Імуногенна композиція проти цирковірусу свиней типу 2 (pcv2)
Номер патенту: 113192
Опубліковано: 26.12.2016
Автори: Чень Хсу-Чун Габріель, Ян Шу-Хсиан, Ко Тсунь-Юньг, Чень Юй-Сань







Формула / Реферат
1. Імуногенна композиція проти цирковірусу свиней типу 2 (PCV2), яка містить антигенний пептид, вибраний щонайменше з одного з:
(a) пептиду відкритої рамки зчитування 2 (ORF2) з PCV2, вибраного з групи, що складається з SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 18, SEQ ID NO: 22, SEQ ID NO: 55 і SEQ ID NO: 57; та
(b) рекомбінантного химерного білка, що включає від аміно-кінця до карбоксильного кінця рекомбінантного химерного білка:
пептид РЕ, що характеризується послідовністю SEQ ID NO: 35;
пептид з ORF2 з PCV2 з (а) та
сигнальний пептид KDEL, що характеризується послідовністю SEQ ID NO: 31.
2. Імуногенна композиція за п. 1, що додатково містить відкриті рамки зчитування (ORF), відмінні від ORF2 з PCV2, де ORF, відмінні від ORF2, включають ORF1 та ORF3.
3. Імуногенна композиція за п. 1, що додатково містить щонайменше один патогенний антиген, вибраний з групи, що включає антиген вірусу свинячого грипу (SIV), антиген вірусу репродуктивного та респіраторного синдрому свиней (PRRSV), антиген мікоплазми, антиген парвовірусу свиней (PPV), антиген бешихи свиней і антиген вірусу псевдосказу.
4. Імуногенна композиція за п. 1, що додатково містить одне або декілька, вибраних з групи, що включає наповнювачі, розчинник, емульгатор, суспендуючі засоби, засіб для розкладання, зв'язувальні засоби, середовище для ліків, стабілізуючі засоби, хелатоутворюючі засоби, розріджувач, загусники, консерванти, засіб, що обумовлює плинність, поверхнево-активну речовину, ад'ювант і біологічні носії.
5. Спосіб отримання фрагмента антигена PCV2, що включає
розташування послідовності ДНК, що кодує пептид ORF2 з PCV2, в біологічній системі експресії, де пептид ORF2 з PCV2 вибраний з групи, що складається з SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 18, SEQ ID NO: 22, SEQ ID NO: 55 і SEQ ID NO: 57; і
експресію послідовності ДНК, що кодує пептид ORF2 з PCV2, з отриманням фрагмента антигену імуногенної композиції проти PCV2.
6. Спосіб за п. 5, де послідовність ДНК, що кодує пептид з ORF2 з PCV2, вибрана з групи, що складається з SEQ ID NO: 5, SEQ ID NO: 7, SEQ ID NO: 9, SEQ ID NO: 11, SEQ ID NO: 17, SEQ ID NO: 21, SEQ ID NO: 54 і SEQ ID NO: 56.
Текст
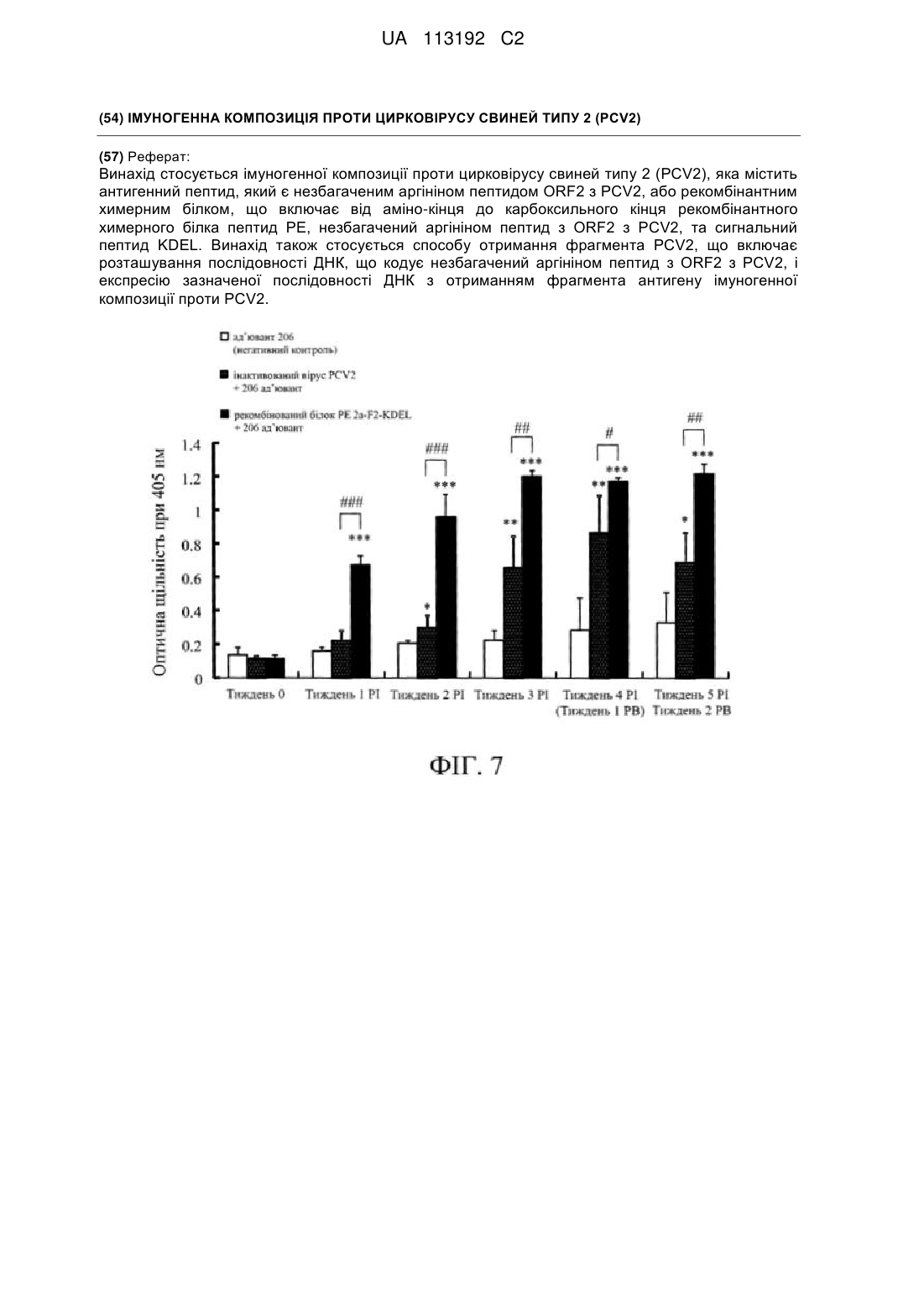
Реферат: Винахід стосується імуногенної композиції проти цирковірусу свиней типу 2 (PCV2), яка містить антигенний пептид, який є незбагаченим аргініном пептидом ORF2 з PCV2, або рекомбінантним химерним білком, що включає від аміно-кінця до карбоксильного кінця рекомбінантного химерного білка пептид РЕ, незбагачений аргініном пептид з ORF2 з PCV2, та сигнальний пептид KDEL. Винахід також стосується способу отримання фрагмента PCV2, що включає розташування послідовності ДНК, що кодує незбагачений аргініном пептид з ORF2 з PCV2, і експресію зазначеної послідовності ДНК з отриманням фрагмента антигену імуногенної композиції проти PCV2. UA 113192 C2 5 10 15 20 25 30 35 40 45 50 55 60 ПЕРЕДУМОВИ ВИНАХОДУ 1. ГАЛУЗЬ ТЕХНІКИ Даний винахід відноситься до субодиничної вакцини проти цирковірусу свиней типу 2 (PCV2), конкретно, до субодиничної вакцини проти PCV2, що містить пептид відкритої рамки зчитування (ORF2) з PCV2, який може надмірно експресуватися у якості антигена, і додатково придатний носій або ад’ювант. 2.ОПИС РІВНЯ ТЕХНІКИ Відомо, що цирковірус свиней типу 2 (PCV2) пов’язаний з синдромом мультисистемного виснаження після відлучення (PMWS) і синдромом дерматиту і нефропатії у свиней (PDNS). PMWS вперше був виявлений у свиней в Канаді в 1991 році, а згодом було повідомлено про його зустрічальність в усьому світі. Синдром призвів до великої втрати в свинарській індустрії у всьому світі. Головні симптоми PMWS включають прогресуючу втрату ваги, тахіпное, диспное, розлиття жовчі і т. д. Візуальні патологічні зміни в тканині включають лімфоцитарний та гранулематозний інфільтрат, лімфаденопатію, лімфоцитарний та гранулематозний гепатит і нефрит. Цирковірус свиней (PCV) вперше був виявлений в лінії клітин нирки свині (PK-15, ATCC CCL33) в 1982 році. Хоча цирковірус свиней може постійно інфікувати клітини PK-15, вірус не спричиняє цитопатичного ефекту (CPE) в інфікованих клітинах PK-15. Навіть, незважаючи на те, що цирковірус свиней може інфікувати свиней, він не призводить до уражень в інфікованих свиней. Вірус назвали цирковірусом свиней типу 1 (PCV1). PCV1 є дванадцятигранним вірусом з одноланцюговою ДНК з кільцевим геномом з 1759 п. о. Отриманий з PK-15 PCV класифікували в сімейство Circoviridae в 1995 році. Отриманий з PK-15 PCV1 вважається апатогенним. На відміну від цього, вірусний мутант, виділений із свиней з PMWS в 1997 році, є патогенним і називається цирковірус свиней типу 2 (PCV2). Синдром мультисистемного виснаження після відлучення (PMWS) є високо контагіозним захворюванням свиней. Він головним чином інфікує поросних свиноматок та їх поросят і серйозно впливає на здоров’я свиней. PCV2 є вірусом з одноланцюговою кільцевою ДНК діаметром 17 нм, і розмір його геному складає 1,76 т. н. Геномний аналіз за допомогою програмного забезпечення показує сумарно 11 відкритих рамок зчитування (ORF), що транскрибуються в напрямках за годинниковою стрілкою та проти годинникової стрілки. Серед 11 ORF, ORF1 і ORF2 імовірно являють собою найбільш важливі гени. Ген ORF1 кодує білки Rep і Rep’, які пов’язані з реплікацією вірусу. Відомо, що ген ORF2 кодує імуногенний структурний білок капсида з PCV2, який застосовують для індукції імунної відповіді в організмів. Інактивована вакцина проти PCV2 є найбільш розповсюдженою комерційно доступною вакциною проти PCV2. Однак розробка інактивованої вакцини потребує, щоб лінії клітин не містили забруднювача, а можливість неповної інактивації вірусу за допомогою хімічних реактивів є найбільш значним недоліком інактивованої вакцини. Іншим недоліком є те, що антигенні структури вірусу можуть бути змінені шляхом хімічної обробки, що призведе до неможливості індукції достатньої імунної відповіді для знищення вірусу і неможливості захисту свиней від інфікування захворюванням. Таким чином, розробка інактивованої вакцини може бути складною і дорогою, а безпека вакцини може викликати питання. На відміну від інактивованої вакцини, в якій цілий вірус є вакцинним антигеном, в субодиничній вакцині застосовують частину білків із патогена у якості білка-антигена і білокантиген уводять щепленням тваринам або людям для індукції імунітету. Субодиничну вакцину можна отримувати шляхом клонування генів, що кодують білки-антигени з патогенів, а потім отримання великих кількостей білків-антигенів за допомогою генної інженерії. Безпека є найбільш значною перевагою субодиничної вакцини, оскільки в ній застосовують частини патогена замість цілого патогена для щеплення свиней з відсутністю проблеми неповної інактивації. В традиційній субодиничній вакцині проти PCV2 застосовують білок ORF2 з PCV2 у якості білка-антигена; тим не менш, рівень експресії повнорозмірного білка ORF2 з PCV2 в прокаріотичних системах експресії є достатньо низьким і не відповідає вимогам отримання вакцини. Таким чином, розробка фрагментів антигена з ORF2 з PCV2, які можуть характеризуватися високим рівнем експресії в біологічних системах експресії, доцільна для комерційного застосування субодиничної вакцини проти PCV2. КОРОТКИЙ ОПИС ВИНАХОДУ Дане розкриття відноситься до послідовностей ДНК, що кодують фрагменти білка з ORF2 з PCV2, і фрагменти білка з ORF2 з PCV2 можуть характеризуватися високим рівнем експресії в біологічних системах експресії. Дане розкриття також відноситься до субодиничної вакцини проти PCV2, в якій фрагменти 1 UA 113192 C2 5 10 15 20 25 30 35 40 45 50 55 60 білка з ORF2 з PCV2, які можуть характеризуватися високим рівнем експресії в біологічних системах експресії, застосовують в якості білків-антигенів для введення щепленням тваринам для індукції відповідного імунітету проти інфекції PCV2 у тварин. Дане розкриття додатково відноситься до субодиничної вакцини, розробленої шляхом генної інженерії для отримання за допомогою простого способу недорогої, високоочищеної, з хорошим рівнем безпеки субодиничної вакцини проти PCV2. У зв’язку з тим фактом, що рівень експресії повнорозмірного білка ORF2 з PCV2 в біологічних системах експресії є достатньо низьким, для досягнення зазначених вище цілей застосовують генну інженерію. Послідовності ДНК (такі як SEQ ID NO: 1), які кодують повнорозмірні білки ORF2 з PCV2, розрізали на фрагменти різних розмірів і фрагменти ДНК вставляли в вектори експресії, а потім експресували в хазяїнах. Вимірювали рівні експресованих білків для визначення того, які фрагменти ДНК можуть давати високі рівні білків в системах експресії білків. Результати аналізу білкової послідовності і досліджень експресії білка показують, що в повнорозмірному білку ORF2 з PCV2 є приблизно 30 аргінінів, з яких щонайменше дві треті аргінінів виявлені на N-кінці білка ORF2 з PCV2, і що чим більша кількість аргінінів на N-кінці буде видалена, тим вище буде рівень експресії фрагмента білка ORF2. Крім того, після того як перші 234 нуклеотиди на 5’-кінці повнорозмірної послідовності ДНК з ORF2 з PCV2 були видалені, фрагмент білка, що кодується фрагментом ДНК, що залишився (тобто від нуклеотиду 235 на 5’-кінці до стоп-кодона), можна експресувати у великому масштабі. Внаслідок цього, субодинична вакцина проти PCV2, представлена в даному розкритті, містить антигенний пептид з PCV2 з придатним носієм або ад’ювантом. Антигенний пептид може експресуватися в великому масштабі в системах експресії і є незбагаченим аргініном пептидом з ORF2 з PCV2. Кількості аргінінів незбагаченого аргініном пептиду з ORF2 з PCV2 складають не більше половини (1/2) кількостей аргінінів збагаченого аргініном домену на N-кінці повнорозмірного ORF2 з PCV2. В одному варіанті здійснення, якщо число аргінінів збагаченого аргініном домену на N-кінці повнорозмірного ORF2 з PCV2 складає 20, то число аргінінів незбагаченого аргініном пептиду з ORF2 з PCV2 становить одне з цілих чисел від 0 до 10. В іншому варіанті здійснення, якщо число аргінінів збагаченого аргініном домену на N-кінці повнорозмірного ORF2 з PCV2 складає 21, то число аргінінів незбагаченого аргініном пептиду з ORF2 з PCV2 становить одне з цілих чисел від 0 до 10. В ще одному варіанті здійснення, якщо число аргінінів збагаченого аргініном домену на N-кінці повнорозмірного ORF2 з PCV2 складає 22, то число аргінінів незбагаченого аргініном пептиду з ORF2 з PCV2 становить одне з цілих чисел від 0 до 11. В одному варіанті здійснення збагачений аргініном домен характеризується залишками 1-78 на N-кінці повнорозмірного ORF2 з PCV2 (який кодується нуклеотидами 1-234 на 5'-кінці повнорозмірної послідовності ДНК з ORF2 з PCV2). В одному варіанті здійснення збагачений аргініном пептид характеризується пептидною послідовністю від залишку 79 до останнього амінокислотного залишку на С-кінці повнорозмірного ORF2 з PCV2 (який кодується послідовністю ДНК від нуклеотиду 235 на 5'-кінці до стоп-кодона на 3'-кінці повнорозмірної послідовності ДНК з ORF2 з PCV2). Розкритий в даному документі ORF2 з PCV2 має повнорозмірну пептидну послідовність, що характеризується щонайменше 80%, переважно 85%, більш переважно 90%, ще більш переважно 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% або 99% гомології з однією з SEQ ID NO: 2, SEQ ID NO: 16, SEQ ID NO: 20, SEQ ID NO: 51 і SEQ ID NO: 53. У переважному варіанті здійснення розкритий в даному документі ORF2 з PCV2 характеризується повнорозмірною пептидною послідовністю однією з SEQ ID NO: 2, SEQ ID NO: 16, SEQ ID NO: 20, SEQ ID NO: 51 і SEQ ID NO: 53. На додаток до цього, дане розкриття відноситься до химерного білка ORF2 з PCV2 в якості антигенного пептиду із застосуванням функціональних елементів рецептор-зв’язувального домену I і трансмембранного домену II, що націлює, екзотоксину А з Pseudomonas aeruginosa (тобто "білка PE") і сигналу затримки в ендоплазматичному ретикулумі (ER) (тобто "сигнального пептиду KDEL"). Білок PE з Pseudomonas aeruginosa можна застосовувати в якості гіда цільового білка до його цільового положення, згідно з наведеним далі способом. Спочатку, рецепторзв’язувальний домен екзотоксину А з Pseudomonas aeruginosa забезпечує зв’язування з рецепторами на мембрані клітин-мішеней (CD8+ Т-клітин), а потім ліганд-рецепторні комплекси проникають в ендосоми клітин-мішеней шляхом ендоцитозу. Після ферментативного відщеплення за допомогою протеази в ендосомах укорочені фрагменти білка (що містять трансмембранний домен, що націлює, і зв’язаний цільовий білок) доставляються в тільця 2 UA 113192 C2 5 10 15 20 25 30 35 40 45 50 55 60 Гольджі та ендоплазматичний ретикулум (ER) і далі переміщуються в цитоплазму клітинмішеней за допомогою трансмембранного домену, що націлює. Крім того, при приєднанні сигнального пептиду KDEL на C-кінець цільового білка цільовий білок буде транспортуватися в ER за допомогою сигнального пептиду KDEL, а потім взаємодіяти в ER. Таким чином, дане розкриття відноситься до химерного білка ORF2 з PCV2 в якості антигенного пептиду шляхом застосування функціональних елементів білка PE і сигнального пептиду KDEL. Білок PE і сигнальний пептид KDEL зливають з фрагментом білка ORF2 з PCV2 на N- та C-кінцях, відповідно, з отриманням химерного білка (PE-фрагмент ORF2-KDEL) для введення щепленням тваринам з метою індукції достатньої імунної відповіді проти інфекції PCV2 у тварин. В деяких варіантах здійснення застосовувані в даному документі антигенні пептиди включають без обмежень фрагменти ORF2 з PCV2. Фрагменти ORF2 мають пептидну послідовність, що характеризується щонайменше 80%, переважно 85%, більш переважно 90%, ще більш переважно 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% або 99% гомології з однією з SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 18, SEQ ID NO: 22, SEQ ID NO: 55 та SEQ ID NO: 57. В переважному варіанті здійснення фрагменти ORF2 з PCV2 мають пептидну послідовність однієї з SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 10, SEQ ID NO: 12, SEQ ID NO: 18, SEQ ID NO: 22, SEQ ID NO: 55 та SEQ ID NO: 57, і були отримані за допомогою генної інженерії. Послідовності ДНК (SEQ ID NO: 5, 7, 9, 11, 17, 21, 54, 56), що кодують фрагменти ORF2 з PCV2, клонували в вектори експресії з отриманням плазмід, що містять фрагмент ДНК, який кодує антигенний пептид. Плазміди потім трансформували в клітини-хазяїни для експресії антигенних пептидів. Розкриті в даному документі химерні білки (PE-фрагмент ORF2-KDEL) отримували за допомогою генної інженерії. Послідовності ДНК (SEQ ID NO: 5, 7, 9, 11, 17, 21, 54, 56), що кодують фрагменти ORF2 з PCV2, послідовність ДНК (SEQ ID NO: 34), що кодує білок PE, і послідовність ДНК (SEQ ID NO: 30), що кодує сигнальний пептид KDEL, клонували у вектори експресії з отриманням плазмід, які містять фрагмент ДНК, що кодує химерний білок. Плазміди потім трансформували в клітини-хазяїни для експресії химерних білків. Вектори експресії включають без обмеження вектори pET і вектори pGEX. В одному варіанті здійснення вектором експресії є pET24a. Системи експресії (клітини-хазяїни) включають без обмеження прокаріотичні системи експресії (такі як Escherichia coli), еукаріотичні системи експресії (такі як клітини тварин (наприклад, клітина CHO) і клітини рослин). В одному варіанті здійснення системою експресії є E. coli. Ад’ювант включає без обмеження водний ад’ювант (такий як гідроксид алюмінію), алюмокалієвий галун, неповний ад’ювант Фрейнда, масляний ад’ювант, розчинний у воді ад’ювант або ад’ювант за типом емульсії "вода-в-маслі-у-воді" (W/O/W). В одному варіанті здійснення ад’ювантом є масляний ад’ювант. Розкрита в даному документі імуногенна композиція додатково містить антигенні пептиди з інших ORF з PCV2. Інші ORF з PCV2 включають без обмеження ORF1 та ORF3. На додаток до цього, розкрита в даному документі імуногенна композиція додатково містить патогенний антиген. Патогенний антиген обраний з групи, що включає антиген вірусу свинячого грипу (SIV), антиген вірусу репродуктивного та респіраторного синдрому свиней (PRRSV), антиген мікоплазми, антиген парвовірусу свиней (PPV), антиген бешихи свиней і антиген вірусу псевдосказу (хвороби Ауєскі). На додаток до цього, розкрита в даному документі імуногенна композиція додатково містить одне або декілька з наступного: наповнювачі, розчинник, емульгатор, суспендуючий засіб, засіб для розкладання, зв’язувальний засіб, середовище для ліків, стабілізуючий засіб, хелатоутворюючий засіб, розріджувач, загусник, консервант, засіб, що обумовлює плинність, поверхнево-активну речовину, ад’ювант і біологічні носії. Значення технічних і наукових термінів, які описані в даному документі, зможе легко зрозуміти кожний фахівець в даній галузі. Даний винахід більш докладно описаний у наведених далі ілюстративних прикладах. Хоча приклади можуть являти собою тільки обрані варіанти здійснення даного винаходу, слід розуміти, що наведені далі приклади є ілюстративними, а не обмежуючими. КОРОТКИЙ ОПИС ГРАФІЧНИХ МАТЕРІАЛІВ На фіг. 1 представлені результати філогенетичного аналізу геномних послідовностей PCV2. На фіг. 2 проілюстрована схематична діаграма фрагментів ORF2 з PCV2, в яких праймерами для ПЛР є pF1, pF2, pF3, pF4, pR1, pR2 та pR3. На фіг. 3 показані результати аналізу SDS-PAGE рекомбінантних білків різних фрагментів 3 UA 113192 C2 5 10 15 20 25 30 35 40 45 50 55 2a-ORF2 з PCV2, експресованих в E. coli. Цілі білки E. coli збирали через 6 годин після індукції за допомогою IPTG і потім розділяли за допомогою 15% SDS-PAGE, в якому доріжки 1-9 показують маркери молекулярної ваги, порожній вектор pET24a (в якості негативного контролю), фрагмент ORF2 2a-F1 з PCV2 (12,7 кДа), фрагмент ORF2 2a-F2 з PCV2 (11,6 кДа), фрагмент ORF2 2a-F3 з PCV2 (12,1 кДа), фрагмент ORF2 2a-F4 з PCV2 (16,5 кДа), фрагмент ORF2 2a-F5 з PCV2 (21,4 кДа), фрагмент ORF2 2a-F6 з PCV2 (20,8 кДа) і повнорозмірний 2a-ORF2 з PCV2 (27,5 кДа), відповідно. На фіг. 4 представлені результати вестерн-блотингу рекомбінантних білків різних фрагментів 2a-ORF2 з PCV2. Доріжки 1-9 показують маркери молекулярної ваги, порожній вектор pET24a (в якості негативного контролю), фрагмент ORF2 2a-F1 з PCV2 (12,7 кДа), фрагмент ORF2 2a-F2 з PCV2 (11,6 кДа), фрагмент ORF2 2a-F3 з PCV2 (12,1 кДа), фрагмент ORF2 2a-F4 з PCV2 (16,5 кДа), фрагмент ORF2 2a-F5 з PCV2 (21,4 кДа), фрагмент ORF2 2a-F6 з PCV2 (20,8 кДа) і повнорозмірний 2a-ORF2 з PCV2 (27,5 кДа), відповідно. На фіг. 5 представлені результати ELISA PCV2 зразків сироватки, зібраних в різні моменти часу у щурів, вакцинованих рекомбінантними білками різних фрагментів 2a-ORF2 з PCV2, відповідно. *, p < 0,05. (PI: після імунізації; PB: після бустер-ін’єкції) На фіг. 6 проілюстровані результати ELISA PCV2 зразків сироватки, зібраних в різні моменти часу у мишей, вакцинованих рекомбінантними білками фрагмента 2a-F2 і рекомбінантними білками фрагмента PE-2a-F2-KDEL, відповідно. *, p < 0,05; **, p < 0,01; ***, p < 0,001; #, p < 0,05; ##, p < 0,01; ###, p < 0,001. (PI: після імунізації; PB: після бустер-ін’єкції). На фіг. 7 показані результати ELISA PCV2 зразків сироватки, зібраних в різні моменти часу у мишей, вакцинованих рекомбінантними білками фрагмента PE-2a-F2-KDEL і вакциною з цілим вірусом PCV2, відповідно. *, p < 0,05; **, p < 0,01; ***, p < 0,001; #, p < 0,05; ##, p < 0,01; ###, p < 0,001. (PI: після імунізації; PB: після бустер-ін’єкції) ДЕТАЛЬНИЙ ОПИС ПЕРЕВАЖНОГО ВАРІАНТА ЗДІЙСНЕННЯ Приклад 1. Будування та визначення антигенних пептидів субодиничної вакцини проти PCV2 У зв’язку з тим фактом, що рівень експресії повнорозмірного білка ORF2 з PCV2 в біологічних системах експресії є досить низьким, послідовності ДНК, що кодують повнорозмірні білки ORF2 з PCV2, розрізали на фрагменти різних розмірів і фрагменти ДНК вбудовували в вектори експресії, а потім експресували в хазяїнах. Рівні експресованих білків вимірювали для визначення того, які фрагменти ДНК можуть давати високі рівні білків в білкових системах експресії. Широко застосовувану прокаріотичну систему експресії (Escherichia coli) застосовували у наведених далі прикладах для аналізу і в ілюстративних цілях. 1. Ампліфікація фрагментів гена ORF2 з PCV2 різних розмірів Повнорозмірний ген ORF2 з PCV2, застосовуваний в даному прикладі, характеризується послідовністю SEQ ID NO: 1. Послідовності гена ORF2 з PCV2, представлені у Wang et al. (Wang et al., Virus Research 2009, Genetic variation analysis of Chinese strains of porcine circovirus type 2.), застосовували в якості стандартних послідовностей для філогенетичного аналізу SEQ ID NO: 1. Результат показав, що послідовність ORF2 з PCV2 (SEQ ID NO: 1) належить до підгрупи PCV2 2a (яка показана на фіг. 1). Праймери (які описані в таблиці 1) призначені для ампліфікації фрагментів різних розмірів гена ORF2 з PCV2 2a (2a-ORF2) за допомогою полімеразної ланцюгової реакції (ПЛР). Як показано на фіг. 2, повнорозмірний ген 2a-ORF2 ампліфікували за допомогою специфічних праймерів pF1 (прямий праймер) і pR1 (зворотний праймер) шляхом ПЛР. Повнорозмірний ген 2a-ORF2 (2a-ORF2) характеризується послідовністю ДНК SEQ ID NO: 1, а фрагменти різних розмірів 2a-ORF2 з PCV2 називали 2a-F1, 2a-F2, 2a-F3, 2a-F4, 2a-F5 і 2a-F6, відповідно. Фрагмент 2a-F1 характеризується послідовністю ДНК SEQ ID NO: 3, яка є нуклеотидами 1-234 на 5’-кінці 2a-ORF2. Фрагмент 2a-F2 характеризується послідовністю ДНК SEQ ID NO: 5, яка є нуклеотидами 235-468 на 5’-кінці 2a-ORF2. Фрагмент 2a-F3 характеризується послідовністю ДНК SEQ ID NO: 7, яка є нуклеотидами 469-699 на 5’-кінці 2aORF2. Фрагмент 2a-F4 характеризується послідовністю ДНК SEQ ID NO: 9, яка є нуклеотидами 349-699 на 5’-кінці 2a-ORF2. Фрагмент 2a-F5 характеризується послідовністю ДНК SEQ ID NO: 11, яка є нуклеотидами 235-699 на 5’-кінці 2a-ORF2. Фрагмент 2a-F6 характеризується послідовністю ДНК SEQ ID NO: 13, яка є нуклеотидами 1-468 на 5’-кінці 2a-ORF2. Усі прямі праймери містили сайт рестрикції Hind III, а всі зворотні праймери містили сайт рестрикції Xho I. Таблиця 1. Перелік послідовностей праймерів для ампліфікації фрагментів гена ORF2 з PCV2 різних розмірів 4 UA 113192 C2 примітка Фрагмент Праймер pF1 ORF2 pR1 pF1 F1 pR3 pF2 F2 pR2 pF3 F3 pR1 pF4 F4 pR1 pF2 F5 pR1 pF1 F6 pR2 Прямий праймер Зворотний праймер Послідовність CCCAAGCTTGCATGACGTATCCAAGGA SEQ ID NO: 23 GGCG CCGCTCGAGGGGTTTAAGTGGGGGGTC SEQ ID NO: 24 TTTA Прямий праймер Зворотний праймер CCCAAGCTTGCATGACGTATCCAAGGA SEQ ID NO: 23 GGCG CCGCTCGAGGTCGTTAATATTAAATCTC SEQ ID NO: 28 ATC Прямий праймер Зворотний праймер CCCAAGCTTGCTTTGTTCCCCCGGGAG SEQ ID NO: 25 GGGG CCGCTCGAGGTAGGAGAAGGGTTGGG SEQ ID NO: 26 GGATT Прямий праймер Зворотний праймер CCCAAGCTTGCCACTCCCGGTACTTTAC SEQ ID NO: 27 CCC CCGCTCGAGGGGTTTAAGTGGGGGGTC SEQ ID NO: 24 TTTA Прямий праймер Зворотний праймер CCCAAGCTTGCGGAGTGGGCTCCACTG SEQ ID NO: 29 CTGT CCGCTCGAGGGGTTTAAGTGGGGGGTC SEQ ID NO: 24 TTTA Прямий праймер Зворотний праймер CCCAAGCTTGCTTTGTTCCCCCGGGAG SEQ ID NO: 25 GGGG CCGCTCGAGGGGTTTAAGTGGGGGGTC SEQ ID NO: 24 TTTA Прямий праймер Зворотний праймер CCCAAGCTTGCATGACGTATCCAAGGA SEQ ID NO: 23 GGCG CCGCTCGAGGTAGGAGAAGGGTTGGG SEQ ID NO: 26 GGATT Примітка: послідовності, позначені знизу “____”, є послідовностями сайту рестрикції Hind III (AAGCTT), а послідовності, позначені знизу “ ”, є послідовностями сайту рестрикції Xho I (CTCGAG). 5 10 15 20 Умови для реакції ПЛР включали 95°C протягом 5 хвилин, 25 циклів 95°C протягом 30 секунд, 55°C протягом 30 секунд, і 72°C протягом 30 секунд, і 72°C протягом 5 хвилин для елонгації. Розміри продуктів ПЛР 2a-ORF2, 2a-F1, 2a-F2, 2a-F3, 2a-F4, 2a-F5 і 2a-F6 становили 699 п. о., 234 п. о., 234 п. о., 231 п. о., 351 п. о., 465 п. о. і 468 п. о., відповідно, і підтверджені за допомогою електрофорезу в 2% агарозному гелі. Після підтвердження, продукти ПЛР очищали за допомогою набору для очищення PCR-M (Viogene). 2. Будування плазмід pET24a, що містять фрагменти ORF2 з PCV2 різних розмірів. Один (1) мкг очищених продуктів ПЛР і 1 мкг вектора експресії pET24a (Novagen) розщеплювали за допомогою двох ферментів рестрикції (New England Biolabs) 1 мкл Hind III і 1 мкл Xho I, відповідно, протягом 8 годин при 37°C. Після реакції розщеплення за допомогою ферментів рестрикції розщеплені продукти ПЛР і вектор експресії pET24 очищали за допомогою системи PCR-M Clean up System (Viogene), відповідно. Очищені продукти ПЛР лігували з очищеним вектором експресії pET24a з отриманням плазмід pET24a-2a-F1, pET24a-2a-F2, pET24a-2a-F3, pET24a-2a-F4, pET24a-2a-F5, pET24a-2a-F6 і pET24a-2a-ORF2, а потім плазміди трансформували в клітини-хазяїни (E. coli). Відбирали трансформантів, що містять продукти ПЛР, і послідовності ДНК підтверджували за допомогою секвенування ДНК. Потім отримували бактерії, що містять зазначені вище плазміди. 3. Експресія білка і підтвердження фрагментів ORF2 з PCV2 різних розмірів 5 UA 113192 C2 5 10 15 20 25 30 35 40 45 50 55 60 Бактерії, що містять зазначені вище плазміди, інкубували в 2 мл середовища LB при 37°C протягом 16-18 годин і потім інокулювали в співвідношенні 1:50 в середовище LB, що містить 25 мкг/мл канаміцину, для наступного інкубування при 37°C в інкубаторі з 200 об./хв. до тих пір, поки O.D. при 600 нм не становила 0,6. Після цього, β-D-тригалактозид (IPTG) додавали до кінцевої концентрації 1 мM і клітини-хазяїни E. coli інкубували при 37°C в інкубаторі з 200 об./хв. протягом ще 6 годин. Потім 1 мл клітин хазяїнів E. coli центрифугували при 10000xg, і осад обробляли шляхом B-PERTM Bacterial Protein Extraction (придбаним у PIERCE) для перевірки того, чи є рекомбінантні білки розчинними білками або знаходяться в тільцях включення, за допомогою наступних етапів. Осад додавали в 40 мкл реагенту, перемішували за допомогою вортексного шейкера протягом 1 хвилини і центрифугували при 10000 xg для відділення розчинних білків (верхня частина) від нерозчинних тілець включень (нижня частина). Розчинні білки розчиняли в 1x буфері для розведення зразка для SDS-PAGE, а осад додавали в 2x буфера для розведення зразка для SDS-PAGE. Зразки поміщали в термоблок з температурою 100°C на 20 хвилин, а потім центрифугували. Супернатант аналізували за допомогою 15% SDSPAGE для вимірювання експресії рекомбінантних білків. Рекомбінантні білки отримували шляхом індукції бактеріальної культури за допомогою IPTG. Рекомбінантний білок 2a-ORF2 характеризується амінокислотною послідовністю SEQ ID NO: 2. Рекомбінантний білок 2a-F1 характеризується амінокислотною послідовністю SEQ ID NO: 4. Рекомбінантний білок 2a-F2 характеризується амінокислотною послідовністю SEQ ID NO: 6. Рекомбінантний білок 2a-F3 характеризується амінокислотною послідовністю SEQ ID NO: 8. Рекомбінантний білок 2a-F4 характеризується амінокислотною послідовністю SEQ ID NO: 10. Рекомбінантний білок 2a-F5характеризується амінокислотною послідовністю SEQ ID NO: 12. Рекомбінантний білок 2a-F6 характеризується амінокислотною послідовністю SEQ ID NO: 14. Експресію рекомбінантних білків аналізували за допомогою SDS-PAGE, і результати показані на фіг. 3, причому доріжка 1 показує маркер молекулярної ваги, доріжка 2 показує негативний контроль (пустий вектор pET24a), доріжка 3 показує рекомбінантний білок 2a-F1 (12,7 кДа), доріжка 4 показує рекомбінантний білок 2a-F2 (11,6 кДа), доріжка 5 показує рекомбінантний білок 2a-F3 (12,1 кДа), доріжка 6 показує рекомбінантний білок 2a-F4 (16,5 кДа), доріжка 7 показує рекомбінантний білок 2a-F5 (21,4 кДа), доріжка 8 показує рекомбінантний білок 2a-F6 (20,8 кДа) і доріжка 9 показує повнорозмірний рекомбінантний білок 2a-ORF2 (27,5 кДа). Серед 6 фрагментів 2a-ORF2 фрагменти 2a-F2, 2a-F3, 2a-F4 і 2a-F5 експресувалися в великих кількостях (фіг. 3, доріжки 4-7). Розраховані розміри фрагментів білків 2a-F2, 2a-F3, 2aF4 і 2a-F5 відповідали їх фактичним розмірам, що становили 11,6 кДа, 12,1 кДа, 16,5 кДа і 21,4 кДа, відповідно. Далі, застосовували вестерн-блотинг для перевірки, чи були експресовані рекомбінантні білки фрагментами 2a-ORF2 з PCV2. Після електрофорезу SDS-PAGE переносили на поліамідну мембрану PVDF. Отриману мембрану блокували за допомогою блокувального буфера, який являє собою TBST (10 мM трис-HCl, pH 8,0, 150 мM NaCl, 0,1% Твін 20), що містить 5% знежиреного молока протягом 1 години при кімнатній температурі для запобігання неспецифічного зв’язування білків. Після цього білки виявляли за допомогою моноклональних антитіл миші, кон’югованих з лужною фосфатазою (AP), до химерних білків з 6X гістидиновою міткою протягом 1 години при кімнатній температурі з наступним шестиразовим промиванням за допомогою TBST, по 5 хвилин кожне. Після промивання додавали субстрат NBT/BCIP (Bio-Rad). Через 10 хвилин реакцію зупиняли шляхом додавання води. Результати вестерн-блотинга показані на фіг. 4, причому доріжка 1 показує маркер молекулярної ваги, доріжка 2 показує негативний контроль (пустий вектор pET24a), доріжка 3 показує рекомбінантний білок 2a-F1 (12,7 кДа), доріжка 4 показує рекомбінантний білок 2a-F2 (11,6 кДа), доріжка 5 показує рекомбінантний білок 2a-F3 (12,1 кДа), доріжка 6 показує рекомбінантний білок 2a-F4 (16,5 кДа), доріжка 7 показує рекомбінантний білок 2a-F5 (21,4 кДа), доріжка 8 показує рекомбінантний білок 2a-F6 (20,8 кДа), і доріжка 9 показує повнорозмірний рекомбінантний білок 2a-ORF2 (27,5 кДа). Результати вестерн-блотинга співпадали з результатами аналізу SDS-PAGE (який показаний на фіг. 3). Повнорозмірний рекомбінантний білок 2a-ORF2 не міг експресуватися (фіг. 4, доріжка 9). Серед 6 фрагментів з 2a-ORF2 фрагменти 2a-F2, 2a-F3, 2a-F4 і 2a-F5 експресувалися в великих кількостях (фіг. 4, доріжки 4-7). Розраховані розміри фрагментів білків 2a-F2, 2a-F3, 2a-F4 і 2a-F5 відповідали їх фактичним розмірам, що становили 11,6 кДа, 12,1 кДа, 16,5 кДа і 21,4 кДа, відповідно. Однак, була відсутня експресія рекомбінантного білка 2a-F1 або рекомбінантного білка 2a-F6 (фіг. 4, доріжки 3 і 8). Оскільки як рекомбінантний білок 2a-F1, так і рекомбінантний білок 2a-F6 містили нуклеотиди 1234 на 5’-кінці повнорозмірної послідовності ДНК з 2a-ORF2 з PCV2 (див. фіг. 2), деякі послідовності в нуклеотидах 1-234 могли зачіпати експресію білка ORF2 з PCV2. Аналіз 6 UA 113192 C2 5 амінокислотних послідовностей фрагмента 2a-F1 (нуклеотиди 1-234 на 5’-кінці 2a-ORF2 з PCV2, SEQ ID NO: 3) і фрагмента 2a-F2 + 2a-F3 (нуклеотиди 235-699 (без стоп-кодона) на 5’-кінці 2aORF2 з PCV2, SEQ ID NO: 11) показував, що кількості аргінінів в амінокислотній послідовності (SEQ ID NO: 4) фрагмента 2a-F1 в два рази або більше перевищували кількості аргінінів в амінокислотній послідовності (SEQ ID NO: 12) фрагмента 2a-F2 + 2a-F3 (як показано в таблиці 2). Аналіз додавання або делеції аргініну показував, що делеція надлишкового аргініну могла підвищувати експресію білка з ORF2 з PCV2. Таблиця 2. Аналіз кількостей аргінінів в амінокислотній послідовності 2a-ORF2 з PCV2 Кодон AGA AGG CGA CGC CGT Загалом 2a-F1 Амінокислота Аргінін Аргінін Аргінін Аргінін Аргінін Кількість 5 3 1 10 2 21 Кодон AGA AGG CGC CGG CGT Загалом 2a-F2 + 2a-F3 Амінокислота Аргінін Аргінін Аргінін Аргінін Аргінін Кількість 3 3 1 1 1 9 10 15 20 25 30 35 40 45 50 Приклад 2. Аналіз імуногенності антигенних пептидів субодиничної вакцини проти PCV2 1. Імунізація щурів Отримували субодиничну вакцину проти PCV2 з рекомбінантними білками 2a-F2, 2a-F3, 2aF4 та 2a-F5, отриманими в прикладі 1, відповідно, і повним ад’ювантом Фрейнда. Щурів вакцинували субодиничною вакциною проти PCV2 для аналізу імуногенності антигенних пептидів субодиничної вакцини проти PCV2. Здорових щурів, які не мали конкретних патогенів, віком від п’яти до шести тижнів рандомно розділяли на 5 груп по 3 щури в кожній. Твердофазний імуноферментний аналіз (ELISA) показував, що всі 15 щурів були негативними за антитілами до PCV2. Кожному щуру з 4 груп вакцинації (групи 1-4) вводили ін’єкцією підшкірно 200 мкг рекомбінантного білка, і загальний об’єм кожної ін’єкції становив 300 мкл при співвідношенні 1:1 (об./об.) білка до ад’ювантної композиції. Щурам з групи 5 вводили ін’єкцією 300 мкл PBS і використовували в якості негативного контролю. Через два тижні після первинної імунізації (p.i.) щурам з 4 груп вакцинації повторно уводили таку ж дозу 4 різних рекомбінантних білків, відповідно. Зразки сироватки збирали через 0, 2, 4 і 8 тижнів після первинної імунізації. Всі зразки сироватки тестували за допомогою ELISA для вимірювання титру антитіл до PCV2. 2. Визначення антитіл до PCV2 за допомогою ELISA В даному прикладі використовували планшети з дев’яноста шістьма лунками, що містять патогенний антиген PCV2 (300 нг/лунка), в якості планшетів для ELISA. Планшети для ELISA промивали 3 рази 50 ммоль/л PBS (pH 7,2), що містить 500 мкл/л Твін-20 (тобто PBST) протягом від 3 до 5 хвилин кожен раз. Для блокування планшетів для ELISA додавали 200 мкл 0,15% блокувального розчину BSA в кожну лунку планшетів для ELISA, і потім планшети для ELISA інкубували протягом 2 годин при 37°C. Після чого планшети для ELISA промивали PBS. Зразки сироватки щурів розводили в п’ятдесят разів (1:50) за допомогою PBS, а потім послідовно розводили в два рази. Кожен зразок мав 8 повторів. Розведені зразки сироватки додавали в лунки планшетів (100 мкл/лунка) для ELISA і планшети інкубували протягом 1 години при 37°C. Після інкубування планшети промивали PBS. Потім в лунки додавали антитіло до IgG щура, кон’юговане з лужною фосфатазою (AP). Після інкубування протягом 1 години при 37°C планшети промивали PBS. Для візуалізації результатів в лунки додавали паранітрофенілфосфат (pNPP). Після інкубації реакцію зупиняли шляхом додавання 1M NaOH. Оптичну щільність кожної лунки зчитували за допомогою пристрою для зчитування оптичної щільності при 405 нм. Кожен зразок аналізували у двох повторностях і значення O.D. від дублів усереднювали. Результати ELISA показані на фіг. 5. Всі рекомбінантні білки 2a-F2, 2a-F3, 2a-F4 та 2a-F5, отримані в прикладі 1, були здатні індукувати сироваткові антитіла до PCV2 у тестованих тварин. Серед рекомбінантних білків рекомбінантний білок 2a-F2 (SEQ ID NO: 6) індукував найбільш високий рівень сироваткового антитіла до PCV2. Для статистичного аналізу всі групи порівнювали з негативним контролем PBS у різних моментах забору зразка, і виявляли значущі відмінності між негативним контролем і кожною групою вакцинації (p
ДивитисяДодаткова інформація
Назва патенту англійськоюPorcine circovirus type-2 subunit vaccine
Автори англійськоюKuo, Tsun-Yung, Chen, Hsu-Chung Gabriel, Yang, Shu-Hsiang, Chen, Yu-San
Автори російськоюКо Тсунь-Юньг, Чэнь Хсу-Чун Габриэль, Ян Шу-Хсиан, Чэнь Юй-Сань
МПК / Мітки
МПК: A61K 39/12, C07K 14/01
Мітки: pcv2, типу, цирковірусу, свиней, композиція, імуногенна
Код посилання
<a href="https://ua.patents.su/42-113192-imunogenna-kompoziciya-proti-cirkovirusu-svinejj-tipu-2-pcv2.html" target="_blank" rel="follow" title="База патентів України">Імуногенна композиція проти цирковірусу свиней типу 2 (pcv2)</a>
Одноразова доза полівалентної комбінованої вакцини, яка включає антиген цирковірусу свиней типу 2

Номер патенту: 99708
Опубліковано: 25.09.2012
Автори: Шеффер Меррілл, Хейз Філліп, Нітцель Грег, Руф Майк Б., Айхмейер Марк
МПК: A61K 39/04, A61K 39/12, C12N 15/866, A61K 39/295
Мітки: типу, вакцини, цирковірусу, одноразова, доза, комбінованої, антиген, полівалентної, свиней, включає, яка
Формула / Реферат:
1. Одноразова доза полівалентної комбінованої вакцини, яка включає антиген цирковірусу свиней типу 2, ефективний для зниження частоти зустрічальності або зниження тяжкості прояву інфекцій цирковірусу свиней типу 2, і принаймні один додатковий компонент імуногенної дії, ефективний відносно іншого організму, що викликає захворювання у свиней, у якій антиген цирковірусу свиней типу 2 є рекомбінантно експресованим ORF2 білком цирковірусу свиней...
Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, вакцина проти багатосистемного синдрому виснаження свиней (бсвс) та набір для вакцинації

Номер патенту: 72891
Опубліковано: 16.05.2005
Автори: Одонне Жан-Крістоф Франсіс, Краковка Джордж Стівен, Аллан Гордон Мур, Мак-Нейлі Френсіс, Міхен Брайан Мартін, Елліс Джон Алберт
МПК: A61P 43/00, A61K 39/295, C07K 14/01, A61P 37/04, A61P 31/22, C12N 7/00, A61P 31/12
Мітки: вакцинації, викликає, виснаження, імунну, препарат, цирковірусу, набір, багатосистемного, синдрому, свиней, парвовірусу, відповідь, імуногенний, бсвс, вакцина
Формула / Реферат:
1. Імуногенний препарат, що викликає імунну відповідь проти парвовірусу та цирковірусу свиней, що включає щонайменше один антиген цирковірусу свиней та щонайменше один антиген парвовірусу свиней у розчиннику або наповнювачі, прийнятному з ветеринарної точки зору.2. Імуногенний препарат за п.1, що включає антиген цирковірусу свиней типу II.3. Імуногенний препарат за п.2, у якому антиген цирковірусу свиней типу II вибраний з...
Ізольований цирковірус свиней, композиції та способи для імунізації свиней проти цирковірусу свиней

Номер патенту: 99495
Опубліковано: 27.08.2012
Автор: Ву Стівен Циту
МПК: C12N 15/00, C07K 14/01, C12Q 1/68, A61K 39/12, C12N 7/00
Мітки: цирковірус, ізольований, цирковірусу, свиней, способи, імунізації, композиції
Формула / Реферат:
1. Ізольований цирковірус свиней, геномна послідовність якого відповідає нуклеотидній послідовності будь-якої з послідовностей SEQ ID NО: 1 або 2.2. Ізольований цирковірус свиней за п. 1, який є цирковірусом свиней типу 2В.3. Ізольований цирковірус свиней за п. 2, що включає ORF2 білка, в якому амінокислотна послідовність ORF2 білка є будь-якою з послідовностей SEQ ID NO: 3 або 4.4. Ізольований цирковірус свиней за п. 1,...
Цирковірус свиней 2 типу (цвс2), імуногенна композиція, що містить його, набір для аналізу та їх застосування

Номер патенту: 110632
Опубліковано: 25.01.2016
Автори: Ко Тсунь-Юньг, У Чунг-Чін, Чень Хсу Чунг Габріель, Чень Хань-Тін
МПК: C12Q 1/70, C12N 15/34, G01N 33/53, G01N 33/569, C07K 14/01, A61K 39/12, C12N 7/01, C07K 16/08, A01N 63/00, C12R 1/93
Мітки: цвс2, свиней, імуногенна, композиція, цирковірус, аналізу, його, містить, застосування, типу, набір
Формула / Реферат:
1. Цирковірус свиней 2 типу (ЦВС2), який містить геномну послідовність SEQ ID NО:1.2. Цирковірус свиней 2 типу (ЦВС2) за п. 1, який був депонований у Китайському центрі колекції типових культур (ССТСС) під номером доступу V201117.3. Цирковірус свиней 2 типу (ЦВС2) за п. 1, де ЦВС2 здатний викликати цитопатичний ефект (СРЕ) у клітинній лінії РК-15 або її похідних клітинних лініях.4. Імуногенна композиція, яка містить...
Зменшення супутніх інфекцій у свиней за допомогою антигену pcv2

Номер патенту: 104129
Опубліковано: 10.01.2014
Автори: Фахінгер Віккі, Ельберс Кнут, Кіксмьоллер Маріон
МПК: A61P 31/12, A61K 39/12, A61P 31/06, A61P 31/10
Мітки: зменшення, свиней, супутніх, інфекцій, антигену, допомогою
Формула / Реферат:
1. Спосіб зниження відсотка супутніх інфекцій у свиней або у череді свиней, що викликаються одним або декількома патогенами, відмінними від PCV2, який полягає в тому, що вводять свині(ям) в ефективній кількості білок ORF-2 PCV2 або імуногенну композицію, яка містить білок ORF-2 PCV2.2. Спосіб за п. 1, який відрізняється тим, що супутня інфекція викликається вірусним, бактеріальним і/або грибним патогеном.3. Спосіб за п. 2, який...