Спосіб визначення рухомості спермієв сільськогосподарських тварин
Номер патенту: 21719
Опубліковано: 20.01.1998
Автори: Мегель Юрій Євгеньович, Зубец Михайло Васильович, Путятін Валерій Петрович




Формула / Реферат
Способ определения подвижности спермиев сельскохозяйственных животных, включающий освещение раствора спермы в проходящем свете, получение оптической информации и ее обработку, включающую кодирование в проекциях, отличающийся тем, что кодирование производят на четыре проекции всей массы спермиев с определением кода изображения каждого спермия, а определение пути и скорости по каждому спермию осуществляют путем вычитания соответствующих кодов проекций из первого одновременно для всей массы спермиев в реальном масштабе времени.
Текст
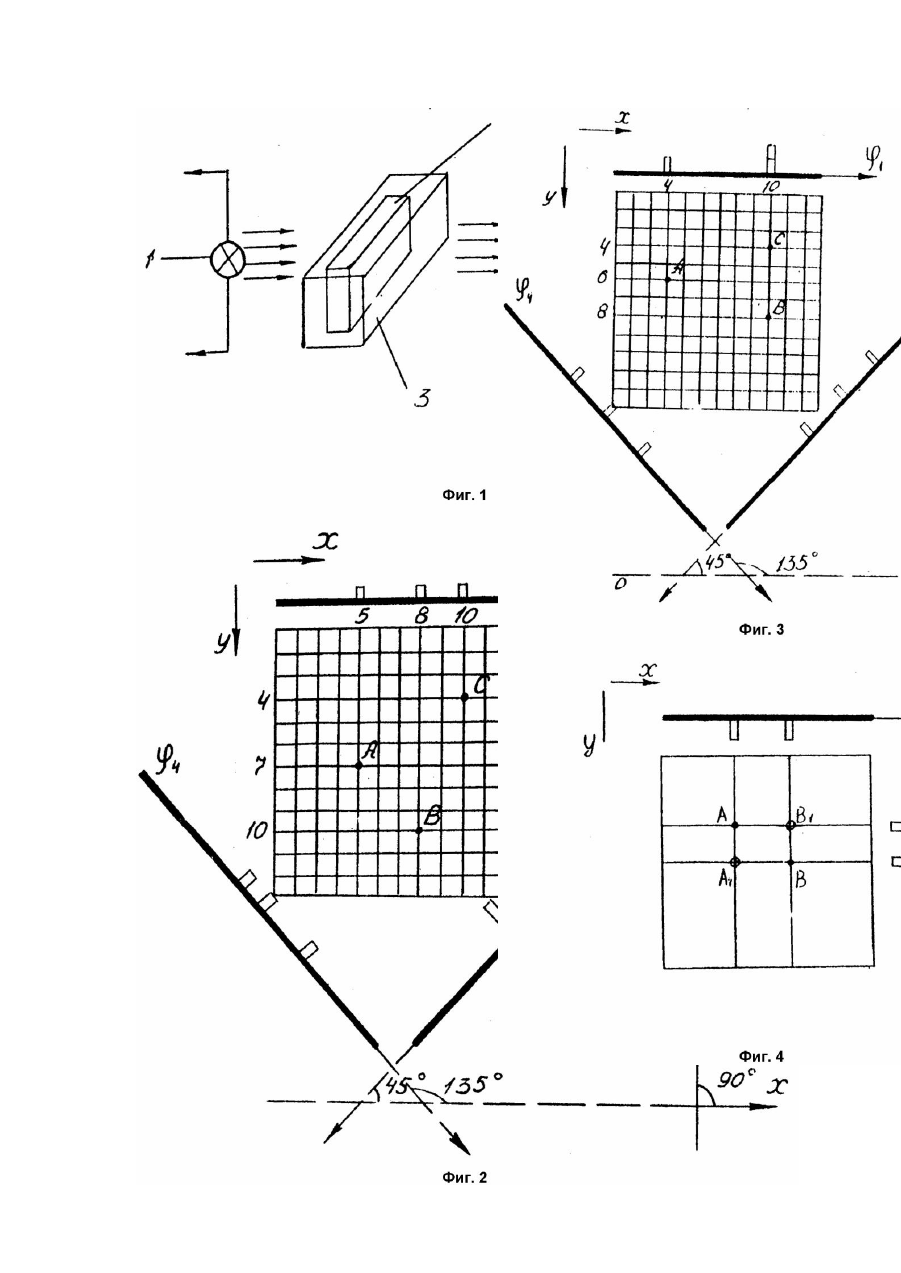
Изобретение относится к сельскому хозяйству, в частности к искусственному осеменению животных, и может быть использовано для оценки качества спермы по подвижности (активности) спермиев. Важнейшим этапом технологии искусственного осеменения животных является объективное и быстрое определение качества биоматериала, полученного от производителя (до замораживания и после размораживания перед искусственным осеменением). Одним из основных качественных показателей спермы является количество спермиев с активным прямолинейным движением. Известен способ определения качества спермиев, включающий смешивайте спермы с питательной средой, содержащей два красителя тиазиновый красный и этидиум бромид. При повреждении мембраны акросомы тиазиновый красный проникает в ак-росому и вызывает ее свечение, а этидиум бромид, проникает через поврежденную ци-топлазматическую мембрану спермия, вызывает свечение его головки. О качестве спермы судят по интенсивности их свечения под люминесцентным микроскопом (А.с. СССР №1329780, кл. A61D7/02, 1987). Недостатком этого способа является большая погрешность и длительность процесса оценки. Это связано с субъективизмом оператора, его утомляемостью и зрительной способностью. Так под микроскопом нормальные спермин будут выглядеть более светлыми на темно-синем фоне, а при повреждении акросомы - светло-салатными, при повреждении цитоплазматической мембраны ярко-малиновыми. Длительность подсчета спермиев с различными оттенками приводят к повышенной утомляемости оператора и снижению его зрительной способности и, естественно, к ошибкам. Известен способ определения подвижности сперматозоидов в эякулятах самцов путем воздействия на них постоянным электрическим током. Для этого в емкость с проводящей электрический ток средой помещают сосуд с раствором спермы, на который воздействуют постоянным током. Сперматозоиды начинают двигаться от отрицательного полюса к положительному и через горизонтальные отверстия в стенках внутреннего сосуда проходят в среду наружной емкости. При этом мертвые сперматозоиды и сперматозоиды с манежным движением не выйдут из внутреннего сосуда. Подсчитав количество сперматозоидов в эякуляте и в среде наружной емкости определяют процент подвижных сперматозоидов в сперме, т.е. ее качество (А.с. СССР №1386186, кл. A61D7/02, 1988). Недостатками известного способа являются длительность (подсчет сперматозоидов под микроскопом) и большая погрешность. На подвижность сперматозоидов, кроме воздействия постоянного тока, сказывается характер температурного поля, наличие градиента температуры в растворе и стенках сосудов, осмотическое давление. Указанные факторы влияют на точность оценки известного способа. Наиболее близким по технической сущности к заявляемому является способ, реализованный в известном устройстве для считывания информации о подвижных микрообъектах в жидком растворе (А.с. СССР №903918, кл. G06K9/46, 1982, прототип). Способ включает освещение раствора спермы в проходящем свете, получение оптической информации и ее обработку. Оптическую информацию в каждый заданный момент времени T фиксируют на видеомагнитофонной ленте. В полученной записи выделяют наиболее характерные кадры. После этого в каждый заданный момент времени T каждому спермию определяют коды координат "x" и "y". При последующей обработке на ЭВМ координат "x" и "y" определяют подвижность спермиев в нереальном масштабе времени (по прошествии события). По сравнению с аналогами данный способ значительно точнее, требует меньшего времени для оценки и не зависит от субъективных качеств оператора. Если необходимо контролировать подвижность большого количества спермиев, то реализация способа вызовет затруднения из-за дефицита памяти и быстродействия устройства. Кроме того, известный способ позволяет вести обработку кодов координат отдельно каждого спермия и при большом их количестве требует больших затрат времени и памяти ЭВМ, что не всегда приемлемо, например, при анализе подвижности спермиев перед искусственным осеменением, когда требуется быстрый анализ в реальном масштабе времени. Задачей изобретения является разработка такого способа определения подвижности спермиев сельскохозяйственных животных, в котором путем введения операций кодирования на четыре проекции всей массы спермиев с определением кода изображения каждого спермия повышается быстродействие способа. Поставленная задача достигается тем, что в способе определения подвижности спермиев сельскохозяйственныхживотных, включающим освещение раствора спермы в проходящем свете, получение оптической информации и ее обработку, включающую кодирование в проекциях, согласно изобретению кодирование производят на четыре проекции всей массы спермиев с определением кода изображения каждого спермия, а определение пути и скорости по каждому спермию осуществляют путем вычитания соответствующих кодов координат из первого одновременно для всей массы спермиев в реальном масштабе времени. Благодаря проецированию местонахождения каждого спермия в заданные моменты времени на четыре проекции повышается точность способа, т.к. любое перемещение спермия будет отражено на проекциях в кодах координат изображений и поступит для дальнейшего анализа. Предлагаемый способ предусматривает обработку информации всего изображения сразу в реальном масштабе времени, что значительно ускоряет процесс определения подвижности спермиев по сравнению с прототипом, где обработка кодов координат каждого микрообъекта осуществляется последовательно и в нереальном масштабе времени. При поиске по патентной и научно-технической литературе не выявлены объекты с признаками, сходными с отличительными признаками заявляемого технического решения, на основании чего можно сделать вывод о соответствии его критерию "существенные отличия". Предлагаемый способ реализуют следующим образом. Концентрированную сперму разбавляют до концентрации, в которой количество спермиев в растворе является статистически значимо. Разбавленную сперму помещают в канюлю с размерами, не затрудняющими движение спермиев. Температура канюли должна соответствова ть температуре, характерной для жизнеспособности спермиев (38 - 40°C). Канюлю с раствором спермы освещают в проходящем свете. Оптическая информация о спермиях в канюле регистрируется, например, микроскопом, обрабатывается, например, преобразуется в электрические сигналы о каждом спермие, а затем в бинарный двоичный код. Потом это бинарное изображение одновременно кодируется на четыре проекции в заданные моменты времени с определением кода изображения каждого спермия. Затем осуществляют вычитание кодов проекций последующих изображений во времени из первого по каждому спермию одновременно для всего массива информации. Все выше перечисленные операции производят в реальном масштабе времени. В результате последней операции получают общую оценку подвижности каждого спермия одновременно для всей массы исследуемого объема, т.е. путь, пройденный каждым спермием и его скорость. Устройство, реализующее способ, иллюстрируется чертежами. На фиг.1 приведена блок-схема устройства, реализующая операции способа; на фиг.2 положение спермиев в начальный момент наблюдения T1 с соответствующим кодированием оптической обработанной информации о них на четыре оси координат; на фиг.3 - то же, но в следующий момент времени T2; на фиг.4 иллюстрируется подтверждение необходимости введения четырех проекций, поскольку двух (проекций только на оси и мало. Это связано с тем, что перемещение, например двух спермиев и (фиг.4) соответственно в точки и не изменяет кодирования на оси и хотя местоположение спермиев изменилось. Устройство содержит источник 1 света, освещающий канюлю 2 с разбавленной спермой самца. Канюля 2 может быть помещена в термостат 3 с прозрачными стенками и теплоносителями. В зависимости от температуры в помещении и учитывая незначительное время исследования каждой канюли (3 - 5сек), термостат 3 может отсутствовать. За канюлей 2, со стороны, противоположной источнику 1, установлен микроскоп 4, совмещенный с телевизионным датчиком 5, содержащим матричный фотоприемник на приборе с зарядовой связью (ПЗС), например, КТМ-1. Датчик 5 своим выходом связан со входом блока 6 кодирования с компаратором. Выход блока 6 соединен с вычитающим узлом 7 с памятью, который связан с анализатором 8, выполненным в виде арифметического устройства с монитором. Способ осуществляется следующим образом При включении источника 1 света канюля 2 освещается. Предположим, что в поле зрения микроскопа 4 попали спермин и Степень увеличения изображения микроскопа 4 выбирается такой, чтобы минимальный из характерных геометрических размеров спермиев совпадал с несколькими элементами матричного фотоприемника на ПЗС в датчике 5. Кроме того, частота съема информации выбирается с учетом того, чтобы время между двумя кадрами обеспесчивало перемещение самых быстрых спермиев на 1 - 2 элемента матричного фотоприемника. Это позволит повысить точность считывания и кодирования информации. Таким образом информация о местонахождении спермиев и в канюле 2 для каждого момента времени порциями (кадрами) с помощью объектива микроскопа 4 проецируется на фоточувствительные секции ПЗС датчика 5, где преобразуется в видеоэлектрические сигнал. Затем эти сигналы поступают на компаратор блока 6, который срабатывает при превышении амплитуды видеосигнала заданного значения, осуществляя таким образом преобразование видеоэлектрических импульсов в бинарный двоичный код. После этого в блоке 6 это бинарное изображение кодируется на четыре проекции. Для идентификации спермиев кодирования их местоположения в плоскостях изображения воспользуемся преобразованием Радона (Хелгасон С. Преобразование Радона. - М.: Мир, 1983). Для такого кодирования информации, находящейся в каждом из кадров (фиг.2) вводится четыре прямые под углами соответственно 0°, 90°, 45 и 135° к оси В начальный момент спермии и (фиг.2) имеют в декартовой системе следующие значения координат где Телекамера (блок 5) раскладывает изображение на прямоугольную сетку-ПЗСматрицу, состоящую из строк и столбцов. Блок 6 фиксирует положение спермиев в момент времени и далее осуществляет кодирование местоположения спермиев на четыре проекции (прямые) (фиг.2). Тогда коды проекций изображений спермиев на прямые определяются следующим образом. Для случая прямой под углом 45° (табл.3) номер разряда (значение координаты), в котором появится "1" (сигнал) в результате кодирования на прямую местоположения спермия определяется как где для рассматриваемого примера (размер кадра 13 ´ 13); - принимает значения т.е. значения координат спермиев на оси соответственно принимает значения Для случая прямой под углом 135° (табл.4) номер разряда (значение координаты), в котором появится "1" (сигнал) в результате кодирования на прямую местоположения спермия определяется как принимает где как и в предыдущем случае значения а соответственно Следует отметить, что число разрядов на прямых и меньше, чем число разрядов на прямых и Это связано с тем, что для первой группы кодов на прямых и проецирование осуществляется на отрезки, равные длине стороны квадрата (кадра), а для второй группы кодов на прямых и - на отрезки, равные длине диагонали квадрата (кадра). Таким образом, в начальный момент времени спермии и (фиг.2) имеют следующие коды проекций изображения спермиев на прямые Значения кодов проекций спермиев в момент времени изображений т е. передают в память блоки 7. Вполне очевидно, что изменение местоположения спермиев изменяет значение кодов - проекций изображений спермиев. Анализ во времени изменения соответствующи х кодов дает возможность оценить подвижность спермиев В момент времени коды проекций (фиг.3) будут иметь значения (таблицы не приведены, т.к. построение их аналогично табл.1 - 4): По результатам анализа кодов и в блоке 8 определяют число подвижных и неподвижных спермиев, а также путь (число дискретов ПЗС-матрицы телекамеры в каждом из направлений) пройденный тем или иным спермием за исследуемый промежуток времени, что дает возможность определить скорость перемещения спермиев. Например, из анализа результирующего кода видно, что спермий за время сместился на один дискрет "влево" по оси (перемещение "1" с пятого разряда кода четвертый разряд кода появление "-1" в в Это обусловило четвертом разряде Эти коды в момент времени также поступают в блок 7, где производят поразрядное вычитание соответствующи х одноименных кодов проекций изображений спермиев в моменты результирующего кода Спермий за то же время переместился на два дискрета "вправо" (перемещение "1" с восьмого времени разряда кода Спермий и в десятый разряд кода остался неподвижным, т.к. в десятом разряде кода появилась "2", свидетельствующая о том, что спермий и лежат на одной прямой, перпендикулярной к оси Это обусловило появление "-1" в десятом разряде результирующего кода Аналогично проводится где - результат разности кодов изображений. Подставив значения соответствующи х кодов и произведя вычитания получим: анализ кодов что дает возможность получить информацию об изменении положения спермиев в други х анализируемых направлениях. Моменты траектории времени исследования выбирает оператор. При выделении каждого спермия в блоке 8 анализируют промежутков результаты двух времени, соседних т.е. На мониторе блока 8 оператор видит траектории подвижных спермиев. Зная пройденный путь каждого спермия (число дискретов ПЗС-матрицы телекамеры) и время, за которое произошло смещение, определяют скорость движения спермиев. Все измерения и преобразования происходят в момент наблюдения за спермиями, т.е. в реальном масштабе времени, что значительно повышает быстродействие предлагаемого способа по сравнению с прототипом. Кроме того, в прототипе производят обработку координат последовательно для каждого отдельного элемента объекта (например, спермия). В предлагаемом способе обрабатывают все изображения сразу, т.е. все спермиев. Таким образом предлагаемый способ увеличивает за счет этого быстродействие в раз, где - число спермиев. Особенно ощутимы преимущества заявляемого способа по сравнению с прототипом при анализе большого количества спермиев, например в условиях промышленной технологии заготовки спермодоз (банка спермодоз), до и после их замораживания. Разрешающая способность аппаратурной реализации зависит от числа ячеек ПЗС-матрицы типа КТМ-1 эти х ячеек 260 ´ 380шт. При необходимости можно использовать ПЗС-матрицы на 1024 ´ 1024 ячеек и более.
ДивитисяДодаткова інформація
Автори англійськоюZubets Mykhailo Vasyliovych, Mehel Yurii Yevhenovych, Putiatin Valerii Petrovych
Автори російськоюЗубец Михаил Васильевич, Мегель Юрий Евгеньевич, Путятин Валерий Петрович
МПК / Мітки
МПК: A61D 19/02
Мітки: сільськогосподарських, визначення, спосіб, спермієв, рухомості, тварин
Код посилання
<a href="https://ua.patents.su/5-21719-sposib-viznachennya-rukhomosti-spermiehv-silskogospodarskikh-tvarin.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення рухомості спермієв сільськогосподарських тварин</a>
Спосіб визначення кровності сільськогосподарських тварин

Номер патенту: 13580
Опубліковано: 25.04.1997
Автори: Романов Леонід Максимович, Цілуйко Григорій Олександрович
МПК: A01K 67/02
Мітки: тварин, сільськогосподарських, визначення, кровності, спосіб
Формула / Реферат:
Способ определения кровности сельскохозяйственных животных, включающий вычисление долей крови, отличающийся тем, что, с целью повышения надежности определения кровности помесных сельскохозяйственных животных и обеспечения возможности определения кровности во всех типах скрещивания, вычисление долей крови осуществляется по процентному соотношению специфических маркерних аллелей систем групп крови, характеризующих каждую исходную...
Спосіб визначення молочної кислоти в рослинних кормах для сільськогосподарських тварин

Номер патенту: 12748
Опубліковано: 28.02.1997
Автори: Шевцова Галина Миколаївна, Малінін Олег Олексійович, Ткаченко Андрій Миколайович
МПК: A23K 1/00
Мітки: визначення, молочної, кормах, спосіб, сільськогосподарських, рослинних, тварин, кислоти
Текст:
...К 4 мл 0,2% раствора молочной кислоты в этиловом спирте добавляли 0,6 мл концентрированной серной кислоты (уд.вес. 1,84) и нагревали смесь на водяной бане 70°С 10 мин. Избыток серной кислоты не оправдано ухудшает условия работы и воспроизводимость. П р и м е р 2. Кб мл 0,2% раствора молочной кислоты в этиловом спирте добавляли 0,4 мл концентрированной серной кислоты (уд.вес.1.84) и нагревали смесь на водяной бане при 70°С 10 мин. Уменьшение...
Спосіб одержання концентрату лактатів для годування сільськогосподарських тварин

Номер патенту: 3289
Опубліковано: 27.12.1994
Автор: Лінд Робєрт Мейнгардтовіч
МПК: A23K 1/08
Мітки: тварин, спосіб, лактатів, одержання, годування, сільськогосподарських, концентрату
Формула / Реферат:
Способ получения концентрата лактатов для кормления сельскохозяйственных животных, предусматривающий термообработку творожной сыворотки и нейтрализацию ее щелочью, отличающийся тем, что в качестве щелочи используют 2М раствор едкого натра, причем термообработку ведут при температуре 32-400С, а нейтрализацию - путем введения едкого натра через равные промежутки времени 18-20 раз в течение 4,5-6,0 часов при рН 6,6-6,8.
Спосіб кріоконсервації сперми сільськогосподарських тварин і пристрій для його здійснення

Номер патенту: 13371
Опубліковано: 28.02.1997
Автори: Зорін Олександр Володимирович, Зубец Михайло Васильович, Зубець Микола Михайлович, Зорін Володимир Михайлович
МПК: A61D 19/00
Мітки: сперми, здійснення, сільськогосподарських, кріоконсервації, тварин, спосіб, пристрій
Формула / Реферат:
(57) 1. Способ криоконсервации спермы сельскохозяйственных животных, включающий загрузку спермодоз в тубы, эквилибрацию их путем выдерживания при (+2) -(+5)°С и последующее замораживание с помощью жидкого азота, отличающийся тем, что эквилибрацию и замораживание туб со спермодозами осуществляют в герметичной камере, причем эквилибрацию при (+2)-(+5)°С проводят в течение 1,5-2 ч. а замораживание с помощью жидкого азота ведут путем подачи...
Спосіб підвищення резистентності молодняка сільськогосподарських тварин

Номер патенту: 14118
Опубліковано: 25.04.1997
Автори: Карвацька Гера Петрівна, Нікітенко Анатолій Мефодійович, Половинка Микола Михайлович, Заіка Леонід Андрійович, Паска Микола Миколайович
МПК: A61K 38/24
Мітки: спосіб, молодняка, тварин, резистентності, сільськогосподарських, підвищення
Формула / Реферат:
Способ повышения резистентности молодняка сельскохозяйственных животных, включающий введение в организм животного гомогената тимуса, отличающийся тем, что, с целью повышения эффективности способа, гомогенат тимуса вводят совместно с ферроглюкином при их массовом соотношении 1:0,5-4,0 в дозе 0,3 мл на 1 кг живой массы тела подкожно.
Попередній патент: Спосіб одержання залізо-вуглецевого розплаву
Наступний патент: Каркас, що трансформується, для тентового покриття залу
Випадковий патент: Спосіб місцевого лікування гнійних ран