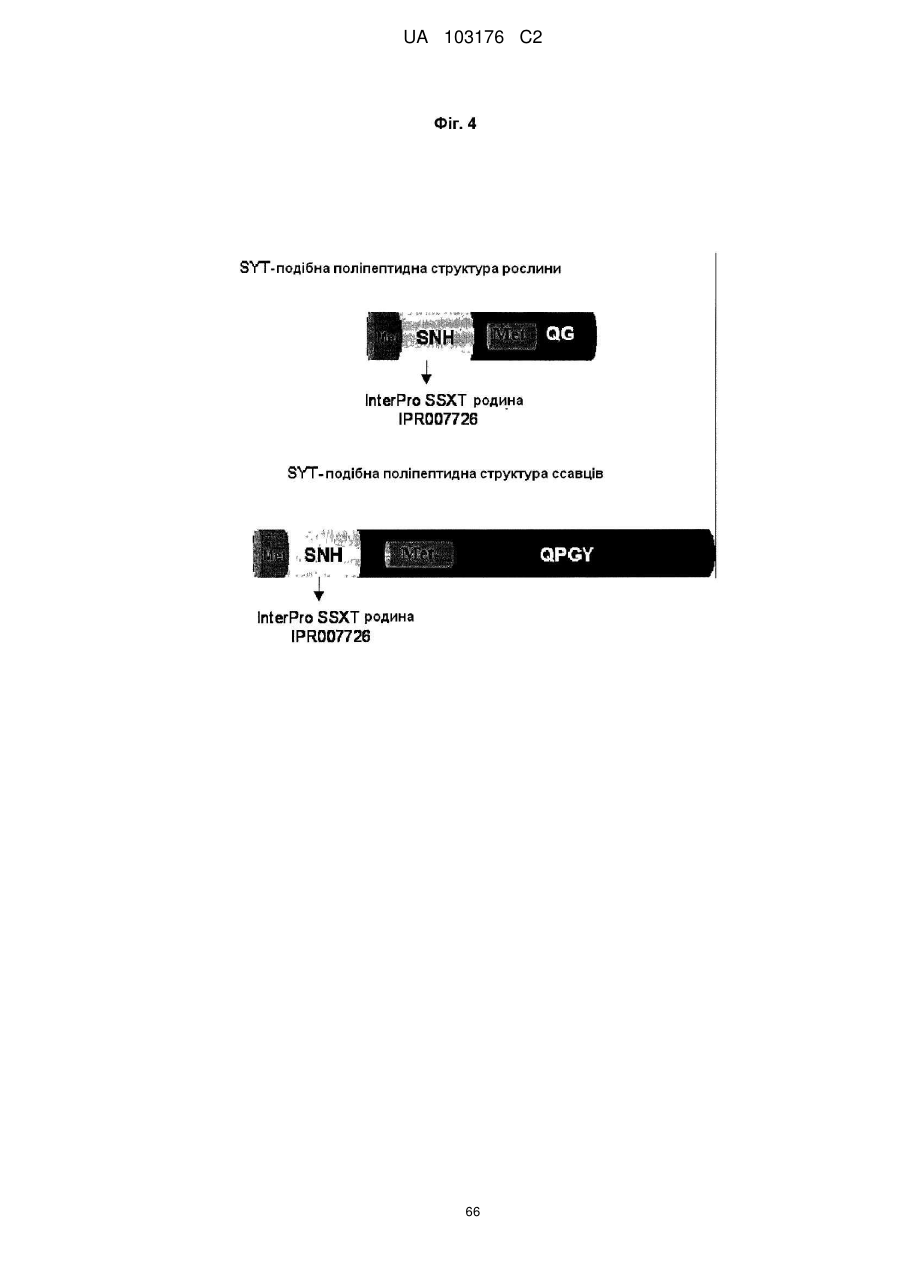
причому вказаний SYT поліпептид містить між N-кінцем та C-кінцем:
(i) SNH домен, ідентичність послідовності якого збільшується на щонайменше 20 % до SNH домену SEQ ID NO:262; та
(ii) Met-збагачений домен; та
(iii) QG-збагачений домен.
2. Спосіб за п. 1, де вказаний GRF поліпептид містить: (i) домен, амінокислотна послідовність якого на щонайменше 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 %, 95 %, 98 %, 99 % або більше ідентична до QLQ домену, представленого SEQ ID NO:115; та (ii) домен, амінокислотна послідовність якого на щонайменше 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 %, 95 %, 98 %, 99 % або більше ідентична до WRC домену, представленого SEQ ID NO:116.
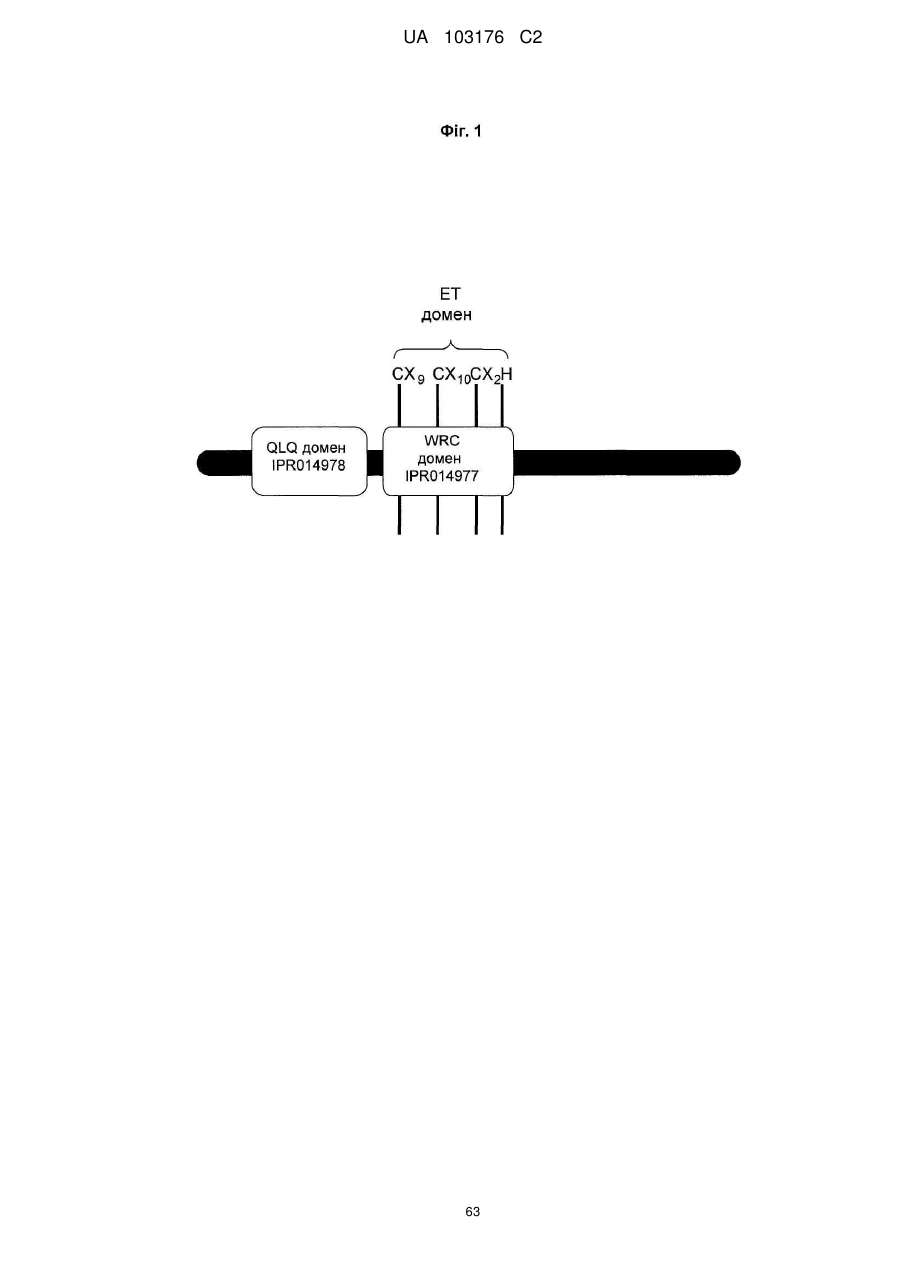
3. Спосіб за пп. 1 або 2, де вказаний GRF поліпептид містить: (i) QLQ домен з InterPro повтором IPR014978 (PFAM повтор PF08880); (ii) WRC домен з InterPro повтором IPR014977 (PFAM повтор PF08879); та (iii) домен ефектора транскрипції (ET), який містить три Cys та один His залишки у консервативному регіоні (CX9CX10CX2H).
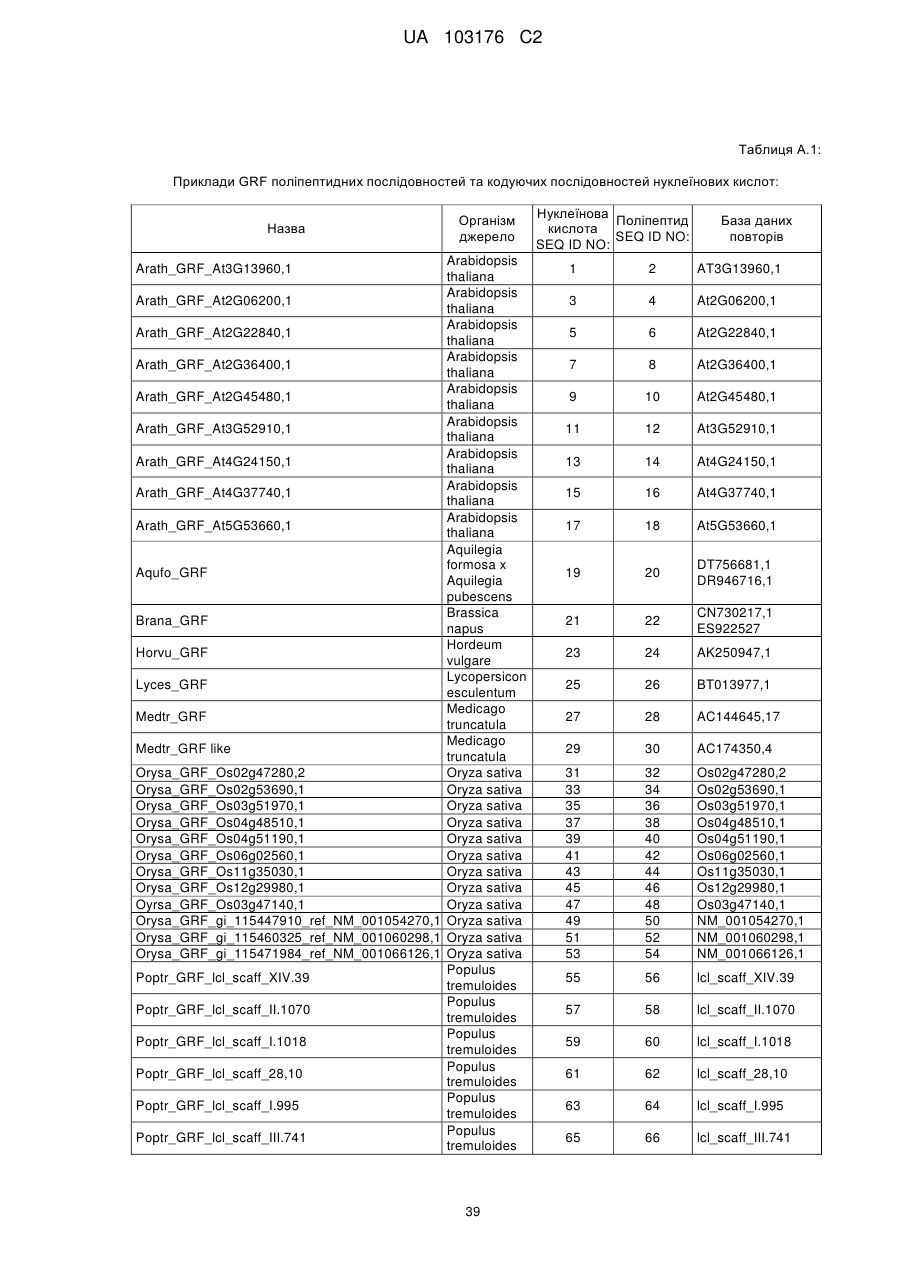
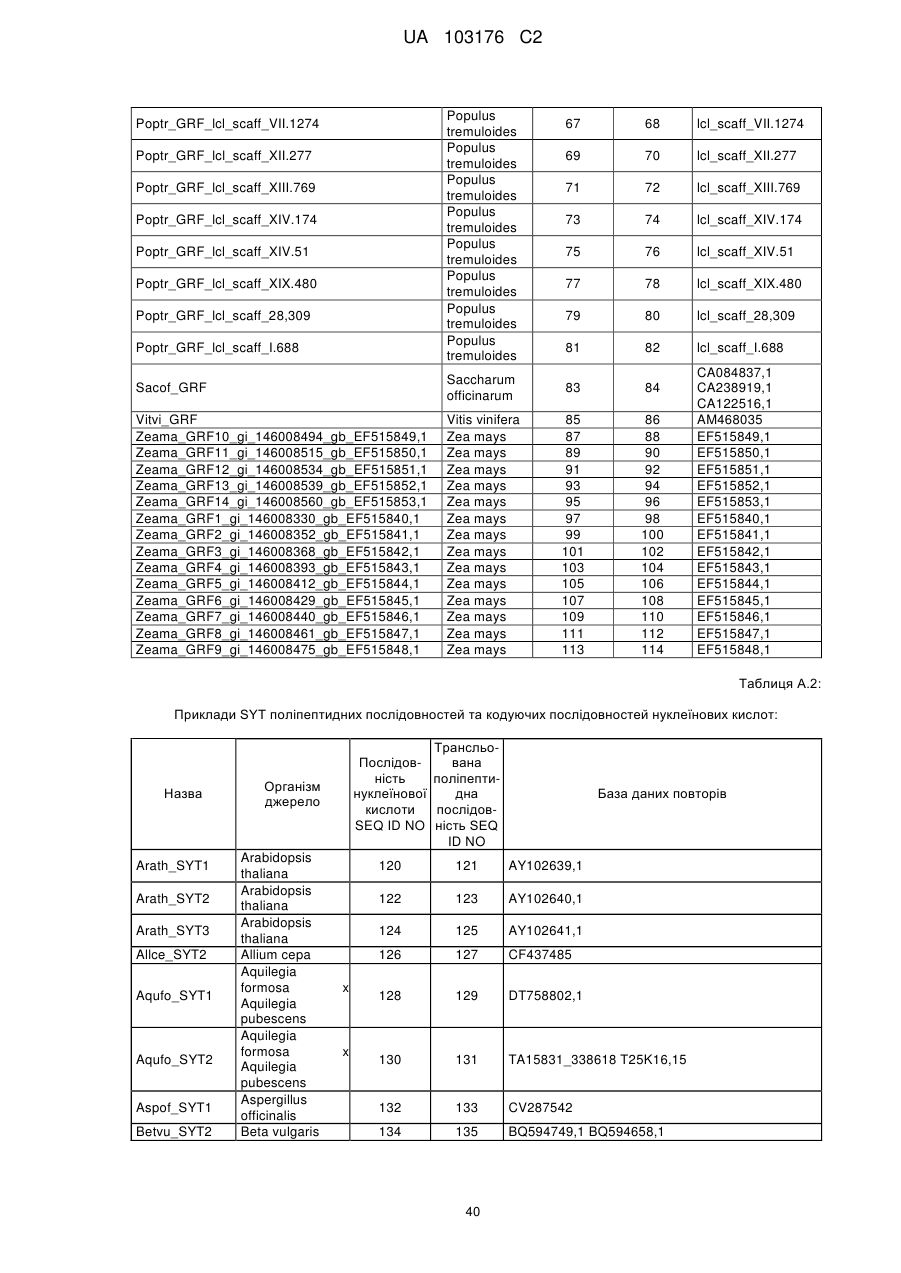
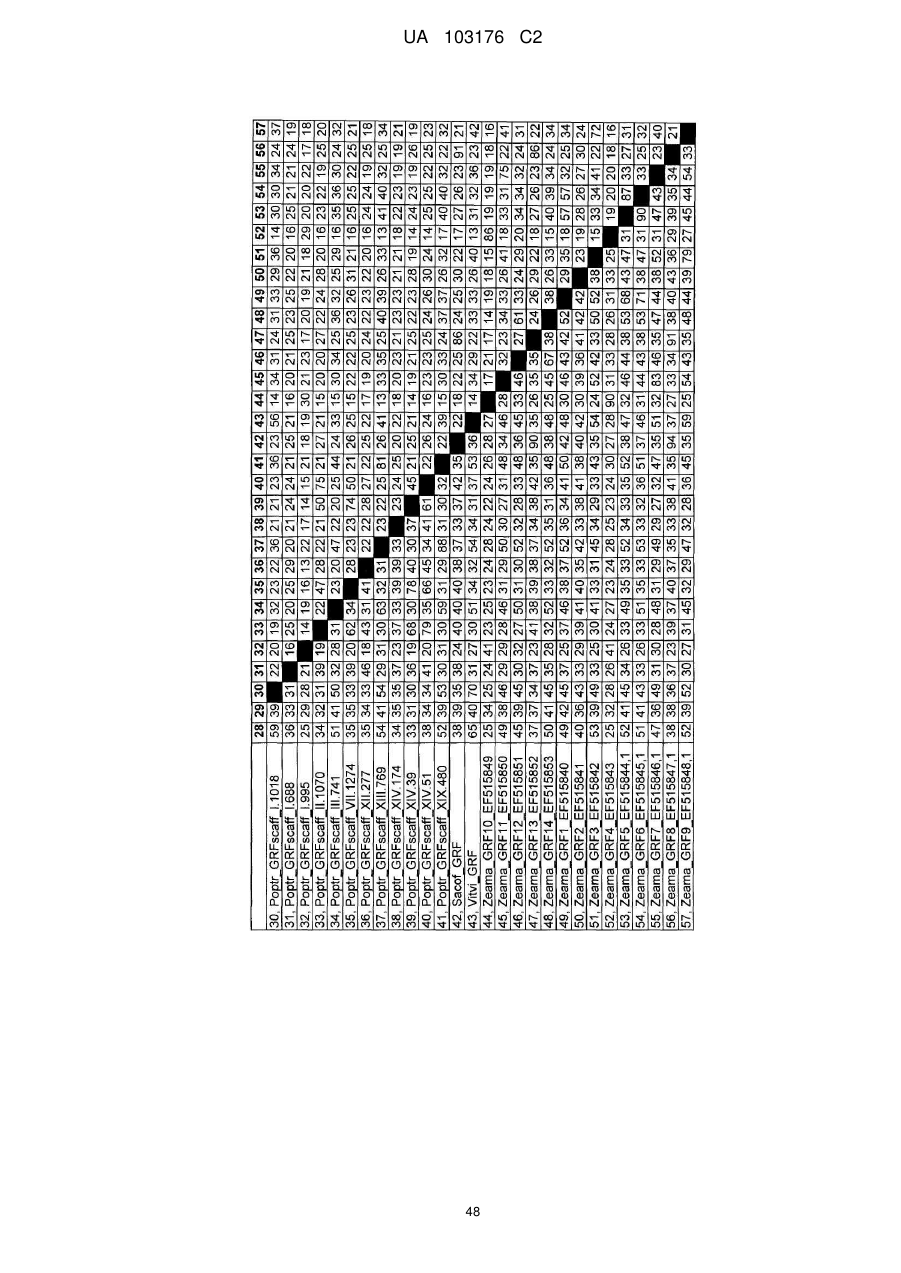
4. Спосіб за будь-яким з попередніх пунктів, де ідентичність амінокислотної послідовності вказаного GRF поліпептиду збільшується у порядку переважності на щонайменше 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 %, 95 %, 98 %, 99 % або більше до GRF поліпептиду, представленого SEQ ID NO:2 або до будь-яких поліпептидних послідовностей, наведених у Таблиці A.1.
5. Спосіб за будь-яким з попередніх пунктів, де вказана послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, представлена будь-якою послідовністю нуклеїнової кислоти SEQ ID NO, наведеною у Таблиці A.1 або їх частиною, або послідовністю, здатною до гібридизації з будь-якою послідовністю нуклеїнової кислоти SEQ ID NO, наведеної у Таблиці A.1.
6. Спосіб за будь-яким з попередніх пунктів, де вказана послідовність нуклеїнової кислоти кодує ортолог або паролог будь-якої GRF поліпептидної послідовності SEQ ID NO, наведеної у Таблиці A.1.
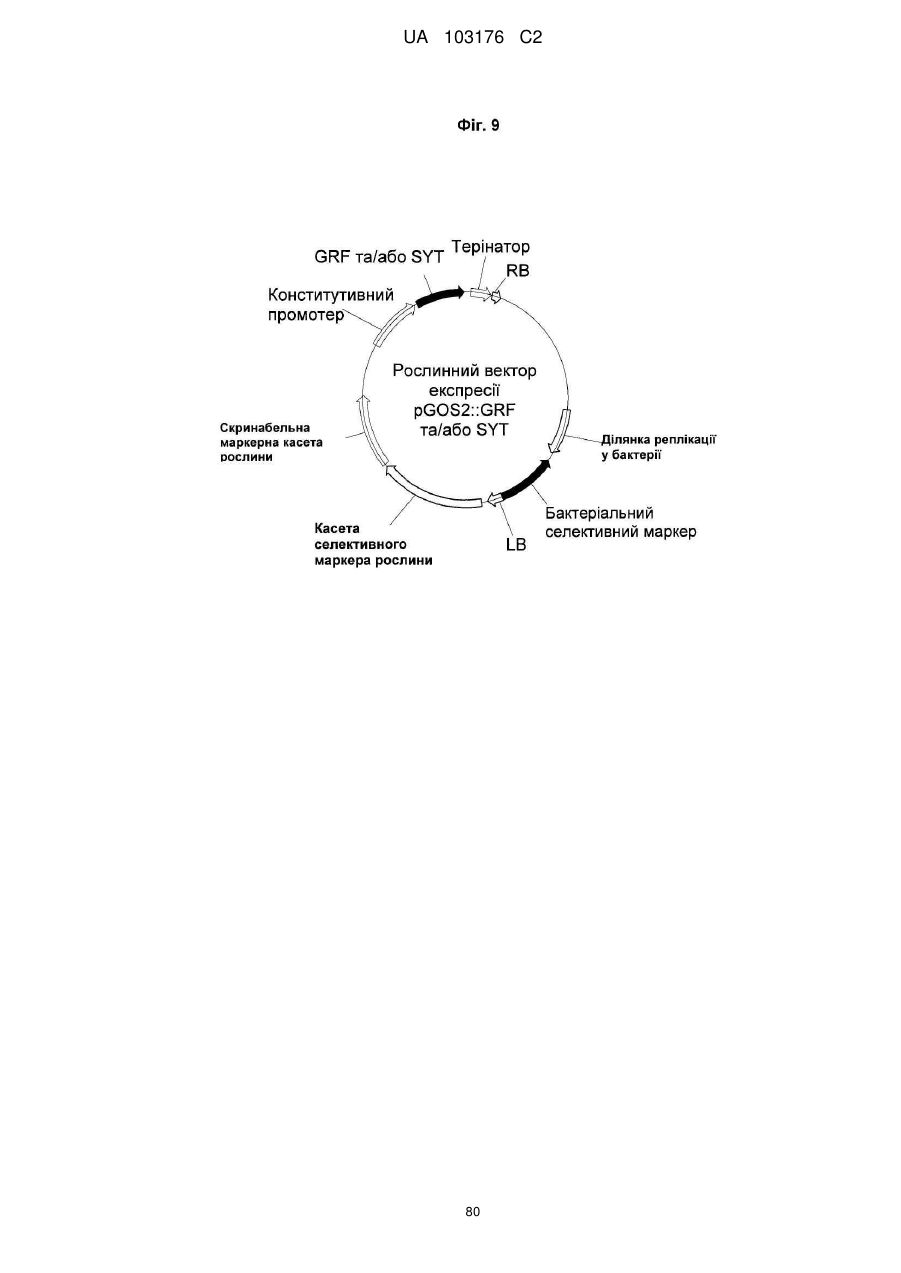
7. Спосіб за будь-яким з попередніх пунктів, де вказана послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, є операбельно зв′язаною з конститутивним промотером, більш бажано з GOS2 промотером, найбільш бажано з GOS2 промотором рису, представленим SEQ ID NO:117.
8. Спосіб за будь-яким з попередніх пунктів, де вказана послідовність нуклеїнової кислоти, яка кодує GRF поліпептид має рослинне походження, бажано походить від дводольної рослини, більш бажано з родини Brassicaceae, найбільш бажано з Arabidopsis thaliana.
9. Спосіб за п. 1, де вказана послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, де вказаний SYT поліпептид містить між N-кінцем та C-кінцем: (i) SNH домен, ідентичність послідовності якого збільшується у порядку переважності на щонайменше 25 %, 30 %, 35 %, 40 %, 45 %, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 %, 91%, 92 %, 93 %, 94 %, 95%, 96 %, 97 %, 98 %, 99 % до SNH домену SEQ ID NO: 262; та (ii) Met-збагачений домен; та (iii) QG-збагачений домен.
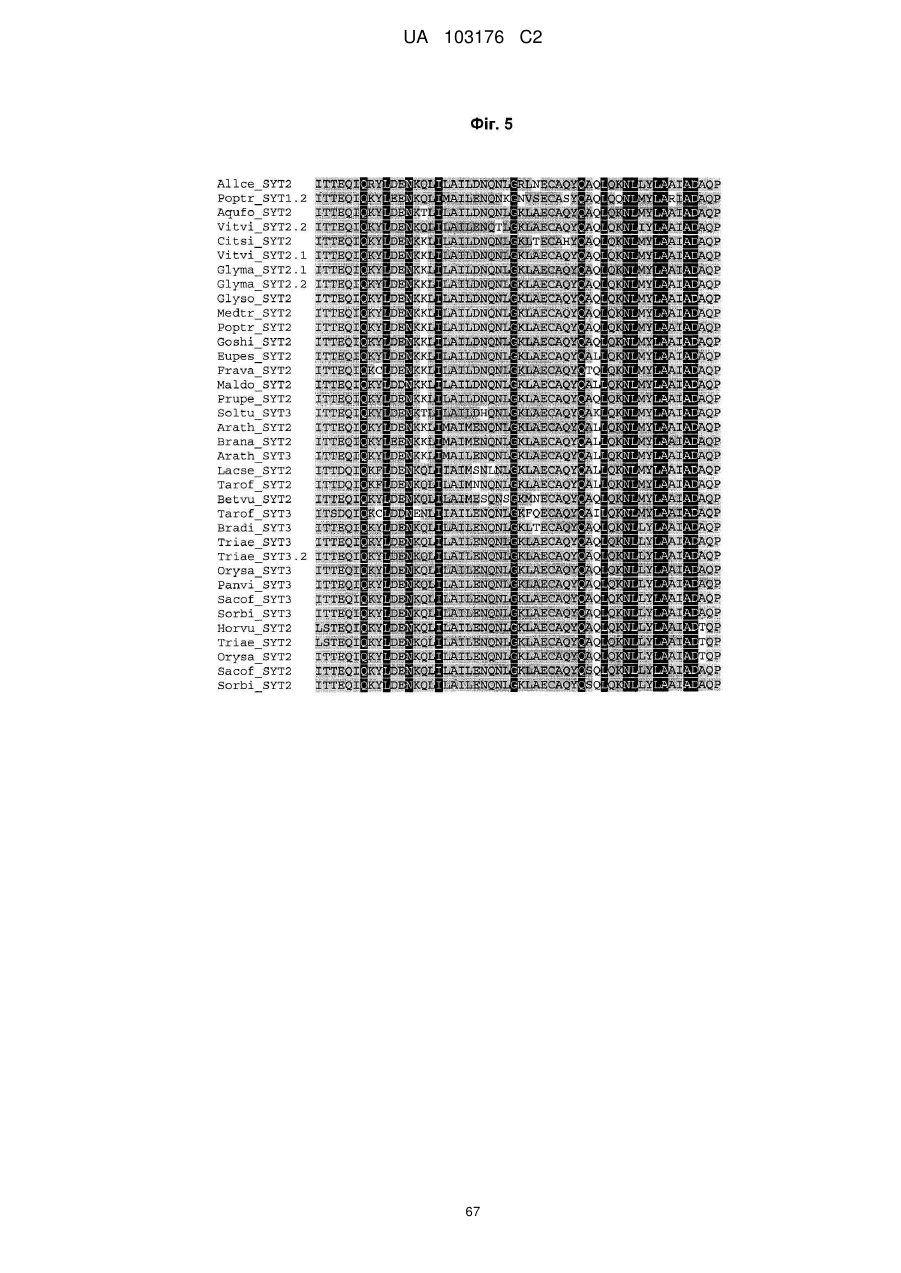
10. Спосіб за будь-яким з пп. 1-9, де вказаний SYT поліпептид додатково містить найбільш консервативні залишки SNH домену, представленого SEQ ID NO:263 та показаного на Фіг. 5.
11. Спосіб за будь-яким з пп. 1-10, де вказаний SYT поліпептид містить домен, ідентичність послідовності якого збільшується у порядку переважності на щонайменше 20 %, 25 %, 30 %, 35 %, 40 %, 45 %, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 8 %, 90 %, 91 %, 92 %, 93 %, 94 %, 95 %, 96 %, 97 %, 98 %, 99 % до SSXT домену з InterPro повтором IPR007726 SEQ ID NO: 264,
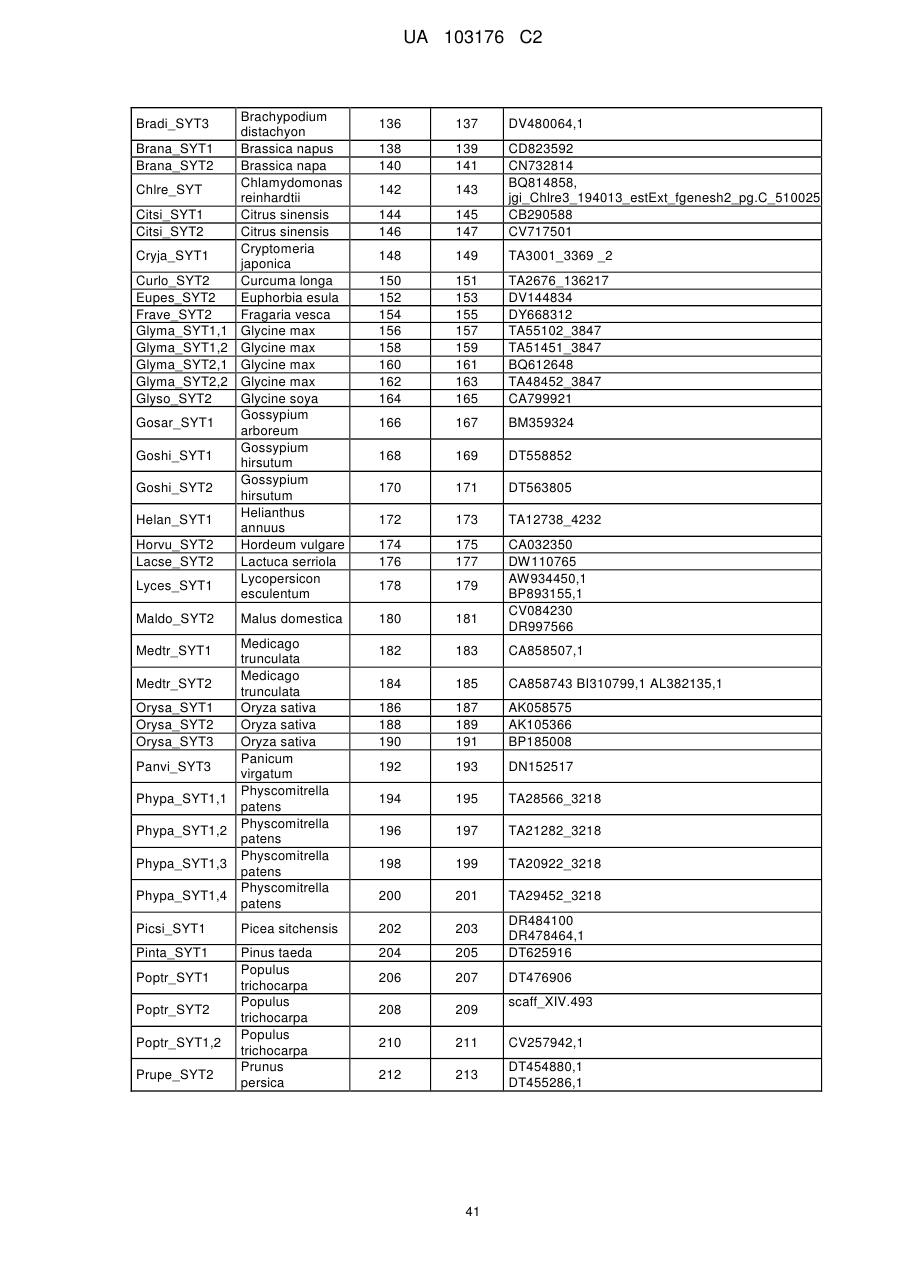
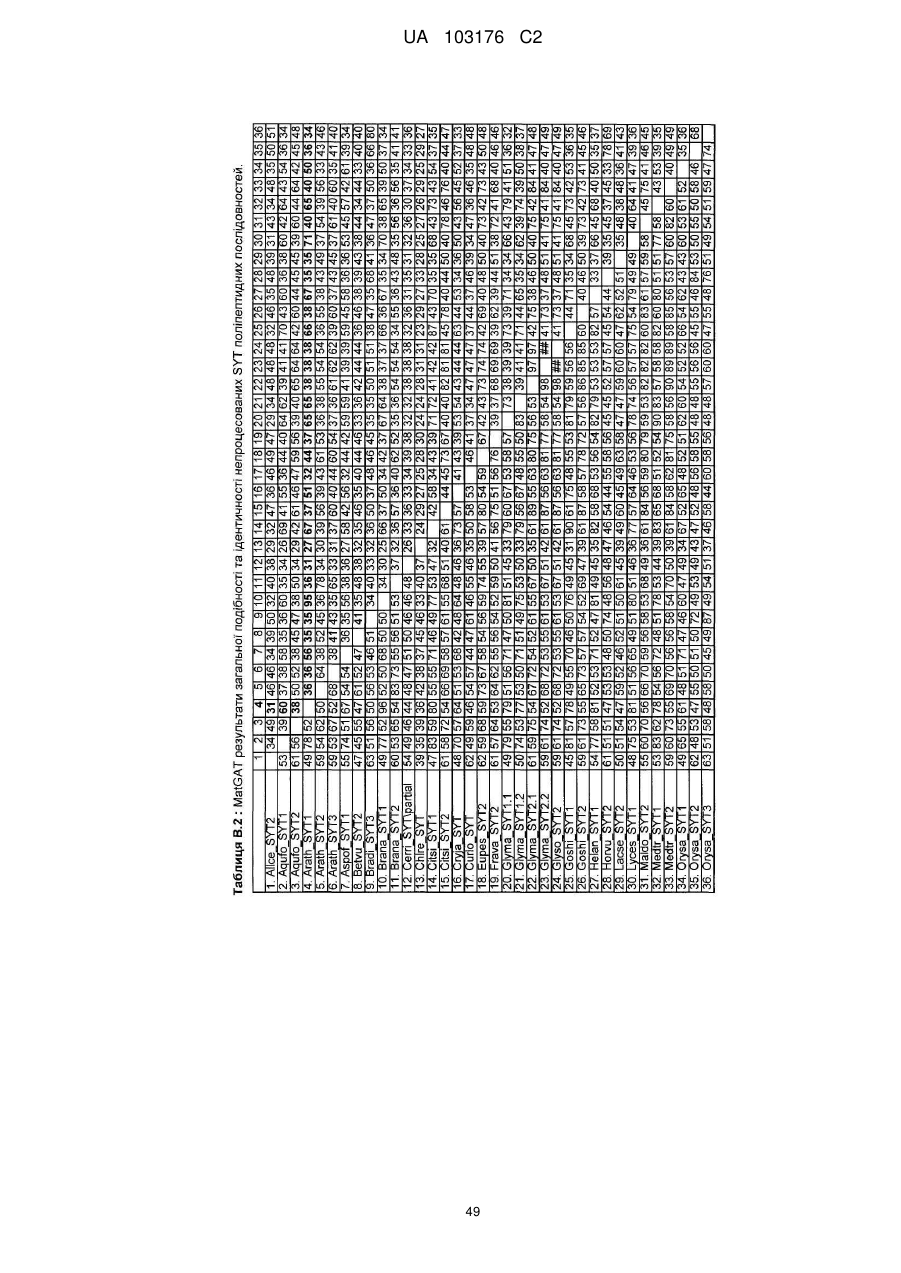
12. Спосіб за будь-яким з пп. 1-10, де ідентичність амінокислотної послідовності вказаного SYT поліпептиду збільшується у порядку переважності на щонайменше 20 %, 25 %, 30 %, 35 %, 40 %, 45 %, 50 %, 55 %, 60 %, 65 %, 70 %, 75 %, 80 %, 85 %, 90 %, 95 %, 98 %, 99 % або більше до SYT поліпептиду, представленого SEQ ID NO: 121 або до будь-яких послідовностей непроцесованих поліпептидів, наведених у Таблиці A.2.
13. Спосіб за будь-яким з пп. 1-12, де вказана послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, представлена будь-якою послідовністю нуклеїнової кислоти SEQ ID NO, наведеною у Таблиці A.2, або її частиною, або послідовністю, здатною до гібридизації з будь-якими послідовностями нуклеїнових кислот SEQ ID NO, наведеними у Таблиці A.2.
14. Спосіб за будь-яким з пп. 1-13, де вказана послідовність нуклеїнової кислоти кодує ортолог або паролог будь-якої послідовності SYT поліпептиду SEQ ID NO, наведеної у Таблиці A.2.
15. Спосіб за будь-яким з пп. 1-14, де вказана послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, є операбельно зв′язаною з конститутивним промотором, більш бажано з GOS2 промотором, найбільш бажано з GOS2 промотором рису, представленого SEQ ID NO: 117.
16. Спосіб за будь-яким з пп. 1-15, де вказана послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, має рослинне походження, бажано походить від дводольної рослини, більш бажано з родини Brassicaceae, найбільш бажано з Arabidopsis thaliana.
17. Спосіб за будь-яким з пп. 1-16, де вказану підвищену експресію отримують шляхом введення та експресії у рослині: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид; та (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид.
18. Спосіб за п. 17, де вказані послідовності нуклеїнових кислот (i) та (ii) послідовно вводять та експресуються у рослині, бажано шляхом схрещування, більш бажано шляхом перетрансформації.
19. Спосіб за п. 18, де вказане схрещування здійснюють між батьківською рослиною жіночої статі, яка містить введену та експресовану ізольовану послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, та батьківською рослиною чоловічої статі, яка містить введену та експресовану ізольовану послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, або реципрокно, та шляхом відбору потомства на наявність та експресію обох трансгенів, де вказана рослина має підвищені показники врожайності відносно до кожної батьківської рослини.
20. Спосіб за п. 18, де вказану перетрансформацію здійснюють шляхом введення та експресування послідовності нуклеїнових кислот, яка кодує GRF поліпептид у рослині, частині рослини або рослинній клітині, яка містить введену та експресовану послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, або реципрокно.
21. Спосіб за п. 17, де вказані послідовності нуклеїнових кислот (i) та (ii) одночасно вводять та експресуються у рослині.
22. Спосіб за п. 21, де вказані послідовності нуклеїнових кислот (i) та (ii) входять до однієї або кількох молекул нуклеїнової кислоти.
23. Спосіб за будь-яким з попередніх пунктів, де вказаним підвищеним показником врожайності є одна або кілька з наступних: (i) підвищена рання сила; (ii) підвищена біомаса надземної частини рослини; (iii) підвищений загальний вихід насіння на рослину; (iv) підвищений показник наповнення насіння; (v) підвищена кількість (наповненого) насіння; (vi) підвищений індекс врожайності; або (vii) підвищена маса тисячі зерен (TKW).
24. Спосіб за будь-яким з попередніх пунктів, де вказана послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, та вказана послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, є операбельно та послідовно зв′язаними з конститутивним промотором, бажано рослинним конститутивним промотором, більш бажано з GOS2 промотором, найбільш бажано з GOS2 промотором рису, представленим SEQ ID NO: 117.
25. Конструкт, який містить:
(a) послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8;
(b) послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16;
(c) одну або кілька контрольних послідовностей, здатних керувати експресією послідовності нуклеїнової кислоти, яка кодує (a) та (b); та необов′язково
(d) послідовність термінації транскрипції.
26. Конструкт за п. 25, де вказаною контрольною послідовністю є щонайменше один промотор, бажано GOS2 промотор, більш бажано GOS2 промотор, представлений SEQ ID NO:117.
27. Суміш конструктів, де щонайменше один конструкт містить:
(a) послідовність нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8;
(b) одну або кілька контрольних послідовностей, здатних керувати експресією послідовності нуклеїнової кислоти (a); та необов′язково
(c) послідовність термінації транскрипції,
та, де щонайменше один інший конструкт містить:
(d) послідовність нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16;
(e) одну або кілька контрольних послідовностей, здатних керувати експресією послідовності нуклеїнової кислоти (d); та, необов′язково,
(f) послідовність термінації транскрипції.
28. Суміш за п. 27, де вказана контрольна послідовність (b) та/або (e) є щонайменше одним конститутивним промотором, бажано GOS2 промотором, більш бажано GOS2 промотором, представленим SEQ ID NO:117.
29. Застосування конструкту за пп. 25 або 26, або суміші конструктів за п. 27, у способі отримання рослин з підвищеними показниками врожайності у порівнянні з рослинами з підвищеною експресією однієї з: (a) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, або (b) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, підвищеними показниками врожайності якої є один або кілька з наступних: (i) підвищена рання сила; (ii) підвищена біомаса надземної частини рослини; (iii) підвищений загальний вихід насіння на рослину; (iv) підвищений показник наповнення насіння; (v) підвищена кількість (наповненого) насіння; (vi) підвищений індекс врожайності; або (vii) підвищена маса тисячі зерен (TKW).
30. Спосіб отримання трансгенних рослин з підвищеними показниками врожайності у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, або (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, який включає:
a. введення та експресію у рослині, частині рослини або рослинній клітині, послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8, під контролем конститутивного промотора; та
b. введення та експресію у рослині, частині рослини або рослинній клітині послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16, під контролем конститутивного промотора; та
c. культивування рослинної клітини, частини рослини або рослини в умовах, які стимулюють ріст та розвиток рослини.
31. Трансгенна рослина з підвищеними показниками врожайності у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид; або (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, отриманими завдяки підвищеній експресії: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8; та (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16.
32. Трансгенна рослина за п. 31, де вказана рослина є злаком або однодольною, або зерновою рослиною, такою як рис, кукурудза, пшениця, ячмінь, просо, жито, тритикале, сорго та овес, або трансгенна рослинна клітина, отримана з вказаної трансгенної рослини.
33. Трансгенна рослинна клітина з підвищеними показниками врожайності у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид; або (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, отриманими завдяки підвищеній експресії: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8; та (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16.
34. Трансгенна рослинна клітина за п. 33, де вказана рослина є злаком або однодольною, або зерновою рослиною, такою як рис, кукурудза, пшениця, ячмінь, просо, жито, тритикале, сорго та овес, або трансгенна рослинна клітина, отримана з вказаної трансгенної рослини.
35. Застосування (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, визначений в будь-якому з пп. 1-6 та 8; та (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, визначений в будь-якому з пп. 1, 9-14 та 16, для підвищення показників врожайності у рослинах у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, або (ii) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, чиї підвищені показники врожайності є однією з наступних: (i) підвищена рання сила; (ii) підвищена біомаса надземної частини рослини; (iii) підвищений загальний вихід насіння на рослину; (iv) підвищений показник наповнення насіння; (v) підвищена кількість (наповненого) насіння; (vi) підвищений індекс врожайності; або (vii) підвищена маса тисячі зерен (TKW).
Текст
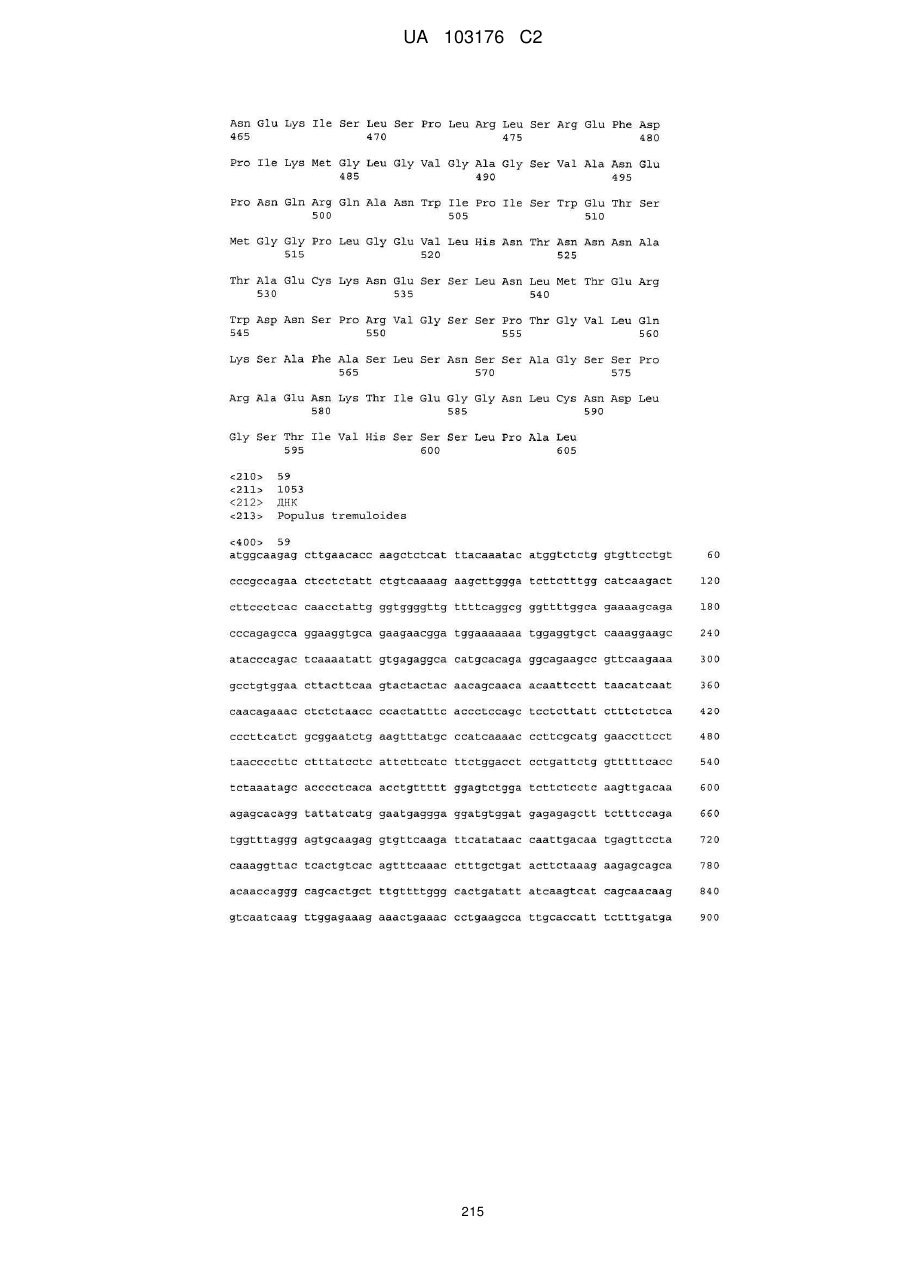
Реферат: Винахід належить до галузі молекулярної біології та стосується способу підвищення показників врожайності рослин шляхом підвищення експресії в рослині: (і) послідовності нуклеїнової кислоти, яка кодує поліпептид фактора регулювання росту (GRF); та (іі) послідовності нуклеїнової кислоти, яка кодує поліпептид транслокації синовіальної саркоми (SYT), де вказана врожайність є підвищеною у порівнянні з рослинами, які мають підвищену експресію однієї з: (і) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид, або (іі) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид. Винахід також стосується рослин з підвищеною експресією UA 103176 C2 (12) UA 103176 C2 (і) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид; та (іі) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид, де вказані рослини мають підвищену врожайність у порівнянні з рослинами з підвищеною експресією однієї з: (і) послідовності нуклеїнової кислоти, яка кодує GRF поліпептид; або (іі) послідовності нуклеїнової кислоти, яка кодує SYT поліпептид. Винахід також стосується конструктів, придатних у способах винаходу. UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 Даний винахід стосується галузі молекулярної біології та способу підвищення різноманітних показників врожайності рослин шляхом підвищення експресії в рослині: (i) послідовності нуклеїнових кислот, яка кодує поліпептид фактору регулювання росту (GRF); та (ii) послідовності нуклеїнових кислот, яка кодує поліпептид транс локації синовіальної саркоми (SYT), де вказана врожайність є підвищеною у порівнянні з рослинами, які мають підвищену експресію однієї з: (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид, або (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид. Даний винахід також стосується рослин з підвищеною експресією (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид; та (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид, де вказані рослини мають підвищену врожайність у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид; або (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид. Винахід також стосується конструктів, придатних у способах винаходу. У світі постійно збільшується населення та постійно зменшується площа доступних оброблюваних земель, тому ведуться дослідження сільськогосподарських паливних культур з метою збільшення ефективності сільського господарства. Традиційні способи покращення зернових та содових культур використовують селекцію для ідентифікації рослин з бажаними характеристиками. Однак, такі технології селекції мають декілька недоліків, зокрема, такі технології потребують значної затрати праці та приводять до того, що рослини часто містять гетерогенні генетичні компоненти, які не завжди приводять до бажаної характеристики, отриманої з батьківських рослин. Переваги молекулярної біології дозволили людству модифікувати зародкову плазму тварин та рослин. Генетична інженерія рослин означає ізоляцію та маніпулювання генетичним матеріалом (зазвичай у формі ДНК або РНК) та наступне введення генетичного матеріалу у рослину. Завдяки таким технологіям з'являються зернові та рослини з різноманітними покращеними економічними, агрономічними або садівничими характеристиками. Особливо економічно переважною характеристикою є підвищена врожайність. Врожайність зазвичай визначають як вихід економічної цінності зернових, що вимірюється. Це може бути представлено у визначеннях кількості та/або якості. Врожайність напряму залежить від кількох факторів, наприклад, кількість та розмір частин рослини, структура рослини (наприклад, кількість відгалужень), насінна продуктивність, відмирання листя тощо. Розвиток коріння, поглинання споживних речовин, толерантність до стресу та рання сила також є важливими факторами при визначенні врожайності. Оптимізація таких факторів призведе до збільшення врожайності зернових. Вихід насіння є особливо важливою характеристикою, з огляду на те, що насіння багатьох рослин є важливим для харчування людини та тварин. Зернові, такі як кукурудза, рис, пшениця, канола та соя, складають більше ніж половину загального вживання калорій, або внаслідок безпосередньої споживання насіння, або внаслідок споживання м'ясних продуктів, отриманих з тварин, які годувались насінням. Також вони є джерелом сахарів, олій та багатьох метаболітів, які використовують у промисловому виробництві. Насіння містить ембріон (джерело нових ростків та коріння) та ендосперм (джерело споживних речовин для ембріонального росту, та протягом проростання, та протягом раннього росту саджанців). Розвиток насіння залежить від багатьох генів, та потребує трансфер метаболітів з коріння, листя та стебла до насіння, що росте. Ендосперм, зокрема, накопичує метаболічні попередники та синтезує їх у депо макромолекул для заповнення зерна. Рослинна біомаса є врожаєм для кормових культур, таких як люцерна, силос та сіно. Було зроблено багато пропозицій щодо врожайності зернових. Головною серед них є оцінка розміру рослини. Розмір рослини можна виміряти багатьма шляхами в залежності від виду та стадії розвитку, проте включають загальну суху масу рослини, суху масу надземної частини рослини, масу свіжої надземної частини рослини, площину листя, об'єм стебла, висоту рослини, діаметр розетки, довжину листя, довжину коріння, вагу коріння, кількість ростків та кількість листя. Багато видів підтримують постійне співвідношення між розміром різних частин рослини на певній стадії розвитку. Ці алометричні співвідношення використовують для екстраполяції одного з таких вимірювань на інше (наприклад, Tittonell et al 2005 Agric Ecosys & Envferum 105: 213). Розмір рослини на ранній стадії розвитку буде корелювати з розміром рослини на подальшій стадії. Більша рослина з більшою площиною листя зазвичай поглинає більше світла та вуглекислого газу, ніж менша рослина, та тому скоріш за все буде мати більшу вагу протягом однакового періоду (Fasoula & Tollenaar 2005 Maydica 50:39). Це є додатковою перевагою до мікро-середовищних та генетичних переваг які повинна досягти рослина з більшим вихідним розміром. Існує сильний генетичний компонент стосовно розміру рослини та швидкості росту 1 UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 (наприклад, ter Steege et al 2005 Plant Physiology 139:1078), та таким чином, діапазон різних генотипів розміру рослини при однакових умовах середовища скоріш за все буде корелювати з розміром рослини при інших умовах (Hittalmani et al 2003 Theoretical Applied Genetics 107:679). Таким чином, стандартні умови використовують як допоміжні, щодо різноманітних та динамічних умов, які спостерігаються у різних місцях розташування та часі, для зернових на полі. Іншою важливою характеристикою багатьох зернових є рання сила. Покращення ранньої сили є важливою метою для сучасних програм по вирощуванню рису, як помірних, так і тропічних культиварів. Довге коріння є важливим для гарного закріплення у ґрунті для рису водних ґрунтів. Коли рис сіють безпосередньо у затоплені поля та, коли рослини повинні швидко з'явитись над водою, довге коріння асоціюється з силою. Коли використовують рядовий посів для гарної схожості саджанців важливим є довші мезокотилідон та колеоптилі. Здатність забезпечити ранню силу у рослин є дуже важливою для сільського господарства. Наприклад, слабка рання сила була бар'єром для введення гібридів кукурудзи (Zea mays L.) на основі зародкової плазми Corn Belt на атлантичному узбережжі Європи. Індекс врожайності, співвідношення виходу насіння до сухої маси надземної частини рослини є відносно стабільним при багатьох умовах навколишнього середовища та, таким чином, існує чітка кореляція між розміром рослини та виходом зерна (наприклад, Rebetzke et al 2002 Crop Science 42:739). Ці процеси по суті є пов'язаними, тому що більшість біомаси зерна залежить від поточної та збереженої листям та стеблом рослини продуктивності фотосинтезу (Gardener et al 1985 Physiology of Crop Plants. Iowa State University Press, pp68-73). Тому, відбір рослин по розміру, навіть на ранній стадії розвитку, використовували як індикатор для майбутньої потенційної врожайності (наприклад, Tittonell et al 2005 Agric Ecosys & Envзалізо 105: 213). При дослідженні впливу генетичних різниць на толерантність до стресу, можливість стандартизувати характеристики ґрунту, температуру, воду та доступність харчових речовин та інтенсивність світла є перевагами теплиць або камер для вирощування рослин у порівнянні з полем. Однак, неприродні обмеження врожайності внаслідок незначного запилення, внаслідок відсутності вітру або комах, або недостатня площа для зрілого коріння або росту пологу, обмежує застосування цих контрольованих умов середовища для дослідження різниць врожайності. Тому визначення розміру рослини на ранній стадії розвитку при стандартизованих умовах у камері для вирощування або теплиці є стандартною методикою для передбачення потенційних генетичних переваг врожайності. Іншою важливою характеристикою є збільшена толерантність до абіотичного стресу. Абіотичний стрес є першочерговою причиною втрати зернових по всьому світу, що знижує врожайність більшості зернових, більше ніж на 50 % (Wang et al. (2003) Planta 218: 1-14). Абіотичний стрес викликається посухою, засоленням ґрунту, екстремальними температурами, хімічною токсичністю, надлишком чи браком споживних речовин (мікроелементів та/або мікроелементів), опроміненням або окислювальним стресом. Здатність збільшувати толерантність рослини до абіотичного стресу бути мати значну економічну перевагу для фермерів по всьому світу та дозволить вирощувати зернові при несприятливих умовах та на територіях, де вирощування зернових в інакшому випадку буде неможливим. Таким чином врожайність зернових можна збільшити шляхом оптимізації одного з вищенаведених факторів. В залежності від кінцевої мети модифікація певної показники врожайності може бути важливішою за інші. Наприклад, для використання, такого як корм або вироблення деревини, або як джерело біопалива, бажаними є збільшення вегетативних частин рослини, та для застосування такого як отримання муки, крохмалю олії, особливо бажаним є збільшення параметрів насіння. Навіть серед параметрів насіння, деякі можуть бути більш важливими, в залежності від застосування. Різноманітні механізми можуть привести до збільшення виходу насіння, або у виді збільшення розміру насіння або у виді збільшення кількості насіння. Одним способом збільшення показників врожайності (вихід насіння та/або біомаса) рослин є модифікація наявних механізмів росту рослини, таких як клітинний цикл або різноманітні сигнальні шляхи які регулюють ріст рослини або механізми захисту. У наш час винайшли, що різноманітні характеристики пробійності рослин збільшують шляхом підвищення експресії у рослині: (i) послідовності нуклеїнових кислот, яка кодує a поліпептид Фактору Регулювання Росту (GRF); таf (ii) послідовності нуклеїнових кислот, яка кодує поліпептид транслокації синовіальної саркоми (SYT), де вказані показники врожайності є підвищеними у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид; або (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид. Підвищені показники врожайності включають одну або кілька: підвищена рання сила, підвищена біомаса надземної частини рослини, підвищений загальний вихід 2 UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 насіння на кожну рослину, підвищений показник наповнення насіння, підвищена кількість (наповненого) насіння, підвищений індекс врожайності та підвищена маса тисячі зерен (TKW). Рівень техніки поліпептидів Фактору Регулювання Росту (GRF) ДНК-зв'язувальні білки є білками, які містять будь-який з багатьох ДНК-зв'язувальних доменів та, таким чином, мають особливу чи загальну афінність до ДНК. ДНК-зв'язувальні білки включають, наприклад, фактори транскрипції, нуклеази, які розщеплюють молекули ДНК, та гістони, які беруть участь у розміщенні ДНК у ядрі клітини. Фактори транскрипції зазвичай представлені білками, які проявляють зв'язувальну афінність до специфічної послідовності ДНК, та які здатні активувати та/або пригнічувати транскрипцію. Геном Arabidopsis thaliana кодує щонайменше 1533 регулятори транскрипції, що складає приблизно ~5,9 % загальної кількості генів (Riechmann et al. (2000) Science 290: 2105-2109). База даних факторів транскрипції рису (DRTF) є колекцією відомих та прогнозованих факторів транскрипції Oryza sativa L. ssp. indica та Oryza sativa L. ssp. japonica, та на даний час містить 2,025 генні моделі передбачуваних факторів транскрипції (TF) у indica та 2,384 у japonica, розподілені між 63 родинами (Gao et al. (2006) Bioinformatics 2006, 22(10):1286-7). Однією з таких родин є родина Фактору Регулювання Росту (GRF) факторів транскрипції, яка є спеціальною у рослин. Ідентифікували щонайменше дев'ять GRF поліпептидів у Arabidopsis thaliana (Kim et al. (2003) Palnt J 36: 94-104), та щонайменше 12 у Oryza sativa (Choi et al. (2004) Plant Cell Physiol 45(7): 897-904). GRF поліпептиди характеризуються наявністю на їх N-термінальній частині щонайменше двох високо консервативних доменах, названих за їх найбільш консервативними амінокислотами у кожному домені: (i) QLQ домен (InterPro повтор IPR014978, PFAM повтор PF08880), де найбільш консервативними амінокислотами домену є Gln–Leu-Gln; та (ii) WRC домен (InterPro повтор IPR014977, PFAM повтор PF08879), де найбільш консервативними амінокислотами домену є Trp-Arg-Cys. WRC домен додатково містить дві відрізняльні структурні характеристики, а саме, WRC домен збагачений основними амінокислотами Lys та Arg, та та додатково містить три Cys та один His залишок у консервативному регіоні (CX9CX10CX2H), позначений як домен ефектора Транскрипції (ET) (Ellerstrom et al. (2005) Plant Molec Biol 59: 663-681). Консервативний регіон залишків цистеїну та гістидину у ЕТ домені нагадує білки "цинкові пальці" (цинк-зв'язуючі). Окрім цього, сигнальна послідовність ядерної локалізації (NLS) зазвичай міститься у GRF поліпептидних послідовностях. Взаємодію деяких GRF поліпептидів з маленькою родиною спів активаторів транскрипції, GRF-взаємодіючих факторів (GIF1-GIF3, також називають транслокацією синовіальної саркоми SYT1-SYT3), продемонстрували, використовуючи метод двогібридної системи дріжджів для дослідження взаємодії (Kim & Kende (2004) Proc Natl Acad Sci 101: 13374-13379). Назву GRF також дали іншому типу поліпептидів, які належать до родини поліпептидів 14-33 (de Vetten & Ferl (1994) Plant Physiol 106: 1593-1604), які взагалі не мають відношення до GRF поліпептидів, які є придатними для використання у способах винаходу. Трансгенні рослини Arabidopsis thaliana трансформовані поліпептидом рису GRF (OsGRF1) під контролем вірусного конститутивного промотера 35S CaMV, демонстрували курчаче листя, значне зменшення подовження первинних суцвіть, та уповільнений вихід у стрілку (van der Knapp et al. (2000) Plant Physiol 122: 695-704). Трансгенні рослини Arabidopsis thaliana з або одним або двома GRF поліпептидами Arabidopsis (AtGRF1 та AtGRF2) розвинули більші листя та сім'ядолі, мали уповільнений вихід у стрілку та були частково стерильними (внаслідок недостатньої кількості життєздатного пилку), у порівнянні з дикими типами рослин (Kim et al. (2003) Palnt J 36: 94-104). У патентній заявці США US2006/0048240, GRF поліпептид Arabidopsis thaliana ідентифіковано як SEQ ID NO: 33421, У патентній заявці США US2007/0022495, GRF поліпептид Arabidopsis thaliana ідентифіковано як SEQ ID NO: 1803 (також у даному документі посилаються як на G1438). Трансгенні рослини Arabidopsis, які надекспресують G1438 використовуючи 35S CaMV промотор, демонструють темно-зелене листя. Рівень техніки поліпептидів транслокації синовіальної саркоми (SYT) SYT є співактиватором транскрипції, який у рослинах утворює функціональний комплекс з активаторами транскрипції родини протеїнів GRF (фактором регуляції росту) (Kim HJ, Kende H (2004) Proc Nat Acad Sc 101: 13374-9). SYT, у даному документі, називають GIF для GRFвзаємодіючого фактору, та AN3 для angustifolia 3 у Horiguchi et al. (2005) Plant J 43: 68-78, GRF активатори транскрипції ділять структурні домени (у N-термінальному регіоні) з SWI/SNF протеїнами у хроматин-реконструюючих комплексах у дріжджів (van der Knaap E et al., (2000) Plant Phys 122: 695-704). Вважають, що спів-активатори транскрипції цих комплексів, беруть участь у рекрутінгу SWI/SNF комплексів для посилюючих та промоторних регіонів для 3 UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 здійснення локального трансформування хроматину (огляд Näär AM et al., (2001) Annu Rev Biochem 70: 475-501). Зміни у локальній структурі хроматину регулюють активацію транскрипції. Зокрема, вважають, що SYT взаємодіє з рослинним комплексом SWI/SNF для впливу на активацію транскрипції гену(ів) мішеней GRF (Kim HJ, Kende H (2004) Proc Nat Acad Sc 101: 13374-9). SYT належить до родини генів, яка складається з трьох членів у Arabidopsis. SYT поліпептид є гомологічним людському SYT. Людський поліпептид SYT є спів-активатором транскрипції (Thaete et al. (1999) Hum Molec Genet 8: 585-591). Людський SYT поліпептид характеризують три домени: (i) N-термінальний SNH (SYT N-термінальний гомологічний) домен, збережений у ссавців, рослин, нематод та риб; (ii) C-термінальний QPGY-збагачений домен, який переважно складається з гліцину, проліну, глютаміну та тирозину, з'являється з різноманітними інтервалами; (iii) Метіонін-збагачений (Met-збагачений) домен, розташований між двома попередніми доменами. У рослині SYT поліпептиди, SNH домен є добре збереженими. C-термінальний домен є збагаченим гліцином та глутаміном, проте не проліном або тирозином. Внаслідок цього його назвали QG-збагаченим доменом у протиріч з QPGY доменом ссавців. Також як і SYT ссавців, Met-збагачений домен ідентифікують по N-термінальному QG домену. QG-збагачений домен може бути по суті залишком C-термінального поліпептиду (мінус SHN домен); Met-збагачений домен зазвичай складає першу половину QG-збагаченого домену (з N-термінального до Cтермінального). Другий Met-збагачений домен може передувати SNH домену у SYT поліпептидах рослин (дивитись Фіг. 1). Продемонстровано, що SYT не функціональні мутанти та трансгенні рослини з пониженою експресією SYT, розвивають маленьке та вузьке листя та пелюстки, які мають менше клітин (Kim HJ, Kende H (2004) Proc Nat Acad Sc 101: 13374-9). Надекспресія AN3 у Arabidopsis thaliana привела до того, що листя рослин збільшилось на 20-30 % у порівнянні з диким типом (Horiguchi et al. (2005) Palnt J 43: 68-78). У японській патентній заявці 2004-350553 описано спосіб регулювання розміру листя у горизонтальному напрямку шляхом контролювання експресії гену AN3. Несподівано виявили, що підвищена експресія у рослині: (i) послідовності нуклеїнових кислот, яка кодує поліпептид Фактору Регулювання Росту (GRF); та (ii) послідовності нуклеїнових кислот, яка кодує поліпептид транслокації синовіальної саркоми (SYT), дає рослини з підвищеними показниками врожайності у порівнянні з рослинами з підвищеною експресією однієї з: (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид; або (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид. Відповідно до одного варіанту здійснення, винахід стосується різноманітних підвищених показників врожайності шляхом збільшення експресії у рослині: (i) послідовності нуклеїнових кислот, яка кодує поліпептид Фактору Регулювання Росту (GRF); та (ii) послідовності нуклеїнових кислот, яка кодує поліпептид транслокації синовіальної саркоми (SYT), де вказані показники врожайності є підвищеними по відношенню до рослин з підвищеною експресією однієї з: (i) послідовності нуклеїнових кислот, яка кодує GRF поліпептид, або (ii) послідовності нуклеїнових кислот, яка кодує SYT поліпептид. Підвищені показники врожайності включають одну або кілька з наступних: підвищена рання сила, підвищена біомаса надземної частини рослини, підвищений загальний вихід насіння на рослину, підвищений показник наповнення насіння, підвищена кількість (наповненого) насіння, підвищений індекс врожайності або підвищена маса тисячі зерен (TKW). Визначення термінів Поліпептид(и)/Протеїн(и) Терміни "поліпептид" та "протеїн" у даному документі використовують взаємозамінно та вони стосуються амінокислот у полімерній формі будь-якої довжини, зв'язані пептидними зв'язками. Полінуклеотид(и)/Нуклеїнова кислота(и)/Послідовність нуклеїнової кислот(и)/послідовність нуклеотидів. Терміни "полінуклеотид(и)", "послідовність нуклеїнової кислот(и)", "послідовність нукледтидів", "нуклеїнова кислота(и)” у даному документі використовують взаємозамінно та вони стосуються нуклеотидів, або рібонуклеотидів або дезоксирібонуклеотидів або їх комбінації, у полімерній не розгалуженій формі будь-якої довжини. Контрольні рослини(а) Вибір придатних контрольних рослин є стандартною частиною експерименту, ці рослини можуть включати дикі типи рослин або відповідні рослини які не містять потрібного гену. 4 UA 103176 C2 5 10 15 20 25 Контрольною рослиною зазвичай є рослина того виду або навіть того ж сорту що й рослина, яку оцінюють. Контрольною рослиною може також бути рослина, яка є нульовою зиготою рослини, яку оцінюють. Термін "контрольна рослина" як використано у даному документі, стосується не тільки всієї рослини, а й частин рослини, включаючи насіння та частини насіння. Гомолог(и) "Гомологи" протеїну включають пептиди, олігопептиди, поліпептиди, протеїни та ферменти, з заміщеннями по амінокислотам, делеціями та/або вставками по відношенню до такого ж не модифікованого протеїну та, які мають однакову біологічну та функціональну активність з не модифікованим протеїном з якого вони походять. Делеція стосується видалення однієї або кількох амінокислот з протеїну. Вставка стосується одного або кількох амінокислотних залишків, які включили у попередньо визначений сайт протеїну. Вставки можуть включати N-термінальні та/або C-термінальні приєднання, а також вставки в рамках послідовності однієї або кількох амінокислот. Зазвичай, вставки в межах амінокислотної послідовності є меншими ніж N- або C-термінальні приєднання, порядком приблизно 1-10 залишків. Приклади N- або C-термінальних приєднань протеїнів або пептидів включають зв'язувальний домен або домен активації активатора транскрипції, як використано у дріжджевій дво-гібридній системі, протеїни поверхні фагу, (гістидин)-6-tag, глутатіон S-трансфераза-tag, протеїн A, протеїн, що зв'язує мальтозу, дигідрофолат редуктаза, ® Tag•100 епітоп, c-myc епітоп, FLAG -епітоп, lacZ, CMP (пептид, що зв'язує калмодулін), HA епітоп, протеїн C епітоп та VSV епітоп. Заміщення стосується заміни амінокислот протеїну іншими амінокислотами з однаковими властивостями (такими як, однакова гідрофобність, гідрофільність, антигенність, схильність до утворення або розриву α-спіральних структур або β-пластинчастих структур). Амінокислотні заміщення зазвичай стосуються одного залишку, проте іноді до кластеру, внаслідок функціонального взаємозв'язку у поліпептиді; вставки зазвичай мають порядок від 1 дo 10 амінокислотних залишків. Амінокислотними заміщеннями зазвичай є заміщення консервативної амінокислоти. Таблиці консервативних заміщень є добре відомими у галузі (дивитись, наприклад, Creighton (1984) Proteins. W.H. Freeman and Company (Eds) та Таблиця 1 нижче). Таблиця 1: Приклади заміщень консервативних амінокислот Залишок Ala Arg Asn Asp Gln Cys Glu Gly His Ile Консервативні заміщення SerLys Gln; His Glu Asn Ser Asp Pro Asn; Gln Leu, Val Залишок Leu Lys Met Phe Ser Thr Trp Tyr Val Консервативні заміщення Ile; Val Arg; Gln Leu; Ile Met; Leu; Tyr Thr; Gly Ser; Val Tyr Trp; Phe Ile; Leu 30 35 40 45 Амінокислотні заміщення, делеції та/або вставки отримують за допомогою технологій синтезу білка, які є добре відомими у галузі, твердо фазний пептидний синтез тощо, або шляхом маніпуляцій з рекомбінантною ДНК. Способи маніпуляцій послідовностями ДНК для отримання варіантів заміщень, вставок або делецій протеїну є добре відомими у галузі. Наприклад, технології отримання мутантних заміщень у попередньо визначених місцях ДНК є добре відомими фахівцям у галузі та включають M13 мутагенез, T7-ген in vitro муиагенез (USB, Cleveland, OH), QuickChange сайт спрямований мутагенез (Stratagene, San Diego, CA), ПЛРопосередкований сайт-спрямований мутагенез або інші сайт-спрямовані протоколи мутагенезу. Похідні "Похідні" включають пептиди, олігопептиди, поліпептиди, які можуть, у порівнянні з послідовністю амінокислот природних форм білку, такого як потрібний білок, містити заміни амінокислот залишками неприродних амінокислот або добавки залишків неприродних амінокислот. "Похідні" протеїну також включають пептиди, олігопептиди, поліпептиди, які включають природні заміщені (глікозильовані, ацильовані, пренильовані, фосфорильовані, міристоільовані, сульфатовані тощо) або неприродно заміщені амінокислотні залишки у 5 UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 порівнянні з амінокислотною послідовністю природної форми поліпептиду. Похідна може також містити один або кілька не амінокислотних замісника або добавки, у порівнянні з амінокислотною послідовністю, похідними якої вони є, наприклад, репортерну молекулу або інший ліганд, ковалентно або нековалентно зв'язаний з амінокислотною послідовністю, наприклад, репортера молекула, яка полегшує виявлення, та амінокислотні залишки, що з'явились неприродним шляхом у амінокислотні послідовності природного білку. Ортолог(и)/Паролог(и) Ортологи та парологи включають еволюційні концепції, які використовували для характеризування спадкового взаємозв'язку генів. Парологи є генами в рамках однакових видів, які з'явились внаслідок дуплікації батьківського гену, ортологи є генами різних організмів, які з'явились внаслідок видоутворення, та також є похідними від загального батьківського гену. Домен Термін "домен" стосується набору амінокислот, збережених у певних положеннях лінії послідовностей еволюційно пов'язаних протеїнів. У той час як амінокислоти у інших положеннях можуть змінюватись між гомологами, амінокислоти, які є сильно консервативними у певних положеннях, вказують на амінокислоти які є природними у структурі, стабільність або функцію протеїну. ЇХ використовують як ідентифікатори для визначення будь-якого поліпептида, щодо того, чи він належить до визначеної родини поліпептидів, завдяки їх високій консервативності у лінії послідовностей родини білків гомологів. Мотив/Консенсусна послідовність/Відбиток Термін "мотив" або "консенсусна послідовність" або "відбиток" стосується коротко консервативної ділянки у послідовності еволюційно пов'язаних протеїнів. Мотиви є часто високо консервативними частинами доменів, проте також можуть включати тільки частину домену, або бути розташованими поза консервативним доменом (якщо всі амінокислоти мотиву випадають з визначеного домену). Гібридизація Термін "гібридизація" як вказано у даному документі, є процесом, де по суті гомологічні комплементарні послідовності нуклеотидів гібридизують одна одну. Процес гібридизації може повністю здійснюватись у розчині, тобто, обидві молекули комплементарних нуклеїнових кислот знаходяться у розчині. Процес гібридизації може здійснюватись з однією молекулою комплементарних нуклеїнових кислот, іммобілізованою на матрикс, такий як магнітні мікрочастинки, сефароза або інша смола. Процес гібридизації більш того може здійснюватись з однією молекулою комплементарних нуклеїнових кислот, іммобілізованою на тверду основу, таку як нітроцелюлоза або нейлонова плівка або, іммобілізованою, наприклад, фотолітографією на, наприклад, кремнієве скло (останнє відоме як матриці або мікроматриці послідовності нуклеїнової кислоти або як чіпи послідовності нуклеїнової кислоти). Для здійснення гібридизація молекули нуклеїнової зазвичай термічно або хімічно денатурують для плавлення подвійної нитки у дві одинарні нитки та/або для видалення "шпильки" або інших вторинних структур для одно спіральних молекул нуклеїнових кислот. Термін "жорсткість" стосується умов при яких здійснюють гібридизацію. Жорсткість гібридизації регулюється умовами, такими як температура, концентрація солі, іонна сила та буферна композиція для гібридизації. Зазвичай, нежорсткі умови означають що температура становить приблизно на 30 °C ніж температура плавлення (T m) для певної послідовності при визначеній іонній силі та pH. Середньо жорсткі умови означають що температура становить приблизно на 30 °C ніж температура плавлення (T m) та сильно жорсткі умови, коли температура на 10 °C нижче, ніж Tm. Гібридизацію з сильно жорсткими умовами використовують для ізолювання гібридизованих послідовностей, які є дуже подібними за послідовністю до бажаної послідовності нуклеїнової кислоти. Однак, послідовності нуклеїнових кислот можуть дещо відрізнятись та все ж кодувати ідентичний поліпептид, завдяки вродженості генетичного коду. Таким чином, умови гібридизації середньої жорсткості деколи потрібні для ідентифікації таких послідовностей молекул нуклеїнової кислоти. Tm означає температуру при визначеній іонній силі та pH, при якій 50 % бажаної послідовності гібридизується у ідеально підібраний зонд. T m залежить від характеристик розчину та основної композиції та довжини зонду. Наприклад, довші послідовності гібридизують специфічно при вищих температурах. Максимальну швидкість гібридизації отримують при температурі від приблизно 16 °C до 32 °C нижче Tm. Наявність моновалентних катіонів у розчині гібридизації зменшує електростатичне відштовхування між двома послідовностями ниток нуклеїнових кислот таким чином стимулюючи утворення гібриду; цей результат видно при концентраціях натрію до 0,4M (при вищих концентраціях цей результат може бути невидимим). Формамід знижує температуру плавлення ДНК-ДНК та ДНК-РНК дуплексів з 0,6 дo 0,7 °C на 6 UA 103176 C2 5 10 15 20 25 30 35 40 45 50 55 60 кожний процент формаміду, та додавання 50 % формам іду дозволяє здійснювати гібридизацію при 30-45 °C, хоча швидкість гібридизації зменшиться. Невідповідність пар основ зменшує швидкість гібридизації та термічну стабільність дуплексів. Для середніх та великих зондів Tm зменшується приблизно до 1 °C на % невідповідності основ. Tm розраховують, використовуючи наступні рівняння, в залежності від типу гібридів: 1) ДНК-ДНК гібриди (Meinkoth and Wahl, Anal. Biochem., 138: 267-284, 1984): + a b c -1Tm=81,5 °C+16,6xlog10[Na ] +0,41x%[G/C ] – 500x[L ] 0,61x% формамід 2) ДНК-РНК або РНК-РНК гібриди: + a b b 2c Tm=79,8+18,5 (log10[Na ] ) + 0,58 (%G/C ) + 11,8 (%G/C ) 820/L d 3) оліго-ДНК або оліго-РНК гібриди: Для
ДивитисяДодаткова інформація
Назва патенту англійськоюPlant with increased yield-related traits and method for its production
Автори англійськоюFrankard, Valerie, Reuzeau, Christophe
Назва патенту російськоюРастение с повышенными показателями урожайности и способ его получения
Автори російськоюФранкард Валерии, Резо Кристоф
МПК / Мітки
МПК: C12N 15/82, C12N 15/29, A01H 5/00
Мітки: спосіб, підвищеними, показниками, одержання, рослина, врожайності
Код посилання
<a href="https://ua.patents.su/373-103176-roslina-z-pidvishhenimi-pokaznikami-vrozhajjnosti-ta-sposib-oderzhannya.html" target="_blank" rel="follow" title="База патентів України">Рослина з підвищеними показниками врожайності та спосіб її одержання</a>
Спосіб одержання помісного молодняку свиней з підвищеними показниками відгодівельних та м’ясних якостей

Номер патенту: 41423
Опубліковано: 25.05.2009
Автори: Войтенко Світлана Леонідівна, Петренко Сергій Миколайович, Пісковий Микола Борисович
МПК: A01K 67/02
Мітки: молодняку, одержання, відгодівельних, помісного, спосіб, якостей, м'ясних, показниками, підвищеними, свиней
Формула / Реферат:
Спосіб одержання помісного молодняку свиней з підвищеними показниками відгодівельних та м'ясних якостей, який характеризується тим, що схрещують свиноматок миргородської породи із кнурами м'ясних генотипів французької селекції (породи п'єтрен та синтетичною лінією Maxter) із наступним одержанням помісного молодняку свиней та подальшою відгодівлею їх для забою.
Спосіб створення сортів озимого тритикале з підвищеними технологічними показниками якості зерна

Номер патенту: 44901
Опубліковано: 26.10.2009
Автори: Панченко Іван Архіпович, Щипак Геннадій Васильович, Чернобаб Раїса Андріївна, Суворова Катерина Юріївна
МПК: A01H 1/00
Мітки: спосіб, якості, технологічними, озимого, показниками, створення, підвищеними, сортів, тритікале, зерна
Формула / Реферат:
Спосіб створення сортів озимого тритикале з підвищеними технологічними показниками якості зерна, який включає досягнення збалансованості клейковинного комплексу, сформованого з відселектованої в різних агрокліматичних умовах сукупності ліній зі складної за типом розвитку і рівнем якості гібридної популяції (ступінчаста внутрішньовидова гібридизація ярих, озимих і дворучок), який відрізняється тим, що для підвищення технологічних властивостей...
Спосіб індикації та оцінки екологічного стану забрудненої важкими металами системи ґрунт-рослина за біохімічними показниками

Номер патенту: 92476
Опубліковано: 10.11.2010
Автори: Фатєєв Анатолій Іванович, Самохвалова Валентина Леонідівна, Якушко Віра Іванівна, Журавльова Інта Михайлівна
МПК: G01N 33/24
Мітки: індикації, екологічного, забрудненої, системі, ґрунт-рослина, важкими, оцінки, металами, спосіб, стану, показниками, біохімічними
Формула / Реферат:
Спосіб індикації та оцінки екологічного стану системи ґрунт-рослина за біохімічними показниками, який включає відбір зразків ґрунту, їх біохімічний аналіз, математичну обробку одержаних результатів, на базі яких визначається ступінь деградації ґрунту, який відрізняється тим, що відбір зразків проводять з ґрунтового шару не більш 35 см, аналіз біохімічних показників визначають і в ґрунті, і в рослинах, за результатами математичної обробки...
Спосіб поліпшення резистентності або толерантності до нематод у рослини та її потомства, молекула днк, злитий протеїн та рослина, що його експресує

Номер патенту: 71901
Опубліковано: 17.01.2005
Автори: Ервін Пітер Едвард, Аткінсон Говард Джон, МакФерсон Майкл Джон
МПК: C12N 15/82, A01H 5/00, C12N 9/02, G01N 33/53, C07K 14/81, G01N 33/50, C12N 9/00, C12N 5/10, G01N 33/15, C07K 19/00, C12N 15/09
Мітки: резистентності, спосіб, толерантності, потомства, протеїн, злитий, експресує, рослини, поліпшення, рослина, нематод, молекула, днк
Формула / Реферат:
1. Спосіб поліпшення резистентності або толерантності до нематод у рослини та її потомства, який включає включення у геном вищезгаданої рослини гена, що кодує злитий протеїн, який включає: перший протеїн або домен протеїну з антипатогенною активністю, пептид-лінкер та другий протеїн або домен протеїну з антипатогенною активністю, у котрого щонайменше один з протеїнів або доменів протеїнів з антипатогенною активністю...
Мутантний штам вірусу гепатиту b, виділена молекула нуклеїнової кислоти, що кодує мутантний головний поверхневий антиген штаму вірусу гепатиту в, вектор та векторна система для одержання поліпептиду, спосіб оде

Номер патенту: 73476
Опубліковано: 15.08.2005
Автори: Ун Чонг Джін, Чен Уей Нінг, Жао Йі, Лім Гек Кіоу
МПК: C07K 14/02, C12P 21/08, A61K 45/00, C12Q 1/68, C12N 15/51, C12N 7/00, G01N 33/576, G01N 33/68, C12P 21/02, A61P 31/20, G01N 33/50, C07K 16/08, C12N 15/09, A61P 1/16, A61K 39/29, A61P 35/00, G01N 33/15
Мітки: векторна, молекула, нуклеїнової, вірусу, кислоти, одержання, антиген, головний, штам, кодує, система, штаму, спосіб, поверхневий, оде, поліпептиду, гепатиту, вектор, мутантний, виділена
Формула / Реферат:
1. Виділений штам вірусу гепатиту В, визначений як штам Оon вірусу гепатиту В людини, що несе поверхневий антиген-'S'-133 (метіонін на треонін), що зберігається під номером доступу № Р97121504, Р97121505 і Р97121506 в Європейській колекції культур клітин від 15 грудня 1997 року.2. Виділена молекула нуклеїнової кислоти, яка кодує поліпептид, що є мутантним головним поверхневим антигеном штаму вірусу гепатиту В, яка відрізняється тим, що...
Попередній патент: Застосування аніоноактивних емульгаторів для підвищення твердості плавлених сирів із вмістом сухих речовин
Наступний патент: Спосіб дозрівання сиру у фользі
Випадковий патент: Різальний елемент