Спосіб вакцинації тварин від інфекційних захворювань
Номер патенту: 50800
Опубліковано: 15.11.2002
Автори: Величко Сергій Володимирович, Науменко Володимир Васильович, Кладницька Лариса Володимирівна



Формула / Реферат
Спосіб вакцинації тварин від інфекційних захворювань, що включає введення антигенного подразника, який відрізняється тим, що антигенний подразник уводять інтракутанно через біологічно активні точки J(XIV) 17 (тань-чжунь), Т(ХПІ) 14 (да-чжуй) у дозі 0,2 мл у 8 - 10-тижневому віці.
Текст
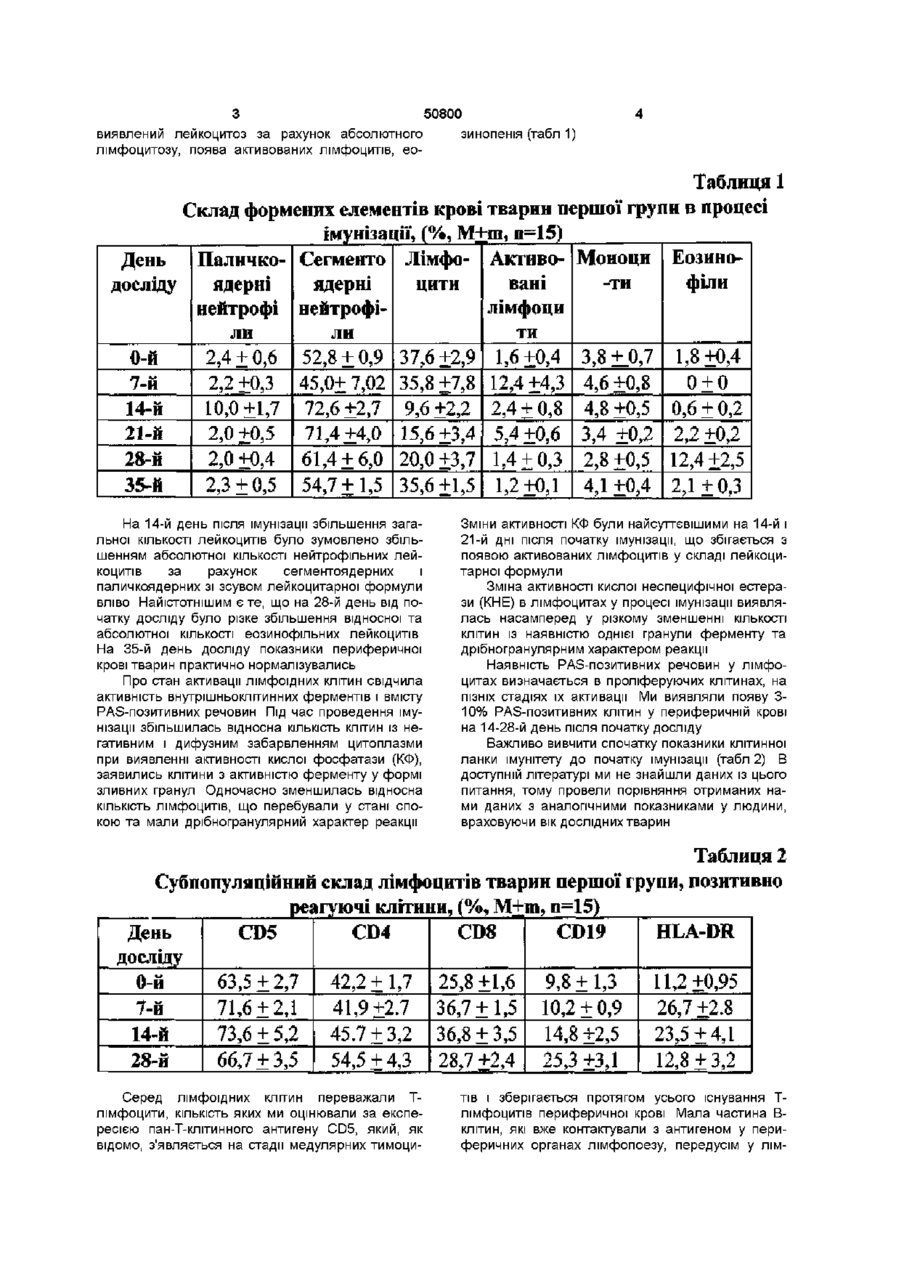
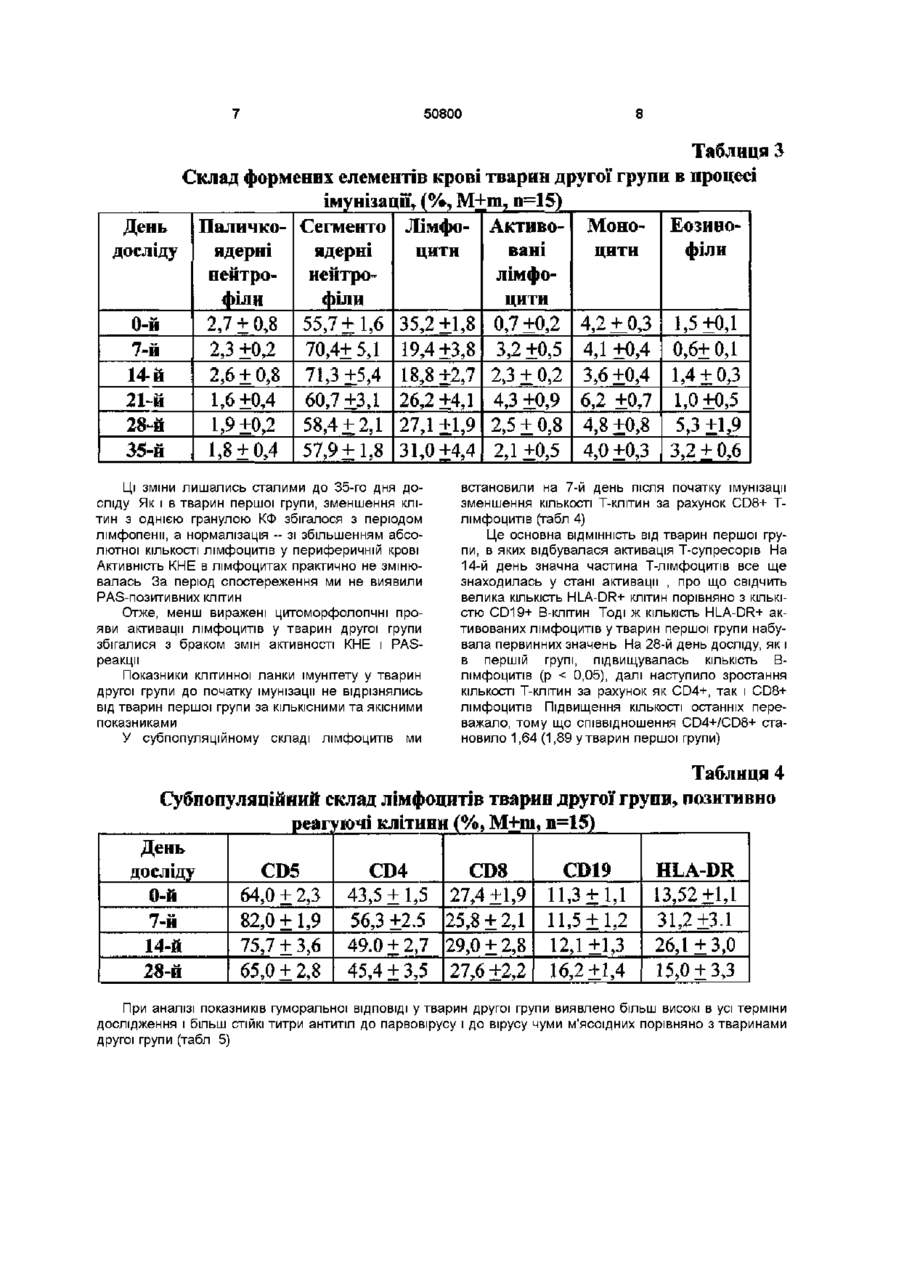
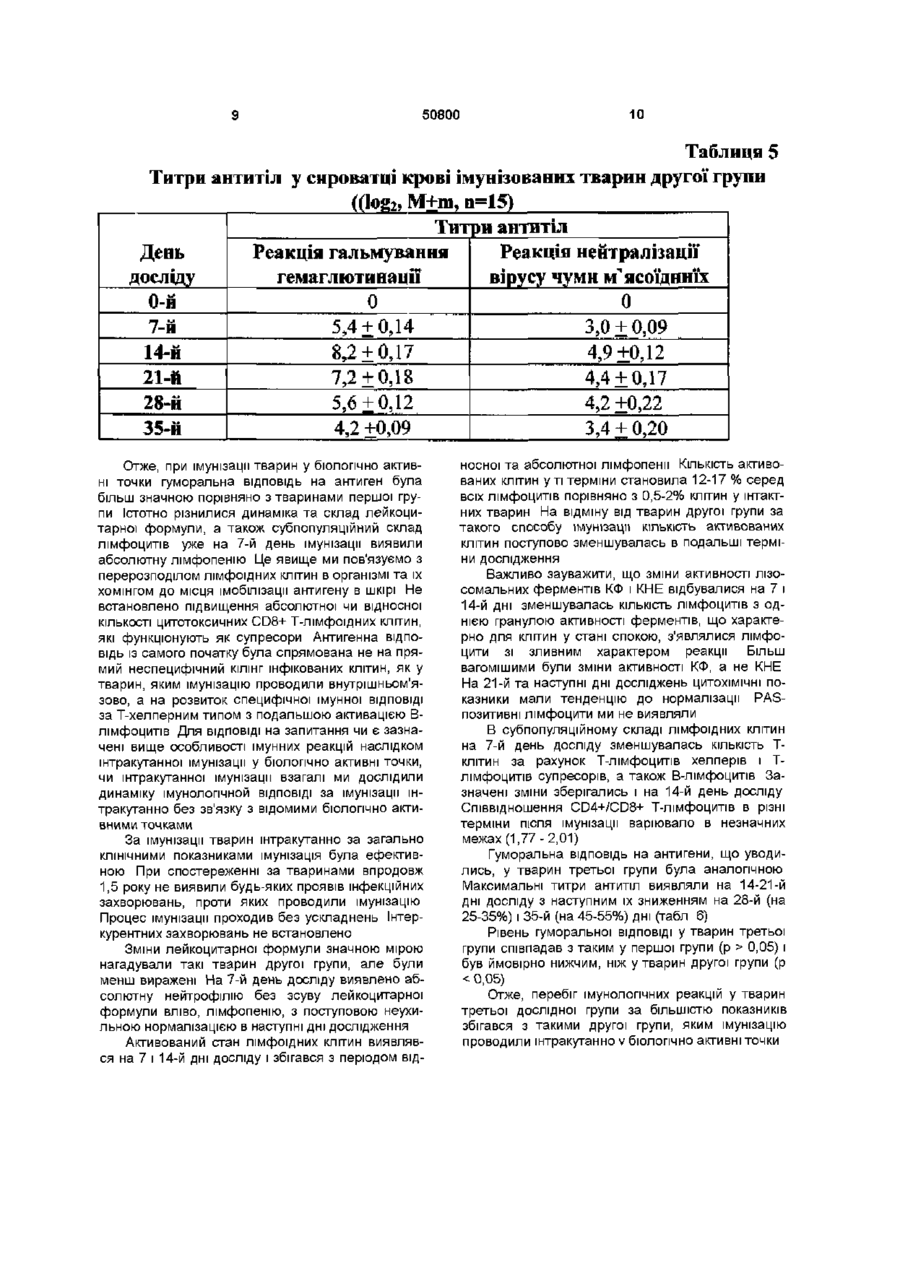
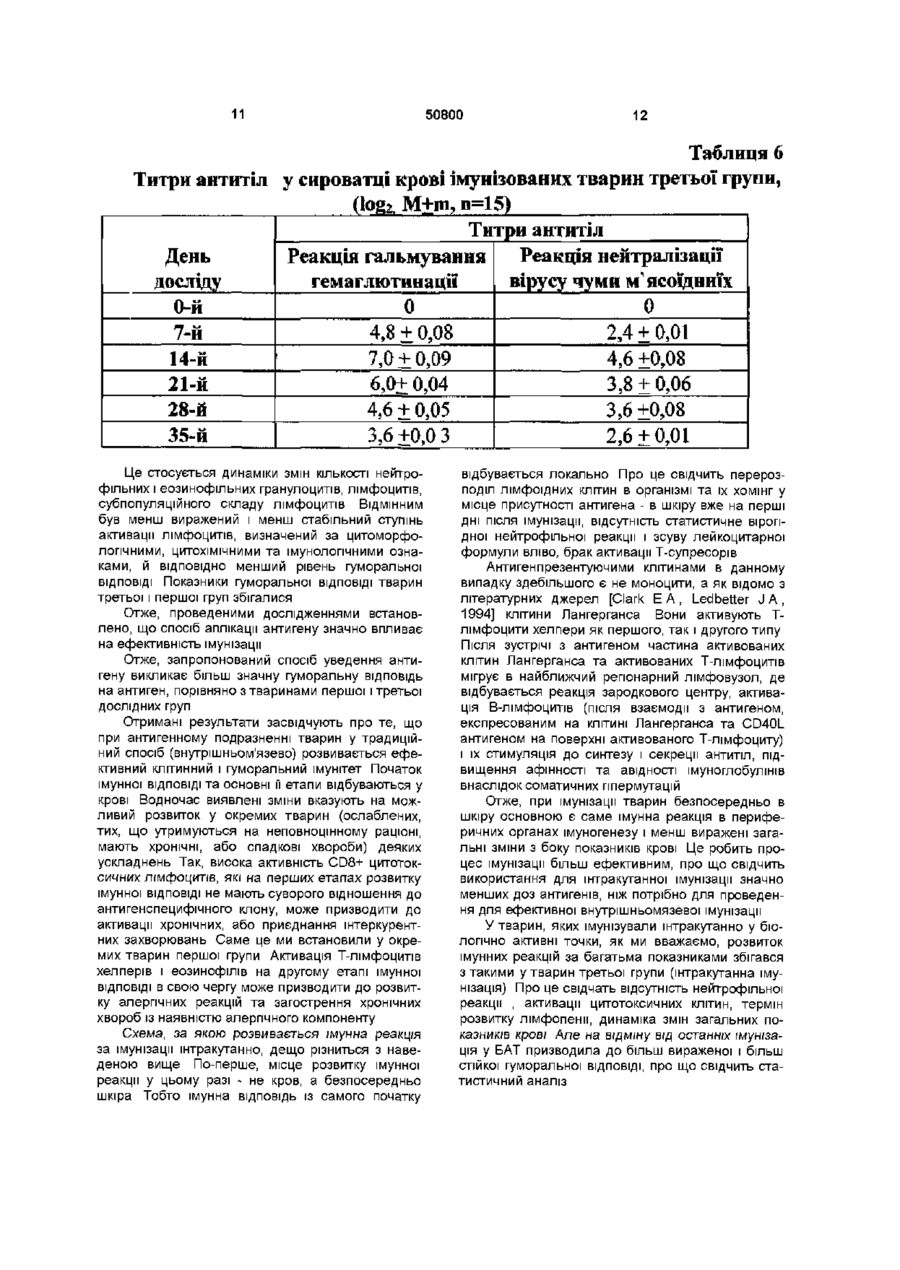
Спосіб вакцинації тварин від інфекційних захворювань, що включає введення антигенного подразника, який відрізняється тим, що антигенний подразник уводять штракутанно через біологічно активні точки J(XIV) 17 (тань-чжунь), Т(ХПІ) 14 (да-чжуй) у дозі 0,2мл у 8 -10-тижневому Винахід відноситься до галузі ветеринарної медицини, зокрема до способів вакцинації тварин Відомий спосіб вакцинації тварин - внутрішньом'язовий Він обумовлює використання вакцинного препарату в дозі 1мл як для дорослих тварин, так і для молодих, внаслідок чого в організм потрапляє певна доза вакцинного препарату [Медуніцин Н В ,1995] Недоліком внутрішньом'язового способу - прототипу є те, що він може призводити до розвитку у окремих тварин деяких ускладнень (активації хронічних або приєднання штеркурентних захворювань, розвитку алергічних реакцій та загострення хронічних хвороб з наявністю алергічного компоненту) Винаходом ставиться завдання щодо стимулювання функціонального стану імунної системи та оптимізацм фізіологічних механізмів імунної ВІДПОВІДІ організму на антигенне подразнення Поставлене винаходом завдання досягається способом вакцинації тварин, зокрема від інфекційних захворювань, що включає введення антигенного подразника інтракутанно через біологічно активні точки J(XIV) 17 (тань-чжунь), Т(ХІІІ) 14 (дачжуй) у дозі 0,2мл у 8-10-тижневому ВІЦІ Таке введення антигену тваринам призводить до більш ефективної та оптимальної ВІДПОВІДІ організму на антигенне подразнення Приклад Дослідним собакам (друга дослідна група п = 15) наносили антигенне подразнення через біологічно активні точки Вакцину "Альфавак 1" (проти чуми м'ясоїдних) уводили інтракутанно у дозі 0,2мл в біологічно активну точку J(XIV) 17 (тань-чжунь) в ДІЛЯНЦІ четвертого міжребер'я на передній серединній лінії Вакцину "Альфавак 3" у дозі 0,2мл уводили інтракутанно у точку Т(ХІІІ) 14 (да-чжуй) на задньому серединному меридіані між сьомим шийним та першим грудним хребцями Розташування точок визначали за анатомотопографічними даними, а також за допомогою автоматичного багатоканального пристрою для пошуку біологічно активних точок [Лувсан Г , 1986, Шульга В О , 1986, Якубов М Г, 1988, Lambardt А Б , 1989] Вакцинацію повторювали через 7 діб в аналогічний спосіб Першій групі контрольних (перша дослідна група) тварин антигени обох вакцин уводили сукупно у дозах по 1мл кожної внутрішньом'язово на внутрішній поверхні стегон Через сім діб в аналогічний спосіб вакцини уводили повторно Іншій контрольній групі (третя дослідна) антигени вакцин уводили інтракутанно у дозі 0,2мл без зв'язку з біологічно активними точками Через сім діб в аналогічний спосіб вакцини уводили повторно Загальний стан тварин першої групи у процесі імунізації був задовільний, випадків захворювання на парвовірусний ентерит, чуму м'ясоїдних, аденовірусну інфекцію протягом 1,5 року спостережень не зафіксовано Однак окремі тварини в ході імунізації хворіли на штеркурентні захворювання дерматит, кон'юнктивіт Ступінь перебігу захворювань був легким Загальні КЛІНІЧНІ показники (температура, пульс, частота дихальних рухів були в межах норми) Загальна КІЛЬКІСТЬ еритроцитів і тромбоцитів, рівень гемоглобіну істотно не змінювались впродовж спостереження На 7 день після імунізації ВІЦІ О о о 0 0 о ю 50800 виявлений лейкоцитоз за рахунок абсолютного лімфоцитозу, поява активованих лімфоцитів, ео зинопенія (табл 1) Таблиця 1 Склад формених елементів крові тварин першої групи в процесі імунізації» (%, М±ш, д=15) День Паличко- Сегменто Лімфо- Активо- Моноци Еозино-ти філи вані ядерні ядерні цити досліду лімфоци нейтрофі нейтрофіти ли ли 2,4 ±0,6 52,8 ±0,9 37,6 ±2,9 1,6+0,4 3,8 +.0,7 1,8 ±0,4 0-й 7-й 2,2 ±0,3 45,0± 7,02 35,8+7,8 12,4+4,3 4,6 +0,8 0+0 10,0 ±1,7 72,6+2,7 9,6 ±2,2 2,4 + 0,8 4,8 ±0,5 0,6 ± 0,2 14-й 21-й 2,0 ±0,5 71,4+4,0 15,6 ±3,4 5,4 +0,6 3,4 ±0,2 2,2 ±0,2 28-й 2,0 ±0,4 61,4 + 6,0 20,0 ±3,7 1,4 ±0,3 2,8 ±0,5 12,4+2,5 35-й 2,3 ± 0,5 54,7 + 1,5 35,6 ±1,5 1,2 ±0,1 4,1 ±0,4 2Д±0 ? 3 На 14-й день після імунізації збільшення загальної КІЛЬКОСТІ лейкоцитів було зумовлено збільшенням абсолютної КІЛЬКОСТІ нейтрофільних лейкоцитів за рахунок сегментоядерних і паличкоядерних зі зсувом лейкоцитарної формули вліво Найістотнішим є те, що на 28-й день від початку досліду було різке збільшення відносної та абсолютної КІЛЬКОСТІ еозинофільних лейкоцитів На 35-й день досліду показники периферичної крові тварин практично нормалізувались Про стан активації лімфоідних клітин свідчила активність внутрішньоклітинних ферментів і вмісту PAS-позитивних речовин Під час проведення імунізації збільшилась відносна КІЛЬКІСТЬ КЛІТИН ІЗ негативним і дифузним забарвленням цитоплазми при виявленні активності кислої фосфатази (КФ), заявились клітини з активністю ферменту у формі зливних гранул Одночасно зменшилась відносна КІЛЬКІСТЬ лімфоцитів, що перебували у стані спокою та мали дрібногранулярний характер реакції Зміни активності КФ були найсуттєвішими на 14-й і 21-й дні після початку імунізації, що збігається з появою активованих лімфоцитів у складі лейкоцитарної формули Зміна активності кислої неспецифічної естерази (КНЕ) в лімфоцитах у процесі імунізації виявлялась насамперед у різкому зменшенні КІЛЬКОСТІ КЛІТИН із наявністю однієї гранули ферменту та дрібногранулярним характером реакції Наявність PAS-позитивних речовин у лімфоцитах визначається в проліферуючих клітинах, на ПІЗНІХ стадіях їх активації Ми виявляли появу 310% PAS-позитивних клітин у периферичній крові на 14-28-й день після початку досліду Важливо вивчити спочатку показники клітинної ланки імунітету до початку імунізації (табл 2) В доступній літературі ми не знайшли даних із цього питання, тому провели порівняння отриманих нами даних з аналогічними показниками у людини, враховуючи вік дослідних тварин Таблиця 2 Субпопуляційний склад лімфоцитів тварин першої групи, позитивно реагуючі клітини, (%, М+т, п=15) HLA-DR День CD4 CD8 CD19 CD5 досліду 63,5+2,7 9,8+1,3 11,2+0,95 0-й 42,2+1,7 25,8+1,6 7-й 71,6 + 2,1 41,9 ±2.7 36,7 ±1,5 10,2 + 0,9 26,7 +2.8 14-й 73,6 ±5,2 45.7 ±3,2 36,8 ±3,5 14,8+2,5 23,5+4,1 28-й 66,7 ±3,5 54,5 ±4,3 28,7+2,4 25,3 ±3,1 12,8+3,2 Серед лімфоідних КЛІТИН переважали Тлімфоцити, КІЛЬКІСТЬ яких ми оцінювали за експересією пан-Т-клітинного антигену CD5, який, як відомо, з'являється на стадії медулярних тимоци тів і зберігається протягом усього існування Тлімфоцитів периферичної крові Мала частина Вклітин, які вже контактували з антигеном у периферичних органах лімфопоезу, передусім у лім 50800 фатичних вузлах, теж може експресувати CD5 антиген, але в незначній КІЛЬКОСТІ, ЩО не може бути виявлена використаними нами методами Тому всі СО5-позитивні лімфоцити (63,5 + 2,7%) ми обгрунтовано вважали Т-лімфоцитами їх відносна КІЛЬКІСТЬ співпадала з КІЛЬКІСТЮ Т-лімфоцитів люди та у ВІЦІ 5-7-років [Berek С , Ziegner M , 1993] Серед Т-лімфоцитів переважали Т-лімфоцити з експресією антигену CD4 (42,2 + 1,7%), функція яких насамперед полягає у допомозі В-клітинам в реалізації ВІДПОВІДІ на антиген CD4 лімфоцити надають В-лімфоцитам корецепторний сигнал, який посилює активаційну його дію, що передається при зв'язуванні імуноглобулинового рецептору В-лімфоциту розчинною формою антигену [Varela F J , Countinho A, Dupire В, 1988] КІЛЬКІСТЬ Тлімфоцитів з експресією антигену CD8 була значно меншою (25,8 + 1,6%) Серед останніх присутні як цитотоксичні Т-лімфоцити, які безпосередньо впливають на клітини, уражені вірусом, так і супресорні Т-клітини, які пригнічують функціональну активність В-клітин на ПІЗНІХ стадіях імуногенезу Потрібно зазначити, що на сучасному рівні можливо провести диференціювання цих двох ПІДТИПІВ за допомогою використання клітинного сортеру і одночасно двох моноклональних антитіл, позначених різними типами флуорохромів, до різних антигенів поверхневої мембрани Але ми не мали змоги використати таке обладнання Співвідношення CD4/CD8 становило в середньому 1,63 + 0,03 Це дещо нижче, ніж аналогічний показник у людини ВІДПОВІДНОГО віку (1,9 з коливаннями 1,5 - 2,9) КІЛЬКІСТЬ В-лімфоцитів, яку ми визначали за експресією пан-В-клітинного антигену CD19, в периферичній крові не перевищувала 15% (в середньому склала 9,8 + 1,3%) Експресія цього антигену не притаманна іншим типам клітин На підставі одночасного визначення експресії антигенів CD19 і HLA-DR ми мали змогу оцінити не тільки КІЛЬКІСТЬ В-клітин, але й КІЛЬКІСТЬ активованих В-лімфоцитів HLA-DR антиген можна виявити на всіх В-лімфоцитах, моноцитах і на Тлімфоцитах, які знаходяться у стані активації Моноцити з легкістю відрізняються від лімфоідних клітин за цитоморфолопчними ознаками, оскільки ми визначали експресію антигенів імунопероксидазним методом і результати реакції враховували під світлооптичним мікроскопом У неімунізованих тварин КІЛЬКІСТЬ CD19+ і HLA-DR+ лімфоцитів практично співпадала, що свідчить про відсутність у них активованих лімфоцитів Т-клітинної ланки імунітету Як видно з даних табл 3, на 7-й день після імунізації підвищилась КІЛЬКІСТЬ активованих Тлімфоцитів, що експресують HLA-DR антигени, та CD8+ лімфоцитів - найімовірніше цитотоксичних Тлімфоцитів ефекторів, що знищують клітини-носм чужорідної інформації За рахунок останніх спів відношення CD4/CD8 клітин зменшується в середньому від 1,63 до 1,24 Цей стан активації імунної системи майже відсутній на 14-й день після початку імунізації Збільшення КІЛЬКОСТІ CD19 - позитивних В-клітин (в середньому на 45-57% порівнянно з первісним станом) відбувається на 28-й день У ті ж самі терміни серед Т-лімфоцитів периферичної крові переважають CD4+ Т-лімфоцити хелпери індуктори Це призводить до зростання індексу CD4/CD8, який становить 1,89 + 2,07 Слід зазначити, що це збігається зі збільшенням КІЛЬКОСТІ еозинофілів у складі лейкоцитарної формули тварин Гуморальна ВІДПОВІДЬ на антигени, що вводились була високою максимальний титр для "Альфавак 3" встановлений на 14-й день — 6,0 ~ 8,0, який знижувався на 28-й день - 4,0 ~ 6,0, а потім на 35-й день (3,0 ~ 4,0) Аналогічною була тенденція щодо титрів антитіл проти чуми м"ясоідних (вакцина "Альфавак 1") Найістотнішу гуморальну ВІДПОВІДЬ виявили на 14-й день (4,0 ~ 5,0), з подальшим зниженням на 28-й день (3,0 ~ 4,0) і 35-й день дослідження (2,0-3,0) Отже, при імунізації у тварин першої групи виявлена ефективна імунна ВІДПОВІДЬ ЯК гуморальна, так і на клітинному рівні Водночас виявлене значне підвищення КІЛЬКОСТІ CD8+ цитотоксичних лімфоцитів на першому етапі і наступна активація Тхелперів і еозинофілів можуть призводити у тварин до активації хронічних і приєднання інтеркурентних захворювань, розвитку алергічних реакцій та загострення хронічних хвороб з наявністю алергічного компоненту Імунізація проходила без ускладнень, поствакцинальний імунітет був стійкий Жодного випадку розвитку інфекційних захворювань або приєднання штеркурентних інфекцій не спостерігали З боку гематологічних показників виявили такі ж самі зміни, як і в тварин першої групи, але ступінь їх прояву був значно нижчим (табл 3) У тварин другої групи ми не встановили фази початкового лімфоцитозу на 7-й день після початку імунізації, а незначне підвищення загальної КІЛЬКОСТІ лейкоцитів було зумовлено нейтрофільними сегментоядерними лейкоцитами До того ж КІЛЬКІСТЬ паличкоядерних гранулоцитів не підвищувалась Активація лімфоідних клітин за цитоморфолопчними ознаками була незначною- 10-14% активованих клітин від загальної КІЛЬКОСТІ лімфоцитів, але зберігалась достатньо довго - до 35-го дня спостереження Етапу еозинопенм не було, еозинофілія на 28-й день після імунізації була меншою, ніжутварин першої групи Під час дослідження активності КФ виявлено різке зменшення відносної КІЛЬКОСТІ клітин з однією гранулою ферменту і появу клітин із зливним характером реакції вже на 7-й день після початку імунізації 50800 Таблиця З Склад формених елементів крові тварин другої групи в процесі імунізації, (%, М+m, п=15) День Паличко- Сегменто Лімфо- Активо- Моно- Еозинофіли цити досліду цити вані ядерні ядерні лімфонейтронейтроцити філи філи 0-й 2,7 + 0,8 55,7 ±1,6 35,2 ±1,8 0,7 ±0,2 4,2 ±0,3 1,5+0,1 7-й 2,3 ±0,2 70,4± 5,1 19,4+3,8 3,2 ±0,5 4,1 ±0,4 0,6±0Д 14 й 2,6 ± 0,8 71,3+5,4 18,8+2,7 2,3 + 0,2 3,6 ±0,4 1,4 + 0,3 21-й 1,6 ±0,4 60,7+ЗД 26,2 ±4,1 4,3 ±0,9 6,2 ±0,7 1,0 ±0,5 28-й 1,9 ±0,2 58,4 + 2,1 27Д ±1,9 2,5 ± 0,8 4,8+0,8 5,3 +1,9 35-й 1,8 ±0,4 57,9 ±1,8 31,0 ±4,4 2,1 +0,5 4,0+0,3 3,2 + 0,6 Ці ЗМІНИ лишались сталими до 35-го дня досліду Як і в тварин першої групи, зменшення клітин з однією гранулою КФ збігалося з періодом лімфопенм, а нормалізація ~ зі збільшенням абсолютної КІЛЬКОСТІ лімфоцитів у периферичній крові Активність КНЕ В лімфоцитах практично не змінювалась За період спостереження ми не виявили PAS-позитивних клітин Отже, менш виражені цитоморфолопчні прояви активації лімфоцитів у тварин другої групи збігалися з браком змін активності КНЕ і PASреакцм Показники клітинної ланки імунітету у тварин другої групи до початку імунізації не відрізнялись від тварин першої групи за КІЛЬКІСНИМИ та якісними показниками У субпопуляційному складі лімфоцитів ми встановили на 7-й день після початку імунізації зменшення КІЛЬКОСТІ Т-клітин за рахунок CD8+ Тлімфоцитів (табл 4) Це основна ВІДМІННІСТЬ від тварин першої групи, в яких відбувалася активація Т-супресорів На 14-й день значна частина Т-лімфоцитів все ще знаходилась у стані активації , про що свідчить велика КІЛЬКІСТЬ HLA-DR+ клітин порівняно з КІЛЬКІСТЮ CD19+ В-клітин Тоді Ж КІЛЬКІСТЬ HLA-DR+ активованих лімфоцитів у тварин першої групи набувала первинних значень На 28-й день досліду, як і в першій групі, підвищувалась КІЛЬКІСТЬ Влімфоцитів (р 0,05) і був ймовірно нижчим, ніж у тварин другої групи (р < 0,05) Отже, перебіг імунологічних реакцій у тварин третьої дослідної групи за більшістю показників збігався з такими другої групи, яким імунізацію проводили штракутанно v біологічно активні точки 11 50800 12 Таблиця 6 Титри антитіл у сироватці крові імунізованих тварин третьої групи, M+m, n=15) Титри антитіл Реакція нейтралізації День Реакція гальмування вірусу чуми м'ясоїдниїх досліду гемаглютинації 0 0 0-й 2,4 + 0,01 7-й 4,8 + 0,08 14-й 7,0 + 0,09 4,6 ±0,08 21-й 6,0+ 0,04 3,8 ±0,06 28-й 4,6 + 0,05 3,6 ±0,08 35-й 3,6+0,0 3 2,6 + 0,01 Це стосується динаміки змін КІЛЬКОСТІ нейтрофільних і еозинофільних гранулоцитів, лімфоцитів, субпопуляційного складу лімфоцитів ВІДМІННИМ був менш виражений і менш стабільний ступінь активації лімфоцитів, визначений за цитоморфолопчними, ЦИТОХІМІЧНИМИ та імунологічними ознаками, й ВІДПОВІДНО менший рівень гуморальної ВІДПОВІДІ Показники гуморальної ВІДПОВІДІ тварин третьої і першої груп збігалися Отже, проведеними дослідженнями встановлено, що спосіб аплікації антигену значно впливає на ефективність імунізації Отже, запропонований спосіб уведення антигену викликає більш значну гуморальну ВІДПОВІДЬ на антиген, порівняно з тваринами першої і третьої дослідних груп Отримані результати засвідчують про те, що при антигенному подразненні тварин у традиційний спосіб (внутрішньом'язево) розвивається ефективний клітинний і гуморальний імунітет Початок імунної ВІДПОВІДІ та основні її етапи відбуваються у крові Водночас виявлені зміни вказують на можливий розвиток у окремих тварин (ослаблених, тих, що утримуються на неповноцінному раціоні, мають хронічні, або спадкові хвороби) деяких ускладнень Так, висока активність CD8+ цитотоксичних лімфоцитів, які на перших етапах розвитку імунної ВІДПОВІДІ не мають суворого відношення до антигенспецифічного клону, може призводити до активації хронічних, або приєднання штеркурентних захворювань Саме це ми встановили у окремих тварин першої групи Активація Т-лімфоцитів хелперів і еозинофілів на другому етапі імунної ВІДПОВІДІ в свою чергу може призводити до розвитку алергічних реакцій та загострення хронічних хвороб із наявністю алергічного компоненту Схема, за якою розвивається імунна реакція за імунізації штракутанно, дещо різниться з наведеною вище По-перше, місце розвитку імунної реакції у цьому разі - не кров, а безпосередньо шкіра Тобто імунна ВІДПОВІДЬ ІЗ самого початку відбувається локально Про це свідчить перерозподіл лімфоідних клітин в організмі та їх хомінг у місце присутності антигена - в шкіру вже на перші дні після імунізації, відсутність статистичне вірогідної нейтрофільної реакції і зсуву лейкоцитарної формули вліво, брак активації Т-супресорів Антигенпрезентуючими клітинами в данному випадку здебільшого є не моноцити, а як відомо з літературних джерел [Clark Е А , Ledbetter J А , 1994] клітини Лангерганса Вони активують Тлімфоцити хелпери як першого, так і другого типу Після зустрічі з антигеном частина активованих клітин Лангерганса та активованих Т-лімфоцитів мігрує в найближчий репонарний лімфовузол, де відбувається реакція зародкового центру, активація В-лімфоцитів (після взаємодії з антигеном, експресованим на КЛІТИНІ Лангерганса та CD40L антигеном на поверхні активованого Т-лімфоциту) і їх стимуляція до синтезу і секреції антитіл, підвищення афінності та ЗВІДНОСТІ імуноглобулінів внаслідок соматичних ппермутацій Отже, при імунізації тварин безпосередньо в шкіру основною є саме імунна реакція в периферичних органах імуногенезу і менш виражені загальні зміни з боку показників крові Це робить процес імунізації більш ефективним, про що свідчить використання для штракутанної імунізації значно менших доз антигенів, ніж потрібно для проведення для ефективної внутрішньомязевої імунізації У тварин, яких імунізували штракутанно у біологічно активні точки, як ми вважаємо, розвиток імунних реакцій за багатьма показниками збігався з такими у тварин третьої групи (інтракутанна імунізація) Про це свідчать відсутність нейтрофільної реакції , активації цитотоксичних клітин, термін розвитку лімфопенм, динаміка змін загальних показників крові Але на відміну від останніх імунізація у БАТ призводила до більш вираженої і більш стійкої гуморальної ВІДПОВІДІ, про що свідчить статистичний аналіз 13 50800 ДП «Український інститут промислової власності» (Укрпатент) вул Сім'ї Хохлових, 15, м Київ, 04119, Україна ( 0 4 4 ) 4 5 6 - 2 0 - 90 ТОВ "Міжнародний науковий комітет" вул Артема, 77, м Київ, 04050, Україна (044)216-32-71 14
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for vaccination of animals against infectious diseases
Автори англійськоюVelychko Serhii Volodymyrovych
Назва патенту російськоюСпособ вакцинации животных против инфекционных заболеваний
Автори російськоюВеличко Сергей Владимирович
МПК / Мітки
МПК: A61K 39/00, A61D 7/00
Мітки: вакцинації, спосіб, захворювань, інфекційних, тварин
Код посилання
<a href="https://ua.patents.su/7-50800-sposib-vakcinaci-tvarin-vid-infekcijjnikh-zakhvoryuvan.html" target="_blank" rel="follow" title="База патентів України">Спосіб вакцинації тварин від інфекційних захворювань</a>
Спосіб одержання препарату для профілактики інфекційних захворювань у свійських тварин

Номер патенту: 10273
Опубліковано: 25.12.1996
Автори: Замковий Віталій Кузьмович, Кіфоренко Євген Іванович, Кулаков Дмітрій Фьодоровіч, Потапов Юрій Миколайович
МПК: A61K 33/34
Мітки: препарату, інфекційних, тварин, одержання, спосіб, свійських, захворювань, профілактики
Формула / Реферат:
Способ получения препарата для профилактики инфекционных заболеваний домашних животных, предусматривающий осуществление электролиза в водной среде с использованием анода, выполненного из материала, являющегося источником антисептических ионов, и размещенного в мембранной оболочке, непроницаемой для упомянутых ионов, до получения в оболочке требуемой концентрации указанных ионов, отличающийся тем, что электролиз осуществляют в...
Спосіб визначення нешкідливості вакцинних препаратів проти інфекційних хвороб тварин

Номер патенту: 41113
Опубліковано: 15.08.2001
Автори: Головко Анатолій Миколайович, Ушкалов Валерій Олександрович, Коваленко Лариса Володимирівна, Нікітенко Юрій Вікторович, Романько Марина Євгеніївна
МПК: A61K 39/00
Мітки: нешкідливості, спосіб, вакцинних, тварин, хвороб, інфекційних, препаратів, визначення
Формула / Реферат:
Спосіб визначення нешкідливості вакцинних препаратів проти інфекційних хвороб тварин, призначений для лабораторної оцінки нешкідливості вакцин, що включає аналіз результатів досліджень крові імунізованих і контрольних тварин та визначення показників, що характеризують рівень накопичення токсичних продуктів перекисного окислення ліпідів та активності антиоксидантної системи, який відрізняється тим, що критерієм оцінки нешкідливості служить...
Мутант коров’ячого вірусу герпесу типу 1 (bhv-1), композиція вакцини для вакцинації тварин для захисту їх від bhv-1, композиція вакцини для вакцинації тварин для захисту їх від патогена

Номер патенту: 40571
Опубліковано: 15.08.2001
Автори: Оірсот Йоханнес Теодорус ван, Рейсевейк Франціскус Антоніус Марія, Має Ооже Кам'єль
МПК: G01N 33/569, A61K 39/245, C12Q 1/70, C12N 15/83, C12N 15/09, C12N 7/00, C12N 15/00, A61K 39/395, C12N 7/01, G01N 33/53, C07K 14/06, C12N 15/869, A61K 39/265, A61P 31/12, C12P 21/08
Мітки: тварин, патогена, типу, герпесу, мутант, вірусу, bhv-1, коров'ячого, вакцинації, вакцини, композиція, захисту
Формула / Реферат:
1.Мутант коровьего вируса герпеса типа 1 (BHV-1), имеющий делецию в gЕ-гене гликопротеина, причем gЕ-ген гликопротеина расположен в US-регионе между сайтами рестрикции Pst 1 и Sma 1, причем указанная делеция позволяет мутанту отличаться серологически от BHV-1 дикого типа.2.Мутант BHV-1 по п.1, отличающийся тем, что указанная делеция в gЕ-гене гликопротеина вызвана аттенуацией.3.Мутант BHV-1 по п. 2, отличающийся тем, что...
Спосіб визначення генетичної схильності до інфекційних захворювань

Номер патенту: 45129
Опубліковано: 15.03.2002
Автори: Дизик Галина Михайлівна, Незгода Ірина Іванівна
МПК: G01N 33/536, G01N 33/569, G01N 33/48, C12N 1/20, C12R 1/00
Мітки: генетично, інфекційних, спосіб, визначення, захворювань, схильності
Формула / Реферат:
1. Спосіб визначення генетичної схильності до інфекційних захворювань за групами крові АВО, що включає моделювання реакції ізогемаглютинації і введення в реагуючу систему полісахаридної фракції мікроорганізмів з подібними до речовини групи крові цукрами, який відрізняється тим, що, крім полісахаридної фракції, в реакцію вводиться ендотоксин відповідного інфекційного збудника з попереднім підбором його субгемолітичної дози шляхом розведення...
Спосіб захисту шовкопряду від інфекційних захворювань

Номер патенту: 12368
Опубліковано: 02.12.1996
Автори: Алексеніцер Майя Леонідівна, Кириченко Іван Олександрович, Головко Валерій Олексійович, Аретинська Тетяна Борисівна
МПК: A01K 67/04
Мітки: шовкопряду, інфекційних, захворювань, захисту, спосіб
Формула / Реферат:
Способ защиты шелкопряда от инфекционных заболеваний, включающий обработку корма гусениц водным раствором ароматической кислоты бензойного ряда, отличающийся тем, что корм гусениц обрабатывают водным раствором бензойной кислоты концентрацией 5-50 мг/л.
Попередній патент: Двигун внутрішнього згоряння (варіанти) і спосіб роботи двигуна внутрішнього згоряння (варіанти)
Наступний патент: Вогнетривка набивна маса
Випадковий патент: Привід круглов`язальної машини