Спосіб забезпечення резистентності до грибкового або грибкоподібного патогену у однодольних рослинах
Номер патенту: 84406
Опубліковано: 27.10.2008
Автори: Когель Карл-Хайнц, Труїлло Марко, Хюккельховен Ральф





Формула / Реферат
1. Спосіб одержання або підвищення стійкості до принаймні одного грибкового або грибоподібного патогену в однодольних рослинах, який відрізняється тим, що він включає наступні стадії:
a) зниження кількості, активності або функції NADPH-оксидази в рослині або в її тканині, органі, частині або клітині, і
b) вибір рослин, які, на відміну або у порівнянні з вихідною рослиною, мають або у яких підвищена стійкість до принаймні одного грибкового або грибкоподібного патогену,
де NADPH-оксидаза кодується
a) поліпептидними послідовностями, що включають SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22, або
b) поліпептидними послідовностями, що мають гомологію принаймні у 50 % з поліпептидом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22.
2. Спосіб за п. 1, де зниження кількості білка, активності або функції NADPH-оксидази здійснюють при застосуванні одного зі способів, вибраних із групи, що включає:
a) введення послідовності дволанцюгової рибонуклеїнової кислоти NADPH-оксидази або експресійної касети, або експресійних касет, що забезпечують її експресію;
b) введення антисмислової послідовності нуклеїнової кислоти NADPH-оксидази або експресійної касети, що забезпечує їх експресію ;
c) введення антисмислової послідовності нуклеїнової кислоти NADPH-оксидази, комбінованої з рибозимом, або експресійної касети, що забезпечує її експресію;
d) введення смислових послідовностей нуклеїнової кислоти NADPH-оксидази для індукції косупресії або експресійної касети, що забезпечує їх експресію;
е) введення ДНК- або білокзв'язувальних факторів проти генів, РНК або білків NADPH‑оксидази або експресійної касети, що забезпечує їх експресію;
f) введення вірусних послідовностей нуклеїнової кислоти, що викликають деструкцію РНК та експресійних конструктів NADPH-оксидази, або експресійної касети, що забезпечує їх експресію;
g) введення конструктів для індукції гомологічної рекомбінації в ендогенних генах NADPH-оксидази, і
h) введення мутацій в ендогенний ген NADPH-оксидази.
3. Спосіб за п. 1 або 2, що включає
(i) стабільну трансформацію рослинної клітини за допомогою рекомбінантної експресійної касети, що містить у функціональному зв'язку з активним у рослині промотором послідовність нуклеїнової кислоти, яка кодує
a) дволанцюгову послідовність рибонуклеїнової кислоти NADPH-оксидази або
b) антисмислову послідовність нуклеїнової кислоти NADPH-оксидази, або
c) антисмислову послідовність нуклеїнової кислоти NADPH-оксидази, комбіновану з рибозимом, або
d) смислову послідовність нуклеїнової кислоти NADPH-оксидази для індукції косупресії, або
e) ДНК- або білокзв'язувальні фактори проти генів, РНК або білків NADPH-оксидази,
f) вірусні послідовності нуклеїнових кислот, що викликають деструкцію РНК NADPH-оксидази,
де NADPH-оксидаза кодується
a) поліпептидними послідовностями, що включають SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22, або
b) поліпептидними послідовностями, що мають гомологію принаймні у 50 % з поліпептидом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22,
(ii) регенерацію рослин з рослинної клітини і
(iii) експресію зазначеної послідовності нуклеїнової кислоти у кількості та протягом часу, достатніх для одержання або підвищення стійкості до патогену в зазначеній рослині.
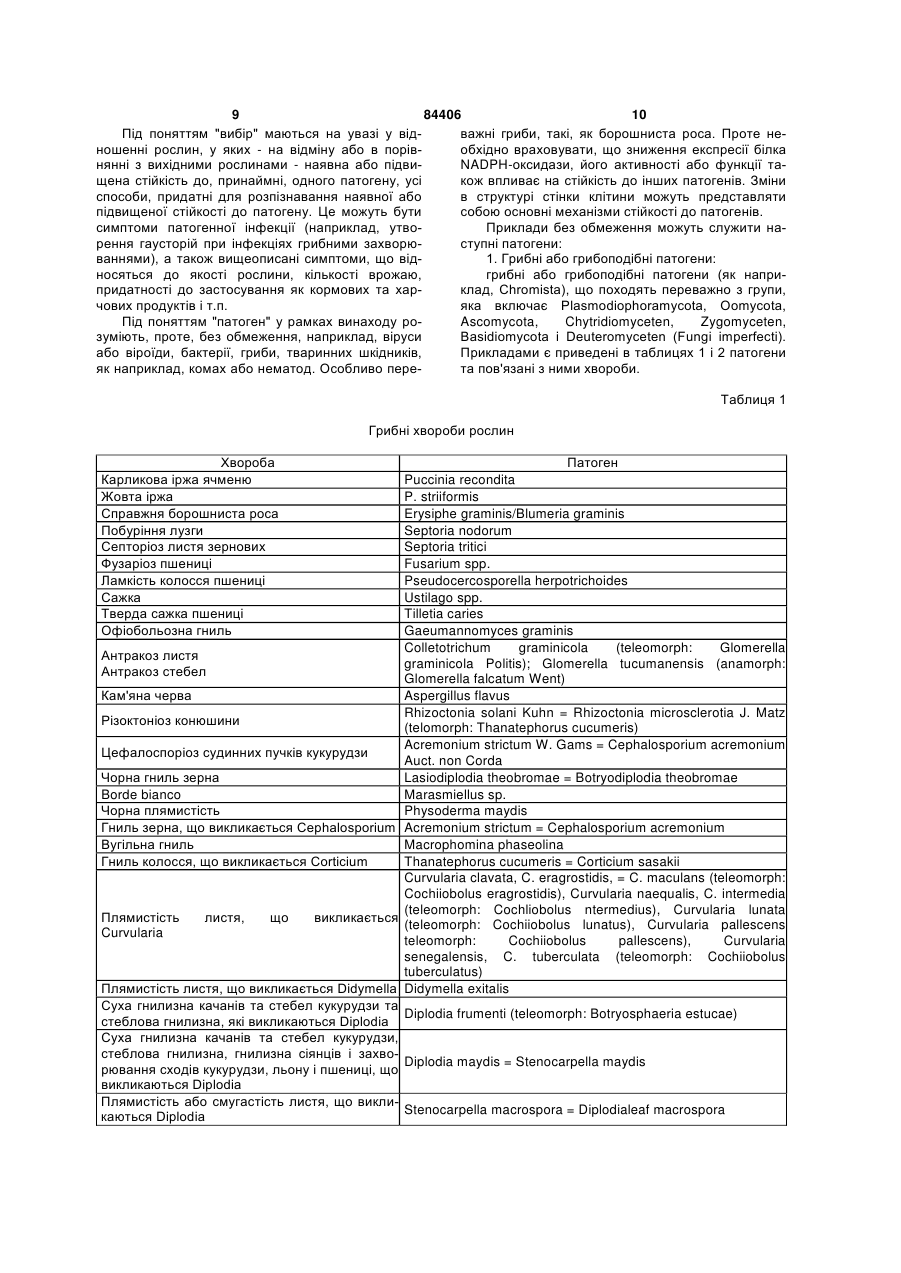
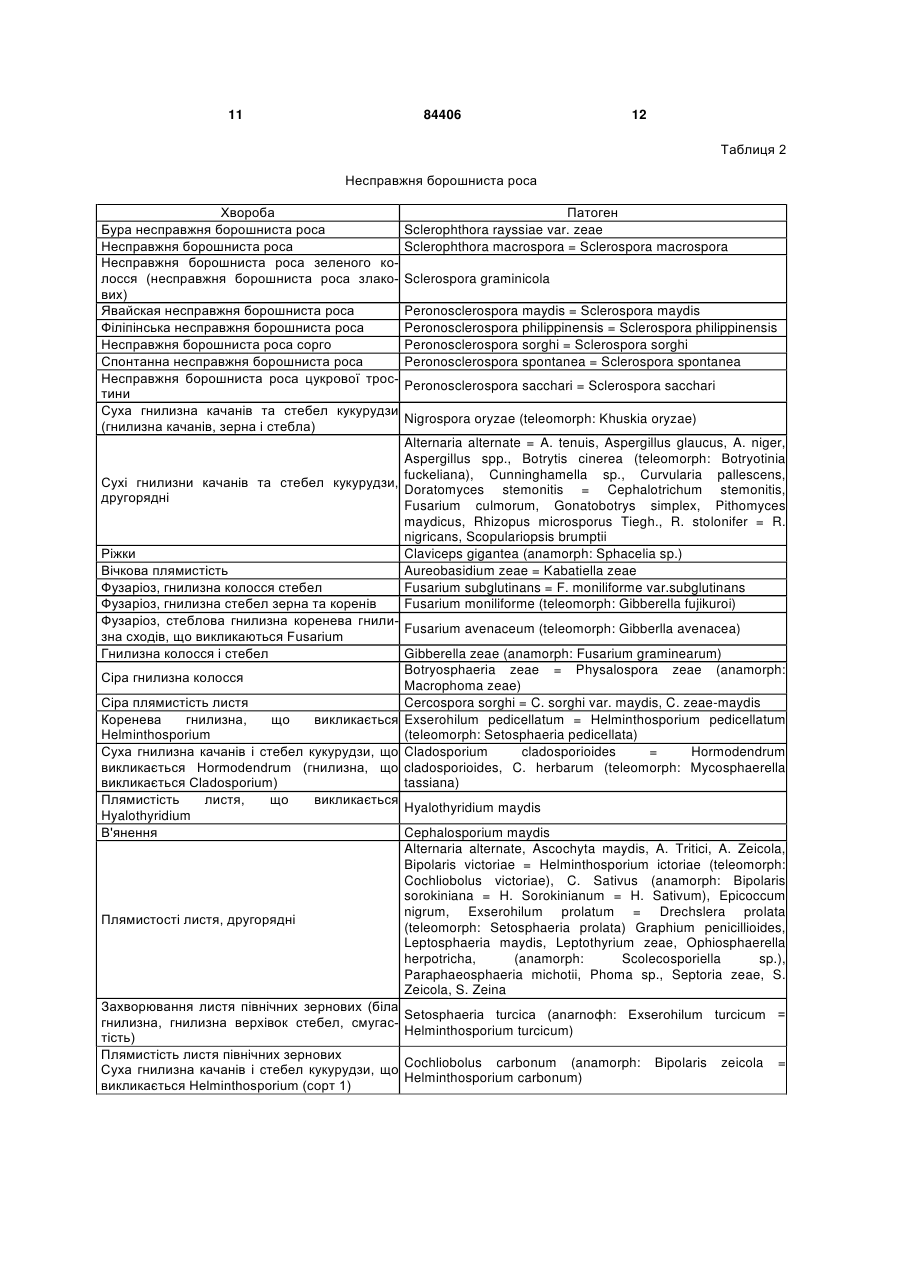
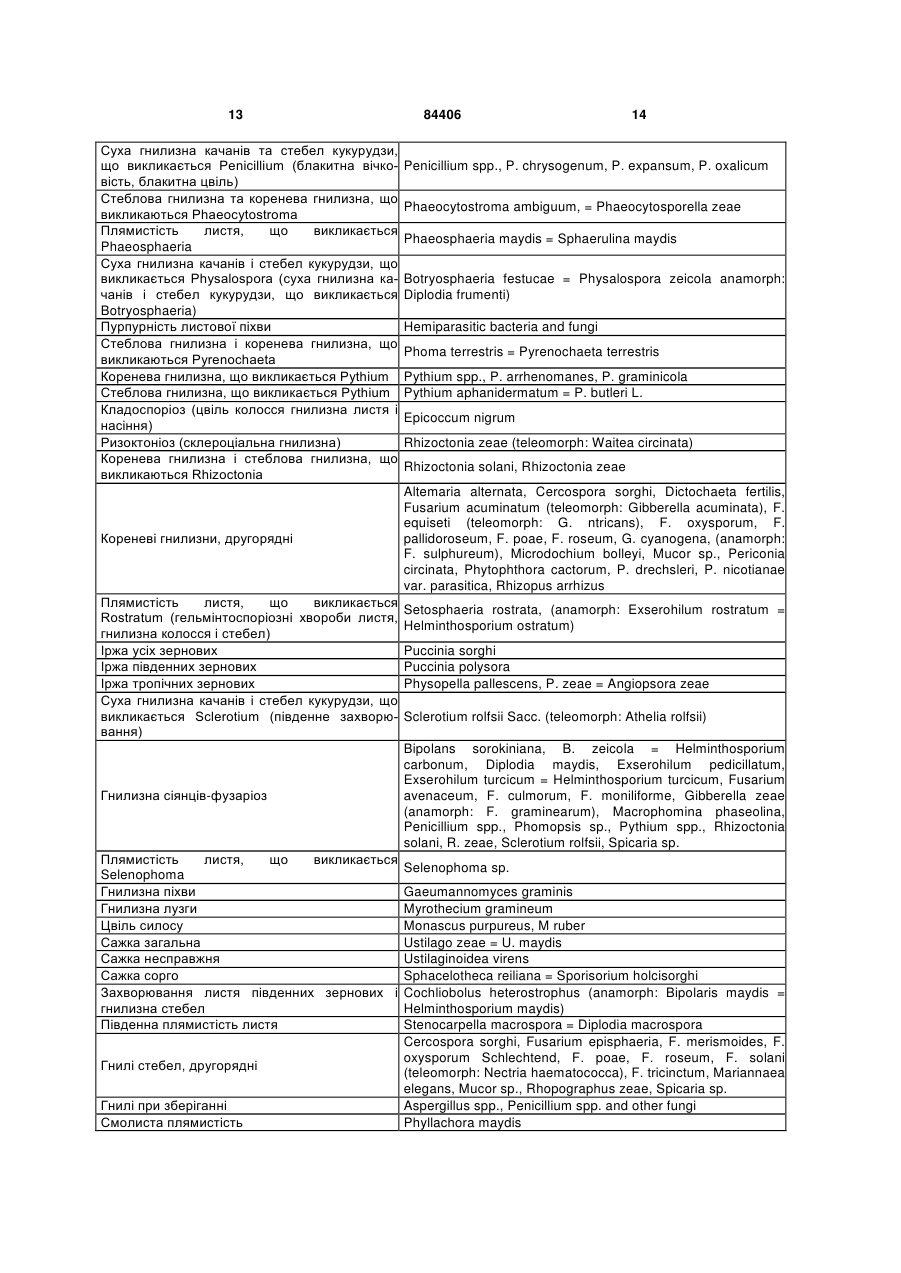
4. Спосіб за одним з пп. 1-3, причому патоген вибраний із групи грибків, що включає Plasmodiophoramycota, Oomycota, Ascomycota, Chytridiomycetes, Zygomycetes, Basidiomycota і Deuteromycetes.
5. Спосіб за одним з пп. 1-4, причому рослина вибрана з групи однодольних рослин, що включає пшеницю, овес, просо, ячмінь, жито, кукурудзу, рис, сорго, тритикале, полбу та цукрову тростину.
6. Молекула дволанцюгової РНК для зниження експресії NADPH-оксидази, яка включає
a) "смисловий" ланцюг РНК, що включає принаймні одну послідовність рибонуклеїнової кислоти, що в основному ідентична принаймні частині "смислового" РНК-транскрипта послідовності нуклеїнової кислоти, яка кодує NADPH-оксидазу, і
b) "антисмисловий" РНК-ланцюг, що в основному комплементарний смисловому РНК-ланцюгу відповідно до пункту а),
причому один з двох ланцюгів РНК кодується принаймні частиною послідовності нуклеїнової кислоти, що кодує послідовність NADPH-оксидази згідно з SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 або 21, або кодується поліпептидною послідовністю, що має гомологію принаймні у 50 % з поліпептидом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22.
7. Дволанцюгова молекула РНК за п. 6, причому обидва ланцюги дволанцюгової РНК ковалентно зв'язані один з одним.
8. Трансгенна експресійна касета, що містить у функціональному зв'язку з функціональним у рослинних організмах промотором послідовність нуклеїнової кислоти, яка кодує дволанцюгову молекулу РНК відповідно до п. 6 або 7.
9. Трансгенна експресійна касета за п. 8, причому функціональним у рослинах промотором є промотор, що індукується патогеном.
10. Трансгенна експресійна касета, що містить принаймні частину послідовності нуклеїнової кислоти, яка кодує NADPH-оксидазу відповідно до SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 або 21, або кодує поліпептидну послідовність, що має гомологію принаймні у 50 % з поліпептидом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22, причому зазначена послідовність нуклеїнової кислоти функціонально зв'язана в антисмисловій орієнтації з функціональним у рослинних організмах промотором.
11. Трансгенна експресійна касета за п. 10, причому функціональним у рослинах промотором є промотор, що індукується патогеном.
Текст
1. Спосіб одержання або підвищення стійкості до принаймні одного грибкового або грибоподібного патогену в однодольних рослинах, який відрізняється тим, що він включає наступні стадії: a) зниження кількості, активності або функції NADPH-оксидази в рослині або в її тканині, органі, частині або клітині, і b) вибір рослин, які, на відміну або у порівнянні з вихідною рослиною, мають або у яких підвищена стійкість до принаймні одного грибкового або грибкоподібного патогену, де NADPH-оксидаза кодується a) поліпептидними послідовностями, що включають SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22, або b) поліпептидними послідовностями, що мають гомологію принаймні у 50 % з поліпептидом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22. 2. Спосіб за п. 1, де зниження кількості білка, активності або функції NADPH-оксидази здійснюють при застосуванні одного зі способів, вибраних із групи, що включає: a) введення послідовності дволанцюгової рибонуклеїнової кислоти NADPH-оксидази або експресійної касети, або експресійних касет, що забезпечують її експресію; b) введення антисмислової послідовності нуклеїнової кислоти NADPH-оксидази або експресійної касети, що забезпечує їх експресію ; c) введення антисмислової послідовності нуклеїнової кислоти NADPH-оксидази, комбінованої з рибозимом, або експресійної касети, що забезпечує її експресію; d) введення смислових послідовностей нуклеїнової кислоти NADPH-оксидази для індукції косупресії або експресійної касети, що забезпечує їх експресію; UA (21) a200501646 (22) 14.07.2003 (24) 27.10.2008 (86) PCT/EP2003/007589, 14.07.2003 (31) 102 33 327.0 (32) 22.07.2002 (33) DE (46) 27.10.2008, Бюл.№ 20, 2008 р. (72) КОГЕЛЬ КАРЛ-ХАЙНЦ, ХЮККЕЛЬХОВЕН РАЛЬФ, ТРУЇЛЛО МАРКО (73) БАСФ ПЛАНТ САЄНС ГМБХ (56) TORRES MIGUEL ANGEL ET AL: "Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response" PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES, Bd. 99, Nr. 1, 8. Januar 2002 (2002-01-08), Seiten 517-522, HUECKELHOVEN RALPH ET AL: "Tissue-specific superoxide generation at interaction sites in resistant and susceptibile near-isogenic barley lines attacked by the powdery mildew fungus (Erysiphe graminis f. sp. hordei)" MOLECULAR PLANT-MICROBE INTERACTIONS, Bd. 11, Nr. 4, April 1998 (1998-04), Seiten 292-300, SAGI MOSHE ET AL: "Superoxide production by plant homologues of the gp91phox NADPH oxidase. Modulation of activity by calcium and by tobacco mosaic virus infection" PLANT PHYSIOLOGY (ROCKVILLE), Bd. 126, Nr. 3, Juli 2001 (2001-07), Seiten 1281-1290, BOLWELL G PAUL ET AL: "The apoplastic oxidative burst in response to biotic stress in plants: A threecomponent system" JOURNAL OF EXPERIMENTAL BOTANY, Bd. 53, Nr. 372, Mai 2002 (2002-05), Seiten 1367-1376, HUECKELHOVEN R ET AL: "Functional studies on the role of reactive oxygen intermediates in the resistance of barley against powdery mildew." PLANT PROTECTION SCIENCE, Bd. 38, Nr. Special Issue 2, 2002, Seiten 458-460, BORDEN STEPHANIE ET AL: "Hydrogen peroxide plays a critical role in the defence response of tomato to Cladosporium fulvum." PHYSIOLOGICAL AND 2 (19) 1 3 84406 4 е) введення ДНК- або білокзв'язувальних факторів 5. Спосіб за одним з пп. 1-4, причому рослина випроти генів, РНК або білків NADPH-оксидази або брана з групи однодольних рослин, що включає експресійної касети, що забезпечує їх експресію; пшеницю, овес, просо, ячмінь, жито, кукурудзу, f) введення вірусних послідовностей нуклеїнової рис, сорго, тритикале, полбу та цукрову тростину. кислоти, що викликають деструкцію РНК та екс6. Молекула дволанцюгової РНК для зниження пресійних конструктів NADPH-оксидази, або ексекспресії NADPH-оксидази, яка включає пресійної касети, що забезпечує їх експресію; a) "смисловий" ланцюг РНК, що включає принаймg) введення конструктів для індукції гомологічної ні одну послідовність рибонуклеїнової кислоти, що рекомбінації в ендогенних генах NADPH-оксидази, в основному ідентична принаймні частині "смислоі вого" РНК-транскрипта послідовності нуклеїнової h) введення мутацій в ендогенний ген NADPHкислоти, яка кодує NADPH-оксидазу, і оксидази. b) "антисмисловий" РНК-ланцюг, що в основному 3. Спосіб за п. 1 або 2, що включає комплементарний смисловому РНК-ланцюгу від(i) стабільну трансформацію рослинної клітини за повідно до пункту а), допомогою рекомбінантної експресійної касети, що причому один з двох ланцюгів РНК кодується примістить у функціональному зв'язку з активним у наймні частиною послідовності нуклеїнової кислорослині промотором послідовність нуклеїнової ти, що кодує послідовність NADPH-оксидази згідно кислоти, яка кодує з SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 або a) дволанцюгову послідовність рибонуклеїнової 21, або кодується поліпептидною послідовністю, кислоти NADPH-оксидази або що має гомологію принаймні у 50 % з поліпептиb) антисмислову послідовність нуклеїнової кислоти дом відповідно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, NADPH-оксидази, або 14, 16, 18, 20 або 22. c) антисмислову послідовність нуклеїнової кислоти 7. Дволанцюгова молекула РНК за п. 6, причому NADPH-оксидази, комбіновану з рибозимом, або обидва ланцюги дволанцюгової РНК ковалентно d) смислову послідовність нуклеїнової кислоти зв'язані один з одним. NADPH-оксидази для індукції косупресії, або 8. Трансгенна експресійна касета, що містить у e) ДНК- або білокзв'язувальні фактори проти генів, функціональному зв'язку з функціональним у росРНК або білків NADPH-оксидази, линних організмах промотором послідовність нукf) вірусні послідовності нуклеїнових кислот, що леїнової кислоти, яка кодує дволанцюгову молекувикликають деструкцію РНК NADPH-оксидази, лу РНК відповідно до п. 6 або 7. де NADPH-оксидаза кодується 9. Трансгенна експресійна касета за п. 8, причому a) поліпептидними послідовностями, що включафункціональним у рослинах промотором є промоють SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 тор, що індукується патогеном. або 22, або 10. Трансгенна експресійна касета, що містить b) поліпептидними послідовностями, що мають принаймні частину послідовності нуклеїнової кисгомологію принаймні у 50 % з поліпептидом відполоти, яка кодує NADPH-оксидазу відповідно до відно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, SEQ ID NOs: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 або 21, 20 або 22, або кодує поліпептидну послідовність, що має го(ii) регенерацію рослин з рослинної клітини і мологію принаймні у 50 % з поліпептидом відпові(iii) експресію зазначеної послідовності нуклеїнової дно до SEQ ID NOs: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 кислоти у кількості та протягом часу, достатніх для або 22, причому зазначена послідовність нуклеїодержання або підвищення стійкості до патогену в нової кислоти функціонально зв'язана в антисмисзазначеній рослині. ловій орієнтації з функціональним у рослинних 4. Спосіб за одним з пп. 1-3, причому патоген виборганізмах промотором. раний із групи грибків, що включає 11. Трансгенна експресійна касета за п. 10, приPlasmodiophoramycota, Oomycota, Ascomycota, чому функціональним у рослинах промотором є Chytridiomycetes, Zygomycetes, Basidiomycota і промотор, що індукується патогеном. Deuteromycetes. Винахід відноситься до способу досягнення або підвищення стійкості до патогенів у рослин за допомогою зниження експресії, активності або функції NADPH-оксидази (НАДФнікотинамідаденіндинуклеотид фосфат). Метою біотехнологічних робіт на рослинах є одержання рослин з новими властивостями, наприклад, для підвищення сільськогосподарської продуктивності. Часто природні захисні механізми рослин від патогенів є недостатніми. Лише ураження фітопатогенними грибами приводять до втрати врожаю на багато мільярдів американських доларів. Внесення чужорідних генів з рослинних, тваринних або мікробних джерел може підсилити захист. Прикладами є захист проти ушкодження гризучими комахами шляхом експресії ендотоксинів Bacillus thuringiensis [див. публікацію Vaeck і ін. (1987) Nature 328: 33-37] або захист від ураження грибами шляхом експресії хітинази з квасолі [див. Broglie і ін. (1991) Science 254: 1194-1197]. Більшість описаних методів забезпечує стійкість тільки до окремих патогенів або до вузького спектра патогенів. Існує невелика кількість засобів, що надають рослинам стійкості до широкого спектра патогенів. Систематична набута стійкість ("systemic acquired resistance"; SAR), що є захисним механізмом при різних взаємодіях рослин і патогенів, може набуватися завдяки застосуванню ендогенних сигнальних речовин, таких, як жасмонат (JA) або саліци 5 84406 6 лова кислота (SA) [див. Ward, і ін. (1991) Plant Cell з диким типом рослин [Torres MA і ін. (2002) Proc 3: 1085-1094; Uknes, і ін. (1992) Plant Cell 4(6): 645Natl Acad Sci USA 99:517-522]. Також й інгубування 656]. Подібні ефекти можуть викликатися такими NADPH-оксидази за допомогою інгібіторів дифенісинтетичними сполуками, як 2,6леніодоніумхлориду (DPI) при застосуванні фізіодихлорізонікотинова кислота (INA) або складний логічно прийнятних концентрацій не справляло S-метиловий ефір бензо(1,2,3)тіадіазол-7ніякого впливу на розвиток патогенних грибів. [Hückelhoven R та Kogei KH (1998) Mol Plant тіокарбонової кислоти (BTH; Віоn) [див. Friedrich і Microbe Interact 11:292- 300]. Фрагмент кДНК фаін. (1996) Plant J. 10(1): 61-70; Lawton і ін. (1996) гоцитарної NADPH-оксидази з ячменю (pNAox, Plant J. 10:71-82]. Експресія високорегульованих у гомолог великої підгрупи gp91phox фагоцитарної рамках SAR білків, "пов'язаних з патогенезом" (PR) NADPH-оксидази) описаний у GenBank під реєстможе частково викликати стійкість до патогенів. раційним номером GenBank Acc.-No.: AJ251717). У ячменю вже давно був описаний Mlo-локус Даний винахід має задачу розробки нових як негативний регулятор захисту від патогенів. способів захисту від патогенів у рослинах, що заВтрата Mlo-гена обумовлює підвищену та неспебезпечують ефективний захист від якомога ширцифічну до видів стійкість до численних видів бошого спектра патогенів в якомога більшій кількості рошнистої роси [Buschges R і ін. (1997) Cell 88: різних видів рослин, таких культурних рослин, що 695-705; Jorgensen JH (1977) Euphytica 26: 55-62; переважно застосовуються у сільському господарLyngkjaer MF і ін. (1995) Plant Pathol 44: 786-790]. стві . Ця задача вирішується способом за винахоСорти ячменю з Mlo-дефіцитом, отримані класичдом. ним розведенням, вже широко застосовуються в Перший об'єкт винаходу включає спосіб одерсільському господарстві. Очевидно, внаслідок режання або підвищення стійкості до, принаймні, цесивності, незважаючи на інтенсивне вирощуодного патогену в рослинах, який відрізняється вання, ця стійкість виявила себе як надзвичайно тим, що він включає наступні стадії стабільна. Mlo-подібна стійкість в інших рослинах, a) зниження кількості, активності або функції насамперед, у зернових, описана не була. МІо-ген NADPH-оксидази в рослині або в її тканині, органі, та різні гомологи з інших видів зернових були іденчастині або клітині і тифіковані та клоновані [Büschges R і ін. (1997) b) відбір рослин, в яких - на відміну або у поріCell 88: 695-705; WO 98/04586; Schulze-Lefert P, внянні з вихідною рослиною - наявна або підвищуVogel J (2000) Trends Plant Sci. 5: 343-348]. У літеється стійкість до, принаймні, одного патогену. ратурі описані різні способи одержання стійкості Несподіваним чином виявилося, що зниження до патогенів при застосуванні цього гена [WO експресії NADPH-оксидази з ячменю (pNAox) в 98/04586; WO 00/01722; WO 99/47552]. Недоліком клітині епідермісу за допомогою специфічної до є те, що викликаний MIo механізм захисту включає послідовності PHК-інтерференції при застосуванні супутню загибель клітин листя [Wolter M і ін. (1993) дволанцюгової pNAox-длРНК (длРНК - дволанцюМоl Gen Genet 239: 122-128]. Крім того, недоліком гова PHK, dsRNA). "мовчання гена" ("Geneє те, що генотипи з Mlo-дефіцитом виявляють надSilencing") значно запобігало ураженню внаслідок чутливість стосовно гемібіотропних патогенів, таBgh-інфекції (що вимірюється за утворенням гауских, як Magnaporte grisea (M. grisea), а також торій). Такий висновок є несподіваним, оскільки Cochliobolus sativus (Bipolaris sorokiniana) [див. пов'язаному з NADPH-оксидазою вивільненню Jarosch B і ін. (1999) Моl Plant Microbe Interact 12: реакційноздатних сполук на основі кисню ("спалах 508-514; Kumar J і ін. (2001) Phytopathology 91: окиснення") приписується в загальному випадку 127-133]. захисна функція. Вивільненню реакційноздатних сполук на осПодібно до MIo зниження експресії NADPHнові кисню (ROS; наприклад, супероксид (O2-), оксидази забезпечує широку стійкість до різних гідроксирадикали і H2O2) приписується важлива ізолятів Blumeria graminis f.sp. hordei. У недовгозахисна функція в реакції на патогени рослин тривалих експериментах на основі "мовчання ге[Wojtaszek P (1997) Biochem J 322: 681-692]. Відомі нів" значно знижується ефективність проникнення різні шляхи, якими клітина виробляє ROS. У мак(утворення гаусторій), а саме більші ніж на 35%. рофагах ссавців необхідно при цьому, зокрема, Цей ефект по своїй силі відповідає досягнутому за назвати NADPH-оксидазу, що може передавати допомогою МІо-дволанцюгової PHK [див. електрони на молекулярний кисень. Гомологічні Schweizer і ін. (2000) Plant J 24:895- 903]. У ячменю ферменти були ідентифіковані також і в рослинах, дикого типу Pallas приблизно 40% проникнення [cм. Lamb та Dixon (1997) Annu Rev Plant Physiol грибів приводить до утворення гаусторій, на проPlant Моl ВіоІ 48: 251]. тивагу до чого зниження експресії NADPHБуло встановлено, що мутації в каталітичних оксидази за допомогою введення дволанцюгової підгрупах NADPH-оксидази в Arabidopsis thaliana PHK NADPH-оксидази (pNAox-dsRNA) складає виявляють знижену акумуляцію реакційноздатних тільки приблизно 25%. Той факт, що на чутливих посередників кисню (ROI). У відношенні гіперчутдо патогену сортах дикого (батьківського) типу, ливої реакції (HR) картина не була гомогенною. таких, як Pallas, можна було спостерігати проникПри використанні подвійного мутанта при інфекції нення на рівні значення приблизно від 40 до 50%, авірулентними бактеріями Pseudomonas syringae пояснюється постійно присутньою базовою стійкісBakterium була виявлена знижена реакція HR, у тю NADPH-оксидази, що по вищевикладеним притой час, як при інфекції вірулентними бактеріями чинам потрібно розуміти як ключовий елемент для Oomyceten Peronospora parasitica була виявлена успішного проникнення патогену, такого як Bgh, у підвищена HR. Ріст як вірулентних, так і авіруленрослинну клітину. тних штамів P.syringae, не змінювався в порівнянні 7 84406 8 Крім того, спосіб має переваги стосовно всіх плю, моркву, буряк, редьку, цукровий буряк, солоспособів, при яких реалізується стійкий до патогедку картоплю, огірок, цикорій салатний, цвітну кану фенотип за допомогою понадекспресії білка, що пусту, брокколі, спаржу, цибулю, часник, селеру, надає стійкості. Відключення гена можна реалізуполуниці, малину, ожину, ананас, авокадо, і різні вати без експресії (чужорідного) білка. В ідеальвиди дерев, чагарників, горіхових і виноградних. ному випадку дезактивується тільки ендогенний Види дерев включають переважно сливові, вишген. Це має переваги при допуску та прийнятті неві, персикові, нектаринові, абрикосові, бананові, споживачем, який часто має упереджену думку яблуневі, грушеві дерева, дерева папайї, манго, щодо рослин з чужорідними білками. Особливо айви. бажаним в цьому зв'язку є застосування індуковаДалі включені декоративні рослини, корисні них промоторів для зниження кількості NADPHабо декоративні дерева, квіткові рослини, квіткові оксидази, активності або функції, що, наприклад, декоративні рослини для зрізання, кущі або дерпри застосуванні індукованих патогенами промонина, як наприклад, але без обмеження, родини торів дозволяє проводити експресію тільки в разі Rosaceae, такі, як троянди, Ericaceae, такі як ропотреби (тобто при ураженні патогенами). додендрони й азалії, Euphorbiaceae, такі, як пуанСпосіб за винаходом може, у принципі, застозеція і кротон, Caryophyllaceae, такі, як гвоздика, совуватися на усіх видах рослин, переважно на Solanaceae, такі, як петунії, Gesneriaceae, такі, як таких, у яких природним чином експримується узамбарські фіалки, Balsaminaceae, такі, як бальNADPH-оксидаза або її функціональний еквівазамін, Orchidaceae, такі, як орхідеї, lridaceae, такі, лент. як гладіолуси, ірис, фрезії і крокуси, Compositae, Під "рослиною" у рамках даного винаходу вартакі, як календула, Geraniaceae, такі, як геранії, то розуміти всі роди та види вищих і нижчих росLiliaceae, такі, як драцена, Могасеае, такі, як фікус, лин рослинного світу. У це поняття включені зрілі Агасеае, такі, як філодендрон та інші. рослини, посівний матеріал, пагони і проростки, а У рамках винаходу бажані такі рослини, що також одержані від них фрагменти, матеріал для застосовуються як продукти харчування та корми, розмноження, рослинні органи, тканини, протоплаособливо переважно однодольні, такі, як вищенасти, калус та інші культури, наприклад, клітинні ведені види зернових. культури, а також всі інші види угруповань рослинОсобливо переважно спосіб за винаходом заних клітин у функціональні або структурні одиниці. стосовують для однодольних рослин із сільськогоПід зрілими рослинами маються на увазі будь-які сподарським значенням, таких, як пшениця, овес, стадії розвитку поза зародком. Під зародком мапросо, ячмінь, жито, кукурудза, рис, гречка, сорго, ється на увазі молода, незріла рослина в ранній трітікале, пшениця-спельта, льон або цукрова тростадії розвитку. Термін "рослина" включає всі одстина. нолітні та багаторічні однодольні та дводольні Під "стійкістю до патогенів" мається на увазі рослини та охоплює без обмеження, наприклад, зниження або ослаблення хворобливих симптомів такі, роди, як Cucurbita, Rosa, Vitis, Juglans, рослини внаслідок ураження патогеном. Симптоми Fragaria, Lotus, Medicago, Onobrychis, Trifolium, можуть бути різного роду. Переважно, вони вклюTrigonella, Vigna, Citrus, Linum, Geranium, Manihot, чають такі симптоми, що приводять безпосередDaucus, Arabidopsis, Brassica, Raphanus, Sinapis, ньо або опосередковано до погіршення якості росAtropa, Capsicum, Datura, Hyoscyamus, лини, зниження врожаю, придатності для Lycopersicon, Nicotiana, Solarium, Petunia, Digitalis, застосування в якості кормового або харчового Majorana, Ciahorium, Helianthus, Lactuca, Bromus, продукту або ж ускладнюють висівання, збір вроAsparagus, Antirrhinum, Heterocallis, Nemesis, жаю або зберігання зібраного врожаю. Pelargonium, Panieum, Pennisetum, Ranunculus, Під поняттями "надання", "наявність", "одерSenecio, Salpiglossis, Cucumis, Browaalia, Glycine, жання" або "підвищення" стійкості до патогену маPisum, Phaseolus, Lolium, Oryza, Zea, Avena, ється на увазі те, що захисний механізм визначеHordeum, Secale, Triticum, Sorghum, Picea і ного виду рослини або сорту внаслідок Populus. застосування способу за винаходом має в порівПоняття "рослина" охоплює переважно однонянні з вихідним типом рослини ("батьківської росдольні культурні рослини, як наприклад, такі види лини"), на якому спосіб за винаходом не застосозернових, як пшениця, ячмінь, жито, просо, трітікавували, при однакових умовах (як наприклад, ле, кукурудза, рис, сорго або овес, а також цукрову кліматичні умови та умови висівання, вид патогену тростину. і т.п.) більш високу стійкість до одного або декільДалі поняття дводольні культурні рослини кох патогенів. При цьому підвищена стійкість певключає, наприклад, реважно виявляється в більш слабко виражених - Brassicacae, такі як рапс, канола, блощичник, симптомах хвороби, причому симптоми хвороби Arabidopsis, види капусти або каноли, бобові, такі, означають поряд з вищенаведеними впливами як соя, люцерна, горох, бобові рослини або земтакож і ефективність проникнення патогену в росляний горіх, лину або рослинну клітину або ж ефективність - Solanaceae, такі, як картопля, тютюн, томати, проліферації в або на них. При цьому симптоми баклажани або перець, Asteraceae, такі, як соняшхвороби знижуються переважно, принаймні, на ник, тагетес, салат або календула, 10% або, принаймні, на 20%, особливо переважно, - Cucurbitaceae, такі, як диня, гарбуз або кабапринаймні, на 40% або 60%, особливо переважно, чки, принаймні, на 70% або 80%, найбільш переважно, а також льон, бавовник, коноплі, конюшину, принаймні, на 90% або 95%. шпинат, льон звичайний, червоний перець, карто 9 84406 10 Під поняттям "вибір" маються на увазі у відважні гриби, такі, як борошниста роса. Проте неношенні рослин, у яких - на відміну або в порівобхідно враховувати, що зниження експресії білка нянні з вихідними рослинами - наявна або підвиNADPH-оксидази, його активності або функції тащена стійкість до, принаймні, одного патогену, усі кож впливає на стійкість до інших патогенів. Зміни способи, придатні для розпізнавання наявної або в структурі стінки клітини можуть представляти підвищеної стійкості до патогену. Це можуть бути собою основні механізми стійкості до патогенів. симптоми патогенної інфекції (наприклад, утвоПриклади без обмеження можуть служити нарення гаусторій при інфекціях грибними захворюступні патогени: ваннями), а також вищеописані симптоми, що від1. Грибні або грибоподібні патогени: носяться до якості рослини, кількості врожаю, грибні або грибоподібні патогени (як наприпридатності до застосування як кормових та харклад, Chromista), що походять переважно з групи, чових продуктів і т.п. яка включає Plasmodiophoramycota, Oomycota, Під поняттям "патоген" у рамках винаходу роAscomycota, Chytridiomyceten, Zygomyceten, зуміють, проте, без обмеження, наприклад, віруси Basidiomycota і Deuteromyceten (Fungi imperfecti). або віроїди, бактерії, гриби, тваринних шкідників, Прикладами є приведені в таблицях 1 і 2 патогени як наприклад, комах або нематод. Особливо перета пов'язані з ними хвороби. Таблиця 1 Грибні хвороби рослин Хвороба Карликова іржа ячменю Жовта іржа Справжня борошниста роса Побуріння лузги Септоріоз листя зернових Фузаріоз пшениці Ламкість колосся пшениці Сажка Тверда сажка пшениці Офіобольозна гниль Патоген Puccinia recondita P. striiformis Erysiphe graminis/Blumeria graminis Septoria nodorum Septoria tritici Fusarium spp. Pseudocercosporella herpotrichoides Ustilago spp. Tilletia caries Gaeumannomyces graminis Colletotrichum graminicola (teleomorph: Glomerella Антракоз листя graminicola Politis); Glomerella tucumanensis (anamorph: Антракоз стебел Glomerella falcatum Went) Кам'яна черва Aspergillus flavus Rhizoctonia solani Kuhn = Rhizoctonia microsclerotia J. Matz Різоктоніоз конюшини (telomorph: Thanatephorus cucumeris) Acremonium strictum W. Gams = Cephalosporium acremonium Цефалоспоріоз судинних пучків кукурудзи Auct. non Corda Чорна гниль зерна Lasiodiplodia theobromae = Botryodiplodia theobromae Borde bianco Marasmiellus sp. Чорна плямистість Physoderma maydis Гниль зерна, що викликається Cephalosporium Acremonium strictum = Cephalosporium acremonium Вугільна гниль Macrophomina phaseolina Гниль колосся, що викликається Corticium Thanatephorus cucumeris = Corticium sasakii Curvularia clavata, C. eragrostidis, = C. maculans (teleomorph: Cochiiobolus eragrostidis), Curvularia naequalis, C. intermedia (teleomorph: Cochliobolus ntermedius), Curvularia lunata Плямистість листя, що викликається (teleomorph: Cochiiobolus lunatus), Curvularia pallescens Curvularia teleomorph: Cochiiobolus pallescens), Curvularia senegalensis, C. tuberculata (teleomorph: Cochiiobolus tuberculatus) Плямистість листя, що викликається Didymella Didymella exitalis Суха гнилизна качанів та стебел кукурудзи та Diplodia frumenti (teleomorph: Botryosphaeria estucae) стеблова гнилизна, які викликаються Diplodia Суха гнилизна качанів та стебел кукурудзи, стеблова гнилизна, гнилизна сіянців і захвоDiplodia maydis = Stenocarpella maydis рювання сходів кукурудзи, льону і пшениці, що викликаються Diplodia Плямистість або смугастість листя, що виклиStenocarpella macrospora = Diplodialeaf macrospora каються Diplodia 11 84406 12 Таблиця 2 Несправжня борошниста роса Хвороба Бура несправжня борошниста роса Несправжня борошниста роса Несправжня борошниста роса зеленого колосся (несправжня борошниста роса злакових) Явайская несправжня борошниста роса Філіпінська несправжня борошниста роса Несправжня борошниста роса сорго Спонтанна несправжня борошниста роса Несправжня борошниста роса цукрової тростини Суха гнилизна качанів та стебел кукурудзи (гнилизна качанів, зерна і стебла) Патоген Sclerophthora rayssiae var. zeae Sclerophthora macrospora = Sclerospora macrospora Sclerospora graminicola Peronosclerospora maydis = Sclerospora maydis Peronosclerospora philippinensis = Sclerospora philippinensis Peronosclerospora sorghi = Sclerospora sorghi Peronosclerospora spontanea = Sclerospora spontanea Peronosclerospora sacchari = Sclerospora sacchari Nigrospora oryzae (teleomorph: Khuskia oryzae) Alternaria alternate = A. tenuis, Aspergillus glaucus, A. niger, Aspergillus spp., Botrytis cinerea (teleomorph: Botryotinia fuckeliana), Cunninghamella sp., Curvularia pallescens, Сухі гнилизни качанів та стебел кукурудзи, Doratomyces stemonitis = Cephalotrichum stemonitis, другорядні Fusarium culmorum, Gonatobotrys simplex, Pithomyces maydicus, Rhizopus microsporus Tiegh., R. stolonifer = R. nigricans, Scopulariopsis brumptii Ріжки Claviceps gigantea (anamorph: Sphacelia sp.) Вічкова плямистість Aureobasidium zeae = Kabatiella zeae Фузаріоз, гнилизна колосся стебел Fusarium subglutinans = F. moniliforme var.subglutinans Фузаріоз, гнилизна стебел зерна та коренів Fusarium moniliforme (teleomorph: Gibberella fujikuroi) Фузаріоз, стеблова гнилизна коренева гнилиFusarium avenaceum (teleomorph: Gibberlla avenacea) зна сходів, що викликаються Fusarium Гнилизна колосся і стебел Gibberella zeae (anamorph: Fusarium graminearum) Botryosphaeria zeae = Physalospora zeae (anamorph: Сіра гнилизна колосся Macrophoma zeae) Сіра плямистість листя Cercospora sorghi = C. sorghi var. maydis, C. zeae-maydis Коренева гнилизна, що викликається Exserohilum pedicellatum = Helminthosporium pedicellatum Helminthosporium (teleomorph: Setosphaeria pedicellata) Суха гнилизна качанів і стебел кукурудзи, що Cladosporium cladosporioides = Hormodendrum викликається Hormodendrum (гнилизна, що cladosporioides, C. herbarum (teleomorph: Mycosphaerella викликається Cladosporium) tassiana) Плямистість листя, що викликається Hyalothyridium maydis Hyalothyridium В'янення Cephalosporium maydis Alternaria alternate, Ascochyta maydis, A. Tritici, A. Zeicola, Bipolaris victoriae = Helminthosporium ictoriae (teleomorph: Cochliobolus victoriae), C. Sativus (anamorph: Bipolaris sorokiniana = H. Sorokinianum = H. Sativum), Epicoccum nigrum, Exserohilum prolatum = Drechslera prolata Плямистості листя, другорядні (teleomorph: Setosphaeria prolata) Graphium penicillioides, Leptosphaeria maydis, Leptothyrium zeae, Ophiosphaerella herpotricha, (anamorph: Scolecosporiella sp.), Paraphaeosphaeria michotii, Phoma sp., Septoria zeae, S. Zeicola, S. Zeina Захворювання листя північних зернових (біла Setosphaeria turcica (anarnoфh: Exserohilum turcicum = гнилизна, гнилизна верхівок стебел, смугасHelminthosporium turcicum) тість) Плямистість листя північних зернових Cochliobolus carbonum (anamorph: Bipolaris zeicola = Суха гнилизна качанів і стебел кукурудзи, що Helminthosporium carbonum) викликається Helminthosporium (сорт 1) 13 Суха гнилизна качанів та стебел кукурудзи, що викликається Penicillium (блакитна вічковість, блакитна цвіль) Стеблова гнилизна та коренева гнилизна, що викликаються Phaeocytostroma Плямистість листя, що викликається Phaeosphaeria Суха гнилизна качанів і стебел кукурудзи, що викликається Physalospora (суха гнилизна качанів і стебел кукурудзи, що викликається Botryosphaeria) Пурпурність листової піхви Стеблова гнилизна і кореневa гнилизна, що викликаються Pyrenochaeta Коренева гнилизна, що викликається Pythium Стеблова гнилизна, що викликається Pythium Кладоспоріоз (цвіль колосся гнилизна листя і насіння) Ризоктоніоз (склероціальна гнилизна) Коренева гнилизна і стеблова гнилизна, що викликаються Rhizoctonia Кореневі гнилизни, другорядні Плямистість листя, що викликається Rostratum (гельмінтоспоріозні хвороби листя, гнилизна колосся і стебел) Іржа усіх зернових Іржа південних зернових Іржа тропічних зернових Суха гнилизна качанів і стебел кукурудзи, що викликається Sclerotium (південнe захворювання) Гнилизна сіянців-фузаріоз Плямистість листя, що викликається Selenophoma Гнилизна піхви Гнилизна лузги Цвіль силосу Сажка загальна Сажка несправжня Сажка сорго Захворювання листя південних зернових і гнилизна стебел Південна плямистість листя Гнилі стебел, другорядні Гнилі при зберіганні Смолиста плямистість 84406 14 Penicillium spp., P. chrysogenum, P. expansum, P. oxalicum Phaeocytostroma ambiguum, = Phaeocytosporella zeae Phaeosphaeria maydis = Sphaerulina maydis Botryosphaeria festucae = Physalospora zeicola anamorph: Diplodia frumenti) Hemiparasitic bacteria and fungi Phoma terrestris = Pyrenochaeta terrestris Pythium spp., P. arrhenomanes, P. graminicola Pythium aphanidermatum = P. butleri L. Epicoccum nigrum Rhizoctonia zeae (teleomorph: Waitea circinata) Rhizoctonia solani, Rhizoctonia zeae Altemaria alternata, Cercospora sorghi, Dictochaeta fertilis, Fusarium acuminatum (teleomorph: Gibberella acuminata), F. equiseti (teleomorph: G. ntricans), F. oxysporum, F. pallidoroseum, F. poae, F. roseum, G. cyanogena, (anamorph: F. sulphureum), Microdochium bolleyi, Mucor sp., Periconia circinata, Phytophthora cactorum, P. drechsleri, P. nicotianae var. parasitica, Rhizopus arrhizus Setosphaeria rostrata, (anamorph: Exserohilum rostratum = Helminthosporium ostratum) Puccinia sorghi Puccinia polysora Physopella pallescens, P. zeae = Angiopsora zeae Sclerotium rolfsii Sacc. (teleomorph: Athelia rolfsii) Bipolans sorokiniana, B. zeicola = Helminthosporium carbonum, Diplodia maydis, Exserohilum pedicillatum, Exserohilum turcicum = Helminthosporium turcicum, Fusarium avenaceum, F. culmorum, F. moniliforme, Gibberella zeae (anamorph: F. graminearum), Macrophomina phaseolina, Penicillium spp., Phomopsis sp., Pythium spp., Rhizoctonia solani, R. zeae, Sclerotium rolfsii, Spicaria sp. Selenophoma sp. Gaeumannomyces graminis Myrothecium gramineum Monascus purpureus, M ruber Ustilago zeae = U. maydis Ustilaginoidea virens Sphacelotheca reiliana = Sporisorium holcisorghi Cochliobolus heterostrophus (anamorph: Bipolaris maydis = Helminthosporium maydis) Stenocarpella macrospora = Diplodia macrospora Cercospora sorghi, Fusarium episphaeria, F. merismoides, F. oxysporum Schlechtend, F. poae, F. roseum, F. solani (teleomorph: Nectria haematococca), F. tricinctum, Mariannaea elegans, Mucor sp., Rhopographus zeae, Spicaria sp. Aspergillus spp., Penicillium spp. and other fungi Phyllachora maydis 15 84406 16 Суха гнилизна качанів і стебел кукурудзи і коренева гнилизна, що викликається Trichoderma viride = T. lignorum teleomorph: Hypocrea sp. Trichoderma Біла суха гнилизна качанів та стебел кукуруStenocarpella maydis = Diplodia zeae дзи, коренів і гнилизна стебел Ascochyta ischaemi, Phyllosticta maydis (teleomorph: Мозаїчна хвороба бавовнику Mycosphaerella zeae-maydis) Зональна плямистість листя Gloeocercospora sorghi особливо переважні Plasmodiophoromycota, такі, як Plasmodiophora brassicae (кіла капусти), Spongospora subterranea (порошиста парша бульб картоплі), Polymyxa graminis (хвороба коренів на злаках та дернині), - Oomycota, такі, як Bremia lactucae (несправжня борошниста роса на салаті), Peronospora (несправжня борошниста роса), на ротиках (P. antirrhini), на цибулі (P. destructor), на шпинаті (P. effusa), на соєвих бобах (P. Manchurica), тютюні ("блакитна гнилизна", P. tabacina), на люцерні посівній та конюшині (P. trifolium), Pseudoperonospora humuli (несправжня борошниста роса хмелю), Plasmopara (несправжня борошниста роса винограду) (P. viticola) та на соняшнику (P. haJstedii), Sclerophtohra macrospora (несправжня борошниста роса злаків і дернини), Pythium (плямистість насіння, наприклад, чорна гнилизна на буряку внаслідок P. debaryanum), Phytophthora infestans (гнилизна, фітофтороз картоплі, фітофтороз томатів і т.п.), Albugo spec, (біла гнилизна коренів і стебел рослин). - Ascomycota, такі як Microdochium nivale (сніжна цвіль жита і пшениці), Fusarium graminearum, Fusarium culmorum (цвіль колосся пшениці), Fusarium oxysporum (фузаріозне в'янення томатів), Blumeria graminis (справжня борошниста роса ячменю (f.sp. hordei) і пшениці (f.sp. tritici)), Erysiphe pisi (борошниста роса горошку), Nectria galligena (нектріозний рак стовбура та гілок плодових дерев), Unicnula necator (справжня борошниста роса виноградної лози), Pseudopeziza tracheiphila (краснуха винограду), Claviceps purpurea (ріжки жита і трав), Gaeumannomyces graminis (офіобольозна гнилизна на пшениці, житі, та інших злаках), Magnaporthe grisea (пірікуляріоз рису), Pyrenophora graminea (смугаста плямистість ячменю), Pyrenophora teres (сітчаста плямистість ячменю), Pyrenophora tritici-repentis (плямистість листя пшениці (септоріоз листя зернових)), Venturia inaequalis (парша яблуні), Sclerotinia sclerotium (склеротиніоз (біла гнилизна) рапсу), Pseudopeziza medicaginis (бура плямистість люцерни, білої та червоної конюшини). - Basidiomycetes, такі, як Typhula incarnata (тифельоз ячменю, жита, пшениці), Ustilago maydis (пухирчаста сажка кукурудзи), Ustilago nuda (пильна сажка ячменю), Ustilago tritici (пильна сажка пшениці, полби), Ustilago avenae (пильна сажка вівса), Rhizoctonia solani (різоктоніоз картоплі), Sphacelotheca spp. (сажка сорго), Melampsora Ііnі (іржа льону), Puccinia graminis (чорна іржа пшениці, ячменю, жита, вівса), Puccinia recondita (бура іржа пшениці), Puccinia dispersa (бура іржа жита), Puccinia hordei (бура іржа ячменю), Puccinia coronata (корончата іржа вівса), Puccinia striiformis (жовта ірм пшениці, ячменю, жита, а також численних злаків), Uromyces appendiculatus (ірн квасолі), Sclerotium rolfsii (біла гнилизна коренів і стовбурів рослин). - Deuteromycetes (Fungi imperfecti), такі, як Septoria nodorum (септоріоз пшени (Septoria tritici), Pseudocercosporella herpotrichoides (гнилизна кореневої шийки пшениі. ячменю, жита), Rynchosporium secalis (плямистість листя жита і ячменю), Alternar solani (альтернарюз картоплі, томатів), Phoma betae (кореневий опік буряка Cercospora beticola (плямистість листя буряка), (Altemaria brassicae (альтернаріс рапсу, капусти інших хрестоцвітих), Verticillium dahliae (гнилизна стебел рапсу Colletotrichum lindemuthianum (антракноз квасолі), Phoma lingam - полягання сход (чорна ніжка капусти; гнилизна стебел або чорна ніжка рапсу), Botrytis cinere (цвілеподібна сіра гнилизна виноградної лози, полуниць, томатів, хмелю і т.п.). Найбільш переважні Phytophthora mfestans (фітофтороз картоплі, фітофторс томатів і т.п.), Microdochium nivale (vormals Fusarium nivale; сніжна цвіль жита пшениці), Fusarium graminearum, Fusarium culmorum (цвіль колосся пшениці), Fusariur oxysporum (в'янення томатів, що спричинене Fusarium), Blumeria graminis (справжн борошниста роса ячменю (f. Sp. Hordei) і пшениці (f. Sp. Tritici)), Magnaporthe grise (пірікуляріоз рису), Sclerotinia sclerotium (біла стеблистість, біла гнилизна рапсу; Septoria nodorum і Septoria tritici (септоріоз пшениці), Altemaria brassicae (альтернаріо рапсу, капусти та інших хрестоцвітих), Phoma lingam (чорна ніжка капусти; гнилизн; стебел або чорна ніжки рапсу). Бактеріальні патогени: В таблиці 3 приведені патогени та пов'язані з ними хвороби рослин я необмежувальні приклади. 17 84406 18 Таблиця 3 Бактеріальні хвороби рослин Хвороба Бактеріальний антракноз листя і стебел Бактеріальна плямистість листя Бактеріальний антракноз стебел Патоген Pseudomonas avenae subsp. Avenae Xanthomonas campestris pv. Holcicola Enterobacter dissolvens = Erwinia dissolvens Erwinia carotovora subsp. carotovora, Erwinia chrysanthemi Офіобольозна гнилизна pv. zeae Бактеріальна смугастість Pseudomonas andropogonis Плямистість Pseudomonas syringae pv. coronafaciens Clavibacter michiganensis subsp. nebraskensis = В'янення (плямистість листя і в'янення) Corynebacterium michiganense pv.andnebraskense Листова плямистість, при якій утворюються Pseudomonas syringae pv порожнечі . syringae Пурпурність листової піхви Hemiparasitic bacteria Гнилизна зерна, сходів Bacillus subtilis Бактеріальне в'янення Pantoea stewartii = Erwinia stewartii Карликовість пшениці (achapparramiento, карликовість маїсу, карликовість маїсу Mesa Spiroplasma kunkelii Central або Rio Grande) Особливо переважні наступні патогенні бактерії: Corynebacterium sepedonicum (кільцева гнилизна бульб картоплі), Erwinia carotovora (бактеріальна чорна ніжка картоплі), Erwinia amylovora (бактеріальний опік грушевих, яблуневих, айвових), Streptomyces scabies (парша картоплі), Pseudomonas syringae pv. tabaci (бактеріальна рябуха тютюну), Pseudomonas syringae pv. phaseolicola (кутаста бактеріальна плямистість квасолі), Pseudomonas syringae pv. tomato (бактеріальна плямистість томатів), Xanthomonas campestris pv. malvacearum (плямистість листя бавовнику) і Xanthomonas campestris pv. oryzae (бактерільна гнилизна рису та інших злаків). 3. Вірусні патогени: Поняття "вірусні патогени" включають усі віруси рослин, такі, як наприклад, вірус мозаїка тютюну або огірків, вірус кільцевої плямистості, некрозний вірус, вірус карликової мозаїки кукурудзи і т.п. В таблиці 4 приведені патогени та пов'язані з ними хвороби рослин як необмежувальні приклади. Таблиця 4 Вірусні хвороби рослин Хвороба Американська смугастість пшениці (смугаста мозаїка пшениці) Смугаста мозаїка ячменю Жовта карликовість ячменю Мозаїка стоколосу Хлоротичная плямистіст злаків Corn chlorotic vein banding (Бразильська мозаїка кукурудзи) Летальний некроз зернових Мозаїчність огірків Хлоротична смугастість, що викликається вірусом Cynodon Мозаїка роговика (дике сорго) Руниста карликовість кукурудзи Хлоротична карликовість кукурудзи Хлоротична плямистість кукурудзи Мозаїчна карликовість кукурудзи Плямистість листя кукурудзи Смугастість кукурудзи Мозаїка кукурудзи (смугастість зерна, листя, Патоген American wheat striate mosaic virus(AWSMV) Barley stripe mosaic virus (BSMV) Barley yellow dwarf virus (BYDV) Brome mosaic virus (BMV) Cereal chlorotic mottle virus (CCMV) Corn chlorotic vein banding virus (CCVBV) Вірусний комплекс із Maize chlorotic mottle virus (MCMV) і Maize dwarf mosaic virus (MDMV) А або В або Wheat streak mosaic virus (WSMV) Вірус мозаїчності огірків (CMV) Cynodon chlorotic streak virus (CCSV) Johnsongrass mosaic virus (JGMV) Mycoplasma- like organism (MLO) associated Maize chlorotic dwarf virus (MCDV) Maize chlorotic mottle virus (MCMV) Maize dwarf mosaic virus (MDMV) strains A, D, E and F Maize leaf fleck virus (MLFV) Maize line virus (MLV) Maize mosaic virus (MMV) 19 enanismo rayado) Плямистість і хлоротична карликовість кукурудзи Прозора кільцева мозаїка кукурудзи Maize raya gruesa maize rayado fino (смугастість) Почервоніння листя і червона штрихуватість кукурудзи Червона штрихуватість кукурудзи Кільцева плямистість (крапчастість) кукурудзи Maize rio IV Нерівна карликовість кукурудзи (nanismo ruvido) Стерильна карликовість кукурудзи Смугастість кукурудзи Штрихуватість кукурудзи (хлоротична штрихуватість кукурудзи, maize hoja blanca) Низькорослість кукурудзи Недорозвинення волот кукурудзи Прожилковий енацій кукурудзи Maize wallaby ear Білизна листя кукурудзи Біла лінійна мозаїка кукурудзи Почервоніння листя проса Північна мозаїка зернових Псевдорозетковість зерна (заляльковування) Стерильна карликовість вівса Чорно-смугаста карликовість рису Штрихуватість рису Мозаїка сорго Хвороба Фіджі цукрової тростини Мозаїка цукрової тростини Плямиста мозаїка пшениці 84406 20 Maize mottle and chlorotic stunt virus Maize pellucid ringspot virus (MPRV) vlaize raya gruesa virus (MRGV) Maize rayado fino virus (MRFV) Mollicute Maize red stripe virus (MRSV) Maize ring mottle virus (MRMV) Maize rio cuarto virus (MRCV) Maize rough dwarf virus (MRDV) (Cereal tillering disease virus) Maize sterile stunt virus (strains of barley yellow striate virus) Maize streak virus (MSV) Maize stripe virus Maize stunting virus Maize tassel abortion virus (MTAV) Maize vein enation virus (MVEV) Maize wallaby ear virus (MWEV) Maize white leaf virus Maize white line mosaic virus (MWLMV) Millet red leaf virus (MRLV) Northern cereal mosaic virus (NCMV) Oat pseudorosette virus Oat sterile dwarf virus (OSDV) Rice black- streaked dwarf virus (RBSDV) Rice stripe virus (RSV) Sorghum mosaic virus (SrMV) (auch: sugarcane mosaic virus (SCMV) Stflmme H, I and M) Sugarcane Fiji disease virus (FDV) Sugarcane mosaic virus (SCMV) strains A, B, D, E, SC, BC, Sabi and MB (formerly MDMV- B) Wheat spot mosaic virus (WSMV) 4. Тваринні шкідники 4.1 Комахи-шкідники: Як необмежувальні приклади можна привести таких жуків, гусінь, вошей або кліщів Переважними є шкідники родини Coieoptera, Diptera, Hymenoptera, Lepidoptera, Mallophaga, Homoptera, Hemiptera, Orthoptera, Thysanoptera. Dermaptera, lsoptera, Anoplura, Siphonaptera, Trichoptera, і т.п. Особливо переважні шкідники Coieoptera і Lepidoptera, такі, як, наприклад, кукурудзяний метелик (European Corn Borer (ECB)), Diabrotica barberi ("блішка довговуса"), Diabrotica undecimpunctata ("південна блішка довговуса"), Diabrotica virgifera ("західна блішка довговуса"), Agrotis ipsilon ("совка іпсілон"), Crymodes devastator ("прозора совка"), Feltia ducens ("тьмяна совка"), Agrotis gladiaria ("світло-бура совка"), Melanotus spp., Aeolus mellillus ("дротяник"), Aeolus mancus ("личинка ковалика пшеничного"), Horistonotus uhlerii ("піщаний дротяник"), Sphenophorus maidis ("довгоносик кукурудзяний"), Sphenophorus zeae ("довгоносик тимофіївки"), Sphenophorus parvulus ("довгоносик тонконогу"), Sphenophorus callosus ("південний зерновий довгоносик"), Phyllogphaga spp.("личинка хруща"), Anuraphis maidiradicis ("попелиця кукурудзяна коренева"), Delia platura ("личинка мухи паросткової"), Colaspis brunnea ("grape colaspis"), Stenolophus lecontei ("зернівки") і Clivinia impressifrons ("lender seedcom beetle"). Далі варто привести наступних: п'явиця червоногруда (Oulema melanopus), муха шведська (Oscinella frit), дротяники (Agrotis lineatus) та попелиця (як, наприклад, попелиця черемхова звичайна Rhopalosiphum padi, попелиця злакова Sitobion avenae). 4.2 Нематоди: Як необмежувальні приклади варто назвати патогени з таблиці 5 та пов'язані з ними хвороби рослин. 21 84406 22 Таблиця 5 Паразитичні нематоди Ураження Довгостилетна нематода Нематода стеблова Свердлильники Haferzystenaelchen ("Cyst") Вівсяна нематода Совка Несправжня галова нематода Ланцет колумбійський Ланцет Ураження Голка Кільце Галова нематода Спіраль Ужалення В'янення коренів Припинення росту Патогенні нематоди Dolichodorus spp., D. Heterocephalus Ditylenchus dipsaci Radopholus similis Heterodera avenae, H. zeae, Punctodera chalcoensis Xiphinema spp., X. americanum, X. mediterraneum Nacobbus dorsalis Hoplolaimus columbus Hoplolaimus spp., H. galeatus Pratylenchus spp., P. brachyurus, P. crenatus, P. hexincisus, P. neglectus, P. penetrans, P. scribneri, P. thornei, P. zeae Longidorus spp., L. breviannulatus Criconemella spp., C. ornata Meloidogyne spp., M. chitwoodi, M. incognita, M. javanica Helicotylenchus spp. Belonolaimus spp., B. longicaudatus Paratrichodorus spp., P. christiei, P. minor, Quinisulcius acutus, Trichodorus spp. Tylenchorhynchus dubius Особливо переважні Globodera rostochiensis і G. pallida (галоутворювальна нематода картоплі, томатів і ін.), Heterodera schachtii (гетеродера цукрового буряка, кормового буряка, рапсу, капусти і т.п. ), Heterodera avenae (вівсяна цистоутворювальна нематода вівса та інших зернових), Ditylenchus dipsaci (нематода стеблова жита, вівса, кукурудзи, конюшини, тютюну, буряка), Anguina tritici (нематода пшенична), фітогельмінтоз (полби жита), Meloidogyne hapla (галова нематода моркви, огірків, салату, томатів, картоплі, цукрового буряка, люцерни). Як переважні для окремих сортів грибних або вірусних патогенів варто назвати, наприклад: 1. Ячмінь: грибні, бактеріальні та вірусні патогени: Puccinia graminis f.sp. hordei (стеблова іржа ячменю), Blumeria (Erysiphe) graminis f.sp. hordei (справжня борошниста роса ячменю), вірус жовтої карликовості ячменю (BYDV), патогенні комахи/нематоди: Ostrinia nubilalis (кукурудзяний метелик); Agrotis ipsilon (совка іпсілон); Schizaphis graminum (попелиця злакова звичайна); Blissus leucopterus leucopterus (клопчерепашка пшенична північно-американська); Acrostemum hilare (зелений клоп-щитник); Euschistus servus (коричневий клоп-щитник); Deliaplatura (личинка мухи паросткової); Mayetiola destructor (гесенська мушка); Petrobia latens (пшеничний кліщ). 2. Соєві боби: грибні, бактеріальні або вірусні патогени: Phytophthora megasperma fsp.glycinea, Macrophomina phaseolina, Rhizoctonia solani, Sclerotinia sclerotiorum, Fusarium oxysporum, Diaporthe phaseolorum var. sojae (Phomopsis sojae), Diaporthe phaseolorum var. caulivora, Sclerotium rolfsii, Cercospora kikuchii, Cercospora sojina, Peronospora manshurica, Colletotrichum dematium (Colletotrichum truncatum), Corynespora cassiicola, Septoria glycines, Phyllosticta sojicola, Alternaria alternata, Pseudomonas syringae p.v. glycinea, Xanthomonas campestris p.v. phaseoli, Microsphaera diffussa, Fusarium semitectum, Phialophora gregata, вірус мозаїк соєвих бобів, Glomerella glycines, вірус кільцевої плямистості тютюну, вірус смугастості тютюну, Phakopsorapachyrhizi, Pythium aphanidermatum, Pythium ultimum, Pythium debaryanum, вірус бронзовості томату, Heterodera glycines Fusarium solani. Патогенні комахи/нематоди: Pseudoplusia includens (п'ядак соєвих бобів); Anticarsia gemmatalis (совка); Plathypena scabra (зелений шкідник конюшини); Ostrinia nubilalis (кукурудзяний метелик); Agrotis ipsilon (парна совка); Spodoptera exigua (ратні хробаки буряка); Heliothis virescens (листовійка-почкоїд бавовнику); Helicoverpa zea (совка бавовняна); Epilachna varivestis (мексиканська зернівка бобова); Myzus persicae (попелиця персикова); Empoasca fabae (товстоголовка); Acrosternum hilare (зелений клоп-щитник); Melanoplus femurrubrum (коник червононогий); Melanoplus differentialis (коник відмітний); Hylemya platura (личинка мухи паросткової); Sericothrips variabilis (трипс соєвих бобів); Thrips tabaci (трипс тютюновий); Tetranychus turkestani (кліщик павутинний атлантичний); Tetranychus urticae (кліщик павутинний двоплямистий); 3. Канола: грибні, бактеріальні або вірусні патогени: Albugo Candida, Alternaria brassicae, Leptosphaeria maculans, Rhizoctonia solani, Sclerotinia sclerotiorum, Mycosphaerella brassiccola, Pythium ultimum, Peronospora parasitica, Fusarium roseum, Alternaria alternata. 4. Люцерна: грибні, бактеріальні або вірусні патогени: Clavibater michiganese subsp. insidiosum, Pythium ultimum, Pythium irregulare, Pythium splendens, Pythium debaryanum, Pythium aphanidermatum, 23 84406 24 Phytophthora megasperma, Peronospora trifoliorum, acremonium, Phytophthora cryptogea, Albugo Phoma medicaginis var. medicaginis, Cercospora tragopogonis. medicaginis, Pseudopeziza medicaginis, патогенні комахи/нематоди: Suleima Leptotrochila medicaginis, Fusarium, Xanthomonas helianthana (трав'яниця соняшникова, листовійка); campestris p.v. alfalfae, Aphanomyces euteiches, Homoeosoma electellum (трав'яниця соняшникова); Stemphylium herbarum, Stemphylium alfalfae. zygogramma exclamationis (совка соняшникова 5. Пшениця: оклична); Bothyrus gibbosus (жук морквяний); грибні, бактеріальні або вірусні патогени: Neolasioptera murtfeldtiana (галиця соняшникова); Pseudomonas syringae p.v. atrofaciens, Urocystis 7. Кукурудза: agropyri, Xanthomonas campestris p.v. translucens, грибні, бактеріальні або вірусні патогени: Pseudomonas syringae p.v. syringae, Alternaria Fusarium moniliforme var. subglutinans, Erwinia alternata, Cladosporium herbarum, Fusarium stewartii, Fusarium moniliforme, Gibberella zeae graminearum, Fusarium avenaceum, Fusarium (Fusarium graminearum), Stenocarpella maydi culmorum, Ustilago tritici, Ascochyta tritici, (Diplodia maydis), Pythium irregulare, Pythium Cephalosporium gramineum, Collotetrichum debaryanum, Pythium graminicola, Pythium graminicola, Erysiphe graminis f.sp. tritici, Puccinia splendens, Pythium ultimum, Pythium graminis f.sp. tritici, Puccinia recondita f.sp. tritici, aphanidermatum, Aspergillus flavus, Bipolaris maydis Puccinia striiformis, Pyrenophora tritici- repentis, O, T (Cochliobolus heterostrophus), Septoria nodorum, Septoria tritici, Septoria avenae, Helminthosporium carbonum I, Il та III (Cochliobolus Pseudocercosporella heфotrichoides, Rhizoctonia carbonum), Exserohilum turcicum I, Il та III, solani, Rhizoctonia cerealis, Gaeumannomyces Helminthosporium pedicellatum, Physoderma maydis, graminis var. tritici, Pythium aphanidermatum, Phyllosticta maydis, Kabatiella maydis, Cercospora Pythium arrhenomanes, Pythium ultimum, Bipolaris sorghi, Ustilago maydis, Puccinia sorghi, Puccinia sorokiniana, вірус жовтої карликовості ячменю, polysora, Macrophomina phaseolina, Penicillium Brome Mosaic Virus, Soil Borne Wheat Mosaic Virus, oxalicum, Nigrospora oryzae, Cladosporium вірус смугастої мозаїки пшениці, вірус веретеноherbarum, Curvularia lunata, Curvularia inaequalis, подібної мозаїки пшениці, американський вірус Curvularia pallescens, Clavibacter michiganese штрихуватої мозаїки пшениці, Claviceps purpurea, subsp. nebraskense, Trichoderma viride, вірус карTilletia tritici, Tilletia laevis, Ustilago tritici, Tilletia ликової мозаїки кукурудзи А та В, вірус смугастої indica, Rhizoctonia solani, Pythium arrhenomannes, мозаїки пшениці, вірус хлоротичної карликовості Pythium gramicola, Pythium aphanidermatum, High кукурудзи, Claviceps sorghi, Pseudonomas avenae, Plains Virus, European wheat striate virus, Puccinia Erwinia chrysanthemi p.v. Zea, Erwinia corotovora, graminis f.sp. tritici (стеблова іржа пшениці), Cornstunt spiroplasma, Diplodia macrospora, Blumeria (Erysiphe) graminis f.sp. tritici (справжня Sclerophthora macrospora, Peronosclerospora борошниста роса). sorghi, Peronosclerospora philippinesis, Патогенні комахи/нематоди: Pseudaletia Peronosclerospora maydis, PeronoscleYospora unipunctata (ратні хробаки); Spodoptera, frugiperda sacchari, Spacelotheca reiliana, Physopella zeae, (совка трав'яна); Elasmopalpus lignosellus (трав'яCephalosporium maydis, Caphalosporium ниця кукурудзяна); Agrotis orthogonia (західна совacremonium, вірус хлоротичної плямистості кукука); Elasmopalpus Zignosellus (трав'яниця кукурурудзи, High Plains Virus, вірус мозаїки кукурудзи, дзяна); Oulema melanopus (п'явиця червоногруда); Maize Rayado Fino Virus, вірус смугастості кукуруHypera punctata (довгоносик конюшиновий); дзи (MSV, вірус качанів кукурудзи), вірус штрихуDiabrotica undecimpunctata howardi (південна блішватості кукурудзи, Maize Rough Dwarf Virus. ка довговуса); російська пшенична попелиця; Патогенні комахи/нематоди: Ostrinia nubilalis Schizaphis graminum (зелений клоп); Macrosiphum (кукурудзяний метелик); Agrotis ipsilon (совка іпсіavenae (попелиця листова); Melanoplus лон); Helicoverpa zea (совка бавовникова); femurrubrum (коник червононогий); Melanoplus Spodoptera frugiperda. (совка трав'яна); Diatraea differentialis (коник відмітний); Melanoplus grandiosella (трав'яниця кукурудзяна південноsanguinipes (коник мігруючий); Mayetiola destructor західна); Elasmopalpus lignosellus (трав'яниця куку(гесенська мушка); Sitodiplosis mosellana (комарик рудзяна); Diatraea saccharalis (трав'яниця цукрової пшеничний); Meromyza americana (личинка америтростини); Diabrotica virgifera (західна блішка довканської меромізи); Hylemya coarctata (муха озиговуса); Diabrotica longicomis barberi (північна бліма); Frankliniella fusca (трипс тютюну), Cephus шка довговуса); Diabrotica undecimpunctata howardi cinctus (пилильник хлібний американський); Асегіа (південна блішка довговуса); Melanotus spp. (дроtulipae (галовий кліщ); тяники); Cyclocephala borealis (північний хрущ, що 6. Соняшник: маскується; личинка хруща); Cyclocephala грибні, бактеріальні або вірусні патогени: immaculata (південний хрущ, що маскується; личиPlasmophora halstedii, Sclerotinia sclerotiorum, Aster нка хруща); Popillia japonica (хрущик японський); Yellows, Septoria helianthi, Phomopsis helianthi, Chaetocnema pulicaria (листоїди), Sphenophorus Alternaria helianthi, Alternaria zinniae, Botrytis maidis (довгоносик кукурудзяний); Rhopalosiphum cinerea, Phoma macdonaldii, Macrophomina maidis (попелиця кукурудзяна листова); Anuraphis phaseolina, Erysiphe cichoracearum, Rhizopus maidiradicis (попелиця кукурудзяна коренева); oryzae, Rhizopus arrhizus, Rhizopus stolonifer, Blissus leucopterus leucopterus (клоп білокрилий); Puccinia helianthi, Verticillium dahliae, Erwinia Melanoplus femurrubrum (коник червононогий); carotovorum p.v. Carotovora, Cephalosporium Melanoplus sanguinipes (коник мігруючий); Hylemva platura (личинка мухи паросткової); Agromyza 25 84406 26 parvicornis (мінуюча мушка); Anaphothrips вий); Nephotettix nigropictus (цикада рисова); obscrurus (трипс безщетинковий); Solenopsis Blissus leucopterus leucopterus (клоп білокрилий); milesta (мураха-злодій); Tetranychus urticae (кліщик Acrosternum hilare (зелений клоп-щитник); павутинний двоплямистий). 11. Рапс: 8. Сорго: патогенні комахи/нематоди: Brevicoryne грибні, бактеріальні або вірусні патогени: brassicae (попелиця капустяна); Phyilotreta Exserohilum turcicum, Colletotrichum graminicola cruciferae (земляні блішки); Mamestra conjgurata (Glomerella graminicola), Cercospora sorghi, (совка Bertha); Plutella xylostella (міль капустяна); Gloeocercospora sorghi, Ascochyta sorghina, Delia ssp. (кореневі хробаки). Pseudomonas syringae p.v. syringae, Xanthomonas Під терміном "NADPH- оксидаза" у рамках виcampestris p.v. holcicola, Pseudomonas находу розуміють всі такі ферменти, що як істотну andropogonis, Puccinia purpurea, Macrophomina властивість мають здатність за допомогою переphaseolina, Perconia circinata, Fusarium monilifonne, носу окремих електронів перетворювати молекуAlternaria alternate, Bipolaris sorghicola, лярний кисень (О2) у супероксид (C2-). Бажаними є Helminthosporium sorghicola, Curvularia lunata, ті ферменти, що описані EC-класом Е.С. 1.23.45.3. Phoma insidiosa, Pseudomonas avenae При цьому NADPH-оксидаза може складатися з (Pseudomonas alboprecipitans), Ramulispora sorghi, одного або декількох пептидів, які можуть бути Ramulispora sorghicola, Phyllachara sacchari, однаковими або різними. Sporisorium reilianum (Sphacelotheca reiliana), Бажаною NADPH-оксидазою є білок флавоциSphacelotheca cruenta, Sporisorium sorghi, тохром, що як простетичну групу включає цитрохSugarcane mosaic H, Maize Dwarf Mosaic Virus А та ром b і/або FAD-rpyny. NADPH-оксидаза може В, Claviceps sorghi, Rhizoctonia solani, Acremonium складатися з 11 гетеродимера, причому strictum, Sclerophthona macrospora, підгрупа представляє собою функціональну підгPeronosclerospora sorghi, Peronosclerospora рупу флавоцитохрому та може включати як глікопphilippinensis, Sclerospora graminicola, Fusarium ротеїнові компоненти переносу електронів (гідроgraminearum, Fusarium oxysporum, Pythium фільний, цитозольний, С-термінальний домен, що antienomanes, Pythium graminicola. містить NADPH і FAD, а також від 4 до 6 NПатогенні комахи/нематоди: Chilo partellus термінальних, ймовірних трансмембранних (свердлильник сорго); Spodoptera frugiperda (совка спіралей, що включають дві комплексовані з гіститрав'яна); Helicoverpa zea (совка бавовнику); дином простетичні групи гему --підгрупа може Elasmopalpus lignosellus (трав'яниця кукурудзяна); включати C-термінальну, багату проліном послідоFeltia subterranea (совка гранульована); вність, що може зв'язувати потенційні цитологічні, Phvllophaga crinita (личинка хруща); Eleodes, активувальні фактори для NADPH-оксидази. ВнаConoderus und Aeolus spp. (дротяник); Oulema слідок зв'язування цитологічних phox білків (наmelanopus (п'явиця червоногруда); Chaetocnema приклад, р47- phox, p67- phox, p40- phox) і р21гас pulicaria (листоїди); Sphenophorus maidis (довгонозв'язувальний GTP-білок може відбуватися актисик кукурудзяний); Rhopalosiphum maidis (попеливація. ця кукурудзяна листова); Siphaflava (попелиця Фахівцеві в даній області відомі численні жовта цукрової тростини); Blissus leucopterus NADPH-оксидази з рослинних організмів (див. сеleucopterus (клоп білокрилий); Contarinia sorghicola ред інших публікацію [Torres MA і ін. (1998) Plant J (галиця соргова); Tetranychus cinnabarinus (кармі14: 365-370)]. Як необмежувальні приклади варто новий павутинний кліщ); Tetranychus urticae (кліщ назвати послідовності з наступними номерами павутинний двоплямистий). GenBnk Acc.-No: AJ251717 (Hordeum vulgare), 9. Бавовник: AP003560 (Oryza sativa var. japonica), AJ320505 патогенні комахи/нематоди: Heliothis virescens (Nicotiana tabacum), AB050660 (Solarium (бавовняна листовійка-брунькоїд); Helicoverpa zea tuberosum), AF088276 (Lycopersicon esculentum), (совка бавовникова); Spodoptera exigua (ратні хроAB008111 (Arabidopsis thaliana; Atrboh F), баки буряка); Pectinophora gossypiella (рожевий AF055357 (Arabidopsis thaliana; Rboh), AJ309006 хробак коробочок бововнику); Anthonomus grandis (Nicotiana tabacum; rboh), AP003271 (Oryza sativa grandis (довгоносик бавовниковий); Aphis gossypii cv. japonica), AF055355 (Arabidopsis thaliana; (попелиця бавовникова); Pseudatomoscelis seriatus Rboh), AF055353 (Arabidopsis thaliana; Rboh). Осо(блішка бавовникова); Trialeurodes abutilonea (бібливо бажані NADPH-оксидази, що включають локрилка); Lygus lineolaris (клоп луговий); послідовності № 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або Melanoplus femurrubrum (коник червононогий); 22. Melanoplus differentialis (коник відмітний); Thrips Послідовності з інших рослин, гомологичні роtabaci (трипс тютюновий); Franklinkiella fusca зкритим у рамках винаходу послідовностям (трипс тютюновий); Tetranychus cinnabarinus (карNADPH-оксидази, можуть бути легко знайдені, міновий павутинний кліщ); Tetranychus urticae наприклад, пошуком у банку даних або перевіркою (кліщик павутинний двоплямистий); банків даних при застосуванні послідовностей 10. Рис: NADPH-оксидази як пошукової послідовності, відпатогенні комахи/нематоди: Diatraea повідно, зонда. Як приклад варто привести при saccharalis (трав'яниця цукрової тростини); цьому послідовності з наступними номерами Spodoptera frugiperda (совка трав'яна); Helicoverpa GenBank Acc.-No.: САС51517.1, AJ251717, Т03973, zea (совка бавовникова); Colaspis brunnea (grape ВАВ68079.1, АР003560, Т02024, САС87256.1, colaspis); Lissorhoptrus oryzophilus (довгоносик риAJ320505, BAB70750.1, AB050660, AF088276_1, совий водний); Sitophilus oryzae (довгоносик рисоNP_564821.1,N M_105079, T00265 AC007764_16, 27 84406 28 NPJ92862.1, NM_117194, AF147783J, AAM28891.1, принаймні, 10%, бажано, принаймні, 10% або, AF506374, CAC84140.1, AJ309006, T51804, принаймні, 20%, особливо бажано, принаймні, NP_199602.1, NM_124165, BAB89740.1, AP003271, 40% або 60%, дуже бажано, принаймні, 70% або AAC39477.1, AF055355, NP_199919.1, NM_124485, 80%, найбільш бажано, принаймні, 90% або 95%. AAC39475.1, AF055353, NP_196356.1, NM_120821, Під поняттям "функція" варто розуміти переNP_194239.1, NM_118641, BAB08369.1, AB015475, важно силу зв'язування NADPH-оксидази із субAAC39478.1, AF055356, AC069143_9, стратом в організмі, тканині, клітині або частині NP_173357.1, NM_101781, NP_172383.1, клітини. Як субстрат придатні низькомолекулярні NM_100780, AAB70398.1, AC000106, AAC39476.1, сполуки, такі, як NADPH або FAD, а також інтеракAF055354, BAB70751.1, AB050661, BAB63664.1, тивні партнери білка NADPH-оксидази. AP003275, AAD24966.1, AF109150. Під "зниженням" функції варто розуміти кількіОсобливо бажана поліпептидна послідовність сне зниження сили зв'язування NADPH-оксидази з, NADPH-оксидази включає, принаймні, один мотив принаймні, одним субстратом в організмі, тканині, послідовностей, вибраний із групи послідовностей, клітині або частині клітини, наприклад, за допомощо включає гою одного з описаних нижче способів, у порівняні) AL (K/R) GL (K/R) ні з вихідним типом того ж роду і виду, на якому іі) DK (N/D XDG (R/K) (UUV) (T/N) E цей спосіб не застосовувався, при однакових інііі) LSASAN ших загальних умовах (як наприклад, умови вироiv) IMEELDP щування, вік рослини і т.п.). Під зниженням варто v) K (F/L) NMA (I/L) (I/V) LXPVCRN розуміти також зміну специфічності стосовно субvi) (E/Q) WHPFSIT страту, яку можна, наприклад, виразити через веvii) S (A/S) PXDD (Q/Y) (L/l) S (I/V) H (V/l/L) R личину kcat/Km. Зниження складає при цьому, viii) DGPYG (S/A)PAGDY принаймні, 10%, бажано, принаймні, 10% або, ix) L (I/V) GLGIGATP принаймні, 20%, особливо бажано, принаймні, x) FYWVTREQGSF 40% або 60%, особливого бажано, принаймні, 70% xi) GVFYCG або 80%, найбільш бажано, принаймні, 90% або Особливо бажана пептидна послідовність 95%. Партнери зв'язування для NADPH-оксидази включає, принаймні, № 2 або 3, більш бажано, можуть бути ідентифіковані, наприклад, за допопринаймні, № 4 або 5, найбільше бажано усі види могою гібридної системи дріжджі-2 відомим фахівпослідовностей, вибрані з групи послідовностей, цеві в даній області способом. що включає і), іі), ііі), iv), v), vi) vii), viii), ix) x) і хі). Способи визначення кількості білка, активності (дані в дужках означають альтернативно можливі NADPH-оксидази або сили зв'язування із субстраамінокислоти в цих положеннях, наприклад, (V/I) том відомі фахівцеві. Наприклад, можна вимірюваозначає те, що в цьому положенні можливий валін ти залежне від NADPH виробництво O2 або H2O2, або ізолейцин). яке інгібується DPI (наприклад, відновленням нітADPH-оксидаза може також означати кожну ро-синього тетразолію [NBT] або цитохрому C. іншу ланку ферментативного комплексу NADPHКількість білка може визначатися, наприклад, імуоксидази, що є важливим для активності NADPHнологічно при застосуванні відповідних антитіл. оксидази. Відповідні способи описані в публікаціях [Yu L та Під терміном "кількість білка" варто розуміти ін. (1999) Blood 94(7):2497-504; Doke N (1983а) кількість поліпептиду NADPH-оксидази в організмі, Physiol Plant Pathol 23:345- 357; Levine A та ін. тканині, клітині або частині клітини. "Зниження" (1994) Cell 79:583-593; Tenhaken R та ін. (1995) кількості білка означає зменшення кількості Ргос Nat Acad Sd USA 92: 4158-4163; Sagi M та NADPH-оксидази в організмі, тканині, клітині або Fluhr R. (2001) Plant Physiol 126(3): 1281-90; частині клітини, наприклад, за допомогою одного з Huckelhoven R та Kogel KH (1998) Mol Plant описаних нижче способів, у порівнянні з вихідним Microbe Interact 11:292-300; а також у джерелах, (диким) типом того ж роду і виду, на якому цей що цитуються у приведених публікаціях]. спосіб не застосовувався, в іншому при однакових Під "функціональними еквівалентами" NADPHзагальних умовах (як наприклад, умови вирощуоксидази варто розуміти такі послідовності, що вання, вік рослини і т.п.). Зниження складає при одержані від NADPH-оксидази, яка включає поліцьому, принаймні, 10%, бажано, принаймні, 10% пептидну послідовність № 2, 4, 6, 8, 10, 12, 14, 16, або, принаймні, 20%, особливо бажано, принаймні, 18, 20 або 22 або гомологичні їм, та мають одна40% або 60%, дуже бажано, принаймні, 70% або кові істотні властивості. 80%, найбільш бажано, принаймні, 90% або 95%. При цьому ефективність стійкості до патогену Під поняттям "активність" варто мати на увазі може відхилятися як вниз, так і вгору в порівнянні здатність NADPH-оксидази перетворювати молезі значенням, отриманим при зниженні однієї з кулярний кисень (O2) у супероксид (O2-). Під "зниNADPH- оксидаз, що включає поліпептидну посліженням" активності варто розуміти зниження загадовність № 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22. льної активності білка NADPH-оксидази в Перевага надається таким функціональним еквіорганізмі, тканині, клітині або частині клітини, навалентам, для яких ефективність стійкості до патоприклад, за допомогою одного з описаних нижче гену, виміряна, наприклад, за ефективністю проспособів, наприклад у порівнянні з вихідним (диникнення патогена (утворення гаусторій), ким) типом того ж роду і виду, на якому цей спосіб відрізняється не більше, ніж на 50%, переважно, не застосовувався, при цьому інші загальні умови на 25%, особливо переважно, на 10%, від порівняє однаковими (як наприклад, умови вирощування, льної величини, отриманої при зменшенні NADPHвік рослини і т.п.). Зниження складає при цьому, оксидази, що включає поліпептидну послідовність 29 84406 30 № 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22. Особливо Під гомологією між двома поліпептидами робажані такі послідовності, при зменшенні яких зуміють ідентичність послідовності амінокислот по ефективність стійкості до патогену кількісно перевсій довжині послідовності, що розраховується за вищує порівняльну величину, отриману при знидопомогою порівняння при використанні програмженні NADPH-оксидази, що включає поліпептидну ного алгоритму GAP (Wisconsin Package Version послідовність № 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 10.0, University of Wisconsin, Genetics Computer 22, більш, ніж на 50 %, переважно, на 100%, особGroup (GCG), Madison, USA; Altschul та ін. (1997) ливо переважно на 500%, найбільш переважно на Nucleic Acids Res. 25:3389ff) при встановленні на1000%. ступних параметрів: Вага гепів: 8 Вага послідовноПорівняння проводиться переважно у порівнясті за виключенням гепів: 2 них умовах. Під "порівняними умовами" варто роСереднє значення відповідності: 2,912 Середзуміти те, що всі загальні умови, як наприклад, нє значення невідповідності: 2,003 умови вирощування або умови племінної роботи, Під послідовністю білка, що має гомологію, умови дослідження (такі, як, наприклад, буфер, принаймні, у 80% з послідовністю №2, розуміють, температура, субстрат, концентрація патогену і наприклад, таку послідовність, що при порівнянні з т.п.) між порівнюваними тестами підтримуються послідовністю №2 відповідно до вищенаведеного ідентичними, а реакційні суміші відрізняються тільпрограмного алгоритму з вищенаведеними параки послідовністю порівнюваних NADPH-оксидаз, метрами має гомологію, принаймні, у 80%. організмом, з якого їх одержаної, у разі потреби, Функціональні еквіваленти, що одержані від патогеном. При виборі патогену для порівняння NADPH-оксидази, яка включає поліпептидну посварто вибирати патоген, найбільш близький до лідовність відповідно до послідовностей № 2, 4, 6, іншого, при урахуванні специфіки виду. 8, 10, 12, 14, 16, 18, 20 або 22, за допомогою заПід "функціональним еквівалентом" розуміють, міщення, інсерції або делеції, мають гомологію, зокрема, природні або штучні мутації NADPHпринаймні, у 60%, бажано, принаймні, 80%, бажаоксидаз, що включають поліпептидну послідовно, принаймні, 90%, особливо бажано, принаймні, ність відповідно до послідовностей № 2, 4, 6, 8, 10, 95%, дуже бажано, принаймні, 98% до одного з 12, 14, 16, 18, 20 або 22, а також гомологічні поліполіпептидів, що включає поліпептидну послідовпептиди з інших рослин, що мають в основному ність відповідно до послідовностей № 2, 4, 6, 8, 10, однакові властивості. Перевага надається гомоло12, 14, 16, 18, 20 або 22 і характеризуються такими гічним поліпептидам з описаних вище переважних ж властивостями. рослин. Послідовності з інших рослин (наприклад, Функціональні еквіваленти, що одержані від з Arabidopsis thaliana), гомологічні розкритим у послідовності нуклеїнової кислоти NADPHрамках даного винаходу послідовностям NADPHоксидази, що включає послідовність згідно № 1, 3, оксидази, можуть бути легко знайдені, наприклад, 5, 7, 9, 11, 13, 15, 17, 19 або 21, шляхом заміщенпошуком у банках даних або перевіркою банків ня, інсерції або делеції, мають гомологію, принайгенів при застосуванні послідовностей NADPHмні, 60%, бажано 80%, бажано, принаймні, 90%, оксидази як пошукових послідовностей, відповідособливо бажано, принаймні, 95 %, дуже бажано, но, зондів Відповідні послідовності приведені вище принаймні, 98 % до поліпептиду за винаходом відз номером банку генів GenBank Ace- No. повідно до послідовностей №: 1, 3, 5, 7, 9, 11, 13, Мутації включають заміщення, приєднання, 15, 17, 19 або 21, та кодують поліпептиди з такими делеції, інверсії або інсерції одного або декількох ж властивостями, що і поліпептид, який включає амінокислотних залишків. Таким чином, винаходом послідовність № 2, 4, 6, 8,10, 12, 14, 16, 18, 20 або охоплюються також і такі поліпептиди, які забезпе22. чують модифікацію поліпептиду, що включає поліПеревірка кДНК- або геномних бібліотек інших пептидну послідовність відповідно до послідовносорганізмів, переважно приведених нижче, що є тей № 2, 4, 6, 8, 10, 12, 14, 16, 18, 20 або 22. придатними як хазяйські організми для трансфорПід гомологією між двома послідовностями нумації видів рослин, із застосуванням описаних у клеїнової кислоти розуміється ідентичність посліпослідовностях № 1, 3, 5, 7, 9, 11, 13, 15, 17, 19 довності нуклеїнової кислоти по всій довжині посабо 21 послідовностей нуклеїнової кислоти або їх лідовності, що розраховується за допомогою фрагментів як зондів, є відомою фахівцеві в даній порівняння при використанні програмного алгориобласті способом для ідентифікації гомології в тму GAP (Wisconsin Package Version 10.0, інших видах. При цьому одержані від послідовносUniversity of Wisconsin, Genetics Computer Group тей нуклеїнової кислоти відповідно до послідовно(GCG), Madison, USA; Altschul та ін. (1997) Nucleic стей № 1, 3, 5, 7-і 9, 11, 13, 15, 17, 19 або 21 зонди Acids Res. 25:3389ff) при встановленні наступних мають довжину, принаймні, 20п.о, бажано, припараметрів: наймні, 50п.о., особливо бажано, принаймні, Вага гепів: 50 Вага послідовності за виключен100п.о., дуже бажано, принаймні, 200п.о., найням гепів: 3 більш бажано, принаймні, 400п.о. Для перевірки Середнє значення відповідності: 10 Середнє бібліотек може також застосовуватися ДНКзначення невідповідності: 0 ланцюг, комплементарна до описаних в № 1, 3, 5, Наприклад, під послідовністю нуклеїнової кис7, 9, 11, 13, 15, 17, 19 або 21 послідовностей. лоти, що має гомологію, принаймні, 80%, розуміФункціональні еквіваленти включають посліють таку послідовність, що при порівнянні з послідовності ДНК, що при стандартних умовах гібридовністю 1 відповідно до вищенаведеного дизуються з послідовностями № 1, 3, 5, 7, 9, 11, програмного алгоритму з вищенаведеними пара13, 15, 17, 19 або 21 нуклеїнової кислоти NADPHметрами має гомологію, принаймні, у 80%. оксидази, з комплементарною до неї послідовніс 31 84406 32 тю нуклеїнової кислоти або з фрагментами вищеоксидази може здійснюватися різними ми спосонаведених послідовностей і як повні послідовності бами. кодують білки, які мають ті ж самі істотні властивоПоняття "зменшення" або "зниження" може бусті, що й поліпептид, який включає послідовності ти широко інтерпретоване в зв'язку з білком № 2, 4, 6, 8, 10, 12, 14, 16,18, 20 або 22. NADPH-оксидази, активністю NADPH-оксидази або Під "стандартними умовами гібридизації" слід функцією NADPH-оксидази та включає часткове розуміти в широких межах жорсткі, а також менш або повне, відключення або блокування функціожорсткі умови гібридизації. Такі умови гібридизації нальності білка NADPH-оксидази в рослині або описані, серед інших, у [публікації Sambrook J, частині, що походить від неї, тканині, органі, клітиFritsch EF, Maniatis T та ін., in Molecular Cloning (A ні або насінні, що ґрунтується на різних клітинноLaboratory Manual), друге видання, Cold Spring біологічних механізмах. Harbor Laboratory Press, 1989, Seiten 9.31-9.57) або Зменшення або зниження в контексті винаходу в публікації Current Protocols in Molecular Biology, включає кількісне зниження білка NADPH-оксидази John Wiley and Sons, N.Y. (1989), 6.3.1-6.3.6]. до суттєво повної відсутності білка NADPHНаприклад, умови під час стадії промивання оксидази (тобто, відсутність доказу активності можуть бути вибрані з умов обмеженої зниженої NADPH-оксидази, відповідно, функції NADPHжорсткості (приблизно з 2Х SSC при 50°C) та підоксидази або відсутність імунологічного доказу вищеної жорсткості (приблизно з 0,2Х SSC при наявності білка NADPH-оксидази). При цьому екс50°C, переважно, при 65°C) (2OX SSC означає: пресія визначеного білка NADPH-оксидази або 0,3моля цитрату натрію, 3моля NaCI, рН 7,0). Крім активності NADPH-оксидази, відповідно, функції того, температура під час стадії промивання може NADPH-оксидази в клітині або організмі, знижена бути підвищена з умов низької жорсткості при кімпереважно більше, ніж на 50%, особливо переванатній температурі, приблизно при 22°С, до більш жно більше, ніж на 80%, особливо переважно бівисоко жорстких умов приблизно при 65°С. Обидльше, ніж на 90%. ва параметри, концентрація солі і температура Відповідно до винаходу охоплені різні способи можуть одночасно варіюватися, один з двох паразниження експресії білка NADPH-оксидази, активметрів також може підтримуватися постійним і ності NADPH-оксидази або функції NADPHваріюватися тільки інший Під час гібридизації мооксидази. Фахівець у даній області знає, що в його жуть застосовуватися денатуруючі реагенти, такі, розпорядженні наявний цілий ряд методів, щоб як, наприклад, формамід або додецилсульфат бажаним чином вплинути на експресію білка натрію (SDS). У присутності 50% формаміду гібриNADPH-оксидази, активність NADPH-оксидази або дизацію проводять переважно при температурі функцію NADPH-оксидази. 42°С. Деякі приклади умов гібридизації і стадії Зниження кількості білка NADPH-оксидази, акпромивання наведені нижче: тивності NADPH-оксидази або функції NADPH(1) Умови гібридизації можуть бути вибрані, оксидази може здійснюватися, наприклад, при наприклад, з наступних: застосуванні наступних способів, які не обмежують a) 4Х SSC при 65°C (у разі потреби, з даний винахід. 100мкг/мл денатурованої, фрагментованоїДНКзі a) введення послідовності нуклеїнової кислоти сперми риб), дволанцюгової PHK NADPH-оксидази (NAoxb) 6X SSC при 45°C, (у разі потреби, з dsRNA) або експресійної касети або експресійних 100мкг/мл денатурованої, фрагментованої ДНК зі касет, що забезпечують її експресію ; сперми риб), b) введення антисмислових послідовностей c) 6Х SSC, 0,5% додецилсульфату натрію, нуклеїнової кислоти NADPH-оксидази або експре50% формаміду при 42°С (у разі потреби, з сійної касети, що забезпечує її експресію. Вклю100мкг/мл денатурованої, фрагментованоїДНКзі чаються такі способи, при яких антисмислова поссперми риб), лідовність нуклеїнової кислоти орієнтована проти d) 4X SSC, 50% формаміду при 42°С (у разі гена NADPH-оксидази (тобто, геномної послідовпотреби, з 100мкг/мл денатурованої, фрагментоності ДНК) або проти транскрипта гена NADPHваної ДНК зі сперми риб), оксидази (тобто, послідовності PHK). Включені e) 2Х або 4Х SSC при 50°C (умови слабкої жотакож і а-аномерні послідовності нуклеїнової кисрсткості), лоти. f) 30-40% формаміду , 2Х або 4Х SSC при c) введення антисмислової послідовності нук42°С(умови слабкої жорсткості). леїнової кислоти NADPH-оксидази, комбінованої з (2) Умови стадії промивання можуть бути вибрибозимом, або експресійної касети, що забезперані, наприклад, з наступних: чує її експресію. а) 0,015M NaCI/0,0015M цитрату натрію /0,1% d) введення смислових послідовностей нукледодецилсульфату натрію при 50°C. їнової кислоти NADPH-оксидази для індукції косуb) 0,1XSSC при 65°С. пресії або експресійної касети, що забезпечує їх c) 0,1Х SSC, 0,5% додецилсульфату натрію, експресію. 50% формаміду при 42°С. e) введення зв'язувальних ДНК або білкових d) 0,2X SSC, 0,1% додецилсульфату натрію факторів, спрямованих проти генів NADPHпри 42°C оксидази, її PHK або білків, або експресійної касеe) 2Х SSC при 65°C (умови слабкої жорсткости, що забезпечує їх експресію. ті). f) введення вірусних послідовностей нуклеїноЗниження експресії білка NADPH-оксидази, аквої кислоти, що викликають деструкцію PHK та тивності NADPH-оксидази або функції NADPH 33 84406 34 експресійних конструктів NADPH-оксидази, або а) один «смисловий» ланцюг PHK, що вклюекспресійної касети, що забезпечує їх експресію чає, принаймні, одну рибонуклеотидну послідовg) введення конструктів для індукції гомологічність, яка в основному ідентична, принаймні, часної рекомбінації на ендогенних генах NADPHтині послідовності нуклеїнової кислоти NADPHоксидази, наприклад, для одержання нокаутних оксидази і мутантів. b) «антисмисловий» ланцюг РНК, що в основh) ведення мутацій в ендогенні гени NADPHному комплементарний, бажано, цілком комплеоксидази для одержання втрати функції (наприментарний, смисловому ланцюгу PHK згідно з а). клад, для генерування стоп-кодона, зсувів у рамці При ще одній із бажаних форм виконання визчитування і т.п.). находу молекула дволанцюгової PHK для зниженПри цьому кожний окремий із приведених споня експресії білка NADPH-оксидази включає собів може приводити до зниження експресії a) "смисловий" РНК-ланцюг, що включає, приNADPH-оксидази, активності NADPH-оксидази або наймні, одну рибонуклеотидну послідовність, що в функції NADPH-оксидази в контексті даного винаосновному ідентична, принаймні, частині "смислоходу. Є можливим також і комбіноване застосувого" РНК-транскрипту послідовності нуклеїнової вання. Інші методи відомі фахівцеві в даній обласкислоти, яка кодує білок NADPH-оксидази і ті і можуть включати обмеження або пригнічення b) "антисмисловий" РНК-ланцюг, що в основпроцесування білка NADPH-оксидази, транспорту ному комплементарний, переважно, цілком комбілка NADPH-оксидази або його мРНК, інгубування плементарний, смисловому РНК-ланцюгу згідно з приєднання рибосом, інгубування РНК-сплайсинга, а) індукцію ферменту, що розщеплює (піддає деструУ відношенні молекул дволанцюгової PHK як кції) PHK, NADPH-оксидазу та/або інгібування трапослідовність нуклеїнової кислоти NADPHнсляційної елонгації або термінації. оксидази бажаною є послідовність відповідно до Окремі бажані способи коротко описуються послідовностей 1, 3, 5, 7, 9, 11,13,15, 17, 19 або нижче. 21. а) Введення послідовності нуклеїнової кислоти Під виразом "в основному ідентична" розумідволанцюгової PHK NADPH-оксидази (NAox ють те, що дволанцюгова РНК-послідовність dsRNA) (dsRNA) може мати також й інсерції, делеції, а Спосіб регуляції гена за допомогою дволанцютакож окремі точкові мутації у порівнянні з цільогової PHK ("double-stranded RNA interference"; вою послідовністю NADPH-оксидази або функціоdsRNAi) неодноразово описаний у зв'язку з тванально еквівалентною послідовністю й усе-таки ринними і рослинними організмами [див., напризабезпечувати ефективне зниження експресії. Пеклад, публікації Matzke MA і ін. (2000) Plant Mol ВіоІ реважно гомологія по вищенаведеному визначен43:401-415; Fire A. і ін (1998) Nature 391:806- 811; ню складає, принаймні, 75%, переважно, принайWO 99/32619; WO 99/53050; WO 00/68374; WO мні, 80%, особливо переважно, принаймні, 90% 00/44914; WO 00/44895; WO 00/49035; WO найбільш переважно 100% між "смисловим" лан00/63364]. На приведені в цих публікаціях способи цюгом інгібувальної дволанцюгової PHK і, принайі методи тут дається посилання. Ефективна супремні, частиною "смислового" РНК-транскрипта поссія генів може мати місце також і при тимчасовій лідовності нуклеїнової кислот, що кодує білок експресії або після тимчасової трансформації, NADPH-оксидази або його функціональний еквіванаприклад, внаслідок біолістичної трансформації лент (відповідно, між "антисмисловим" ланцюгом і [див. Schweizer P і ін. (2000) Plant J 2000 24: 895комплементарним ланцюгом послідовності нуклеї903]. Способи регуляції гена за допомогою дволанової кислоти, що кодує білок NADPH-оксидази нцюгової PHK ґрунтуються на тому феномені, що або його функціональний еквівалент). при одночасному введенні комплементарного лаДовжина часткового відрізка складає, принайнцюга і протиланцюга (ланцюга-) транскрипта гена мні, 10 основ, переважно, принаймні, 25 основ, одержують високоефективне пригнічення експресії особливо переважно, принаймні, 50 основ, особвідповідного гена. Отриманий фенотип дуже схоливого переважно, принаймні, 100 основ, найжий з таким для відповідного нокаутного мутанту більш переважно, принаймні, 200 основ або, при[див. публікацію Waterhouse PMi ін. (1998) Proc наймні, 300 основ. Natl Acad Sci USA 95:13959-64]. Альтернативно "в основному ідентична" двоСпосіб регуляції гена за допомогою дволанцюланцюгова PHK може також бути визначена як гової PHK (dsRNAi) виявив себе особливо ефектипослідовність нуклеїнової кислоти, що здатна гібвним для зниження експресії NADPH-оксидази. Як ридизуватися з частиною накопичувального білка приклад, серед інших, композиції dsRNAi, що опигенного транскрипта (наприклад, у 400ммоль NaCI, суються в [WO 99/32619], чітко перевершують кла40ммоль піперазин-N,N'-біс(2-етансульфокислоти) сичні антисмисловіі композиції. (PIPES) pH 6,4, 1ммоль етилендіамінотетраоцтоЩе один об'єкт винаходу відноситься до молевої кислоти при 50°C або 70°C протягом 12 до 16 кул дволанцюгової PHK (dsRNA-молекул), що при годин). введенні в рослину (або в одержану від неї клітиПід "в основному комплементарна" розуміють ну, тканину, орган або насіння) приводять до зните, що антисмисловий РНК-ланцюг може мати таження продукції NADPH-оксидази. кож й інсерції, делеції, а також окремі точкові муМолекула дволанцюгової PHK для зниження тації у порівнянні з комплементом смислового експресії білка NADPH-оксидази відрізняється тим, РНК-ланцюга. Переважно гомологія між антисмисщо наявні ловим РНК-ланцюгом і комплементом смислового РНК-ланцюга складає, принаймні, 80%, переваж 35 84406 36 но, принаймні, 90%, особливо переважно, принайc) схрещуванням двох рослин, кожна з яких мні, 95%, у більшості випадків, переважно 100%. трансформована одним вектором, причому одна Під "частиною смислового РНК-транскрипта" включає експресійні касети зі "смисловим" ланцюпослідовності нуклеїнової кислоти, що кодує білок гом, а інша включає експресійні касети з "антисмиNADPH-оксидази або його функціональний еквівасловим" ланцюгом. лент, розуміють фрагменти PHK або мРНК, що Утворення РНК-дуплекса може ініціюватися транскрибовані з використанням кодуючої посліабо поза клітиною або усередині неї. Як описано в довності білка NADPH-оксидази або її функціона[WO 99/53050], дволанцюгова PHK може охоплюльного еквіваленту, переважно, геном NADPHвати шпилькову структуру, в якій "смисловий" і оксидази. При цьому фрагменти мають переважно "антисмисловий" ланцюги зв'язані „лінкером" (надовжину послідовності, принаймні, 20 основ, переприклад, інтроном). Бажаними є аутокомплеменважно, принаймні, 50 основ, особливо переважно, тарні структури дволанцюгової PHK, оскільки вони принаймні, 100 основ, особливого переважно, вимагають експресії одного конструкта та включапринаймні, 200 основ, у більшості випадків, приють комплементарні ланцюги завжди в еквімолярнаймні, 500 основ. Сюди включені також і цілком ному співвідношенні. транскрибовані PHK або мРНК. Експресійні касети, що кодують "антисмислоПри цьому мається на увазі також і застосувий" або "смисловий" ланцюг дволанцюгової PHK вання молекул дволанцюгової PHK за винаходом або аутокомплементарний ланцюг дволанцюгової в способі за винаходом для одержання стійкості до PHK вбудовують переважно у вектор та стабільно патогену у рослин. вбудовують описаними нижче способами (наприДволанцюгова PHK може складатися з одного клад, із застосуванням селективних маркерів) у або декількох ланцюгів полімеризованих рибонукгеном рослини, щоб забезпечити довготривалу леотидів. Можуть також бути наявними модифікаекспресію дволанцюгової PHK. ції як цукор-фосфатного кістяка, так і нуклеозидів. Дволанцюгова PHK може вводитися в кількосТак, наприклад, фосфодіефірні складні зв'язки ті, що дозволяє одержати, принаймні, одну копію природної PHK можуть бути модифіковані таким на клітину. Більш високі кількості (наприклад, причином, що вони охоплюють, принаймні, азот або наймні, 5, 10, 100, 500 або 1000 копій на клітину) гетероатом сірки. Основи можуть бути модифікоможуть забезпечити ефективне зниження. вані таким чином, що, обмежується активність, Як уже було згадано вище, 100%-ої ідентичнонаприклад, аденозиндезамінази. Ці та інші модисті дволанцюгової PHK із генним транскриптом фікації описані нижче при описуванні способів стаNADPH-оксидази функціонально еквівалентного білізації антисмислової PHK. гена не є обов'язковою для того, щоб досягти ефеПриродно, що для досягнення тієї ж мети в ктивного зниження експресії NADPH-оксидази. клітину або організм можуть бути введені також Тому перевага полягає в тому, що спосіб допускає кілька індивідуальних молекул дволанцюгової відхилення послідовностей, які одержані внаслідок PHK, що включають один з вищенаведених відрізгенетичних мутацій, поліморфізмів або еволюційків послідовності рибонуклеїнової кислоти. них дивергенцій. Таким чином, наприклад, можлиДволанцюгова PHK може бути отримана ферво за допомогою дволанцюгової PHK, генерованої ментативним, а також частково або повністю хіміз використанням послідовності NADPH-оксидази чно-синтетичним способом. одного організму, інгібувати експресію NADPHДволанцюгова структура PHK може бути утвооксидази в іншому організмі. Висока гомологія між рена, з використанням двох комплементарних, послідовностями NADPH-оксидази з рису, кукуруокремих РНК-ланцюгів або, що переважно, з викодзи та ячменю має як наслідок високий ступінь ристанням одиничного, аутокомплементарного консервативності цього білка у рослині, так, що РНК-ланцюга. експресія дволанцюгової PHK, яка походить від При одиничному, аутокомплементарному ланоднієї з описаних послідовностей NADPH-оксидази цюгу 'смислова" і 'антисмислова" послідовності згідно 1, 3, 5, 7, 9, 11, 13, 15, 17,19 або 21, має можуть бути зв'язані сполучною послідовністю також переважний ефект в інших видах рослин. ("лінкером") та утворюють, наприклад, шпилькову Також внаслідок високої гомології між окремиструктуру. Сполучна послідовність може бути пеми білками NADPH-оксидази та їх функціональниреважно інтроном, що після синтезу дволанцюгоми еквівалентами можна одержати єдину дволанвої PHK видаляється в процесі сплайсингу. цюгову PHK, що генерована виходячи з визначеної Послідовність нуклеїнової кислоти, що кодує послідовності NADPH-оксидази одного організму, дволанцюгову PHK, може мати ще й інші елеменінгібувати експресію інших гомологічних білків ти, як, наприклад, сигнали термінації транскрипції NADPH-оксидази і/або їх функціональних еквіваабо сигнали поліаденілування. лентів того ж організму або ж експресію білків Якщо два ланцюги дволанцюгової PHK повинNADPH-оксидази в інших споріднених видах. Для ні бути об'єднані в одній клітині або в одній рослицієї мети дволанцюгова PHK включає переважно ні, то це може здійснюватися різними способами: ділянки послідовності генного транскрипта a) трансформацією клітини або рослини векNADPH-оксидази, що відповідають консервативтором, що включає обидві експресійні касети, ним ділянкам. Ці згадані консервативні ділянки b) котрансформацією клітини або рослини можуть бути легко одержані з порівняння послідодвома векторами, причому один вектор включає вностей. експресійні касети зі 'смисловим" ланцюгом, а інДволанцюгова PHK може синтезуватися або in ший вектор включає експресійні касети з "антисvivo, або in vitro. Для цього ДНК-послідовність, яка мисловим" ланцюгом. кодує дволанцюгову PHK, можна вводити в екс 37 84406 38 пресійну касету під контролем, принаймні, одного жуть мати довжину, наприклад, у 5, 10, 15, 20, 25, генетичного регуляторного елемента (такого, як 30, 35, 40, 45 або 50 нуклеотидів, проте вони монаприклад, як промотор, енхансер, сайленсер, жуть бути і більш довгими і включати, принаймні, сплайсинг-донор або сплайсинг-акцептор, сигнал 100, 200, 500, 1000, 2000 або 5000 нуклеотидів. поліаденілування). Відповідні бажані конструкції Антисмислові послідовності нуклеїнової кислоти описані нижче. Поліаденілування при цьому не можуть бути рекомбінантно експримовані або синпотрібно, також не обов'язково мати елементи для тезовані хімічно, відповідно, ферментативно з заініціювання трансляції. стосуванням відомих фахівцеві в даній області Дволанцюгова PHK може бути синтезована хіспособів. При хімічному синтезі можуть застосовумічно або ферментативно. Для цього можуть завати природні або модифіковані нуклеотиди. Мостосовуватися клітинні РНК-полімерази або РНКдифіковані нуклеотиди можуть надавати антисмиполімерази бактеріофагів (як, наприклад, Т3-, Т7словим послідовностям нуклеїнової кислоти або SP6 РНК-полімерази). Відповідні способи для підвищену біохімічну стабільність та можуть приекспресії PHK in vitro описані, наприклад, у [патенводити до підвищеної фізичної стабільності дуплетах WO 97/32016; US 5,593,874; US 5,698,425, US кса, утвореного з антисмислової послідовності 5,712,135, US 5,789,214, US 5,804,693]. Синтезонуклеїнової кислоти і смислової цільової послідоввана хімічно або ферментативно дволанцюгова ності. Наприклад, можуть застосовуватися похідні PHK перед її введенням у клітину, тканину або фосфотіоатів і заміщені акридином нуклеотиди, організм може бути цілком або частково отримана такі, як 5-фторурацил, 5-бромурацил, 5з реакційної суміші екстракцією, осадженням, елехлорурацил, 5-йодурацил, гіпоксантин, ксантин, 4ктрофорезом, хроматографією або комбінацією ацетилцитозин, 5-(карбоксигідроксиметил)урацил, цих способів. Дволанцюгова PHK може бути вве5-карбоксиметиламінометил-2-тіоуридин, 5дена безпосередньо в клітину або ж введена позакарбоксиметиламінометилурацил, дигідроурацил, клітинно (наприклад, в інтерстиціальний простір). -D-галактозилквеозин, інозин, N-6Бажана рослина стабільно трансформується ізопентениладенін, 1-метилгуанін, 1-метилінозин, експресійним конструктом, що реалізує експресію 2,2-диметилгуанін, 2-метиладенін, 2-метилгуанін, дволанцюгової PHK. Відповідні способи будуть 3-метилцитозин, 5-метилцитозин, N-6-аденін, 7описані нижче. метил гуанін, 5-метиламінометилурацил, 5b) Введення антисмислової послідовності нукметоксіамінометил-2-тіоурацил, -Dлеїнової кислоти NADPH-оксидази манозилквеозин, 5'-метоксикарбоксиметилурацил, Способи супресії визначеного білка шляхом 5-метоксіурацил, 2-метилтю-N6-ізопентеніладенін, запобігання акумуляції його мРНК за допомогою урацил-5-оксіоцтова кислота, псевдоурацил, квео"антисмислової" технології неодноразово описані зин, 2-тіоцитозин, 5-метил-2-тіоурацил, 2також стосовно до рослин [див. публікації Sheehy і тіоурацил, 4-тіоурацил, 5-метилурацил, складний ін. (1988) Proc Natl Acad Sci USA 85: 8805- 8809; метиленовий ефір урацил-5-оксіоцтової кислоти, US 4,801,340; Mol JN і ін. (1990) FEBS Lett урацил-5-оксіоцтова кислота, 5-метил-2-тіоурацил, 268(2):427-430]. Антисмислова молекула нуклеї3-(3-аміно-3-N-2-карбоксипропіл)урацил та 2,6нової кислоти гібридизується, відповідно, зв'язудіамінопурин. ється з клітинною мРНК та/або геномною ДНК, що При ще одній бажаній формі виконання екскодує цільовий білок NADPH-оксидази, який підляпресія білка NADPH-оксидази може інгібуватися гає супресії. Внаслідок цього пригнічується транспослідовностями нуклеїнової кислоти, що комплекрипція і/або трансляція цільового білка. Гібридиментарні регуляторній ділянці гена NADPHзація може здійснюватися звичайним чином через оксидази (наприклад, промотору та/або енхансеру утворення стабільного дуплекса або, у випадку NADPH-оксидази), та утворюють трьохспіральні геномної ДНК, через зв'язування антисмислової структури з наявною там подвійною спіраллю ДНК, молекули нуклеїнової кислоти з дуплексом геномтак що знижується транскрипція гена NADPHної ДНК за допомогою специфічної взаємодії у оксидази. Відповідні способи описані в літературі великій петлі ДНК-спіралі. [див. наприклад, публікації Helene C (1991) Антисмислова послідовність нуклеїнової кисAnticancer Drug Res 6(6):569- 84; Helene C і ін. лоти, придатна для зниження експресії білка (1992) Ann NY Acad Sci 660:27-36; Maher LJ (1992) NADPH-оксидази, може виробляється при застоBioassays 14(12):807-815]. суванні послідовності нуклеїнової кислоти, що коПри ще одній бажаній формі виконання антисдує цей білок, наприклад, послідовності нуклеїномислова молекула нуклеїнової кислоти може бути вої кислоти, що включає послідовність 1, 3, 5, 7, 9, а-аномерною нуклеїновою кислотою. Подібні 11, 13, 15, 17, 19 або 21, або послідовності нуклеїаномерні молекули нуклеїнової кислоти утворюють нової кислоти, у відповідність із правилом Уотсона специфічні дволанцюгові гібриди з комплементарта Кріка. Антисмислова послідовність нуклеїнової ною PHK, в яких, на відміну від звичайних ркислоти може бути комплементарна усій транскнуклеїнових кислот, обидві ланцюги є паралельні рибованій мРНК відповідного білка, може бути один одному. [див. публікацію Gautier C і ін. (1987) обмежена кодуючою ділянкою, або складатися Nucleic Acids Res 15:6625- 6641]. Антисмислова тільки з олігонуклеотиду, що частково комплеменмолекула нуклеїнової кислоти може також містити тарний кодуючій або некодуючій послідовності 2'-O-метилрибонуклеотиди [див. публікацію lnoue мРНК. Таким чином, олігонуклеотид може бути та ін. (1987) Nucleic Acids Res 15.6131-6148] або комплементарним, наприклад, ділянці, що вклюхимерні аналоги РНК-ДНК [див. lnoue та ін. (1987) чає старт трансляції для відповідного білка. АнтиFEBS Lett 215:327-330]. смислові послідовності нуклеїнової кислоти мо 39 84406 40 с) Введення антисмислової послідовності нук1774; Smith і ін. (1990) Mol Gen Genet 224: 447-481; леїнової кислоти NADPH-оксидази, комбінованої з Napoli і ін. (1990) Plant Cell 2:279- 289, Van der Krol рибозимом. і ін. (1990) Plant Cell 2: 291-99]. При цьому введеБажано, коли вищеописаний антисмисловий ний конструкт може цілком або частково предстаметод може бути поєднаний з рибозимним спосовляти собою гомологічний ген, який піддають знибом. Каталітичні молекули PHK або рибозими можувальній регуляції. При цьому проводити жуть бути підігнані до будь-який цільової PHK та трансляцію не потрібно. Застосування цієї технорозщеплюють фосфодіефірний кістяк у специфічлогії на рослинах описане, наприклад, у [публікації них положеннях, внаслідок чого функціонально Napoli і ін. (1990) The Plant Cell 2: 279-289 або в дезактивується цільова PHK [див. Tanner NK патенті US 5,034,323.] (1999) FEMS Microbiol Rev 23(3):257-275]. Рибозим Переважно косупресія реалізується при застосам внаслідок цього не модифікується, він є здатсуванні послідовності, що в основному ідентична, ним аналогічно розщеплювати ще й інші молекули принаймні, частині послідовності нуклеїнової кисцільової PHK, внаслідок чого він набуває властилоти, яка кодує білок NADPH-оксидази або його вості ферменту. Вбудовування рибозимних посліфункціональний еквівалент, наприклад, послідовдовностей у "антисмислові" PHK надає цим "антиності нуклеїнової кислоти 1, 3, 5, 7, 9, 11, 13, 15, смисловим" PHK ферментативної властивості 17, 19 або 21. розщеплювати PHK та підвищує їх ефективність e) Введення сполучних ДНК або білкових факпри інактивації цільової PHK. Одержання і застоторів проти генів, PHK або білків NADPH-оксидази. сування відповідних рибозим-"антисмислових" Зниження експресії генів NADPH-оксидази моРНК-молекул описано, наприклад, у [публікації жливо проводити також за допомогою специфічHaseloff та ін. (1988) Nature 334: 585-591]. них, сполучних ДНК-факторів, наприклад, типу Таким чином, можуть застосовуватися рибофакторів транскрипції "цинкові пальці". Ці фактори зими [наприклад, "молотоголові" рибозими, з пуброзміщуються на геномній послідовності ендогенлікації Haselhoff та Gerlach (1988) Nature 334:585ного цільового гена, переважно в регуляторних 591], щоб каталітично розщеплювати мРНК ферзонах, і викликають репресію ендогенного гена. менту, який піддають супримуванню, наприклад, Застосування такого способу забезпечує зниження NADPH-оксидази, і перешкоджати трансляції. Техекспресії ендогенного гена NADPH-оксидази без нологія рибозимів може підвищити ефективність необхідності генетичних маніпуляцій з цією посліантисмислової стратегії. Спосіб експресії рибозидовністю. Відповідні способи для одержання відмів для зниження визначених білків описаний у повідних факторів описані в літературі [див. публі[EP 0291533, EP 0321201, EP 0360257]. Експресія кації Dreier B і ін. (2001) J ВіоІ Chem 276(31):29466рибозимів у рослинних клітинах також вже була 78; Dreier B і ін. (2000) J Mol ВіоІ 303(4):489-502; описана [див. публікації Steinecke P і ін. (1992) Beerli RR і ін. (2000) Proc Natl Acad Sci USA 97 (4): EMBO J 11(4):1525-1530; de Feyter R і ін. (1996) 1495-500; Beerli RR і ін. (2000) J ВіоІ Chem Mol Gen Genet. 250(3):329-338]. Придатні цільові 275(42):32617-32627; Segal DJ та Barbas CF 3rd. послідовності та рибозими, як це описано, напри(2000) Curr Opin Chem ВҐоІ 4(1):34-39; Kang JS та клад, у [публікаціях "Steinecke P, Ribozymes, Kim JS (2000) J Biol Chem 275(12):8742-8748; Beerli Methods in Cell Biology 50, Galbraith і ін. ред, RR і ін. (1998) Ргос Natl Acad Sci USA 95(25): Academic Press, Inc. (1995), стор.449-460"], можуть 14628-14633; Кіт JS і ін. (1997) Ргос Natl Acad Sci бути визначені розрахунком вторинних структур USA 94(8):3616-3620; Klug A (1999) J Mol ВіоІ рибозимних та цільових РНК, а також шляхом їх 293(2):215-218; Tsai SY і ін. (1998) Adv Drug Deliv взаємодії, [див. Bayley CC і ін. (1992) Plant Mol ВіоІ. Rev 30(1-3):23-31; Mapp AK і ін. (2000) Ргос Natl 18(2):353- 361; Lloyd AM та Davis RW і ін. (1994) Acad Sci USA 97(8):3930-3935; Sharrocks AD і ін. Mol Gen Genet. 242(6): 653-657]. Так, наприклад, (1997) lnt J Biochem Cell Biol 29(12):1371-1387; можуть бути сконструйовані похідні вбудованої Zhang L і ін. (2000) J Biol Chem 275(43):33850послідовності PHK Tetrahymena L-19 IVS, що ма33860]. ють комплементарні ділянки з мРНК білка NADPHСелекція цих факторів може здійснюватися оксидази, який піддають супримуванню [див. також при застосуванні будь-якої ділянки гена NADPHпатенти US 4,987,071 і US 5,116,742]. Альтернатиоксидази. Бажано, коли ця ділянка знаходиться в вно до цього такі рибозими можуть бути ідентифіобласті промоторної ділянки. Для пригнічення гена ковані шляхом процесу селекції з бібліотеки можвін може знаходитися у ділянці, що кодує екзон ливих рибозимів [див. Bartel D та Szostak JW або інтрон. Відповідні ділянки доступні фахівцеві в (1993) Science 261:1411-1418]. даній області шляхом пошуку в банках генів або, d) Введення смислової послідовності нуклеївиходячи з кДНК NADPH-оксидази, гена якої не нової кислоти NADPH-оксидази для індукції косупмає в банку генів, шляхом перевірки геномних бібресії. ліотек щодо відповідних геномних клонів. Спосіб, Експресія послідовності нуклеїнової кислоти у що необхідний для цього, відомий фахівцеві в дасмисловій орієнтації NADPH-оксидази може приній області. водити до косупресії відповідного гомологічного, Далі, у клітину можуть бути введені фактори, ендогенного гена. Експресія смислової PHK із гоякі самі інгібують цільовий білок NADPH-оксидази. мологією до ендогенного гена може знизити або Факторами, які інгібують білок, можуть бути, навиключити його експресію, аналогічно описаному приклад, аптамери [див. публікацію Famulok M та для антисмислових композицій [див. публікації Mayer G (1999) Curr Top Microbiol Immunol 243:123Jorgensen і ін. (1996) Plant Mol ВіоІ 31(5):957-973; 36] або антитіла, відповідно, фрагменти антитіл, Goring і ін. (1991) Proc Natl Acad Sci USA 88:1770або одноланцюгові антитіла. Одержання цих фак 41 84406 42 торів відоме фахівцеві в даній області. Так, напризмін, проте експресія (транскрипція та/або трансклад, цитоплазматичне scFv-антитіло використоляція) знижується або усувається. вувалося для модуляції активності білка фітохроПри звичайній гомологічній рекомбінації змінему А у генетично змінених рослинах тютюну [див. на ділянка на її 5'- та 3'-кінці фланкована послідопублікації Owen M і ін. (1992) Biotechnology (N Y) вностями нуклеїнової кислоти, які повинні мати 10(7): 790-794; Franken E і ін. (1997) Curr Opin достатню довжину для можливості рекомбінації. Biotechnol 8(4): 411-416; Whitelam (1996) Trend Довжина, як правило, лежить в інтервалі від декіPlant Sci 1: 286-272]. лькох сотень основ до декількох кілооснов [див. Експресія гена може також інгібуватися спеціпублікацію Thomas KR та Capecchi MR (1987) Cell ально розробленими, низькомолекулярними син51: 503; Strepp і ін. (1998) Proc Natl Acad Sci USA тетичними сполуками, наприклад, поліамідного 95(8): 4368-4373]. Для гомологічної рекомбінації типу. [див. Dervan PB та BürIi RW (1999) Current організм-хазяїн, наприклад, рослина, трансформуOpinion у Chemical Biology 3: 688-693; Gottesfeld ється рекомбінаційним конструктом при застосуJM і ін. (2000) Gene Expr 9(1-2): 77-91]. Ці олігомеванні описаних нижче способів, а клони, які успішри складаються з таких компонентів, як 3но рекомбіновані, піддають селекції при (диметиламіно)пропіламін, N-метил-3застосуванні, наприклад, стійкості до антибіотика гідроксипірол, N-метилімідазол та N-метилпіроли, і або гербіциду. можуть підганятися до кожної ділянки дволанцюГомологічна рекомбінація є відносно рідким гової ДНК таким чином, що вони специфічно посявищем у вищих еукаріотах, насамперед, у рослилідовності зв'язують велику петлю та блокують нах. Переважають випадкові інтеграції в геном експресію послідовностей гена в ній. Відповідні хазяїна. Можливість видалення випадково інтегроспособи відомі й описані в літературі [див. серед ваної послідовності та, у такий спосіб, збагачення інших, публікації Bremer RE і ін. (2001) Bioorg Med клітинних клонів правильно здійсненою гомологічChem. 9(8): 2093-103; Ansari AZ і ін. (2001) Chem ною рекомбінантністю, одержують при застосуBiol. 8(6): 583-92; Gottesfeld JM і ін. (2001) J Mol ванні специфічної послідовності рекомбінантної ВіоІ. 309(3): 615-29; Wurtz NR і ін. (2001) Org Lett системи, описаної в [US 6,110,736], за допомогою 3(8): 1201-3; Wang CC і ін. (2001) Bioorg Med Chem якої можуть бути знову делетовані неспецифічно 9(3): 653-7; Urbach AR та Dervan PB (2001) Ргос інтегровані послідовності, це полегшує проведенNatl Acad Sci USA 98(8): 4343-8; Chiang SY і ін. ня селекцію успішно інтегрованих послідовностей (2000) J Biol Chem. 275(32): 24246-54]. за допомогою явищ гомологічної рекомбінації. Моf) Введення вірусних послідовностей нуклеїноже застосовуватися безліч специфічних до послівої кислоти та експресійних конструкцій, що виклидовностей рекомбінантних систем, наприклад, кають деструкцію PHK NADPH-оксидази ними може служити Сrе/Іох-система бактеріофага Експресія NADPH-оксидази може бути ефекР1, FLP/FRT-система дріжджів, Gin-рекомбіназа тивно реалізована за допомогою індукції специфіфага , Pin-рекомбіназа з E. coli і R/RS - система чної деструкції PHK NADPH-оксидази в рослині за pSR1-плазміди. Бажаною є система Cre/Іох бактедопомогою вірусної експресійної системи ріофага Р1 і FLP/FRT-система дріжджів. Рекомбиі(Amplikon) [Angell, SM і ін. (1999) Plant J. 20(3): нантні системи FLP/FRT і сrе/lox уже застосовува357-362]. Ці системи, що позначаються також як лися в рослинних системах [див. публікацію Odell і "VIGS" (viral induced gene silencing), вводять у росін. (1990) Mol Gen Genet 223: 369-378]. линні послідовності нуклеїнової кислоти, які мають h) Введення мутацій в ендогенні гени NADPHгомологію з транскриптами, що піддають супресії оксидази для одержання функціональної втрати за допомогою вірусних векторів. Транскрипція, (наприклад, генерування стоп-кодона, зсуву в ратаким чином, відключається, очевидно, внаслідок мці зчитування і т п.). захисних механізмів проти вірусу, які виникають у Іншими придатними методами зниження актирослині. Відповідні методи описані в літературі вності NADPH-оксидази є введення нонсенс[див. публікації Ratcliff F і ін. (2001) Plant J 25(2): мутацій в ендогенні гени NADPH-оксидази, напри237-45; Fagard M та Vaucheret H (2000) Plant Mol клад, за допомогою введення РНК/ДНКBiol 43(2-3): 285-93; Anandalakshmi R і ін. (1998) олігонуклеотидів у рослину [див. публікацію Zhu і Proc Natl Acad Sci USA 95(22): 13079- 84; Ruiz MT ін. (2000) Nat Biotechnoi 18(5): 555-558], а також (1998) Plant Cell 10(6): 937-46]. генерування нокаутних мутантів за допомогою, g) Введення конструктів для індукції гомологінаприклад, мутагенезу на основі Т-ДНК [див. пубчної рекомбінації ендогенних генів NADPHлікацію Koncz і ін. (1992) Plant Mol ВіоІ 20(5): 963оксидази, наприклад, для одержання нокаутних 976], ENU (N-етил-N-нітрозосечовина)-мутагенезу мутантів. або гомологічної рекомбінації [див. публікацію Для одержання гомологічних рекомбінантних Hohn В та Puchta (1999) H Ргос Natl Acad Sci USA організмів зі зниженою активністю NADPH96: 8321-8323]. Точкові мутації можуть генеруватиоксидази застосовують, наприклад, конструкт нукся також і за допомогою ДНК-РНК-гібридів, що леїнової кислоти, що містить, принаймні, частину відомі також як "химеропласти" [див. публікацію ендогенного гена NADPH-оксидази, який за допоCole-Strauss і ін. (1999) Nucl Acids Res 27(5): 1323могою делеції, приєднання або заміщення, при1330; Kmiec (1999) Gene Therapy American наймні, одного нуклеотида змінюється таким чиScientist 87(3): 240-247]. ном, що функціональність знижується або цілком Методи регуляції гена за допомогою дволанусувається. Зміна може стосуватися також і регуцюгової PHK (dsRNAi), косупресії за допомогою ляторних елементів (наприклад, промоторів) гена, смислової PHK і "VIGS" ("індукованого вірусом так, що кодуюча послідовність залишається без мовчання генів") позначають також як "пост 43 84406 44 транскрипційне мовчання генів" (PTGS). PTGSклад, дволанцюгової PHK NADPH-оксидази або способи, а також зниження функції NADPH"антисмислової PHK NADPH-оксидази переважно оксидази або активності домінантно-негативних в рослині або його частині, тканині, органі або наваріантів NADPH-оксидази є особливо бажаним, сінні. оскільки вимоги до гомології між ендогенним геУ зазначених експресійних конструктах моленом, який піддають супресії, та трансгенно екскула нуклеїнової кислоти, експресія якої (транскпримованою смисловою або дволанцюговою РНКрипція і, у разі потреби, трансляція) генерує "антипослідовністю (наприклад, між ендогенним геном NADPН-оксидаза"-сполуки, знаходиться переважта його домінантно-негативним варіантом) є більш но у функціональній залежності від, принаймні, низькими, ніж, наприклад, у випадку класичної одного генетично регуляторного елемента (наприантисмислової суміші. Відповідні критерії гомології клад, промотора), що забезпечує експресію в орприведені при описуванні способу длРНКі взагалі ганізмі, переважно, у рослині. Якщо експресійний переносяться на спосіб PTGS або домінантноконструкт повинен бути введений безпосередньо в негативні суміші. Внаслідок високої гомології між рослину і "анти-NADPН-оксидаза"-сполуки (наприбілками NADPH-оксидази з кукурудзи, рису і ячмеклад, дволанцюгова PHK NADPH-оксидази) поню можна припускати високий ступінь консервативинні там вироблятися, то кращими є специфічні вності цього білка в рослинах. У такий спосіб при для рослини генетичні регуляторні елементи (назастосуванні послідовностей нуклеїнової кислоти приклад, промотори). "Анти-NADPН-оксидаза"NADPH-оксидази з кукурудзи, рису або ячменю сполуки можуть, проте, вироблятися ще й в інших може бути також ефективно супримована експреорганізмах або in vitro і тоді вводитися в рослину сія гомологічних білків NADPH-оксидази в інших (як описано, у прикладах 6 і 7). У цьому випадку видах без того, щоб було обов'язково необхідне бажаними є всі прокаріотичні та еукаріотичні геневиділення та визначення структури наявних там тичні регуляторні елементи (наприклад, промотогомологів NADPH-оксидази. Це значно полегшує ри), що дозволяють проводити експресію у вибраробочий процес, аналогічно можна при застосуному для виробництва організмі. ванні домінантно-негативних варіантів білка Під функціональним зв'язком розуміють, наNADPH-оксидази з рису, кукурудзи або ячменю приклад, послідовне розташування промотору ефективно знижувати або інгібувати функстосовно послідовності нуклеїнової кислоти, що цію/активність його гомологів в інших видах роспіддається експресії (наприклад, "анти-NADPНлин. оксидаза"-сполуки) і, у разі потреби, стосовно інУсі речовини та сполуки, що безпосередньо ших регуляторних елементів, таких, як термінатор, або опосередковано викликають зниження кількотак, що кожний з регуляторних елементів може сті білка, кількості PHK, активності гена або активвиконувати свою функцію при трансгенній експреності білка NADPH-оксидази, позначені нижче як сії послідовності нуклеїнової кислоти, в залежності "анти-NADPН-оксидаза"-сполуки. Поняття "антивід положення послідовностей нуклеїнової кислоти NADPH-оксидаза''-сполуки включає застосовувані стосовно смислової або антисмислової PHK. Для у вищенаведених способах послідовності нуклеїцього не обов'язково потрібний безпосередній нової кислоти, пептиди, білки або інші фактори. зв'язок у хімічному контексті. Генетичні регуляторні Поняття "введення" у рамках даного винаходу послідовності, такі, як, наприклад, енхансерні посвключає всі способи, що придатні для введення лідовності, можуть виконувати свою функцію щодо "анти-NADPН-оксидаза"-сполуки безпосередньо цільової послідовності також і з більш віддалених або опосередковано в рослину або клітину, частиположень або навіть з інших ДНК-молекул. Бажану рослини, тканину, орган або насіння, або для ними є положення, в яких послідовність, що підгенерування їх там. При цьому маються на увазі як дають трансгенній експресії, розташована за посбезпосередні, так і опосередковані способи. Ввелідовністю, що діє як промотор, так, що обидві дення може приводити до тимчасової (минучої) послідовності ковалентно зв'язані одна з одною. присутності "анти-NADPH-оксидаза"-сполуки (наБажана відстань між промоторною послідовністю і приклад, дволанцюгової PHK) або ж до тривалої послідовністю, яку піддають трансгенній експресії, (стабільної). складає менше, ніж 200 пар основ. Особливо баУ відповідності з різною природою вищеописажано використовувати менше, ніж 100 пар основ, них сумішей "анти-NADPH-оксидаза"-сполуки модуже бажано менше, ніж 50 пар основ. жуть виконувати свою функцію безпосередньо Одержання функціонального зв'язку, а також (наприклад, за допомогою інсерції в ендогенний одержання експресійної касети може бути реаліген NADPH-оксидази). Функція може виконуватися зоване за допомогою звичайної техніки рекомбінатакож і опосередковано після транскрипції в PHK ції та клонування, як вони описані, наприклад, у (наприклад, при антисмислових композиціях) або [публікаціях Maniatis T, Fritsch EF та Sambrook J після транскрипції та трансляції білка (наприклад, (1989) Molecular Cloning: A Laboratory Manual, Cold при сполучних факторах). "Анти-NADPНSpring Harbor Laboratory, Cold Spring Harbor (NY), в оксидаза"-сполуки, які діють як безпосередньо, так Silhavy TJ, Berman ML та Enquist LW (1984) і опосередковано, включені в даний винахід. Experiments with Gene Fusions, Cold Spring Harbor Введення включає, наприклад, такі способи, як Laboratory, Cold Spring Harbor (NY), в Ausubel FM трансфекція, трансдукція або трансформація. та ін. (1987) Current Protocols in Molecular Biology, «Анти-NADPН-оксидаза»-сполуки включають, Greene Publishing Assoc. та Wiley lnterscience та таким чином, наприклад, рекомбінантні експресійні Gelvin і ін. (1990) В: Plant Molecular Biology Manual]. конструкти, що обумовлюють експресію (тобто Між обома послідовностями можуть бути розташотранскрипцію і, у разі потреби, трансляцію), напривані також й інші послідовності, які, наприклад, 45 84406 46 мають функцію лінкера з визначеними сайтами конститутивний промотор особливо бажаним є рестрикційного ферменту або сигнального пептипромотор гена нітрілази-1 (nit1) з A. thaliana [див. ду. Інсерція послідовностей може також приводити GenBank Acc.-№:Y07648.2, Nukleotide 2456-4340, до експресії гібридних білків. Бажано, коли експреHillebrand і ін. (1996) Gene 170: 197-200]. сійна касета, що складається зі зв'язку промотора і b) Тканиноспецифічні промотори послідовності нуклеїнової кислоти, яку піддають Бажаними є промотори зі специфічністю до експресії, може бути інтегрована у вектор і може пиляка, насінних зачатків, квіток, листя, стебел, бути вбудована, наприклад, за допомогою транскоріння та насіння. формації в геном рослини. Специфічні до насіння промотори включають, Під експресійної касетою варто також розуміти наприклад, промотор фазеоліну [див. US такі конструкції, при яких промотор, наприклад, за 5,504,200; Bustos MM і ін. (1989) Plant Cell 1(9): допомогою гомологічної рекомбінації, розташову839-53], 2S альбуміну [див. Joseffson LG і ін (1987) ють за ендогенним геном NADPH-оксидази та за J Biol Chem 262: 12196-12201], легуміну [див. допомогою експресії антисмислової PHK NADPHShirsat A і ін. (1989) Mol Gen Genet 215(2): 326оксидази приводить до зниження білка NADPH331], білка USP [невідомий білок насіння; Baumlein оксидази відповідно до винаходу. Аналогічно до H і ін. (1991) Mol Gen Genet 225(3): 459-67], напіцього також і "анти-^АОРН-оксидаза"-сполуки (нанового гена [US 5,608,152; Stalberg K та ін. (1996) приклад, послідовність нуклеїнової кислоти, що L Planta 199: 515-519], білка, що зв'язує сахарозу кодує дволанцюгову PHK NADPH-оксидази або [WO 00/26388], або промотор легуміну В4 [Le4; антисмислову PHK NADPH-оксидази), можуть бути Baumlein H і ін. (1991) Mol Gen Genet 225: 121-128; розташовані за ендогенним промотором таким Baeumlein і ін. (1992) Plant Journal 2(2): 233-9; чином, що виникає той самий ефект. Обидва ріFiedler U і ін. (1995) Biotechnology (NY) 13(10): шення приводять до одержання експресійної касе1090f], промотор олеозину з Arabidopsis [WO ти відповідно до винаходу. 98/45461], Все4-промотор з Brassica [WO Під специфічними для рослин промоторами 91/13980]. Іншими придатними специфічними для варто розуміти в принципі будь-який промотор, що насіння промоторами є промотори генів, що кодуможе регулювати експресію генів, зокрема, чужоють глютенін високої молекулярної ваги "High рідних генів, у рослинах або частинах рослин, кліMolecular Weight Glutenin" (HMWG), гліадин, фертинах, тканинах, культурах. При цьому експресія мент галуження, АДФпірофосфатазу глюкози може бути конститутивною, індукованою або за(AGP-азу) або синтазу крохмалю. Бажаними є лежною від розвитку. промотори, що дозволяють проводити специфічну Бажаними є: для насіння експресію в однодольних рослинах, а) конститутивні промотори таких, як кукурудза, ячмінь, пшениця, жито, рис і Бажаними є вектори, що дозволяють проводит.п. Бажано можуть використовуватися промотори ти конститутивну експресію в рослинах [див. публігена Ipt2 або Ipt1 [WO 95/15389, WO 95/23230] або кацію Benfey і ін.(1989) EMBO J 8: 2195-2202]. Під промотори, описані в [WO 99/16890] (промотори "конститутивним" промотором варто розуміти тагордеїнового гена, глутелінового гена, оризинового кий промотор, що забезпечує експресію в численгена, проламінового гена, гліадининового гена, них, переважно, у всіх тканинах, протягом триваглютелінового гена, зеїнового гена, казиринового лого періоду розвитку рослини, переважно, на всіх гена або секалінового гена). стадіях розвитку рослини. Зокрема, застосовують Специфічні до бульб, накопичувальних коренів промотор рослини або промотор, що походить від або коренів промотори, такі, як, наприклад, проморослинного вірусу. Зокрема, бажаним є промотор тор пататину класу І (В33), промотор інгібітора 35S-транскрипта вірусу мозаїки цвітної капусти катепсину D з картоплі. CaMV [див. публікації Franck і ін. (1980) Cell 21: Специфічні до листя промотори включають, 285-294; Odell і ін. (1985) Nature 313: 810-812; наприклад, цитозольну FBP-азу картоплі [WO Shewmaker і ін. (1985) Virology 140: 281-288; 97/05900], промотор малої субодиниці Rubisco Gardner і ін. (1986) Plant Mol ВіоІ 6: 221-228] або (рибу лоза-1,5-бісфософаткарбоксилази) або пропромотор 19S CaMV [див. US 5,352,605; WO мотор ST-LSI картоплі [Stockhaus і ін. (1989) EMBO 84/02913; Benfey і ін. (1989) EMBO J 8: 2195-2202]. J 8: 2445-2451]. Особливо бажаними є специфічні Іншим придатним конститутивним промотором є до епідерміса промотори, як наприклад, промотор промотор малої субодиниці Rubisco [US гена OXLP (білок, подібний до оксалат-оксидази); 4,962,028], промотор легуміну (GenBank Acc.-№ [Wei і ін. (1998) Plant Mol. ВіоІ. 36:101-112]. X03677), промотор нопалінсинтази з Специфічні до квітів промотори включають, Agrobacterium, TR-подвійний промотор, промотор наприклад, промотор фітоенсинтази [WO OCS (октопінсинтази) з Agrobacterium, промотор 92/16635] або промотор гена P-rr [WO 98/22593]. убіквітину [див. Holtorf S і ін. (1995) Plant Mol ВіоІ Специфічні пилякам промотори включають, 29:637-649], промотор убіквітину 1 [див. наприклад, 5126- промотор [US 5,689,049, US Christensen і ін. (1992) Plant Mol ВіоІ 18:675-689; 5,689,051], промотор glob-l і промотор -зеїну. Bruce та ін. (1989) Proc Natl Acad Sci USA 86:9692с) Промотори, здатні до хімічної індукції 9696], промотор Smas, промотор дегідрогенази Експресійні касети можуть також містити прокоричного спирту [US 5,683,439], промотори вакумотор, здатний до хімічної індукції [див. оглядову олярних підгруп АТФази або промотор збагаченого статтю: Gatz і ін. (1997) Annu Rev Plant Physiol проліном білка з пшениці [WO 91/13991], а також Plant Mol Biol 48: 89-108], за допомогою якого можінші промотори генів, конститутивна експресія яких на керувати експресією екзогенного гена в рослині у рослинах відома фахівцеві в даній області. Як у визначений момент часу. Можуть також застосо 47 84406 48 вуватися подібні промотори, як наприклад, промотюну і Arabidopsis [EP-A 0332104, WO 98/03536] тор PRP1 [Ward і ін. (1993) Plant Mol ВіоІ 22: 361також придатні як індуковані патогеном промотори. 366], промотор, що індукується саліциловою кисБажаними є "кислі PR-5"-(aPR5)-промотори з ячлотою [WO 95/19443], промотор, що індукується меню [див. Schweizer і ін. (1997) Plant Physiol бензолсульфонамідом [EP 0388186], промотор, що 114:79- 88] і пшениці [див. Rebmann і ін. (1991) індукується тетрацикліном [див. Gatz і ін. (1992) Plant Mol ВіоІ 16: 329-331] оскільки вони особливо Plant J 2: 397-404], промотор, що індукується абсспецифічно індукуються патогеном, aPR5-білки цизовою кислотою [EP 0335528], відповідно, пронакопичуються приблизно через 4-6 годин після мотор, що індукується етанолом або циклогексаураження патогеном і виявляють дуже малу фононом [WO 93/21334]. ву експресію [WO 99/66057]. Початок для досягd) Промотори, що індукуються стресом або панення підвищеної специфічності, що індукується тогеном патогеном, покладено одержанням синтетичних Крім того, бажаними є промотори, що індукупромоторів з комбінацій відомих чутливих до патоється біотичним або абіотичним стресом, як нагену елементів [див. Rushton і ін. (2002) Plant Cell приклад, промотори, які індукуються гена патогена 14, 749-762; WO 00/01830; WO 99/66057]. Інші здаPRP1 (відповідно, промотор гена gst1) наприклад, тні до індукці їпатогеном промотори з різних видів з картоплі [WO 96/28561; Ward і ін. (1993) Plant Mol відомі фахівцеві в даній області [EP-A 1165794; ВіоІ 22: 361-366], промотор hsp70- або hsp80- з EP-А 1062356; EP-А 1041148; EP-A 1032684]. томатів, що індукується теплом [US 5,187,267], e) Залежні від розвитку промотори промотор альфа-амілази з картоплі, що індукуєтьІншими придатними промоторами є, наприся холодом [WO 96/12814], промотор PPDK, що клад, промотори, специфічні до зрілості плодів, індукується світлом або pinll-промотор, що індукутакі, як, наприклад, специфічний до зрілості плоду ється пошкодженнями [EP-A 0375091]. Іншими промотор з томатів [WO 94/21794, EP 409625]. промоторами, які індукуються патогеном, є промоЗалежні від розвитку промотори включають частитор Fis1-промотор з льону [WO 96/34949], Vstiну тканиноспецифічних промоторів, оскільки утвопромотор [Schubert і ін. (1997) Plant Mol ВіоІ рення окремих тканин відбувається в залежності 34:417-426], також EAS4 промотор сесквітерпенвід розвитку рослини. циклази з тютюну [US 6,100,451]. Особливо бажаними є конститутивні, а також Промотори, що індукуються патогеном, вклюспецифічні до листя і/або стебел промотори, що чають промотори генів, що індукуються внаслідок індукуються патогеном, специфічні до епідермісу, ураження патогеном, як наприклад, гени PR-білків, причому промотори, які індукуються патогеном, є бажаними в більшій мірі. SAR-білків, -1,3-глюканази, хітинази і т.п. [див. Інші промотори можуть бути функціонально наприклад, публікації Redolfi і ін. (1983) Neth J зв'язані з послідовністю нуклеїнової кислоти, яку Plant Pathol 89: 245-254; Uknes, і ін. (1992) Plant піддають експресії, що дозволяє проводити ексCell 4: 645-656; Van Loon (1985) Plant Mol Viral 4: пресію в інших тканинах рослин або в інших орга111-116; Marineau і ін. (1987) Plant Mol ВіоІ 9: 335нізмах, наприклад, бактеріях Е.соlі. Як промотори 342; Matton і ін. (1987) Molecular Plant- Microbe рослин в основному придатні усі вищенаведені Interactions 2: 325-342; Somssich і ін. (1986) Proc промотори. Natl Acad Sci USA 83: 2427-2430; Somssich і ін. Послідовності нуклеїнової кислоти, що міс(1988) Mol Gen Genetics 2: 93-98; Chen і ін. (1996) тяться в експресійних касетах або векторах за Plant J 10: 955-966; Zhang та Sing (1994) Proc Natl винаходом, можуть бути функціонально зв'язані з Acad Sci USA 91: 2507-2511; Warner, і ін. (1993) іншими генетичними регуляторними послідовносPlant J 3: 191-201; Siebertzi ін. (1989) Plant Cell 1: тями поряд із промоторами. Поняття генетичних 961-968 (1989]). регуляторних послідовностей варто розуміти шиВключені також промотори, які індукуються роко і під ним маються на увазі всі такі послідовпошкодженнями, такі, як промотор pinll-гена [див. ності, що впливають на формування та функцію публікації Ryan (1990) Ann Rev Phytopath 28: 425експресійної касети за винаходом. Генетичні регу449; Duan і ін. (1996) Nat Biotech 14: 494-498], ляторні послідовності модифікують, наприклад, wun1- та wun2-генів [US 5,428,148], win1- та win2транскрипцію і трансляцію в прокаріотичних і еукагенів [Stanford та ін. (1989) Mol Gen Genet 215: 200ріотичних організмах. Переважно експресійні касе208], системіну [див. McGurl і ін. (1992) Science ти за винаходом включають на 5'-проксимальному 225: 1570-1573], WIP1-гена [див. Rohmeier і ін. кінці послідовності нуклеїнової кислоти, яку підда(1993) Plant Mol ВіоІ 22: 783-792; Eckelkamp і ін. ють трансгенній експресії, промотор зі специфічні(1993) FEBS Letters 323: 73-76], МРІ-гена [Corderok стю до ембріонального епідермісу і/або квітки, а на та ін. (1994) Plant J 6(2): 141-150] і т.п. 3'-дистальному кінці послідовність термінації як Джерелом для інших промоторів, що індукудодаткові генетичні контрольної послідовності, а ються патогеном, є родина PR-генів. Ряд елементакож, у разі потреби, інші звичайні регуляторні тів у цих промоторах виявили себе як бажані. Так, елементи, а саме функціонально зв'язані з послінаприклад, область від -364 до -288 у промоторі довністю нуклеїнової кислоти, яку піддають ексPR-2d саліцилат- специфічності [див. Buchel і ін. пресії. (1996) Plant Mol ВіоІ 30, 493-504]. Послідовність 5'Генетичні регуляторні послідовності включаТСАТСТТСТТ-3' повторно виявляється в промотоють також інші промотори, промоторні елементи рі -1,3-глюканази ячменю та у більш, ніж у 30 інабо мінімальні промотори, що можуть модифікуваших генах, що індукується стресом. Ця ділянка ти властивості регуляції експресії. Так, наприклад, зв'язує в тютюні нуклеарний білок, надлишок якого на основі генетичних регуляторних послідовностей підвищується саліцилатом. PR-1-промотори з тю 49 84406 50 тканиноспецифічна експресія може додатково ні фланкуючі послідовності приєднуються до ціздійснюватися в залежності від визначених стрельового гена (Іох-послідовності), що пізніше досових факторів. Відповідні елементи описані, назволяє проводити видалення за допомогою сгеприклад, для водного стресу, стресу абсцизової рекомбінази. кислоти [Lam E та Chua NH, J ВюІ Chem 1991; Експресійна касета та одержані від неї вектори 266(26): 17131-17135] і теплового стресу [Schoffl F можуть містити ще інші функціональні елементи та ін., Molecular and General Genetics 217(2-3): 246Поняття функціональні елементи варто розуміти в 53, 1989]. широкому змісті і під ними маються на увазі всі У принципі можуть застосовуватися всі приротакі елементи, що впливають на одержання, розмдні промотори з їх регуляторними послідовностяноження або функцію експресійних касет, векторів ми, приведеними вище для способу згідно з винаабо трансгенних організмів за винаходом. Як не ходом. Крім того, можуть застосовуватися обмежувальні приклади варто привести наступні: синтетичні промотори. а) селективні маркери, що надають стійкості Генетичні регуляторні послідовності включадо інгібітору метаболізму, такому, як 2ють далі також і 5'-області, що не транслюються, дезоксиглюкоза-6-фосфат [WO 98/45456], антибіоінтрони, або некодуючу 3'-ділянкутенів, як, напритики або біоциди, переважно, гербіциди, як наприклад, інтрон актину-1 , або інтрони 1, 2 та 6 Adh1-S клад, канаміцин, G 418, блеоміцин, гігроміцин або [див. у загальних рисах публікацію: The Maize фосфінотрицин і т.п. Особливо бажані селективні Handbook, Chapter 116, Freeling та Walbot, Eds., маркери представляють собою такі, що додають Springer, New York (1994)]. Було встановлено, що стійкості до гербіцидів. Як приклад варто привести вони можуть мати важливу функцію при регуляції послідовності ДНК, що кодують фосфінотрицинекспресії гена. У такий спосіб було встановлено, ацетилтрансферази (PAT) та інактивують інгібітощо 5'-нетрансльовані послідовності можуть підсири глютамінсинтази (ген bar і pat), гени 5лювати нестійку експресію гетерологічних генів. енолпірувілшикімат-3-фосфатсинтази (гени EPSPПрикладами підсилювачів трансляції є 5'-лідерна синтази), що надають стійкості до гліфосату (Ыпослідовність з вірусу мозаїки тютюну [див. публі(фосфонометил)гліцин), що являє собою ген дох, кацію Gallie і ін. (1987) Nucl Acids Res 15: 8693який кодує ферменти, які розкладають гліфозат 8711] і подібні до неї. Вони можуть сприяти ткани(гліфозатоксидоредуктаза), ген deh (який кодує носпецифічності. [див. Rouster J і ін. (1998) Plant J дегалогеназу, що інактивує далапон), ацетолакта15: 435-440]. тсинтази, які інактивують сульфонілсечовину та Експресійна касета може переважно містити імідазолінон, а також гени bхn, що кодують феродну або декілька так званих "енхансерних посліменти нітрилази, які розкладають бромоксиніл, довностей", функціонально зв'язаних із промотоaasa-ген, що надає стійкості до антибіотика апекром, що забезпечують підвищену трансгенну екстиноміцину, ген стрептоміцинфосфотрансферази пресію послідовності нуклеїнової кислоти. Також і (SPT), що забезпечує стійкість до стрептоміцину, на 3'-кінці послідовностей нуклеїнової кислоти, яку ген неоміцинфосфотрансферази (NPTII), що надає піддають трансгенній експресії, можуть бути вбустійкості до канаміцину та генетицидину, ген гігродовані додаткові послідовності, такі, як інші регуміцинфосфотрансферази (HPT), що надає стійкосляторні елементи або термінальні послідовності ті до гігроміцину, ген ацетолактатсинтази (ALS), Послідовності нуклеїнової кислоти, які піддають що надає стійкості до гербіцидів на основі сультрансгенній експресії, можуть міститися в генному фонілсечовини (наприклад, ALS-варіанти з мутаціконструкті в одній або декількох копіях. єю, наприклад, S4 та/або Нrа). Придатними як контрольні послідовності сигb) репортерні гени, які кодують білки, що легко налами поліаденілування є рослинні сигнали полівизначаються кількісно, та за допомогою власного аденілування, переважно такі, що, в основному, забарвлення або ферментативної активності завідповідають Т-ДНК сигналам поліаденілування з безпечують оцінку ефективності траскрипції або Agrobacterium tumefaciens, зокрема, гена 3 Т-ДНК місця та моменту експресії. Особливо кращі при (октопінсинтази) Ті-плазміди pTiACHS [див. Gielen і цьому є репортерні білки [див публікацію ін. (1984) EMBO J 3:835 ff] або їх функціональні Schenborn E, Groskreutz D. Mol Biotechnol. 1999; еквіваленти. Прикладами особливо придатних 13(1): 29-44], такі, як флуоресціюючий зеленим термінаторних послідовностей є термінатор OCS білок (GFP) [див. Sheen і ін.(1995) Plant Journal (октопінсинтази) і термінатор NOS (нопалінсинта8(5): 777-784; Haseloff і ін. (1997) Proc Natl Acad Sci зи). USA 94(6): 2122-2127; Reichel і ін.(1996) Proc Natl Під контрольними послідовностями варто далі Acad Sci USA 93(12): 5888-5893; Тіаn і ін. (1997) розуміти такі, що дозволяють проводити гомологіPlant Cell Rep 16: 267-271; WO 97/41228; Chui WL і чну рекомбінацію, відповідно, інсерцію в геном ін. (1996) Curr Biol 6:325-330; Leffel SM і ін. (1997) організме хазяїна або дозволяють проводити виBiotechniques. 23(5): 912-8], хлорамфеніколтрансдалення з генома. При гомологічній рекомбінації фераза, люцифераза [див. Ow і ін. (1986) Science природний промотор одного визначеного гена мо234: 856-859; Millar і ін. (1992) Plant Mol Biol Rep же, наприклад, бути замінений промотором зі спе10: 324-414], екворин [див. Prasher і ін. (1985) цифічністю до ембріонального епідермісу та/або Biochem Biophys Res Commun 126(3): 1259-1268], квітки. Такі методи, як cre/lox-технологія дозволя-галактозидаза, ген R-локусу (який кодує білок, ють проводити тканиноспецифічне, видалення що регулює виробництво пігменту антоціану (заекспресійної касети з генома організма хазяїна, що барвлення у червоний колір) у рослинних тканинах індукується при певних умовах [див. Sauer B і в такий спосіб дозволяє проводити безпосередній (1998) Methods. 14(4): 381-92]. При цьому визначеаналіз активності промотора без додання додат 51 84406 52 кових допоміжних або хромогенних речовин, [див. Nature 327: 70-73; Howell і ін. (1980) Science публікації; Dellaporta і ін., In: Chromosome Structure 208:1265; Horsch і ін.(1985) Science 227: 1229and Function: Impact of New Concepts, 18th Stadler 1231; DeBlock і ін. (1989) Plant Physiology 91: 694Genetics Symposium, 11: 263-282, 1988], особливо 701; Methods for Plant Molecular Biology (Weissbach бажана глюкуронідаза [див. Jefferson і ін., EMBO J. та Weissbach, eds.) Academic Press Inc. (1988); та 1987, 6, стор.3901-3907]. Methods in Plant Molecular Biology (SchulerTa c) реплікаційні стрибки, що забезпечують розZielinski, eds.) Academic Press Inc. (1989)]. множення експресійних касет або векторів за виЦі описані способи для трансформації та ренаходом, наприклад, в Е.соlі. Як приклад варто генерації рослин з рослинних тканин або рослинпривести ORI (джерело ДНК-реплікації), pBR322 них клітин використовуються для тимчасової або огі або Р15А оrі [див. Sambrook і ін.: Molecular стабільної трансформації. Придатними методами Cloning. A Laboratory Manual, 2nd ed. Cold Spring є, насамперед, трансформація протопластів шляHarbor Laboratory Press, Cold Spring Harbor, NY, хом індукованого поліетиленгліколем поглинання 1989]. ДНК, біолістичний спосіб за допомогою генної гарd) елементи, що потрібні для опосередкованої мати, так званий метод бомбардування частинкаагробактеріями трансформації рослини, як наприми, електропорація, інкубація сухих ембріонів в клад, праве та ліве обмеження Т-ДНК або virрозчині ДНК або мікроін'єкція. ділянка. Поряд з технікою безпосередньою трансфорВведення експресійної касети за винаходом в мації може здійснюватися трансформація шляхом організм або його клітини, тканини, органи, частибактеріальної ін'єкції агробактеріями Agrobacterium ни або насіння (переважно в рослини, відповідно tumefaciens або Agrobacterium rhizogenes. Опосерослинні клітини, тканини, органи, частини або редкована агробактеріями трансформація найнасіння), може здійснюватися при застосуванні краще підходить для клітин дводольних рослин. векторів, в яких містяться експресійні касети. ЕксТакі способи описані, наприклад, у [публікації пресійна касета може вводитися у вектор (наприHorsch RB і ін. (1985) Science 225: 1229f]. клад, плазміду) через придатний сайт рестрикції. Якщо застосовують агробактерії, то експресійПлазміда, що утворилася, вводиться спочатку в ну касету варто інтегрувати в спеціальну плазміду, Е.соlі. Правильно трансформовані Е.соlі піддають або в проміжний вектор ("shuttle" або "intermediate" селекції, вирощуються та одержують рекомбінантвектор) або бінарний вектор. Якщо для трансфорну плазміду відомими фахівцеві в даній області мації застосовують плазміди Ті або Ri, то, принайметодами. Аналіз рестрикції та секвенування можмні, праве обмеження, проте, в більшості випадків, на використовувати для перевірки ступеня клонуправе і ліве обмеження плазміди Ті або Ri T- ДНК вання. поєднують як фланкуючу ділянку з експресійною Векторами можуть бути, наприклад, плазміди, касетою, яку піддають введенню. косміди, фаги, віруси або ж агробактерії. При баПереважно застосовуються бінарні вектори. жаній формі виконання винаходу введення ексБінарні вектори можуть бути репліковані як в пресійної касети реалізується за допомогою плазЕ.coli, так і в агробактерії. Вони містять, як правимідного вектора. Бажані такі вектори, що ло, селективний маркерний ген та лінкер або полізабезпечують стабільну інтеграцію експресійної лінкер, що фланкований правою та лівою обмежукасети в геном хазяїна. вальною послідовністю Т-ДНК. Вони можуть бути Одержання трансформованого організму (відтрансформовані безпосередньо в агробактерії. повідно, трансформованої клітини або тканини) [див. Holsters і ін. (1978) Mol Gen Genet 163: 181вимагає внесення відповідної ДНК, PHK або білка 187]. Селективний маркерний ген дозволяє провоу відповідну клітину-хазяїн. дити селекцію трансформованих агробактерій і, Для здійснення цього процесу, що називається наприклад, є геном nptll, що надає стійкості до трансформацією (або трансдукцією, відповідно, канаміцину. Виступаючі в цьому випадку як оргатрансфекцією), наявний ряд методів [див. публіканізм-хазяїн агробактерії повинні вже містити плазцію Keown і ін. (1990) Methods in Enzymology 185: міду з vir-областю. Це потрібно для передачі Т527-537]. Так, наприклад, ДНК або РНК може бути ДНК у рослинну клітину. Трансформовані в такий введена безпосередньо мікроін'єкцією або бомбаспосіб агробактерії можуть застосовуватися для рдуванням покритими ДНК-мікрочастинками. Татрансформації рослинних клітин. Застосування Ткож клітина може бути перетворена у хімічноДНК для трансформації рослинних клітин було проникну, наприклад, за допомогою поліетиленгліінтенсивно досліджене та описане [див. публікації колю, так, що ДНК може попадати в клітину за доEP 120516; Hoekema, In: The Binary Plant Vector помогою дифузії. Трансформація ДНК може також System, Offsetdrukkerij Kanters B.V., Alblasserdam, здійснюватися шляхом злиття протопластів з інChapter V; An і ін. (1985) EMBO J 4: 277-287]. Різні шими, елементами, що містять ДНК, такими, як бінарні вектори відомі та частково можуть бути мініклітини, клітини, лізосоми або ліпосоми. Електодержані комерційними шляхом, як, наприклад, ропорація є одним з подальших способів введення рВІ101.2 або pBIN19 (Clontech Laboratories, Inc. ДНК, при якому клітини стають знову проникними USA). за допомогою електричного імпульсу. Відповідні Інші придатні для експресії в рослинах промоспособи описані в літературі [наприклад, у публітори описані, наприклад, у [публікаціях Rogers і ін. каціях Bilang і ін. (1991) Gene 100:247-250; Scheid і (1987) Meth in Enzymol 153: 253-277; Schardl і ін. ін. (1991) Mol Gen Genet 228: 104-112; Guerche і ін. (1987) Gene 61: 1-11; Berger і ін. (1989) Proc Natl (1987) Plant Science 52: 111-116; Neuhause і ін. Acad Sci USA 86: 8402-8406]. (1987) Theor Appl Genet 75: 30-36; Klein і ін. (1987) 53 84406 54 Безпосередні способи трансформації придатні бактерій, нематод і т.п.), стійкість до стресу або для будь-якого організму і типу клітин. інші властивості рослин. Приклади приведені, сеУ випадку ін'єкції або електропорації ДНК, відред інших, у [публікації Dunwell JM, Transgenic повідно, PHK, у рослинні клітини немає ніяких осоapproaches to crop improvement, J Exp Bot. 2000; 51 бливих вимог у відношенні застосовуваної плазміSpec No; стор.487-96]. ди. Можуть застосовуватися прості плазміди, такі, Під поняттям "трансгенний" варто розуміти, як ряд pUC. Якщо з трансформованих клітин понаприклад, щодо послідовності нуклеїнової кисловинні бути регенеровані цілі рослини, то потрібно, ти, експресійної касети або вектора, що містить щоб плазміда містила додатковий, селективний зазначену послідовність нуклеїнової кислоти, або маркерний ген. організму, трансформованого за допомогою цієї Стабільно трансформовані клітини, тобто такі, послідовності нуклеїнової кислоти, експресійної які містять введену ДНК, інтегровану в ДНК клітикасети або вектора, усі такі отримані генетичними ни-хазяїна, можуть відокремлюватися від нетранспособами конструкції, в яких або сформованих, якщо селективний маркер є складоa) послідовність нуклеїнової кислоти NADPHвою частиною введеної ДНК. Як маркер може оксидази або виступати, наприклад, будь-який ген, що може b) функціонально зв'язана з послідовністю нунадавати стійкості до антибіотика або гербіциду клеїнової кислоти NADPH-оксидази генетична ре(такого, як, наприклад, канаміцин, G 418, блеомігуляторна послідовність, наприклад, промотор, цин, гігроміцин або фосфінотрицин і т.п.) (див. виабо ще). Трансформовані клітини, що експримують c) (а) та (b) такий маркер, виживають в присутності концентзнаходяться не в їх природному генетичному рацій відповідного антибіотика або гербіциду, які оточенні або модифіковані генетичними методами, знищують нетрансформований вихідний тип. Припричому модифікацією може бути заміщенням, клади наведені вище і включають переважно ген приєднанням, делецією, інверсією або інсерцією bar, що надає стійкості до гербіциду фосфінотриодного або декількох нуклеотидних залишків. Під цину [див. Rathore KS і ін. (1993) Plant Mol ВіоІ природним генетичним оточенням мають на увазі 21(5): 871-884], ген nptll, що надає стійксті до каприродний хромосомний локус в організмі похонаміцину, ген hpt, що надає стійкості до гігроміцидження або присутність у геномной бібліотеці. У ну, або ген EPSP, що надає стійкості до гербіциду випадку геномної бібліотеки природне генетичне гліфозату. Селективний маркер дозволяє провооточення послідовності нуклеїнової кислоти передити відокремлення трансформованих клітин від важно, принаймні, частково ще збережене. Отонетрансформованих [див. McCormick і ін. (1986) чення фланкує послідовність нуклеїнової кислоти, Plant Cell Reports 5: 81-84]. Отримані рослини мопринаймні, в одному місці і має довжину послідовжуть вирощуватися і схрещуватися звичайним ності, принаймні, 50п.о., переважно, принаймні, способом. Два або три покоління повинні культи500п.о., особливо переважно, принаймні, 1000п.о., вуватися, щоб забезпечити стабільну геномну іннайбільш переважно, принаймні, 5000п.о. Наявна теграцію, яка успадковується. в природі експресійна касета, наприклад, природВищенаведені способи описані, наприклад, у на комбінація промотору NADPH-оксидази з відпо[публікаціях Jenes B і ін.(1993) Techniques for Gene відним геном NADPH-оксидази стає трансгенною Transfer, in: Transgenic Plants, Vol. 1, Engineering експресійною касетою, якщо вона змінена неприand Utilization, видану SD Kung та R Wu, Academic родним, синтетичним (штучним) способом, наприPress, стр.128-143, а також у Potrykus (1991) Annu клад, мутагенезом. Відповідні способи описані в Rev Plant Physiol Plant Molec Biol 42: 205-225]. Балітературі [US 5,565,350; WO 00/15815; див. також жано, коли конструкція, яку піддають експримувище]. ванню, клонується у вектор, що придатний для Ще одним об'єктом винаходу є трансгенний трансформації агробактерій Agrobacterium організм, трасформований, принаймні, однією поtumefaciens, наприклад, рВіn19 [див. Bevan і ін. слідовністю нуклеїнової кислоти за винаходом, (1984) Nucl Acids Res 12:8711 і далі]. експресійною касетою або вектором за винаходом, Якщо створюють трансформовану рослинну а також клітини, клітинні культури, тканини, частиклітину, то можна одержати цільну рослину при ни, як наприклад, у випадку рослинних організмів, застосуванні відомих фахівцеві в даній області листя, коренів і т.п., або матеріалу для розмноспособів. При цьому, наприклад, виходять з калусження, який одержують від таких організмів. Поних культур. З цієї ще не оформленої клітинної няття організм варто розуміти широко і під ним маси можна індукувати утворення пагонів та коремаються на увазі прокаріотичні та еукаріотичні нів відомим способом. Отримані пагони можуть організми, переважно, бактерії, дріжджі, гриби, висаджуватися і вирощуватися. тваринні та рослинні організми. Фахівцеві в даній області відомі також способи Бажаними є для регенерації частин рослин і цілих рослин з a) гриби, такі, як Aspergillus, Eremothecium, клітин рослин. Наприклад, для цього застосовуTrichoderma, Ashbya, Neurospora, Fusarium, ються способи, описані в [публікаціях Fennell і ін. Beauveria або інші, описані в [Indian Chem Eng. (1992) Plant Cell Rep. 11: 567-570; Stoeger і ін Section B. VoI 37, No 1,2 (1995) стор.15, таблиця 6 (1995) Plant Cell Rep. 14: 273-278; Jahne і ін. (1994) гриби]. Особливо бажані філаментний Theor Appl Genet 89: 525-533]. Hemiascomycet Ashbya gossypii або Eremothecium Спосіб за винаходом може бути комбінований ashbyii. з іншими способами, що впливають на стійкість до b) дріжджі, такі, як Candida, Saccharomyces, патогену (наприклад, стійкість до комах, грибів, Hansenula або Рісhіа, особливо бажані 55 84406 56 Saccharomyces cerevisiae або Pichia pastoris нокислоти, цукор, жирні кислоти, природні та син(ATCC Accession No. 201178), тетичні смакові речовини, ароматичні речовини та c) рослини відповідно до вищенаведеного вибарвники. Особливо бажаним є виробництво тозначення „рослина" коферолів і токотриєнолів, а також каротиноїдів. d) хребетні та безхребетні тварини. Особливо Вирощування трансформованих організмів, а табажаними хребетними є такі ссавці, як собака, кож виділення з хазяйських організмів, відповідно, кішка, вівця, коза, курка, миша, пацюк, корова або із середовища вирощування, здійснюють відомими кінь. Бажані тваринні клітини включають клітини фахівцеві в даній області способами. Виробництво CHO, COS, HEK293. Бажані клітини безхребетних фармацевтичних продуктів, таких, як антитіла або включають клітини комах, такі, як клітини вакцини, описане, наприклад, у [публікації Hood Drosophila S2 і Spodoptera Sf9 або Sf21, EE, Jilka JM (1999) Curr Оріп Biotechnol 10(4): 382e) прокаріоти, такі, як грам-позитивні або грам6; Ma JK, Vine ND (1999) Curr Top Microbiol негативні бактерії, такі, як Acetobacter, Immunol 236: 275-92]. Gluconobacter, Corynebacterium, Brevibacterium, Послідовності: Bacillus, Clostridium, Cyanobacter, Escherichia (на1. Послідов.: 1: послідовність нуклеїнової киссамперед, Escherichia coli), Serratia, лоти, що кодує NADPH-оксидазу з ячменю Staphylococcus, Aerobacter, Alcaligenes, Penicillium (Hordeum vulgare). або Klebsiella. 2. Послідов.: 2: послідовність амінокислот, що Як трансгенні організми бажаними є хазяйські кодує NADPH-оксидазу з ячменю (Hordeum організми та вихідні організми, насамперед, росvulgare). лини відповідно до вищенаведеного визначення. У 3. Послідов.: 3: послідовність нуклеїнової кисрамках винаходу вони включають усі роди та види лоти, що кодує NADPH-оксидазу з рису (Oryza вищих і нижчих рослин рослинного світу. Далі sativa). включені зрілі рослини, посівний матеріал, парост4. Послідов.: 4: послідовність амінокислот, що ки і зародки, а також одержані від них частини, кодує NADPH-оксидазу з рису (Oryza sativa). матеріал для розмноження та культури, напри5. Послідов.: 5: послідовність нуклеїнової кисклад, клітинні культури. Під зрілими рослинами лоти, що кодує NADPH-оксидазу з Nicotiana маються на увазі рослини в будь-якій стадії після tabacum стадії проростків. Під проростками розуміють мо6. Послідов.: 6: послідовність амінокислот, що лоду, незрілу рослину в ранній стадії розвитку. кодує NADPH-оксидазу з Nicotiana tabacum Зокрема, бажаними як хазяйські організми є рос7. Послідов.: 7: послідовність нуклеїнової кислини, на яких можна застосовувати спосіб за вилоти, що кодує NADPH-оксидазу з картоплі находом для одержання стійкості до патогену від(Solanum tuberosum) повідно до вищенаведених критеріїв. Особливо 8. Послідов.: 8: послідовність амінокислот, що бажаними однодольними рослинами є такі, як кодує NADPH-оксидазу з картоплі (Solanum пшениця, овес, просо, ячмінь, жито, кукурудза, tuberosum) рис, гречка, сорго, трітікале, полба, льон, цукрова 9. Послідов.: 9: послідовність нуклеїнової кистростина, дводольні культурні рослини, такі, як лоти, що кодує NADPH-оксидазу з томату рапс, канола, блощичник, Arabidopsis, види капус(Lycopersicon esculentum) ти, соя, люцерна, горох, бобові, земляний горіх, 10. Послідов.: 10: послідовність амінокислот, картопля, тютюн, томати, баклажан, перець, сощо кодує NADPH-оксидазу з томату (Lycopersicon няшник, тагетес, салат, календула, диня, гарбуз esculentum) або кабачки. 11. Послідов: 11: послідовність нуклеїнової киОдержання трансгенних організмів може бути слоти, що кодує NADPH-оксидазу з Arabidopsis реалізоване вищеописаними способами трансфоthaliana (RbohF) рмації або трансфекції організмів. 12. Послідов.: 12: послідовність амінокислот, Ще одним об'єктом винаходу є застосування що кодує NADPH-оксидазу з Arabidopsis thaliana трансгенних організмів за винаходом і клітин, що (RbohF) одержані від них, клітинних культур, частин, на13. Послідов.: 13: послідовність нуклеїнової приклад, у випадку трансгенних рослинних органікислоти, що кодує NADPH-оксидазу з Arabidopsis змів корені, листя і т.п., і трансгенного матеріалу thaliana (RbohD) розмноження, такого, як посівний матеріал або 14. Послідов.: 14: послідовність амінокислот, плоди, для одержання засобів харчування, кормів, що кодує NADPH-оксидазу з Arabidopsis thaliana фармацевтичних продуктів або тонких хімікатів. (RbohD) Далі бажаний спосіб рекомбінантного одер15. Послідов: 15: послідовність нуклеїнової кижання фармацевтичних продуктів або тонких хіміслоти, що кодує NADPH-оксидазу з Nicotiana катів в хазяйських організмах, відповідно до якого tabacum (rboh) організм-хазяїн трансформують однієї або декіль16. Послідов.: 16: послідовність амінокислот, кома вищеописаними експресійними касетами, а що кодує NADPH-оксидазу з Nicotiana tabacum експресійна касета містить один або декілька (rboh) структурних генів, що кодують бажані тонкі хімікати 17. Послідов.: 17: послідовність нуклеїнової або каталізують біосинтез бажаних тонких хімікакислоти, що кодує NADPH-оксидазу з рису (Oryza тів, трансформований організм-хазяїн вирощують sativa var. japonica) та бажаний тонкий хімікат виділяють із середови18. Послідов.: 18: послідовність амінокислот, ща вирощування. Цей спосіб застосовується для що кодує NADPH-оксидазу з рису (Oryza sativa var. таких тонких хімікатів, як ферменти, вітаміни, аміjaponica) 57 84406 58 19 Послідов.: 19: послідовність нуклеїнової киФіг. «РНК інтерференція» з pNAox дволанцюслоти, що кодує NADPH-оксидазу з Arabidopsis говою PHK знижує ефективність проникнення thaliana (RbohC) справжньої борошнистої роси Bgh6 у ячменю. 20. Послідов.: 20: послідовність амінокислот, Відносна ефективність проникнення (RPE) вищо кодує NADPH-оксидазу з Arabidopsis thaliana значається в шести індивідуальних експериментах (RbohC) при інокуляції за допомогою Bgh з ячменю cv 21. Послідов.: 21: послідовність нуклеїнової Pallas. Відносна ефективність проникнення RPE кислоти, що кодує NADPH- і оксидазу з Arabidopsis розраховується як різниця ефективності проникthaliana (RbohA) I нення PE для трансформованих рNAох-длРНК 22. Послідов.: 22: послідовність амінокислот, клітин та ефективності проникнення PE для контщо кодує NADPH-оксидазу з Arabidopsis thaliana рольних, трансформованих дволанцюговою PHK (RbohA) клітин (тут: середня ефективність проникнення 23. Послідов.: 23: олігонуклеотидний праймер складає 57%). Процентне значення RPE (%-RPE) 5' NAOX 5'-GARCAAGGCTCTTTTGATTG-3' розраховують з RPE шляхом віднімання одиниці 24. Послідов.: 24: олігонуклеотидний праймер та множенням на 100. 3' Naox 5'-GAAATGCTCCTTATGGAATTC-3' RPE [РЕ для трансформо ваних pNAо - длРНК клітин ] [РЕ для трансформо ваних контрольн ою длРНК клітин ] %-RPE=100*(RPE-1) Колонки «1»-«5» представляють собою %-RPE (тобто, відхилення ефективності проникнення від середнього значення ефективності проникнення контролю) при оцінці, принаймні, 100 місць взаємодії для кожного незалежного експерименту. Клонка «m» представляє собою середню відносну ефективність проникнення %-RPE експериментів. Колонка похибок показує стандартну похибку. "Контроль-длРНК" показує паралельні експерименти з контрольною дволанцюговою PHK. «pNAox-длРНК» представляють собою експерименти з дволанцюговою PHK NADPH-оксидази ячменю. %-RPE було значно нижче в клітинах, що бомбардувалися pNAox-длРНК (характеристика р=0,0054), у порівнянні з клітинами, що бомбардувалися контрольною длРНК (TR означає людську дволанцюгову PHK рецептора щитовидної залози). Приклади Загальні способи: Хімічний синтез олігонуклеотидів може здійснюватися, наприклад, відомим способом, за фосфоамідитному способом [див. публікацію Voet, Voet, 2. Auflage, Wiley Press New York, стор.896897]. Проведені в рамках даного винаходу стадії клонування, такі, як, наприклад, рестрикційне розщеплення, гель-електрофорез на агарозі, очищення ДНК-фрагментів, перенос нуклеїнової кислоти на нітроцелюлозну та нейлонову мембрани, зв'язування ДНК-фрагментів, трансформація клітин E. coli, вирощування бактерій, розмноження фагів та аналіз послідовності рекомбінантної ДНК проводять так, як описано в [публікації Sambrook і ін. (1989) Cold Spring Harbor Laboratory Press; ISBN 087969-309-6]. Секвенування рекомбінантних ДНКмолекул здійснюють на лазерному ДНКсеквенаторі з детектором флуоресценції фірми MWG-Licor згідно зі способом Зангера [див. Sangeri ін. (1977) Proc Natl Acad Sci USA 74: 54635467]. Приклад 1 Рослини, патогени та інокуляція Сорт ячменю Pallas був-наданий Lisa Munk, Department of Plant Pathology, Royal Veterinary and Agricultural University, Копенгаген, Данія. Його одержання описане в літературі див. [Klster P і ін. (1986) Crop Sci 26: 903-907]. 5 зерен пророщенного протягом 12-36 годин у темряві на фільтрувальному папері посівного матеріалу, який викладають на край чотиригранного горщика (8х8см) із землею "Fruhstorfer Erde" типу P, покривають землею та регулярно поливають водопровідною водою. Усі рослини культивують у вегетаційних кліматичних камерах або кліматронах при температурі від 16 до 18°С, з відносною вологістю повітря від 50 до 60% і з циклами 16 годин світла/8 годин темряви при 3000 відповідно, 5000лк (щільність потоку фотонів 50, відповідно, 60мкмолей-1м-2) протягом 5-8 днів і застосовують у тестах на стадії проростків. При тестах, в яких аплікацію проводять на первинному листі, застосовують цілком розвинуті рослини. Перед проведенням трансгенних експериментів по трансфекції рослини культивують у вегетаційних кліматичних камерах або кліматронах при денній температурі 24°С, нічній температурі 20°C з відносною вологістю повітря від 50 до 60% з 16годинним світлим/8-годинним темним періодами при 30000лк. Для інокуляції рослин ячменю застосовують справжню борошнисту росу ВІumеrіа graminis (DC) Speer f.sp. hordei Em. Marchal der Rasse А6 (Wiberg A (1974) Hereditas 77: 89-148) (Bgh6). Вона була надана Інститутом по біометрії в місті Гісен, Німеччина. Вирощування інокуляту проводять у вегетаційних кліматичних камерах за тих самих умов, що описані вище для рослин, за допомогою передачі конідій з ураженого рослинного матеріалу на 7-денні рослини ячменю cv. Golden Promise при щільності зараження 100конідій/мм2. Інокуляцію за допомогою Bgh6 проводять із застосуванням 7-денних проростків стряхуванням конідій з вже уражених рослин в інокуляційних баштах з прибл. 100конідій/мм2 (якщо не зазначено інакше). Приклад 2 Клонування pNAox кДНКпослідовності з ячменю Необхідна для виділення HvpNAox кДНК, її клонування, секвенування та одержання зондів 59 84406 60 кДНК-фрагментів одержують за допомогою поліосаджують центрифугуванням протягом 30 хвилин меразної ланцюгової реакції зі зворотною транскпри 20000g і при - 4°С. Осад завантажують у риптазою (RT-PCR) при застосуванні "одностадій10мкл ТЕ буфера. Відповідну реакційну суміш заного набору RT-PCR" (фірма Life Technologies, стосовують безпосередньо для транскрипції in Карлсруе, Німеччина, або фірма Qiagen, Гільден, vitro за допомогою Е7-РНК-полімерази, відповідно, Німеччина). Для цього застосовують загальну PHK SP6- PHK-полімерази. РНК-полімерази одержують із сіянців ячменю як матрицю. PHK виділяють з на фірмі Roche Molecular Biology, Маннгейм, Німеячменю сорту Pallas через 3, 5 і 7 днів після проччина. ростання. Крім того, PHK із Pallas та зворотносхКожна транскрипційна композиція містить в рещених ліній з mlo5, MIg або МІа12 виділяють об'ємі 40мкл: через 1, 2 і 5 днів після інокуляції за допомогою 2мкл лінеаризованої плазмідної ДНК (1мкг) Bgh6 на 7 день після проростання. Для RT-PCR 2мкл NTP (25ммоль) (1,25мМ кожного NTP) застосовують наступні праймери: 4мкл 10 х реакційного буфера (фірми Roche 5' NAOX: 5'-GARCAAGGCTCTTTTGATTG-3' Molecular Biology), (послідов. 23) 1мкл інгібітора РНКази (27 одиниць; фірми 3' Naox: 5'-GAAATGCTCCTTATGGAATTC-3' Roche Molecular Biology), (послідов. 24) 2мкл РНК-полімерази (40од.), Для реакції (25мкл суміші) застосовують по 29 мкл DEPC-води 1000нг від загальної PHK, 0,4ммоль дезоксинуклеПісля інкубації протягом 2 годин при темпераозидтрифосфату, по 0,6ммолів праймерів OPN-1 турі 37°С по одній частині реакційної суміші продута OPN-2, 10мкл інгібітора РНКази і 1мкл суміші ктів транскрипції "смислового"-, відповідно, "антиферментів у 1 х RT-буфері (набір для одноетапної смислового"-ланцюга змішують, протягом 5 RT-PCR, Qiagen, Hilden). хвилин, денатурують при 95°C і після цього під час Застосовують наступну температурну програохолодження протягом 30 хвилин до заключної му (PTC-100TM Modell 96V; MJ Research, Inc., температури 37° гібридизують один з одним ("відWatertown, Massachussetts): палювання"). Альтернативно, можна після денату1 цикл протягом 30хв. при 50°C рації реакційну суміш, що складається із смисло1 цикл протягом 150сек при 94°C вого та антисмислового ланцюга охолоджувати 30 циклів протягом 45сек при 94°С, протягом протягом 30 хвилин при -20°C. Осад білка, що 1хв при 55°C і протягом 2хв. при 72°С утворюється під час денатурації і гібридизації, не1 цикл протягом 7хв. при 72°C тривалим центрифугуванням при 20.800g роздіПродукт полімеразної ланцюгової реакції розляють та надосадову рідину застосовують безподіляють за допомогою 2% ваг./об. гельсередньо для покриття вольфрамових частинок, електрофорезу на агарозі. Одержують продукт RT(див. нижче). Для аналізу 1мкл розчину кожного PCR 378п.о.(послідовність 1), що кодує частину ланцюга PHK і дволанцюгової PHK розділяють на відкритої рамки зчитування NADPH-оксидази. Віднеденатурованому агарозному гелі. Успішна гібповідну кДНК виділяють з агарозного гелю та клоридизація виявляється зсувом смуг у бік більш нують у pGEM-T-вектор (фірма Promega, Маннвисокої молекулярної ваги в порівнянні з окремими хейм, Німеччина) за допомогою Т-понадлігування. ланцюгами. кДНК секвенують, виходячи з плазмідної ДНК при 4мкл дволанцюгової PHK осаджують етанолом застосуванні набору для секвенування "Thermo (доданням 6мкл води, 1мкл 3М розчину ацетату Sequenase Fluorescent Labeled Primer Cycle натрію і 25мкл етанолу, а також центрифугуванSequencing Kit" (фірми Amersham, Фрейбург, Німеням, принаймні, протягом 5 хвилин при 20000g та ччина). Конструкт позначають як pGEM-T-pNAox. температурі 4°C) та ресуспендують у 500мкл води. Приклад 3 Синтез in vitro pNAox дволанцюгоВимірюють спектр абсорбції між 230 і 300нм, відвої РНК повідно, абсорбцію при 280 і 260нм, щоб визначиПлазміда pGEM-T-pNAox, яку застосовують ти чистоту та концентрацію дволанцюгової PHK. для транскрипції in vitro, містить промотор Т7 і SP6 Як правило, одержують від 80 до 100мкг дволанна кожному кінці вбудованої послідовності нуклеїцюгової PHK зі співвідношенням оптичної густини нової кислоти, що дозволяє проводити синтез OD260/OD280 від 1,80 до 1,95. У разі потреби можна смислової, відповідно, антисмислової PHK. Плазпроводити перетравлювання ДНКазою I, що, проміда може бути лінеаризована за допомогою прите, не справляє значного впливу на наступні редатних рестрикційних ферментів (АраІ для полімезультати. рази SP6 і Pst для полімерази Т7), щоб Як контрольна дволанцюгова PHK служить забезпечити коректну транскрипцію вбудованої дволанцюгова PHK людського тироїдного рецеппослідовності нуклеїнової кислоти і перешкодити тора (вихідний вектор p7betaSal [див. публікацію зчитуванню у векторних послідовностях. Для цього Norman C і ін. (1988) Cell 55(6): 989-1003], що за10мкг плазмідної pGEM-T-pNAox ДНК вирізають за безпечується др. Баніахмадом, Інститут генетики в допомогою АраІ для полімерази SP6 і Pst для пом. Гісен, Німеччина, послідовність інсерта описана лімерази 11. Вирізані плазміди екстрагують у під GenBank Acc.-No.:NM_000461). Для одержання 200мкл води з однаковими об'ємами фесмислової PHK перетравлюють плазміду за допонол/хлороформ/ізоаміловий спирт, переносять у могою Pvull, для одержання антисмислової PHK нову реакційну ємкість Еппендорфа (в якому неперетравлюють за допомогою Hindlll і PHK транскмає РНКази) і центрифугують протягом 5 хвилин. рибують потім за допомогою РНК-полімерази 17, 180мкл плазмідного розчину перемішують з відповідно SP6. Окремі стадії способу одержання 420мкл етанолу, ставлять на лід та після цього
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for providing of resistibility to fungal or fungaceous pathogen in monocytoledon
Автори англійськоюKogel Karl-Hainz, Hukkelhoven Ralf, Truillo Marco
Назва патенту російськоюСпособ обеспечения резистентности к грибковому или грибоподобному патогену в однодольных растениях
Автори російськоюКогель Карл-Хайнц, Хюккельховен Ральф, Труилло Марко
МПК / Мітки
МПК: C12N 15/63, C12N 15/82, A01H 5/00
Мітки: забезпечення, грибкового, патогену, резистентності, грибкоподібного, рослинах, спосіб, однодольних
Код посилання
<a href="https://ua.patents.su/97-84406-sposib-zabezpechennya-rezistentnosti-do-gribkovogo-abo-gribkopodibnogo-patogenu-u-odnodolnikh-roslinakh.html" target="_blank" rel="follow" title="База патентів України">Спосіб забезпечення резистентності до грибкового або грибкоподібного патогену у однодольних рослинах</a>
Спосіб визначення зараженості картоплі збудником раку на ранніх етапах розвитку патогену
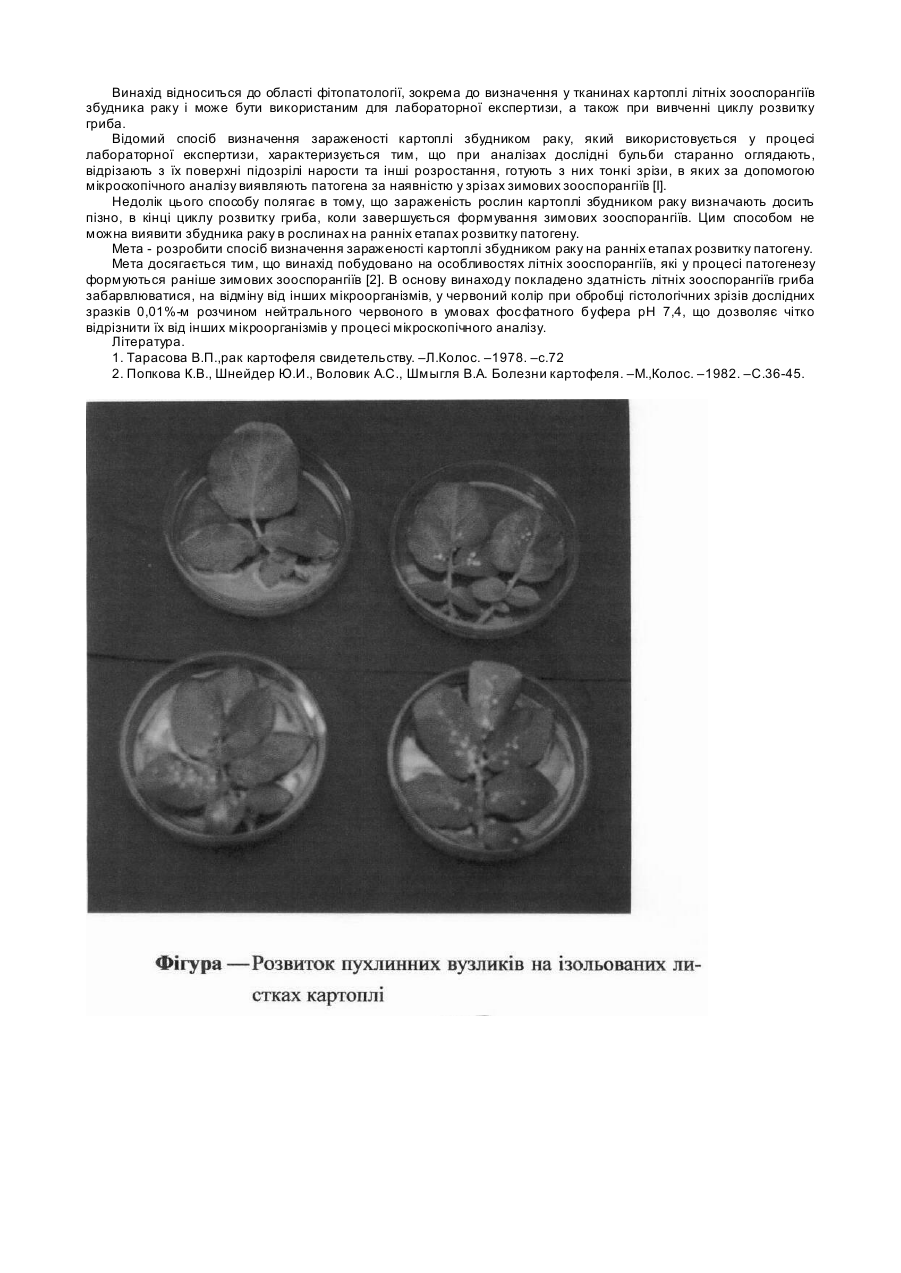
Номер патенту: 64564
Опубліковано: 16.02.2004
Автори: Голик Іван Васильович, Мельник Павло Олексійович, Зеля Аурелія Георгіївна, Мовчан Олексій Миколайович, Хома Інга Павлівна, Мілашевська Жанна Іванівна
МПК: A01N 59/00, A01P 3/00
Мітки: ранніх, етапах, визначення, розвитку, раку, збудником, зараженості, спосіб, патогену, картоплі
Формула / Реферат:
Спосіб визначення зараженості картоплі збудником раку на ранніх етапах розвитку патогену, який включає використання особливостей структурних елементів різних стадій розвитку патогену, який відрізняється тим, що визначення гриба здійснюють за особливостями літніх зооспорангіїв, які розвиваються раніше зимових і, на відміну від інших мікроорганізмів, здатні забарвлюватися в червоний колір в гістологічних зрізах при обробці останніх 0,01%-м...
Спосіб підвищення токсичної дії гербіциду “центуріон” для захисту посівів буряків від однодольних бур’янів

Номер патенту: 60675
Опубліковано: 16.05.2005
Автори: Гізбуллін Наіль Гайфулович, Санчес Антоніо, Гонтаренко Світлана Миколаївна
МПК: A01N 43/00, A01B 79/02
Мітки: токсичної, спосіб, захисту, бур'янів, центуріон, посівів, дії, гербіциду, підвищення, однодольних, буряків
Формула / Реферат:
1. Спосіб визначення виникнення механічних напруг допустимої величини і напряму їх дії в листовому ізотропному матеріалі конструкцій, по якому виготовляють для досліджень на розтягування із листового матеріалу суцільний зразок, отримують діаграму його деформування, визначають по діаграмі допустиму величину механічних напруг, який відрізняється тим, що із листового матеріалу виготовляють партію зразків, подібних суцільному зразку, в центрі...
Мікробіцидна композиція і спосіб контролю та інгібування розвитку грибків на рослинах

Номер патенту: 49841
Опубліковано: 15.10.2002
Автори: Кнауф-Бейтер Гертруд, Кюнг Рут Беатрис
МПК: A01N 43/653
Мітки: грибків, інгібування, мікробіцидна, рослинах, контролю, розвитку, спосіб, композиція
Формула / Реферат:
1. Фітомікробіцидна композиція, що включає принаймні два активні компоненти, яка відрізняється тим, що компонент І є сполукою, яку вибирають з групи, до складу якої входять(Іа) 1-[3-(4-трет-бутилфеніл)-2-метилпропіл]піперидин ("Фенпропідин"),(Іb) цис-4[3-(4-трет-бутилфеніл)-2-метилпропіл]-2,6-диметилморфолін ("Фенпропіморф"), або одна з їх солей чи комплексів з металами кожної зі сполук, а компонент II є...
Спосіб спектрофотометричного визначення гідроксикоричної кислоти та її похідних у лікарських рослинах та їх препаратах

Номер патенту: 54887
Опубліковано: 17.03.2003
Автори: Ліпковська Наталія Олександрівна, Крушинська Олена Анатоліївна, Запорожець Ольга Антонівна, Барвінченко Валентина Миколаївна
МПК: G01N 21/63, G01N 33/483
Мітки: похідних, кислоти, визначення, рослинах, гідроксикоричної, лікарських, спосіб, препаратах, спектрофотометричного
Формула / Реферат:
Спосіб спектрофотометричного визначення гідроксикоричної кислоти та її похідних у лікарських рослинах та їх препаратах шляхом вимірювання поглинання розчину, який відрізняється тим, що пробу попередньо обробляють 0,045 - 0,225 М розчином солі алюмінію (III) при рН 4,5 - 5,5 і вимірюють поглинання при 355 нм проти розчину порівняння, який містить усі ті ж компоненти, крім солі алюмінію (III).
Спосіб визначення фосфатів в рослинах

Номер патенту: 11803
Опубліковано: 25.12.1996
Автори: Пучіхін Кім Олександрович, Носенко Петро Володимирович, Гусаковська Ніна Семенівна
МПК: G01N 33/50
Мітки: фосфатів, рослинах, визначення, спосіб
Формула / Реферат:
Способ определения фосфатов в растениях, включающий срез части растения, выделение из псе сока, обработку части растения и сока раствором молибденовокислого аммония в кислой среде с последующей обработкой дифференцирующими окраску веществами и оценкой результатов по цветной шкале, отличающийся тем, что, с целью повышения чувствительности способа, в качестве дифференцирующих окраску веществ используют 0,2-0,5%-ный водный раствор двухлористого...
Попередній патент: Спосіб лікування потягу до алкогольних напоїв та/або тютюнових продуктів та набір
Наступний патент: Ацетил 2-гідрокси-1,3 діаміноалкани
Випадковий патент: Спосіб моніторингу асинхронних двигунів і пристрій для його здійснення