Взаємодія moraxella catarrhalis з епітеліальними клітинами, позаклітинними матриксними білками і системою комплементу







Формула / Реферат
1. Пептид, що складається з послідовності ID NO: 2, або його продукт гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин.
2. Пептид, що складається з послідовності ID NO: 3, або його продукт гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин.
3. Застосування принаймні одного пептиду за будь-яким з пп. 1-2 для отримання лікарського засобу для лікування або профілактики інфекції, викликаної Moraxella catarrhalis.
4. Застосування за п. 3 для профілактики або лікування запалення середнього вуха, синуситу або інфекцій нижніх дихальних шляхів.
5. Ліганд, що включає домен, що зв'язує фібронектин і складається з амінокислотної послідовності, вибраної з групи, яка складається з послідовності ID NO: 2 або послідовності ID NО:3, або його продукту гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин.
6. Злитий білок, що включає один або декілька лігандів за п. 5.
7. Лікарський засіб, що містить один або декілька лігандів за п. 5 або п. 6 і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів.
8. Вакцина, що містить один або декілька лігандів за п. 5 або 6 і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів.
9. Спосіб лікування або профілактики у індивідуума інфекції, викликаної Moraxella catarrhalis, що включає введення фармацевтично ефективної кількості лікарського засобу за п. 7 або вакцини за п. 8.
10. Послідовність нуклеїнової кислоти, яка кодує пептид за п. 1 або 2.
11. Поліпептид або зрізаний поліпептид, що включає принаймні одну з консервативних послідовностей послідовності ID NО:2 або послідовності ID NО:3 причому зрізання зберігає функцію зв’язування ламініну і/або функцію зв’язування фібронектину і/або функцію зв’язування С3; або його продукту гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин.
Текст
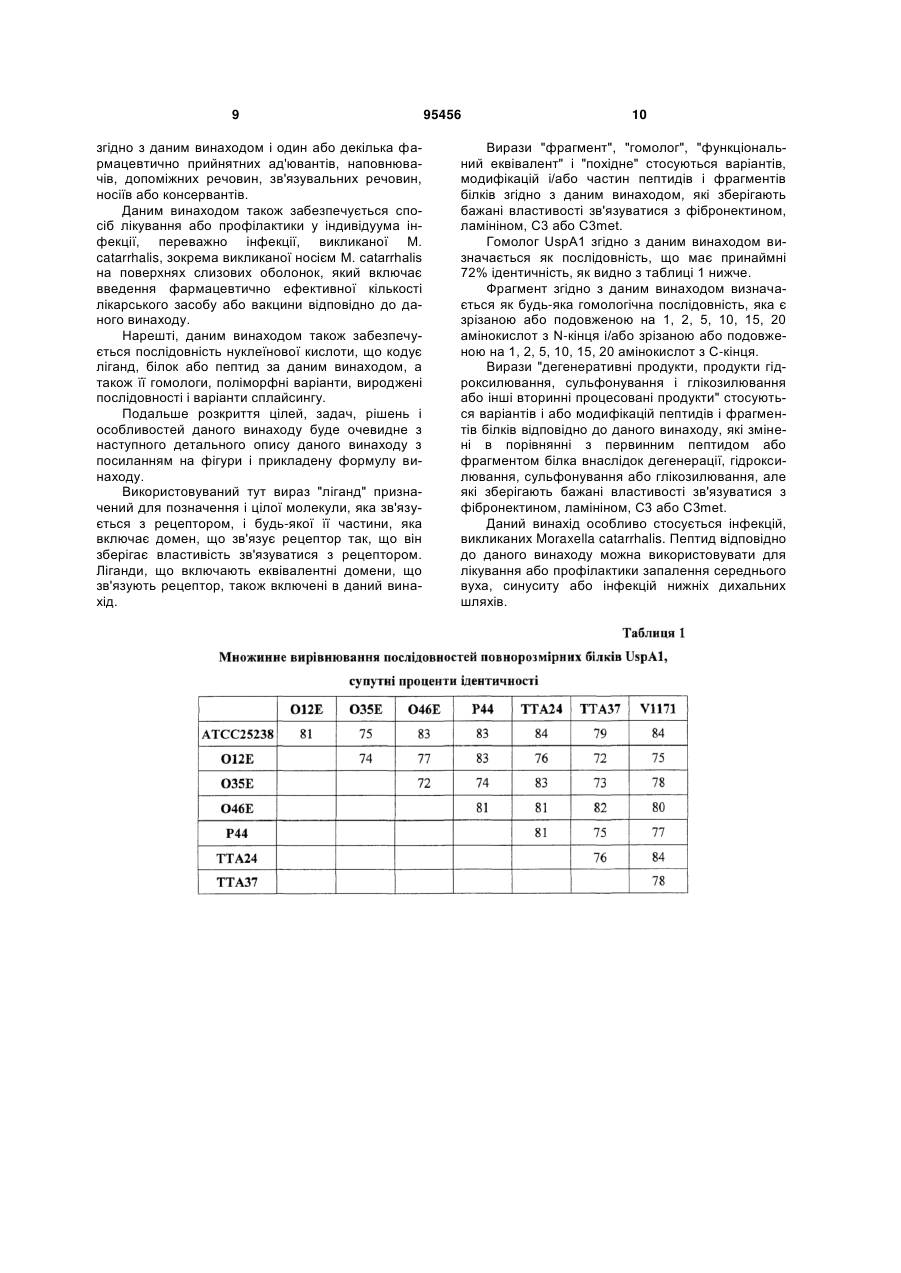
1. Пептид, що складається з послідовності ID NO: 2, або його продукт гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин. 2. Пептид, що складається з послідовності ID NO: 3, або його продукт гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин. 3. Застосування принаймні одного пептиду за будь-яким з пп. 1-2 для отримання лікарського засобу для лікування або профілактики інфекції, викликаної Moraxella catarrhalis. 4. Застосування за п. 3 для профілактики або лікування запалення середнього вуха, синуситу або інфекцій нижніх дихальних шляхів. 5. Ліганд, що включає домен, що зв'язує фібронектин і складається з амінокислотної послідовності, вибраної з групи, яка складається з послідовності ID NO: 2 або послідовності ID NО:3, або його продукту гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин. 6. Злитий білок, що включає один або декілька лігандів за п. 5. 7. Лікарський засіб, що містить один або декілька лігандів за п. 5 або п. 6 і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів. UA (21) a200802951 (22) 08.08.2006 (24) 10.08.2011 (86) PCT/SE2006/000931, 08.08.2006 (31) 60/706,745 (32) 10.08.2005 (33) US (31) 60/707,148 (32) 11.08.2005 (33) US (46) 10.08.2011, Бюл.№ 15, 2011 р. (72) ФОРСГРЕН АРНЕ, SE, РІЄСБЕКК КРІСТІАН, SE (73) АРНЕ ФОРСГРЕН АБ, SE (56) BLOM A.M.: 'Strategies developed by bacteria and virus for protection from the human complement system' SCAND. J. CLIN. LAB. INVEST. vol. 64, 2004, с. 479 - 495. NORDSTRo"M T. ET AL.: 'The Emerging Pathogen Moraxella catarrhalis Interacts with Complement Inhibitor C4b Binding Protein through Ubiquitous Surface Proteins A1 and A2' THE JOURNAL OF IMMUNOLOGY vol. 173, 2004, с. 4598 - 4606. NORDSTRo"M T.: 'Lund University Dissertations', 01 April 2005, SCRIPTA ACADEMICA LUNDENSIA article 'Moraxella catarrhalis outer membrane proteins and interactions with the human immune system', с. 1 - 4. NORDSTRo"M T. ET AL.: 'Ionic binding of C3 to the human pathogen Moraxella catarrhalis is a unique mechanism for combating innate immunity' J. IMMUNOL. vol. 175, no. 6, 15 September 2005, с. 3628 - 3636. AEBI C. ET AL.: 'A Protective Epitope of Moraxella catarrhalis Is Encoded by Two Different Genes' INFECTION AND IMMUNITY vol. 65, no. 11, November 1997, с. 4367 - 4377. MCMICHAEL J.C. ET AL.: 'Isolation and Characterization of Two Proteins from Moraxella catarrhalis That Bear a Common Epitope' INFECTION AND IMMUNITY vol. 66, no. 9, September 1998, с. 4374 - 4381. TAN T.T. ET AL.: 'The Respiratory Pathogen Moraxella catarrhalis Adheres to Epithelial Cells by Interacting with Fibronectin through Ubiquitous 2 (19) 1 3 95456 4 8. Вакцина, що містить один або декілька лігандів за п. 5 або 6 і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів. 9. Спосіб лікування або профілактики у індивідуума інфекції, викликаної Moraxella catarrhalis, що включає введення фармацевтично ефективної кількості лікарського засобу за п. 7 або вакцини за п. 8. 10. Послідовність нуклеїнової кислоти, яка кодує пептид за п. 1 або 2. 11. Поліпептид або зрізаний поліпептид, що включає принаймні одну з консервативних послідовностей послідовності ID NО:2 або послідовності ID NО:3, причому зрізання зберігає функцію зв’язування ламініну і/або функцію зв’язування фібронектину і/або функцію зв’язування С3; або його продукту гідроксилювання, сульфонування або глікозилювання, який зберігає властивості зв'язувати фібронектин. Галузь техніки Даний винахід стосується Moraxella catarrhalis і їх здатності взаємодіяти з епітеліальними клітинами через позаклітинні матриксні білки, такі як фібронектин і ламінін, а також їх здатності інгібувати систему комплементу. Взаємодія з цими позаклітинними білками використовується для приготування вакцин. Рівень техніки Здатність зв'язуватися з епітеліальними клітинами має величезне значення для декількох видів бактерій. Наприклад, Staphylococcus aureus і Streptococcus pyogenes мають білки, що зв'язують фібронектин (FnBP) з спорідненою організацією послідовностей. Ці FnBP відомі як мікробні поверхневі компоненти, що впізнають адгезивні матриксні молекули (MSCRAMM). При зв'язуванні з фібронектином вони використовують модульну структуру фібронектину, що утворює обширні тандемні бета-блискавки [27, 39, 47, 73]. Призначення фібронектину полягає в опосередковуванні адгезії бактерії і проникнення в клітини хазяїна. Важливий патоген слизової оболонки Moraxella catarrhalis є третьою провідною бактерійною причиною гострого запалення середнього вуха у дітей після Streptococcus pneumoniae і Haemophilus influenzae [14, 40, 55]. Μ. catarrhalis також є одним з найбільш частих мешканців глотки здорових дітей. Більш того, М. catarrhalis також є частою причиною синуситу і інфекцій нижніх дихальних шляхів у дорослих з хронічним обструктивним захворюванням легень (COPD) [74]. Ймовірно, успіх цього виду у хворих COPD частково пов'язаний з наявністю у нього великого набору адгезинів. В останні роки дослідження фокусувалося на білках зовнішньої мембрани і їх взаємодіях з хазяїном, що є людиною [6, 48, 56]. Деякі з цих білків зовнішньої мембрани, в тому числі серед інших той, що зв'язує IgD M. catarrhalis (MID, що також позначається Hag), білок CD, білок адгезії М. catarrhalis (McaP) і повсюдні поверхневі білки (Usp), мають, очевидно, адгезивні функції [1, 22, 33, 48, 61, 81, 84]. Суть винаходу У зв'язку з тим фактом, що М. catarrhalis є, як виявлено, провідною причиною інфікування верхніх і нижніх дихальних шляхів, існує поточна необхідність в розробці вакцин, які можна використати проти М. catarrhalis. Тому мета даного винаходу полягала у виявленні того, яким чином М. catarrhalis взаємодіє з епітеліальними клітинами в організмі і впливає на імунну систему. Таким способом можна розробити речовини, які можуть діяти як вакцини проти Μ catarrhalis. У цьому дослідженні автори даного винаходу, використовуючи мутанти М. catarrhalis, що походять з клінічних ізолятів, змогли показати, що і UspA1, і А2 зв'язують фібронектин і ламінін. Крім того, автори даного винаходу змогли показати, що М. catarrhalis блокує класичний шлях системи активації комплементу, а також пояснити, яким чином вони здійснюють блокування. Багато які бактерії адгезуються до епітеліальних клітин через MSCRAMM, що зв'язують фібронектин [54, 77]. Pseudomonas aeruginosa має FnBP, який зв'язується з асоційованим з клітиною фібронектином на епітеліальних клітинах порожнини носа [69]. Блокування взаємодій бактеріїбілок фібронектин може допомогти тканині хазяїна подолати інфекцію. Дійсно було показано, що антитіла проти FnBP S. aureus приводять до швидкого знешкодження бактерії у інфікованих мишей [71]. Рекомбінантні зрізані білки UspA1/A2 разом з більш маленькими фрагментами, що перекривають цілу молекулу, перевірили відповідно до даного винаходу на зв'язування з фібронектином. І UspA1, і А2 зв'язували фібронектин, і виявлено, що домени, що зв'язують фібронектин, розташо299-452 165-318 вані межах UspA1 і UspA2 . Ці два зрізаних білки, обидва, інгібували зв'язування М. catarrhalis з епітеліальними клітинами кон'юнктиви Чанг в мірі, схожій з інгібуванням з використанням антитіл проти фібронектину. Дані спостереження показали, що і UspA1, і А2 М. catarrhalis залучені в адгезію до епітеліальних клітин через асоційований з клітиною фібронектин. Отже, як можливі кандидати, які потрібно включати у вакцину проти М. catarrhalis, пропонуються біологічно активні сайти 299-452 165-318 в межах UspA1 і UspA2 . Крім того, автори даного винаходу вивчили і охарактеризували зв'язування Μ. catarrhalis з ламініном. М. catarrhalis є частою причиною загострень інфекцій у хворих на COPD. Ймовірно, успіх цього виду у хворих з COPD частково пов'язаний з наявністю у нього великого набору адгезинів. Крім того, існують патологічні зміни, такі як втрата цілісності епітелію з оголенням базальної мембрани, 5 де у курців потовщений сам шар ламініну [4]. Показано, що деякі патогени здатні зв'язувати ламінін, і це може робити внесок в їх здатність адгезуватися до таких пошкоджених і оголених поверхонь слизової оболонки. Ці патогени включають патогени, які, як відомо, спричиняють значуще захворювання дихальних шляхів, такі як серед інших S. aureus і P. aeruginosa [7, 63]. Автори даного винаходу змогли показати, що повсюдний поверхневий білок (Usp) A1 і А2 Μ. catarrhalis також зв'язується з ламініном. Домени UspA1 і А2, що зв'язують ламінін, були виявлені, серед інших, 50-491 30в межах N-кінцевих половин UspA1 і UspA2 351 . Ці домени також містять домени, що зв'язують фібронектин. Однак, найменші фрагменти, які 299-452 165-318 зв'язували фібронектин, UspA1 і UspA2 , не зв'язували в будь-якій помітній мірі ламінін. Фрагменти, менші, ніж N-кінцева половина UspA1 50-491 (UspA1 ), втрачають всю свою здатність зв'язувати ламінін, в той час як у випадку UspA2, тіль30-170 ки UspA2 зв'язував ламінін, хоч на більш низькому рівні, ніж цілий рекомбінантний білок 30-539 (UspA2 ). Ці відкриття кажуть про те, що різні частини молекули можуть мати різні функціональні 50-770 ролі. Також виявлено, що UspA1 має властивості зв'язувати ламінін. При порівнянні найменших областей UspA1 і А2, що зв'язують ламінін, автори даного винаходу виявили, що існує, однак, незначна подібність у 30-170 вигляді амінокислотної гомології, між UspA2 і 50-491 UspA1 (дані не показані). Це не дивно, оскільки відомий той факт, що обидва білки мають структуру сферичної головки у вигляді "льодяника", незважаючи на те, що їх N-кінцеві половини ідентичні тільки на 22% [2, 32]. Як можливі кандидати, які потрібно включати у вакцину проти М. catarrhalis, пропонуються біоло50-770 30гічно активні сайти в межах UspA1 і UspA2 539 . Нарешті, автори даного винаходу вивчили взаємодію між повсюдними поверхневими білками А1 і А2 М. catarrhalis і вродженою імунною системою і виявили, що М. catarrhalis блокує систему комплементу. Система комплементу є однією з перших ліній вродженого захисту проти патогенних мікроорганізмів, і активація цієї системи приводить до каскаду осадження білків на поверхні бактерій, що приводить до утворення мембрано-атакуючого комплексу або опсонізації патогена з подальшим фагоцитозом [85, 86]. Одним з найбільш важливих білків комплементу є C3, який присутній в кровотоці в концентрації, схожій з концентрацією деяких імуноглобулінів (1-1,2 мг/мл). C3 не тільки відіграє вирішальну роль як опсонін, але також є спільною зв'язувальною ланкою між класичним, лектиновим і альтернативним шляхами активації комплементу. Альтернативний шлях функціонує як петля посилення для класичного і лектинового шляхів, і його можна також спонтанно активувати за допомогою ковалентного приєднання C3 до поверхні мікробної клітини при відсутності інгібіторів комплементу. Для осадження C3 потрібна присутність внутрішнього тіоефірного зв'язку, утвореного в нативному 1010 білку поблизу сульфгідрильної групи (Cys ) і 1012 карбонільної групи глутаміну (Gln ) на 95456 6 ланцюгу C3 [76]. Протеолітичне відщеплення з амінокінця -ланцюга C3 пептиду із 77 залишків приводить до утворення C3а (анафілатоксину) і C3b. Приєднання C3b потім здійснюється через ковалентний зв'язок між карбонільною групою метастабільного тіоефіру і або групами білків -NH2 або -ОН, або вуглеводними структурами на поверхні активатора [36, 37]. Виявлено, що UspA1 і А2 М. catarrhalis нековалентно і залежним від дози способом зв'язуються з третім компонентом комплементу (C3) з обробленої EDTA сироватки і обробленого метиламіном C3 (C3met). Виявлено, що 50-770 30-539 UspA1 і UspA2 зв'язуються з C3 і C3met. Виявлено, що область UspA2, що зв'язує C3, роз200-458 ташована головним чином в UspA2 . Однак UspA1, як виявлено, відіграє мінорну роль в цих взаємодіях. Як можливі кандидати, які потрібно включати у вакцину проти М. catarrhalis, пропону50-77 ються біологічно активні сайти в межах UspA1 і 30-539 UspA2 . Сімейство UpsA складається з UspA1 (молекулярна маса 88 кДа), UspA2 (62 кДа) і гібридного білка UspA2H (92 кДа) [2, 43]. Ці білки мігрують у вигляді високомолекулярних комплексів при електрофорезі в SDS-PAGE, є відносно консервативними і, отже, важливими вакцинними кандидатами. Амінокислотні послідовності UspA1 і А2 ідентичні на 43% і мають 140 амінокислотних залишків, які ідентичні на 93% [2]. У ряду з 108 ізолятів М. catarrhalis з носоглотки новонароджених із запаленням середнього вуха гени uspA1 і uspA2 виявлені в 107 (99%) і 108 (100%) ізолятах, відповідно. Двадцять один відсоток новонароджених були ідентифіковані як такі, що мають гібридний варіант гена UspA2H [50]. Крім того відомо, що природно набуті антитіла проти UspA1 і А2 є бактерицидними [15]. Сімейству білків UpsA приписується декілька функцій. Експресія UpsA є істотною для прикріплення М. catarrhalis до епітеліальних клітин кон'юнктиви Чанг і до епітеліальних клітин гортані Нер-2 [43, 49]. У більш недавньому дослідженні виявлено, що UpsA зв'язується з клітинними адгезивними молекулами (СЕАСАМ), що належать до канцероембріональних антигенів, що експресуються лінією епітеліальних клітин легені А549 [31]. Також показано, що очищений UspA1 зв'язує фібронектин в дот-блот аналізах, в той час як очищений UspA2 не зв'язує [49]. І UspA1, і А2 можуть відігравати важливі ролі в стійкості сироватки до М. catarrhalis [1, 5, 58, 60]. У даному винаході продемонстровано, що і UspA1, і А2 є детермінантами зв'язування М. catarrhalis з фібронектином і ламініном в клінічних ізолятах М. catarrhalis ВВН18 і RH4. Цікаво, що обидва рекомбінантних UspA1 і А2, що походять з М. catarrhalis Bc5, зв'язували фібронектин в однаковій мірі. Домени, що зв'язують фібронектин, виявлені в межах амінокислотних залишків 229-452 UspA1 і 165-318 UspA2. Послідовності цих двох доменів ідентичні по 31 амінокислотному залишку. Важливо, що зрізані фрагменти білків, що містять ці залишки в UspA1 і UspA2, були здатні інгібувати зв'язування М. catarrhalis з епітеліальними клітинами Чанг, що говорить про те, що взаємодії з ци 7 ми клітинами здійснювалися через асоційований з клітиною фібронектин. Домени, що зв'язують ламінін, виявили в межах відмічених вище амінокислотних залишків. За допомогою аналізів зв'язування з використанням рекомбінантних білків виявили, що головні зв'язувальні області розташовані в N-кінцевих частинах, де обидва білки утворюють сферичну головку. Бактерійні фактори, що опосередковують адгезію до тканини, і компоненти позаклітинного матриксу (ЕСМ), групуються разом в одне сімейство, зване "мікробні поверхневі компоненти, що розпізнають адгезивні матриксні молекули" (MSCRAMM). Оскільки і UspA1, і UspA2 зв'язують фібронектин і ламінін, ці білки можна визначити MSCRAMM. Відповідно до одного аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:1, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:2, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. У відповідності з наступним аспектом даним винаходом забезпечується пептид, що має послідовність ID NO:3, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:4, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. У відповідності з наступним аспектом даним винаходом забезпечується пептид, що має послідовність ID NO:5, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. У відповідності з наступним аспектом даним винаходом забезпечується пептид, що має послідовність ID NO:6, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:7, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. 95456 8 Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:8, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:9, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується пептид, що має послідовність ID NO:10, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Відповідно до іншого аспекту даним винаходом забезпечується застосування принаймні одного пептиду згідно з даним винаходом для отримання лікарського засобу для лікування або профілактики інфекції, переважно інфекції, викликаної М. catarrhalis, зокрема викликаної носійством М. catarrhalis на поверхнях слизових оболонок. Відповідно до іншого аспекту даним винаходом, крім того, забезпечується ліганд, що включає домен, що зв'язує фібронектин, який складається з амінокислотної послідовності, вибраної з групи, яка складається з послідовності ID NO:1, послідовності ID NO:2 і послідовностей ID NO:3, і його фрагментів, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Крім того, даним винаходом забезпечується ліганд, що включає домен, що зв'язує ламінін, який складається з амінокислотної послідовності, вибраної з групи, яка складається з послідовності ID NO:4 - послідовності ID NO:8, і його фрагменти, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Крім того, даним винаходом забезпечується ліганд, що включає домен, що зв'язує C3 або C3met, який складається з амінокислотної послідовності, вибраної з групи, яка складається з послідовності ID NO:4, послідовності ID NO:6, послідовності ID NO:9 і послідовностей ID NO:10, і його фрагментів, гомологи, функціональні еквіваленти, похідні, дегенеративні продукти або продукти гідроксилювання, сульфонування або глікозилювання і інші вторинні процесовані продукти. Крім того, даним винаходом забезпечується лікарський засіб, що містить один або декілька лігандів відповідно до даного винаходу і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів. Крім того, даним винаходом забезпечується вакцина, що містить один або декілька лігандів 9 згідно з даним винаходом і один або декілька фармацевтично прийнятних ад'ювантів, наповнювачів, допоміжних речовин, зв'язувальних речовин, носіїв або консервантів. Даним винаходом також забезпечується спосіб лікування або профілактики у індивідуума інфекції, переважно інфекції, викликаної М. catarrhalis, зокрема викликаної носієм М. catarrhalis на поверхнях слизових оболонок, який включає введення фармацевтично ефективної кількості лікарського засобу або вакцини відповідно до даного винаходу. Нарешті, даним винаходом також забезпечується послідовність нуклеїнової кислоти, що кодує ліганд, білок або пептид за даним винаходом, а також її гомологи, поліморфні варіанти, вироджені послідовності і варіанти сплайсингу. Подальше розкриття цілей, задач, рішень і особливостей даного винаходу буде очевидне з наступного детального опису даного винаходу з посиланням на фігури і прикладену формулу винаходу. Використовуваний тут вираз "ліганд" призначений для позначення і цілої молекули, яка зв'язується з рецептором, і будь-якої її частини, яка включає домен, що зв'язує рецептор так, що він зберігає властивість зв'язуватися з рецептором. Ліганди, що включають еквівалентні домени, що зв'язують рецептор, також включені в даний винахід. 95456 10 Вирази "фрагмент", "гомолог", "функціональний еквівалент" і "похідне" стосуються варіантів, модифікацій і/або частин пептидів і фрагментів білків згідно з даним винаходом, які зберігають бажані властивості зв'язуватися з фібронектином, ламініном, C3 або C3met. Гомолог UspA1 згідно з даним винаходом визначається як послідовність, що має принаймні 72% ідентичність, як видно з таблиці 1 нижче. Фрагмент згідно з даним винаходом визначається як будь-яка гомологічна послідовність, яка є зрізаною або подовженою на 1, 2, 5, 10, 15, 20 амінокислот з N-кінця і/або зрізаною або подовженою на 1, 2, 5, 10, 15, 20 амінокислот з С-кінця. Вирази "дегенеративні продукти, продукти гідроксилювання, сульфонування і глікозилювання або інші вторинні процесовані продукти" стосуються варіантів і або модифікацій пептидів і фрагментів білків відповідно до даного винаходу, які змінені в порівнянні з первинним пептидом або фрагментом білка внаслідок дегенерації, гідроксилювання, сульфонування або глікозилювання, але які зберігають бажані властивості зв'язуватися з фібронектином, ламініном, C3 або C3met. Даний винахід особливо стосується інфекцій, викликаних Moraxella catarrhalis. Пептид відповідно до даного винаходу можна використовувати для лікування або профілактики запалення середнього вуха, синуситу або інфекцій нижніх дихальних шляхів. 11 Відповідно даним винаходом забезпечується ліганд, що виділяється з білка зовнішньої мембрани Moraxella catarrhalis, який має властивість зв'язуватися з ламініном і/або фібронектином і/або C3, причому вказаний ліганд являє собою поліпептид, що включає амінокислотну послідовність, вибрану з групи, яка складається з SEQ ID NO: 1-10, що походять з представлених нижче послідовностей повнорозмірних UspA1 і UspA2 Moraxella 95456 12 catarrhalis ВС5, або що складаються з такої амінокислотної послідовності, або його фрагмент, гомолог, функціональний еквівалент, похідне, дегенеративний продукт або продукт гідроксилювання, сульфонування або глікозилювання або інший вторинний процесований продукт. Повнорозмірний UspA1 з штаму ВС5 Moraxella catarrhalis: 13 У переважному варіанті здійснення даного винаходу ліганд являє собою поліпептид [або зрізаний поліпептид в порівнянні з поліпептидом дикого типу], що включає амінокислотну послідовність, вибрану з групи, яка складається з SEQ ID NO: 110, або що складається з такої амінокислотної послідовності, або його фрагмент, гомолог, функціональний еквівалент, похідне, дегенеративний продукт або продукт гідроксилювання, сульфонування або глікозилювання або інший вторинний процесований продукт. Термін "ліганд" використовується тут для позначення і цілої молекули, яка зв'язується з ламініном і/або фібронектином і/або C3, і будь-якої її частини, яка включає домен, що зв'язує ламінін і/або фібронектин і/або C3 так, що він зберігає відповідну зв'язувальну властивість. Таким чином, "ліганд" включає молекули, які складаються тільки із домену, що зв'язує ламінін і/або фібронектин і/або C3, тобто області або областей пептиду, необхідної для зв'язування. Для цілей даного винаходу властивості поліпептиду зв'язувати ламінін, фібронектин або C3 можна встановити таким чином. Поліпептиди можна помітити йодом або іншими радіоактивними сполуками і перевірити на зв'язування в радіоіму 95456 14 ноаналізах (РІА) у вигляді рідкої або твердої фази (наприклад, дот-блотів). Крім того, поліпептиди можна проаналізувати на зв'язування за допомогою твердофазних імуноферментних аналізів (ELISA) або проточної цитометрії з використанням відповідних антитіл і систем виявлення. Взаємодії між поліпептидами і ламініном, фібронектином або C3 можна, крім того, дослідити за допомогою резонансу поверхневих плазмонів (Віаcore). Приклади методів детально наведені в розділі "Матеріали і методи". В іншому переважному варіанті здійснення даного винаходу поліпептид [або зрізаний поліпептид в порівнянні з поліпептидом дикого типу] включає принаймні одну з консервативних послідовностей з SEQ ID NO: 1-10, які ідентифіковані при продемонстрованому тут вирівнюванні, або складається з принаймні однієї такої консервативної послідовності. Отже, в цьому варіанті здійснення даного винаходу поліпептид [або поліпептид, зрізаний в порівнянні з поліпептидом дикого типу] включає або складається з принаймні однієї з наступних послідовностей: З UspA1 (консервативних фрагментів домену, що зв'язує фібронектин - '/' відділяє альтернативні вибори амінокислоти в будь-якому положенні) 15 95456 16 3 UspA2 (консервативних фрагментів домену, що зв'язує фібронектин - '/' відділяє альтернативні вибори амінокислоти в будь-якому положенні) З UspA2 (консервативних фрагментів домену, що зв'язує C3 - '/' відділяє альтернативні вибори амінокислоти в будь-якому положенні) Зрозуміло, що ліганди даного винаходу, що є поліпептидами, можуть включати домен, що зв'язує ламінін і/або фібронектин і/або C3 послідовності, що згадується тут, модифікованої додаванням або делецією амінокислотних залишків до або з послідовності, що згадується тут з одного з двох або з обох N- і С-кінців, причому модифіковані пептиди зберігають здатність зв'язувати ламінін і/або фібронектин і/або C3, відповідно. Відповідно даним винаходом, крім того, забезпечується ліганд, що включає або складається з поліпептиду, до якого додані або з якого делетовані 50, 40, 30, 20, 10, 5, 3 або 1 амінокислотний залишок послідовності, що згадується тут, з одного з двох або з обох Ν- і С-кінців, причому вказаний модифікований пептид зберігає здатність зв'язувати ламінін і/або фібронектин і/або C3; і/або індукувати імунну відповідь проти не модифікованого пептиду. Під подовженням розуміється подовження послідовності з використанням пептиду з повнорозмірної амінокислотної послідовності, з якої він отриманий. Що стосується фрагментів поліпептидів даного винаходу, то в даному винаході можна викори стати фрагменти будь-якого розміру (засновані на гомологічних послідовностях/консервативних областях/функціональних доменах, що обговорюються тут) при умові, що ці фрагменти зберігають здатність зв'язувати ламінін і/або фібронектин і/або C3. Може бути бажаним виділення мінімального пептиду, що містить тільки такі області, які потрібні для зв'язування з рецептором. Ліганди, що є поліпептидами відповідно до даного винаходу можна отримати з відомих білків UspA1 або UspA2 Moraxella catarrhalis за допомогою усікання одного з двох або обох N- і С-кінців. Зрізані поліпептиди не є повнорозмірними нативними молекулами UspA1 або UspA2. Відповідно даним винаходом, крім того, забезпечується послідовність UspA1 дикого типу, в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160 і т.п. до 298 амінокислот з N-кінця і/або в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160, 180, 200 і т.п. до 450 амінокислот з С-кінця. Переважно зрізані поліпептиди зберігають функцію зв'язування з фібронектином (необов'язково також зв'язування з ламініном і/або C3). 17 95456 18 Відповідно даним винаходом, крім того, забезпечується послідовність UspA2 дикого типу, в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160, 164 амінокислоти з N-кінця і/або в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 180, 200 і т.п. до 312 амінокислот з С-кінця. Переважно зрі зані поліпептиди зберігають функцію зв'язування з фібронектином (необов'язково також зв'язування з ламініном і/або C3). Можливі зрізані поліпептиди можна вибрати з зрізаних поліпептидів, представлених в наступній таблиці, всі з яких знаходяться в межах об'єму даного винаходу. Відповідно даним винаходом, крім того, забезпечується послідовність UspA2 дикого типу, в якій відсутні принаймні (або точно) 5, 10, 15, 20, 25 або 29 амінокислот з N-кінця і/або в якій відсутні принаймні (або точно) 20, 30, 40, 50,60, 70, 80, 100, 120, 140, 160, 180, 200 і т.п. до 453 амінокислот з С-кінця. Переважно зрізані поліпептиди зберігають функцію зв'язування з ламініном (необов'язково також зв'язування з фібронектином і/або C3). Можливі зрізані поліпептиди можна вибрати з зрізаних поліпептидів, представлених в наступній таблиці, всі з яких знаходяться в межах об'єму даного винаходу. 19 Відповідно даним винаходом, крім того, забезпечується послідовність UspA2 дикого типу, в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160 і т.п. до 301 амінокислоти з N-кінця і/або в якій відсутні принаймні (або точно) 20, 30, 40, 50, 60, 70, 80, 100, 120, 140, 160 або 172 амінокислоти з С-кінця. Пере 95456 20 важно зрізані поліпептиди зберігають функцію зв'язування з C3 (необов'язково також зв'язування з фібронектином і/або ламініном). Можливі зрізані поліпептиди можна вибрати з зрізаних поліпептидів, представлених в наступній таблиці, всі з яких знаходяться в межах об'єму даного винаходу. 21 Відомі послідовності UspA1 дикого типу, які можна піддати усіканню таким чином, являють собою послідовності штамів АТСС2538 (МХ2; номер доступу в GenBank - AAD43495), Р44 (AAN84895), О35Е (ААВ96359), ТТА37 (AAF40122), О12Е (AAF40118), О46Е (AAF36416), VI171 (AAD43469), ТТА24 (AAD43467) (дивись таблицю 1/Фігура 19) або ВС5 (дивись вище). Відомі послідовності UspA2 дикого типу, які можна піддати усіканню таким чином, являють собою послідовності штамів 03 5Е (номер доступу в GenBank - О4407), МС317 (номер доступу в GenBank - Q58XP4), Е22 (номер доступу в GenBank - Q848S1), V1122 (номер доступу в GenBank - Q848S2), Р44 (номер доступу в GenBank - Q8GH86), ТТА37 (номер доступу в GenBank - Q9L961), О46Е (номер доступу в GenBank -Q9L962), О12Е (номер доступу в GenBank - Q9L963), VI171 (номер доступу в GenBank - Q9XD51), ТТА24 (номер доступу в GenBank - Q9XD53), SP12-5 (номер доступу в 95456 22 GenBank - Q8RTB2), АТСС25238 (номер доступу в GenBank - Q9XD55) (дивись таблицю 2/Фігура 20) або ВС5 (Forsgren_UspA2) (дивись вище). Ідеально, якщо зрізаний поліпептид UspA1 або UspA2 цього варіанту здійснення даного винаходу включає або складається з амінокислотної послідовності, вибраної з групи, яка складається з SEQ ID NO: 1-10, або її фрагмента, гомолога, функціонального еквівалента, похідного, дегенеративного продукту або продукту гідроксилювання, сульфонування або глікозилювання або іншого вторинного процесованого продукту; або включає або складається з принаймні однієї з консервативних послідовностей всередині цих областей, які ідентифіковані при продемонстрованому тут вирівнюванні, наприклад: З UspA1 (консервативних фрагментів домену, що зв'язує фібронектин - V відділяє альтернативні вибори амінокислоти в якому-небудь положенні) 23 З UspA2 (консервативних фрагментів домену, що зв'язує фібронектин - V відділяє альтернатив 95456 24 ні вибори амінокислоти в якому-небудь положенні) 3 UspA2 (консервативних фрагментів домену, що зв'язує C3 - V відділяє альтернативні вибори амінокислоти в якому-небудь положенні) Може бути прийнятним отримання злитих білків, що містять ліганди, що є поліпептидами, описані тут. Відповідно в додатковому варіанті здійснення даним винаходом забезпечуються злиті білки, що включають ліганди, що є поліпептидами відповідно до даного винаходу. Переважно злитий білок відповідно до цього варіанту здійснення даного винаходу ідентичний менш ніж на 50% будь-якій відомій повнорозмірній послідовності протягом повної довжини. Такі злиті білки можуть складати похідне поліпептидів даного винаходу. Крім того, в похідних можуть використовуватися поліпептиди даного винаходу як носій для ковалентного приєднання пептидних або сахаридних складових. Їх можна з'єднати, наприклад, з пневмококовими капсульними олігосахаридами або полісахаридами або ліпоолігосахаридами Moraxella catarrhalis або ліпоолігосахаридами Haemophilus influenzae, що не типуються. Гомологічні пептиди даного винаходу можна ідентифікувати за допомогою порівняння послідовностей. Переважно гомологічні пептиди ідентичні на принаймні 60%, більш переважно на принаймні 70%, 80%, 90%, 95% або 99%, в порядку наростання переваги, розкритій тут пептидній послідовності або її фрагментам або зрізаним поліпептидам даного винаходу протягом їх повної довжини. Переважно гомологічний пептид зберігає здатність зв'язувати фібронектин і/або ламінін і/або C3; і/або індукувати імунну відповідь проти розкритих тут пептидних послідовностей або їх фрагментів. На Фіг. 19 і 20 показано вирівнювання пептидних послідовностей UspA1 і UspA2 різного походження, при яких вказані області послідовності, які можна модифікувати з утвореннямгомологічних послідовностей при збереженні функції (тобто здатності зв'язувати фібронектин і/або ламінін і/або C3). Пептидами, гомологічними пептидам SEQ ID NO: 1-10 ВС5, є, наприклад, такі послідовності, які відповідають послідовності ВС5, з інших штамів, поданих на Фіг. 19 і 20. Вакцини даного винаходу Поліпептиди/пептиди/функціональні домени/гомологи/фрагменти/зрізані поліпептиди/похідні даного винаходу в ідеалі готують у вигляді вакцини, що містить ефективну кількість вказаного(их) компонента(ів) і фармацевтично прийнятну допоміжну речовину. Вакцини даного винаходу можна використовувати для призначення пацієнту для профілактики або лікування інфекцій, викликаних Moraxella catarrhalis, або запалення середнього вуха, або синуситу, або інфекцій нижніх дихальних шляхів, їх можна вводити будь-яким відомим способом, в тому числі внутрішньом'язово, парентерально, на слизові оболонки і інтраназально. Комбіновані вакцини даного винаходу Вакцини даного винаходу можна скомбінувати з іншими антигенами Moraxella catarrhalis для профілактики або лікування відмічених вище захворювань. Автори даного винаходу, зокрема, виявили, що Moraxella catarrhalis має принаймні два способи гальмування імунної системи хазяїна від наступу на організм. У доповненні до взаємодії з C3 (і С4ВР), що згадується в наведених нижче прикладах, М. catarrhalis має сильну спорідненість до розчинного і зв'язаного з мембраною IgD людини завдяки білку MID (також відомому як ОМР106). Moraxella-залежне зв'язування IgD з Влімфоцитами приводить до синтезу поліклональних імуноглобулінів, який може перешкоджати продукції специфічних моноклональних антитіл проти Moraxella. Той факт, що М. catarrhalis гальмує імунну систему людини декількома способами, може служити поясненням того, чому М. catarrhalis є таким частим мешканцем дихальних шляхів. Автори даного винаходу вважають, що комбінація антигенів, залучених до функції зв'язування IgD (MID) і функції зв'язування C3 (UspA1 і/або UspA2), може забезпечити імуногенну композицію, що збільшує захисні здатності хазяїна проти гальмування бактеріями роду Moraxella імунної системи людини, забезпечуючи таким чином по 25 силене зниження носійства М. catarrhalis на поверхнях слизових оболонок. Отже, подальшим аспектом даного винаходу є вакцинна композиція, що містить ефективну кількість UspA1 і/або UspA2 (особливо останнього) (наприклад повнорозмірних поліпептидів або поліпептидів/пептидів/функціональних доменів/гомологів/фрагментів/зрізаних поліпептидів/похідних даного винаходу, описаних тут, які переважно зберігають функцію зв'язування C3) в комбінації з ефективною кількістю білка MID (наприклад повнорозмірних поліпептидів або їх поліпептидів/ пептидів/ функціональних доменів/гомологів/фрагментів/зрізаних поліпептидів/похідних, які переважно зберігають функцію зв'язування IgD людини) і фармацевтично прийнятну допоміжну речовину. Білок MID і його гомологи/фрагменти/зрізані поліпептиди, що зв'язують IgD, описані в заявці WO 03/004651 (включеній сюди за допомогою посилання). Особливо прийнятним для цієї мети фрагментом є поліпептид, що включає (або що складається з) Р2-фрагмент, описаний в заявці WO 03/004651, або послідовності, ідентичні з ним на 60, 70, 80, 90, 95, 99%, які переважно зберігають активність зв'язування IgD людини. Компоненти MID і UspA цієї комбінованої вакцини можуть бути відділені один від одного, або вони можуть бути відповідним чином злиті разом з допомогою відомих методів молекулярної біології. Короткий опис креслень На Фіг. 1 демонструються тринадцять штамів М. catarrhalis, перевірених на зв'язування з фібронектином (А). Сильне зв'язування фібронектину корелювало з експресією UspA1/A2, визначеною з допомогою рАb проти UspA1/A2 (B-I). Профілі проточної цитометрії М. catarrhalis BBH18 дикого типу і дефіцитних у відношенні UspA1/A2 мутантів демонструють залежне від UspA1/A2 зв'язування з розчинним фібронектином. Представлені профілі клінічного ізоляту дикого типу (В і F) і відповідних мутантів, позбавлених UspA1 (С і G) або UspA2 (D і Н), і подвійних мутантів (Е і І), у яких відсутній і UspA1, і UspA2. Бактерії інкубували з кролячими антитілами проти UspA1/A2 або фібронектином, а потім з рАb проти фібронектину. Згодом додавали кролячі FITC-кон'юговані рАb з подальшим аналізом проточною цитометрією. Продемонстрований типовий експеримент з трьох із середньою інтенсивністю флуоресценції (MFI) для кожного профілю. На Фіг. 2 демонструється, що дефіцитні у відношенні UspA2 мутанти М. catarrhalis RH4 не 125 зв'язують І-мічений фібронектин. Як негативний контроль, що не зв'язує фібронектин, вклюІ25 чили Е. coli BL21. Бактерії інкубували з Іміченим фібронектином з подальшими декількома промиваннями і аналізували в гаммалічильнику. За 100% прийняли зв'язування фібронектину з RH4 дикого типу, що експресує і UspA1, і А2. Показані середні значення трьох незалежних експериментів. Планки погрішностей представляють стандартні відхилення (SD). Схо 95456 26 жі результати були отримані з М. catarrhalis BBH18. На Фіг. 3 демонструються фотографії, які підтверджують, що мутанти М. catarrhalis, позбавлені UspA1 і UspA2, не зв'язуються з іммобілізованим фібронектином. М. catarrhalis дикого типу міг адгезуватися до покритого фібронектином предметного скла з високою густиною (А). Мутант М. catarrhalis uspA1 також зберігав високу густину адгезії (В), в той час як М. catarrhalis uspA2 і подвійні мутанти uspA1/A2 слабко адгезувалися (С і D). Предметне скло покривали фібронектином і інкубували з М. catarrhalis RH4 і його відповідними мутантами UspA1/A2. Після декількох промивань бактерії забарвлювали за Грамом. Фіг. 4 являє собою графік, на якому демонструється, що рекомбінантні UspA1 і А2 зв'язуються з фібронектином залежним від дози спосо50-770 30-539 бом. Для UspA1 і UspA2 показане специфічне зв'язування фібронектину. Обома білками UpsA (40 нМ) покривали мікротитраційні планшети і інкубували із зростаючими концентраціями фібронектину, з подальшим виявленням з допомогою кролячих рАb проти фібронектину людини і HRP-кон'югованих антикролячих рАb. Показані середні значення трьох окремих експериментів, а планки погрішностей вказують стандартні відхилення (SD). Фіг. 5. Активні домени UspA1 і UspA2, що зв'язують фібронектин знаходяться між амінокислотами 299-452 і 165-318, відповідно. Показані зрізані білки, отримані з UspA1 (А) і UspA2 (В). Всі фрагменти перевіряли на зв'язування з фібронектином в ELISA. Сорок нМ кожного зрізаного фрагмента покривали мікротитраційні планшети і інкубували з 80 мкг/мл і 120 мкг/мл фібронектину для UspA1 і UspA2, відповідно. Зв'язаний фібронектин виявляли з допомогою кролячих рАb проти фібронектину, а потім HRP-кон'югованих антикролячих рАb. Показані результати трьох наборів експериментів. Планки погрішностей представляють стандартні відхилення (SD). На Фіг. 6 демонструється послідовність відповідно до послідовності ID NO:1 і гомологія пос299-452 165-318 лідовностей між UspA1 і UspA2 . У дужках знаходяться 31 ідентичних амінокислотних залишків. На Фіг. 7 демонструється, що зрізані фрагме50-491 299-452 нти UspA1 і UspA1 конкурентно інгібують залежне від UpsA зв'язування М. catarrhalis з фібронектином. Як негативні контролі включали подвійні мутанти (uspA1/A2 M catarrhalis, які не зв'язують фібронектин. Рекомбінантні білки UspA1 заздалегідь інкубували з 2 мг/100 мл фібронектину до інкубації з М. catarrhalis. Представлені середні значення флуоресценції (MFI) М. catarrhalis зі зв'язаним фібронектином, що виявляється з допомогою FITC-кон'югованих рАb проти фібронектину в аналізі проточною цитометрі50-491 299-452 єю. UspA1 і UspA1 приводили до інгібування на 95% і 63%, відповідно. Планки погрішностей представляють середні значення ± стандартні відхилення (SD) трьох незалежних експериментів. 27 95456 299-452 На Фіг. 8 демонструється, що UspA1 і 165-318 UspA2 інгібують адгезію М. catarrhalis до клітин кон'юнктиви Чанг через асоційований з клітинами фібронектин. Епітеліальні клітини Чанг експресують фібронектин на поверхні, що виявлено з допомогою рАb проти фібронектину і проточної цитометрії (А). Попередня інкубація із біл299-452 ками, що зв'язують фібронектин, UspA1 , 165-318 UspA2 або рАb проти фібронектину приводила до значного зменшення зв'язування М. catarrhalis RH4 в порівнянні з контрольними ре433-580 30-177) комбінантними білками (UspA1 і UspA2 і контрольним антитілом (мАb проти ІСАМ1) (В). Р
ДивитисяДодаткова інформація
Назва патенту англійськоюInteraction of moraxella catarrhalis with epithelial cells, extracellular matrix proteins and the complement system
Автори англійськоюForsgren Arne, Forsgren, Arne, Riesbeck Kristian
Назва патенту російськоюВзаимодействие moraxella catarrhalis с эпителиальными клетками, внеклеточными матриксными белками и системой комплемента
Автори російськоюФорсгрен Арне, Риесбекк Кристиан
МПК / Мітки
МПК: C07K 14/21, A61K 39/104, A61P 31/04
Мітки: білками, комплементу, позаклітинними, клітинами, moraxella, матриксними, catarrhalis, взаємодія, системою, епітеліальними
Код посилання
<a href="https://ua.patents.su/72-95456-vzaehmodiya-moraxella-catarrhalis-z-epitelialnimi-klitinami-pozaklitinnimi-matriksnimi-bilkami-i-sistemoyu-komplementu.html" target="_blank" rel="follow" title="База патентів України">Взаємодія moraxella catarrhalis з епітеліальними клітинами, позаклітинними матриксними білками і системою комплементу</a>
Виділений поліпептид омр106 зовнішньої мембрани moraxella catarrhalis, його пептидний фрагмент та молекули днк, що їх кодують, виділене антитіло, вакцина (варіанти) та антигенна композиція (варіанти), а також с

Номер патенту: 68332
Опубліковано: 16.08.2004
Автори: Такер Кеннет, Плосіла Лаура
МПК: C12Q 1/68, C12R 1/36, C07K 14/21, A61P 31/04, G01N 33/53, C07K 16/12, A61K 39/095, C12N 15/31
Мітки: омр106, молекули, moraxella, днк, зовнішньої, поліпептид, антитіло, також, фрагмент, виділене, пептидний, мембрани, композиція, кодують, антигенна, вакцина, catarrhalis, варіанти, виділений
Формула / Реферат:
1. Выделенный полипептид ОМР106, который является полипептидом наружной мембраны Moraxella catarrhalis и имеет молекулярную массу приблизительно от 180 кДа до приблизительно 230 кДа, как определено электрофорезом в SDS-полиакриламидном геле с использованием миозина скелетных мышц кролика и -галактозидазы Е.соlі в качестве стандартов молекулярных масс 200 кДа и 116,25 кДа,...
Спосіб моделювання дозованої дії цитостатику колхіцину на щурів різного віку та маси тіла для блокування проходження мітотичного циклу епітеліальними клітинами на стадії мітозу

Номер патенту: 9388
Опубліковано: 15.09.2005
Автор: Смірнов Сергій Миколайович
МПК: A61K 31/165
Мітки: стадії, циклу, щурів, віку, дії, мітозу, цитостатику, мітотичного, спосіб, епітеліальними, блокування, маси, моделювання, різного, колхіцину, тіла, клітинами, проходження, дозованої
Формула / Реферат:
1. Спосіб моделювання дозованої дії цитостатику колхіцину на щурів різного віку та маси тіла для блокування проходження мітотичного циклу епітеліальними клітинами на стадії мітозу, що включає зменшення дози препарату при використанні експериментальних щурів перших трьох днів життя, які є особливо чутливими до токсичного впливу колхіцину.2. Спосіб за п. 1, який відрізняється тим, що доза колхіцину знижується з 2,5 мг/кг до 1,5 мг/кг...
Спосіб одержання лістеріозної сироватки іеквм для реакції зв’язування комплементу

Номер патенту: 8749
Опубліковано: 15.08.2005
Автори: Сербиненко Тетяна Миколаївна, Соловйов Сергій Тихонович, Стегній Борис Тимофійович
МПК: A61K 39/02
Мітки: іеквм, зв'язування, комплементу, спосіб, сироватки, реакції, лістеріозної, одержання
Формула / Реферат:
Спосіб одержання сироватки, що включає вирощування бактерійної маси на живильному середовищі, виготовлення антигену, гіперімунізацію продуцентів одержаним антигеном з наступним забором крові та відокремленням цільного продукту, який відрізняється тим, що використовують як бактерійну масу штами лістерій "Буринь", "Тернопіль", "Кріль № 17", "№ 1".
Дендример-пег з чотирма гілками для кон’югування з білками і пептидами
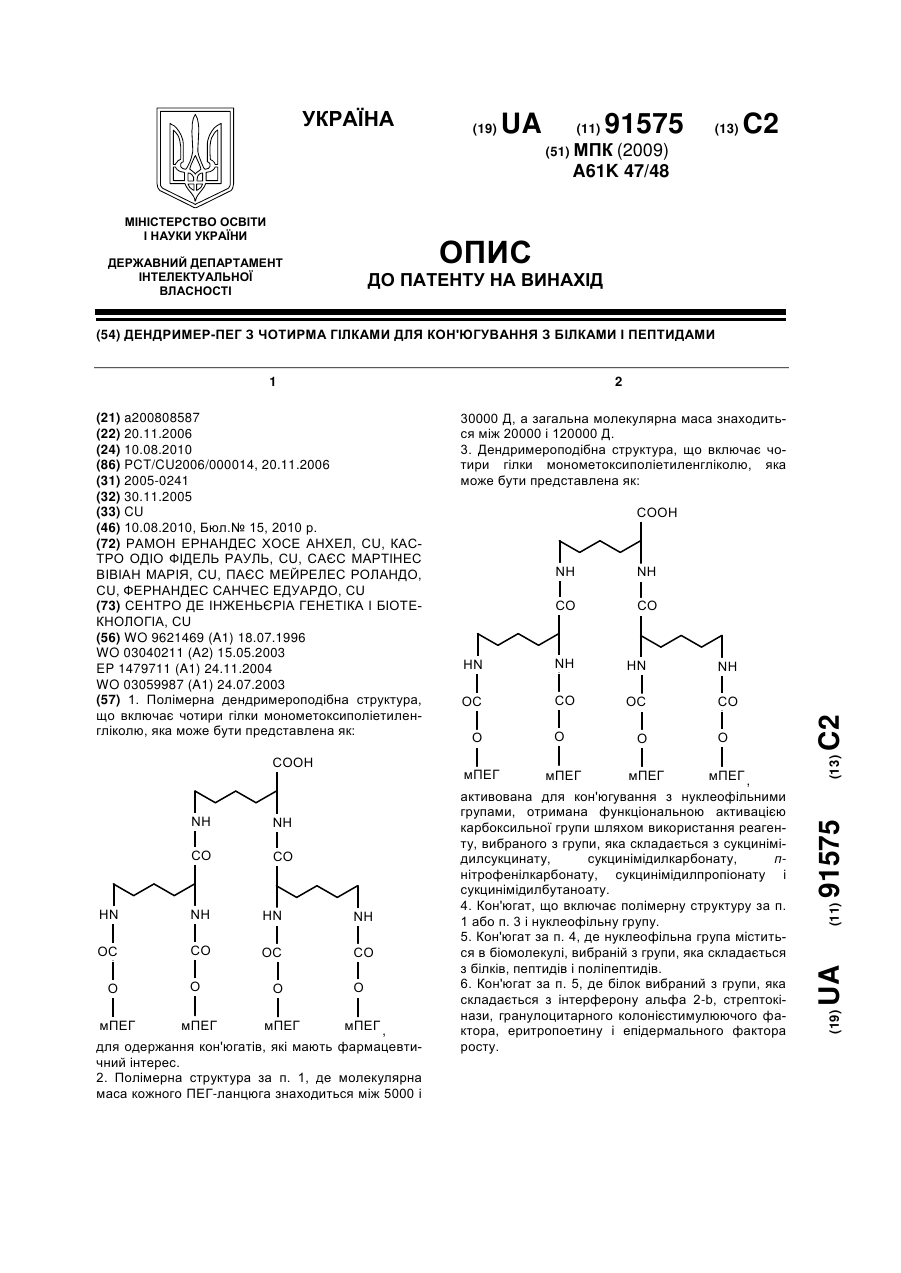
Номер патенту: 91575
Опубліковано: 10.08.2010
Автори: Кастро Одіо Фідель Рауль, Паєс Мейрелес Роландо, Рамон Ернандес Хосе Анхел, Фернандес Санчес Едуардо, Саєс Мартінес Вівіан Марія
МПК: A61K 47/48
Мітки: пептидами, дендример-пег, білками, чотирма, кон'югування, гілками
Формула / Реферат:
1. Полімерна дендримероподібна структура, що включає чотири гілки монометоксиполіетиленгліколю, яка може бути представлена як:,для одержання кон'югатів, які мають фармацевтичний інтерес.2. Полімерна структура за п. 1, де молекулярна маса кожного ПЕГ-ланцюга знаходиться між 5000 і 30000 Д, а загальна молекулярна маса знаходиться між 20000 і 120000...
Сироватка гемолітична для реакції зв’язування комплементу

Номер патенту: 58697
Опубліковано: 15.08.2003
Автори: Стегній Борис Тимофійович, Галіщев Микола Гнатович, Вовк Сергій Іванович, Бабкін Анатолій Федорович
МПК: A61K 39/00
Мітки: гемолітична, комплементу, сироватка, реакції, зв'язування
Формула / Реферат:
Сироватка гемолітична для реакції зв’язування комплементу, що одержана шляхом гіперімунізації кролів суспензією еритроцитів баранів з наступною консервацією її, яка відрізняється тим, що сироватку після одержання стабілізують магнієм сульфату у кінцевій концентрації 4%, консервують розчином фенолу (С6Н5OН) та хімічно чистим гліцерином (НОСН2СН/ОН-СН2СН) при наступному співвідношенні компонентів, мас. %: фенол ...
Попередній патент: Макроциклічні інгібітори реплікації вірусу гепатиту с
Наступний патент: Антиміостатинове моноклональне антитіло
Випадковий патент: Спосіб визначення електрокінетичного потенціалу пухирців газу в рідині