Виділене моноклональне антитіло людини, яке зв’язується з сd20, та його застосування
Номер патенту: 93478
Опубліковано: 25.02.2011
Автори: Баадсгаард Оле Д. М. С., Паррен Пауль, Руулс Зігрід, Тілінг Джесіка, Хуанг Хайчун, Ван де Вінкель Ян Г. Й., Гленні Мартін, Петерсен Йорген



































































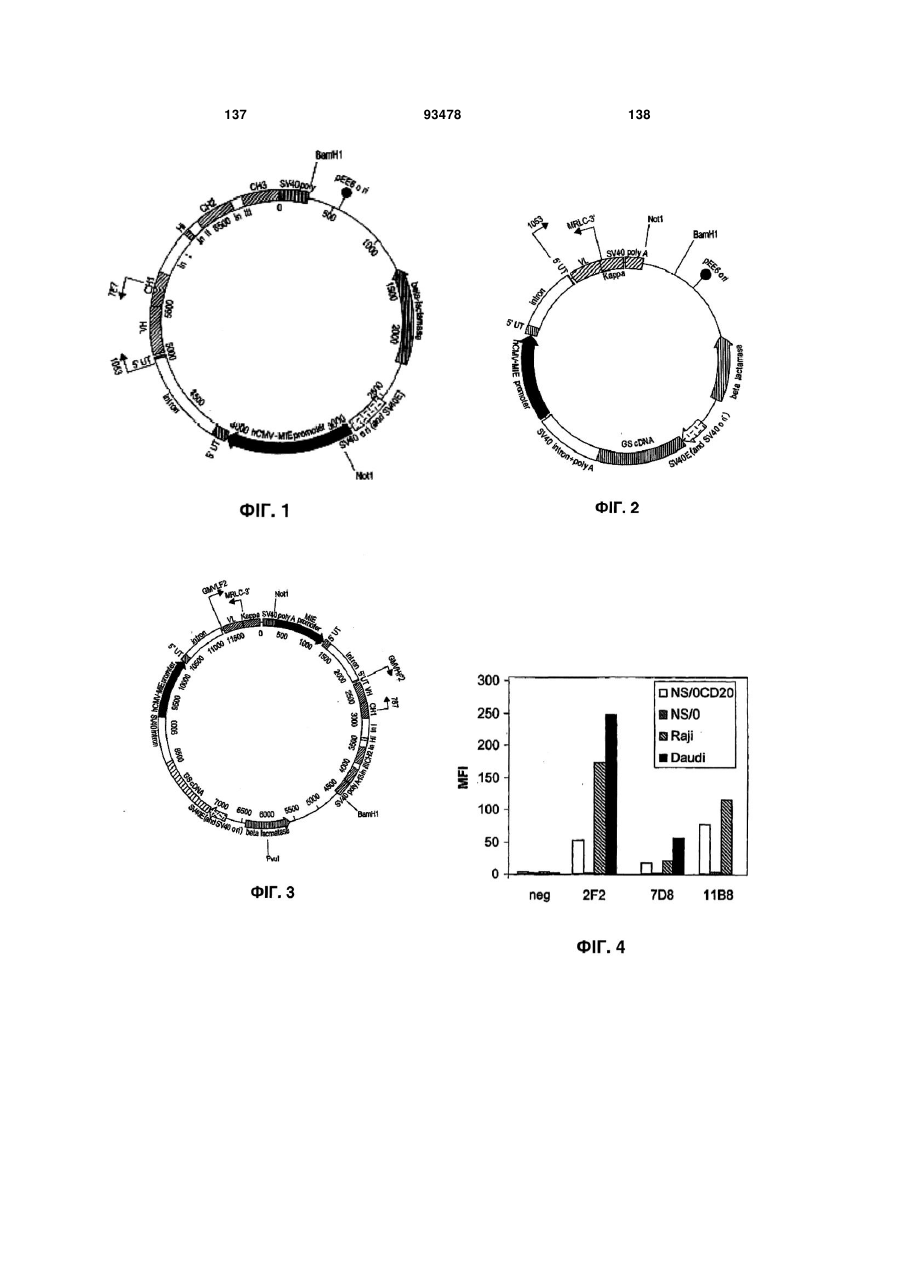
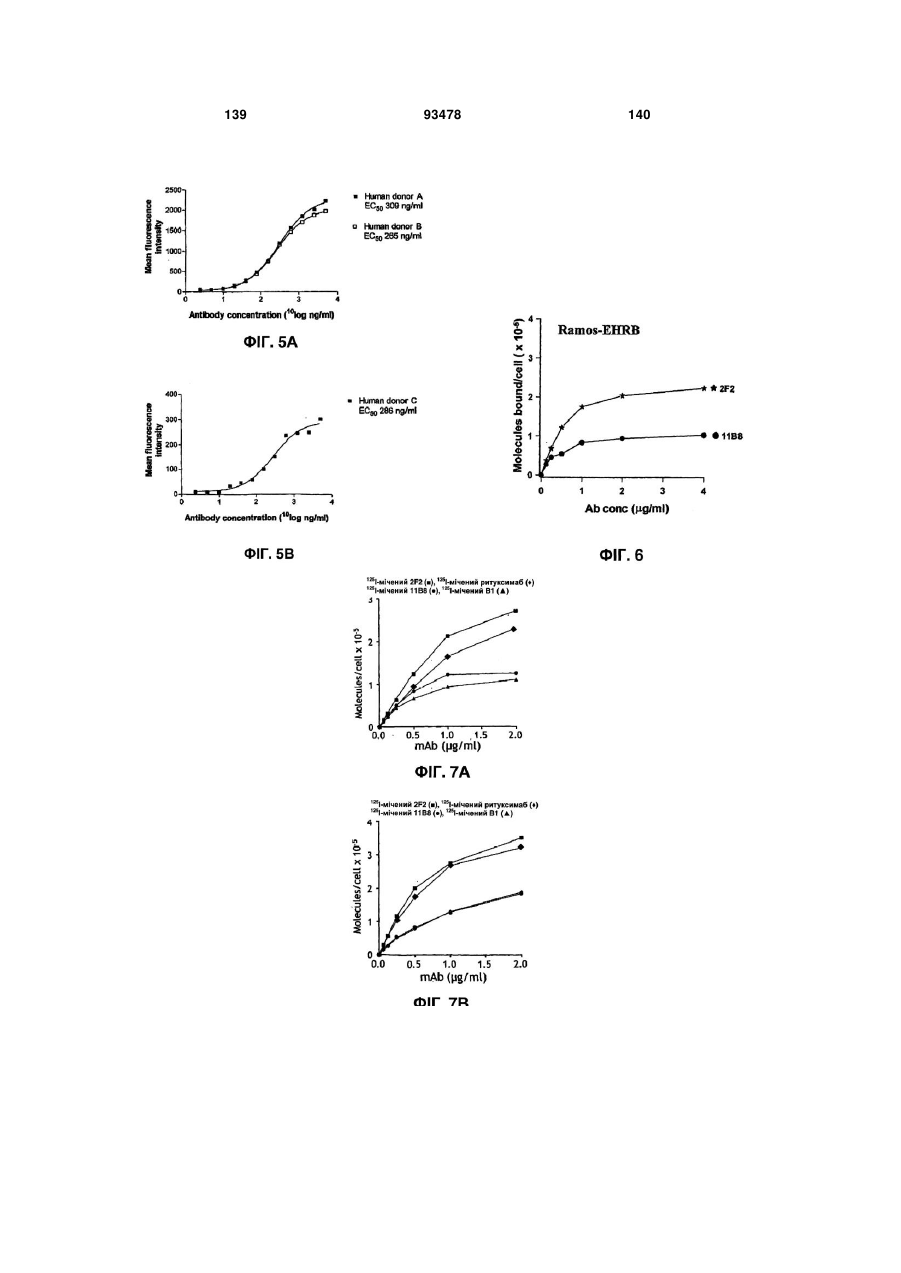
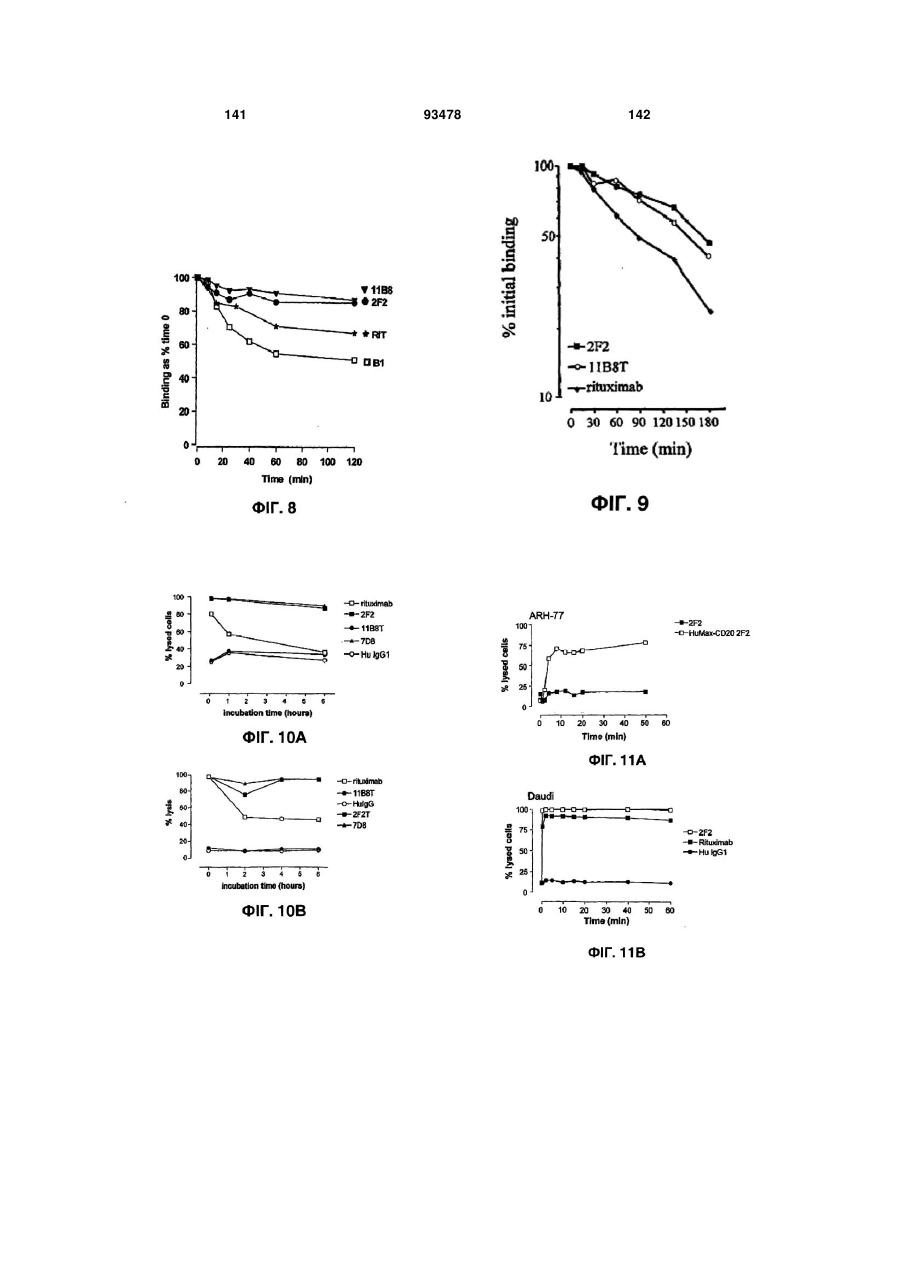
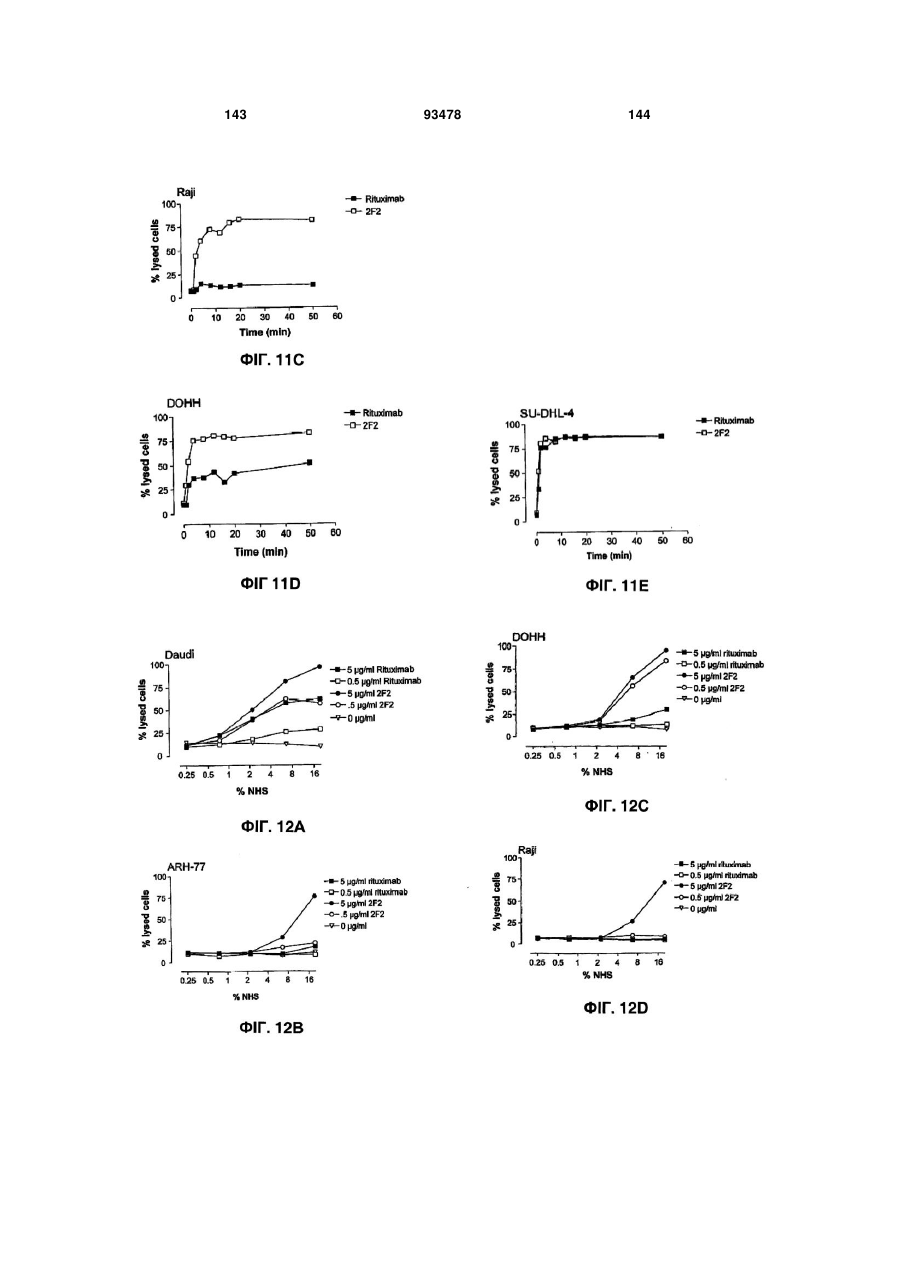
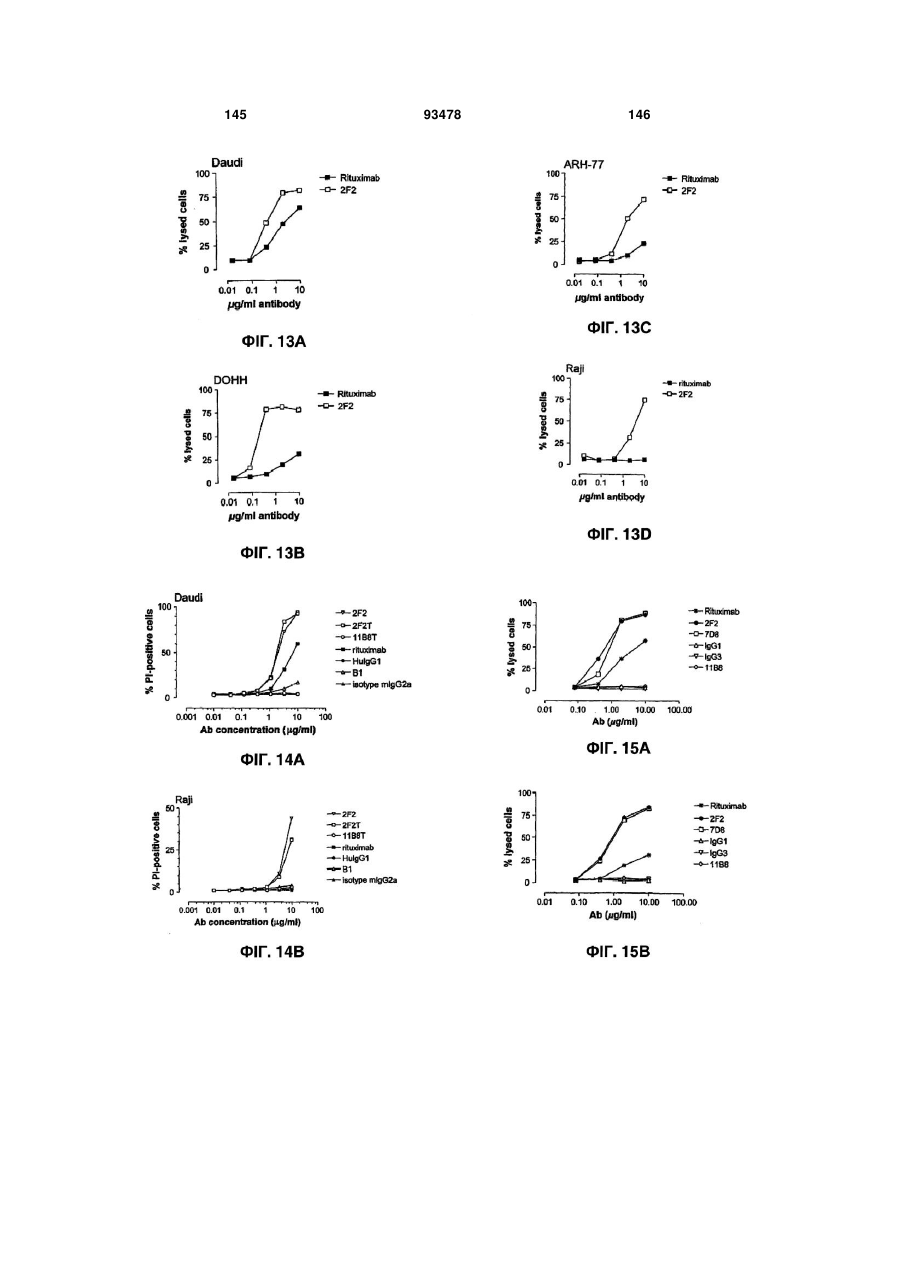
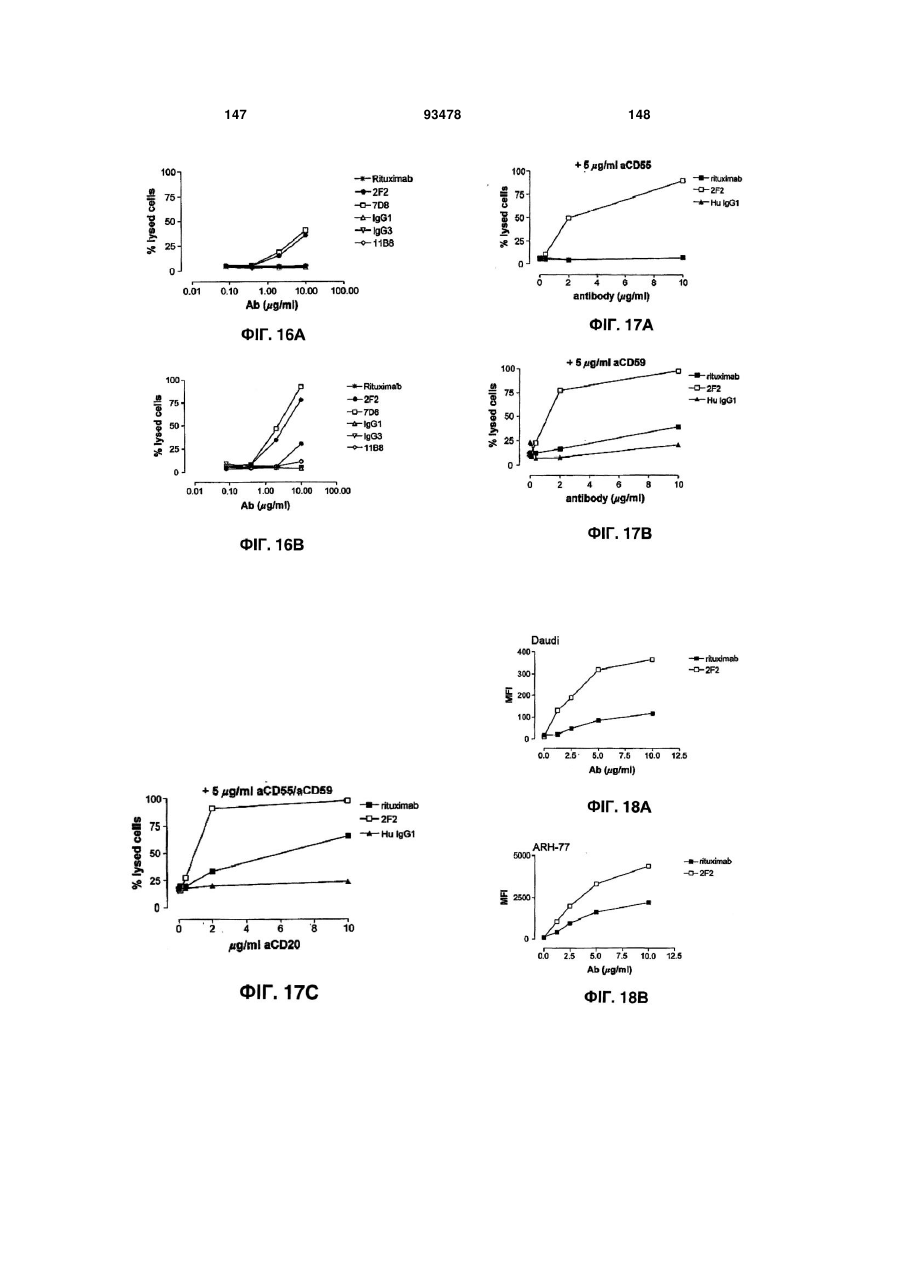
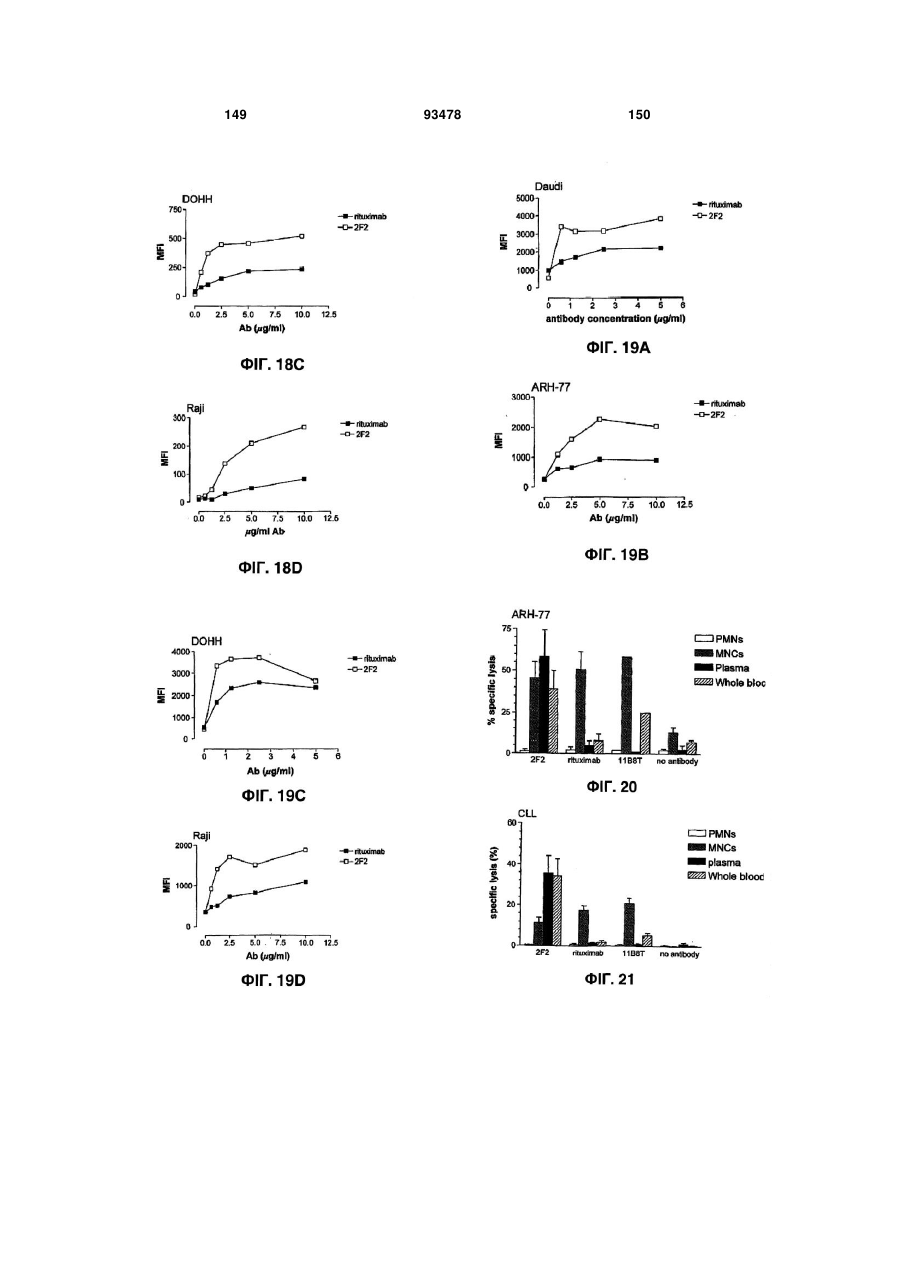















Формула / Реферат
1. Виділене моноклональне антитіло людини, яке зв’язується з CD20 людини і з епітопом на CD20, що не включає або не потребує амінокислотного залишка проліну у положенні 172, але що включає або потребує амінокислотні залишки аспарагіну у положенні 163 та у положенні 166 і яке включає:
(і) CDR1-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:13,
CDR2-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:14,
CDR3-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:15,
CDR1-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:16,
CDR2-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:17 і
CDR3-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:18, або
(іі) CDR1-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:19,
CDR2-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:20,
CDR3-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:21,
CDR1-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:22,
CDR2-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:23 і
CDR3-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:24.
2. Антитіло за п. 1, яке має щонайменше одну з наступних характеристик, вибраних з групи, що складається з:
(i) вибирається з групи, що складається з IgG1-антитіла, IgG2-антитіла, IgG3-антитіла, IgG4-антитіла, IgM-антитіла, IgA1-антитіла, IgA2-антитіла, секреторного IgA-антитіла, IgD-антитіла і IgE-антитіла;
(ii) дисоціюється з CD20 людини із константою швидкості дисоціації (kd) приблизно 10-5 сек-1 або менше;
(iii) зв’язується з CD20 людини із константою афінності (KD) приблизно 5 нмоль або менше;
(iv) здатне індукувати комплемент-залежну цитотоксичність (CDC) клітин, які експресують CD20, у присутності комплементу;
(v) здатне індукувати комплемент-залежну цитотоксичність (CDC) клітин, які експресують CD20 і високі рівні CD55 і/або CD59, у присутності комплементу;
(vi) здатне індукувати апоптоз клітин, які експресують CD20;
(vii) здатне індукувати залежну від антитіл клітинну цитотоксичність (ADCC) клітин, які експресують CD20, у присутності клітин-ефекторів;
(viii) здатне індукувати гомотипічну адгезію клітин, які експресують CD20;
(ix) здатне транслокуватися у ліпідні маси під час зв’язування з CD20;
(x) здатне пролонгувати виживання суб’єкта, який має пухлинні клітини, що експресують CD20;
(xi) здатне знищувати клітини, які експресують CD20; і
(xii) здатне знищувати клітини, які експресують низькі рівні CD20 (клітини CD20low).
3. Антитіло за п. 1, яке здатне знищувати периферійні B-клітини, які експресують низькі рівні CD20 (В-клітини CD20low), до невиявлюваних рівнів протягом більше ніж 50 днів у мавп cynomolgus при дозуванні 6,25 мг/кг на день протягом 4 днів підряд.
4. Антитіло за п. 1, яке зв’язується з CD20, що кодується:
(i) нуклеїновими кислотами важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:1 і SEQ ID NO:3 відповідно;
(ii) нуклеїновими кислотами важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:5 і SEQ ID NO:7 відповідно.
5. Антитіло за п. 1, яке зв’язується з CD20, що має:
(i) варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 і SEQ ID NO:4 відповідно;
(ii) варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:6 і SEQ ID NO:8 відповідно.
6. Антитіло за будь-яким із попередніх пунктів, яке являє собою інтактне антитіло, вибране з групи, що складається з інтактного IgG1-антитіла, інтактного IgG2-антитіла, інтактного IgG3-антитіла, інтактного IgG4-антитіла, інтактного IgM-антитіла, інтактного IgА1-антитіла, інтактного IgА2-антитіла, інтактного секреторного IgA-антитіла, інтактного IgD-антитіла і інтактного IgE-антитіла, причому зазначене антитіло глікозильоване в еукаріотичній клітині.
7. Антитіло за будь-яким із попередніх пунктів, яке являє собою фрагмент антитіла або одноланцюгове антитіло.
8. Антитіло за будь-яким з попередніх пп. 1-7, яке містить послідовність амінокислот варіабельної області важкого ланцюга імуноглобуліну, дериватизовану з гаметичної послідовності VH3-09/JH6b (послідовність SEQ ID NO:56) людини, і послідовність амінокислот варіабельної області легкого ланцюга імуноглобуліну, дериватизовану з гаметичної послідовності VL-L6/JK5 (послідовність SEQ ID NO:57) людини, причому зазначене антитіло людини зв’язується з CD20 людини.
9. Антитіло за будь-яким з пп. 1-7, яке додатково містить хелатуючий лінкер для прикріплення радіоактивного ізотопу.
10. Трансфектома, яка продукує моноклональне антитіло людини, яке зв’язується з CD20 людини і яке кодується нуклеїновими кислотами важкого ланцюга імуноглобуліну (IgG) людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:1 та 3 або в послідовностях SEQ ID NO:5 та 7 відповідно.
11. Трансфектома, яка продукує моноклональне антитіло людини, яке зв’язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну (IgG) і каппа легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно.
12. Еукаріотична або прокаріотична клітина-хазяїн, яка продукує моноклональне антитіло людини, яке зв’язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну і легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно.
13. Трансгенна тварина, відмінна від людини, або рослина, яка продукує моноклональне антитіло людини, яке зв’язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну і легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно.
14. Спосіб продукування моноклонального антитіла людини, що зв’язується з CD20 людини, за будь-яким з попередніх пп. 1-7, який включає:
імунізацію трансгенної тварини, відмінної від людини, що має геном, який містить трансген важкого ланцюга імуноглобуліну людини і трансген легкого ланцюга імуноглобуліну людини, CD20 людини або клітиною, яка експресує CD20 людини, завдяки чому антитіла продукуються В-клітинами тварини;
виділення В-клітин тварини;
злиття В-клітин з клітинами мієломи з метою утворення іммортальних клітин гібридоми, які секретують моноклональні антитіла людини, що є специфічними до CD20 людини; і
виділення моноклональних антитіл людини, що є специфічними до CD20, з культурального супернатанту гібридоми або з трансфектоми, дериватизованої з такої гібридоми.
15. Спосіб за п. 14, відповідно до якого імунізація здійснюється за допомогою клітин, які були трансфіковані CD20 людини.
16. Композиція, яка містить антитіло людини за будь-яким з пп. 1-7 і фармацевтично прийнятний носій.
17. Композиція за п. 16, яка містить перше антитіло, що має варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 і SEQ ID NO:4 відповідно, і друге антитіло, що має варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:10 і SEQ ID NO:12 відповідно.
18. Композиція за будь-яким з пп. 16, 17, яка додатково містить терапевтичний агент.
19. Імунокон’югат, який містить антитіло за будь-яким з пп. 1-7, зшите з цитотоксичним агентом, радіоактивним ізотопом або лікарським засобом.
20. Біспецифічна молекула, яка містить антитіло за будь-яким з пп. 1-7 і має специфічність зв’язування до клітини-ефектора людини.
21. Біспецифічна молекула за п. 20, яка містить антитіло відповідно до будь-якого з пп. 1-7 і має специфічність зв’язування до Fc-рецептора людини або специфічність зв’язування до T-клітинного рецептора, наприклад CD3.
22. Спосіб інгібування росту клітини, що експресує CD20, або знищення клітини, що експресує CD20, який включає введення клітини в контакт з ефективною кількістю антитіла відповідно до будь-якого з пунктів 1-7, завдяки чому ріст клітини інгібується або має місце знищення клітини.
23. Спосіб за п. 22, відповідно до якого клітина являє собою B-лімфоцит або пухлинну клітину.
24. Спосіб лікування або запобігання захворювання або розладу, при якому клітини експресують CD20, який включає застосування до суб’єкту антитіла людини, відповідно до будь-якого з пп. 1-7, композиції, відповідно до будь-якого з пп. 16-18, імунокон’югата відповідно до п. 19, або біспецифічної молекули відповідно до будь-якого з п. 20-21, експресуючого вектора відповідно до будь-якого з пп. 43, 44 у кількості, яка є ефективною для лікування або запобігання захворювання.
25. Спосіб за п. 24, відповідно до якого захворювання являє собою В-клітинну лімфому.
26. Спосіб за п. 25, відповідно до якого захворювання являє собою В-клітинну лімфому не Ходжкіна.
27. Спосіб за п. 25, відповідно до якого захворювання вибране з групи, що складається з лімфобластичного лейкозу/лімфоми прекурсорних В-клітин і неоплазм зрілих В-клітин, наприклад, хронічного лімфоцитарного лейкозу В-клітин (CLL)/малолімфоцитарної лімфоми (SLL), пролімфоцитарного лейкозу В-клітин, лімфоплазмоцитарної лімфоми, лімфоми мантієподібних клітин (MCL), фолікулярної лімфоми (FL), шкірної лімфоми центрального фолікула, лімфоми В-клітин маргінальної зони (типу MALT (у лімфоїдній тканині слизових оболонок), вузлового і селезінкового типу), лейкозу ворсистих клітин, дифузної лімфоми великих В-клітин, лімфоми Беркіта, плазмоцитоми, мієломи плазмоцидів, пост-трансплантаційного лімфопроліферативного розладу, макроглобулінемії Вальденстрьома і анапластичної лімфоми великих клітин (ALCL).
28. Спосіб за п. 27, відповідно до якого захворювання являє собою фолікулярну лімфому (FL) або хронічний лімфоцитарний лейкоз В-клітин (CLL)/малолімфоцитарну лімфому (SLL).
29. Спосіб за п. 25, відповідно до якого захворювання вибране з групи, що складається з лімфоматоїдного гранулематозу, первинної ефузійної лімфоми, інтраваскулярної лімфоми великих В-клітин, медіастинальної лімфоми великих В-клітин, захворювань важких ланцюгів (включаючи γ-, μ- і α-захворювання), лімфом, індукованих лікуванням імуносупресантами, наприклад, циклоспорин-індукованої лімфоми і метотрексат-індукованої лімфоми.
30. Спосіб лікування або запобігання імунного захворювання, при якому імунокомпетентні клітини експресують CD20, який включає застосування до суб’єкта антитіла, відповідно до будь-якого з пп. 1-7, композиції, відповідно до будь-якого з пп.в 16-18, імунокон’югата відповідно до п. 19, або біспецифічної молекули відповідно до будь-якого з пп. 20-21, експресуючого вектора відповідно до будь-якого з пп. 43-44 у кількості, яка є ефективною для лікування або запобігання імунного захворювання.
31. Спосіб за п. 30, відповідно до якого лікування включає знищення B-клітин, які продукують антитіла проти аутологічних антигенів.
32. Спосіб за п. 30, відповідно до якого захворювання або розлад вибраний з групи, що складається з псоріазу, псоріатичного артриту, дерматиту, системних склеродерми і склерозу, запального захворювання кишечнику (IBD), хвороби Крона, виразкового коліту, синдрому дихальної недостатності, менінгіту, енцефаліту, увеїту, гломерулонефриту, екземи, астми, атеросклерозу, недостатності адгезії лейкоцитів, розсіяного склерозу, синдрому Рейно, синдрому Шегрена, ювенільного діабету, хвороби Рейтера, хвороби Бехчета, імунокомплексного нефриту, нефропатії IgA, полінейропатії IgM, імуноопосердкованої тромбоцитопенії, наприклад гострої ідіопатичної тромбоцитопенічної пурпури і хронічної ідіопатичної тромбоцитопенічної пурпури, гемолітичної анемії, злоякісної міастенії, вовчакового нефриту, системного вовчакового еритематозу, ревматоїдного артриту (RA), атопічного дерматиту, пухирчатки, хвороби Грейвса, тиреоїдиту Хасімото, гранулематозу Вегенера, синдрому Омена, хронічної ниркової недостатності, гострого інфекційного мононуклеозу, ВІЛ і захворювань, пов’язаних з вірусом герпесу.
33. Спосіб за п. 32, відповідно до якого аутоімунне захворювання являє собою ревматоїдний артрит (RA).
34. Спосіб за п. 30 відповідно до якого захворювання являє собою запальний, імунний і/або аутоімунний розлад, вибраний з виразкового коліту, хвороби Крона, ювенільного діабету, розсіяного склерозу, імуноопосердкованої тромбоцитопенії, наприклад гострої ідіопатичної тромбоцитопенічної пурпури і хронічної ідіопатичної тромбоцитопенічної пурпури, гемолітичної анемії, злоякісної міастенії, системного склерозу і пухирчатки звичайної.
35. Спосіб за п. 30, відповідно до якого захворювання являє собою запальний, імунний і/або аутоімунний розлад, вибраний з запального захворювання кишечнику (IBD), виразкового коліту, хвороби Крона і розсіяного склерозу.
36. Спосіб за будь-яким з пп. 22-35, який додатково включає окреме застосування до суб’єкту іншого терапевтичного агента.
37. Спосіб за п. 36, відповідно до якого терапевтичний агент являє собою
(i) цитотоксичний агент або радіотоксичний агент;
(ii) імуносупресант; або
(iii) імунологічний модулюючий агент, наприклад цитокін або хемокін.
38. Спосіб за п. 36, відповідно до якого терапевтичний агент вибраний з групи, що складається з доксорубіцину, цисплатину, блеоміцину, кармустину, хлорамбуцилу і циклофосфаміду.
39. Спосіб за п. 36, відповідно до якого терапевтичний агент вибраний з групи, що складається з антитіл проти CD25, антитіл проти CD19, антитіл проти CD21, антитіл проти CD22, антитіл проти CD37, антитіл проти CD38, антитіл проти IL6R, антитіл проти IL8, антитіл проти IL15, антитіл проти IL15R, антитіл проти CD4, анти-CD11a, антитіл проти альфа-4/бета-1 інтегрин (VLA4), CTLA4-Ig і антитіл проти C3b(i).
40. Спосіб виявлення іn vitro присутності CD20-антигену або клітини, яка експресує CD20, у зразку, який включає:
введення зразка в контакт з антитілом за будь-яким з пп. 1-7 в умовах, які дозволяють утворити комплекс між антитілом і CD20; і
виявлення утворення комплексу.
41. Набір для виявлення присутності CD20-антигену або клітини, яка експресує CD20, у зразку, який містить антитіло за будь-яким з пп. 1-7.
42. Спосіб виявлення іn vivo присутності CD20-антигену або клітини, яка експресує CD20, у суб’єкта, який включає:
введення антитіла за будь-яким з пп. 1-7 суб’єкту в умовах, які дозволяють утворити комплекс між антитілом і CD20; і виявлення утвореного комплексу.
43. Експресуючий вектор, кодуючий моноклональне антитіло людини, яке зв’язується з CD20 людини, який містить послідовність нуклеотидів, яка кодує варіабельну область важкого ланцюга імуноглобуліну, що містить послідовність нуклеотидів, вибрану з групи, що складається з послідовностей нуклеотидів, як це зазначено в послідовностях SEQ ID NO:1 і 3, і варіабельну область легкого ланцюга імуноглобуліну, що містить послідовність нуклеотидів, вибрану з групи, що складається з послідовностей нуклеотидів, як це зазначено у послідовностях SEQ ID NO:5 і 7.
44. Експресуючий вектор, кодуючий моноклональне антитіло людини, яке зв’язується з CD20 людини, який містить послідовність нуклеотидів, яка кодує варіабельну область важкого ланцюга імуноглобуліну, що містить послідовність амінокислот, вибрану з групи, що складається з послідовностей амінокислот, як це зазначено в послідовностях SEQ ID NO:2 та 4, і варіабельну область легкого ланцюга імуноглобуліну, що містить послідовність амінокислот, вибрану з групи, що складається з послідовностей амінокислот, як це зазначено у послідовностях SEQ ID NO:6 та 8.
45. Фармацевтична композиція, яка містить експресуючий вектор за будь-яким з пп. 43-44 і фармацевтично прийнятний носій.
46. Антиідіотипічне антитіло, яке зв’язується з антитілом за будь-яким з пп. 1-7.
47. Антиідіотипічне антитіло у відповідності до п. 46, яке зв’язується з 2F2 або 7D8.
48. Використання антиідіотипічного антитіла за п. 46 або 47 для виявлення рівня вмісту моноклонального антитіла людини проти CD20 у зразку.
Текст
1. Виділене моноклональне антитіло людини, яке зв‘язується з CD20 людини і з епітопом на CD20, що не включає або не потребує амінокислотного залишка проліну у положенні 172, але що включає або потребує амінокислотні залишки аспарагіну у положенні 163 та у положенні 166 і яке включає: (і) CDR1-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:13, CDR2-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:14, CDR3-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:15, CDR1-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:16, CDR2-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:17 і CDR3-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:18, або (іі) CDR1-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:19, CDR2-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:20, CDR3-ділянку важкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:21, CDR1-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:22, CDR2-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:23 і CDR3-ділянку легкого ланцюга, що має амінокислотну послідовність, зазначену в SEQ ID NO:24. 2. Антитіло за п. 1, яке має щонайменше одну з наступних характеристик, вибраних з групи, що складається з: (i) вибирається з групи, що складається з IgG1-антитіла, IgG2-антитіла, IgG3-антитіла, IgG4-антитіла, IgM-антитіла, IgA1-антитіла, IgA2антитіла, секреторного IgA-антитіла, IgD-антитіла і IgE-антитіла; UA (21) a200504550 (22) 17.10.2003 (24) 25.02.2011 (86) PCT/US2003/033057, 17.10.2003 (31) 60/419,163 (32) 17.10.2002 (33) US (31) 60/460,028 (32) 02.04.2003 (33) US (46) 25.02.2011, Бюл.№ 4, 2011 р. (72) ТІЛІНГ ДЖЕСІКА, NL/NL, РУУЛС ЗІГРІД, NL/NL, ГЛЕННІ МАРТІН, GB/GB, ВАН ДЕ ВІНКЕЛЬ ЯН Г. Й., NL/NL, ПАРРЕН ПАУЛЬ, NL/NL, ПЕТЕРСЕН ЙОРГЕН, DK/DK, БААДСГААРД ОЛЕ Д. М. С., SE, ХУАНГ ХАЙЧУН, US/US (73) ДЖЕНМАБ А/С, DK (56) US A1 20020064823, 30.05.2002. WO A2 0212437, 17.02.2002. US A1 2002009427, 24.01.2002. WO A 0222212, 21.03.2002. WO A 9411026, 26.05.1994. US A 6120767, 19.09.2000. WO A 0009160, 24.02.2000. US A 5770429, 23.06.1998. GAZZANO-SANTORO H ET AL: "A non-radioactive complement-dependent cytotoxicity assay for antiCD20 monoclonal antibody" JOURNAL OF IMMUNOLOGICAL METHODS, ELSEVIER SCIENCE PUBLISHERS B.V.,AMSTERDAM, NL, vol. 202, no. 2, 28 March 1997 (1997-03-28), pages 163171, XP004126276 ISSN: 0022-1759. EDWARDS J C W ET AL: "Sustained improvement in rheumatoid arthritis following a protocol designed to deplete B lymphocytes" RHEUMATOLOGY, OXFORD UNIVERSITY PRESS, LONDON, GB, vol. 40, no. 2, February 2001 (2001-02), pages 205-211, XP002348931 ISSN: 1462-0324. VAN SPRIEL A B ET AL: "Immunotherapeutic perspective for bispecific antibodies" IMMUNOLOGY TODAY, ELSEVIER PUBLICATIONS, CAMBRIDGE, GB, vol. 21, no. 8, 1 August 2000 (2000-08-01), pages 391-397, XP004215167 ISSN: 0167-5699. 2 (19) 1 3 (ii) дисоціюється з CD20 людини із константою швидкості дисоціації (kd) приблизно 10-5 сек-1 або менше; (iii) зв‘язується з CD20 людини із константою афінності (KD) приблизно 5 нмоль або менше; (iv) здатне індукувати комплементзалежну цитотоксичність (CDC) клітин, які експресують CD20, у присутності комплементу; (v) здатне індукувати комплементзалежну цитотоксичність (CDC) клітин, які експресують CD20 і високі рівні CD55 і/або CD59, у присутності комплементу; (vi) здатне індукувати апоптоз клітин, які експресують CD20; (vii) здатне індукувати залежну від антитіл клітинну цитотоксичність (ADCC) клітин, які експресують CD20, у присутності клітин-ефекторів; (viii) здатне індукувати гомотипічну адгезію клітин, які експресують CD20; (ix) здатне транслокуватися у ліпідні маси під час зв‘язування з CD20; (x) здатне пролонгувати виживання суб‘єкта, який має пухлинні клітини, що експресують CD20; (xi) здатне знищувати клітини, які експресують CD20; і (xii) здатне знищувати клітини, які експресують низькі рівні CD20 (клітини CD20low). 3. Антитіло за п. 1, яке здатне знищувати периферійні B-клітини, які експресують низькі рівні CD20 low (В-клітини CD20 ), до невиявлюваних рівнів протягом більше ніж 50 днів у мавп cynomolgus при дозуванні 6,25 мг/кг на день протягом 4 днів підряд. 4. Антитіло за п. 1, яке зв‘язується з CD20, що кодується: (i) нуклеїновими кислотами важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:1 і SEQ ID NO:3 відповідно; (ii) нуклеїновими кислотами важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:5 і SEQ ID NO:7 відповідно. 5. Антитіло за п. 1, яке зв‘язується з CD20, що має: (i) варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 і SEQ ID NO:4 відповідно; (ii) варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:6 і SEQ ID NO:8 відповідно. 6. Антитіло за будь-яким із попередніх пунктів, яке являє собою інтактне антитіло, вибране з групи, що складається з інтактного IgG1-антитіла, інтактного IgG2-антитіла, інтактного IgG3-антитіла, інтактного IgG4-антитіла, інтактного IgM-антитіла, інта 93478 4 ктного IgА1-антитіла, інтактного IgА2-антитіла, інтактного секреторного IgA-антитіла, інтактного IgD-антитіла і інтактного IgE-антитіла, причому зазначене антитіло глікозильоване в еукаріотичній клітині. 7. Антитіло за будь-яким із попередніх пунктів, яке являє собою фрагмент антитіла або одноланцюгове антитіло. 8. Антитіло за будь-яким з попередніх пп. 1-7, яке містить послідовність амінокислот варіабельної області важкого ланцюга імуноглобуліну, дериватизовану з гаметичної послідовності VH3-09/JH6b (послідовність SEQ ID NO:56) людини, і послідовність амінокислот варіабельної області легкого ланцюга імуноглобуліну, дериватизовану з гаметичної послідовності VL-L6/JK5 (послідовність SEQ ID NO:57) людини, причому зазначене антитіло людини зв‘язується з CD20 людини. 9. Антитіло за будь-яким з пп. 1-7, яке додатково містить хелатуючий лінкер для прикріплення радіоактивного ізотопу. 10. Трансфектома, яка продукує моноклональне антитіло людини, яке зв‘язується з CD20 людини і яке кодується нуклеїновими кислотами важкого ланцюга імуноглобуліну (IgG) людини і каппа легкого ланцюга імуноглобуліну людини, які містять у своїх варіабельних областях послідовності нуклеотидів, як зазначено в послідовностях SEQ ID NO:1 та 3 або в послідовностях SEQ ID NO:5 та 7 відповідно. 11. Трансфектома, яка продукує моноклональне антитіло людини, яке зв‘язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну (IgG) і каппа легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно. 12. Еукаріотична або прокаріотична клітина-хазяїн, яка продукує моноклональне антитіло людини, яке зв‘язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну і легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно. 13. Трансгенна тварина, відмінна від людини, або рослина, яка продукує моноклональне антитіло людини, яке зв‘язується з CD20 людини, що має варіабельні області важкого ланцюга імуноглобуліну і легкого ланцюга імуноглобуліну, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 та 4 або в послідовностях SEQ ID NO:6 та 8 відповідно. 14. Спосіб продукування моноклонального антитіла людини, що зв‘язується з CD20 людини, за будь-яким з попередніх пп. 1-7, який включає: імунізацію трансгенної тварини, відмінної від людини, що має геном, який містить трансген важкого ланцюга імуноглобуліну людини і трансген легкого ланцюга імуноглобуліну людини, CD20 людини або клітиною, яка експресує CD20 людини, завдяки чому антитіла продукуються В-клітинами тварини; виділення В-клітин тварини; 5 злиття В-клітин з клітинами мієломи з метою утворення іммортальних клітин гібридоми, які секретують моноклональні антитіла людини, що є специфічними до CD20 людини; і виділення моноклональних антитіл людини, що є специфічними до CD20, з культурального супернатанту гібридоми або з трансфектоми, дериватизованої з такої гібридоми. 15. Спосіб за п. 14, відповідно до якого імунізація здійснюється за допомогою клітин, які були трансфіковані CD20 людини. 16. Композиція, яка містить антитіло людини за будь-яким з пп. 1-7 і фармацевтично прийнятний носій. 17. Композиція за п. 16, яка містить перше антитіло, що має варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:2 і SEQ ID NO:4 відповідно, і друге антитіло, що має варіабельні області важкого ланцюга імуноглобуліну людини і каппа легкого ланцюга імуноглобуліну людини, які містять послідовності амінокислот, як зазначено в послідовностях SEQ ID NO:10 і SEQ ID NO:12 відповідно. 18. Композиція за будь-яким з пп. 16, 17, яка додатково містить терапевтичний агент. 19. Імунокон‘югат, який містить антитіло за будьяким з пп. 1-7, зшите з цитотоксичним агентом, радіоактивним ізотопом або лікарським засобом. 20. Біспецифічна молекула, яка містить антитіло за будь-яким з пп. 1-7 і має специфічність зв‘язування до клітини-ефектора людини. 21. Біспецифічна молекула за п. 20, яка містить антитіло відповідно до будь-якого з пп. 1-7 і має специфічність зв‘язування до Fc-рецептора людини або специфічність зв‘язування до T-клітинного рецептора, наприклад CD3. 22. Спосіб інгібування росту клітини, що експресує CD20, або знищення клітини, що експресує CD20, який включає введення клітини в контакт з ефективною кількістю антитіла відповідно до будь-якого з пунктів 1-7, завдяки чому ріст клітини інгібується або має місце знищення клітини. 23. Спосіб за п. 22, відповідно до якого клітина являє собою B-лімфоцит або пухлинну клітину. 24. Спосіб лікування або запобігання захворювання або розладу, при якому клітини експресують CD20, який включає застосування до суб‘єкту антитіла людини, відповідно до будь-якого з пп. 1-7, композиції, відповідно до будь-якого з пп. 16-18, імунокон‘югата відповідно до п. 19, або біспецифічної молекули відповідно до будь-якого з п. 20-21, експресуючого вектора відповідно до будь-якого з пп. 43, 44 у кількості, яка є ефективною для лікування або запобігання захворювання. 25. Спосіб за п. 24, відповідно до якого захворювання являє собою В-клітинну лімфому. 26. Спосіб за п. 25, відповідно до якого захворювання являє собою В-клітинну лімфому не Ходжкіна. 27. Спосіб за п. 25, відповідно до якого захворювання вибране з групи, що складається з лімфобластичного лейкозу/лімфоми прекурсорних Вклітин і неоплазм зрілих В-клітин, наприклад, хро 93478 6 нічного лімфоцитарного лейкозу В-клітин (CLL)/малолімфоцитарної лімфоми (SLL), пролімфоцитарного лейкозу В-клітин, лімфоплазмоцитарної лімфоми, лімфоми мантієподібних клітин (MCL), фолікулярної лімфоми (FL), шкірної лімфоми центрального фолікула, лімфоми В-клітин маргінальної зони (типу MALT (у лімфоїдній тканині слизових оболонок), вузлового і селезінкового типу), лейкозу ворсистих клітин, дифузної лімфоми великих В-клітин, лімфоми Беркіта, плазмоцитоми, мієломи плазмоцидів, пост-трансплантаційного лімфопроліферативного розладу, макроглобулінемії Вальденстрьома і анапластичної лімфоми великих клітин (ALCL). 28. Спосіб за п. 27, відповідно до якого захворювання являє собою фолікулярну лімфому (FL) або хронічний лімфоцитарний лейкоз В-клітин (CLL)/малолімфоцитарну лімфому (SLL). 29. Спосіб за п. 25, відповідно до якого захворювання вибране з групи, що складається з лімфоматоїдного гранулематозу, первинної ефузійної лімфоми, інтраваскулярної лімфоми великих Вклітин, медіастинальної лімфоми великих В-клітин, захворювань важких ланцюгів (включаючи γ-, μ- і α-захворювання), лімфом, індукованих лікуванням імуносупресантами, наприклад, циклоспориніндукованої лімфоми і метотрексат-індукованої лімфоми. 30. Спосіб лікування або запобігання імунного захворювання, при якому імунокомпетентні клітини експресують CD20, який включає застосування до суб‘єкта антитіла, відповідно до будь-якого з пп. 17, композиції, відповідно до будь-якого з пп. 16-18, імунокон‘югата відповідно до п. 19, або біспецифічної молекули відповідно до будь-якого з пп. 2021, експресуючого вектора відповідно до будьякого з пп. 43-44 у кількості, яка є ефективною для лікування або запобігання імунного захворювання. 31. Спосіб за п. 30, відповідно до якого лікування включає знищення B-клітин, які продукують антитіла проти аутологічних антигенів. 32. Спосіб за п. 30, відповідно до якого захворювання або розлад вибраний з групи, що складається з псоріазу, псоріатичного артриту, дерматиту, системних склеродерми і склерозу, запального захворювання кишечнику (IBD), хвороби Крона, виразкового коліту, синдрому дихальної недостатності, менінгіту, енцефаліту, увеїту, гломерулонефриту, екземи, астми, атеросклерозу, недостатності адгезії лейкоцитів, розсіяного склерозу, синдрому Рейно, синдрому Шегрена, ювенільного діабету, хвороби Рейтера, хвороби Бехчета, імунокомплексного нефриту, нефропатії IgA, полінейропатії IgM, імуноопосердкованої тромбоцитопенії, наприклад гострої ідіопатичної тромбоцитопенічної пурпури і хронічної ідіопатичної тромбоцитопенічної пурпури, гемолітичної анемії, злоякісної міастенії, вовчакового нефриту, системного вовчакового еритематозу, ревматоїдного артриту (RA), атопічного дерматиту, пухирчатки, хвороби Грейвса, тиреоїдиту Хасімото, гранулематозу Вегенера, синдрому Омена, хронічної ниркової недостатності, гострого інфекційного мононуклеозу, ВІЛ і захворювань, пов‘язаних з вірусом герпесу. 7 93478 8 33. Спосіб за п. 32, відповідно до якого аутоімунне захворювання являє собою ревматоїдний артрит (RA). 34. Спосіб за п. 30 відповідно до якого захворювання являє собою запальний, імунний і/або аутоімунний розлад, вибраний з виразкового коліту, хвороби Крона, ювенільного діабету, розсіяного склерозу, імуноопосердкованої тромбоцитопенії, наприклад гострої ідіопатичної тромбоцитопенічної пурпури і хронічної ідіопатичної тромбоцитопенічної пурпури, гемолітичної анемії, злоякісної міастенії, системного склерозу і пухирчатки звичайної. 35. Спосіб за п. 30, відповідно до якого захворювання являє собою запальний, імунний і/або аутоімунний розлад, вибраний з запального захворювання кишечнику (IBD), виразкового коліту, хвороби Крона і розсіяного склерозу. 36. Спосіб за будь-яким з пп. 22-35, який додатково включає окреме застосування до суб‘єкту іншого терапевтичного агента. 37. Спосіб за п. 36, відповідно до якого терапевтичний агент являє собою (i) цитотоксичний агент або радіотоксичний агент; (ii) імуносупресант; або (iii) імунологічний модулюючий агент, наприклад цитокін або хемокін. 38. Спосіб за п. 36, відповідно до якого терапевтичний агент вибраний з групи, що складається з доксорубіцину, цисплатину, блеоміцину, кармустину, хлорамбуцилу і циклофосфаміду. 39. Спосіб за п. 36, відповідно до якого терапевтичний агент вибраний з групи, що складається з антитіл проти CD25, антитіл проти CD19, антитіл проти CD21, антитіл проти CD22, антитіл проти CD37, антитіл проти CD38, антитіл проти IL6R, антитіл проти IL8, антитіл проти IL15, антитіл проти IL15R, антитіл проти CD4, анти-CD11a, антитіл проти альфа-4/бета-1 інтегрин (VLA4), CTLA4-Ig і антитіл проти C3b(i). 40. Спосіб виявлення іn vitro присутності CD20антигену або клітини, яка експресує CD20, у зразку, який включає: введення зразка в контакт з антитілом за будьяким з пп. 1-7 в умовах, які дозволяють утворити комплекс між антитілом і CD20; і виявлення утворення комплексу. 41. Набір для виявлення присутності CD20антигену або клітини, яка експресує CD20, у зразку, який містить антитіло за будь-яким з пп. 1-7. 42. Спосіб виявлення іn vivo присутності CD20антигену або клітини, яка експресує CD20, у суб‘єкта, який включає: введення антитіла за будь-яким з пп. 1-7 суб‘єкту в умовах, які дозволяють утворити комплекс між антитілом і CD20; і виявлення утвореного комплексу. 43. Експресуючий вектор, кодуючий моноклональне антитіло людини, яке зв‘язується з CD20 людини, який містить послідовність нуклеотидів, яка кодує варіабельну область важкого ланцюга імуноглобуліну, що містить послідовність нуклеотидів, вибрану з групи, що складається з послідовностей нуклеотидів, як це зазначено в послідовностях SEQ ID NO:1 і 3, і варіабельну область легкого ланцюга імуноглобуліну, що містить послідовність нуклеотидів, вибрану з групи, що складається з послідовностей нуклеотидів, як це зазначено у послідовностях SEQ ID NO:5 і 7. 44. Експресуючий вектор, кодуючий моноклональне антитіло людини, яке зв‘язується з CD20 людини, який містить послідовність нуклеотидів, яка кодує варіабельну область важкого ланцюга імуноглобуліну, що містить послідовність амінокислот, вибрану з групи, що складається з послідовностей амінокислот, як це зазначено в послідовностях SEQ ID NO:2 та 4, і варіабельну область легкого ланцюга імуноглобуліну, що містить послідовність амінокислот, вибрану з групи, що складається з послідовностей амінокислот, як це зазначено у послідовностях SEQ ID NO:6 та 8. 45. Фармацевтична композиція, яка містить експресуючий вектор за будь-яким з пп. 43-44 і фармацевтично прийнятний носій. 46. Антиідіотипічне антитіло, яке зв‘язується з антитілом за будь-яким з пп. 1-7. 47. Антиідіотипічне антитіло у відповідності до п. 46, яке зв‘язується з 2F2 або 7D8. 48. Використання антиідіотипічного антитіла за п. 46 або 47 для виявлення рівня вмісту моноклонального антитіла людини проти CD20 у зразку. У цій заявці заявлено пріоритет на основі попередньої заявки США Nо.: 60/419163, поданої 17 жовтня 2002 року, і попередньої заявки США No.: 60/460028, поданої 2 квітня 2003 року, обидві під назвою ЛЮДСЬКІ МОНОКЛОНАЛЬНІ АНТИТІЛА ДО CD20, і обидві включені до цієї заявки в повному об'ємі. Передумови створення винаходу Молекула CD20 (яка також називається людський В-лімфоцит-обмежений диференційований антиген або Вр35) представляє собою гідрофобний трансмембранний білок з молекулярною масою приблизно 35 kD, який міститься на пре-В і зрілих В-лімфоцитах (Valentine та ін. (1989) J. Вiol. Chem. 264(19):11282-11287; і Einfield та ін. (1988) ЕМВО J. 7(3):711-717). CD20 виявлений на поверхні більше, ніж 90% В-клітин периферійних кров'яних судин або лімфоїдних органів, і встановлено, що він експресується на стадії початкового пре-Вклітинного розвитку і запишається в них до диференціації клітин плазми. CD20 присутній як в нормальних В-клітинах, так і в пухлинних В-клітинах. Зокрема, CD20 експресується більше ніж у 90% Вклітин неходжкінської лімфоми (NHL) (Anderson та ін. (1984) Blood 63(6):1424-1433), але не виявлений у гемопоетичних стовбурових клітинах, про-Вклітинах, нормальних клітинах плазми або в інших нормальних тканинах (Tedder та ін. (1985) J. Immunol. 135(2):973-979). 9 Ділянка білка CD20, яка складається з 85 амінокислот і має карбоксильний кінець, розташована всередині цитоплазми. Довжина цієї ділянки помітно відрізняється від властивих для В-клітин поверхневих структур, таких як важкі ланцюги IgM, IgD і IgG або ланцюги антигенів гістосумісності класу II або , які мають відносно короткі інтрацитоплазматичні ділянки, що складаються з 3, 3, 28, 15 і, відповідно, 16 амінокислот (Komaromy та ін. (1983) NAR 11:6775-6785). Із 61 амінокислот з карбоксильним кінцем 21 представляють собою кислотні залишки, тоді як лише 2 є основними, що свідчить про те, що ця ділянка має сильний результуючий негативний заряд. Номер доступу в The GenBank NP_690605. Існує думка, що CD20 може використовуватись для регулювання ранніх стадій активаційного і диференційного процесу у В-клітин (Tedder та ін. (1986) Eur.J, Immunol. 16:881-887) і може виконувати функції іонних кальцієвих каналів (Tedder та ін. (1990) J. Cell. Biochem. 14D:195). Незважаючи на невизначеність щодо реальної ролі CD20 у сприянні проліферації і/або диференціації В-клітин, він є важливим об'єктом терапії з використанням антитіл, направленої на боротьбу з В-клітинами, з якими пов'язані ракові та аутоімунні захворювання, або їх знищенням. Зокрема, експресія CD20 на пухлинних клітинах, наприклад NHL, робить його важливим об'єктом для терапії з використанням антитіл проти СР20-позитивних неопластичних клітин. Проте, тоді як отримані на сьогодні результати чітко вказують на те, що CD20 може використовуватись в імунотерапії, вони також вказують на те, що наявні мишачі і химерні антитіла не є ідеальними терапевтичними агентами. Відповідно, існує потреба в удосконалених терапевтичних антитілах проти CD20, які були б ефективними щодо профілактики і/або лікування ряду захворювань, пов'язаних з клітинами, які експресують CD20. Короткий виклад суті винаходу Цей винахід стосується покращених лікарських агентів на основі антитіл для лікування і/або профілактики захворювань, пов'язаних з клітинами, які експресують CD20, включно з пухлинними захворюваннями, та імунними захворюваннями, включно з аутоімунними захворюваннями. Антитіла, які є предметом цього винаходу, удосконалені у тому, що вони є повністю людськими, а відтак є потенційно менш імуногенними для пацієнтів. Як проілюстровано у цьому винаході, людські антитіла згідно з винаходом виступають медіаторами знищення В-клітин за рахунок експресії CD20 через ряд механізмів. Відповідно до одного з варіантів людські антитіла згідно з винаходом індукують комплементарну залежну цитотоксичність (CDC), наприклад принаймні приблизно 20% CDCопосередкований лізис, переважно приблизно 30% CDC-опосередкований лізис і більш переважно 4050%-опосередкований лізис, такий як у випадку клітин хронічної В-лейкемії (B-CLL). Відповідно до ще одного варіанту людські антитіла згідно з винаходом індукують апоптоз клітин, які експресують CD20. Відповідно до ще одного варіанту людські 93478 10 антитіла згідно з винаходом індукують гомотипічну адгезію клітин, які експресують CD20. Крім того, людські антитіла згідно з винаходом можуть індукувати залежну від антитіл клітинну цитотоксичність (ADCC) у клітин, які експресують CD20 у присутності людських клітин-ефекторів (наприклад, моноцити, мононуклеарні клітини, NK-клітини (натуральні клітини-кіллери) і ПМН). Крім того, людські антитіла згідно з винаходом можуть індукувати фагоцитоз клітин, які експресують CD20 у присутності макрофагів. Людські моноклональні антитіла згідно з винаходом можуть діяти відповідно до одного або більше з таких механізмів. Приклади клітин, які можуть умертвлятись людськими антитілами згідно з винаходом, включають, але не обмежуються цим переліком, В-клітини, які експресують CD20, такі як канцерогенні В-клітини і Вклітини, з якими пов'язані аутоімунні захворювання. В особливо переважному варіанті винаходу людські антитіла використовують для опосередкування умертвлення В-лімфоцитів у ході лікуванні лімфоми, наприклад В-клітинної не лімфоми Ходжкіна. Людські антитіла згідно з винаходом включають антитіла lgG1 (наприклад, lgG1,k). lgG3 (наприклад, lgG3, k і lgG4 (наприклад, lgG4, k). Проте, винахід також включає інші ізотипи антитіл і включає lgG2, IgM, lgA1, lgA2, секреторні антитіла IgA, IgD i lgE. Такі антитіла можуть представляти собою повні антитіла або їх антиген-зв'язуючі фрагменти, які включають, наприклад, Fab, F(ab')2, Fv, одноланцюгові Fv-фрагменти або біспецифічні антитіла. Крім того, такі антиген-зв'язуючі фрагменти включають синтезовані білки імоноглобуліну із зв'язуючим доменом, які містять (і) зв'язуючий поліпептидний домен (такий як змінна ділянка важкого ланцюга або змінна ділянка легкого ланцюга), злитий з поліпептидом із зв'язуючої ділянки імуноглобуліну, (іі) незмінна ділянка імуноглобулінового важкого ланцюга СН2, злита зі зв'язуючою ділянкою, і (ііі) незмінна ділянка імуноглобулінового важкого ланцюга СН3, злита з незмінною ділянкою СН2. Такі синтезовані білки імуноглобуліну із зв'язуючими доменами більш детально описані в заявках US 2003/0118592 і US 2003/0133939. Особливо переважні антитіла згідно з винаходом включають антитіла 11В8, 2F2 і 7D8, які кодуються людськими нуклеїновими кислотами з важкими ланцюгами та каппа-нуклеїновими кислотами з легкими ланцюгами, які включають нуклеотидні послідовності у змінних ділянках, як показано у ІД. № ПОСЛІДОВНОСТІ: 1, 5 або 9 і ІД. № ПОСЛІДОВНОСТІ: 3, 7 або, відповідно, 11, та їх консервативні модифікації. Відповідно до ще одного варіанту людські антитіла характеризуються тим, що вони мають змінні ділянки з важкими ланцюгами і каппалегкими ланцюгами, які складаються з амінокислотних послідовностей як показано в ІД. № ПОСЛІДОВНОСТІ: 2, 6 або 10 і ІД. № ПОСЛІДОВНОСТІ: 4, 8 або, відповідно, 12, та їх консервативні модифікації. Відповідно до ще одного варіанту людські антитіла характеризуються тим, що вони мають змінні ділянки з важкими ланцюгами і каппалегкими ланцюгами, які принаймні на 90% є гомо 11 логічними, переважно принаймні на 95% гомологічними і більш переважно принаймні на 98% або принаймні на 99% гомологічними до амінокислотних послідовностей яке показано у ІД. № ПОСЛІДОВНОСТІ: 2 і ІД. № ПОСЛІДОВНОСТІ: 4; ІД. № ПОСЛІДОВНОСТІ: 6 і, відповідно, ІД. № ПОСЛІДОВНОСТІ: 8; або ІД. № ПОСЛІДОВНОСТІ: 10 і, відповідно, ІД. № ПОСЛІДОВНОСТІ: 12. Інші переважні людські антитіла згідно з винаходом включають антитіла, які містять CDR-домен, який має ділянку CDR1 з людським важким і легким ланцюгом, ділянку CDR2 з важким і легким ланцюгом і ділянку CDR3 з людським важким і легким ланцюгом, де (a) ділянки CDR1, CDR2 і CDR3 людського важкого ланцюга містять амінокислотну послідовність, вибрану з групи, яка складається з амінокислотних послідовностей CDR1, CDR2 і CDR3, показаних на Фігурах 53, 55 або 57 (ІД. № ПОСЛІДОВНОСТІ: 13-15, 19-21 і 25-27), та консервативні модифікації послідовностей, і (b) ділянки CDR1, CDR2 і CDR3 людського легкого ланцюга містять амінокислотну послідовність, вибрану з групи, яка складається з амінокислотних послідовностей CDR1, CDR2 і CDR3, показаних на Фігурах 53, 55 або 57 (ІД. № ПОСЛІДОВНОСТІ: 16-18, 22-24 і 28-30), та консервативні модифікації послідовностей. Також цей винахід включає антитіла, які дисоційовані від CD20 з рівноважною константою дисоціації (KD) приблизно 1-10 нМ або менше. Такі антитіла також включають антитіла, які не вступають у перехресні реакції зі спорідненими клітинноповерхневими антигенами, а відтак не інгібують їх функції. Відповідно до ще одного варіанту людські антитіла проти CD20 можуть характеризуватися однією або більше властивостями: a) специфічність до людського CD20; b) подібність у зв'язуванні до CD20 (КD) приблизно 10 нМ або менше, переважно приблизно 5 нМ або менше і більш переважно приблизно 1-3 нМ або менше, що визначається відповідно до експерименту на зв'язування, описаного у Прикладі 5 (Фігура 9) цього опису; c) константа швидкості дисоціації (kd) від CD20 приблизно 10-4 сек-1 або менше, переважно приблизно 10-5 сек-1 або менше і більш переважно приблизно 10-6 сек-1 або менше, що визначається з допомогою експерименту на визначення швидкості дисоціації, описаного у Прикладі 5 (Фігура 9); d) здатність виступати медіатором високого рівня CDC або на CD55/59 негативних, або на CD55/59 позитивних клітинах; е) здатність до транслокації в накопичення ліпідів при зв'язуванні з CD20; f) здатність інгібувати ріст клітин, які експресують CD20; g) здатність індукувати апоптоз клітин, які експресують CD20; h) здатність індукувати гомотипову адгезію клітин, які експресують CD20; і) здатність індукувати ADCC у клітин, які експресують CD20 у присутності клітин ефекторів; 93478 12 j) здатність пролонгувати життя суб'єкта, який має пухлинні клітини, що експресують CD20; к) здатність знищувати клітини, які експресують CD20; і/або І) здатність знищувати клітини, які експресують низькі рівні CD20 (клітини CD20low). Людські антитіла проти CD20 згідно з винаходом можуть бути дериватизовано зв'язані з або співекспресовані з іншими зв'язуючими компонентами. Відповідно до переважного варіанту винахід стосуєтьсябіспецифічної або мультиспецифічної молекули, яка містить принаймні перший специфічний зв'язуючий компонент для CD20 (наприклад, людське антитіло проти CD20 або його міметик) та другий специфічний зв'язуючий компонент для людської клітини-ефектора, такий як специфічний зв'язуючий компонент для Fc-рецептора (наприклад, людський рецептор Fс, такий як FcRI, або людський рецептор Fc) або Т-клітинний рецептор, наприклад CD3. Відповідно цей винахід охоплює біспецифічні і мультиспецифічні молекули, які зв'язуються як з людським CD20, так і з Fc-рецептором або Тклітинним рецептором наприклад CD3. Приклади Fc-рецепторів включають людський рецептор IgG, Fc-гамма рецептор (FcR), такий як FcRI (CD64), FcRII (CD32) і FcRIІІ (CD16). Також можна застосовувати інші Fc-рецептори, такі як людські lgAрецептори (наприклад FcRI). Рецептор переважно розташовується на поверхні клітини-ефектора, наприклад моноцити макрофага або активованої мононуклеарної клітини. Відповідно до переважного варіанту винаходу біспецифічні і мультиспецифічні молекули зв'язуються з Fc-рецептором у мі відмінному від імуноглобулінового місця зв'язування Fc-рецептора (наприклад, IgG або IgA). Тому зв'язування біспецифічних і мультиспецифічних молекул не блокується фізіологічними рівнями імуноглобулінів. Відповідно до ще одного аспекту людські антитіла проти CD20 згідно з винаходом дериватизовані, зв'язані або ко-експресовані з іншою поліфункціональною молекулою, наприклад, з іншим пептидом або білком (наприклад, з фрагментом Fab'). Наприклад, антитіло згідно з винаходом може бути функціонально зв'язаним (наприклад, хімічним сполученням, генетичним злиттям, нековалентною асоціацією або іншим шляхом) з однією або більше молекулярними структурами, такими як інше антитіло (наприклад, з утворенням біспецифічного або мультиспецифічного антитіла), цитотоксин, клітинний ліганд або антиген (наприклад, з одержанням імунокон'югату, такого як імунотоксин). Антитіло згідно з винаходом може бути зв'язане з іншими терапевтичними компонентами, наприклад, з радіоізотопом, дрібномолекулярною протираковою лікарською речовиною, протизапальним агентом або імуносупресивним агентом. Відповідно цей винахід охоплює цілий ряд кон'югатів антитіл, біспецифічних і мультиспецифічних молекул і злитих білків, всі з яких зв'язуються з клітинами, які експресують CD20, і які можуть використовуватись для зв'язування інших молекул до таких клітин. 13 Відповідно до ще одного аспекту винахід стосується композицій, наприклад фармацевтичних і діагностичних композицій/наборів, які включають фармацевтично прийнятний носій та одне або комбінацію людських моноклональних антитіл згідно з винаходом. Відповідно до переважного варіанту така композиція включає комбінацію антитіл, які зв'язуються з окремими епітопами або які мають специфічні функціональні властивості, такі як індукування CDC і індукування апоптозу. Людські антитіла, імунокон'югати, біспецифічні і мультиспецифічні молекули і композиції згідно з цим винаходом можуть застосовуватись у ряді методик для інгібування росту клітин, які експресують CD20, і/або умертвлення клітин, які експресують CD20, шляхом введення таких клітин у взаємодію з ефективною кількістю антитіла, імунокон'югата, біспецифічної/мультиспецифічної молекули або композиції, внаслідок чого ріст клітин інгбіується і/або клітини гинуть. Відповідно до одного варіанту спосіб полягає в умертвленні клітин, які експресують CD20 у присутності клітинефекторів, наприклад за рахунок CDC, апоптозу, ADCC, фагоцитозу або поєднання двох або більше із названих механізмів. Клітини переважно умертвлюють або інгібують без умертвлення або інгібування активності клітин, які не експресують CD20, але які можуть, наприклад, експресувати структурно споріднені антигени на поверхні клітин (тобто без вступання в перехресну реакцію зі спорідненими, але функціонально відмінними поверхневими антигенами клітин). Клітини, які експресують CD20, які можуть інгібуватися або умертвлятися з використанням людських антитіл згідно з винаходом, включають, наприклад, пухлинні В-клітини. Відповідно людські антитіла згідно з винаходом можуть використовуватись для лікування і/або профілактики ряду захворювань, пов'язаних з клітинами, які експресують CD20, шляхом їх призначення пацієнтам, які страждають від таких захворювань. Приклади захворювань, які можуть лікуватися (наприклад, полегшуватися) або запобігатися, включають, але не обмежуються цим переліком пухлинні та імунні захворювання, наприклад аутоімунні захворювання. Приклади пухлинних захворювань, які можуть лікуватися і/або запобігатися, включають В-клітинну лімфому, наприклад, NHL, включно з прекурсорною Вклітинною лімфобластичною лейкемією/лімфомою і зрілими В-клітинними неоплазмами, такими як хронічний В-клітинний лімфолейкоз (СLL)/Вклітинна лімфома (SLL), В-клітинний пролімфолейкоз, лімфоплазмацитрана лімфома, лімфома клітин, які одягають спорангій (MCL), фолікулярна лімфома (FL), включно з ранньою, середньою та пізньою FL, шкірна лімфома клітин центрального фолікула, В-клітинна лімфома краєвого поясу (у лімфоїдній тканині слизових оболонок (MALT),y вузлах і селезінці), лейкоз ворсистих клітин, дифузна лімфома великих пре-В лімфоцитів, лімфома Беркіта, плазмацитома, хвороба Калера, посттрансплантаційне лімфопроліферативне захворювання, хвороба Вальденстрьома (макроглобулінемія) і анапластична крупноклітинна лімфома (ALCL). Приклади імунних захворювань, пов'яза 93478 14 них з В-клітинами, які експресують CD20, які можуть лікуватися і/або запобігатися згідно з винаходом, включають псоріаз, псоріатичний артрит, дерматит, системну склеродерму і склероз, запальне захворювання кишечника (IBD), хворобу Крона, виразковий коліт, синдром дихальної недостатності, менінгіт, енцефаліт, увеїт, гломерулонефрит, екзема, астма, атеросклероз, недостатність злипання лейкоцитів, розсіяний склероз, синдром Рейно, синдром Шегрена, діабет у молодих людей, хвороба Рейтера, хвороба Бехччета, імунокомплексний нефрит, нефропатію ІgА, полінейропатію ІgМ, імуно-обумовлена тромбоцитопенію, таку як гостра ідіопатична тромбоцитопенічна пурпура і хронічна ідіопатична тромбоцитопенічна пурпура, гемолітична анемія, астенічний бульбарний параліч, люпус-нефріт, системний люпусериматоз, ревматоїдний артрит (RA), атопічний дерматит, пемфігус, хвороба Грейвса, тіреоїдит Хашімото, гранульоматоз Венегера, синдром Оменна, хронічна ниркова недостатність, гострий інфекційний мононуклеоз, ВІЛ і захворювання, пов'язані з вірусом герпеса. Додаткові приклади включають тяжкі форми гострого синдрому дихальної недостатності і хореоретиніт. Додаткові приклади включають захворювання і розлади, зумовлені інфікуванням В-клітин вірусом, таким як вірус Епштейна-Барра (EBV). Відповідно до переважного варіанту винаходу суб'єкту, якому призначають антитіла, додатково призначають хіміотерапевтичний агент, опромінення або агент, який модулює, наприклад, посилює або інгібує експресію або активність Fcрецептора, наприклад, Fc -рецептора або Fc рецептора, такого як цитокін. Типові цитокіни для призначення під час лікування включають фактор стимулювання гранулоцитної колонії (G-CSF), фактор стимулювання гранулоцитно-макрофагової колонії (GM-CSF), інтерферону (IFN-) і фактор некрозу пухлини (TNF). Типові терапевтичні агенти включають, окрім інших, анти-неопластичні агенти, такі як доксорубіцин, цисплатин, блеоміцин, кармустин, хлорамбуцил і циклофосфамід. Відповідно до ще одного аспекту даний винахід стосується способу виявлення in vitro або in vivo присутності CD20 у зразку або у індивіда, наприклад, для діагностики пов'язаного з CD20 захворювання, переважно на ранній стадії. Це також може бути корисним для моніторингу протікання захворювання та ефективності лікування, а також для визначення дози антитіла для призначення. Методика in vivo може бути здійснена з використанням методики формування зображень, такої як PET (позитронна емісійна томографія) або SPECT (однофотонна емісійна комп'ютерна томографія). Відповідно до одного варіанту це досягається за рахунок введення у взаємодію тестованого зразка, необов'язково разом із контрольним зразком, з людським моноклональним антитілом згідно з винаходом за умов, які сприяють утворенню комплексу між антитілом і CD20. Після цього визначають утворення комплексу (наприклад, з використанням аналіз FACS або вестерн-блоттингу). У випадку використання разом із тестованим зразком контрольного зразка, утворення комплексу виявляєть 15 ся в обох зразках, і будь-яка значна відмінність у формуванні комплексів між зразками свідчить про присутність CD20 у тестованому зразку. Відповідно до ще одного аспекту винахід стосується трансгенного ссавця, відмінного від людини, такого як трансгенна миша, який експресує людські моноклональні антитіла, що зв'язуються з CD20. Відповідно до переважного варіанту трансгенним ссавцем, відмінним від людини, є трансгенна миша з геномом, який включає людський трансген з важким ланцюгом і людський трансген з легким ланцюгом, який кодує весь або частину антитіла згідно з винаходом. Такий трансгенний ссавець, відмінний від людини, може бути імунізований очищеним або збагаченим препаратом на основі антигену CD20 і/або клітин, які експресують CD20. Переважно такий трансгенний ссавець, відмінний від людини, наприклад трансгенна миша, здатен виробляти множинні ізотипи людських моноклональних антитіл до CD20 (наприклад, IgG, IgA і/або IgM) за рахунок рекомбінації V-D-J і переключення ізотипів. Переключення ізотипів може відбуватися як, наприклад, класичним, так і не класичним шляхом переключення. Відповідно, згідно із ще одним аспектом винахід стосується ізольованих В-клітин трансгенного ссавця, відмінного від людини, як описувалося вище, наприклад, трансгенної миші, який виробляє людські антитіла проти CD20. Ізольовані В-клітини можуть бути законсервовані за рахунок злиття із законсервованою клітиною з одержанням джерела (наприклад, гібридоми) людських антитіл проти CD20. Такі гібридоми (наприклад, ті, що виробляють людські антитіла проти CD20) також входять в об'єм правової охорони цього винаходу. Як показано у прикладах цього винаходу, людські антитіла згідно з винаходом можуть бути безпоcередньо одержані з гібридом, які експресують таке антитіло, або вони можуть бути клоновані і рекомбінантним шляхом експресовані у клітинухазяїна (наприклад, СНО-клітину, NS/0-клітину або лімфоцит). Додаткові приклади клітин-хазяїв включають мікроорганізми, такі як Е. соlі, та гриби, такі як дріжджі. Як альтернатива, вони можуть вироблятися через рекомбінантний механізм у трансгенного ссавця, відмінного від людини, або рослини. Відповідно, згідно з іншим аспектом цей винахід стосується способів одержання людських моноклональних антитіл, здатних зв'язуватись з людським CD20. Відповідно до одного варіанту такий спосіб включає імунізацію трансгенного ссавця, відмінного від людини, наприклад, трансгенної миші, як описувалося раніше (наприклад, зумовлюючи часткове або повне кодування геномом, який включає людський трансген з важким ланцюгом і людський трансген з легким ланцюгом, антитіла проти CD20), очищеним або збагаченим препаратом людського антигену CD20 і/або клітинами, які експресують людський CD20. Після цього одержують В-клітини (наприклад, В-клітини селезінки) ссавця і зливають їх з клітинами мієломи з одержанням «безсмертних» клітин гібридом, які секретують людські моноклональні антитіла проти CD20. 93478 16 Відповідно до ще одного аспекту винахід стосується молекул нуклеїнової кислоти, які кодують людські антитіла проти CD20 (наприклад, їх змінні ділянки), а також рекомбінантних векторів експресії, які включають нуклеїнові кислоти згідно з винаходом, та клітин-хазяїв, трансфікованих такими векторами. Способи одержання антитіл культивуванням клітин-хазяїв також входять до об'єму цього винаходу. Особливо переважні нуклеїнові кислоти згідно з винаходом включають нуклеотидні послідовності, показані на ІД. № ПОСЛІДОВНОСТІ: 1, 5 або 9 і ІД. № ПОСЛІДОВНОСТІ: 3, 7 або 11, які кодують важкі і легкі ланцюги людських антитіл проти CD20, а саме 2F2, 7D8 і 11В8. Інші ознаки та переваги цього винаходу будуть очевидними з наведеного далі детального опису та формули винаходу. Короткий опис креслень На Фігурі 1 показаний змінний вектор з важким ланцюгом pCON1f, який використовують для рекомбінантного продукування людських моноклональних антитіл 2F2 і 11В8. На Фігурі 2 показаний змінний вектор з легким ланцюгом рСОNк, який використовують для рекомбінантного продукування 2F2 і 11В8. На Фігурі 3 показаний клонуючий вектор з двома генами (pCON1f/k2F2), який використовують для рекомбінантного продукування 2F2 і 11В8. На Фігурі 4 показана крива, на якій порівнюється зв'язування людських моноклональних антитіл 2F2, 7D8 і 11В8 з Raji, Daudi і CD20, трансфікованими NS/0-клітинами і материнськими NS/0клітинами, з допомогою проточної цитометрії. На Фігурах 5А і 5В показане зв'язування 2F2 з PBMCs від трьох людей-донорів з використанням проточної цитометрії. На Фігурі 6 показаний графік, який порівнює подібність зв'язування 125І-міченого 2F2 і 125Іміченого 11В8 з клітинами Рамоса-EHRB. На Фігурах 7А і 7В показане зв'язування 125Іміченого 2F2 і 125І-міченого 11В8 порівняно з 125Іміченим ритуксимабом (химерним антитілом проти CD20, IDEC) і 125І-міченим В1 (позначення В1 відповідає неміченій формі Веххаr™, що представляє собою 131І-мічене мишаче антитіло проти CD20, Coulter) з клітинами Рамоса-EHRB (А) і клітинами Дауді (В). На Фігурі 8 показана діаграма, на якій порівнюється швидкість дисоціації 125I-міченого 11В8Т, 125 І-міченого 2F2,125І-міченого ритуксимабу (RIT) і 125 І-міченого В1. На Фігурі 9 показана швидкість дисоціації фрагментів F(ab')2 у 2F2, 11В8Т і ритуксимабу в клітинах Рамоса-EHRB. На Фігурах 10А і 10В показана комплементарна залежна токсичність (CDC) у 2F2T, 11В8Т, 7D8, ритуксимабу і ізотипічного контрольного антитіла (HuMab-KLH) клітин Дауді (А) і клітин SU-DHL-4 (В) у різні моменти часу (функціональні відхилення) з використанням проточної цитометрії. На Фігурах 11А-Е представлена кінетика комплементарної залежної токсичності (CDC), індукована 2F2 і ритуксимабом у різних лініях клітин, з використанням проточної цитометрії. 17 На Фігурах 12A-D представлена CDC, індукована 2F2 і ритуксимабом у різних ліній клітин, через функцію концентрації комплемента (нормальна людська сироватка (NHS)) при двох різних концентраціях антитіл, використовуючи проточну цитометрію. На Фігурах 13A-D показана залежна від концентрації індукція CDC під впливом 2F2 і ритуксимабу у різних клітинних лініях з використанням проточної цитометрії. На Фігурах 14А і 14В показана залежна від концентрації індукція токсичності CDC під дією 2F2, 2F2T, 11В8Т, В1 і ритуксимабу в клітинах Дауді (А) і клітинах Раджі (В). На Фігурах 15А і 15В представлені діаграми, на яких порівнюється комплементарна залежна токсичність клітин Дауді (клітини, які експресують низькі рівні CD55/59) під дією моноклонапьних антитіл 2F2, 7D8 і 11В8 та ритуксимабу; (А) показує відсоток лізису непромитих клітин і (В) показує відсоток лізису клітин, які промивали перед додаванням сироватки. На Фігурах 16А і 16В представлені діаграми, які порівнюють комплементарну залежну токсичність клітин Раджі (клітини, які експресують значні кількості CD55/59), викликану людськими моноклональними антитілами 2F2, 7D8 і 11В8 та ритуксимабом; (A) вказує на відсоток лізису клітин, які не блокуються антитілами проти CD55 і CD59, і (B) вказує на відсоток лізису клітин, блокованих антитілами проти CD55 і CD59. На Фігурах 17А-С проілюстрована роль CD55 і CD59 в токсичності CDC, індукованій 2F2 і ритуксимабом у клітинах Раджі. (А) вказує на відсоток розчинених клітин при додаванні антитіла проти CD55, (В) вказує на відсоток розчинених клітин при додаванні антитіл проти CD59, і (С) вказує на відсоток розчинених клітин при додаванні антитіл проти CD55 і CD59. На Фігурах 18A-D показане зв'язування комплементного фактора С1q з допомогою 2F2 і ритуксимабу у різних лініях клітин як показують дані проточної цитометрії. На Фігурах 19A-D показане відкладення фрагмента комплементного фактора С4с під дією 2F2 і ритуксимабу у різних клітинних лініях, як показують дані проточної цитометрії. На Фігурі 20 показаний лізис of ARH-77 cells by 2F2, ритуксимаб, і 11В8Т у присутності of PMNs, MNCs, plasma or whole blood. На Фігурі 21 показаний лізис клітин B-CLL під дією 2F2, ритуксимабу і 11В8Т у присутності PMNs, MNCs, плазми абонезбираної крові. На Фігурі 22 показаний лізис клітин HCL (лейкозу ворсистих клітин) під дією 2F2, ритуксимабу і 11В8Т у пристуності PMNs, MNCs, плазми або незбираної крові. На Фігурі 23 показаний лізис клітин B-ALL під дією 2F2 і ритуксимабу у присутності PMNs, MNCs, плазми або незбираної крові. На Фігурі 24 показаний лізис клітин фолікулярної лімфоми (FL) під дією 2F2, ритуксимабу і 11В8Т у присутності PMNs, MNCs, плазми або незбираної крові. 93478 18 На Фігурі 25 показаний лізис клітин лімфоми клітин, які одягають спорангій, під дією 2F2, ритуксимабу і 11В8Т у пристуності PMNs, MNCs, плазми або незбираної крові. На Фігурі 26 показаний залежний від концентрації лізис клітин ARH-77 під дією 2F2 і ритуксимабу у пристуності незбираної крові. На Фігурі 27 показаний опосередкований MNC лізис клітин ARH-77 під дією 2F2T, 11В8Т і ритуксимабу. На Фігурі 28 показаний опосередекований MNC лізис клітин Раджі під дією 2F2T, 11В8Т і ритуксимабу. На Фігурах 29А, В і С представлені діаграми, на яких показана кластеризація CD20 25 у ліпідних масах під час інкубування з 2F2, 7D8 або 11В8, складені з використанням аналізу FRET (резонансного переносу енергії флуоресценції) і аналізу на нерозчинність Triton-X. На Фігурі 30 показане кластерування CD20 у ліпідних масах під час інкубування з 2F2, ритуксимабом або 11В8Т, з використанням аналізу FRET. На Фігурі 31 показане співвідношення CD20, які залишаються у фракції нерозчинної маси після обробки з використанням Triton Х-100 (ТХ) і інкубування з використанням 2F2, ритуксимабу або 11В8Т. На Фігурі 32 показаний розподіл CD20 між масовими і немасовими мембранними фракціями при стимулюванні клітин Дауді з допомогою 2F2, ритуксимабу або 11В8Т. На Фігурах 33A-G показаний апоптоз клітин Дауді під дією 2F2, 7D8 і 11В8, що отримано з допомогою проточної цитометрії. На Фігурі 34 показане індукування апоптозу клітин Раджі з допомогою 2F2, 11В8Т, ритуксимабу або В1 з використанням проточної цитометрії. На Фігурі 35А показане індукування апоптозу клітин Дауді з допомогою 2F2T, 11В8Т, ритуксимабу або В1 з використанням прточної цитометрії. На Фігурі 35В показана рання стадія і пізня стадія апоптозу клітин Дауді під дією людських моноклональних антитіл 2F2T, 11В8Т, ритуксимабу і В1 з використанням проточної цитометрії. На Фігурах 36А-Е показана гомотипова адгезія клітин Рамоса -EHRB під дією 2F2, 7D8 і 11В8 з використанням оптичної мікроскопії. На Фігурі 37 показана гомотипова адгезія клітин Дауді під дією 2F2, ритуксимабу і В1 з використанням оптичної мікроскопії. На Фігурі 38 наведена діаграма, на якій показаний відсоток виживання у мишей SCID, яким ін'єкцією призначали клітини Дауді і лікували 2F2 або 7D8. На Фігурі 39 показаний відсоток виживання мишей SCID, яким ін'єкцією призначали клітини Тануе і лікували 2F2, ритуксимабом або В1. На Фігурі 40 показаний відсоток виживання мишей SCID, яким вводили ін'єкцію клітин Дауді і лікували 2F2 або ритуксимабом в різних концентраціях. На Фігурі 41 показаний відсоток виживання мишей SCID, яким вводили ін'єкцію клітин Дауді і лікували 11В8Т або В1. 19 На Фігурі 42 показане біолюмінісцентне зображення пухлинних клітин у мишей SCID на 39 день (31 день після призначення 10 мкг В1, ритуксимабу, 11В8Т, 2F2T або hulgG1). Біолюмінісценція представлена у червоному кольорі (темні зони у миші) (інтенсивність світла > 50 фотонів на 5 хв.) у вигляді накладки на чорно-білому зображенні тіла миші. На Фігурі 43 показана пухлинна маса у кожній миші, кількісний аналіз якої проводився на 25, 32, 39 і 46 день після призначення на 8 день 10 мкг В1, ритуксимабу, 11В8Т, 2F2T або hulgG1, інтегруючи світлові сигнали над поверхнею тіла. На Фігурах 44А-С показаний проточний цитометричний аналіз клітин CD20+ у периферичній крові мавпам cynomolgus після внутрішньовенного призначення 2F2 або ритуксимабу в різних дозах, 4 х 1.25 мк/кг (А), 4 х 6.25 мг/кг (В) або 4 х 12.50 мг/кг (С). На Фігурах 45А-С показаний проточний цитометричний аналіз клітин CD21+ у периферичній крові мавпам cynomolgus після внутрішньовенного призначення 2F2 або ритуксимабу у різних дозах, 4 х 1.25 мк/кг (А), 4 х 6.25 мг/кг (В) або 4 х 12.50 мг/кг (С). На Фігурах 46А-С показаний проточний цитометричний аналіз клітин CD20+ у лімфатичних вузлах мавп cynomolgus після внутрішньовенного призначення 2F2 або ритуксимабу у різних дозах, 4 х 1.25 мк/кг (А), 4 х 6.25 мг/кг (В) або 4 х 12.50 мг/кг (С). На Фігурах 47А-С показаний проточний цитометричний аналіз клітин, які експресують CD20lowCD23+CD40high у периферичній крові мавп cynomolgus після внутрішньовенного призначення 2F2 або ритуксимабу у різних дозах, 4 х 1.25 мк/кг (А), 4 х 6.25 мг/кг (В) або 4 х 12.50 мг/кг (С). На Фігурах 48А-Е показане зв'язування ритуксимабу (A), 2F2 (В), 11В8 (С), В1 (D) або ізотипового контрольного антитіла (Е) з клітинами СНО, які експресують CD20 дикого типу (WT), мутантні CD20 (АхР), або як WT CD20, так і мутантні CD20 (АхР), про що свідчать дані проточної цитометрії. На Фігурах 49A-F показаний відсоток зв'язування 2F2,11В8Т, В1 або ритуксимабу з мутантним P172S порівняно з WT CD20 (А), відсоток зв'язування 2F2T, 11В8Т, В1, CAT (CAT 13.6E12, мишаче моноклональне lgG2A антитіло проти CD20, Diatec.Com), контрольного ізотипового антитіла (KLH) або ритуксимабу з мутантним CD20 (АхР) порівняно з WT CD20 (В), відсоток зв'язування 2F2, 11В8Т, В1 або ритуксимабу з мутантним N166D порівняно з WT CD20 (С), відоток зв'язування 2F2T, CAT або ритуксимабу з мутантним N166D порівняно з WT CD20 (D), відсоток зв'язування 2F2T, 2F2, 11B8T, B1 або ритуксимабу з мутантним N163D порвняно з WT CD20 (Е) і відсоток зв'язування 2F2T, CAT або ритуксимабу з мутантним N163D порівняно з WT CD20 (F). На Фігурі 50 показане зв'язування 2F2T, 7D8 і ізотипового контрольного антитіла, за даними імуноферментного твердофазного аналізу (ELISA), з трьома анти-ідіотиповими антитілами, aнти-2F2 sab 1.1, aнти-2F2 sab 1.2 і aнти-2F2 sab 1.3, порівняно 3 2F2. 93478 20 На Фігурі 51 показане зв'язування 11В8Т, за даними імуноферментного твердофазного аналізу (ELISA), з анти-ідіотиповими антитілами, анти11В8Т sab 2.2, анти-11В8Т sab 2.3, анти-11В8Т sab 2.4, анти-11В8Т sab 2.5, і анти-11В8Т sab 2.6, порівняно з 10 11В8Т, але не зв'язування з антиідіотиповими антитілами проти 2F2. На Фігурах 52А-С показане залежне від дозування зв'язування 2F2T, за даними аналізу ELISA, з трьома анти-ідіотиповими антитілами, aнти-2F2 sab 1.1 (A), aнти-2F2 sab 1.2 (В) і aнти-2F2 sab 1.3 (С), порівняно з 2F2. На Фігурі 53 показана амінокислотна послідовність (ІД. № ПОСЛІДОВНОСТІ:2) важкого ланцюга V ділянки амінокислотної послідовності (ІД. № ПОСЛІДОВНОСТІ:4) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 2F2 з позначеними ділянками CDR. На Фігурі 54 показана нуклеотидна послідовність (ІД. № ПОСЛІДОВНОСТІ:1) важкого ланцюга V ділянки нуклеотидної послідовності (ІД. № ПОСЛІДОВНОСТІ:3) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 2F2. На Фігурі 55 показана амінокислотна послідовність (ІД. № ПОСЛІДОВНОСТІ:6) важкого ланцюга V ділянки і амінокислотна послідовність (ІД. № ПОСЛІДОВНОСТІ:8) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 7D8 з позначеними ділянками CDR. На Фігурі 56 показана нуклеотидна послідовність (ІД. № ПОСЛІДОВНОСТІ:5) важкого ланцюга V ділянки і нуклеотидна послідовність (ІД. № ПОСЛІДОВНОСТІ:7) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 7D8. На Фігурі 57 показана амінокислотна послідовність (ІД. № ПОСЛІДОВНОСТІ:10) важкого ланцюга V ділянки і амінокислотна послідовність (ІД. № ПОСЛІДОВНОСТІ:12) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 11В8 з позначеними ділянками CDR. На Фігурі 58 показана нуклеотидна послідовність (ІД. № ПОСЛІДОВНОСТІ:9) важкого ланцюга V ділянки і нуклеотидна послідовність (ІД. № ПОСЛІДОВНОСТІ:11) легкого (каппа) ланцюга V ділянки людського моноклонального антитіла 11В8. Детальний опис винаходу Цей винахід стосується удосконалених лікарських засобів на основі антитіл для лікування і діагностики ряду захворювань, пов'язаних з клітинами, які експресують CD20. Лікарські засоби згідно з винаходом включають ізольовані людські моноклональні антитіла, які специфічно зв'язуються з епітопом, присутнім на CD20. Ізольовані людські моноклональні антитіла, які входять в об'єм цього винаходу, включають антитіла ІgА, lgG1-4, IgE, IgM I lgD. Відповідно до одного варіанту втілення винаходу в якості антитіла використовують антитіло lgG1, k або ізотип lgG1,. Відповідно до ще одного варіанту в якості антитіла використовують антитіло lgG3, більш переважно lgG3, k або ізотип lgG3, . Відповідно до ще одного варіанту втілення винаходу в якості антитіла викорисотовують антитіло lgG4, більш переважно lgG4, k або ізотип lgG4, . 21 Відповідно до ще одного варіанту використовують антитіло ІgА1 або ІgА2. Відповідно до ще одного варіанту використовують антитіло ІgМ. Відповідно до одного варіанту людські антитіла виробляються трансгенним ссавцем, відмінним від людини, наприклад, трансгенною мишою, здатною продукувати множинні ізотопи людських моноклональних антитіл до CD20 за рахунок рекомбінації V-D-J і переключення ізотипів. Відповідно, до аспектів цього винаходу входять не тільки антитіла, фрагменти антитіл і фармацевтичні композиції на їх основі, але й трансгенні ссавці, відмінні від людини, В-клітини, трансфектоми клітин-хазяїв і гібридоми, які продукують моноклональні антитіла. Такою трансгенною твариною, яка продукує поліклонапьні антитіла, як описано у заявці США US 2003/0017534, також може бути кріль. Відповідно, винахід також включає людські поліклональні антитіла, які специфічно зв'язуються з CD20. Відповідно до одного з варіантів винахід стосується полікпональних антитіл, які зв'язуються з епітопом на поверхні CD20, (і) що не містить або не вимагає амінокислотного залишка проліну у положенні 172; (іі) що не містить або не вимагає амінокислотних залишків аланіну у положенні 170 або проліну у положенні 172; (ііі) що містить або вимагає амінокислотні залишки аспарагіну у положенні 163 і аспарагіну у положенні 166; (iv) що не містить або не вимагає амінокислотного залишка проліну у положенні 172, але що містить або вимагає амінокислотні залишки аспарагіну у положенні 163 і аспарагіну у положенні 166; або (v) що не містить або не вимагає амінокислотного залишка аланіну у положенні 170 або проліну у положенні 172, але що містить або вимагає амінокислотні залишки аспарагіну у положенні 163 і аспарагіну у положенні 166. Відповідно до ще одного варіанту винаходу він стосується людських поліклональних антитіл, які мають одну або більше із наведених характеристик: (і) зв'язуються з мутантними P172S CD20 (пролін у положенні 172, мутований у серин) принаймні з тією самою спорідненістю, що і з людським CD20; (іі) зв'язуються з мутантним АхР (аланін у положенні 170, мутований у серин, а пролін у положенні 172 мутований у серин) принаймні з тією ж спорідненістю, що й до людського CD20; (ііі) виявляють знижений ступінь зв'язування, що становить 50% або вище, з мутантним N166D (аспарагін у положенні 166, мутований в аспарагінову кислоту) до людського CD20 при концентрації антитіла 10 мкг/мл; і/або (iv) виявляють знижений ступінь зв'язування, що становить 50% або більше, з мутованим N163D (аспарагін у положенні 163, мутований в аспарагінову кислоту) порівняно з людським CD20 при концентрації антитіла 10 мкг/мл. Відповідно до ще одного варіанту винахід стосується людських поліклональних антитіл, які зв'язуються з епітопом у першій малій зовнішньоклітиинній петлі людського CD20. Відповідно до ще одного варіанту винахід також стосується людських поліклональних антитіл, які зв'язуються з дискретним епітопом на CD20. Відповідно до ще од 93478 22 ного варіанту винахід стосується людських поліклональних антитіл, які зв'язуються з дискретним епітопом на CD20, який має частину першої малої зовнішньоклітинної петлі і частину другої зовнішньоклітинної петлі. Відповідно до ще одного варіанту винахід стосується людських поліклональних антитіл, які зв'язуються з дискретним епітопом на CD20, який має залишки AGIYAP першої малої зовнішньоклітинної петлі і зал MESLNFIRAHTPYI другої зовнішньоклітинної петлі. Також винахід включає способи застосування антитіл згідно з винаходом виявлення клітин, які експресують CD20. Також він стосуєтсья способів застосування антитіл згідно з винаходом для блокування або інгібування активності, індукованої СD20, наприклад, проліферативної і/або диференційної активності, які застосовуються в лікуванні розладів, пов'язаних з CD20, таких як пухлинорідні захворювання (наприклад клітинна лімфома) і аутоімунні захворювання (наприклад, ревматоїдний артрит, хвороба Крона і грануломатоз Вегенера). Для того, щоб суть цього винаходу була зрозумілою, спочатку слід дати визначення певним термінам. Додаткові визначення наведені у детальному описі. Терміни "CD20" і "антиген CD20" вживаються в цьому описі поперемінно і включають будь-які варіанти, ізоформи і різновиди, гомологічні людському CD20, які природним шляхом експресуютсья клітинами або на клітинах, трансфікованих геном СD20. Зв'язування антитіла згідно з винаходом з антигеном CD20 опосередковує умертвлення клітин, які експресують CD20 (наприклад, пухлинна клітина), за рахунок дезактивації СD20. Умертвлення клітин, які експресують CD20, може відбуватися через один або більш механізм: Комплементарна залежна цитотоксичність (CDC) клітин, які експресують CD20; Апоптоз клітин, які експресують CD20; Фагоцитоз клітини-ефектора у клітин, які експресують CD20; або залежна від антитіл клітинна цитотоксичність клітини-ефектора у клітин, які експресують CD20. Синоніми CD20, відповідно до галузі техніки, включають В-лімфоцитний антиген CD20, Влімфоцитний поверхневий антиген В1, Leu-16, Bp35, BM5 і LF5. В контексті цього опису поняття "інгібує ріст" (наприклад, щодо клітин) вживається на позначення будь-якого здатного бути визначеним зниження темпу росту клітин при їх взаємодії з антитілом проти CD20 порівняно з ростом таких самих клітин, які не перебувають у контакті з антитілом проти CD20, наприклад, інгібування росту клітинної культури на принаймні приблизно 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 99% або 100%. Такий спад у рості клітин може відбуватися через цілий ряд механізмів, наприклад, фагоцитоз клітин-ефекторів, ADCC, CDC, і/або апоптоз. Термін "маса" вживається на позначення збагачених сфінголіпідами і холестерином мембранних мікродоменів, розміщених у зовнішній зоні плазмової мембрани клітини. Здатність деяких білків об'єднуватися в межах таких доменів може позначатися на їх функціях. Наприклад, трансло 23 кація молекул CD20 у ліпідні rafts, після їх зв'язування людськими антитілами згідно з винаходом, утворює комплекси антитіл проти антигену CD20 з високою густиною в мембранах плазми. Така висока густина комплексів антитіл проти антигену CD20 може зробити можливою ефективну активацію комплементної системи під час комплементарної залежної цитотоксичності. Термін "антитіло" як він вживається у цьому винаході включає повні антитіла і будь-який антигенний зв'язуючий фрагмент (тобто, "частина, яка зв‘язується з антигеном ") або його окремий ланцюг. "Антитіло" вживається на позначення глікопротеїну, який містить принаймні два важких (Н) ланцюги і два легких (L) ланцюги, зв'язаних між собою дисульфідними зв'язками, або його частини, яка зв'язується з антигеном. Кожен важкий ланцюг містить змінну ділянку ланцюга (яка в опис позначена абревіатурою VH) і сталу ділянку ланцюга. Кожен легкий ланцюг містить змінну ділянку ланцюга (позначена абревіатурою VL) і сталу ділянку ланцюга. Ділянки VH і VL можуть далі підрозділятися на ділянки гіперперемінності, які називаються ділянки, що визначають комплементарність (CDR), між якими вкраплені ділянки, які є більш сталими, які називаються рамковими ділянками (FR). Кожна із VH і VL складається з трьох CDR і чотирьох FR, які мають наступний склад від амінокінця до карбокси-кінця: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Змінні ділянки важких і легких ланцюгів містять зв'язуючий домен, який взаємодіє з антигеном. Сталі ділянки антитіл можуть опосередковувати зв'язування імуноглобуліну з тканинами-хазяями або факторами, включно з різними клітинами імунної системи (наприклад, клітинамиефекторами) і першим компонентом (Clq) класичної комплементної системи. Термін "частина, яка зв'язується з антигеном" антитіла (або просто "частина антитіла ") в контексті цього опису вживається на позначення одного або більше фрагментів антитіла, який зберігає здатність специфічно зв'язуватись з антигеном (наприклад, CD20). Було показано, що функцію зв'язування антитіла з антигеном може виконувати фрагмент повного антитіла. Приклади зв'язувальних фрагментів, які охоплюються терміном "частина антитіла, яка зв'язується з антигеном " включають (і) фрагмент Fab, моновалентний фрагмент, який містить VL, VH, CL і Cн1 домени; (іі) фрагмент F(ab')2, двовалентний фрагмент, який містить два фрагменти Fab, зв'язані дисульфідним містком у ділянці з'єднання; (ііі) фрагмент Fd, який містить домени VH і СН1 (iv) фрагмент Fv, який містить VL і VH домени єдиного плеча антитіла, (v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), який містить домен VH; (vi) ізольовану ділянку, яка визначає комплементарність (CDR), і (vii) комбінацію двох або більше ізольованих CDR, до яких необов'язково може бути приєднаний синтезований лінкер. Крім того, хоча обидва домени фрагмента Fv, а саме VL і VH, кодуються двома окремими генами, вони можуть об'єднуватись з допомогою рекомбінантних методик, через синтезований лінкер, який робить можливим утворення ними єдиного протеїнового ланцюга, в якому ділянки VL і VH об'єдну 93478 24 ються, утворюючи моновалентні молекули (відомі як єдиний ланцюг Fv (scFv); див. наприклад, Bird et al. (1988) Science 242:423-426; і Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такі одноланцюгові антитіла також охоплюються терміном "частина антитіла, яка зв'язується з антигеном". Ще один приклад - це зв'язуючий домен, який представляє собою протеїни, злиті з імуноглобуліном, які містять (і) зв'язуючий поліпептидний домен, злитий з поліпептидом у ділянці поліпептидної петлі імуноглобуліну, (іі) сталу ділянку СН2 імуноглобулінового важкого ланцюга, злиту з ділянкою петлі, і (ііі) сталу ділянку СН3 імуноглобулінового важкого ланцюга, злиту зі сталою ділянкою СН2. Зв'язуючий поліпептидний домен може представляти собою змінну ділянку важкого ланцюга або змінну ділянку легкого ланцюга. Злиті з імнуглобуліном протеїни зв'язуючого домену додатково описані у заявках US 2003/0118592 і US 2003/0133939. Ці фрагменти антитіл одержують з допомогою відомих методик, відомих фахівцям у цій галузі, і скринінг на корисність таких антитіл здійснюють таким самим чином як і щодо перовинних антитіл. Термін "епітоп" означає протеїнову детермінанту, здатну специфічно зв'язуватися з антитілом. Епітопи зазвичай складаються з хімічно активних поверхневих групувань молекул, таких як амінокислоти або бічні ланцюги цукрози і зазвичай мають специфічні тривимірні структурні властивості, а також специфічні зарядні властивості. Конформаційні і некоформаційні епітопи відрізняються тим, що зв'язування з першим із згаданих втрачається у присутності денатурованих розчинників. Термін "дискретний епітоп" в контексті цього винаходу вживається на позначення конформаційного епітопу на протеїновому антигені, який утворений принаймні двома окремими ділянками у первинній послідовності протеїну. Термін "біспецифічна молекула " вживається на позначення будь-якого агента, наприклад, протеїну, пептиду, або протеїнового чи пептидного комплексу, який має дві зв'язувальних специфічності. Наприклад, може зв'язуватися або взаємодіяти з (а) поверхнево-клітинним антигеном (b) Fcрецептором на поверхні клітини-ефектора. Термін "мультиспецифічна молекула " або "гетероспецифічна молекула " вживається на позначення будьякого агента, наприклад, протеїну, пептиду, або протеїнового чи пептидного комплексу, який має більше ніж дві різні зв'язувальні специфічності. Наприклад така молекула може зв'язуватись або взаємодіяти з (а) поверхнево-клітинним антигеном, (b) Fc-рецептором на поверхні клітиниефектора і (с) принаймні одним іншим компонентом. Відповідно, винахід включає, але не обмежується цим переліком, біспецифічні, триспецифічні, тетраспецифічні та інші мультиспецифічні молекули, направлені на поверхнево-клітинні антигени, такі як CD20, та на інші мішені, такі як Fcрецептори на клітинах-ефекторах. Термін "біспецифічні антитіла" також включає діатіла. Діатіла є бівалентними, біспецифічними антитілами, в яких домени VH і VL експресуються 25 на одному поліпептидному ланцюгу, але при цьому бере участь лінкер, який є надто коротким, щоб зробити можливим спарювання двох доменів в одному ланцюгу, внаслідок чого домени вимушені паруватися з комплементарними доменами з іншого ланцюга, внаслідок чого утворюються два зв'язувальних сайти антигена (див., наприклад, Holliger, P., et al. (1993) Рrос. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R.J., et al. (1994) Structured 121-1123). Термін "похідні людського антитіла" вживається на позначення будь-якої модифікованої форми антитіла, наприклад, кон'югату антитіла та іншого агента або антитіла. У контексті цього винаходу людське антитіло є «похідним» від певної зародкової лінії, якщо таке антитіло одержують із системи, в якій використовують послідовності людського імуноглобуліну, наприклад, шляхом імунізації трансгенної миші, яка є носієм генів людського імуноглобуліну, або шляхом скринінгу бібліотеки генів людського імуноглобуліну, і в якому вибране людське антитіло принаймні на 90%, більш переважно на 95%, ще більш переважно на 96%, 97%, 98% або 99% ідентичне за амінокислотною послідовністю з амінокислотною послідовністю, яка кодується імуноглобуліновим геном зародкової лінії. Як правило, людське антитіло, похідне від певної послідовності людської зародкової лінії, має не більше, ніж 10 відмінностей в амінокислотах, більш переважно не більше, ніж 5 відмінностей, або навіть ще більш переважно не більше, ніж 4, 3, 2 або 1 амінокислотну відмінність від амінокислотної послідовності, яка кодується імуноглобуліновим геном зародкової лінії. У контексті цього винаходу термін «гетероантитіла» вживається на позначення двох або більше антитіл, їх похідних або ділянок, що зв'язуються з антигеном, з'єднаних одна з одною, принаймні дві з яких мають різні специфічності. Такі різні специфічності включають зв'язувальну специфічність щодо Fc-рецептора на клітині-ефекторі і зв'язувальну специфічність щодо антигену або епітопу на клітині-мішені, наприклад, пухлинній клітині. У контексті цього винаходу термін "людське антитіло" вживається на позначення антитіл, які мають змінні і сталі ділянки, похідні від імуноглобулінових послідовностей людської зародкової лінії. Людські антитіла згідно з винаходом можуть включати амінокислотні залишки, які не кодуються імуноглобуліновими послідовностями людської зародкової лінії (наприклад, мутації, які є результатом випадкового або специфічного щодо сайту мутагенезу in vitro або соматичної мутації in vivo). Однак, термін "людське антитіло " у контексті цього винаходу не включає антитіла, в яких послідовності CDR, які є похідними від зародкової лінії інших різновидів ссавців, таких як миша, були привиті на людські рамкові послідовності. Терміни "моноклональне антитіло" або "композиція моноклонального антитіла" в контексті цього винаходу вживаються на позначення препарату на основі молекул антитіла однієї молекулярної композиції. Композиція на основі моноклонального антитіла виявляє єдину зв'язувальну специфіч 93478 26 ність щодо і спорідненість з певним епітопом. Відповідно, термін "людське моноклональне антитіло" вживається на позначення антитіл, які виявляють єдину зв'язувальну специфічність, які мають змінні та сталі ділянки, що є похідними від імуноглобулінових послідовностей людської зародкової лінії. Відповідно до одного варіанту людські моноклональні антитіла продукуються гібридомою, яка включає В-клітину, отриману від трансгенного або трансхромосомного ссавця відмінного від людини, наприклад, від трансгенної миші, яка має геном, що містить трансген людського важкого ланцюга та трансген легкого ланцюга, злитого із законсервованою клітиною. Термін "рекомбінантне людське антитіло " у контексті цього винаходу включає всі людські антитіла, які одержані, експресуються, створюються або виділяються рекомбінантним способом, такі як (а) антитіла, виділені у ссавця (наприклад, миші), який є трансгенним або трансхромосомним щодо генів людського імуноглобуліну, або одержана з них гібридома (більш детально описана у нижченаведеному Розділі І), (b) антитіла, виділені з клітини-хазяїна, трансформованої для експресії такого антитіла, наприклад, з трансфектоми, (с) антитіла, виділені з бібліотеки рекомбінантних комбінаторних людських антитіл, і (d) антитіла, одержані, експресовані, утворені або виділені будь-яким іншим способом, який полягає у сплетенні послідовностей гену людського імуноглобуліну з іншими ДНК-послідовностями. Такі рекомбінантні людські антитіла мають змінні і сталі ділянки, які є похідними від імуноглобулінових послідовностей людської зародкової лінії. В деяких варіантах, проте, такі рекомбінантні людські антитіла можуть піддаватися мутагенезу in vitro (або, коли використовують тварину, трансгенну щодо людських Іg- послідовностей, соматичному мутагенезу in vivo), тож, відповідно, амінокислотні послідовності VH та VL ділянок рекомбінантних антитіл є послідовностями, які, незважаючи на те, що вони є похідними від і спорідненими з VH і VL послідовностями людської зародкової лінії, можуть природно не існувати і зародковій лінії людського антитіла in vivo. Термін "трансфектома" у контексті цього винаходу вживається на позначення рекомбінантних еукаріотичних клітин-хазяїв, які експресують антитіло, таких як СНО-клітини, клітини NS/0, клітини НЕК293, рослинні клітини або гриби, включно з клітинами дріжджів. У контексті цього винаходу "гетерологічне антитіло " визначається відносно трансгенного нелюдського організму, який виробляє таке антитіло. Цей термін вживається на позначення антитіла, яке має амінокислотну послідовність або кодуючу послідовність нуклеїнових кислот, яка відповідає послідовності, присутній в організмі, який не є трансгенним ссавцем, відмінним від людини, і загалом є організмом, відмінним від трансгенного ссавця, відмінного від людини. У контексті цього винаходу "гетерогібридне антитіло" означає антитіло, яке має легкий і важкий ланцюги, які походять від різних організмів. Наприклад, антитіло, яке має людський важкий 27 ланцюг, зв'язаний з мишачим легким ланцюгом, є гетеро гібридним антитілом. Приклади гетерогібридних антитіл включають химерні і олюднені антитіла, які описувалися вище. У контексті цього винаходу "виділене антитіло" вживається на позначення антитіла, яке є по суті вільним від інших антитіл, які мають різні антигенні специфічності (наприклад, виділене антитіло, яке специфічно зв'язується з CD20, є по суті вільним від антитіл, які специфічно зв'язуються з антигенами, відмінними від CD20). Виділене антитіло, яке специфічно зв'язується з епітопом, ізоформом або варіантом людського CD20, однак може виявляти перехресну реактивність щодо інших споріднених антигенів, наприклад, антигенів інших різновидностей (наприклад, гомологи різновиду CD20). Крім того, виділене антитіло може бути по суті вільним від іншого клітинного матеріалу і/або хімічних сполук. Відповідно до одного варіанту винаходу комбінація «виділених» моноклонапьних антитіл, які мають різні специфічності, об'єднані у чітко визначеній композиції. У контексті цього винаходу термін "специфічне зв'язування" вживається на позначення антитіла, яке зв'язується із заданим антигеном. Як правило, таке антитіло зв'язується із афінітетом, який відповідає КD з приблизно 1 х 10-7 М або менше, і зв'язується із заданим антигеном із афінітетом, який відповідає KD, який принаймні на два порядки нижчий, ніж його афінітет щодо зв‘язування із неспецифічним антигеном (наприклад, BSA, казеїн), відмінним від заданого антигену або близько спорідненого антигену. Вирази "антитіло, яке розпізнає антиген" і" антитіло, специфічне щодо антигену " вживаються як синонімічні вирази до виразу "антитіло, яке специфічно зв'язується з антигеном". У контексті цього винаходу термін ‗‘Kd" (sec-1) вживається на позначення константи швидкості дисоціації певної взаємодії антитіло-антиген. Згадана величина також позначається як величина кoff. Термін "ка" (М-1 х sec-1) у контексті цього винаходу вживається на позначення константи швидкості зв'язування для взаємодії певного антитіла і антигена. Термін "KD" (M) у контексті цього винаходу вживається на позначення константи рівноваги дисоціації для взаємодії певного антитіла і антигена. Термін "КА" (М-1) у контексті цього винаходу вживається на позначення константи рівноваги зв‘язування для взаємодії певного антитіла і антигена і одержується діленням ка на кd. У контексті цього винаходу "ізотип" вживається на позначення класу антитіл (наприклад, ІgМ або IgGI), які кодуються генами сталої ділянки важкого ланцюга. У контексті цього винаходу "переключення ізотипів " вживається на позначення феномена, який полягає у тому, що клас або ізотип антитіла змінюється з одного Іg-класу на інші Іg-класи. У контексті цього винаходу "непереключений ізотип" позначає ізотиповий клас важкого ланцюга, 93478 28 який одержується в результаті відсутності будьякого переключення ізотипів; ген СН, який кодує непереключений ізотип, як правило, є першим СНгеном, розташованим безпосередньо під функціонально перебудованого гену VDJ. Переключення ізотипів було класифіковане як класичне і некласичне переключення ізотипів. Класичне переключення ізотипів відбувається через рекомбінантні явища, з якими пов'язана принаймні одна ділянка послідовності трансгенц, яка бере участь у переключенні. Некласичне переключення може відбуватись, наприклад, шляхом гомологічної рекомбінації між людським і людським (5-пов'язана делеція). Альтернативні некласичні механізми переключення, такі як інтертрансгенна і/або інтерхромосомна рекомбінація, можуть мати місце і виконувати переключення ізотипів. У контексті цього винаходу термін "послідовність, яка бере участь у переключенні" вживається на позначення ДНК-послідовностей, які відповідають за рекомбінацію, з 5 допомогою якої відбувається переключення. "Світч-донорна " послідовність, як правило ділянка переключення , буде знаходитись у положенні 5' (тобто зверху) від ділянки-конструкта, яка видаляється під час рекомбінації. Ділянка "світч-акцептора" знаходиться між ділянкою конструкта, що видаляється, і замінною сталою ділянкою (наприклад, , ε, і т.д.). Оскільки специфічного сайту, де завжди відбувалась би рекомбінація, немає, то на основі конструкту, як правило, не можна передбачити, якою буде остаточна генна послідовність. У контексті цього винаходу "схема глікозиліювання" визначається як схема карбогідратних одиниць, які ковалентно приєднані до протеїну, зокрема до імуноглобулінового (антитіло) протеїну. Схему глікозиліювання гетерологічного антитіла можна охарактеризувати як по суті подібну до схеми глікозиліювання, яка виникає природним шляхом у антитіл, які продукуються різновидами трансгенних ссавців, відмінних від людини, тоді як фахівцям у цій галузі відомо, що схема глікозиліювання гетерологічного антитіла є більш подібною до схеми, яку виявляють види трансгенних ссавців, відмінних від людини, аніж ті види, від яких були одержані СН-гени трансгену. Термін "той, що зустрічається у природі" у контексті цього винаходу вживається на позначення того факту, що об'єкт може зустрічатися у природі. Наприклад, поліпептид або поліпептидна послідовність, присутня в організмі (включно з вірусами), яка може бути виділена із природного джерела, і яка не була навмисне модифікована людиною в лабораторних умовах є такою, що зустрічається у природі. Термін "перегрупований" у контексті цього винаходу вживається на позначення конфігурації важкого ланцюга або легкого ланцюга імуноглобулінового локусу, в якому V-сегмент розміщений безпосередньо по сусідству з D-J або J-сегментом у структурі, яка кодує по суті повний VH або, відповідно, VL домен. Перегрупований локус гену імуноглобуліну (антитіла) може бути ідентифікований шляхом порівняння із зародковою лінією ДНК, а 29 саме перегрупований локум має принаймні один рекомбінований гептамер/нонамерний гомологічний елемент. Термін "неперегрупований" або "конфігурація зародкової лінії " у контексті цього винаходу і щодо сегменту V вживається на позначення структури, в якій V-сегмент не є рекомбінованим, а отже розташованим безпосередньо по сусідству з сегментами D або J. Термін "молекула нуклеїнової кислоти " у контексті цього винаходу вживається на позначення молекул ДНК і молекул РНК. Молекула нуклеїнової кислоти може бути одноланцюговою або дволанцюговою, але переважно представляє собою дволанцюгову ДНК. Термін "виділена молекула нуклеїнової кислоти" у контексті цього винаходу щодо нуклеїнових кислот, які кодують повні антитіла або частини антитіл (наприклад, VH, VL, CDR3), які зв'язуються з CD20, вживається на позначення молекули нуклеїнової кислоти, в якій нуклеотидні послідовності, які кодують незаймане антитіло або частину антитіла, не містять інших нуклеотидних послідовностей, які кодують цілі антитіола або частини антитіл, які зв'язуються з антигенами, відмінними від CD20, і які можуть природним шляхом розміщуватися з обох боків нуклеїнової кислоти у людській геномній ДНК. Відповідно до одного варіанту людське антитіло проти CD20 включає нуклеотидну або амінокислотну послідовність 2F2, 7D8 або 11В8, а також змінні ділянки важкого ланцюга (VH) і легкого ланцюга (VL), які мають послідовності, представлені в in ІД. №ПОСЛІДОВНОСТІ: 1, 5 або 9, і, відповідно, ІД. № ПОСЛІДОВНОСТІ: 3, 7 або 11. Як описано у цьому винаходу, послідовності, представлені у ІД. № ПОСЛІДОВНОСТІ: 1-30 включають "консервативні модифікації послідовностей," тобто модифікації нуклеотидних і амінокислотних послідовностей, які значною мірою не позначаються на або не змінюють властивості зв'язування антитіла, яке кодується нуклеотидною послідовністю або яке містить амінокислотну послідовність. Такі консервативні модифікації послідовності включають нуклеотидні і амінокислотні заміщення, включення і делеції. Модифікації можуть бути запроваджені у ІД. № ПОСЛІДОВНОСТІ:1-30 з допомогою стандартних методик, відомих у цій галузі техніки, таких як сайт-спрямований мутагенез і опосередкований полімеразною ланцюговою реакцією (PCR) мутагенез. Консервативні амінокислотні заміщення включають заміщення, в яких амінокислотний залишок заміщений амінокислотним залишком із подібним бічним ланцюгом. Сімейства амінокислотних залашків з подібними бічними ланцюгами були описані в рівні техніки. Такі сімейства включають амінокислоти з основними бічним 5 ланцюгами (наприклад, лізин, аргінін, гістидин), кислотні бічні ланцюги (наприклад, аспарагінова кислота, глутамінова кислота), незаряджені полярні бічні ланцюги (наприклад, гліцин, аспарагін, глутамін, серин, треонін, тірозин, цистеїн, триптофан), неполярні бічні ланцюги (наприклад, аланін, валін, лейцин, ізолейцин, пролін, фенілаланін, метіонін), бета-розгалужені ланцюги (наприклад, треонін, валін, ізолейцин) і ароматичні 93478 30 бічні ланцюги (наприклад, тірозин, фенілаланін, триптофан, гістидин). Таким чином, наперед визначений несуттєвий амінокислотний залишок у людському антитілі проти CD20 переважно заміщується іншим амінокислотним залишком із того ж самого сімейства бічних ланцюгів. Цей винахід також включає "похідні" амінокислотних послідовностей, описаних в ІД. № ПОСЛІДОВНОСТІ: 1-30 і їх консервативні модифікації, в яких один або більше амінокислотний залишок був дериватизований, наприклад шляхом ацилювання або глікозилювання без значного впливу на зв'язувальні характеристики антитіла, яке містить амінокислотні послідовності. Крім того, цей винахід включає антитіла, в яких зміни були здійснені місце в ділянці Fc з метою зміни функціональних і фармакокінетичних властивостей антитіл. Такі зміни можуть зумовлювати підвищення або зниження зв'язування з C1q або з CD, FcR і ADCC. Заміщення можуть бути здійснені, наприклад в одному або більше амінокислотному залишку у положеннях 234, 235, 236, 237, 297, 318, 320 і 322 сталої ділянки важкого ланцюга, внаслідок чого відбувається зміна у функції ефектора, тоді як зберігається здатність зв‘язуватися з антигеном, порівняно з немодифікованим антитілом, порівн. US 5,624,821 і US 5,648,260. Період напіврозпаду антитіл in vivo також може бути покращений шляхом модифікації епітопа реутилізаційного рецептора сталого домену Іg або подібного до Іg сталого домену, внаслідок чого молекула не містить незайманий домен СН2 або незайману ділянку Fc домену Ig, порівн. US 6,121,022 і US 6,194,551. Крім того, період напіврозпаду може бути подовжений шляхом впровадження мутацій в ділянці Fc, наприклад шляхом заміщення треоніну лейцином у положенні 252, шляхом заміщення треоніну серином у положенні 254 або шляхом заміщення треоніну фенілаланіном у положенні 256, порівн. US 6,277,375. Крім того, схема глікозилювання для антитіл може бути модифікована з метою зміни ефекторної функції у антитіл. Наприклад, такі антитіла можуть експресуватися у трансфектомі, яка не додає фукозну одиницю, яка зазвичай приєднується до Asn у положенні 297 ділянки Fc з метою посилення спорідненості ділянки Fcз FcRIII, що, у свою чергу, призводить до підвищеної ADCC антитіл у присутності NK-клітин, порівн. Shield et al. (2002) JBC, 277:26733. Крім того, для зміни CDC може здійснюватись модифікація галактосилювання. Альтернативно, відповідно до ще одного варіанту, мутації можуть впроваджуватись довільно вздовж усієї або частини кодуючої послідовності антитіла проти CD20, наприклад, з допомогою насичувального мутагенезу, і одержані в результаті модифіковані антитіла проти CD20 можуть перевірятись на зв'язувальну активність. Відповідно, антитіла, які кодуються (змінною ділянкою важкого і легкого ланцюга) нуклеотидними послідовностями, описаними у цьому винаході і/або містять (змінну ділянку важкого і легкого ланцюга) описані в цій заявці амінокислотні послідовності (а саме ІД. № ПОСЛІДОВНОСТІ: 1-30), вклю 31 чають по суті подібні антитіла, які кодуються або які містять подібні послідовності, які були модифіковані консервативним шляхом. Більш детальний опис того, як такі по суті подібні антитіла можуть генеруватися на основі часткових (тобто змінних ділянок важкого і легкого ланцюгів) послідовностей, представлених в ІД. № ПОСЛІДОВНОСТІ: 130. Щодо нуклеїнових кислот, то термін "суттєва гомологічність" вживається на позначення двох нуклеїнових кислот або їх вказаних послідовностей, які при оптимальному розташуванні і порівнянні є ідентичними, маючи належні нуклеотидні вставки або делеції, принаймні у 80% нуклеотидів, зазвичай принаймні у приблизно від 90% до 95%, і більш переважно у принаймні приблизно від 98% до 99.5% нуклеотидів. Альтернативно суттєва гомологічність має місце, коли сегменти гібридизуються при селективних умовах гібридизації до комплементу ланцюга. Щодо нуклеотидних і амінокислотних послідовностей термін "гомологія" вказує на ступінь ідентичності між двома послідовностями нуклеїнових і амінокислот при їх оптимальному розташуванні і зіставленні відповідних вставок і делецій. Альтернативно суттєва гомологія має місце, коли сегменти ДНК гібридизуються в умовах селективної гібридизації до комплементу ланцюга. Відсоток ідентичності між двома послідовностями представляє собою функцію числа ідентичних позицій, які мають такі послідовності (тобто % гомології= # ідентичних позицій/загальне # позицій х 100), беручи до уваги число прогалин та довжину кожної такої прогалини, які мають впроваджуватися для оптимального розташування двох послідовностей. Порівняння послідовностей та визначення відсотка ідентичності між двома послідовностями може бути здійснене з використанням математичного алгоритму, що описано у наведених далі прикладах, які не носять обмежувального характеру. Відсоток ідентичності між двома нуклеотидними послідовностями може бути визначений з використанням програми GAP у пакеті програмного забезпечення GCG (доступне на сайті http://www.qcq.com). з використанням матриці NWSgapdna.CMP і прогалину масою 40, 50, 60, 70 або 80 і масою по довжині 1, 2, 3, 4, 5 або 6. Відсоток ідентичності між двома нуклеотидними і амінокислотними послідовностями також може визначатися з використанням алгоритму Е. Меєрса і У. Міллера (Comput. Appl. Biosci., 4:11-17 (1988)), який був включений у програму ALIGN (версія 2.0), використовуючи таблицю маси залишків РАМ120, похибку довжини прогалини 12 і похибку прогалини 4. Крім того, відсоток ідентичності між двома амінокислотними послідовностями може бути визначений з допомогою алгоритму Нідлмана і Вунша (J. Моl. ВіоІ. 48:444-453 (1970)), який був включений до програми GAP у пакеті програмного забезпечення GCG (доступне на сайті http://www.gcg.corn), використовуючи матрицю Blossum 62 або РАМ250, і прогалину масою 16,14,12,10, 8, 6 або 4 і масою по довжині 1, 2, 3, 4, 5 або 6. 93478 32 Послідовності нуклеїнових кислот і протеїнів згідно з винаходом можуть також використовуватись як "послідовність запиту " для проведення пошуку у відкритих для громадськості базах даних, наприклад з метою ідентифікації споріднених послідовностей. Такі пошуки можуть проводитись з допомогою програм NBLAST і XBLAST (версія 2.0 згідно Altschul, et al. (1990) J. Моl. ВіоІ. 215:403-10. Нуклеотидні пошуки BLAST можуть виконуватись з допомогою програми NBLAST, рахунок = 100, довжина запиту = 12 для того, щоб отримати нуклеотидні послідовності, гомологічні з молекулами нуклеїнових кислот згідно з винаходом. Протеїнові пошуки BLAST можуть виконуватись з допомогою програми XBLAST, рахунок = 50, довжина запиту = 3, щоб одержати амінокислотні послідовності, гомологічні з протеїновими молекулами згідно з винаходом. З метою одержання вивірки прогалин з метою зіставлення може використовуватись Gapped BLAST як описано в Altschul et al., (1997) Nucleic Acids Res. 25(17):3389-3402. При використанні програм BLAST і Gapped ВLAST можуть використовуватись вихідні установки відповідних програм (наприклад, XBLAST I NBLAST). Див. http://www.ncbi.nlm.nih.gov. Нуклеїнові кислоти можуть бути присутні у цілих клітинах, у клітинних лізатах або у частково очищеній чи по суті чистій формі. Нуклеїнова кислота є «виділеною» або «по суті очищеною», коли вона є відділеною від інших клітинних компонентів або інших домішок наприклад, інших клітинних нуклеїнових кислот або протеїнів з допомогою стандартних методик, включно з обробкою лугом/додецилсульфатом натрію, CsCI бандінгом, колонковою хроматографією, електрофорезом з агарозовим гелем та іншими добре відомими у цій галузі методиками. Див., F. Ausubel, et al., ed. Current Protocols in Molecular Biology (Сучасні протоколи у молекулярній біології), Greene Publishing і Wiley Interscience New York (1987). Композиції нуклеїнових кислот згідно з винаходом, які часто отримують із рідної послідовності (за винятком модифікованих сайтів рестрикції та под.), як з кДНК, геномної послідовності або їх суміші, може піддаватись мутації відповідно до стандартних методик одержання генних послідовностей. У випадку кодуючих послідовностей такі мутації можуть при бажанні позначатися на амінокислотних послідовностях. Зокрема, ДНКпослідовності, які є по суті гомологічними з або похідними від рідних V, D, J, констант, перемикачів та інших подібних послідовностей, описаних у цій заявці, входять в об'єм правової охорони винаходу (де "похідні" іозначає, що послідовність є ідентичною абомодифікованою з іншої послідовності). Нуклеїнова кислота "операбельно зв'язана", якщо вона розміщена у функціональному зв'язку зі іншою послідовністю нуклеїнової кислоти. Наприклад, промотор або енхансер операбельно зв'язаний з кодуючою послідовністю, якщо він впливає на транскрипцію такої послідовності. Щодо транскрипції регуляторних послідовностей, операбельно зв'язана означає, що ДНК-послідовності, будучи зв'язаними, є суміжними і у випадку, коли необхідно приєднати дві кодуючі ділянки протеїнів, вони є 33 суміжними і розміщеними у зчитувальній рамці. Щодо послідовностей-перемикачів, операбельно зв'язаний означає, що послідовності здатні впливати на перемикальну рекомбінацію. Термін "вектор" у контексті цього винаходу вживається на позначення молекули нуклеїнової кислоти, здатної транспортувати іншу нуклеїнову кислоту, з якою вона була зв'язана. Одним типом вектора є «плазміда», яка вживається на позначення кругової дволанцюгової ДНК-петлі, на яку можуть накладатися додаткові сегменти ДНК. Ще один різновид вектора - це вірусний вектор, в якому додаткові сегменти ДНК можуть накладатися на вірусний геном. Деякі вектори мають здатність до автономної реплікації у клітині-хазяїні, в яку вони вживлені (наприклад, бактеріальні вектори, які мають бактеріальну природу реплікації, і епісомнів вектори ссавців). Інші вектори (наприклад, неепісомні ветори ссавців) можуть бути інтегровані у геном клітини-хазяїна при їх вживленні у літинухазяїн, і відтак реплікованими уздовж геномахазяїна. Крім того, деякі вектори здатні направляти експресію генів, з якими вони оперативно зв'язані. Такі вектори у тексті цього опису згадуються як "рекомбінантні вектори експресії" (або просто "вектори експресії "). Загалом вектори експресії, які використовуються в рекомбінантних ДНКметодиках, часто представляють собою плазміди. У тексті цього опису терміни "плазміда" і "вектор" можуть вживатися як синоніми, оскільки плазміда є найбільш часто вживаною формою вектора. Проте, винахід включає інші форми векторів експресії, такі як вірусні вектори (наприклад, реплікаційні дефективні ретровіруси, аденовіруси і аденопов'язані віруси), які виконують еквіваленті функції. Термін "рекомбінантна клітина-хазяїн " (або просто "клітина-хазяїн ") у контексті цього винаходу вживається на позначення клітини, в яку був вживлений рекомбінантний вектор експресії. Слід мати на увазі, що такі терміни вживаються на позначення не тільки певної клітини, але й потомства такої клітини. Оскільки у наступних поколіннях можуть 5 мати місце певні модифікації внаслідок мутації або впливу навколишнього середовища, таке потомство може фактично не бути ідентичним материнській клітині, але все ж входять в об'єм поняття «клітина-хазяїн» у контексті цього винаходу. Рекомбінантні клітини-хазяї включають, наприклад, трансфектоми, такі як СНО-клітини, NS/0клітини і лімфоцитарні клітини. У контексті цього винаходу термін "суб'єкт" включає будь-яку людину або ссавця, відмінного від людини. Термін "ссавець, відмінний від людини " включає всіх хребетних, наприклад, ссавців і відмінних від людини ссавців, таких як нелюдиноподібні примати, вівці, собаки, корови, кури, земноводні, рептилії і т.д. Термін "трансгенна відмінна від людини тварина" вживається на позначення тварини, відмінної від людини, яка має геном, який включає один або більше людських трансгенів або трансхромосом з важкими і/або легкими ланцюгами (або інтегрованих, або не інтегрованиху природну геномну ДНК тварини) і здатний експресувати повністю 93478 34 людські антитіла. Наприклад, трансгенна миша може мати людський трансген з легким ланцюгом та або людський трансген з важким ланцюгом, або трансхромосому з важким ланцюгом, завдяки чому така миша виробляє людські антитіла проти CD20 за умови її імунізації антигеном CD20 і/або клітинами, які експресують CD20. Людський трансген з важким ланцюгом може бути інтегрований у хромосомну ДНК мині, як у випадку з трансгенною твариною, наприклад, мишею HuMAb, такою як НСо7 або НСо12 миша, або ж людський трансген з важким ланцюгом може утримуватися поза хромосомами, як у випадку з трансхромосомними (наприклад, KM) мишами, описаними в заявці WO 02/43478. Такі трансгенні і хромосомні миші здатні продукувати множинні ізотипи людських моноклональних антитіл проти CD20 (наприклад, IgG, IgA і/або IgE) за рахунок рекомбінації V-D-J і переключення ізотипів. Різні аспекти цього винаходу описані у наведених далі розділах. І. Продукування людських антитіл проти CD20 Людські моноклональні антитіла згідно з винаходом можуть продукуватися з допомогою ряду методик, включно з традиційною методикою для стандартних моноклональних антитіл, наприклад, стандартної методики гібридизації соматичних клітин за Колером і Мілштейном (Kohler і Milstein, Nature 256: 495 (1975)). Хоча перевага віддається методикам гібридизації соматичних клітин, в принципі, можуть використовуватися інші методики продукування моноклональних антитіл, наприклад, методики вірусної або онкогенної трансформації В-лімфоцитів або методик відображення фагів з використанням бібліотек генів людських антитіл. Переважною тваринною системою для одержання гібридом, які секретують людські моноклональні антитіла, є система мишей. Продукування гібридом у миші є досить добре розробленою процедурою. Протоколи імунізації та методик виділення імунізованих спленоцитів для злиття віжомі з рівня техніки. Партнери по злиттю (наприклад, клітини мишачої мієломи) та процедури злиття також відомі. Відповідно до переважного варіанту людські моноклональні антитіла проти CD20 можуть генеруватися з використанням частин трансгенних і трансхромосомних мишей, які є носіями людської імунної системи, а не мишачої системи. Такі трансгенні і трансхромосомні миші включають мишей, які у цьому описі згадуються як миші HuMAb і, відповідно, миші KM, і разом називаються «трансгенні миші». Миша HuMAb містить мінілокуси гену людського імнуглобуліну, який кодує неперегруповану людську важку ( і ) і к легку імуноглобулінову послідовність разом з мутаціями-мішенями, які дезактивують ендогенні докуси і к ланцюга (Lonberg, N. et al. 25 (1994) Nature 368 (6474): 856859). Відповідно, миші проявляють знижену експресію мишачого ІgМ або к, і у відповідь на імунізацію вживлені людські трансгени з важкими і легкими ланцюгами зазначають класичного переключення і соматичної мутації, генеруючи високо споріднені людські IgG к моноклональні 35 антитіла (Lonberg, N. et al. (1994), supra; reviewed in Lonberg, N. (1994) Handbook of Experimental Pharmacology (Підручник експериментальної фармакології) 113:49-101; Lonberg, N. і Huszar, D. (1995) Intern. Rev. Immunol. Vol. 13: 65-93, і Harding, F. і Lonberg, N. (1995) Ann. N.Y. Acad. Sci 764: 536-546). Виведення миші HuMAb детально описане у Taylor, L. et al. (1992) Nucleic Acids Research 20:6287-6295; Chen, J. et al (1993) International Immunology 5: 647-656; Tuaillon et al. (1994) J. Immunol. 152:2912-2920; Lonberg et al., (1994) Nature 368(6474): 856-859; Lonberg, N. (1994) Handbook of Experimental Pharmacology 113:49-101; Taylor, L. et al. (1994) International Immunology 6: 579-591; Lonberg, N. і Huszar, D. (1995) Intern. Rev. Immunol. Vol. 13:65-93; Harding, F. і Lonberg, N. (1995) Ann. N.Y. Acad. Sci 764:536546; Fishwild, D. et al. (1996) Nature Biotechnology 14:845-851. Див. Також патенти США № 5,545,806; 5,569,825; 5,625,126; 5,633,425; 5,789,650; 5,877,397; 5,661,016; 5,814,318; 5,874,299 і 5,770,429 (у всіх цих патентах власником є Lonberg і Кау), а також патент США 5,545,807, виданий Surani та ін.; WO 98/24884, WO 94/25585, WO 93/1227, WO 92/22645, WO 92/03918 і WO 01/09187. KM миша має людську трансхромосому з важким ланцюгом і людський трансген з легким каппа ланцюгом. Гени ендогенної миші з важким і легким ланцюгом також були порушені у мишей KM, внаслідок чого їх імунізація призводить до продукування людських імуноглобулінів, а не мишачих імуноглобулінів. Конструювання мишей КМ та їх застосування для вирощування людських імуноглобулінів детально описане у заявці WO 02/43478. Імунізація Для генерування повністю людських моноклональних антитіл проти CD20, трансгенні або трансхромосомні миші, які мають гени людського імуноглобуліну (наприклад, миші НСо12, НСо7 або KM) можуть бути імунізовані препаратом, збагаченим антигеном CD20 і/або клітинами, які експресують CD20, як описано, наприклад, у Lonberg et al. (1994), див. вище; Fishwild et al. (1996), див. вище і WO 98/24884. Альтернативно миші можуть бути імунізовані ДНК, яка кодує людський CD20. Переважно при першому вприскуванні використовують мишей віком 6-12 тижнів. Наприклад, для внутрішньочеревинної імнузації мишей HuMAb може використовуватись препарат, збагачений антигеном CD20 (5-50 мкг). У випадку, якщо імунізації, в яких використовують очищений або збагачений препарат на основі антигену CD20, не зумовлюють вироблення антитіл, миші також можуть імунізуватися клітинами, які експресують CD20, наприклад, клітинною лінією для стимулювання імунної реакції. Накопичений досвід з різними антигенами показав, що трансгенні миші HuMAb найкраще реагують, коли перша імунізація здійснюється внутрішньочеревинним (IP) або підшкірним шляхом (SC) і призначаються клітини, які експресують CD20 у повному ад'юванті Фройнда, після чого щотижня здійснюють внутрішньочеревинні імунізації (до 10 раз) з призначенням клітин, які експресують CD20 93478 36 у фосфатно-соляному буферному розчині. Імунна реакція може контролюватися упродовж протоколювання імунізації, при якій зразки плазми одержують з допомогою ретроорбітальних відсмоктувачів. Плазма може сортуватись з допомогою активованого флюоресцентного аналізатора клітин (FACS) (як описано далі), і миші з достатніми титрами людського імуноглобуліну проти CD20 можуть використовуватись для злиття. Миші можуть додатково одержувати клітини, які експресують CD20, за 3 дні до їх умертвлення та видалення селезінки. Генерування гібридом, які виробляють людські моноклональні антитіла проти CD20 Для генерування гібридом, які виробляють людські моноклональні антитіла проти людського CD20 спленоцити і клітини лімфатичних вузлів від імунізованих мишей можуть ізолюватися і зливатися з відповідною законсервованою клітинною лінією, такою як клітинна лінія мишачої мієломи. Одержані гібридоми можуть аналізуватись на вироблення антиген-специфічних антитіл. Наприклад, одноклітинні суспензії селезінчатих лімфоцитів від імунізованих мишей можуть зливатися з несекреторними клітинами мієломи миші SP2/0Ag8.653 (ATCC, CRL 1580) з 50% PEG (мас/об). Клітини можуть висіватися приблизно у кількості 1 х 105 на лунку на плоскодонному мікротитрувальному планшеті з наступним інкубуванням протягом двох тижнів у селективному середовищі, яке, окрім звичайних реагентів, містить 10% сироватки зародкового клона, 5-10% клонуючого фактора гібридоми (IGEN) і 1Х HAT (Sigma). Приблизно через два тижні клітини можуть культивуватися у середовищі, в якому HAT заміщений на НТ. Окремі лунки можуть аналізватися з допомогою імуноферментного твердофазного аналізу ELISA для людських каппа-легких ланцюгів, що містять антитіла, і з допомогою аналізу FACS з використанням клітин, які експресують CD20 для специфічності CD20. При досягненні інтенсивного росту гібридоми середовище може аналізуватися через 10-14 днів. Гібридоми, які секретують антитіла, можуть бути перепосіяні, ще раз вивчені, і якщо вони все ще є позитивними щодо людського IgG, моноклональні антитіла проти CD20 можуть субклонуватись принаймні двічі, обмежуючи при цьому розведення. Після цього стійкі субклони можуть культивуватись in vitro для генерування антитіл у середовищі культури тканини для зняття характеристик. Генерування трансфектом, які виробляють людські моноклональні антитіла проти CD20 Людські антитіла зігдно з винаходом також можуть вироблятися трансфектомою клітинихазяїна з допомогою, наприклад, комбінації технологій рекомбінантних ДНК і методик трансфекції генів, які добре відомі в цій галузі техніки (Morrison, S. (1985) Science 229:1202). Наприклад, відповідно до одного варіанту гени, які представляють інтерес, наприклад, гени людських антитіл, можуть замикатися у вектор експресії, такий як еукаріотична плазміда експресії, така як та, що використовується системою експресії генів GS, описаною в заявці WO 87/04462, 37 WO 89/01036 і ЕР 338 841, або іншою системою експресії, відомою в цій галузі техніки. Очищена плазміда з клонованими генами антитіла може вводитись в еукаріотичні клітини-хазяї, такі як клітини СНО, клітини NS/0 або клітини НЕК293, або, як альтернатива, інші еукаріотичні клітини, такі як клітини рослинного походження, клітини грибів або дріжджів. Метод, з допомогою якого вводять такі гени, може бути одним із методів, описаних в цій галузі техніки, таким як електропорація, ліпофектин, ліпофектамін або інші. Після введення цих генів антитіл у клітини-хазяї, клітини, які експресують таке антитіло, можуть бути ідентифіковані і відібрані. Ці клітини представляють собою трансфектоми, які після цього можуть бути ампліфіковані до рівня експресії і удосконалені для продукування антитіл. Рекомбінантні антитіла можуть бути виділені і очищені від таких супернатантів і/або клітин. Додаткові рекомбінантні засоби для продукування людських моноклональних антитіл проти CD20 Альтернативно клоновані гени антитіл можуть експресуватися в інших системах експресії, включно з прокаріотичними клітинами, такими як мікроорганізми, такі як Е. соlі для продукування одноланцюгових Fv-антитіл, водорості, а також клітини комах. Крім того, такі антитіла можуть вироблятися у трансгенних тварин, відмінних від людини, як, наприклад, в овечому молоці і кролиному молоці або в курячих яйцях, або у трансгенних рослин. Див., наприклад Verma, R., et al. (1998). Antibody engineering: Comparison of bacterial, yeast, insect і mammalian expression systems (Одержання антитіл методами генної інженерії: Порівняння бактеріальної, дріжджевої систем експресій та системи експресій комах і ссавців). J.lmmunol.Meth. 216:165-181; Pollock, et al. (1999). Transgenic milk as a method for the production of recombinant антитіла. (Трансгенне молоко як засіб одержання рекомбінантних антитіл) J.lmmunol.Meth. 231:147157; та Fischer, R., et al. (1999). Molecular farming of recombinant антитіла in plants (Вирощування рекомбінантних антитіл у рослин на молекулярному рівні). Biol.Chem. 380:825-839. Застосування часткових послідовностей антитіл для експресії цілих антитіл Антитіла взаємодіють з антигенами-мішенями, головним чином, через амінокислотні залишки, які розміщені у шести ділянках, що визначають комплементарність важких і легких ланцюгів (CDRs). З цієї причини амінокислотні послідовності в межах таких CDR-ділянок є більш різнотиповими між індивідуальними антитілами, ніж послідовності поза такими CDR-ділянками. Оскільки CDRпослідовності відповідають за більшість взаємодій антитіло-антиген, можна експресувати рекомбінантні антитіла, які повторюють властивості специфічних природних антитіл за рахунок конструювання веторів експресії, які включають CDRпослідовності із специфічних природних антитіл, привитих на рамкові послідовності від різних антитіл з різними властивостями (див, наприклад, Riechmann, L. et. al. (1998) Nature 332:323-327; Jones, P. et al. (1986) Nature 321:522-525; і Queen, 93478 38 С et al. (1989) Proc. Natl. Acad. Sci.. U.S.A. 86:10029-10033). Такі рамкові послідовності можуть бути отримані із громадських баз ДНК, які включають зародкові генні послідовності антитіла. Такі зародкові послідовності відрізняються від зрілих генних послідовностей ; антитіл, тому що вони не включають повністю укомплектовані змінні гени, які утворюються 5 приєднанням V(D)J під час дозрівання В-клітини. Зародкові генні послідовності також відрізняються від послідовностей антитіла із високоспорідненим вторинним репертуаром у змінній ділянці. Наприклад, соматичні мутації досить нечасто зустрічаються в аміно-термінальній частині рамкової ділянки 1 і в карбокситермінальній ділянці 4. Крім того, багато соматичних мутацій суттєво не змінюють зв'язувальних властивостей антитіла. З цієї причини немає необхідності одержувати повну послідовність ДНК певного антитіла для того, щоб відтворити ціле рекомбінантне антитіло із властивостями зв'язування, подібними до первинного антитіла (див. WO 99/45962). Часткова послідовність важкого і легкого ланцюга, яка поширена у CDR-ділянках, є зазвичай достатньою з такою метою. Таку часткову послідовність використовують для визначення, яка змінна ділянка зародкової лінії і сегменти приєднаного гену визначили змінні гени рекомбінованого антитіла. Послідовність зародкової лінії після цього використовують для заповнення прогалин у змінних ділянках. Послідовності-лідери важкого і легкого ланцюга розколюються під час визрівання протеїну і не визначають властивостей кінцевого антитіла. З метою додавання пропущених послідовностей клоновані послідовності кДНК можуть поєднуватися із синтетичними олігонуклеотидами через механізм лігування або ПЛР-ампліфікації. Альтернативно вся змінна ділянка може бути синтезована у вигляді набору коротких накладних олігонуклеотидів і об'єднана ПЛР-ампліфікацією, утворюючи повністю синтетичний клон змінної ділянки. Цей процес має певні переваги, такі як видалення або включення відповідних сайтів рестрикції або оптимізація певних кодонів. Нуклеотидні послідовності транскриптів важкого і легкого ланцюга із гібридом використовують для конструювання групи накладних олігонуклеотидів для створення синтетичних Vпослідовностей з ідентичними амінокислотними кодуючими властивостями до природних послідовностей. Синтетичні послідовності важкого і каппа ланцюга можуть відрізнятися від природних послідовностей трьома показниками: ланцюги повторюваних нуклеотидних основ перериваються для спрощення синтезу олігонуклеотидів і ПЛРампліфікацію; включені оптимальні сайти ініціації трансляції відповідно до правил Козака (Kozak (1991) J. Biol. Chem. 266L19867019870); і сайти Hindlll сконструйовані над сайтами ініціації трансляції. Для змінних ділянок як важкого, так і легкого ланцюга, оптиізовані кодуючі і відповідні некодуючі ниткові послідовності розбиваються на 30 - 50 нуклеотидів приблизно посередині відповідного некодуючого олігонуклеотиду. Таким чином, для кожного ланцюга олігонуклеоити можуть згруповуватися 39 у накладні двониткові структури які обплатають сегменти 150 - 400 нуклеотидів. Після цього такі фонди використовують як моделі для одержання продуктів ПЛР-ампліфікації 150 - 400 нуклеотидів. Зазвичай одну змінну ділянку групи олігонуклеотидів розбивають на два фонди, які окремо ампліфікують для одержання двох накладних ПЛРпродуктів. Після цього такі накладні продукти об'єднують ПЛР-ампліфікацією з одержанням повної змінної ділянки. Також може бути бажано включити накладний фрагмент сталої ділянки важкого або легкого ланцюга (включно з сайтом Bbsl каппалегкого ланцюга або сайт Agel у гамма важкому ланцюгу) в ПЛР-ампліфікації для генерування фрагментів, які можуть легко клонуватися в конструкти вектора експресії. Реконструйовані змінні ділянки важкого і легкого ланцюга після цього об'єднували з клонованим промотором, лідерною послідовністю, послідовністю ініціації трансляції, сталою ділянкою, 3' нетрансльованою послідовністю, послідовністю поліадениліції і послідовністю термінування транскрипції з одержанням конструктів векторів експресії. Конструкти експресії важкого і легкого ланцюга можуть бути об'єднані у спільний вектор, котрансфіковані, серійно трансфіковані або окремо трансфіковані у клітини-хазяїна, які після того зливаються з утворенням клітини-хазяїна, яка експресує обидва ланцюги. Плазміди, які використовуються у конструюванні векторів експресії для людського IgGk описані далі. Такі плазміни сконструйовані таким чином, щоб ПЛР-ампліфіковані кДНК послідовності V важкого і V каппа легкого ланцюга могли використовуватися для реконструювання повних мінігенів важкого і легкого ланцюга. Такі плазміди можуть використовуватися для експресування повністю людського або химерного IgG1,k або lgG4,k антитіла. Подібні плазміди можуть конструюватися для експресії інших ізотипів важкого ланцюга або для експресії антитіла, яке містить лямбда легкі ланцюги. Таким чином, у ще одному аспекті винаходу структурні риси людських антитіла проти CD20, наприклад, 11В8, 2F2 або 7D8, використовуються для створення структурно 5 споріднених людських антитіл проти CD20, які зберігають принаймні одну функціональну властивість антитіла згідно з винаходом, таку як зв'язування з CD20. Більш точно, одна або більше CDR-ділянки антитіла 2F2, 7D8 або 11В8 може рекомбінантно об'єднуватися з відомими людськими рамковими ділянками і CDRs, утворюючи додаткові сконструйовані рекомбінацією людські антитіла проти CD20 згідно з винаходом. Відповідно, в інщому варінаті винахід стосується способу одержання людського антитіла проти CD20, який полягає у тому, що: одержують антитіло, яке містить (1) рамкові ділянки людського важкого ланцюга і CDRs людського важкого ланцюга, де принаймні одна із CDRділянок людського важкого ланцюга включає амінокислотну послідовність, вибрану з амінокислотнх послідовностей CDRs, показаних на Фігурах 53, 55 або 57 (або відповідних амінокислотних за 93478 40 лишків в ІД. № ПОСЛІДОВНОСТЕЙ:13-15, 19-21 або 25-27); і (2) рамкові ділянки людського легкого ланцюга і CDRs людського легкого ланцюга, де принаймні одна із CDR-ділянок людського легкого ланцюга включає амінокислотну послідовність, вибрану з амінокислотних послідовностей CDRs, показаних на Фігурах 53, 55 або 57 (або відповідних амінокислотних залишків в ІД. № ПОСЛІДОВНОСТЕЙ: 16-18, 22-24, or 28-30); де антитіло зберігає здатність зв'язуватись з CD20. Здатність антитіла зв'язуватись з CD20 може визначатися з допомогою стандартних аналізів на зв'язування, таких як описані у Прикладах (наприклад, FACS аналіз). Оскільки добре відомо, що CDR3 домени важкого і легкого ланцюгів антитіла відіграють особливо важливу роль у зв'язувальній специфічності/спорідненості антитіла щодо антигену, рекомбінантні антитіла згідно з винаходом, одержані вищеописаним способом, переважно містять домени CDR3 важкого і легкого ланцюга антитіл 2F2, 7D8 або 11В8. Такі антитіла додатково можуть включати домени CDR2 антитіл 2F2, 7D8 або 11В8. Такі антитіла The антитіла додатково можуть включати домени CDR1 антитіл 2F2, 7D8 або 11В8. Відповідно, цей винахід додатково стосується антитіл проти CD20, які місттять: (1) рамкові ділянки людського важкого ланцюга, CDR1-ділянку людського важкого ланцюга, СDR2-ділянку людського важкого ланцюга і СDR3-ділянку людського важкого ланцюга, де CDR3-ділянка людського важкого ланцюга відповідає CDR3 антитіл 2F2, 7D8 або 11В8 як показано на Фігурах 53, 55 або 57 (або відповідним амінокислотним залишкам, показаним в ІД. № ПОСЛІДОВНОСТЕЙ: 15, 21 або 27); і (2) рамкові ділянки людського легкого ланцюга, CDR1-ділянку людського легкого ланцюга, СDR2ділянку людського легкого ланцюга і CDR3- ділянку людського легкого ланцюга, де СDR3-ділянка людського легкого ланцюга відповідає CDR3 антитіл 2F2, 7D8 або 11В8 як показано на Фігурах 53, 55 або 57 (або відповідним амінокислотним залишкам, показаним в ІД. № ПОСЛІДОВНОСТЕЙ:18, 24 або 30), де таке антитіло зв'язується з CD20. Таке антитіло може додатково включати CDR2 важкого ланцюга і/або CDR2 легкого ланцюга антитіла 2F2, 7D8 або 11В8. Таке антитіло може додатково включати CDR1 важкого ланцюга і/або CDR1 легкого ланцюга антитіла 2F2, 7D8 або 11В8. Переважно CDR1, 2 і/або 3 вищеописаного одержаного методами генної інженерії антитіла включають ідентичні амінокислотні послідовності, які відповідають послідовностям 2F2, 7D8 або 11В8, описаним у цій заявці. Однак, для середнього фахівця у цій галузі очевидно, що можливі деякі відхилення від точного відтворення CDRпослідовностей антитіл 2F2, 7D8 або 11В8, при яких все ще зберігається здатність антитіла ефективно зв'язуватися з CD20 (наприклад, консервативні заміщення). Відповідно, згідно із ще одним варіантом сконструйовані антитіла можуть складатися з однієї або більше ділянок CDR, які, наприклад, на 90%, 95%, 98% або 99.5% ідентичні з од 41 нією або більше ділянками CDR антитіл 2F2, 7D8 або 11В8. Окрім простого зв'язування з CD20, сконструйовані антитіла, такі як описані вище, можуть бути вибрані на підставі наявності у них інших функціональних властивостей антитіл згідно з винаходом, таких як: (1) низький ступінь дисоціації від CD20; (2) високий рівень спорідненості щодо зв'язування з CD20; (3) зв'язування з унікальним епітопом на CD20 і/або зв'язування у певній орієнтації відносно CD20, і/або зв'язування із спеціальною формою CD20; (4) опосередкування високого рівня CDC або на СD55/59-негативних, або на СD55/59позитивних клітинах; (5) траслокація на ліпідні маси під час зв'язування з CD20; (6) інгібування росту клітин, які експресують CD20; (7) індукування апоптозу клітин, які експресують CD20; (8) індукування гомотипової адгезії клітин, які експресують CD20; (9) продовження життя суб'єкта, який має пухлинні клітини, що екаспресують CD20; (10) опосередкування ADCC СD20-мішеней при змішуванні з відповідними клітинамиефекторами; (11) здатність зменшувати кількість клітин, які експресують CD20; і/або (12) здатність зменшувати кількість клітин, які експресують низький рівень CD20 (клітин CD20low). Характеристики зв'язування людських моноклональних антитіл з CD20 Для очищення людських антитіл проти CD20 вибрані гібридоми можуть вирощуватись у дволітрових обертальних пробірках для очищення моноклональних антитіл. Супернатанти можуть відфільтровуватись і концентруватись перед афінною хроматографією з протеїном А-сефароза (для антитіл ізотипу lgG1) (Pharmacia, Piscataway, NJ) або з проти-людською покритою IgG сефарозоб або протеїно G-сефарозою у випадку з антитілами ізотипу lgG3. Елюйовані IgG можуть аналізуватись гель-електрофорезом та ВЕРХ з метою забезпечення чистоти. Буферний розчин може бути замінений на фосфатно-сольовий буферний розчин, і його концентрація може визначатись з допомогою OD280 з коефіцієнтом поглинання 1.43. Моноклонапьні антитіла можуть ділитися на аліквоти і зберігатись при температурі -80 °С. Для встановлення, чи зв'язуються селективні людські антитіла проти CD20 з унікальними епітопами, може використовуватись сайт-направлений або мультисайт-направлений мутагенез. Для визначення ізотипу очищених антитіл може проводитись аналіз ELISA на ізотипи. Лунки мікротитрувальних планшетів можуть покриватись 10 мкг/мл анти-людським Іg на ніч при температурі 4 °С. Після блокування з використанням 5% бичачого сироваткового альбуміну (BSA), планшети реагують з 10 мкг/мл моноклональних антитіл або очищених контрольних зразків ізотипів при темпе 93478 42 ратурі навколишнього середовища протягом двох годин. Після цього лунки можуть водити у взаємодію або з людським IgG1, lgG2, lgG3 або lgG4, або з людськими lgM-специфічними лужними фосфатаза-кон, at ambient temperature for two hours югованими зондами. Після промивання пластинки проявляють з допомогою субстрату pNPP (1 мг/мл) і аналізують при OD 405-650. Щоб продемонструвати присутність антитіл проти CD20 у сироватках імунізованих мишей або зв‘язування моноклональних антитіл з живими клітинами, які експресують CD20 може застосовуватись проточна цитометрія. Коротко кажучи, клітинні лінії, які експресують CD20 (ріст за стандартних умов росту), змішують з різними концентраціями моноклональних антитіл у фосфатно-сольовому буферному розчині, який містить 0.1% бичачого сироваткового альбуміну (BSA) і 0.02% натрій-азиду, і інкубують при температурі 4 °С протягом 30 хвилин. Після промивання клітини вводять у взаємодію з міченими флюоресцеїном анти-людським антитілом IgG за тих самих умов, за яких проводиться забарвлювання первинних антитіл. Одержані зразки можуть аналізуватись проточною цитометрією з допомогою активованого флюоресценцією аналізатора клітин, застосовуючи властивості світла і бокового розсіювання для стробування одиничних живих клітин. Альтернативним аналізом, в якому застосовується флюоресцентна мікроскопія, який може використовуватись як доповнення або заміна згаданому аналізу, є проточна цитометрія. Клітини можуть забарвлюватись таким самим чином, як описувалось вище, і аналізуватись з допомогою методів флюоресцентної мікроскопії. Такий метод дозволяє зробити видимими окремі літини, але, можливо, знижує чутливість залежно від густини антигену. Людські антитіла IgGs проти CD20 можуть додатково аналізуватись на реакційну здатність щодо антигену CD20 з допомогою вестерн-блоттингу. Коротше кажучи, екстракти клітин, які експресують CD20, можуть бути одержані і оброблені електрофорезом в гелі на основі поліакриламіду додецилсульфату натрію. Після електрофорезу розділені антитіла переносять на нітроцелюлозні мембрани, блокують 20% мишачою сироваткою і зондують моноклональними антитілами, які підлягають аналізу. Зв'язування людського IgG може визначатися з допомогою анти-людської лужної IgG фосфатази і проявлятися з допомогою субстратних таблеток BCIP/NBT (Sigma Chem. Co., St. Louis, MO). Фагоцитарна та кіперна клітинна активність людських моноклональних антитіл проти CD20 Окрім специфічного зв'язування з CD20, людські моноклональні антитіла проти CD20 можуть аналізуватись на їх здатність опосередковувати фагоцитоз та убивання клітин, які експресують CD20. Аналіз активності моноклональних антитіл in vitro забезпечує початковий скринінг перед тестуванням моделей in vivo. Коротше кажучи, поліморфонуклеарні клітини (PMNs), NK-клітини, моноцити або інші клітини-ефектори від здорових донорів можуть очищуватись з допомогою центрифугування в градієнті густини FiсоІІ 43 Hypaque з наступним лізисом домішкових еритроцитів. Промиті поліморфонуклеарні клітини можуть суспендуватись в індикаторі числа обертів за хвилину з 10% дезактивованою теплом сироваткою 51 плоду корови і змішуватись з Сr-міченими клітинами, які експресують CD20, при різних співвідношеннях кількості клітин ефекторів до кількості пухлинних клітин (-клітини-ефекторигпухлинні клітини). Після цього можуть додаватись очищені людські антитіла IgGs проти CD20 у різних концентраціях. В якості негативних контрольних зразків модуть використовуватись неспоріднені людські IgG. Аналізи можуть проводитись протягом від 4 до 20 годин при температурі 37 °С залежно від типу застосовуваних клітин-ефекторів. Зразки можуть аналізуватись на цитологію шляхом визначення вивільнення 51Сr у супернатант культури. Моноклональні антитіла проти CD20 також можуть аналізуватись у поєднанні одне з одним з метою встановлення, чи посилюється цитоліз у випадку з групою моноклональних антитіл. Людські моноклональні антитіла, які зв‘язуються з CD20, також можуть аналізуватись на моделі in vivo (наприклад, у мишей) для визначення їх ефективності у контролюванні росту пухлинних клітин, які експресують CD20. Такі антитіла можуть, наприклад, вибиратися на основі наступних критеріїв, які не є вичерпними: 1. зв'язування і живими клітинами, які експресують CD20; 2. низький ступінь дисоціації від CD20; 3. висока спорідненість у зв'язуванні з CD20; 4. зв'язування з унікальним епітопом на CD20; і/або зв'язування у специфічній орієнтації з CD20, і/або зв'язування зі специфічною формою CD20; 5. опсонізація клітин, які експресують CD20; 6. опосередкування інгібування росту, фагоцитоз і/або убивання клітин, які експресують CD20, у присутності людських клітин-ефекторів; 7. здатність індукувати CDC як на СD55/СD59негативних, так і на CD55/CD59-позитивних клітинах; 8. здатність індукувати гомотипову адгезію; 9. здатність індукувати транслокацію на ліпідних масах при зв'язуванні з CD20; 10. здатність індукувати апоптоз; 11. здатність індукувати ADCC на клітинах, які експресують CD20; 12. здатність зменшувати кількість клітин, які експресують CD20; і/або 13. здатність зменшувати кількість клітин, які експресують низькі рівні CD20 (клітини CD20low). Переважні людські моноклональні антитіла згідно з винаходом відповіднають одному або більше із вищеназваних критеріїв. Людські моноклональні антитіла проти CD20 можуть аналізуватись на їх здатність 25 опосередковувати комплементарну залежну цитотоксичність (CDC) з допомогою ряду відомих методик. Наприклад, сироватку для комплементу можна одержати із крові здорових суб'єктів, яка може бути центрифугована і збирана. Для визначення CDC-активності різних mAbs, можуть застосовуватись різні методики. Наприклад, вивільнення 51Сr або підвищена проникність мембран може оціню 93478 44 ватись з допомогою аналізу на виключення йодиду пропідію. Коротше кажучи, клітини-мішені можуть промиватись і ресуспендуватись в RPMI з 1% бичачим сироватковим альбуміном BSA при 1 х 6 10 /мл. До клітин можуть додаватися різні концентрації mAb, і їм дають зв'язатися протягом 10-15 хвилин при кімнатній температурі. Після цього може бути додана сироватка з доведенням концентрації до 20% (об./об.), і клітини можуть інкубуватись при 37 °С протягом 45 хвилин. Всі клітини із кожного зразка можуть додаватись до розчину Рl в активованій флюоресценцією трубці-аналізаторі клітин (FACS). Після цього суміш може одразу ж аналізуватись проточною цитометрією з допомогою проточного цитометра FACScalibur і з допомогою програмного забезпечення CellQuest pro (BD Biosciences, Mountain view, CA). Для аналізу здатності ініціювати апоптоз людські моноклональні антитіла проти CD20 можуть, наприклад, інкубуватись з СD20-позитивними пухлинними клітинами, наприклад, клітинами Дауді при температурі 37 °С протягом 20 годин. Такі клітини можуть збиратись, промиватись у зв'язуючому буфері Annexin-V-FITC (BD biosciences) і мітитись з допомогою Annexin V-FITC (BD biosciences) протягом 15 хвилин у темряві при температурі 4 °С. Всі клітини із кожного зразка можуть додаватись до розчину РІ (10 мкг/мл у фосфатносольовому буферному розчині (PBS)) у активованій флюоресценцією трубці-аналізаторі клітин (FACS) і одразу ж аналізуватись проточною цитометрією (як описано вище). Відповідно до одного з варіантів винаходу людські моноклональні антитіла використовуються комбінації одного з одним, наприклад, у формі фармацевтичної композиції, яка містить два або більше людських моноклональних антитіла проти CD20. Наприклад, людські моноклональні антитіла проти CD20, які мають відмінні, але комплементарні активності, можуть поєднуватись в одній лікарській формі для досягнення бажаного терапевтичного або діагностичного ефекту. Відповідно до переважного варіанту така композиція включає людське моноклональне антитіло проти CD20, яке опосередковує комплементарну залежну цитотоксичність (CDC), у поєднанні з іншим людським моноклональним антитілом проти CD20, яке індукує апоптоз. Відповідно до ще одного варіанту така композиція включає людське моноклональне антитіло проти CD20, яке опосередковує високоефективне убивання клітин-мішеней у присутності клітин-ефекторів, у поєднанні з іншим людським моноклональним антитілом проти CD20, яке інгібує ріст клітин, які експресують CD20. II. Виведення трансгенних тварин, відмінних від людини, які генерують людські моноклональні антитіла проти CD20 Відповідно до ще одного аспекту винахід стосується трансгенних і трансхромосомних тварин, відмінних від людини, таких як трансгенні або трансхромосомні миші, які здатні експресувати людські антитіла, які специфічно зв'язуютсья з CD20. Відповідно до одного з варіантів винахід стосується трансгенної або транс хромосомної миші, яка має геном, що включає людський трасген з важким 45 ланцюгом, завдяки якому миша виробляє людські антитіла проти CD20 в результаті імунізації клітинами, які експресують CD20. Людський трансген з важким ланцюгом може бути включений у хромосомну ДНК миші, як у випадку з трансгенною, наприклад мишею HuMAb, що детально описано у цій заявці і проілюстровано прикладами. Альтернативно людський трансген з важким ланцюгом може зберігатись поза хромосомами, як у випадку з трансхромосомною (наприклад, KM) мишею, що описано у заявці WO 02/43478. Такі трансгенні і трансхромосомні тварини здатні виробляти множинні ізотипи людських моноклональних антитіл проти CD20 (наприклад, IgG, IgA і/або IgE) за рахунок V-D-J/V-J рекомбінації і переключення ізотипів. Конструкція трансгенної або транс хромосомної тварини, відмінної від людини, яка реагує на стимуляцію чужорідним геном з гетерологічним репертуаром антитіл, потребує, щоб гетерологічні трансгени імуноглобуліну, носієм яких є така тварина, функціонували коректно упродовж всього шляху розвитку В-клітин. Це, наприклад, включає переключення ізотипів в гетерологічному трансгені з важким ланцюгом. Відповідно, трансгени конструюються таким чином, щоб можна було індукувати переключення ізотипів, а також, щоб гени антитіл мали одну або більше із наведених властивостей: (1) високий рівень клітинно-специфічної експресії, (2) функціональне перегрупування генів, (3) активація і реакція на алельні виключення, (4) експресія достатнього первинного репертуару, (5) сигнальна трансдукція, (6) соматична гіпермутація і (7) домінування положення трансгену антитіла під час імунної реакції. Немає необхідності у тому, щоб задовольнялись усі критерії. Наприклад, у тих варіантах, в яких положення ендогенного імуноглобуліну у трансгенних тварин функціонально порушені, трансген не має активувати виключення алелі. Крім того, у тих варіантах, в яких такий трансген включає функціонально перегрупований ген імуноглобуліну з важким і/або легким ланцюгом, у відповідності другому критерію функціонального перегрупування гену немає необхідності, принаймні щодо трансгену, який вже був перегрупований. Щодо основ молекулярної імунології див. Fundamental Immunology (Фундаментальна імунологія), 2nd edition (1989), Paul William E., ed. Raven Press, N.Y. У деяких варіантах трансгенні або трансхромосомні тварини, які генерували людські моноклональні антитіла згідно з винаходом, містили у зародковій лінії перегруповані, неперегруповані або комбінацію перегрупованих і непергрупованих трансгенів імуноглобуліну з важкими і легкими ланцюгами. Кожен із трансгенів з важким ланцюгом містить принаймні один ген Сн. Крім того, трансген з важким ланцюгом може містити функціональні послідовності переключення ізотипів, які здатні підтримувати переключення ізотипів гетерологічного трансгену, який коду множинні гени Сн в В-клітинах трансгенної тварини. Такі послідовності переключення можуть представляти собою природні послідовності, які зустрічаються у положенні імуноглобулінової зародкової лінії у видів, які є джерелом генів трансгену Сн, або ж такі послідо 93478 46 вності переключення можуть бути похідними від послідовностей, які зустрічаються у видів, які отримують конструкт трансгену (трансгенна тварина). Наприклад, людський конструкт трансгену, який використовується для виведення трансгенної миші, може викликати більш часте переключення ізотипів, якщо він включатиме послідовності переключення, подібні до тих, які зустрічаються у природі у мишей у положенні важкого ланцюга, оскільки мишачі послідовності переключення, ймовірно, оптимізовані для функціонування з мишачою системою ензиму переключення рекомбіназою, тоді як людські послідовності переключення до цього не пристосовані. Послідовності переключення можуть бути виділені і клоновані з допомогою традиційних методик, або ж вони можуть бути синтезовані de novo з накладних синтетичних олігонуклеотидів, сконструйованих на основі опублікованої інформації щодо послідовностей у ділянці переключення в імуноглобулінах (Mills et al., Nucl. Acids Res. 15:73057316 (1991); Sideras et at., Intl. Immunol. 1:631-642 (1989)). У випадку кожного із далі згаданих трансгенних тварин трасгени імуноглобуліну з функціонально перегрупованими важкими і легкими ланцюгами виявляються у значній фракції В-клітин трансгенної тварини (принаймні 10 відсотків). Трансгени, які застосовують для виведення трансгенних тварин, відмінних від людини, згідно з винаходом включають трансген з важким ланцюгом, який включає ДНК, що кодує принаймні один змінний сегмент гену, один з'єднуючий сегмент гену і принаймні один генний сегмент у сталій ділянці. Імуноглобуліновий трансген з легким ланцюгом включає ДНК, яка кодує принаймні один змінний сегмент гену, один з'єднуючий сегмент гену і принаймні один генний сегмент у сталій ділянці. Генні сегменти, які кодують генні сегменти з легкими і важкими ланцюгами, є гетерологічними з генними сегментами у трансгенної тварини у тому, що вони одержуються з або відповідають ДНК, яка кодує імуноглобулінові генні сегменти з важким і легким ланцюгом і присутні у видів, до яких не входять трансгенні тварини, відмінні від людини. Відповідно до одного аспекту цього винаходу странсген сконструйований таким чином, що індивідуальні генні сегменти у ньому не перегруповуються, тобто не угруповані для кодування функціонального легкого або важкого імуноглобулінового ланцюга. Такі неперегруповані трансгени підтримують рекомбінацію генних сегментів V, D і J (функціональне перегрупування) і переважно підтримують включення всього або частини генного сегменту D-ділянки в одержаному перегрупованому імуноглобуліновому важкому ланцюгу у трансгенної тварини під дією антигену CD20. Відповідно до альтернативного варіанту трансгени включають не перегрупований "міні-локус". Такі трансгени зазвичай включають значну частину С, D і J сегментів, також підгрупу генних V сегментів. У таких трансгенних конструктів різні регуляторні послідовності, наприклад промотори, генипосилювачі, ділянки, що беруть участь у переключенні класів, послідовності донорів сплайсованих фрагментів і акцепторів сплайсованих фрагментів 47 процесованої РНК, рекомбінаційні сигнали і подібні, включаючи відповідні послідовності, похідні від гетерологічної ДНК. Такі регуляторні послідовності можуть бути включені у трансген від такого ж самого або спорідненого виду тварини, відмінної від людини, яку використовують у цьому винаході. Наприклад, генні сегменти людського імуноглобуліну можуть поєднуватись у трансгені із імуноглобуліновою послідовність гену-посилювача гризуна для застосування на трансгенній миші. Альтренативно у такий трансген можуть бути включені синтезовані регуляторні послідовності, тоді як у такому трансгені синтетичні регуляторні послідовності не є гомологічними з функціональною ДНКпослідовністю, яка, як відомо, зустрічається у геномах ссавців. Синтетичні регуляторні послідовності конструюються за правилами консенсусу, такими як, наприклад, правила визначення допустимих послідовностей у сайтах акцепторів сплайсованих фрагментів або у мотиві промотора/генапосилювача. Наприклад, мінілокус включає частину локусу геномного імуноглобуліну, який має принаймні одну внутрішню (тобто таку, що знаходиться не на кінці частини) делецію несуттєвої частини ДНК ДНК (наприклад, вбудована послідовність; інтрон або його частина), на відміну від існуючого в природі локусу Іg зародкової лінії. Переважні трансгенні і трансхромосомні тварини, відмінні від людей, наприклад, миші демонструють продукування імуноглобуліну з широким репертуаром, в ідеалі по суті подібним до людського після регулювання об'єму. Переважно репертуар наближається до продемонстрованого людиною після регулювання об'єму, зазвичай із різноманітністю принаймні приблизно максимум 10%, переважно 25 - 50% або більше. Загалом виробляється принаймні тисяча різних імуноглобулінів (в ідеалі IgG), переважно від 104 до 106 або більше, що залежить від числа різних V, J і D ділянок, включених у геном миші і активованих додатковим розмаїттям, що генерується вибірковими нуклеотидними вставками перегрупованих генних сегментів V(-D-)J і ділянках з'єднання. Як правило, імуноглобуліни демонструють спорідненість (KD) щодо попередньо вибраних антигенів 10-7 М, наприклад менше 10-8 М, 10-9 М або 10-10 М, або навіть нижче цього значення. Трансгенні і трансхромосомні тварини, відмінні від людей, наприклад, миші, як описано вище, можуть імунізуватись, наприклад, клітинами, які експресують CD20. Альтернативно трансгенні тварини можуть бути імунізовані ДНК, яка кодує людський ; CD20. В результаті такої імунізації тварини продукують В-клітини, які зазнають переключення класів через механізм свіч-рекомбінації (цис-переключення) і експресують імуноглобуліни, які здатні реагувати з CD20. Такі імуноглобуліни можуть бути людськими антитілами (також згадувані як "антитіла з людським послідовностями"), в яких поліпептиди з важким і легким ланцюгом кодуються людськими трансгенними послідовностями, які можуть включати послідовності, похідні від соматичної мутації і рекомбінаторних з'єднань у Vділянці, а також кодовані зародковою лінією послідовності. Такі людські антитіла можуть вважатися 93478 48 по суті ідентичними поліпептидній послідовності, яка кодується людською VL і JL або генними сегментами VH, DH і JH, хоча в результаті соматичної мутації і різниці рекомбінаційних з'єднань V-J і VD-J можуть бути присутні інші послідовності. Змінні ділянки кожного ланцюга антитіла принаймні на 80 відсотків подібні до людських зародкових ліній V, J і, у випадку важких ланцюгів, D, генних сегментів; часто принаймні на 85 відсотків подібні до послідовностей людської зародкової лінії, присутніх на трансгені; часто на 90 або 95 відсотків або більше подібні до послідовностей людської зародкової лінії, присутніх на трансгені. Однак, оскільки послідовності, відмінні від послідовностей зародкової лінії, включаються через механізм соматичної мутації і приєднання VJ і VDJ, антитіла до людських послідовностей часто мають послідовності змінних ділянок, які не кодуються людськими генними сегментами V, D або J, як це було виявлено у людських трансгенах у зародковій лінії миші. Зазвичай такі послідовності зародкової лінії (або позиції окремих нуклеотидів) згруповуються а або біля CDR, або у ділянках, в яких, як відомо, згруповуються соматичні мутації. Ще один аспект винаходу стосується В-літин, похідних від трансгенних або трансхромосомних тварин, відмінних від людини, як описано у цій заявці. Такі В-клітини можуть застосовуватись для генерування гібридом, які експресують людські моноклональні антитіла, що мають властивості зв'язування, високо споріднені з (наприклад, конс-7 татна рівноваги дисоціації (КD) менше, ніж 10 М) людським CD20. Таким чином, відповідно до ще одного варіанту винахід стосується гібридоми, яка виробляє людське антитіло із подібністю (KD) менше 10-7 М, наприклад менше 10-8 М, 10-9 М або 10-10 М або навіть менше при визначенні з допомогою аналізу Скетчарда клітин, ;які експресують CD20, використовуючи мічені радіоактивним ізотопом моноклональні антитіла,або при визначення половини максимальної концентрації зв'язування з допомогою аналізу з використанням активованого флюоресценцією аналізатора клітин. У контексті цього винаходу моноклональне антитіло включає легкий ланцюг людської послідовності, який складається з (1) змінної ділянки легкого ланцюга, яка включає поліпептидну послідовність, яка по суті ідентична з поліпептидною послідовністю, яка кодується людським генним VL -сегментом і людським JL -сегментом, і (2) сталої ділянки легкого ланцюга, яка кодується людським генним сегментом СL; і Важкого ланцюга людської послідовності, яка складається з (1) змінної ділянки важкого ланцюга, яка має поліпептидну послідовність, по суті ідентичну з поліпептидною послідовністю, яка кодується людським генним сегментом VH, D-ділянкою і людським сегментом JH, і (2) сталої ділянки, яка кодується людським генним сегментом Сн. Розвиток вискоспоріднених людських моноклональних антитіл проти CD20 може бути прискорений з допомогою методики розширення репертуару генних сегментів у людській змінній ділянці у трансгенної тварини, відмінної від людини, яка має геном, який включає інтегрований людський транс 49 ген імуноглобуліну, при цьому така методика полягає у включенні до генома трансгену V-гену, який містить генні сегменти V-ділянки, відсутні у згаданому трансгені людського імуноглобуліну. Часто в якості трансгену V-ділянки виступає штучна хромосома дріжджів, яка містить частину ряду сегментів людського гену VH або VL (VK), що може мати місце в природі у людському геномі або може траплятись в результаті сплайсування, яке індивідуально здійснюють з використанням рекомбінантних методик, що може включати позачергові або пропущені сегменти V-гену. Часто принаймні п'ять або більше функціональних сегментів V-гену містяться на YAC. У такому варіанті можна вивести трансгенну тварину з допомогою V-методики розширення репертуару, в якому тварина експресує імуноглобуліновий ланцюг, який включає послідовність змінної ділянки, яка кодується генним сегментом V-ділянки, присутнім на трансгені V-ділянки, і Сділянки, яка кодується на людському трансгені Іg. З допомогою V-метод розширення репертуару, можуть бути одержані трансгенні тварини, які мають принаймні відмінних V-генів; також можуть бути одержані тварини, які мають принаймні приблизно 24 V-гени і більше. Деякі сегменти V-гену можуть бути нефункціональними (наприклад, псевдогени і подібні). Такі сегменти можуть зберігатися або при бажанні селективно видалятися з допомогою рекомбінантних методик , відомих фахівцям у цій галузі. Одразу ж після конструювання зародкової лінії, яка містить функціональну YAC з розширеним репертуаром V-сегменту, який по суті відсутній у людському трансгені Іg, що містить генні сегменти J і С, така риса може бути поширена і впроваджена і інші генетичні фони, включно з фоном, в якому функціональна YAC, що має розширений репертуар V-сегмента, проявляється у зародковій лінії тварини, відмінної від людини, яка має відмінний людський трансген Іg. Поліфункціональні YAC з розширеним репертуаром V-сегменту можуть виводитися у зародковій лінії для взаємодії з людським трансгеном Ig (або множинними трансгенами Ig). Хоча ці трансгени називаються тут YACтрансгенами, такі трансгени, будучи інтегрованими у геном, можуть не містити послідовностей дріжджів, таких як ті, що необхідні для автономної реплікації у дріжджах. Такі послідовності можуть необов'язково видалятися з допомогою методик генної інженерії (наприклад, рестрикційне розчинення і імпульсний електрофорез у гелі або інша придатна методика) після того, як у реплікації дріжджів більше немає необхідності (тобто перед введенням у ES-клітину миші або мишачу прозиготу). Методики поширення риси експресії послідовності людського імуноглобуліну включають виведення трансгенної тварини, яка має людський трансген(и) Ig transgene(s), а також необов'язково має функціональний YAC із розширеним репертуаром V-сегмента. Як сегменти гену VH, так і сегменти гену VL можуть бути наявними у YAC. Трансгенна тварина може бути виведена з будь-яким генним фоном, юажаним для фахівця, включно з фоном, в якому закріплені інші людські трансгени, включно з людськими трансгенами Ig і/або транс 93478 50 генами, які кодують інші людські лімфоцитарні протеїни. Винахід також стосується послідовностей людського імуноглобуліну з високим ступенем спорідненості, які продукуються трансгенною мишею, у якої трансген YAC має V-ділянку з розширеним репертуаром. Хоча наведений далі опис розкриває переважний варіант трансгенної тварини згідно з винаходом, можливі також інші варіанти, які об'єднуються у три категорії: I. Трансгенні тварини, які мають імуноглобуліновий трансген з неперегрупованим важким і перегрупованим легким ланцюгом; II. Трансгенні тварини, які мають імуноглобуліновий трансген з неперегрупованим важким і легким ланцюгом; і III. Трансгенні тварини, які мають імуноглобуліновий трансген з перегрупованим важким і неперегрупованим легким ланцюгом. Серед вищеназваних категорій за ступенем переважності ці категорії можуть бути перелічені у такому порядку II > І > III, де гени ендогенного легкого ланцюга (або принаймні К-ген) були відкинуті шляхом гомологічної рекомбінації (або іншим способом), і І > II > III, де гени ендогенного легкого ланцюга не були відкинуті і мають домінуватися аллельним виключенням. III. Біспецифічні/мультиспецифічні молекули, які зв'язуються з CD20 Відповідно до ще одного варіанту винаходу людські моноклональні антитіла проти CD20 можуть бути дериватизовані або зв'язані з іншою функціональною молекулою, наприклад з іншим пептидом або протеїном (наприклад, з фрагментом Fab'), генеруючи біспецифічну або мультиспецифічну молекулу, яка зв‘язується з багатьма зв'язувальними сайтами або епітопами-мішенями. Наприклад, антитіло згідно з винаходом може бути функціонально зв'язане (наприклад, хімічним сполученням, злиттям генів, нековалентним зв'язком та іншим чином) з однією або більше зв'язуючими молекулами, такими як інше антитіло, пептид або зв'язуючий міметик. Відповідно, цей винахід стосується біспецифічних і мультиспецифічних молекул, які мають принаймні одну першу зв'язувальну специфічність щодо CD20 і другу зв'язувальну специфічність щодо другого епітопа-мішені. Відповідно до особливо переважного варіанту винаходу другим епітопом-мішенню є Fc-рецептор, наприклад, людський FcRI (CD64) або людський Fc-рецептор (CD89), або рецептор Т-клітин, наприклад CD3. У зв'язку з цим винахід включає біспецифічні і мультиспецифічні молекули, здатні зв'язуватися як з клітинами-ефекторами, які експресують FcR, FcR або FcR (наприклад, моноцити, макрофаги або полімононуклеарні клітини (PMNs)), такі і з клітинами-мішенями, які експресують CD20. Такі біспецифічні і мультиспецифічні молекули спрямовують клітини, що експресують CD20, на клітини-ефектори і, подібно до людських моноклональних антитіл згідно з винаходом, збуджують активність клітин-ефекторів, які опосередковуються Fc-рецептором, а саме фагоцитоз клітин, які експресують CD20, залежну від антитіл клітинну 51 цитотоксичність (ADCC), вивільнення цитокінів або генерування супероксидного аніону. Біспецифічні і мультиспецифічні молекули згідно з винаходом можуть додатково включати третю зв'язуючу специфічність, окрім анти-Fc зв'язуючої специфічності і анти-CD20 зв'язуючої специфічності. Відповідно до ще одного варіанту третя зв'язуюча специфічність представляє собою частину анти-посилюючого фактора (EF), наприклад, молекулу, яка зв'язується з поверхнею протеїну, який бере участь у цитотоксичній активності, і тим посилює імунну реакцію на клітини-мішені. Частина антипосилюючого фактора може представляти собою фрагмент антитіла або ліганд, який зв'язується з певною молекулою, наприклад, антигеном або рецептором, завдяки чому посилюється ефект детермінант зв'язування для Fсрецептора або антигену клітин-мішеней. Частина антипосилюючого фактора може зв'язуватися з Fc -рецептором або антигеном клітини-мішені. Альтернативно частина антипосилюючого фактора може зв'язуватися з одиницею, яка відмінна від одиниці, з якою зв'язуються перша і друга специфічності. Наприклад, частина антипосилюючого фактора може зв'язуватися з цитотоксичними Тклітинами (наприклад через CD2, CD3, CD8, CD28, CD4, CD40, ІСАМ-1 або іншу імунну клітину, завдяки чому посилюється імунна реакція на клітинимішені). Відповідно до ще одного варіанту біспецифічні і мультиспецифічні молекули згідно з винаходом в якості зв'язуючої специфічності включають принаймні одне антитіло, включно з, наприклад, Fab, Fab', F(ab')2, Fv або одноланцюгове Fv. Таке антитіло також може представляти собою димер з важким або легким ланцюгом або будь-який його мінімальний фрагмент, такий як Fv або одноланцюговий конструкт, описаний у Ladner et аl., патент США 4,946,778. Також таке антитіло може представляти собою зв'язуючий домен, який представляє собою злитий протеїн імуноглобуліну, як описано в заявках США 2003/0118592 і US 2003/0133939. Відповідно до одного варіанту біспецифічні і мультиспецифічні молекули згідно з винаходом включають зв'язуючу специфічність для FcR або FcR, присутніх на поверхні клітини-ефектора, та другу зв'язуючу специфічність для антигену клітинмішеней, наприклад, CD20. Відповідно до одного з варіантів зв'язуюча специфічність щодо Fc-рецептора забезпечується людським моноклональним антитілом, зв'язування якого не заблоковане людським імуноглобуліном G (IgG). У контексті цього винаходу термін "рецептор IgG" вживається на позначення будь-якого з восьми у-ланцюгових генів, розташованих на хромосомі 1. Ці гени загалом кодують дванадцять трансмембран або ізоформ розчинного рецептора, які групуються у три класи рецептора Fc: FcRI (CD64), FcRII(CD32) і FcRIII (CD16). Відповідно до одного з переважних варіантів рецептор Fc високо споріднений з людським рецептором FcRl. Продукування і характеристика цих переважних моноклональних антитіл описані Fanger et al. у міжнародній заявці WO 88/00052 і патенті США 93478 52 4,954,617. Ці антитіла зв'язуються з епітопом FcRI, FcRII або FcRIII на сайті, який відрізняється від зв'язувального сайту Fc рецептора, і, відповідно, їх зв'язування суттєво не блокується фізіологічними рівнями IgG. Специфічними антитілами проти FcRI, які використовуються згідно з винаходом, є mAb 22, mAb 32, mAb 44, mAb 62 і mAb 197. Відповідно до ще одного варіанту антитіло проти рецептора Fey представляє собою олюднену форму моноклонального антитіла 22 (Н22). Продукування і характеристики антитіла Н22 описані у Graziano, R.F. etal. (1995) J. Immunol 155 (10): 4996-5002 і заявці WO 94/10332. Клітинна лінія, яка продукує антитіло Н22, була депонована в Американській колекції типів культур 4 листопада 1992 року за позначенням HA022CL1, і має номер доступу CRL 11177. Відповідно до інших переважних варіантів зв'язуюча специфічність щодо Fc-рецептора забезпечується антитілом, яке зв'язується з людським рецептором ІdА, наприклад, рецептором Fcальфа (FcRI (CD89)), зв'язування якого переважно не блокується людським імуноглобуліном А (ІgА). Термі "рецептор ІgА" охоплює генний продукт а-гену (FcRI), розміщеного на хромосомі 19. Відомо, що цей ген кодує декілька альтернативно сплайсованих трансмембранних ізоформ від 55 до 110 kDa. FcRI (CD89) конститутивно експресується на моноцитах/макрофагах, еозинофільних і нейтрофільних гранулоцитах, але не в популяціях неефекторних клітин. FcRI має середню спорідненість щодо ІgА1 і ІgА2, яка зростає під дією цитокінів, таких як G-CSF або GM-CSF (Morton, Н.С. et al. (1996) Critical Reviews in Immunology 16:423440). Чотири FcRI-специфічні моноклональні антитіла, ідентифіковані як A3, А59, А62 і А77, які зв'язуються з FcRI поза лігандним зв'язуючим доменом ІgА, були описані у Monteiro, R.C. et al. (1992) J. Immunol. 148:1764. FcRI і FcRI є переважними трігерними рецепторами, які застосовуються у цьому винаході, тому що вони (1) експресуються переважно на імунних клітинах-ефекторах, наприклад, моноцитах, PMNs, макрофагах і дендровидних клітинах; (2) експресуються у великих об'ємах (наприклад, 5,000-100,000 на клітину); (3) є медіаторами цитотоксичної активності (наприклад, залежної від антитіл цитотоксичності АDСС, фагоцитозу); (4) опосередковують посилені антигенні властивості у антигенів, включно з само-антигенами, які націлені на них. Відповідно до ще одного варіанту біспецифічна молекула складається з двох людських моноклональних антитіл згідно з винаходом, які мають комплементарну функціональну активність, таку як одне антитіло, яке головним чином індукує комплементарну залежну цитотоксичність, а друге антитіло головним чином індукує апоптоз, наприклад 2F2 у поєднанні з 11В8. Відповідно до інших варіантів біспецифічні і мультиспецифічні молекули згідно з винаходом додатково включають зв'язуючу специфічність, яка розпізнає, наприклад, зв'язується з антигеном клітини-мішені, наприклад, CD20. Відповідно до пе 53 реважного варіанту зв'язуюча специфічність забезпечується людським моноклональним антитілом згідно з винаходом. Термін "специфічне антитіло клітини-ефектора " у контексті цього винаходу означає антитіло або фрагмент функціонального антитіла, яке зв'язується з Fc-рецептором клітин-ефекторів. Переважні антитіла, які застосовуються у цьому винаході, зв'язуються з Fc-рецептором клітин-ефекторів у сайті, який не зв'язаний ендогенним імуноглобуліном. 5 У контексті цього винаходу термін "клітинаефектор" вживається на позначення імунної клітини, яка бере участь в ефекторній фазі імунної реакції, на противагу когнітивній фазі і фазі активації імунної реакції. Приклади імунних клітин включають клітину мієлоїдного або лімфоїдного походження, наприклад, лімфоцити (наприклад, Вклітини і Т-клітини, включно з цитолітичними Тклітинами (CTLs)), клітинами-кілерами, природними клітинами-кілерами, макрофагами, моноцитами, еозинофільними клітинами, нейтрофільними клітинами, полімононуклеарними клітинами, гранулоцитами, гепариноцитами і базофільними клітинами. Деякі клітини-ефектори експресують специфічні Fc-рецептори і виконують специфічні імунні функції. Відповідно до переважних варіантів клітина-ефектор здатна індукувати залежну від антитіл клітинну цитотоксичність (ADCC), наприклад, нейтрофільна клітина, здатна індукувати ADCC. Наприклад, моноцити, макрофаги, які експресують FcR, беруть участь у специфічному умертвленні клітин-мішеней і піддаванні антигенів дії інших компонентів імунної системи або у зв'язуванні з клітинами, які представляють антигени. Відповідно до інших варіантів клітина-ефектор може фагоцитувати антиген-мішень, клітинумішень або мікроорганізм. Експресія певного FcR на клітині-ефекторі може регулюватись гуморальними факторами, таким як цитокіни. Наприклад, було виявлено, що експресія FcRI може підвищуватись з допомогою інтерферону гамма (IFN-). Така посилена експресія підвищує цитотоксичну активність клітин, які є носіями FcRI, проти мішеней. Клітина-ефектор може фагоцитувати або лігувати антиген-мішень або клітину-мішень. "Клітина-мішень" означає будь-яку небажану клітину у суб'єкта (наприклад, у людини або тварини), на яку може бути направлена дія композиції (наприклад, людське моноклонапьне антитіло, біспецифічна або мультиспецифічна молекула) згідно з винаходом. Відповідно до переважних варіантів, клітина-мішень представляє собою клітину, яка експресує або експресує CD20 у значних кількостях. Клітини, які експресують CD20, зазвичай включають В-клітини і В-клітинні пухлини. Тоді як віддається перевага людським моноклональним антитілам, інші антитіла, які можуть використовуватись згідно з винаходом у біспецифічних або мультиспецифічних молекулах, є мишачі, химерні і олюднені моноклональні антитіла. Такі мишачі, химерні і олюднені моноклональні антитіла можуть бути одержані з допомогою методик, відомих фахівцям у цій галузі. 93478 54 Біспецифічні і мультиспецифічні молекули згідно з винаходом можуть бути одержані з викорстанням хімічних методик (див. наприклад, D. М. Kranz et al. (1981) Рrос. Natl. Acad. Sci. USA 78:5807), "полідомних" методик (див. патент США 4,474,893, виданий Reading), або методик рекомбінантної ДНК. Зокрема, біспецифічні і мультиспецифічні молекули згідно з винаходом можуть бути одержані кон'югуванням складових зв'язувальних специфічностей, наприклад, зв'язувальних специфічностей проти FcR і проти CD20, застосовуючи методики, відомі у цій галузі техніки і описані у нижченаведених прикладах. Наприклад, кожна зв'язувальна специфічність біспецифічної і мультиспецифічної молекули може генеруватися окремо і після цього кон'югуватися одна з одною. Коли зв'язувальними специфічностями є протеїни або пептиди, може застосовуватись ряд зв'язувальних або зшиваючих агентів для ковалентної кон'югації. Приклади зшиваючих агентів включають протеїн А, карбодіімід, N-сукцинімідил-S-ацетилтіоацетат (SATA), 5,5'-дітіобіс (2-нітробензойну кислоту) (DTNB), офенілендімалеїмід (oPDM), N-сукцинімідил-3-(2піридилдитіо)пропіонат (SPDP) і сульфосукцинімідил 4-(N-малеїмідометил)циклогексан-1 карбоксилат (сульфо-SMCC) (див. наприклад, Karpovsky et al. (1984) J. Exp. Med. 160:1686; Liu, MA et al. (1985) Proc. Natl. Acad. Sci. USA 82:8648). Інші методики включаюь ті, які були описані у Paulus (Behring Ins. Mitt. (1985) No. 78,118-132); Brennan et al. (Science (1985) 229:81 -83) і Glennie et al. (J. Immunol. (1987) 139: 2367-2375). Переважними кон'югуючими агентами є SATA і сульфоSMCC, обидва постачаються компанією Pierce Chemical Co. (Рокфорд, Ілінойс). Коли зв'язуючими специфічностями є антитіла, вони можуть кон'югуватися через сульфгідрильний зв'язок С-термінальних шарнірних ділянок двох важких ланцюгів. В особливо переважному варіанті шарнірна ділянка модифікована таким чином, що вона містить непарне число сульфгідрильних залишків, переважно один, перед кон'югуванням. Альтернативно обидві зв'язувальні специфічності можуть кодуватися у тому ж самому векторі і експресуватися та комплектуватися у тій же клітині-хазяїні. Ця методика є особливо ефективною, коли біспецифічна або мультиспецифічна молекула представляє собою mAb х mAb, mAb x Fab, Fab x F(ab')2 або ліганд х Fab злитий протеїн. Біспецифічна і мультиспецифічна молекула згідно з винаходом, наприклад, біспецифічна молекула може бути одноланцюговою молекулою, такою як одноланцюгове біспецифічне антитіло, одноланцюгова біспецифічна молекула, яка включає одне одноланцбгове антитіло і зв'язуючу детермінанту, або одноланцюгова біспецифічна молекула, яка містить дві зв'язуючі детермінанти. Біспецифічні і мультиспецифічні молекули також можуть бути одноланцюговими молекулами або можуть містити принаймні дві одноланцюгові молекули. Методики одержання бі- і мультиспецифічних молекул описані, наприклад у патентах США US 5,260,203; US 5,455,030; US 4,881,175; US 5,132,405; US 55 5,091,513; US 5,476,786; US 5,013,653; US 5,258,498; і US 5,482,858. Зв'язування біспецифічних і мультиспецифічних молекул з їх специфічними мішенями може підтверджуватись імуноферментним аналізом (ELISA), радіоімуноаналізом (RIA), аналіз з використанням активованого флюоресценцією клітинного сортера, біоаналізу (наприклад, інгібування росту), або вестерн-блотінг аналізу. Кожен із названих аналізів загалом визначає присутність комплексів протеїн-антитіло, які представляють особливий інтерес завдяки застосуванню міченого реагента (наприклад, антитіла), специфічного щодо комплексу, який представляє інтерес. Наприклад, комплекси FcR-антитіла можуть виявлятися, застосовуючи, наприклад, ензим-зв'язане антитіло або фрагмент антитіла, який розпізнає і специфічно звмязується з комплексами анттіло-FcR. Альтернативно такі комплекси можуть виявлятися, застосовуючи будь-який із ряду імуноаналізів. Наприклад, антитіло може бути мічене радіоактивним ізотопом і може використовуватись у радіоімунологічному аналізі (RIA) (див., наприклад, Weintraub, В., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986) (Принципи радіоімунологічного аналізу, Сьомий навчальний курс з методик радіолігандного аналізу). Радіоактивний ізотоп може бути виявлений з допомогою таких засобів як р- лічильник або лічильник сцинтиляції, або з допомогою авторадіографії. IV. Імунокон'югати Відповідно до ще одного аспекту цей винахід стосується людського моноклонального антитіла проти CD20, кон'югованого з терапевтичною частиною, такою як цитотоксин, лікарською речовиною (наприклад, імуносупресант) або радіоізотоп. Такі кон'югати в цьому описі згадуються як "імунокон'югати". Імунокон'югати, які включають один або більше цитотоксинів, називаються "імунотоксинами." Цитотоксин або цитотоксичний агент включає будь-який агент, який згубний (наприклад, убиває) для клітин. Приклади включають таксол, цитохалазин В, граміцидин D, етидію бромід, еметін, мітоміцин, етопозид, тенопозид, вінкристин, вінбластин, колхіцин, доксорубіцин, даунорубіцин, дигідроксиантрациндіон, мітоксантрон, мітраміцин, актиноміцин D, 1-дегідротестостерон, глюкокортикоїди, прокаїн, тетракаїн, лідокаїн, пропранолол, пуроміцин та їх аналоги або гомологи. Придатні терапевтичні агенти для одержанн імунокон'югатів згідно з винаходом включають, але не обмежуються цим переліком, антиметаболіти (наприклад, метотрексат, 6-меркаптопурин, 6тіогуанін, цитарабін, флударабін, 5фторурацилдекарбазин), алкілуючі агенти (наприклад, мехлоретамін, тіоепахлорамбуцил, мелфалан, кармустин (BSNU) і ломустин (CCNU), циклофосфамід, бусульфан, дибромманітол, стрептозотоцин, мітоміцин С і цис-дихлордіамін платина (II) (DDP) цисплатина), антрацикліни (наприклад, даунорубіцин (колишній дауноміцин) і доксорубіцин), антибіотики (наприклад, дактиноміцин (колишній актиноміцин), блеоміцин, мітрамі 93478 56 цин і антраміцин (АМС)), а також анти-мітотичні агенти (наприклад, вінкристин і вінблатин). Відповідно до переважного варіанту терапевтичний агент представляє собою цитотоксичний агент або радіотоксичний агент. Відповідно до ще одного варіанту терапевтичним агентом є імуносупресант. Відповідно до ще одного варіанту терапевтичним агентом є GM-CSF. Відповідно до переважного варіанту терапевтичним агентом е доксорубіцин, цисплатин, блеоміцин, сульфат, кармустин, хлорамбуцил, циклофосфамід або рицин А. Антитіла згідно з винаходом також можуть кон'югуватись з радіоізотопами, наприклад, йодом131, ітрієм-90 або індієм-111, для одержання цитотоксичних радіофармацевтичних засобів для лікування пов'язаних з CD20 захворювань, таких як рак. Такі кон'югати антитіл згідно з винаходом можуть застосовуватись для модифікування заданої біологічної реакції, при цьому частина, яка представляє собою лікарську речовину, не повинна розглядатися як така, що обмежує використання класичних хімічних терапевтичних засобів. Наприклад така частина лікарського засобу може бути протеїном або поліпептидом, який має бажані біологічні властивості. Такі протеїни можуть включати, наприклад, ферментно активний токсин або його активний фрагмент, такий як абрин, рисин А, псевдомональний екзотоксин або токсин дифтерії; протеїн, такий як фактор некрозу пухлини або інтерферон -; або модифікатори біологічної реакції, такі як, наприклад, лімфокіни, інтерлейкін-1("IL1"), інтерлейкін-2 ("IL-2"), інтерлейкін-6 ("IL-6"), стимулюючий фактор колонії макрофагів гранулоцитів ("GM-CSF"), стимулюючий фактор колонії гранулоцитів ("G-CSF") або інші фактори росту. Методики кон'югування таких терапевтичних частин з антитілами є добре відомими, див., наприклад, Arnon et al., "Моноклональні антитіла For Immunotargeting Of Drugs In Cancer Therapy" (Моноклональні антитіла для імунонацілювання лікарських засобів у терапії ракових захворювань), у Моноклональні антитіла І Cancer Therapy (Моноклональні антитіла і терапія ракових захворювань), Reisfeld et al. (eds.), crop. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Антитіла For Drug Delivery" (Антитіла для доставки лікарських засобів), у Controlled Drug Delivery (Контрольована доставка лікарських засобів) (2-е вид.), Robinson et al. (eds.), crop. 623-53 (Marcel Dekker, Inc.1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review" (Антитіла у ролі носіїв цитотоксичних агентів у терапії ракових захворювань: огляд), у Моноклональні антитіла '84: Biological I Clinical Applications (Моноклональні антитіла '84: біологічне та клінічне застосування), Pinchera et al. (eds.), crop. 475-506 (1985); "Аналіз, Results, I Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy" (Аналіз, результати і перспектива терапевтичного застосування мічених радіоактивними ізотопами антитіл у терапії ракових захворювань), у Моноклональні антитіла For Cancer Detection I Therapy (Моноклональні антитіла для діагностики та терапії ракових захворювань), Baldwin et al. (eds.), стор. 303-16 (Academic Press 1985), і Thorpe et al., "The 57 Preparation I Cytotoxic Properties Антитіла-Toxin Conjugates" (Одержання і цитотокисчні властивості кон'югатів антитіло-токсин), Immunol. Rev., 62:11958(1982). Відповідно до ще одного варіанту людські моноклональні антитіла згідно з винаходом приєднані до лінкера-хелатора, наприклад тіуксетану, який робить можливим кон'югування антитіла з радіоактивним ізотопом. V. Фармацевтичні композиції Відповідно до ще одного аспекту цей винахід стосується композиції, наприклад, фармацевтичної композиції, яка містить одне або комбінацію людських моноклональних антитіл згідно з винаходом. Такі фармацевтичні композиції можуть бути одержані з використанням фармацевтично прийнятних носіїв або розбавників, а також будь-яких відомих ад'ювантів і наповнювачів відповідно до традиційних методик, таких як описані у Remington: The Science і Practice of Pharmacy, 19th Edition, Gennaro, Ed., Mack Publishing Co., Easton, PA, 1995 (Теорія і практика фармакології, 19-е видання). Відповідно до одного з варіантів композиції включають комбінацію множинних (наприклад, два або більше) виділених людських антитіл згідно з винаходом, які діють відповідно до різних механізмів, наприклад одне антитіло, яке головним чином функціонує, індукуючи комплементарну залежну цитотоксичність, у поєднанні з іншим антитілом, головною функцією якого є індукування апоптозу. Фармацевтичні композиції згідно з винаходом також можуть призначатися в комбінованій терапії, тобто у поєднанні з іншими агентами. Наприклад, комбінована терапія може включати композицію згідно з винаходом з принаймні одним протизапальним агентом або принаймні одним імуносупресантом. У одному із варіантів такі терапевтичні агенти включають один або більше протизапальні агенти, такі як стероїдні лікарські засоби або нестероїдні протизапальні засоби. Переважні агенти, наприклад, включають аспірин та інші саліцилати, інгібітори Сох-2, такі як рофекоксиб (Vioxx) і целекоксиб (Celebrex), нестероїдні протизапальні засоби, такі як ібупрофен (Motrin, Advil), фенопрофен (Nalfon), напроксен (Naprosyn), суліндак (Clinoril), диклофенак (Voltaren), піроксикам (Feldene), кетопрофен (Orudis), дифлунізал (Dolobid), набуметон (Relafen), етодолак (Lodine), оксапрозин (Daypro) та індометацин (Indocin). Відповідно до іншого варіанту такі терапевтичні агенти включають один або більше БПРП, такий як метотрексат (Rheumatrex), гідроксихлорохін (Plaquenil), сульфасалазин (Asulfidine), інгібітори синтезу піримідину, наприклад лефлуномід (Arava), блокатори IL-1 рецептора, наприклад анакінра (Kineret) і блокатори TNF-, наприклад етанерцепт (Enbrel), інфліксимаб (Remicade) і адалімумаб. Відповідно до іншого варіанту такі терапевтичні агенти включають один або більше імуносупресантів, таких як циклоспорин (Sandimmune, Neoral) і азатіоприн (Imural). Відповідно до ще одного варіанту такі терапевтичні агенти включають один або більше хіміоте 93478 58 рапевтичні агенти, такі як доксорубіцин (Adriamycin), цисплатин (Platinol), блеоміцин (Blenoxane), кармустин (Gliadel), циклофосфамід (Cytoxan, Procytox, Neosar) і хлорамбуцил (Leukeran). Відповідно до ще одного варіанту людські антитіла згідно з винаходом можуть призначатися у поєднанні з хлорамбуцилом і преднізолоном; циклофосфамідом і преднізолоном; циклофосфамідом, вінкристином і преднізолоном; циклофосфамідом, вінкристином, доксорубіцином і преднізоном; флударабіном і антрацикліном; або у поєднанні з іншим традиційними багатоліковими режимами для не лімфоми Ходжкіна, як описано наприклад у Non-Hodgkin's Lymphomas: Making sense of Diagnosis, Treatment, і Options (Неходжкінська лімфома: Розуміння діагностики, лікування і рішень), Lorraine Johnston, 1999, O'Reilly і Associates, Inc. Відповідно до ще одного варіанту людські антитіла можуть призначатися у поєднанні з радіотерапією і/або трансплантацією периферійних стовбурових клітин або кісткового мозку. Відповідно до ще одного варіанту людські антитіла можуть призначатися у поєднанні з одним або більше антитілами, вибраними з антитіл проти CD25, антитіл проти CD19, антитіл проти CD21, антитіл проти CD22, антитіл проти CD37, антитіл проти CD38, антитіл проти IL6R, антитіл проти ІL8, антитіл проти IL15, антитіл проти ІL15R, антитіл проти CD4, антитіл проти CD11a (наприклад ефалізумаб), антитіла проти альфа-4/бета-1 інтегрину (VLA4) (наприклад, наталізумаб) і CTLA4-lg. В одному із варіантів людські моноклональні антитіла призначають у поєднанні з антитілом проти CD25 для лікування бульозного пемфігоїду, наприклад у пацієнтів з реакцією «трансплантат проти хазяїна». Відповідно до ще одного варіанту людські моноклональні антитіла призначають у поєднанні з одним або більше антитілами, вибраними з антиітл проти CD19, антитіл проти CD21, антитіл проти CD22, антитіл проти CD37 і антитіл проти CD38 для лікування злоякісних захворювань. У ще одному переважному варіанті людські антитіла призначають у поєднанні з одним або більше антитілами, вибраними з антитіл проти IL6R, антитіл проти IL8, антитіл проти IL15, антитіл проти IL15R, антитіл проти CD4, антитіл проти CD11a (наприклад ефалізумаб), антитіл проти альфа-4/бета-1 інтегрину (VLA4) (наприклад, наталізумаб) і CTLA4-lg для лікування запальних захворювань. Відповідно до ще одного варіанту людські антитіла можуть призначатись у поєднанні з антитілом проти С3b(і) для посилення активацію комплемента. У контексті цього винаходу "фармацевтично прийнятний носій " вживається на позначення будь-якого і всіх розчинників, дисперсійних середовищ, покриттів, антибактеріальних і протигрибкових агентів, ізотонічних агентів і агентів, що затримують абсорбцію, і подібних, які фізіологічно сумісними. Переважно такий носій є придатним для 25 внутрішньовенного, внутрішньом'язового, 59 підшкірного, парентерального, спінального або епідермального призначення (наприклад, з допомогою ін'єкції або інфузії). Залежно від способу призначення, активна сполука, а саме антитіло, біспецифічна і мультиспецифічна молекула, може бути покрита матеріалом для захисту сполуки від дії кислот та інших природних факторів, які можуть дезактивувати сполуку. Термін "фармацевтично прийнятна сіль" вживається на позначення солі, яка зберігає бажану біологічну активність вихідної сполуки і не викликає жодних небажаних токсикологічних ефектів (див., наприклад, Berge, S.M., et al. (1977) J. Pharm. Sci. 66:1-19). Приклади таких солей включають кислотно-адитивні солі і солі приєднання основ. 5 Кислотно-адитивні солі включають солі, похідні від нетоксичних неорганічних кислот, таких як хлорводнева кислота, азотна, фосфорна, сірчана, бромводнева, йодводнева, фосфорна кислота і подібні, а також нетоксичних органічних кислот, таких як аліфатичні моно- і дикарбонові кислоти, феніл-заміщені алканові кислоти, гідроксиалканові кислоти, ароматичні кислоти, аліфатичні і ароматичні сульфонові кислоти і подібні. Солі приєднання основ включають солі, похідні від лужноземельних металів, таких як натрій, калій, магній, кальцій і подібні, а також від нетоксичних органічних амінів, таких як N,N'-дибензилетилендіамін, Nметилглюкамін, хлорпрокаїн, холін, діетаноламін, етилендіамін, прокаїн і подібні. Композиція згідно з винаходом може призначатися з допомогою ряду методик, відомих з рівня техніки. Як відомо фахівцеві у цій галузі, шлях і/або спосіб призначення відрізняється залежно від бажаних результатів. Активні сполуки можуть бути одержані разом з носієм, який захищатиме їх від швидкого вивільнення, а саме у вигляді композицій контрольованого вивільнення, включно з імплантатами, трансдермальними пластирями і мікрокапсулбованими системами доставки. Можуть застосовуватись біорозкладні біосумісні полімери, такі як етиленвінілацетат, поліангідриди, полігліколева кислота, колаген, поліортоестери і полімолочна кислота. Методики приготування таких композицій загалом відомі фахівцям. Див., наприклад, Sustained і Controlled Release Drug Delivery Systems (Системи доставки ліків пролонгованого і контрольованого вивільнення), J.R. Robinson, ed, Marcel Dekker, Inc., New York, 1978. Для призначення сполуки згідно з винаходом певним способом може бути необхідним покрити сполуку або призначати одночасно з матеріалом для запобігання її дезактивації. Наприклад, така сполука може призначатися суб'єкту у відповідному носії, наприклад, ліпосомах або розбавнику. Фармацевтично прийнятні розбавники включають сольові і водні забуферені розчини. Ліпосоми включають емульсії типу вода- у- маслі- у-воді CGF, а також традиційні ліпосоми (Strejan etal. (1984) J. Neuroimmunol. 7:27). Фармацевтично прийнятні носії включають стерильні водні розчини або дисперсії і стерильні порошки для негайного приготування стерильного розчину для ін'єкцій або дисперсії. Використання такого середовища і агентів для фармацевтично 93478 60 активних речовин відоме із рівня техніки. За винятком лише тих випадків, коли будь-яке традиційне середовище або агент є несумісними з активною сполукою, їх застосування у фармацевтичних композиціях згідно з винаходом передбачене. У композиції також можуть бути включені додаткові активні сполуки. Терапевтичні композиції зазвичай повинні бути стерильними і стійкими в умовах виготовлення і зберігання. Композиція може бути приготована у вигляді розчину, мікроемульсії, ліпосом або іншої придатної форми для високої концентрації лікарського засобу. Носієм може бути розчинним або дисперсійне середовище, яке містить, наприклад, воду, етанол, поліол (наприклад, гліцерин, пропіленгліколь і рідкий поліетиленгліколь та подібні), і їх придатні суміші. Необхідна текучість може підтримуватись, наприклад, за рахунок використання покриття, такого як лецитин, за рахунок підтримання необхідного розміру частинок у випадку дисперсії і завдяки використанню поверхневоактивних сполук. У багатьох випадках переважним було б включення у композицію ізотонічних агентів, наприклад цукру, поліспиртів, таких як манітол, сорбітол або хлорид натрію. Пролонгована абсорбція композицій для ін'єкцій може бути досягнута за рахунок включення у композицію агента, який уповільнює абсорбцію, наприклад моностеаратів і желатину. Стерильні розчини для ін'єкцій можуть бути одержані додаванням активної сполуки у необхідній кількості у відповідний розчинник з одним або більше вищеназваними інгредієнтами за необхідністю з наступним мікрофільтруванням для стерилізації. Зазвичай дисперсії одержують додаванням активної сполуки у стерильний носій, який містить основне середовище для дисперсії та інші необхідні інгредієнти із вищеназваних. У випадку стерильних порошків для приготування стерильних розчинів для ін'єкцій переважними методиками приготування є вакуумне висушування і висушування заморожуванням (ліофілізація), з допомогою яких одержують порошок активного інгредієнта, а також будь-якого іншого бажаного інгредієнта із попередньо відфільтрованого стерилізованого розчину. Режими дозування підбираються таким чином, щоб забезпечити оптимальну бажану реакцію (наприклад, терапевтичний ефект). Наприклад, може призначатися один болюс, може призначатися декілька розділених доз протягом певного часу або доза може пропорційно збільшуватись або зменшуватись за вимогою певної терапевтичної ситуації. Особливо переважним для парентеральних композицій є їх виготовлення у вигляді дозованої лікарської форми для спрощення призначення та кратності доз. Термін дозована лікарська форма у контексті цього винаходу вживається на позначення фізично дискретних одиниць, придатних для застосування у вигляді одиничних доз для суб'єктів, які проходять лікування. Кожна одинична дозована форма містить задану кількість активної сполуки, яка має забезпечувати бажаний терапевтичний ефект, у поєднанні з необхідним фармацевтичним носієм. Характеристики дозованих лікар
ДивитисяДодаткова інформація
Назва патенту англійськоюIsolated human monoclonal antibody which binds to human cd20 and use thereof
Автори англійськоюTeeling Jessica, Ruuls Sigrid, Glennie Martin, Van De Winkel Jan G.J., Parren Paul, Petersen Joergen, Baadsgaard Ole D.M.Sc., Huang Haichun
Назва патенту російськоюВыделенное моноклональное антитело человека, kotopoe связывается c cd20, и его применение
Автори російськоюТилинг Джесика, Руулс Зигрид, Гленни Мартин, Ван де Винкель Ян Г.Й., Паррен Пауль, Петерсен Йорген, Баадсгаард Оле Д.M.C., Хуанг Хайчун
МПК / Мітки
МПК: A61P 35/00, C07K 16/28, A61K 39/395, C07K 16/42
Мітки: сd20, виділене, яке, зв'язується, людини, застосування, моноклональне, антитіло
Код посилання
<a href="https://ua.patents.su/98-93478-vidilene-monoklonalne-antitilo-lyudini-yake-zvyazuehtsya-z-sd20-ta-jjogo-zastosuvannya.html" target="_blank" rel="follow" title="База патентів України">Виділене моноклональне антитіло людини, яке зв’язується з сd20, та його застосування</a>
Моноклональне антитіло, яке спричиняє апоптоз кров’яних клітин, що містять ядро та інтегрин-пов’язаний протеїн людини (іпп), фрагмент моноклонального антитіла, гібридома, яка продукує моноклональне антитіло (ва

Номер патенту: 72192
Опубліковано: 15.02.2005
Автори: Уно Шінсуке, Фукушіма Наоші
МПК: C07K 16/28, C12P 21/08, C12N 5/20
Мітки: моноклонального, гібридома, клітин, людини, ядро, ва, іпп, яка, антитіло, спричиняє, моноклональне, яке, кров'яних, апоптоз, антитіла, продукує, фрагмент, інтегрин-пов'язаний, містять, протеїн
Формула / Реферат:
1. Моноклональне антитіло, яке зв’язується із інтегрин-пов’язаним протеїном людини (ІПП) і спричиняє апоптоз кров'яних клітин, що містять ядро та ІПП.2. Моноклональне антитіло за п.1, яке відрізняється тим, що антитіло отримують способом, що включає:а) імунізацію тварин з використанням ІПП людини як сенсибілізуючого антигена,б) одержання гібридом;в) відбір антитіл, що розпізнають ІПП людини та спричиняють апоптоз...
Виділене моноклональне антитіло людини, яке специфічно зв’язується з cd25 людини та інгібує зв’язування il-2 з сd25

Номер патенту: 90082
Опубліковано: 12.04.2010
Автори: Уільямс Деніс Лі, Шуурман Янін, Паррен Пауль, Ван де Вінкель Ян Г.Й., Петерсен Йорген, Хавеніт Катаріна Емануель Герарда, Баадсгаард Оле Д.М.С.
МПК: A61K 51/10, A61P 7/00, A61P 35/00, A61K 39/395, C07K 16/28, G01N 33/577
Мітки: людини, моноклональне, інгібує, зв'язується, антитіло, виділене, яке, специфічно, зв'язування, сd25
Формула / Реферат:
1. Виділене моноклональне антитіло людини, яке специфічно зв'язується з СD25 людини та інгібує зв’язування IL-2 з CD25, вибране з групи, що містить:а) варіабельну ділянку важкого ланцюга, представлену в SEQ ID NO: 2, та варіабельну ділянку легкого ланцюга, представлену в SEQ ID NO: 4;b) варіабельну ділянку важкого ланцюга, представлену в SEQ ID NO: 6, та варіабельну ділянку легкого ланцюга, представлену в SEQ ID NO: 8;с)...
Людське моноклональне антитіло, що специфічно зв’язується з cd40, фармацевтична композиція, що його містить, та застосування антитіла для виробництва лікарського засобу для лікування cd40-негативної пухлини у людини

Номер патенту: 84539
Опубліковано: 10.11.2008
Автори: Фенг Ксіао, Джіа Ксіао-Чі, Бедіан Вахе, Корвалан Хосе, Глейдью Роналд П.
МПК: C12N 15/13, A61K 39/395, C07K 16/28, C12N 5/20, C12N 15/63, A61P 35/00, A61P 37/00
Мітки: cd40, лікування, виробництва, людське, cd40-негативної, специфічно, містить, композиція, моноклональне, засобу, пухлини, лікарського, застосування, фармацевтична, антитіло, антитіла, зв'язується, людини
Формула / Реферат:
1. Людське моноклональне антитіло або його антигензв’язувальна частина, які специфічно зв’язуються з людським CD40, причому вказане антитіло або його антигензв’язувальна ділянка є агоністом CD40 і де антитіло містить важкий ланцюг і легкий ланцюг, і при цьому амінокислотні послідовності CDR1, CDR2 і CDR3 вказаного важкого ланцюга і CDR1, CDR2 і CDR3 вказаного легкого ланцюга вибрані з групи, яка складається з(a) амінокислотних...
Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент до рецептора ліпопротеїнів низької щільності (лпнщ) людини, спосіб їх одержання

Номер патенту: 78489
Опубліковано: 10.04.2007
Автори: Бельзер Ілана, Дреано Мішель, Смоларскі Моше, Йонах Начум, Суісса Дані, Антонетті Франческо
МПК: C12P 21/08, C07K 16/28, G01N 33/68, G01N 33/577, C12N 5/20
Мітки: ліпопротеїнів, повністю, одержання, фрагмент, антиідіотипічного, моноклональне, людини, химерне, рецептора, антитіло, гуманізоване, щільності, лпнщ, антитіла, низької, спосіб
Формула / Реферат:
1. Моноклональне антитіло, химерне антитіло, повністю гуманізоване антитіло, антитіло до антиідіотипічного антитіла або його фрагмент, що специфічно розпізнає і зв'язує розчинний рецептор ліпопротеїнів низької щільності (ЛПНЩ)людини і його фрагменти, а також здатне інгібувати реплікацію вірусу гепатиту С, і яке одержують шляхом імунізації тварин формою +291 розчинного рецептора ЛПНЩ людини, тобто формою розчинного рецептора людини, що містить...
Виділене антитіло, яке специфічно зв’язує gdf-8, та його застосування

Номер патенту: 91815
Опубліковано: 10.09.2010
Автори: Фельдман Гейтруіда М., Вольфман Нейл М., Сонг Кенінг, Гроув-Бріджес Крісті, Девіс Монік В., Філд Енн, Расселл Керолайн, Вальге-Архер Фія
МПК: A61K 39/395, A61P 3/00, A61P 21/00, C07K 16/22
Мітки: виділене, специфічно, яке, gdf-8, застосування, антитіло, зв'язує
Формула / Реферат:
1. Виділене антитіло, яке специфічно зв'язує GDF-8, що містить щонайменше одну амінокислотну послідовність, вибрану із групи, що складається з:a) SEQ ID NO:14;b) SEQ ID NO:26;c) амінокислотної послідовності, що містить SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:35 і SEQ ID NO:36, де одна або декілька амінокислот у SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:33, SEQ ID NO:34, SEQ ID NO:35 або SEQ ID NO:36...
Попередній патент: Спосіб діагностики депресії
Наступний патент: Фармацевтичний алергенний препарат
Випадковий патент: Спосіб фунгіцидної і бактерицидної обробки грени дубового шовкопряда