Спосіб визначення ферментативної активності пероксидази рослин
Номер патенту: 82489
Опубліковано: 12.08.2013






Формула / Реферат
Спосіб визначення ферментативної активності пероксидази рослин, що містить приготування реакційної суміші з використанням субстрату-відновника (3,3',5,5'-тетраметилбензидину дигідрохлориду моногідрату), субстрату-окислювача (перекису водню), ферментативного препарату, буферного розчину, реєстрацію спектрофотометричним методом значень оптичної густини при довжині хвилі 650 нм, визначення концентрації білка в ферментативному препараті, розрахунок питомої активності пероксидази, який відрізняється тим, що перед приготуванням реакційної суміші проводять приготування неочищених тканинних ферментативних екстрактів цитоплазматичної та іонозв'язаної пероксидази, які використовують як ферментативні препарати, для приготування субстрату-окислювача використовують гідроперит, готують два цитратно-фосфатних буферних розчини з рН 4,8 з концентрацією 0,05 М і 0,2 М, і в другий розчин додатково вводять хлорид калію в кількості 1 М, причому концентрацію білка встановлюють методом Bredford з Coomassi Briliant Blue G250.
Текст
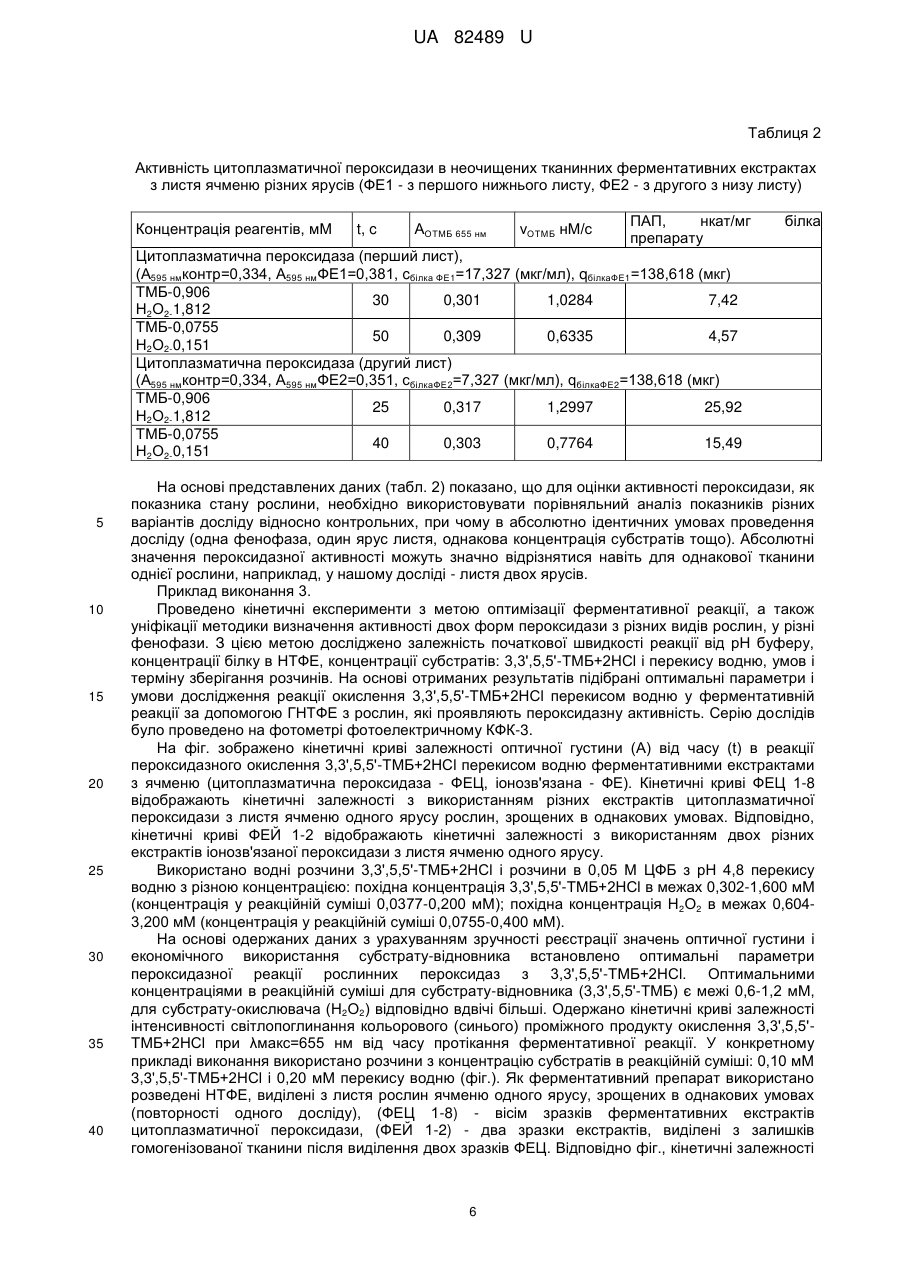
Реферат: UA 82489 U UA 82489 U 5 10 15 20 25 30 35 40 45 50 55 60 Корисна модель належить до біохімічної області і може бути використана у дослідженнях з фізіології та біохімії рослин, в мікології, екології, у галузях сільського і лісового господарства, а також в інших сферах науково-практичної діяльності, що потребують визначення активності пероксидази рослин для моніторингу їх стану. Пероксидаза (КФ.1.11.17.; донор:Н2О2-оксидоредуктаза) є індуцибельним ферментом, індукторами якого можуть бути різноманітні фізичні, хімічні і біологічні чинники. Експериментальні дані різних наукових шкіл свідчать про те, що пероксидаза пов'язана з багатьма важливими метаболічними перетвореннями [1]. Відомо, що активність ферменту зростає при багатьох змінах і порушеннях метаболізму рослин, а деякі її ізоензими у відповідь на стрес синтезуються de novo. Пероксидаза присутня у різних компартментах клітини і є важливим агентом окислювально-відновної системи [2], відіграє провідну роль у процесах детоксикації агресивних форм активного кисню [3, 4], диханні [5] і формуванні захисної реакції організму до дії широкого спектру біогенних і абіогенних стресорів [4-11]. Загальновідомо, що оксидоредуктази, зокрема пероксидази, грають важливу роль у процесі адаптації. Рівень активності ферменту є інформативним показником формування стійкості до різних стресорів. Вивчено роль і механізми участі пероксидази у формуванні "окислювального вибуху" [4, 6, 7, 11]. Досліджено роль пероксидази в метаболічних перетвореннях [9, 10, 12]. Таким чином, активність пероксидази, зміни ізоферментного складу, кількісних характеристик і активності різних форм і ізоформ ферменту під впливом різноманітних чинників є інформативним показником стану будь-якого організму, зокрема рослинного. Пероксидаза є індуцибельним і найбільш філогенетично старим ферментом [12], має широку субстратну специфічність [1, 2, 12], що підкреслює її значущу роль у метаболізмі, в підтриманні гомеостазу, в адаптації і життєдіяльності в цілому. Крім того, пероксидазна реакція окиснення різних субстратів, які мають окрашені продукти, широко використовується як маркер в імуноферментному каталізі. Також ця реакція є модельною у каталізі при вивченні інгібіторів тощо. Дотепер на рослинних об'єктах (неочищених і очищених препаратах) широко застосовується швидкий метод визначення пероксидазної активості з субстратом бензидином (метод A.M. Бояркина) [13], а також різні його модифікації [14, 15]. Головний недолік цих способів канцерогенність бензидину. Використовуються також інші фенольні і аміно-фенольні похідні як субстрати, продукти окислення яких мають максимум поглинання у видимій області: пірогалол, гваякол, о-дианізидин, о-фенілендиамін та інші [16, 17]. Головні недоліки цих способів неприємний запах і нестабільність при зберіганні гваякола, токсичність о-дианізидину, офенілендиаміну [15]. У зв'язку з цим було вирішено задачу корисної моделі більш безпечного і чутливого субстрату ферментативної пероксидазної реакції окислення пероксиду водню 3,3',5,5'-тетраметилбензидину (3,3',5,5'-ТМБ), який є не канцерогенною і не мутагенною похідною бензидину за рахунок метилювання [18, 19]. Але препарат є більш дорогим, ніж бензидин. Широко використовується у дослідницькій роботі з каталізу, у криміналістиці (виявлення крові), медицині (біохімія людини, тварин, імуногістохімія), імуноферментному аналізі, кількісному визначенні антиоксидантів тощо. Головний недолік 3,3',5,5'-ТМБ - погана розчинність у воді та буферах, необхідних для одержання ферментативного екстракту, тому похідний розчин 3,3',5,5'-ТМБ) готують з 0,2 М НСl. Переваги має водорозчинна похідна тетраметилбензидину - 3,3',5,5'-тетраметилбензидин дигідрохлорид гідрат (моногідрат або дигідрат) (3,3',5,5'-ТМБ+2HСІ). Цей субстрат широко використовується як у теоретичних дослідженнях так і для практичних цілей: кінетичні дослідження [15], медична біохімія [19], імуноферментний аналіз [16]. Використання 3,3',5,5'-ТМБ+2НСl підвищує чутливість реакції у чотири рази в порівнянні з бензидином [18]. Однак, не дивлячись на велику кількість досліджень з використанням 3,3',5,5'-ТМБ+2HСl як субстрату для визначення пероксидазної активності, в літературі не представлено даних для визначення активності рослинних пероксидаз у неочищеному тканинному ферментативному екстракті з даним субстратом. На даний час доведено, що завдяки швидкості та інформативності методики, оцінка активності рослинних пероксидаз саме в неочищеному тканинному екстракті широко затребувана для теоретичних і прикладних досліджень у багатьох науко-практичних напрямах: біохімії, екології, сільському і лісовому господарстві, рекультивації тощо. Найбільш близьким по технічній суті та результату, який досягається, є спосіб визначення активності пероксидази спектрофоториметричним методом на основі визначення швидкості утворення кольорового (синього) продукту окислення субстрату пероксидази 3,3',5,5'-ТМБ+2НСl, який має максимум світлопоглинання λ=655 нм і високий молярний коефіцієнт світлопоглинання -1 -1 ε655=39000 М см [20]. Недоліком прототипу є те, що він використовується для очищених ферментативних препаратів (пероксидаза хрону та інших) і не використовується для визначення активності пероксидази в неочищених ферментативних тканинних екстрактах з 1 UA 82489 U 5 10 15 20 25 30 35 рослин, що є швидким і інформативним методом моніторингу стану рослин за умов дії різних біогенних і абіогенних чинників. Іншим недоліком є те, що як субстрат у реакції пероксидазного окислення використовують 3,3',5,5'-ТМБ+2НСl фірми "Sigma" США, який є високо вартісним, тому не використовується широко у практиці сільськогосподарських та інших досліджень на рослинах. В основу корисної моделі поставлено задачу удосконалення способу визначення ферментативної активності пероксидази рослин у неочищених тканинних ферментативних екстрактах (НТФЕ) з використанням 3,3',5,5'-ТМБ+2НСl виробника "Вітротест" (Україна) як субстрату-відновника, що дає можливість економічно, швидко і з високою точністю отримувати інформативні показники, які можуть бути використані у різних галузях науково-практичних досліджень: біохімічних, біологічних, екологічних та сільськогосподарських. Поставлена задача вирішується за рахунок того, що спосіб визначення ферментативної активності пероксидази рослин, який містить приготування реакційної суміші з використанням субстрату-відновника (3,3',5,5'-тетраметилбензидину дигідрохлориду моногідрату), субстратуокислювача (перекису водню), ферментативного препарату, буферного розчину, реєстрацію спектрофотометричним методом значень оптичної густини при довжині хвилі 655 нм, визначення концентрації білку в ферментативному препараті, розрахунок питомої активності пероксидази, згідно з корисною моделлю, перед приготуванням реакційної суміші включає приготування неочищених тканинних ферментативних екстрактів цитоплазматичної та іонозв'язаної пероксидази, які використовують як ферментативні препарати, для приготування субстрату-окислювача використовують гідроперит, готують два цитратно-фосфатних буферних розчини з рН 4,8 з концентрацією 0,05 М і 0,2 М, і в другий розчин додатково вводять хлорид калію в кількості 1 М, причому концентрацію білку встановлюють методом Bredford з Coomassi Briliant Blue G 250. Пропонована методика є новою тому, що вона невідома з об'єкта дослідження: пара фермент+субстрат-відновник, в якій ферментативним препаратом є неочищений ферментативний екстракт з рослинної тканини, а субстратом-відновником є 3,3',5,5'тетраметилбензидин дигідрохлорид моногідрат. Крім того, з метою підвищення економічності методики використано субстрат пероксидазної реакції 3,3',5,5'-ТМБ+2НСl вітчизняного виробника "Вітротест" (Україна), який є менш вартісним, ніж імпортні реактиви фірм "Sigma" США і "Serva" Німеччина. Зручність методики полягає також у тому, що вона не потребує складного обладнання і може бути проведена з використанням не тільки спектрофотометрів (СФ з ультрафіолетовою і видимою областями), але і фотометра фотоелектричного КФК-3. Субстрат-відновник: 3,3',5,5'-тетраметилбензидин дигідрохлорид моногідрат, C16H20N22НСlН2О, М 331,3. Утворення проміжного продукту окислення 3,3',5,5'-тетраметилбензидину синього кольору з максимумом світлопоглинання при λ=652 нм представлене схематично [21]. 40 Для ідентифікації, оцінки чистоти, впливу умов і терміну зберігання реактивів і препаратів, а також придатності запропонованого способу визначення пероксидазної активності за технологічними можливостями з ціллю забезпечення достовірних результатів за відомими 2 UA 82489 U 5 10 15 20 25 30 35 40 45 50 55 60 методиками проведено дослідження спектральних характеристик субстрату-відновника (3,3',5,5'-тетраметилбензидину дигідрохлориду моногідрату (ТМБ) фірми "Вітротест" (Україна), субстрату-окислювача (перекису водню), ферментативних препаратів (НТФЕ - ферментативних екстрактів цитоплазматичної (ФЕЦ) та іонозв'язаної (ФЕЙ) пероксидази з тканини рослин, одержаних за методикою К.З. Гамбурга [14] з модифікацією), стандартних білкових препаратів (СТБ) (бичачого сироваткового альбуміну (БСА), альбуміну людини). Спектрофотометричні дослідження проводили на спектрофотометрах СФ-26 і СФ-2000, а також на фотометрі фотоелектричному КФК-3. Одержані дані свідчать про ідентичність спектральних характеристик 3,3',5,5'-ТМБ+2НСl виробника "Вітротест" (Україна) зарубіжним аналогам фірм "Sigma" (США) та "Serva" (Німеччина). Збереження розчинів 3,3',5,5'-ТМБ+2НСl на протязі доби у посуді з темного скла приводить до утворення забарвлених продуктів окислення синього кольору. Це встановлено спектрофотометричним методом на підставі зменшення оптичної густини розчинів 3,3',5,5'-ТМБ при λ=285 нм, і зростання (від нульового значення) оптичної густини у максимумі світлопоглинання продукту окислення ТМБ при λ=655 нм. Для одержання коректних результатів необхідно готувати розчин 3,3',5,5'-ТМБ+2НСl у день проведення експерименту. Треба також +3 враховувати, що присутність іонів заліза (Fe ) у дистильованій воді прискорює окислення 3,3,5,5'-ТМБ, тому для приготування розчинів краще використовувати бідистильовану воду. Спектрофотометричним методом оцінено вплив агрегатного стану використаного реагенту субстрату-окислювача (твердий гідроперит і розчин пергідролю), а також терміну зберігання на концентрацію розчину перекису водню. Початковий 0,5 %-ний розчин перекису водню готували з гідропериту (вага таблетки 1,5 г містить Н2О2 приблизно 33 % вагових). Робочий розчин з концентрацією (1,47 мМ), готували в 0,05 М цитратно-фосфатному буфері (ЦФБ) з рН 4,8. = -1 -1 Концентрацію встановлювали спектрофотометричним методом при λ=230 нм (ε 230 72,4 М см ) [15]. Збереження розчинів перекису водню на протязі доби приводить до зменшення концентрації реагенту, тому рекомендовано готувати розчин перекису водню також в день експерименту. Встановлено спектрофотометричним методом, що НТФЕ з листя ячменю двох форм пероксидази - цитоплазматичної та іонозв'язаної - мають різні спектральні характеристики за низкою причин (структурно-хімічні відмінності двох форм пероксидази, різний склад і вміст в екстрактах високо і низькомолекулярних компонентів рослинної тканини тощо). У зв'язку з цим спектрофотометричний метод визначення білку в УФ області (λ mах=280 нм), а також у максимумі полоси Соре з використанням молярного коефіцієнта світлопоглинання очищеного ферментативного білку [20], не придатний для НТФЕ, оскільки не забезпечує точність результатів. Для розрахунку питомої активності пероксидази (ПАП) було визначено кількість білку в НТФЕ з рослинної тканини методом Bredford [22, 23] с Coomassi Briliant Blue G 250 фірми "Sigma". Метод дозволяє визначати білок у мікрокількостях (від 2 мкг/мл до 120 мкг/мл). Цей метод є більш точним і коректним для досліджування неочищених ферментативних екстрактів з тканини рослин у порівнянні з методом Lowry et al. [24], який є дуже чутливим до присутності у розчині білку сторонніх відновників, наприклад, фенольних сполук, які містяться в рослинних тканинних екстрактах. Як стандартний білок було використано бичачий сировотковий альбумін (БСА) і альбумін людини (АЛ). З метою підвищення точності доцільно використовувати твердий ліофільно висушений БСА. На підставі одержаних даних було побудовано градуювальний графік, представлена формула розрахунку кількості білка у ферментативних екстрактах. Приклад конкретного виконання 1 Проведено визначення ферментативної активності двох форм пероксидази: цитоплазматичної і іонозв'язаної, виділених із рослинної тканини за методикою, яка представляє модифікацію методики К.З. Гамбурга з співавторами [14]. Використано цитратнофосфатний буфер (ЦФБ) з рН в межах 4,7-5,2, який широко використовується для екстрагування пероксидаз і найбільш придатний у роботі з ТМБ [19, 20], замість ацетатного буферу, необхідного для роботи з бензидином як субстрат. Як ферментативні препарати використовували неочищені тканинні ферментативні екстракти (НТФЕ або скорочено ФЕ), які проявляли пероксидазну активність: ферментативні екстракти цитоплазматичної пероксидази (ФЕЦ), ферментативні екстракти іонозв'язаної пероксидази (ФЕЙ). Приготування НТФЕ цитоплазматичної та іонозв'язаної пероксидази з рослинної тканини проводили за модифікованою методикою К.3. Гамбурга [14]. Для приготування ферментативного екстракту цитоплазматичної пероксидази (ФЕЦ) брали 0,5 г або 1 г рослинної 3 UA 82489 U 5 10 15 20 25 30 35 40 45 50 55 60 тканини, яку гомогенізували при t=+2-4 °C у фарфоровій ступці з тертим склом з ЦФБ з низькою іонною силою (0,05 М ЦФБ), витримували 3 хв у холодильнику, відділяли від твердого залишку центрифугуванням (8 тис. об/хв протягом 5 хв). Повторювали стадії екстракції і центрифугування 5 разів. Загальний об'єм ФЕЦ після п'ятьох центрифугувань складав 25 мл або 50 мл. Кількість рослинної тканини і об'єми екстракту підбирали індивідуально в залежності від виду рослини та фенофази (залежно від щільності тканини потрібна різна кількість тертого скла для гомогенізації тканини). Після виділення цитоплазматичної пероксидази до залишку гомогенізованої тканини додавали 3-5 мл ЦФБ з високою іонною силою (0,2 М ЦФБ з 1 М КСl). Подальшим екстрагуванням і центрифугуванням виділяли іонозв'язану пероксидазу. Загальний об'єм (ФЕЙ) після двох центрифугувань складав 10 мл або 25 мл. Для приготування реакційної суміші використовували ФЕ з різною концентрацією білку, які одержано шляхом розведення відповідним буфером. Вивчено вплив рН ЦФБ на кількість білків при екстрагуванні, які проявляли пероксидазну активність, а також на швидкість ферментативної реакції. Встановлено, що оптимальними є значення рН у межах 4,7-5,2. При значеннях рН нижче і вище вказаного діапазону швидкість пероксидазної реакції знижується. У нашому досліді використано ЦФБ з рН 4,8. Реакцію проводили при t=+22-25 °C в кюветі з довжиною кювети (l) l=2 см. Реакційна суміш загальним об'ємом 8 мл вміщувала 1 мл ФЕ (ФЕЦ або ФЕЙ), 1 мл 3,3',5,5'-ТМБ+2НСl, 5 мл ЦФБ і 1 мл перекису водню. В розчин порівняння (контроль) замість Н 2О2 додавали 1 мл ЦФБ. Спочатку в кювету вносили ФЕ, ТМБ і ЦФБ перемішували, витримували 2 хвилини. Потім додавали Н2О2 одночасно увімкнувши секундомір. Якщо замість дозатору використовують піпетку, вона повинна мати широкий отвір для швидкого витікання рідини і перемішування суміші. Виміри оптичної густини проводили на КФК-3 при λ=655 нм двома способами: перший фіксували час досягнення розчином значень оптичної густини (А) 0,300±0,02 або 0,350±0,02; другий - фіксували значення А через 10 с від додавання розчину Н 2О2 до реакційної суміші з інтервалом 5 с до 45 с. Концентрацію ФЕ підбирали шляхом розведення таким чином, щоб вищевказані значення оптичної густини досягалися за період 15-45 секунд. Для розрахунку питомої активності цитоплазматичної та іонозв'язаної пероксидази (ПАФЕЦ, ПАФЕЙ) визначали концентрацію білку у розчинах ФЕЦ і ФЕЙ, які було безпосередньо використано для приготування реакційної суміші, методом Bredford [22, 23] с Coomassi Briliant Blue G 250. Розрахунок проводили за виведеною формулою, використовуючи показники градуювальника за стандартним білком (СТБ) БСА. Розраховували показники питомої активності пероксидази (ПАП): цитоплазматичної (ПАФЕЦ) і іонозв'язаної (ПАФЕЙ) пероксидази за формулами (1 або 2). Формула (1) виведена на підставі формули (2), в якій для спрощення користування введено константи (для представлених умов проведення дослідів). ПАП=А655 нм ОТМБ/PR (А595 нмФЕ-А595 нм контр)t, (нкат/мг б-ку ФЕ), де (1) А655 нм ОТМБ - оптична густина реакційної суміші через час t від початку реакції; t - час досягнення реєстуємого значення А655 нм ОТМБ, (с); -1 R - константа, R=ε655 нм ОТМБ1, (М ), де (1а) -1 -1 ε655 нм ОТМБ=39 000 (М см ), молярний коефіцієнт світлопоглинання ОТМБ; 1 - довжина кювети, (см); Р - константа при обчисленні білку за градуювальником залежності оптичної густини А (за віссю ординат (У)) від концентрації білку (мкг/мл) за віссю абсцис (X). Р=(ціну поділки X)/tg α ціна поділки У, де (1б) tg α=(А595 нм СТБ-А595 нм контр) ціну поділки X/сСТБ ціна поділки У, де (1в) сСТБ - концентрація стандартного білку, (мкг/мл); А595 нм СТБ - оптична густина розчину стандартного білку з сСТБ; А595 нм контр - оптична густина контролю. Питома пероксидазна активність (ПАП, або ПАФЕЦ і ПАФЕЙ для цитоплазматичних і іонозв'язаних ферментативних екстрактів) є відношенням швидкості утворення проміжного продукту окислення ТМБ до кількості білку ФЕ (qФЕ) у реакційній суміші: ПАП=νОТМБ/qФЕ=А655 нм OTMБV/εОТМБ1t сФЕV=А655 нм ОТМБ/εОТМБ1t cФЕ, (нкат/мг б-ку ФЕ), де (2) νОТМБ=cОТМБ V/t=АОТМБ 655 нм V/εОТМБ1t, (моль/с), де (2а) νОТМБ - початкова швидкість реакції, (моль/с); V - об'єм реакційної суміші в кюветі, (л); t - час досягнення реєстуємого значення А655 нм ОТМБ, (с); cОТМБ=АОТМБ 655 нм/ε655 нм ОТМБ1 (М), де (2б) cОТМБ - концентрація ОТМБ у кюветі (М); А655 нм ОТМБ - оптична густина реакційної суміші через час t від початку реакції; 4 UA 82489 U -1 5 10 15 20 -1 ε655 нм ОТМБ=39000 М см ), молярний коефіцієнт світлопоглинання ОТМБ; 1 - довжина кювети (см); qФЕ=сФЕV, (мг), де (2в) qОТМБ - кількість речовини (ОТМБ) у кюветі в момент реєстрації значення А595 нмФЕ; V - об'єм реакційної суміші в кюветі, (л); cФЕ=(А595 нмФЕ-А595 ммконтр)ціну под-ки X/tgαціна под-ки У, (мкг/мл), де (2 г) А595 нмФЕ - оптична густина розчину ФЕЦ або ФЕЙ; А595 нмконтр - оптична густина контролю; tg α=(А595 нм СТБ-А595 нм контр) ціну поділки X/сСТБ ціна поділки У, де (1в) сСТБ - концентрація стандартного білку, (мкг/мл); А595 нм СТБ - оптична густина розчину стандартного білку з с СТБ; А595 нмконтр - оптична густина контролю. За Міжнародною класифікацією активність ферментів оцінюється в одиницях Е: 1Е=1 мкмоль субстрату (продукту окислення) / 1 хв За одиницю активності в ферментології також прийнято 1 катал - 1 одиниця ферменту, що каталізує реакцію зі швидкістю 1 моль/сек. 7 6 1 катал-1 кат=610 Е (або 6010 Е) Для виведеної формули коректно первісну швидкість виражати у нкат. Приклад конкретного виконання визначення питомої активності пероксидази ПАФЕЦ і ПАФЕЙ за експериментальними і розрахунковими даними за виведеними формулами представлено в таблиці 1. Таблиця 1 Активність цитоплазматичної та іонозв'язаної пероксидази в неочищених тканинних ферментативних екстрактах з листя пшениці Концентрація ТМБ, г/л t, c АОТМБ 655 нм νОТМБ нМ/с ПАП, нкат/мг білка препарату Цитоплазматична пероксидаза (А595 нмконтр=3,339, А595 нмФЕ=0,360, сбілка ФЕ, =7,742 (мкг/мл), qбілка ФЕ, =61,936 (мкг) 0,5 13 0,310 2,4461 39,49 0,4 15 0,308 2,1047 33,98 0,3 16 0,298 1,9091 30,82 0,2 20 0,305 1,5631 25,24 0,1 29 0,306 1,0816 17,46 Іонозв'язана пероксидаза (А595 нмконтр=0,339, А595 нм ФЕ=,0,379, cбілка ФЕ, =14,747 (мкг/мл), qбілка ФЕ, =117,972 (мкг) 0,5 15 0,308 2,1047 17,84 0,4 25 0,282 1,1562 9,80 0,3 30 0,302 1,0318 8,75 25 Приклад конкретного виконання 2. Проведено визначення активності цитоплазматичної пероксидази в НТФЕ з листя ячменю першого і другого ярусів з метою з'ясувати, чи існує достовірна різниця у досліджуваному показнику при використанні субстрату-відновнику 3,3',5,5'-ТМБ+2НСl за запропонованою методикою. 5 UA 82489 U Таблиця 2 Активність цитоплазматичної пероксидази в неочищених тканинних ферментативних екстрактах з листя ячменю різних ярусів (ФЕ1 - з першого нижнього листу, ФЕ2 - з другого з низу листу) Концентрація реагентів, мМ t, c АОТМБ 655 нм νОТМБ нМ/с ПАП, нкат/мг препарату білка Цитоплазматична пероксидаза (перший лист), (А595 нмконтр=0,334, А595 нмФЕ1=0,381, cбілка ФЕ1=17,327 (мкг/мл), qбілкаФЕ1=138,618 (мкг) ТМБ-0,906 30 0,301 1,0284 7,42 Н2О2-1,812 ТМБ-0,0755 50 0,309 0,6335 4,57 Н2О2-0,151 Цитоплазматична пероксидаза (другий лист) (А595 нмконтр=0,334, А595 нмФЕ2=0,351, сбілкаФЕ2=7,327 (мкг/мл), qбілкаФЕ2=138,618 (мкг) ТМБ-0,906 25 0,317 1,2997 25,92 Н2О2-1,812 ТМБ-0,0755 40 0,303 0,7764 15,49 Н2О2-0,151 5 10 15 20 25 30 35 40 На основі представлених даних (табл. 2) показано, що для оцінки активності пероксидази, як показника стану рослини, необхідно використовувати порівняльний аналіз показників різних варіантів досліду відносно контрольних, при чому в абсолютно ідентичних умовах проведення досліду (одна фенофаза, один ярус листя, однакова концентрація субстратів тощо). Абсолютні значення пероксидазної активності можуть значно відрізнятися навіть для однакової тканини однієї рослини, наприклад, у нашому досліді - листя двох ярусів. Приклад виконання 3. Проведено кінетичні експерименти з метою оптимізації ферментативної реакції, а також уніфікації методики визначення активності двох форм пероксидази з різних видів рослин, у різні фенофази. З цією метою досліджено залежність початкової швидкості реакції від рН буферу, концентрації білку в НТФЕ, концентрації субстратів: 3,3',5,5'-ТМБ+2НСl і перекису водню, умов і терміну зберігання розчинів. На основі отриманих результатів підібрані оптимальні параметри і умови дослідження реакції окислення 3,3',5,5'-ТМБ+2НСl перекисом водню у ферментативній реакції за допомогою ГНТФЕ з рослин, які проявляють пероксидазну активність. Серію дослідів було проведено на фотометрі фотоелектричному КФК-3. На фіг. зображено кінетичні криві залежності оптичної густини (А) від часу (t) в реакції пероксидазного окислення 3,3',5,5'-ТМБ+2НСl перекисом водню ферментативними екстрактами з ячменю (цитоплазматична пероксидаза - ФЕЦ, іонозв'язана - ФЕ). Кінетичні криві ФЕЦ 1-8 відображають кінетичні залежності з використанням різних екстрактів цитоплазматичної пероксидази з листя ячменю одного ярусу рослин, зрощених в однакових умовах. Відповідно, кінетичні криві ФЕЙ 1-2 відображають кінетичні залежності з використанням двох різних екстрактів іонозв'язаної пероксидази з листя ячменю одного ярусу. Використано водні розчини 3,3',5,5'-ТМБ+2НСl і розчини в 0,05 М ЦФБ з рН 4,8 перекису водню з різною концентрацією: похідна концентрація 3,3',5,5'-ТМБ+2НСl в межах 0,302-1,600 мМ (концентрація у реакційній суміші 0,0377-0,200 мМ); похідна концентрація Н2О2 в межах 0,6043,200 мМ (концентрація у реакційній суміші 0,0755-0,400 мМ). На основі одержаних даних з урахуванням зручності реєстрації значень оптичної густини і економічного використання субстрату-відновника встановлено оптимальні параметри пероксидазної реакції рослинних пероксидаз з 3,3',5,5'-ТМБ+2НСl. Оптимальними концентраціями в реакційній суміші для субстрату-відновника (3,3',5,5'-ТМБ) є межі 0,6-1,2 мМ, для субстрату-окислювача (Н2О2) відповідно вдвічі більші. Одержано кінетичні криві залежності інтенсивності світлопоглинання кольорового (синього) проміжного продукту окислення 3,3',5,5'ТМБ+2НСl при λмакс=655 нм від часу протікання ферментативної реакції. У конкретному прикладі виконання використано розчини з концентрацію субстратів в реакційній суміші: 0,10 мМ 3,3',5,5'-ТМБ+2НСl і 0,20 мМ перекису водню (фіг.). Як ферментативний препарат використано розведені НТФЕ, виділені з листя рослин ячменю одного ярусу, зрощених в однакових умовах (повторності одного досліду), (ФЕЦ 1-8) - вісім зразків ферментативних екстрактів цитоплазматичної пероксидази, (ФЕЙ 1-2) - два зразки екстрактів, виділені з залишків гомогенізованої тканини після виділення двох зразків ФЕЦ. Відповідно фіг., кінетичні залежності 6 UA 82489 U 5 10 15 20 25 30 35 40 45 50 55 накопичення і розпаду проміжного продукту окислення 3,3',5,5'-ТМБ+2НСl для цитоплазматичної та іонозв'язаної пероксидази мають різний характер, однак у початковий період реакції, який використовують для розрахунку початкової швидкості реакції νO, кінетичні криві співпадають. Таким чином, заявлений спосіб визначення пероксидазної активності двох форм пероксидази (цитоплазматичної та іонозв'язаної) у неочищених тканинних ферментативних екстрактах придатний за технологічними можливостями і забезпечує коректні показники дослідження. Для отримання максимально достовірних показників для різних рослинних об'єктів необхідно підбирати концентрації білку в НТФЕ, а також концентрації субстратів, щоб значення оптичної густини 0,300-0,350 було досягнуто за 15-45 сек. Джерела інформації:, які використані при складанні заявки: 1. Газарян И.Г. Пероксидазы растений - механизм действия и белковая инженерия: дис. дра хим. наук / И.Г. Газарян. - М.:МГУ, 1996.-305 с. 2. Метелица Д.И. Моделирование окислительно-восстановительных ферментов / Метелица. Д.И. - Минск: Наука и техника, 1984.-235 с. 3. Гарифзянов А.Р. Образование и физиологические реакции активных форм кислорода в клетках растений / А.Р. Гарифзянов, Н.Н. Жуков, В.В. Иванищев // Современные проблемы науки и образования.-2011. - № 2. - С. 26-34. 4. Минибаева Ф.В. Активные формы кислорода и ионная проницаемость плазмалеммы в растительных клетках при стрессе: автореф. дис. на соиск. учен. степ, д-ра биол. наук. Минибаева Фарида Вилевна / Ф.В. Минибаева; С.-Петерб. гос. ун-т. - Санкт-Петербург.-2005.-42 с. 5. Рубин Б.А. Физиология и биохимия дыхания растений: учеб. пособие для студентов биол. специальностей ун-тов / Б.А. Рубин, М.Е. Ладыгина. - М.: Изд-во Моск. ун-та.-1974.-312 с. 6. Похомова В.М. Основные положения современной теории стресса и неспецифический адаптационный синдром у растений / Похомова В.М // Цитология - Т. 37-№ 1-2.-1975. - С. 66-91. 7. Пронина Н.Б. Экологические стрессы (причины, классификация, тестирование, физиолого-биохимические механизмы). / Пронина Н.Б // Физ., хим., биотич. Стрессы и адаптация к ним живых организмов. - Московск. С. - х. акад. им К.А. Тимирязева. - М - 2000.-310 с. 8. Изменение активности пероксидази клеток картофеля при патогенезе кольцевой гнили. / [Граскова И.А., Владимирова С.В., Колисниченко А.В. и др.] // Вісник Харківського національного аграрного університету. Серія "Біологія" - 2002. - № 9(1). - С. 37-44. 9. Виноградова Е.Н. Физиолого-биохимические реакции растений в условиях техногенно загрязненной среды в Донбассе: автореф. дис. на соискание ученой степени кандидата биологических наук.: спец. 03.00.12. / Е.Н. Виноградова; Киевский нац. ун-т имени Т. Шевченка. - Киев, 2010.-42. С. 10. Кучеренко В.П. Фермент пероксидаза і зимостійкість рослин. Монографія / В.П. Кучеренко, А.В. Капустин // Київський нац. ун-т ім. Тараса Шевченка, Ботанічний сад ім. акад… О.В. Фоміна. - Київ: Фітосоціоцентр, 2004.-116 с. 11. Вторинний оксидний стрес як елемент загальної адаптивної відповіді рослин на дію несприятливих факторів довкілля. / Н.Ю. Таран, О.А. Оканенко, Л.М. Бацманова, М.М. Мусієнко. // Физиология и биохимия культурниых растений.-2004. - Т. 36. - № 1. - С. 3-14. 12. Андреева В.А. Фермент пероксидаза. / Андреева В.А. - М.: "Наука".-1988.-128 с. 13. Бояркин А.H. Быстрый метод определения активности пероксидази / Бояркин А.Н. // Биохимия. - Т. 16, вып. 4.-1951. - С. 352-357. 14. Гамбург К.З., Подолякина Л.А., Ситнева В.М. Изучение активности пероксидазы и ИУКоксидазы в суспензионных культурах тканей табака и сои // Физиология растений. - Т. 24, вып. 3.-1977. - С. 542-547. 15. Савенкова М.И. Оптимизация окисления ароматических аминов. / Савенкова М.И., Курченко В.П., Метелица Д.И. // Изв. АН БССР. - Сер. хим. наук. 1983. - № 5. - С. 41-47. 16. Метелица Д.И. Оптимизация использования пероксидази хрена и ее антител в иммуноферментном анализе. / Метелица Д.И., Савенкова М.И., Курченко В.П // Прикладная биохимия и микробиология.-1987. - Т. 23. - вып. 1. - С. 116-124. 17. Патент на корисну модель № 38623 України, МПК А01Н 15/00. Спосіб індукції пероксидазної активності базидіоміцетів Pleurotus ostreatus (Jacq.: Fr) Kumm. та Lentinula edodes (Berk.) Sing, аскорбіновою кислотою / Федотов О.В., Волошко Т.С. (Україна); Донецький національний університет (Україна) / опубл. 12.01.2009., бюл. № 1. 18. Holland V.R. and others. A safer substitute for benzidine in the detection of blood / V.R. Holland and others / Tetrahedron. - V. 30.-1974. - P. 3299-3302. 7 UA 82489 U 5 10 15 19. Liem H.H. act. Quantitative determination of hemoglobin cytochemical staining for peroxidase using 3,3',5,5'-tetramethylbenzidine dihydrochloride, a safe substitute for benzidine / H.H. Liem act. // Analytical biochemistry.-1979. - V. 98. - P. 388-393. 20. Ингибирование пероксидазного окисления 3,3',5,5'-тетраметилбензидина аминофенолами. / Наумчик И.В., Карасева Е.И., Метелица Д.И. и др. // Биохимия.-2005. - Т. 70, вып. 3. - С. 397-405 (прототип). 21. Josephy P.D. and others. The horseradish peroxidase-catalyzed oxidation of 3,3',5,5'tetramethylbenzidine. / P.D. Josephy and others // J. of Biological chemistry.-1982. - V. 257. - № 7. P. 3669-3675. 22. Spector T. Refinement of the Coomassie Blue method of protein quantitation. / T. Spector // Analytical biochemistry.-1978. - V. 86-P. 142-146. 23. Скоупс P. Методы очистки белков / P. Скоупс. - M.: Мир, 1985.-392 с. 24. Методы биохимического исследования растений / Ермаков А.И. Арасимович В.В, Смирнова-Иконникова М.И; под ред. А.И. Ермакова. - Ленинград: Агропромиздат, 1987. - С. 107108. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 20 25 30 Спосіб визначення ферментативної активності пероксидази рослин, що містить приготування реакційної суміші з використанням субстрату-відновника (3,3',5,5'-тетраметилбензидину дигідрохлориду моногідрату), субстрату-окислювача (перекису водню), ферментативного препарату, буферного розчину, реєстрацію спектрофотометричним методом значень оптичної густини при довжині хвилі 650 нм, визначення концентрації білка в ферментативному препараті, розрахунок питомої активності пероксидази, який відрізняється тим, що перед приготуванням реакційної суміші проводять приготування неочищених тканинних ферментативних екстрактів цитоплазматичної та іонозв'язаної пероксидази, які використовують як ферментативні препарати, для приготування субстрату-окислювача використовують гідроперит, готують два цитратно-фосфатних буферних розчини з рН 4,8 з концентрацією 0,05 М і 0,2 М, і в другий розчин додатково вводять хлорид калію в кількості 1 М, причому концентрацію білка встановлюють методом Bredford з Coomassi Briliant Blue G250. Комп’ютерна верстка Д. Шеверун Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 8
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for determination of plant peroxidase enzymatic activity
Автори англійськоюRyktor Iryna Anatoliivna, Zubkova Yuliia Mykolaivna
Назва патенту російськоюСпособ определения ферментативной активности пероксидазы растений
Автори російськоюРиктор Ирина Анатольевна, Зубкова Юлия Николаевна
МПК / Мітки
МПК: C12N 9/02
Мітки: активності, пероксидази, рослин, визначення, ферментативної, спосіб
Код посилання
<a href="https://ua.patents.su/10-82489-sposib-viznachennya-fermentativno-aktivnosti-peroksidazi-roslin.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення ферментативної активності пероксидази рослин</a>
Спосіб визначення активності тоніну у біологічних рідинах

Номер патенту: 37647
Опубліковано: 16.02.2004
Автор: Самохіна Любов Михайлівна
МПК: G01N 33/48, C12Q 1/37, G01N 33/543, C12Q 1/28
Мітки: визначення, спосіб, біологічних, активності, рідинах, тоніну
Формула / Реферат:
Спосіб визначення активності тоніну у біологічних рідинах, який полягає у тому, що проводять кон'югацію маркерного ферменту з субстратом білкової природи, що містить аргінін, іммобілізацію отриманого комплексу на поверхні полістиролової твердої фази, а також протеолітичну реакцію з наступним усуненням продуктів реакції відмиванням і визначення залишкової активності маркерного ферменту, який відрізняється тим, що субстрат, який містить...
Спосіб визначення активності макрофагальної еластази

Номер патенту: 28914
Опубліковано: 25.12.2007
Автори: Кубишкін Анатолій Володимирович, Пальона Юлія Володимирівна, Фомочкина Ірина Іванівна
МПК: G01N 33/50
Мітки: макрофагальної, еластази, визначення, спосіб, активності
Формула / Реферат:
Спосіб визначення активності макрофагальної еластази, що включає використання синтетичного субстрату, який відрізняється тим, що визначають еластазоподібну активність, потім у присутності інгібітора діізопропілфлюорофосфату визначають залишкову активність макрофагальної еластази.
Спосіб іммобілізації пероксидази

Номер патенту: 17733
Опубліковано: 16.10.2006
Автори: Севастьянов Олег Всеволодович, Романовська Ірина Ігорівна, Осійчук Ольга Володимирівна
МПК: C12N 11/00
Мітки: пероксидази, спосіб, іммобілізації
Формула / Реферат:
Спосіб іммобілізації пероксидази, що включає зв'язування її з целюлозною матрицею, який відрізняється тим, що як целюлозну матрицю використовують гідратцелюлозну мембрану, попередньо оброблену періодатом натрію, а зв'язування здійснюють шляхом інкубації суміші гідратцелюлозної мембрани і пероксидази протягом 18-20 год., після чого до суміші додають борогідрид натрію і витримують 10-12 год.
Спосіб визначення активності хімази у біологічних рідинах

Номер патенту: 32131
Опубліковано: 15.12.2000
Автор: Самохіна Любов Михайлівна
МПК: C12Q 1/37, G01N 33/49
Мітки: біологічних, рідинах, спосіб, хімази, активності, визначення
Текст:
...вносять у лунки полістиролової плашки з імобілізованим комплексом пероксидаза-субстрат у контрольні зразки , вносять лише буфер. Інкубують при 37 °С протягом 15 хвилин. 4. Продукти реакції усувають відмиванням як вказано раніше . 5. Визначають залишкову активність маркерного фермента . Для цього у лунки плашок вносять суміш яка містить ортофені , лендіамін (ОФД) і пергідроль у 0.1 М цитратному буфері рН 4.5-4.6. 6. Зупиняють реакцію...
Спосіб визначення активності n-ацетил-b-d-глюкозамінідази в спермальній плазмі

Номер патенту: 17441
Опубліковано: 15.09.2006
Автори: Нікуліна Галина Григорівна, Лучицький Віталій Євгенович, Імшинецька Людмила Павлівна, Мигаль Людмила Якимівна
МПК: G01N 33/48
Мітки: визначення, спосіб, плазми, спермальній, активності, n-ацетил-b-d-глюкозамінідази
Формула / Реферат:
Спосіб визначення активності N-ацетил-β-D-глюкозамінідази в спермальній плазмі, який включає додавання до нативної біологічної рідини нітратного буфера та розчину субстрату, інкубацію реакційної суміші при 37°С, припинення ферментативної реакції додаванням розчину вуглекислого натрію, визначення оптичної густини отриманого розчину фотоколориметричним методом та обчислення результату у відносних одиницях, який відрізняється тим, що...