Спосіб підвищення стійкості рослини до абіотичного стресу








Формула / Реферат
1. Спосіб підвищення стійкості рослини до абіотичного стресу, який включає стадії, на яких:
a) вводять в рослинну клітину касету експресії, причому касета експресії містить полінуклеотид, функціонально пов'язаний з промотором, який забезпечує транскрипцію зазначеного полінуклеотиду в рослині, де зазначений полінуклеотид кодує поліпептид, причому зазначений поліпептид має трегалоза-6-фосфатфосфатазну активність, де поліпептид був модифікований так, що він має знижену ферментативну активність у порівнянні з немодифікованим трегалоза-6-фосфатфосфатазою;
б) регенерують рослину з рослинної клітини із стадії а), і, таким чином,
в) одержують рослину з підвищеною стійкістю до абіотичного стресу, причому зазначений абіотичний стрес являє собою стрес, викликаний нестачею води, де рослиною є маїс; і
де зазначена модифікації вибрана із групи, яка складається із: OsT6PP-H244D, OsT6PP-A102V та I129F.
2. Спосіб за п. 1, в якому стрес, викликаний нестачею води, обумовлений посухою.
Текст
Реферат: Винахід належить до способу підвищення стійкості рослини до абіотичного стресу шляхом введення в рослину полінуклеотиду, який кодує поліпептид, що має трегалоза-6фосфатфосфатазну активність, де поліпептид модифікований так, що він має знижену ферментативну активність у порівнянні з немодифікованим трегалоза-6-фосфатфосфатазою. UA 115768 C2 (12) UA 115768 C2 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 Галузь техніки, до якої належить винахід [001] Даний винахід належить, загалом, до галузі молекулярної біології й розглядає різні полінуклеотиди, поліпептиди й способи застосування, які можуть використовуватися для збільшення врожайності трансгенних рослин. Трансгенні рослини, які містять будь-який з полінуклеотидів або поліпептидів, описаних у даному документі, можуть проявляти будь-яку з ознак, що включають збільшену врожайність, збільшену стійкість до абіотичного стресу, збільшений клітинний ріст, збільшений коефіцієнт використання води й збільшений коефіцієнт використання поживних речовин. Передумови винаходу [002] Збільшення населення світу й виснаження запасу орних земель, доступних для сільського господарства, активізує потребу в дослідженні в галузі підвищення ефективності сільського господарства. Традиційні способи поліпшення сільськогосподарських і садових культур застосовують методики селекційного розведення для виявлення рослин, що мають необхідні характеристики. Однак такі методики селекційного розведення мають певні недоліки, а саме, що ці методики часто є трудомісткими й дають у результаті рослини, які часто містять гетерогенні генетичні компоненти, що не завжди може приводити до передачі необхідної ознаки від батьківських рослин. Успіхи в молекулярній біології дозволили людству модифікувати ідіоплазму тварин і рослин. Генна інженерія рослин передбачає виділення й маніпуляцію з генетичним матеріалом (як правило, у формі ДНК або РНК) і наступне введення цього генетичного матеріалу в геном рослини. Така технологія має здатність надавати сільськогосподарським культурам або рослинам різні поліпшені економічні, агрономічні або садівничі ознаки. Короткий опис винаходу [003] Викладений нижче короткий опис перераховує деякі варіанти здійснення об'єктів даного винаходу, а також, у багатьох випадках, перераховує зміни й перестановки цих варіантів здійснення. Даний короткий опис є винятково ілюстративним щодо численних і різноманітних варіантів здійснення. Згадування однієї або декількох репрезентативних ознак указаного варіанта здійснення також є ілюстративним. Такий варіант здійснення може типово мати місце зі згаданою ознакою(ами) або без неї; аналогічно, ці ознаки можуть застосовуватися щодо інших варіантів здійснення даного винаходу, незалежно від того, перераховані вони в короткому описі винаходу, чи ні. Щоб уникнути надмірного повторення, даний короткий опис не перераховує або припускає всі можливі комбінації таких ознак. [004] Даний винахід передбачає нуклеотидні послідовності, що кодують поліпептиди трегалозного шляху, які при трансгенній експресії в рослині збільшують урожайність. Поліпептиди T6PP, описані в даному документі, містять модифікації, які змінюють активність білків трегалоза-6-фосфатфосфатаз (T6PP), причому активність знижується в порівнянні з немодифікованою T6PP. Модифіковані T6PP можуть містити консенсусну послідовність, проілюстровану в SEQ ID NO: 9, і можуть мати щонайменше одну модифіковану амінокислоту. Модифікована T6PP може мати модифікацію, створену в консервативній амінокислоті, включаючи консервативні амінокислоти, які описані в консенсусній послідовності SEQ ID NO: 9. Модифіковані T6PP можуть викликати in vitro активацію T6PP дикого типу. Модифікація в T6PP може знаходитись в межах щонайменше одного з домену CAP, фосфатазного домену, Aфосфатазного домену або B-фосфатазного домену. T6PP, що мають зменшене зв'язування із субстратом трегалоза-6-фосфатом (T6P), демонструють збільшену врожайність і підвищену стресостійкість при трансгенній експресії в рослинах, а також мають підвищену стійкість до стресу, включаючи посуху. Рослини, які експресують модифіковану T6PP, включають однодольні або дводольні рослини, в тому числі рослини, вибрані із групи, що складається з маїсу, цукрового очерету, сої, рису, сорго або пшениці. Модифікації поліпептидів T6PP можуть включати одну з амінокислотних замін, амінокислотних делецій або амінокислотних вставок або їх комбінацію. Модифіковані T6PP можуть являтися членом надсімейства дегалогеназ галогенокислот (HAD) фосфатаз. Ферментативна активність T6PP може бути знижена щонайменше на 50 %, 60 %, 70 %, 80 %, 90 %, 95 %, 99 % або 100 %. [005] Будь-який з описаних поліпептидів T6PP може застосовуватися для одержання полінуклеотидів. Полінуклеотиди, що кодують модифіковані поліпептиди T6PP, можна вводити в клітину-хазяїна. Клітина-хазяїн може включати рослинну клітину. Полінуклеотиди можуть бути включені в касети експресії, які забезпечують транскрипцію полінуклеотида в рослині. Касети експресії можуть включати промотор, який експресує полінуклеотид у репродуктивній тканині рослини. Крім того, репродуктивна тканина може бути вибрана з групи, що складається з тканини колоска, вузла качана, тканини приквітка, меристемної тканини колоска, тканини квітконіжки суцвіття й тканини незрілої квітки. Промотор, який експресує полінуклеотиди T6PP, 1 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 може включати промотор OsMADS або промотор OsMADS6. Екстракт, що містить полінуклеотиди або поліпептиди, можна отримати з будь-чого з клітин-хазяїнів, рослин, частин рослини або тканин рослини. [006] Способи, розкриті в даному документі, додатково включають уведення в рослини полінуклеотидів, наприклад, як розкрито в даному документі. Трансгенні рослини, що містять полінуклеотиди, розкриті в даному документі, можуть проявляти збільшений клітинний ріст, збільшену міць рослин і/або сходів, збільшену врожайність, збільшену вагу насіння, збільшений коефіцієнт використання води та/або збільшену біомасу. Передбачається, що рослини, отримані за допомогою способів, розкритих у даному документі, мають підвищену стійкість до абіотичного стресу. Трансгенні рослини, описані в даному документі, можуть давати більш високу врожайність, як відносно врожайності біомаси, так і врожаю зерна рослини. Один аспект даного винаходу передбачає різні модифікації, які можна здійснювати щодо будь-якої даної генної послідовності T6PP, що може використовуватися в трансгенних рослинах для забезпечення збільшеної врожайності. Альтернативно, щоб забезпечити в рослини збільшену врожайність та стресостійкість, можна застосовувати інші численні способи для збільшення рівнів T6P. [007] Способи збільшення врожайності, збільшення стійкості рослини до абіотичного стресу або зменшення безпліддя в рослини, збільшення кількості качанів на рослину та/або зерен на рослину в умовах дефіциту води можуть включати введення у рослинну клітину касети експресії, що містить модифіковану T6PP, а потім одержання трансгенної рослини. Рослина може являти собою однодольну рослину, наприклад, може являти собою рослину маїсу, рису, пшениці, сорго, цукрового очерету або газонної трави. Це може бути дводольна рослина, така як соя. Для способів підвищення стійкості рослини до абіотичного стресу стрес може бути вибраний з групи, що складається зі стресу, викликаного нестачею води, теплового стресу або стресу, викликаного низькими температурами. Стрес, викликаний нестачею води, може бути обумовлений посухою. [008] Крім того, включені способи модифікації в рослині рівня трегалоза-6-фосфату шляхом ідентифікації поліпептиду в трегалозному шляху; модифікації поліпептиду так, щоб він мав змінену ферментативну активність; уведення в рослину касети експресії, що містить полінуклеотид, який кодує модифікований поліпептид, і одержання рослини з модифікованими рівнями трегалоза-6-фосфату. Поліпептид може мати активність, вибрану з групи, що складається з трегалоза-6-фосфат-синтазної; трегалоза-6-фосфатфосфатазної та трегалазної. Модифіковані рівні трегалоза-6-фосфату можуть приводити до збільшення експресії генів у пентозофосфатному шунтовому шляху під час стресу, викликаного посухою, у тканині вузла качана рослини. Додатково або альтернативно, модифіковані рівні трегалоза-6-фосфату можуть обумовлювати збільшення рівня карбоангідрази в тканині вузла качана рослини. Модифікований поліпептид може експресуватися у репродуктивних тканинах рослини, в тому числі у тканині колоска, вузлі качана, тканині приквітки, меристемній тканині колоска, тканині квітконіжки суцвіття й тканині незрілої квітки. Модифікований поліпептид модифікованого ферменту трегалозного шляху може мати знижену ферментативну активність і/або обумовлювати in vitro активацію трегалоза-6-фосфатфосфатази дикого типу. [009] Ці та інші ознаки, цілі й переваги даного винаходу стануть зрозумілішими з наступного опису. В описі наведене посилання на супровідні послідовності, що є його частиною та у яких показані з метою ілюстрації, а не обмеження, варіанти здійснення даного винаходу. Опис переважних варіантів здійснення не передбачений для обмеження даного винаходу, що охоплює всі модифікації, еквіваленти й альтернативи. Отже, необхідно привести посилання на варіанти здійснення, перераховані в даному документі для інтерпретації об'єму даного винаходу. Короткий опис послідовностей у переліку послідовностей nt= нуклеотидна послідовність pt= білкова послідовність SEQ ID NO: 1 трегалоза-6-фосфатфосфатаза (nt), OsT6PP-WT SEQ ID NO: 2 трегалоза-6-фосфатфосфатаза (pt), OsT6PP-WT SEQ ID NO: 3 трегалоза-6-фосфатфосфатаза - одинарна модифікація (nt), OsT6PP-H244D SEQ ID NO: 4 трегалоза-6-фосфатфосфатаза – одинарна модифікація (pt), OsT6PP-H244D SEQ ID NO: 5 трегалоза-6-фосфатфосфатаза - подвійна модифікація a (nt), OsT6PP-I129F SEQ ID NO: 6 трегалоза-6-фосфатфосфатаза – подвійна модифікація a (pt), 2 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 OsT6PP-I129F SEQ ID NO: 7 трегалоза-6-фосфатфосфатаза - потрійна модифікація b (nt) SEQ ID NO: 8 трегалоза-6-фосфатфосфатаза – потрійна модифікація b (pt) SEQ ID NO: 9 консенсусна послідовність трегалоза-6-фосфатфосфатази SEQ ID NO: 10 фосфатазний домен трегалоза-6-фосфатфосфатази SEQ ID NO: 11 A-фосфатазний бокс трегалоза-6-фосфатфосфатази SEQ ID NO: 12 B-фосфатазний бокс трегалоза-6-фосфатфосфатази SEQ ID NO: 13 промотор OsMADS6 SEQ ID NO: 14 трегалоза-6-фосфатфосфатаза Arabidopsis thaliana 19925 SEQ ID NO: 15 трегалоза-6-фосфатфосфатаза Arabidopsis thaliana 19926 SEQ ID NO: 16 трегалоза-6-фосфатфосфатаза Oryza sativa 19924 SEQ ID NO: 17 подібна до трегалоза-6-фосфатфосфатази Thermoplasma acidophilum (Q9HIW7) Опис графічних матеріалів [010] На Фіг. 1A-1C показане вихідне вирівнювання послідовностей T6PP з Arabidopsis thaliana (19925; SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924; SEQ ID NO: 16), Oryza sativa (15777; OsT6PP-WT дикого типу; SEQ ID NO: 2) і Thermoplasma acidophilum (Q9HIW7; SEQ ID NO: 17). Послідовності вирівнювали за допомогою Vector NTI із застосуванням способу ClustalW. [011] На Фіг. 2 показане філогенетичне дерево, отримане від вирівнювання множинних білків T6PP. [012] На Фіг. 3 показаний опис трегалозного шляху. Трегалозний шлях складається з двох біосинтетичних ферментів, трегалоза-6-фосфатсинтази (TPS) і трегалоза-6-фосфатфосфатази (T6PP), і одного гідролізуючого ферменту, трегалази, що залучений в розщеплення. [013] На Фіг. 4 показаний трегалозний шлях з додатковими продуктами низхідних стадій. UDPG одержують безпосередньо за допомогою сахарозосинтази; G6P одержують із глюкози за допомогою інвертази й гексокінази або за допомогою фруктози й фруктоза-6-фосфату (F6P) за рахунок застосування сахарозосинтази, фруктокінази й фосфоглюкозаізомерази. UDPG і G6P також являють собою два центральні активовані попередники, з яких зрештою можуть походити багато клітинних функціональних сполук. UDPG є попередником для основних полісахаридів клітинної стінки й для гліколіпідів. G6P є попередником для синтезу крохмалю, NADPH за рахунок окисної частини пентозофосфатного шляху, а ATP за рахунок гліколізу, циклу Кребса й окисного фосфорилування. Крім того, G6P може перетворюватися на фруктоза-6-фосфат і разом з UDPG може застосовуватися для синтезу сахароза-6-фосфату, а потім сахарози. Будучи зробленим з UDPG і G6P, T6P перебуває на перетинах основних вуглеводних потоків у рослинах. [014] На Фіг. 5 описаний вплив зв'язування T6P з SnRK1. SnRK1 є гетеротримерним білком та являє собою рослинний гомолог протеїнкінази тварин, що активується AMP, й протеїнкінази дріжджів, що не ферментує сахарозу (SnF1). [015] На Фіг. 6 показане залучення SnRK1 у численні метаболічні шляхи в рослинах. [016] На Фіг. 7 показане стрічкове графічне зображення гомологічної моделі T6PP рису. Фермент містить два домени: α/β-гідролазний домен та менший "домен lid". Активний сайт знаходиться у верхній частині α/β-гідролазного домену (пляма); щілина для зв'язування із субстратом перебуває на границі між двома доменами. Ці два домени з'єднані гнучким лінкером, що дозволяє домену lid відкриватися й закриватися під час каталізу. Оскільки матриця для гомологічного моделювання, бактеріальний T6PP-споріднений білок (код PDB 1U02), являє собою апофермент з порожнім активним сайтом, імовірно, що відносна орієнтація домену lid та гідролазного домену дещо відрізняється в справжньому фермент-субстратному комплексі. [017] На Фіг. 8 показане розташування групи 6-фосфату T6P. Один з атомів кисню групи 6фосфату розташований в екваторіальному положенні координаційної сфери Mg2+. Asp121, що акцептує фосфатну групу під час каталізу з утворенням проміжної сполуки з ковалентним зв'язком, приймає аксіального положення. Lys289 безпосередньо зв'язаний водневим зв'язком з фосфатною групою, а His181 перебуває в безпосередній близькості. Обидва залишки оптимально розташовані для стабілізації негативного заряду, що утворюється на фосфатному фрагменті під час каталізу. [018] На Фіг. 9 показані здатні до обертання зв'язки в субстраті T6P. У той час як положення й орієнтація піранозного кільця, що є найближчим до групи 6-фосфату, точно визначена за допомогою фіксованої фосфатної групи, існує істотна гнучкість між першим і другим кільцями внаслідок наявності двох здатних до обертання глікозидних зв'язків (стрілки). Діапазон 3 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 можливих положень для другого кільця отримували шляхом обертання двох зв'язків. Конформації, у яких друге кільце перетинається з білком, відкидали. [019] На Фіг. 10 показана консенсусна послідовність для T6PP (SEQ ID NO: 9). Залишки, виділені жирним шрифтом і підкреслені, визначають висококонсервативні ділянки. Положення в консенсусній послідовності, що марковані "X", указують на ділянки мінливості, де будь-яка інша окрема буква відповідає загальноприйнятому однобуквеному позначенню амінокислот, загальновживаному в даній галузі техніки. Підкреслені амінокислотні залишки вказують на ділянки, які можна модифікувати для конструювання поліпептидів T6PP, що забезпечують збільшену врожайність і/або підвищену витривалість до стресу у трансгенної рослини. Залишки, позначені DYDGTLSPIV, кодують B-фосфатазний бокс. [020] На Фіг. 11A-11C показане остаточне вирівнювання послідовностей T6PP для Arabidopsis thaliana (19925; SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924; SEQ ID NO: 16) і Oryza sativa (15777; OsT6PP-WT дикого типу; SEQ ID NO: 2), а також T6PP-спорідненого білка з Thermoplasma acidophilum (Q9HIW7; SEQ ID NO: 17). Докладний опис винаходу [021] При здійсненні даного винаходу на практиці будуть використовуватися, якщо не зазначене інше, загальноприйняті методики ботаніки, мікробіології, вирощування тканин, молекулярної біології, хімії, біохімії, генетики кількісних ознак рослин, статистики й технології рекомбінантної ДНК, що знаходяться у межах навичок фахівців у даній галузі. Такі методики роз'яснюються в повному обʼємі в літературі. Див., наприклад, Langenheim and Thimann, (1982) Botany: Plant Biology and Its Relation to Human Affairs, John Wiley; Cell Culture and Somatic Cell Genetics of Plants, vol. 1, Vasil, ed. (1984); Stanier, et al., (1986) The Microbial World, 5th ed., Prentice-Hall; Dhringra and Sinclair, (1985) Basic Plant Pathology Methods, CRC Press; Maniatis, et al., (1982) Molecular Cloning: A Laboratory Manual; DNA Cloning, vols. I and II, Glover, ed. (1985); Oligonucleotide Synthesis, Gait, ed. (1984); Nucleic Acid Hybridization, Hames and Higgins, eds. (1984); і серію Methods in Enzymology, Colowick and Kaplan, eds, Academic Press, Inc., San Diego, Calif. [022] Одиниці, префікси й символи можуть виражатися в їхній формі, прийнятній у системі SI. Якщо не зазначене інше, нуклеїнові кислоти записані зліва направо в орієнтації 5′ - 3′; амінокислотні послідовності записані зліва направо в орієнтації від аміно- до карбокси-кінця, відповідно. Числові діапазони містять у собі числові значення, що визначають діапазон. Амінокислоти в даному документі можуть позначатися або їхніми загальновідомими трибуквеними символами, або однобуквеними символами, які рекомендовані Комісією з біохімічної номенклатури IUPAC-IUB. Аналогічно, нуклеотиди можуть позначатися їхніми загальноприйнятими однобуквеними кодами. Вирази, що визначаються нижче, більш повно визначені за допомогою посилання на опис винаходу в цілому. [023] Якщо не визначене інше, усі технічні й наукові терміни, які застосовуються в даному документі, мають таке саме значення, що звичайно розуміється фахівцем у галузі, до якої належить даний винахід. [024] Слід розуміти, що даний винахід не обмежений конкретною методологією, протоколами, клітинними лініями, видами або родами рослин, конструктами й реагентами, фактично описаними. Слід також розуміти, що термінологія, яка застосовується в даному документі, представлена винятково з метою опису конкретних варіантів здійснення і не передбачена для обмеження об'єму даного винаходу. [025] Форми однини, які застосовуються в даному документі, включають посилання на форми множини, якщо контекст явно не вказує на інше. Таким чином, наприклад, посилання на "вектор" являє собою посилання на один або кілька векторів і включає їхні еквіваленти, відомі фахівцям у даній галузі. [026] Вираз "приблизно" використовується в даному документі для позначення наближено, грубо, близько або в районі. Коли вираз "приблизно" використовується разом з числовим діапазоном, він модифікує цей діапазон шляхом розширення границь вище й нижче зазначених числових значень. Загалом, вираз "приблизно" використовується в даному документі для модифікації числового значення вище й нижче зазначеного значення за допомогою відхилення на 20 відсотків. [027] Слово "або", як застосовується в даному документі, означає будь-якого члена конкретного переліку, а також включає будь-яку комбінацію членів з даного переліку. [028] Вирази "містить", "що містить", "включає", "що включає", "що має" і споріднені з ними за значенням слова означають "включаючи, але без обмеження". Вираз "що складається з" означає "включаючи й обмежуючись". 4 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 [029] Вираз "що складається практично з" означає, що композиція, спосіб або структура можуть включати додаткові інгредієнти, етапи та/або частини, але тільки якщо додаткові інгредієнти, етапи та/або частини не суттєво змінюють основні й нові характеристики заявленої композиції, способу або структури. [030] У всіх випадках, коли у даному документі вказується числовий діапазон, мається на увазі, що він включає будь-яке перераховане число (дробове або ціле) у межах вказаного діапазону. Фрази "у діапазоні/знаходиться у діапазони між" першим вказаним числом і другим вказаним числом і "у діапазоні/знаходиться у діапазоні від "першого вказаного числа" до "другого вказаного числа застосовуються в даному документі взаємозамінно та, мається на увазі, що вони включають перше й друге вказані числа і всі дробові та цілі числа між ними. Вираз "спосіб", як застосовується в даному документі, стосується способів дії, засобів, методик і процедур для здійснення даного завдання, включаючи, але без обмеження, такі способи дії, засоби, методики й процедури, або відомі, або які легко розробити на основі відомих способів дії, засобів, технік і процедур фахівцям в хімічній, фармакологічній, біологічній, біохімічній та медичній галузях техніки. Слід розуміти, що певні ознаки даного винаходу, описані для ясності в контексті окремих варіантів здійснення, можуть також бути представлені в комбінації в одному варіанті здійснення. Навпаки, різні ознаки даного винаходу, описані для стислості в контексті одного варіанта здійснення, можуть також бути представлені окремо, або в будь-якій придатній субкомбінації, або у формі, що придатна у будь-якому іншому описаному варіанті здійснення даного винаходу. Певні ознаки, описані в контексті різних варіантів здійснення, не повинні розглядатися як суттєві ознаки цих варіантів здійснення, якщо тільки варіант здійснення є незастосовним без цих елементів. [031] Під виразом "мікроб" розуміють будь-який мікроорганізм (включаючи як еукаріотичні, так і прокаріотичні мікроорганізми), такий як гриби, дріжджі, бактерії, актиноміцети, водорості й найпростіші, а також інші одноклітинні структури. [032] Під виразом "ампліфікований" розуміють створення множинних копій послідовності нуклеїнової кислоти або множинних копій, комплементарних послідовності нуклеїнової кислоти, із застосуванням щонайменше однієї з послідовностей нуклеїнової кислоти у якості матриці. Системи ампліфікації включають систему полімеразної ланцюгової реакції (ПЛР), систему лігазної ланцюгової реакції (ЛЛР), ампліфікацію на основі послідовності нуклеїнової кислоти (NASBA, Cangene, Міссіссога, Онтаріо), системи O-бета-реплікази, систему ампліфікації на основі транскрипції (TAS) та ампліфікацію з переміщенням ланцюга (SDA). Див., наприклад, Diagnostic Molecular Microbiology: Principles and Applications, Persing, et al., eds., American Society for Microbiology, Washington, D.C. (1993). Продукт ампліфікації має назву амплікон. [033] Вираз "консервативно модифіковані варіанти" застосовується як щодо амінокислотних послідовностей, так і щодо послідовностей нуклеїнових кислот. Що стосується конкретних послідовностей нуклеїнової кислоти, консервативно модифікованими варіантами називають такі нуклеїнові кислоти, які кодують ідентичні або консервативно модифіковані варіанти амінокислотних послідовностей. Внаслідок виродженості генетичного коду велика кількість функціонально ідентичних нуклеїнових кислот кодують будь-який даний білок. Наприклад, усі з кодонів GCA, GCC, GCG і GCU кодують амінокислоту аланін. Таким чином, у будь-якому положенні, де кодоном визначено аланін, кодон може бути змінений на будь-який з описаних відповідних кодонів без зміни поліпептиду, що кодується. Такі варіації нуклеїнових кислот являють собою "мовчазні варіації" і представляють собою один вид консервативно модифікованої варіації. Кожна послідовність нуклеїнової кислоти в даному документі, яка кодує поліпептид, також описує кожну можливу мовчазну варіацію нуклеїнової кислоти. Фахівцю в даній галузі буде зрозуміло, що кожний кодон у нуклеїновій кислоті (за винятком AUG, який, як правило, є єдиним кодоном для метіоніну; єдиним виключенням є Micrococcus rubens, для якого метіоніновим кодоном є GTG (Ishizuka, et al., (1993) J. Gen. Microbiol. 139:425-32) може бути модифікований з одержанням функціонально ідентичної молекули. Відповідно, кожна мовчазна варіація нуклеїнової кислоти, що кодує поліпептид згідно з даним винаходом, передбачається в кожній описаній поліпептидній послідовності й включена в даний документ за допомогою посилання. [034] "Контрольна рослина" або "контроль", як застосовується в даному документі, може являти собою нетрансгенну рослину батьківської лінії, яку застосовували в даному винаході для створення трансгенної рослини. Контрольна рослина в деяких випадках може являти собою лінію трансгенних рослин, що включають порожній вектор або маркерний ген, але не містять рекомбінантний полінуклеотид згідно з даним винаходом, який експресується в трансгенній рослині, що підлягає оцінюванню. В інших випадках контрольна рослина являє собою трансгенну рослину, яка експресує ген за допомогою конститутивного промотора. Як правило, 5 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 контрольна рослина являє собою рослину тієї ж лінії або сорту, що й трансгенна рослина, що підлягає тестуванню, яка не має специфічної рекомбінантної ДНК, що забезпечує ознаку, якою характеризується трансгенна рослина. Така рослина-попередник, яка не містить такої специфічної рекомбінантної ДНК, що забезпечує ознаку, може являти собою природну рослину, рослину дикого типу, елітну нетрансгенну рослину або трансгенну рослину без специфічної рекомбінантної ДНК, що забезпечує ознаку, якою характеризується трансгенна рослина. Рослина-попередник, яка не містить специфічної рекомбінантної ДНК, що забезпечує ознаку, може являти собою сибс трансгенної рослини, яка має специфічну рекомбінантну ДНК, що забезпечує ознаку. Така рослина-попередник, що є сибсом, може включати іншу рекомбінантну ДНК. [035] Що стосується амінокислотних послідовностей, фахівцю в даній галузі буде зрозуміло, що окремі заміни, делеції або вставки в послідовність нуклеїнової кислоти, пептидну, поліпептидну або білкову послідовність, які змінюють, додають або видаляють одну амінокислоту або невеликий відсоток амінокислот у послідовності, що кодується, приводять до "консервативно модифікованого варіанту", коли зміна відбувається в результаті заміщення амінокислоти хімічно подібною амінокислотою. Таким чином, в такий спосіб можна змінювати будь-яке число амінокислотних залишків, вибране з групи цілих чисел, що складається з 1–15. Відповідно, можна зробити, наприклад, 1, 2, 3, 4, 5, 7 або 10 змін. Консервативно модифіковані варіанти, як правило, забезпечують біологічну активність, аналогічну такій немодифікованої поліпептидної послідовності, з якої вони походять. Наприклад, субстратна специфічність, ферментативна активність або зв'язування з лігандом/рецептором, як правило, становить щонайменше 30 %, 40 %, 50 %, 60 %, 70 %, 80 % або 90 %, переважно 60-90 % від нативного білка для його нативного субстрату. Таблиці консервативних замін, у яких наведені функціонально подібні амінокислоти, добре відомі в даній галузі техніки. [036] Кожна з наступних шести груп містить амінокислоти, що представляють собою консервативні заміни одна одної: 1) Аланін (A), Серин (S), Треонін (T); 2) Аспарагінова кислота (D), Глутамінова кислота (E); 3) Аспарагін (N), Глутамін (Q); 4) Аргінін (R), Лізин (K); 5) Ізолейцин (I), Лейцин (L), Метіонін (M), Валін (V) і 6) Фенілаланін (F), Тирозин (Y), Триптофан (W). 7) Див. також Creighton, Proteins, W.H. Freeman and Co. (1984). [037] Як застосовується в даному документі, вирази "модифікований" або "модифікація" взаємозамінно стосуються спланованих або випадкових замін, делецій або вставок у послідовність нуклеїнової кислоти, пептидну, поліпептидну або білкову послідовність, які змінюють, додають або видаляють щонайменше один амінокислотний залишок у межах даного поліпептиду. Вираз "модифікована T6PP", як застосовується в даному документі, стосується будь-якої нуклеїнової кислоти, що кодує T6PP або пептиди, поліпептиди або білок, що має T6PP активність, кожний з яких був модифікований таким чином, що отримана T6PP забезпечує модифіковану T6PP активність і/або модифіковане зв'язування з T6P, що приводить до збільшеної врожайності та/або стійкості до абіотичного стресу в рослини у порівнянні з немодифікованою T6PP. [038] Як застосовується в даному документі, "гомологічне положення" стосується положення однієї або декількох амінокислот у поліпептиді або однієї або декількох пар основ у полінуклеотидній послідовності, які знаходяться в аналогічному або еквівалентному положенні в другому поліпептиді або полінуклеотиді, що є ортологом, паралогом або гомологом вихідної послідовності. Положення амінокислот може знаходитися в одній функціональній ділянці двох білків, але може не являти собою строго відповідне числове положення амінокислоти у двох поліпептидних послідовностях. Гомологічне положення амінокислот у двох білках може визначатися за допомогою декількох способів, добре відомих у даній галузі техніки, в тому числі, наприклад, вирівнювання послідовностей (наприклад, BLAST), тривимірного моделювання білків (див., наприклад, Sander C. and Scheider R., (1991) PROTEINS: Structure, Function and Genetics 9:56-68) і подібного. [039] Під "що кодує" або "що кодується" стосовно певної нуклеїнової кислоти розуміють, що вона містить інформацію для трансляції в певний білок. Нуклеїнова кислота, що кодує білок, може містити послідовності, які не транслюються (наприклад, інтрони), серед ділянок нуклеїнової кислоти, які транслюються, або може не містити такі проміжні послідовності, які не транслюються (наприклад, як у кДНК). Інформація, за якої кодується білок, визначається шляхом застосування кодонів. Як правило, амінокислотна послідовність кодується нуклеїновою 6 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 кислотою із застосуванням "універсального" генетичного коду. Однак варіанти універсального коду, такі як представлені в мітохондріях деяких рослин, тварин і грибів, бактерії Mycoplasma capricolumn (Yamao, et al., (1985)Proc. Natl. Acad. Sci. USA 82:2306-9) або інфузорії Macronucleus, можуть застосовуватися, коли нуклеїнову кислоту експресують із застосуванням цих організмів. [040] Якщо нуклеїнову кислоту одержують або змінюють синтетичним шляхом, то можна отримати перевагу з огляду на використання відомих переважних кодонів у передбачуваного хазяїна, у якого буде експресуватися нуклеїнова кислота. Наприклад, хоча послідовності нуклеїнової кислоти за даним винаходом можна експресувати як у видах однодольних, так і дводольних рослин, послідовності можна модифікувати з урахуванням специфічного переважного використання кодонів і переважного вмісту GC в однодольних рослин або дводольних рослин, оскільки було показано, що ці переваги відрізняються (Murray, et al., (1989) Nucleic Acids Res. 17:477-98 і в даний документ включена за допомогою посилання). Таким чином, переважний кодон маїсу для конкретної амінокислоти можна отримати від відомих генних послідовностей з маїсу. Частота використання кодону маїсу для 28 генів з рослин маїсу наведена в Таблиці 4 у Murray, et al., вище. [041] Як застосовується в даному документі, "гетерологічний" у відношенні нуклеїнової кислоти являє собою нуклеїнову кислоту, яка походить з чужорідного виду, або, якщо вона походить із того самого виду, то є суттєво модифікованою в порівнянні з її нативною формою в композиції та/або геномному локусі шляхом навмисного втручання людини. Наприклад, промотор, функціонально пов'язаний з гетерологічним структурним геном, походить від виду, що відрізняється від того, від якого отримали структурний ген, або, якщо він походить від того самого виду, то один або обидва є суттєво модифікованими в порівнянні з їхньою вихідною формою. Гетерологічний білок може брати початок від чужорідного виду, або, якщо він походить від того самого виду, то він є суттєво модифікованим у порівнянні з його вихідною формою шляхом навмисного втручання людини. [042] Під "клітиною-хазяїном" розуміють клітину, яка містить послідовність гетерологічної нуклеїнової кислоти за даним винаходом, яка містить вектор і підтримує реплікацію та/або експресію вектора експресії. Клітини-хазяї можуть являти собою прокаріотичні клітини, такі як E. coli, або еукаріотичні клітини, такі як клітини дріжджів, комах, рослин, амфібій або ссавців. Переважно, клітини-хазяї являють собою клітини однодольних або дводольних рослин, в тому числі, але без обмеження, маїсу, сорго, соняшника, сої, пшениці, люцерни, рису, бавовнику, каноли, ячменю, проса й томата. Особливо переважною клітиною-хазяїном з однодольної рослини є клітина-хазяїн маїсу. [043] Вираз "гібридизаційний комплекс ” включає посилання на дуплексну структуру нуклеїнової кислоти, що утворена двома одноланцюговими послідовностями нуклеїнової кислоти, які селективно гібридизовані одна з одною. [044] Вираз "введений" передбачає вставлення нуклеїнової кислоти в клітину за допомогою будь-якого способу, такого як "трансфекція", "трансформація" або "трансдукція", і він включає посилання на вбудовування нуклеїнової кислоти в еукаріотичну або прокаріотичну клітину, де нуклеїнова кислота може вбудовуватися в геном клітини (наприклад, в хромосомну, плазмідну, пластидну або мітохондріальну ДНК), перетворюватися на автономний реплікон, в якості частини мініхромосоми, або тимчасово експресуватися (наприклад, трансфікована мРНК). [045] Як застосовується в даному документі, вираз "пакет генів" стосується введення двох або більше генів у геном організму. У певних аспектах даного винаходу може бути бажаним здійснювати стекінг будь-якого гену, пов'язаного з абіотичним стресом (наприклад, білки холодового шоку, гени, пов'язані з реакцією ABA), з T6PP, що описана в даному документі. Аналогічно, також може бути бажаним здійснювати стекінг генів трегалозного шляху, як описано в даному документі, з генами, що забезпечують витривалість до впливу комах, витривалість до хвороб, збільшену врожайність або будь-яку іншу сприятливу ознаку (наприклад, збільшену висоту рослини тощо), відому у даній галузі техніки. Альтернативно, у трансгенних рослин, що містять модифікований ген трегалозного шляху, можна здійснювати стекінг з алелями нативної ознаки, які забезпечують додаткові ознаки, такі як поліпшене використання води, підвищена витривалість до хвороб і подібне. В одному варіанті здійснення у рослин, які експресують модифіковані гени трегалозного шляху, здійснюють стекінг з алелями, які описані в WO2011/079277. Можна здійснювати стекінг ознак шляхом введення касет експресії із множинними генами або розмноження/схрещування рослин з однією або декількома ознаками з іншими рослинами, які мають одну або декілька додаткових ознак. [046] Вираз "виділений" стосується матеріалу, такого як нуклеїнова кислота або білок, який головним чином або практично не містить компонентів, що звичайно супроводжують його або 7 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 взаємодіють з ним при знаходженні його в природному оточенні. Виділений матеріал необов'язково містить матеріал, що не зустрічається разом з матеріалом у його природному оточенні. Нуклеїнові кислоти, які є "виділеними", як визначено в даному документі, також називають "гетерологічними" нуклеїновими кислотами. Якщо інше не заявлене, то вираз "нуклеїнова кислота NUE" означає нуклеїнову кислоту, яка містить полінуклеотид ("полінуклеотид NUE"), що кодує поліпептид NUE повної або часткової довжини. [047] Як застосовується в даному документі, "нуклеїнова кислота" включає посилання на дезоксирибонуклеотидний або рибонуклеотидний полімер у формі одинарного або подвійного ланцюга та, якщо немає обмежень, охоплює відомі аналоги, які мають основну властивість природних нуклеотидів, оскільки вони гібридизуються з одноланцюговими нуклеїновими кислотами в такий спосіб, що аналогічний нуклеотидам, які зустрічаються в природі (наприклад, пептидні нуклеїнові кислоти). [048] Під "бібліотекою нуклеїнових кислот" розуміють сукупність виділених молекул ДНК або РНК, які в одному випадку включають основну представленість всієї частини генома певного організму, яка транскрибується. Про конструювання ілюстративних бібліотек нуклеїнових кислот, таких як бібліотеки геномних та кДНК, повідомляється в стандартній літературі з молекулярної біології, такій як Berger and Kimmel, (1987) Guide To Molecular Cloning Techniques, із серії Methods in Enzymology, vol. 152, Academic Press, Inc., San Diego, Calif.; Sambrook, et al., (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., vols. 1-3; і Current Protocols in Molecular Biology, Ausubel, et al., eds, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc. (1994 Supplement). В іншому випадку "бібліотеку нуклеїнових кислот", як визначено в даному документі, можна також розуміти як таку, що представляє бібліотеки, які містять передбачену частину, або скоріше, які фактично представляють не весь геном певного організму. Наприклад, малі РНК, мРНК і метильована ДНК. Бібліотека нуклеїнових кислот, як визначається в даному документі, може також охоплювати варіанти конкретної молекули (наприклад, сукупність варіантів для конкретного білка). [049] Як застосовується в даному документі, "функціонально пов'язаний" включає посилання на функціональний зв'язок між першою послідовністю, такою як промотор, і другою послідовністю, де промоторна послідовність ініціює й опосередковує транскрипцію ДНК, що відповідає другій послідовності. Звичайно "функціонально пов'язаний" означає, що послідовності нуклеїнових кислот, які є пов'язаними, є суміжними та, якщо є необхідність об'єднати дві області, що кодують білок, вони є суміжними та перебувають в одній рамці зчитування. [050] Як застосовується в даному документі, вираз "рослина" включає посилання на цілі рослини, органи рослини (наприклад, листки, стебла, корені тощо), насіння й рослинні клітини та їхнє потомство. Рослинна клітина, як застосовується в даному документі, включає без обмеження насіння, суспензійні культури, зародки, меристематичні ділянки, калюсну тканину, листки, корені, пагони, гаметофіти, спорофіти, пилок і мікроспори. Клас рослин, які можна застосовувати в способах за даним винаходом, як правило, настільки широкий, як клас вищих рослин, які піддаються методикам трансформації, включаючи як однодольні, так і дводольні рослини, у тому числі види з родів: Cucurbita, Rosa, Vitis, Juglans, Fragaria, Lotus, Medicago, Onobrychis, Trifolium, Trigonella, Vigna, Citrus, Linum, Geranium, Manihot, Daucus, Arabidopsis, Brassica, Raphanus, Sinapis, Atropa, Capsicum, Datura, Hyoscyamus, Lycopersicon, Nicotiana, Solanum, Petunia, Digitalis, Majorana, Ciahorium, Helianthus, Lactuca, Bromus, Asparagus, Antirrhinum, Heterocallis, Nemesis, Pelargonium, Panieum, Pennisetum, Ranunculus, Senecio, Salpiglossis, Cucumis, Browaalia, Glycine, Pisum, Phaseolus, Lolium, Oryza, Avena, Hordeum, Secale, Allium і Triticum. Особливо переважною рослиною є Zea mays. [051] Як застосовується в даному документі, "врожайність" може включати посилання на бушелі на акр зернових культур при збиранні, з поправкою на вологість зерна (для маїсу, наприклад, типово становить 15 %), та об'єм утвореної біомаси (для кормових культур, таких як люцерна, й розмір коренів рослини для сільськогосподарських культур, що дають кілька врожаїв на рік). Вологість зерна вимірюють у зерні при збиранні. Скориговану натурну вагу зерна визначають як вагу у фунтах на бушель, з поправкою на рівень вологості зерна при збиранні. Біомасу вимірюють як вагу утвореного придатного для збирання рослинного матеріалу. На врожайність можуть впливати багато властивостей, у тому числі без обмеження висота рослини, кількість бобів, положення боба на рослині, кількість міжвузлів, частота розтріскування бобів, розмір зерна, ефективність утворення бульбочок і фіксації азоту, ефективність асиміляції поживних речовин, асиміляції вуглецю, структура рослини, відсоток проростання насіння, міць сходів і ювенільні ознаки. На врожайність також може впливати ефективність проростання 8 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 (включаючи проростання в стресових умовах), швидкість росту (включаючи швидкість росту в стресових умовах), кількість качанів, кількість насінин на качан, розмір насіння, склад насіння (крохмаль, олія, білок) і характеристики наливу насіння. Врожайність рослини можна вимірювати різними способами, включаючи натурну вагу, кількість насіння на рослину, вагу насіння, кількість насіння на одиницю площі (тобто насіння, або вага насіння на акр), у бушелях на акр, тоннах на акр або кілограмах на гектар. Наприклад, урожайність кукурудзи можна вимірювати як вихід обмолочених кукурудзяних зерен на одиницю продуктивної площі, наприклад, у бушелях на акр або метричних тоннах на гектар, про що часто повідомляють на основі поправки на вологість, наприклад, на вологість 15,5 відсотка. Більше того, бушель кукурудзи визначається за законом штату Айова як 56 фунтів за вагою, при цьому корисний коефіцієнт перетворення для врожаю кукурудзи такий: 100 бушелів на акр еквівалентні 6,272 метричної тонни на гектар. В даній галузі техніки загальноприйнятими на практиці є й інші вимірювання врожайності. У певних варіантах здійснення даного винаходу врожайність може бути збільшена в стресових умовах та/або безстресових умовах. [052] Як застосовується в даному документі, "полінуклеотид" включає посилання на дезоксирибополінуклеотид, рибополінуклеотид або їхні аналоги, які мають основну властивість природного рибонуклеотиду, оскільки вони гібридизуються за жорстких умов гібридизації з практично такою самою нуклеотидною послідовністю, як і нуклеотиди, що зустрічаються в природі, та/або забезпечують трансляцію в таку саму амінокислоту(и), як і нуклеотид(и), що зустрічається в природі. Полінуклеотид може бути нативним або гетерологічним структурним або регуляторним геном повної довжини або його підпослідовністю. Якщо не зазначене інше, вираз включає посилання на певну послідовність, а також її комплементарну послідовність. Таким чином, ДНК або РНК із каркасами, модифікованими задля стабільності або з інших причин, являють собою "полінуклеотиди", як цей вираз передбачений в даному документі. Більше того, ДНК або РНК, що містять незвичайні основи, такі як інозин, або модифіковані основи, такі як тритильовані основи, наведені лише два приклади, є полінуклеотидами, як даний вираз застосовують у даному документі. Слід розуміти, що у відношенні ДНК і РНК була зроблена велика різноманітність модифікацій, які слугують для багатьох корисних цілей, відомих фахівцям у даній галузі. Вираз "полінуклеотид", як він застосовується в даному документі, охоплює такі хімічно, ферментативно або метаболічно модифіковані форми полінуклеотидів, а також хімічні форми ДНК і РНК, що характерні для вірусів і клітин, включаючи серед іншого прості й складні клітини. [053] Вирази "поліпептид", "пептид" і "білок" застосовуються в даному документі взаємозамінно для позначення полімеру з амінокислотних залишків. Вираз застосовують до амінокислотних полімерів, у яких один або декілька амінокислотних залишків є штучним хімічним аналогом відповідної амінокислоти, що зустрічається в природі, а також до полімерів з амінокислот, що зустрічаються в природі. [054] Як застосовується у даному документі, "промотор" включає посилання на ділянку ДНК вище від початку транскрипції та залучений у розпізнавання й зв'язування РНК-полімерази й інших білків для ініціації транскрипції. "Рослинний промотор" являє собою промотор, здатний ініціювати транскрипцію в рослинних клітинах. Ілюстративні рослинні промотори включають без обмеження такі, які одержують із рослин, рослинних вірусів і бактерій, що містять гени, які експресуються в рослинних клітинах, такі як Agrobacterium або Rhizobium. Прикладами є промотори, які переважно ініціюють транскрипцію в певних тканинах, таких як листки, корені, насіння, жилки, судини ксилеми, трахеїди або склеренхіма. Такі промотори називають "тканинопереважними". Специфічний для "типу клітин" промотор у першу чергу управляє експресією в певних клітинних типах в одному або декількох органах, наприклад, клітинах провідної тканини у коренях або листках. Промотор, "який індукується" або "який регулюється", являє собою промотор, що перебуває під контролем навколишнього середовища. Приклади умов навколишнього середовища, які можуть впливати на транскрипцію за допомогою промоторів, які індукуються, включають анаеробні умови або наявність світла. Іншим типом промотора є промотор, що регулюється стадією розвитку, наприклад, промотор, який управляє експресією під час розвитку пилку. Тканинопереважні, специфічні для типу клітин промотори, промотори, що регулюються стадією розвитку, й промотори, які індукуються, складають клас "неконститутивних" промоторів. "Конститутивний" промотор являє собою промотор, який є активним в більшості клітин за більшості умов навколишнього середовища. [055] Будь-яка придатна промоторна послідовність може застосовуватися у конструкті з нуклеїнових кислот за даним винаходом. Згідно з деякими варіантами здійснення даного винаходу промотор являє собою конститутивний промотор, тканиноспецифічний промотор, або промотор, що індукується абіотичним стресом. 9 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 [056] Придатні конститутивні промотори включають, наприклад, промотор 35S CaMV (SEQ ID NO:1546; Odell et al., Nature 313:810-812, 1985); промотор At6669 Arabidopsis (SEQ ID NO:1652; див. PCT публікацію № WO04081173A2); Ubi 1 маїсу (Christensen et al., Plant Mol. Biol. 18:675-689, 1992); актину рису (McElroy et al., Plant Cell 2:163-171, 1990); pEMU (Last et al., Theor. Appl. Genet. 81:581-588, 1991); 19S CaMV (Nilsson et al., Physiol. Plant 100:456-462, 1997); GOS2 (de Pater et al., Plant J November; 2(6):837-44, 1992); убіквітину (Christensen et al., Plant Mol. Biol. 18: 675-689, 1992); циклофіліну рису (Bucholz et al., Plant Mol Biol. 25(5):837-43, 1994); гістону H3 маїсу (Lepetit et al., Mol. Gen. Genet. 231: 276-285, 1992); актину 2 (An et al., Plant J. 10(1);107-121, 1996), конститутивний промотор кореневого кінчика CT2 (SEQ ID NO:1535; див. також PCT заявку № IL/2005/000627) і синтетичний Super MAS (Ni et al., The Plant Journal 7: 66176, 1995). Інші конститутивні промотори включають описані в патентах США №№ 5659026, 5608149; 5608144; 5604121; 5569597; 5466785; 5399680; 5268463 і 5608142. [057] Придатні тканиноспецифічні промотори включають, але без обмеження, специфічні для листків промотори [такі як описані, наприклад, Yamamoto et al., Plant J. 12:255-265, 1997; Kwon et al., Plant Physiol. 105:357-67, 1994; Yamamoto et al., Plant Cell Physiol. 35:773-778, 1994; Gotor et al., Plant J. 3:509-18, 1993; Orozco et al., Plant Mol. Biol. 23:1129-1138, 1993; і Matsuoka et al., Proc. Natl. Acad. Sci. USA 90:9586-9590, 1993], насінняпереважні промотори [наприклад, з специфічних для насіння генів (Simon, et al., Plant Mol. Biol. 5. 191, 1985; Scofield, et al., J. Biol. Chem. 262: 12202, 1987; Baszczynski, et al., Plant Mol. Biol. 14: 633, 1990), альбуміну бразильського горіха (Pearson' et al., Plant Mol. Biol. 18: 235-245, 1992), легуміну (Ellis, et al. Plant Mol. Biol. 10: 203-214, 1988), глютеліну (рис) (Takaiwa, et al., Mol. Gen. Genet. 208: 15-22, 1986; Takaiwa, et al., FEBS Letts. 221: 43-47, 1987), зеїну (Matzke et al., Plant Mol Biol, 143).323-32 1990), napA (Stalberg, et al., Planta 199: 515-519, 1996), SPA пшениці (Albanietal, Plant Cell, 9: 171-184, 1997), олеозину соняшника (Cummins, et al., Plant Mol. Biol. 19: 873-876, 1992)], специфічні для ендосперму промотори [наприклад, LMW і HMW пшениці, глютеніну-1 (Mol Gen Genet 216:81-90, 1989; NAR 17:461-2), гліадинів a, b і g пшениці (EMBO3:1409-15, 1984), промотор ltrl ячменю, гордеїну B1, C, D ячменю (Theor Appl Gen 98:1253-62, 1999; Plant J 4:34355, 1993; Mol Gen Genet 250:750-60, 1996), DOF ячменю (Mena et al., The Plant Journal, 116(1): 53-62, 1998), Biz2 (EP99106056.7), синтетичний промотор (Vicente-Carbajosa et al., Plant J. 13: 629-640, 1998), NRP33 проламіну рису, глобуліну Glb-1 рису (Wu et al., Plant Cell Physiology 39(8) 885-889, 1998), REB/OHP-1 альфа-глобуліну рису (Nakase et al. Plant Mol. Biol. 33: 513-S22, 1997), ADP-глюкози-PP рису (Trans Res 6:157-68, 1997), сімейства генів ESR маїсу (Plant J 12:235-46, 1997), гамма-кафірину сорго (Plant Mol. Biol 32:1029-35, 1996)], специфічні для зародків промотори [наприклад, OSH1 рису (Sato et al., Proc. Nati. Acad. Sci. USA, 93: 81178122), KNOX (Postma-Haarsma of al, Plant Mol. Biol. 39:257-71, 1999), олеозину рису (Wu et at, J. Biochem., 123:386, 1998)] і специфічні для квіток промотори [наприклад, AtPRP4, халкон-синтази (chsA) (Van der Meer, et al., Plant Mol. Biol. 15, 95-109, 1990), LAT52 (Twell et al., Mol. Gen Genet. 217:240-245; 1989), апетала-3; репродуктивних тканин рослини [наприклад, промотори OsMADS (заявка на патент США № 2007/0006344)]. [058] Придатні промотори, що індукуються абіотичним стресом, включають, але без обмеження, промотори, що індукуються сіллю, такі як RD29A (Yamaguchi-Shinozalei et al., Mol. Gen. Genet. 236:331-340, 1993); промотори, що індукуються посухою, такі як промотор гена rab17 маїсу (Pla et. al., Plant Mol. Biol. 21:259-266, 1993), промотор гена rab28 маїсу (Busk et. al., Plant J. 11:1285-1295, 1997) і промотор гена Ivr2 маїсу (Pelleschi et. al., Plant Mol. Biol. 39:373380, 1999); промотори, що індукуються тепловим впливом, такі як промотор білка теплового шоку hsp 80 томата (патент США № 5187267). [059] Мається на увазі, що вираз "ферментативна активність" включає деметилювання, гідроксилювання, епоксидування, N-окиснення, сульфокиснення, N-, S- і O-деалкілування, десульфатацію, деамінування й відновлення азо, нітро й N-оксидних груп. Вираз "нуклеїнова кислота" стосується дезоксирибонуклеотидного або рибонуклеотидного полімеру в одноланцюговій або дволанцюговій формі, або сенсового або антисенсового, і якщо немає інших обмежень, який охоплює відомі аналоги природних нуклеотидів, що гібридизуються з нуклеїновими кислотами способом, подібними такому у нуклеотидів, що зустрічаються в природі. Якщо не зазначене інше, конкретна послідовність нуклеїнової кислоти включає її комплементарну послідовність. [060] "Структурний ген" являє собою таку частину гена, що містить сегмент ДНК, який кодує білок, поліпептид або його частину, та виключає 5′-послідовність, що управляє ініціацією транскрипції. Альтернативно, структурний ген може кодувати продукт, який не транслюється. Структурний ген може являти собою ген, який у нормі зустрічається в клітині, або ген, який у нормі не зустрічається в клітині або місці в клітині, куди його вводять, у випадку чого його 10 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 називають "гетерологічним геном". Гетерологічний ген може походити повністю або частково від будь-якого джерела, відомого в даній галузі техніки, включаючи бактеріальний геном або епісому, еукаріотичну, ядерну або плазмідну ДНК, кДНК, вірусну ДНК або хімічно синтезовану ДНК. Структурний ген може містити одну або декілька модифікацій, які можуть впливати на біологічну активність або її характеристики, біологічну активність або хімічну структуру продукту експресії, швидкість експресії або спосіб контролю експресії. Такі модифікації включають без обмеження мутації, вставки, делеції й заміни одного або декількох нуклеотидів. Структурний ген може бути безперервною кодуючою послідовністю, або він може включати один або декілька інтронів, обмежених відповідними точками сплайсингу. Структурний ген може бути таким, що транслюється, або таким, що не транслюється, в тому числі геном в антисенсовій орієнтації. Структурний ген може бути складеним з сегментів, що отримані з декількох джерел і з декількох генних послідовностей (таких, що зустрічаються в природі, або синтетичних, де синтетичною називається ДНК, яка є хімічно синтезованою). [061] "Отриманий з" використовують у значенні взятий, здобутий, одержаний, виведений, реплікований або такий, що походить із джерела (хімічного та/або біологічного). Похідне можна одержувати за допомогою хімічної або біологічної маніпуляції (включаючи, але без обмеження, заміну, додавання, вставку, делецію, екстракцію, виділення, мутацію та реплікацію) вихідного джерела. [062] "Хімічно синтезована" відносно послідовності ДНК означає, що частини складових нуклеотидів збирали in vitro. Неавтоматизований хімічний синтез ДНК може здійснюватися із застосуванням загальноприйнятих процедур (Caruthers, Methodology of DNA and RNA Sequencing, (1983), Weissman (ed.), Praeger Publishers, New York, Chapter 1); автоматизований хімічний синтез можна проводити із застосуванням одного з ряду комерційно доступних інструментів. [063] Як застосовується в даному документі, "рекомбінантний" включає посилання на клітину або вектор, який був модифікований шляхом уведення гетерологічної нуклеїнової кислоти, або посилання на те, що клітина отримана із клітини, модифікованої в такий спосіб. Таким чином, наприклад, рекомбінантні клітини експресують гени, які не зустрічаються в такій самій формі в нативній (нерекомбінантній) формі клітини, або експресують нативні гени, які в інших випадках експресуються аномально, надекспресуються або не експресуються взагалі внаслідок навмисного втручання людини, або можуть мати знижену або пригнічену експресію нативного гена. Вираз "рекомбінантний", як застосовується в даному документі, не охоплює зміну клітини або вектора за допомогою подій, що зустрічаються в природі (наприклад, спонтанної мутації, природної трансформації/трансдукції/транспозиції), таких як події, що відбуваються без навмисного втручання людини. [064] Як застосовується в даному документі, вираз "касета експресії" являє собою конструкт нуклеїнової кислоти, утворений рекомбінантно або синтетично, з серії певних елементів нуклеїнових кислот, який робить можливою транскрипцію конкретної нуклеїнової кислоти в цільовій клітині. Касета експресії може бути вбудована в плазмідну, хромосомну, мітохондріальну ДНК, пластидну ДНК, фрагмент вірусу або нуклеїнової кислоти. Типово, частина касети експресії у векторі експресії включає серед інших послідовностей нуклеїнову кислоту, що підлягає транскрипції, і промотор. [065] Вирази "залишок", або "амінокислотний залишок", або "амінокислота" використовуються в даному документі взаємозамінно для позначення амінокислоти, яка вбудовується у білок, поліпептид або пептид (узагальнено "білок"). Амінокислота може являти собою амінокислоту, що зустрічається в природі, та, якщо немає інших обмежень, може включати відомі аналоги природних амінокислот, які можуть функціонувати способом, подібним такому в амінокислот, що зустрічаються в природі. [066] Вираз "селективно гібридизується" включає посилання на гібридизацію, за жорстких умов гібридизації, послідовності нуклеїнової кислоти з певною цільовою послідовністю нуклеїнової кислоти в помітно більшому ступені (наприклад, щонайменше в 2 рази в порівнянні з фоновим рівнем), ніж її гібридизація з нецільовими послідовностями нуклеїнової кислоти і з істотним виключенням нецільових нуклеїнових кислот. Послідовності, що селективно гібридизуються, як правило, мають приблизно щонайменше 40 % ідентичність послідовності, переважно 60-90 % ідентичність послідовності та найбільш переважно 100 % ідентичність послідовності (тобто, є комплементарними) одна одній. [067] Вирази “ жорсткі умови" або “ жорсткі умови гібридизації" включають посилання на умови, за яких зонд буде гібридизуватися зі своєю цільовою послідовністю у помітно більшому ступені, ніж з іншими послідовностями (наприклад, щонайменше в 2 рази в порівнянні з фоновим рівнем). Жорсткі умови залежать від послідовності та будуть відрізнятися за різних 11 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 обставин. За допомогою контролю жорсткості умов гібридизації та/або відмивання можна ідентифікувати цільові послідовності, які можуть бути на 100 % комплементарні до зонду (гомологічне зондування). Альтернативно, умови жорсткості можна регулювати задля забезпечення деякої розбіжності у послідовностях, щоб виявляти більш низькі ступені подібності (гетерологічне зондування). Оптимально, зонд має приблизно 500 нуклеотидів у довжину, але його довжина може значно варіювати від менш ніж 500 нуклеотидів до такої, що дорівнює повній довжині цільової послідовності. [068] Як правило, жорсткі умови будуть такими, за яких концентрація солі становить менш ніж приблизно 1,5 M концентрацію іонів Na, типово приблизно 0,01–1,0 M концентрацію іонів Na (або інших солей) при pH 7,0–8,3, а температура становить щонайменше приблизно 30 °C для коротких зондів (наприклад, 10-50 нуклеотидів) і щонайменше приблизно 60 °C для довгих зондів (наприклад, більш ніж 50 нуклеотидів). Жорсткі умови можуть також досягатися шляхом додавання дестабілізуючих засобів, таких як формамід або розчин Денхардта. Ілюстративні умови низької жорсткості включають гібридизацію з буферним розчином із 30-35 % формаміду, 1 M NaCl, 1 % SDS (додецилсульфат натрію) при 37 °C і відмивання в 1× - 2×SSC (20×SSC=3,0 M NaCl/0,3 M тринатрійцитрат) при 50-55 °C. Ілюстративні умови помірної жорсткості включають гібридизацію в 40-45 % формаміді, 1 M NaCl, 1 % SDS при 37 °C і відмивання в 0,5× - 1×SSC при 55-60 °C. Ілюстративні умови високої жорсткості включають гібридизацію в 50 % формаміді, 1 M NaCl, 1 % SDS при 37 °C і відмивання в 0,1×SSC при 60-65 °C. Специфічність типово залежить від постгібридизаційних відмивань, причому критичними факторами є іонна сила й температура кінцевого розчину для відмивання. Для гібридів ДНК-ДНК Tm можна приблизно розрахувати з рівняння Meinkoth and Wahl, (1984) Anal. Biochem., 138:267-84: Tm=81,5 °C+16,6 (log M)+0,41 (% GC)−0,61 (% форм.)--500/L; де M являє собою молярність моновалентних катіонів, % GC являє собою процентне відношення гуанозинових і цитозинових нуклеотидів у ДНК, % форм. являє собою процент формаміду в розчині гібридизації,і L являє собою довжину гібрида у парах основ. Tm являє собою температуру (за певної іонної сили й pH), за якої 50 % комплементарної цільової послідовності гібридизується з абсолютно співпадаючим зондом. Tm знижують на приблизно 1 °C для кожного 1 % розбіжності; таким чином, Tm, умови гібридизації та/або відмивання можна регулювати для гібридизації з послідовностями необхідної ідентичності. Наприклад, якщо бажаними є послідовності з >90 % ідентичністю, то Tm можна знизити на 10 °C. Загалом, жорсткі умови вибирають так, щоб вони були на приблизно 5 °C нижче, ніж температурна точка плавлення (Tm) для конкретної послідовності та комплементарної до неї послідовності при заданій іонній силі й pH. Однак за умов сильної жорсткості можуть використовуватися гібридизація та/або відмивання при температурі на 1, 2, 3 або 4 °C нижче, ніж температурна точка плавлення (Tm); за умов помірної жорсткості можуть використовуватися гібридизація та/або відмивання при температурі на 6, 7, 8, 9 або 10 °C нижче, ніж температурна точка плавлення (Tm); за умов низької жорсткості можуть використовуватися гібридизація та/або відмивання при температурі на 11, 12, 13, 14, 15 або 20 °C нижче, ніж температурна точка плавлення (Tm). Застосовуючи рівняння, склади для гібридизації й відмивання та необхідну Tm, фахівцям буде очевидно, що варіації у жорсткості гібридизації та/або розчинах відмивання практично описані. Якщо необхідний ступінь розбіжності дає в результаті Tm меншу ніж 45 °C (водний розчин) або 32 °C (розчин формаміду), віддають перевагу збільшенню концентрації SSC таким чином, щоб можна було застосувати більш високу температуру. Докладний посібник з гібридизації нуклеїнових кислот можна знайти в Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, part I, chapter 2, "Overview of principles of hybridization and the strategy of nucleic acid probe assays, ” Elsevier, New York (1993); і Current Protocols in Molecular Biology, chapter 2, Ausubel, et al., eds, Greene Publishing and Wiley-Interscience, New York (1995). Якщо не зазначене інше, у даній заявці високу жорсткість визначають як гібридизацію в 4×SSC, 5× розчині Денхардта (5 г фіколу, 5 г полівінілпіролідону, 5 г альбуміну бичачої сироватки в 500 мл води), 0,1 мг/мл кип'яченої ДНК з молок лососевих і 25 мM фосфату натрію при 65 °C і відмивання в 0,1×SSC, 0,1 % SDS при 65 °C. [069] Як застосовується в даному документі, "трансгенна рослина" включає посилання на рослину, яка містить у своєму геномі гетерологічний полінуклеотид. Як правило, гетерологічний полінуклеотид стабільно інтегрований у геном, так що полінуклеотид передається наступним поколінням. Гетерологічний полінуклеотид може бути інтегрований у геном окремо або як частина рекомбінантної касети експресії. "Трансгенний", як застосовується у даному документі, включає будь-яку клітину, клітинну лінію, калюс, тканину, частину рослини або рослину, генотип яких був змінений через наявність гетерологічної нуклеїнової кислоти, включаючи трансгенні організми, початково змінені таким чином, а також трансгенні організми, отримані шляхом 12 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 статевих схрещувань або безстатевого розмноження з початкових трансгенних організмів. Вираз "трансгенний", як застосовується в даному документі, не охоплює зміну генома (хромосомного або позахромосомного) за допомогою традиційних способів селекції рослин або за допомогою подій, що зустрічаються в природі, таких як випадкове перехресне запилення, нерекомбінантна вірусна інфекція, нерекомбінантна бактеріальна трансформація, нерекомбінантна транспозиція або спонтанна мутація. [070] Як застосовується в даному документі, "вектор" включає посилання на нуклеїнову кислоту, яку застосовують у трансфекції клітини-хазяїна, та у яку можна ввести полінуклеотид. Вектори найчастіше є репліконами. Вектори експресії забезпечують транскрипцію нуклеїнової кислоти, уведеної в них. [071] "Надекспресія" стосується рівня експресії в трансгенних організмах, який перевищує рівні експресії в нормальних або нетрансформованих організмах. [072] "Рослинна тканина" включає диференційовані та недиференційовані тканини рослин, включаючи, але без обмеження, корені, стебла, пагони, листки, пилок, насіння, пухлинну тканину й різні форми клітин, а також культури, такі як окремі клітини, протопласт, зародки й калюсна тканина. Рослинна тканина може знаходитись в рослинах або в органі, культурі тканин або клітин. [073] "Переважною експресією", "переважаючою транскрипцією" або "переважною транскрипцією" взаємозамінно називають експресію генних продуктів, які переважно експресуються на більш високому рівні в одній або декількох рослинних тканинах (просторове обмеження) і/або на одній або декількох стадіях розвитку рослини (часове обмеження), тоді як в інших тканинах/стадіях розвитку має місце відносно низький рівень експресії. [074] "Первинним трансформантом" і "поколінням T0" називають трансгенні рослини, які належать до того самого генетичного покоління, що і тканина, яка була першочергово трансформована (тобто такі, що не проходять через мейоз і запліднення після трансформації). "Вторинними трансформантами" і "поколіннями T1, T2, T3 тощо" називають трансгенні рослини, що отримані з первинних трансформантів шляхом одного або декількох мейотичних циклів і циклів запилення. Вони можуть отримуватись шляхом самозапилення первинних або вторинних трансформантів або схрещувань первинних або вторинних трансформантів з іншими трансформованими або нетрансформованими рослинами. [075] "Селективним маркерним геном" називають ген, експресія якого в рослинній клітині надає клітині селективної переваги. Селективна перевага, яку мають клітини, трансформовані за допомогою селективного маркерного гена, може бути обумовлена їхньою здатністю до росту в присутності негативного селективного засобу, такого як антибіотик або гербіцид, у порівнянні зі здатністю до росту нетрансформованих клітин. Селективна перевага, яку мають трансформовані клітини, може також бути обумовлена їхньою посиленою здатністю, у порівнянні з нетрансформованими клітинами, використовувати додану сполуку як поживну речовину, фактор росту або джерело енергії. Селективна перевага, яку має трансформована клітина, може також бути обумовлена втратою гена, який був у неї раніше, в так званій "негативній селекції". При цьому додають сполуку, що є токсичною лише для клітин, які не втратили специфічний ген (негативний селективний маркерний ген), присутній у батьківській клітині (типово, трансген). [076] Вираз "трансформація" стосується перенесення фрагмента нуклеїнової кислоти в геном клітини-хазяїна, що приводить до генетично стабільного успадкування. "Тимчасово трансформованими" називають клітини, у які були введені трансгени й чужорідна ДНК (наприклад, за допомогою таких способів, як опосередкована Agrobacterium трансформація або біолістичне бомбардування), але які не відбирали для стабільного зберігання. "Стабільно трансформованими" називають клітини, які були відібрані й регенеровані на селекційному середовищі після трансформації. [077] "Трансформованим/трансгенним/рекомбінантним" називають організм-хазяїн, такий як бактерія або рослина, у який була введена молекула гетерологічної нуклеїнової кислоти. Молекула нуклеїнової кислоти може бути стабільно інтегрованою в геном хазяїна, або молекула нуклеїнової кислоти також може бути присутня у якості позахромосомної молекули. Така позахромосомна молекула може бути здатна до самореплікації. Передбачається, що трансформовані клітини, тканини або рослини охоплюють не лише кінцевий продукт процесу трансформації, але також їхнє трансгенне потомство. "Нетрансформованим", "нетрансгенним" або "нерекомбінантним" хазяїном називають організм дикого типу, наприклад, бактерію або рослину, яка не містить молекулу гетерологічної нуклеїнової кислоти. [078] Вираз "трансляційна енхансерна послідовність" стосується такої частини послідовності ДНК гена між промотором і кодуючою послідовністю, яка транскрибується в РНК і наявна у 13 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 повністю процесованій мРНК вище (5') трансляційного старт-кодону. Трансляційна енхансерна послідовність може впливати на процесінг первинного транскрипта в мРНК, стабільність мРНК або ефективність трансляції. "Візуально помітним маркером" називають ген, експресія якого не забезпечує перевагу трансформованій клітині, але може зробити так, що її можна виявляти або візуально помічати. Приклади візуально помітних маркерів включають без обмеження βглюкуронідазу (GUS), люциферазу (LUC) і зелений флюоресцентний білок (GFP). [079] "Диким типом" називають нормальний ген, вірус або організм, які зустрічаються в природі без будь-якої мутації або модифікації. [080] Як застосовується в даному документі, "рослинний матеріал", "частина рослини" або "рослинна тканина" означає рослинні клітини, протопласти рослин, тканинні культури рослинних клітин, з яких можна регенерувати рослини, калюси рослин, рослини, що мають спільну кореневу систему, і рослинні клітини, які є інтактними в рослинах або частинах рослин, таких як зародки, пилок, насінні зачатки, насіння, листки, квітки, гілки, плоди, зерна, качани, стрижні кукурудзяних качанів, оболонки, стебла, корені, кореневі кінчики, пиляки, бульби, ризоми й подібне. [081] Як застосовується в даному документі, "білковим екстрактом" називають частковий або загальний білок, екстрагований із частини рослини. Способи екстракції рослинного білка добре відомі в даній галузі техніки. [082] Як застосовується в даному документі, "зразком рослини" називають або інтактну, або неінтактну (наприклад, розмелену тканину насіння або рослини, порубану тканину рослини, ліофілізовану тканину) рослинну тканину. Це також може бути екстракт, що містить інтактну або неінтактну тканину насіння або рослини. [083] Наступні вирази застосовують для опису відношень послідовностей між двома або більшою кількістю нуклеїнових кислот, або полінуклеотидів, або поліпептидів: (a) "еталонна послідовність", (b) "вікно порівняння", (c) "ідентичність послідовностей", (d) "процент ідентичності послідовностей" і (e) "істотна ідентичність". [084] Як застосовується в даному документі, "еталонна послідовність" являє собою задану послідовність, яку застосовують у якості основи для порівняння послідовностей. Еталонна послідовність може являти собою частину певної послідовності або певну послідовність повністю; наприклад, у вигляді сегмента послідовності кДНК або генної послідовності повної довжини або повної послідовності кДНК або генної послідовності. [085] Як застосовується в даному документі, "вікно порівняння" включає посилання на безперервний і визначений сегмент полінуклеотидної послідовності, у якому полінуклеотидна послідовність може порівнюватися з еталонною послідовністю, та де частина полінуклеотидної послідовності у вікні порівняння може містити додавання або делеції (тобто гепи) у порівнянні з еталонною послідовністю (яка не містить додавання або делеції) для оптимального вирівнювання двох послідовностей. Як правило, вікно порівняння становить щонайменше 20 суміжних нуклеотидів у довжину та необов'язково може мати 30, 40, 50 і 100 або більше. Фахівцям у даній галузі зрозуміло, що для уникнення високої подібності з еталонною послідовністю внаслідок включення гепів у полінуклеотидну послідовність зазвичай вводиться штраф за введення гепа й віднімається від числа збігів. [086] В даній галузі техніки добре відомі способи вирівнювання нуклеотидних і амінокислотних послідовностей для порівняння. Алгоритм локальної гомології (BESTFIT) Smith and Waterman, (1981) Adv. Appl. Math 2:482, може здійснювати оптимальне вирівнювання послідовностей для порівняння; за допомогою алгоритму гомологічного вирівнювання (GAP) Needleman and Wunsch, (1970) J. Mol. Biol. 48:443-53; за допомогою способу пошуку за подібністю (Tfasta and Fasta) Pearson and Lipman, (1988) Proc. Natl. Acad. Sci. USA 85:2444; за допомогою комп'ютеризованих реалізацій цих алгоритмів, включаючи, але без обмеження: CLUSTAL у програмі PC/Gene від Intelligenetics, Mountain View, Calif., GAP, BESTFIT, BLAST, FASTA і TFASTA у пакеті програмного забезпечення Wisconsin Genetics, версія 8 (доступна від Genetics Computer Group (програми GCG® (Accelrys, Inc., San Diego, Calif.)). Програма CLUSTAL добре описана Higgins and Sharp, (1988) Gene 73:237-44; Higgins and Sharp, (1989) CABIOS 5:151-3; Corpet, et al., (1988) Nucleic Acids Res. 16:10881-90; Huang, et al., (1992) Computer Applications in the Biosciences 8:155-65; і Pearson, et al., (1994) Meth. Mol. Biol. 24:307-31. Кращою програмою для застосування у оптимальному глобальному вирівнюванні множинних послідовностей є PileUp (Feng and Doolittle, (1987) J. Mol. Evol., 25:351-60, яка аналогічна способу, описаному Higgins and Sharp, (1989) CABIOS 5:151-53, і включена в даний документ за допомогою посилання). Сімейство програм BLAST, які можна застосовувати для пошуків за подібністю в базах даних, включає: BLASTN для нуклеотидних запитуваних послідовностей відносно бази даних нуклеотидних послідовностей; BLASTX для нуклеотидних запитуваних 14 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовностей відносно бази даних білкових послідовностей; BLASTP для білкових запитуваних послідовностей відносно бази даних білкових послідовностей; TBLASTN для білкових запитуваних послідовностей відносно бази даних нуклеотидних послідовностей і TBLASTX для нуклеотидних запитуваних послідовностей відносно бази даних нуклеотидних послідовностей. Див. Current Protocols in Molecular Biology, розділ 19, Ausubel et al., eds., Greene Publishing and Wiley-Interscience, New York (1995). [087] GAP застосовує алгоритм Needleman and Wunsch, вище, для знаходження вирівнювання двох повних послідовностей, яке максимізує число збігів і мінімізує число гепів. GAP розглядає всі можливі вирівнювання й положення гепів і створює вирівнювання з найбільшим числом основ, що збіглися, і найменшою кількістю гепів. Вона дає змогу ввести штраф за введення гепа й штраф за продовження гепа в одиницях основ, що збіглися. GAP повинен отримувати вигоду від числа штрафів за введення гепа за рахунок збігів для кожного гепа, який він уводить. Якщо вибирають штраф за продовження гепа більше ніж нуль, то GAP повинен, крім того, отримувати вигоду від кожного введеного гепа у розмірі довжини гепа, помноженої на штраф за продовження гепа. Значення за замовчуванням штрафу за введення гепа й значення штрафу за продовження гепа у версії 10 пакета програмного забезпечення Wisconsin Genetics становлять 8 і 2, відповідно. Штрафи за введення гепа й продовження гепа можуть виражатися як ціле число, яке вибране з групи цілих чисел, що складається з 0-100. Таким чином, наприклад, штрафи за введення гепа й продовження гепа можуть являти собою 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40 і 50 або більше. [088] GAP є одним членом сімейства найкращих вирівнювань. Може існувати багато членів цього сімейства, але жоден інший член не має кращої якості. GAP відображає чотири показники оцінки вирівнювань: якість, співвідношення, ідентичність і подібність. Якість являє собою показник, максимізований для вирівнювання послідовностей. Співвідношення являє собою якість, поділену на число основ у коротшому сегменті. Відсоток ідентичності являє собою відсоток символів, які фактично збігаються. Відсоток подібності являє собою відсоток символів, які є подібними. Символи, які знаходяться навпроти гепів, не враховують. Подібність підраховують, коли значення у матриці замін для пари символів становить більше або дорівнює 0,50, граничному значенню подібності. Матрицею замін, яка застосовується у версії 10 пакета програмного забезпечення Wisconsin Genetics, є BLOSUM62 (див., Henikoff and Henikoff, (1989) Proc. Natl. Acad. Sci. USA 89:10915). [089] Якщо не зазначене інше, значеннями ідентичності/подібності послідовностей, представленими в даному документі, називають значення, що отримані із застосуванням пакета програм BLAST 2.0 із застосуванням параметрів за замовчуванням (Altschul, et al., (1997) Nucleic Acids Res. 25:3389-402). [090] Як буде зрозуміло фахівцям у даній галузі, пошуки за допомогою BLAST припускають, що білки можуть моделюватися як випадкові послідовності. Проте, більшість існуючих білків містять ділянки з невипадковими послідовностями, які можуть являти собою гомополімерні тракти, короткоперіодичні повтори або ділянки, доповнені однією або декількома амінокислотами. Такі ділянки низької складності можуть вирівнюватися між неспорідненими білками, навіть якщо інші ділянки білка є повністю несхожими. Ряд програм-фільтрів для ділянок низької складності можна застосовувати з метою зменшення таких вирівнювань низької складності. Наприклад, фільтри для низької складності SEG (Wooten and Federhen, (1993) Comput. Chem. 17:149-63) і XNU (Claverie and States, (1993) Comput. Chem. 17:191-201) можна застосовувати окремо або в комбінації. [091] Як застосовується в даному документі, "ідентичність послідовностей" або "ідентичність" у контексті двох послідовностей нуклеїнової кислоти або поліпептидних послідовностей включає посилання на залишки у двох послідовностях, які є однаковими при вирівнюванні для максимальної відповідності на певному вікні порівняння. Коли відсоток ідентичності послідовностей застосовують по відношенню до білків, слід усвідомлювати, що положення залишків, які не є ідентичними, часто відрізняються консервативними амінокислотними замінами, де амінокислотні залишки заміщені на інші амінокислотні залишки з подібними хімічними властивостями (наприклад, зарядом або гідрофобністю) і, внаслідок цього, не змінюють функціональні властивості молекули. Якщо послідовності відрізняються за консервативними замінами, то відсоток ідентичності послідовностей можна регулювати убік підвищення, зробивши поправку на консервативну природу заміни. Про послідовності, що відрізняються за такими консервативними замінами, говорять, що вони мають "подібність послідовностей" або "подібність". Засоби для здійснення такого регулювання добре відомі фахівцям у даній галузі. Типово це включає виставлення балу консервативній заміні як частковій, а не повній розбіжності, тим самим збільшуючи процент ідентичності послідовностей. 15 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 Таким чином, наприклад, якщо ідентичній амінокислоті виставляють бал 1, а неконсервативній заміні виставляють бал нуль, то консервативній заміні виставляють бал від нуля до 1. Виставлення балів консервативним замінам розраховують, наприклад, згідно з алгоритмом Meyers and Miller, (1988) Computer Applic. Biol. Sci. 4:11-17, наприклад, як реалізовано в програмі PC/GENE (Intelligenetics, Маунтін Вью, Каліфорнія, США). [092] Як застосовується в даному документі, "відсоток ідентичності послідовностей" означає значення, яке визначають за допомогою порівняння двох оптимально вирівняних послідовностей у вікні порівняння, де частина полінуклеотидної послідовності у вікні порівняння може містити додавання або делеції (тобто гепи) у порівнянні з еталонною послідовністю (яка не містить додавань або делецій) для оптимального вирівнювання двох послідовностей. Процент розраховують шляхом визначення числа положень, у яких ідентична основа нуклеїнової кислоти або амінокислотний залишок зустрічається в обох послідовностях з одержанням числа положень, що збіглися, ділення числа положень, що збіглися, на загальне число положень у вікні порівняння й множення результату на 100 з одержанням відсотка ідентичності послідовностей. [093] Вираз "істотна ідентичність" полінуклеотидних послідовностей означає, що полінуклеотид містить послідовність, яка має 50-100 % ідентичність послідовностей, переважно щонайменше 50 % ідентичність послідовностей, переважно щонайменше 60 % ідентичність послідовностей, переважно щонайменше 70 %, більш переважно щонайменше 80 %, більш переважно щонайменше 90 % і найбільш переважно щонайменше 95 % у порівнянні з еталонною послідовністю, із застосуванням однієї з описаних програм вирівнювання із застосуванням стандартних параметрів. Фахівцю в даній галузі буде зрозуміло, що ці значення можуть бути відповідним чином скориговані для визначення відповідної ідентичності білків, які кодуються двома нуклеотидними послідовностями, із урахуванням виродженості кодонів, амінокислотної подібності, розташування рамки зчитування й подібного. Істотна ідентичність амінокислотних послідовностей для цих цілей зазвичай означає ідентичність послідовностей, що дорівнює 55-100 %, переважно щонайменше 55 %, переважно щонайменше 60 %, більш переважно щонайменше 70 %, 80 %, 90 % і найбільше переважно щонайменше 95 %. [094] Іншою ознакою того, що нуклеотидні послідовності є суттєво ідентичними, є гібридизація двох молекул одна з одною за жорстких умов. Виродженість генетичного коду забезпечує більшість амінокислотних замін, що ведуть до різноманітності в нуклеотидній послідовності, що кодує одну й ту саму амінокислоту, тому не виключено, що послідовність ДНК може кодувати такий самий поліпептид, але не гібридизуватися одна з одною за жорстких умов. Це може відбуватися, наприклад, коли копія нуклеїнової кислоти створюється із застосуванням максимальної виродженості кодону, дозволеної генетичним кодом. Одна ознака того, що дві послідовності нуклеїнової кислоти є суттєво ідентичними, заключається в тому, що поліпептид, який кодує перша нуклеїнова кислота, є імунологічно перехресним з поліпептидом, який кодує друга нуклеїнова кислота. [095] Фраза "абіотичний стрес", як застосовується в даному документі, стосується будь-якого несприятливого ефекту на метаболізм, ріст, розмноження та/або життєздатність рослини з боку абіотичних факторів (тобто водозабезпеченості, теплового впливу, холоду і т.д.). Відповідно, абіотичний стрес може бути викликаний субоптимальними навколишніми умовами зростання, такими як, наприклад, засоленість, водне голодування, водний дефіцит, посуха, повінь, заморожування, низька або висока температура (наприклад, охолодження або надмірне нагрівання), забруднення токсичними хімічними речовинами, токсичність важких металів, анаеробіоз, недостатність поживних речовин, надлишок поживних речовин, атмосферне забруднення або УФ-опромінення. [096] Фраза "стійкість до абіотичного стресу", як застосовується в даному документі, стосується здатності рослини витримувати абіотичний стрес, не зазнаючи істотної зміни в метаболізмі, рості, продуктивності та/або життєздатності. [097] Як застосовується в даному документі, "водний дефіцит" означає період, коли вода, що є доступною для рослини, не поповнюється зі швидкістю, з якою вона поглинається рослиною. Тривалий період водного дефіциту в розмовній мові називається посухою. Відсутність дощу або зрошення миттєво може не призводити до стресу, викликаного нестачею води, якщо є доступний запас ґрунтових вод для підтримки швидкості росту рослин. Рослини, які вирощують в ґрунті з достатнім запасом ґрунтових вод, можуть переживати дні без дощу або зрошення без несприятливих впливів на врожайність. Рослини, які вирощують в сухому ґрунті, у випадку мінімальних періодів водного дефіциту, ймовірно, зазнають несприятливих впливів. Стрес, викликаний важким водним дефіцитом, може спричинити зів'янення й загибель рослин; помірна посуха може знижувати врожайність, заримувати ріст або стримувати розвиток. Рослини можуть 16 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 відновлюватися після декількох періодів стресу, викликаного водним дефіцитом, без істотного впливу на врожайність. Однак водний дефіцит під час запилення може зменшувати або знижувати врожайність. Таким чином, корисним періодом у життєвому циклі кукурудзи, наприклад, для спостереження за реакцією або стійкістю до водного дефіциту, є пізня вегетативна стадія росту перед появою волоті або перехід до розвитку репродуктивних органів. Стійкість до водного дефіциту визначають шляхом порівняння з контрольними рослинами. Наприклад, рослини за даним винаходом можуть продукувати більш високий урожай, ніж контрольні рослини за умови впливу водного дефіциту. У лабораторії та у польових випробуваннях посуху можна імітувати шляхом надання рослинам за даним винаходом й контрольним рослинам меншої кількості води, ніж надають контрольним рослинам з достатнім рівнем поливу, й визначення відмінностей в ознаках. Наприклад, урожайність можна розділити на такі компоненти як рослин на акр, кількість рядів зерен на качан, кількість качанів на рослину, кількість зерен на рослину й вага на зерно. Безпліддя або зменшене утворення насіння може також підвищуватися за умов стресу, викликаного нестачею води. Один аспект даного винаходу передбачає рослини, які надекспресують гени, розкриті в даному документі, що забезпечують більш високу стійкість до водного дефіциту. [098] Як застосовується в даному документі, фраза "оптимізація води" стосується будь-якого показника рослини, його частин або його структури, який можна виміряти та/або кількісно визначати, щоб оцінити ступінь або швидкість росту й розвитку рослин за різних умов водозабезпеченості. У зв'язку із цим, "ознака оптимізації води" являє собою будь-яку ознаку, яка, як можна показати, впливає на врожайність рослини за різних поєднань умов росту, повʼязаних з водозабезпеченістю. Ілюстративні показники оптимізації води являють собою врожайність зерна за стандартного відсотка вологості (YGSMN), вологість зерна при збиранні (GMSTP), вагу зерна на ділянку (GWTPN) і процентне відновлення врожаю (PYREC). [099] Як застосовується в даному документі, фрази "посухостійкість" і "посухостійкий" стосуються здатності рослин витримувати та/або успішно розростатися за умов, коли водозабезпеченість є субоптимальною. Як правило, рослину відмічають як "посухостійку", якщо вона проявляє "посилену посухостійкість". Як застосовується в даному документі, фраза "посилена посухостійкість" стосується вимірюваного поліпшення, посилення або збільшення одного або декількох фенотипів оптимізації води в порівнянні з однією або декількома контрольними рослинами. [0100] Коефіцієнт використання води (WUE) являє собою параметр, який часто застосовують для оцінки співвідношення між поглинанням води й уловлюванням CO2/ростом (Kramer, 1983, Water Relations of Plants, Academic Press p. 405). WUE визначали і вимірювали численними способами. Один підхід полягає в обчисленні відношення сухої ваги цілої рослини до ваги води, яку рослина поглинала протягом свого життя (Chu et al., 1992, Oecologia 89:580). Інший варіант полягає в застосуванні більш короткого часового інтервалу, коли вимірюються накопичення біомаси й використання води (Mian et al., 1998, Crop Sci. 38:390). Інший підхід полягає у використанні вимірювань від окремих частин рослини, наприклад, вимірювання росту і використання води лише надземних частин (Nienhuis et al 1994 Amer J Bot 81:943). WUE також був визначений як відношення уловлювання CO2 до втрати води листком або частиною листка за рахунок випаровування, що часто вимірюється протягом дуже короткого періоду часу (наприклад, секунди/хвилини) (Kramer, 1983, p. 406). Відношення 13C/12C, зафіксованого в рослинній тканині та виміряного за допомогою мас-спектрометра для вимірювання співвідношення ізотопів, також застосовувалося для оцінки WUE у рослин, які застосовують C-3 фотосинтез (Martin et al., 1999, Crop Sci. 1775). Як застосовується в даному документі, вираз "коефіцієнт використання води" стосується кількості органічних речовин, продукованих рослиною, поділеної на кількість води, використаної рослиною при їх продукуванні, тобто сухої ваги рослини по відношенню до використання рослиною води. Як застосовується в даному документі, вираз "суха вага" стосується будь-чого у рослині, за винятком води, і включає, наприклад, вуглеводи, білки, олії й мінеральні поживні речовини. Передбачається, що трансгенні рослини, отримані за допомогою способів, описаних у даному документі, будуть забезпечувати збільшення коефіцієнта використання води. [0101] Фраза "біотичний стрес", як застосовується в даному документі, стосується будь-якого несприятливого ефекту на метаболізм, ріст, розмноження та/або життєздатність рослини з боку біотичних факторів (тобто тиск з боку комах, захворювання тощо). [0102] Фраза "стійкість до біотичного стресу", як застосовується в даному документі, стосується здатності рослини витримувати біотичний стрес, не зазнаючи істотної зміни у метаболізмі, рості, розмноженні та/або життєздатності. 17 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 [0103] Як застосовується в даному документі, фраза "рослинна біомаса" стосується кількості (яку вимірюють в грамах повітряно-сухої або сухої тканини) тканини, отриманої від рослини у вегетаційний період, яка може також визначати або впливати на врожайність рослин або врожайність на площу вирощування. [0104] Як застосовується в даному документі, фраза "міць рослин" стосується кількості (яку вимірюють за вагою) тканини, продукованої рослиною за визначений час. Отже, збільшена міць може визначати або впливати на врожайність рослин або врожайність за час вирощування або на площі вирощування. [0105] Вираз "рання міць" стосується активного здорового добре збалансованого росту особливо під час ранніх стадій росту рослини, і вона може бути результатом кращої пристосованості рослини внаслідок того, наприклад, що рослини є краще адаптованими до свого навколишнього середовища (оптимізують використання джерел енергії й розподіляють між пагоном й коренем). Рослини, що мають ранню міць, також демонструють підвищене виживання сходів і краще вкорінення сільськогосподарської культури, що часто приводить до вельми однорідних полів (наприклад, сільськогосподарські культури, що ростуть однаковим чином, а саме, сільськогосподарські культури, що досягають різних стадій розвитку в фактично один і той самий час) і часто до більш високих урожаїв. Отже, ранню міць можна визначати за допомогою вимірювання різних факторів, таких як вага тисячі зерен, відсоткове проростання, відсоток появи сходів, ріст сходів, висота сходів, довжина кореня, біомаса кореня й пагона та багато іншого. [0106] Як застосовується в даному документі, "міць сходів" стосується характеристики рослини, згідно якої рослина з'являється з ґрунту швидше, має підвищену швидкість проростання (тобто, проростає швидше), має більш швидкий і більш масштабний ріст сходів і/або проростає швидше в умовах холоду в порівнянні з диким типом або контролем за аналогічних умов. Міць сходів часто визначали як таку, що включає властивості насіння, які визначають "потенціал для швидкої, однорідної появи й розвитку нормальних сходів у широкому діапазоні польових умов". [0107] Життєвий цикл квітучих рослин в цілому можна розділити на три фази росту: вегетативну фазу, фазу бутонізації й фазу цвітіння (пізня фаза бутонізації). У вегетативній фазі апікальна меристема пагона (SAM) утворює листки, які пізніше будуть забезпечувати джерела, необхідні для створення фертильного потомства. Після одержання відповідних сигналів від навколишнього середовища й сигналів стадії розвитку рослина переходить до росту квітки, або до репродуктивного росту, і SAM вступає у фазу бутонізації (I) і дає початок суцвіттю з зачатками квіток. Під час цієї фази напрямок розвитку SAM і вторинних пагонів, які виникають у пазухах листків, визначається набором генів ідентичності меристем, деякі з яких запобігають, а деякі з яких стимулюють розвиток квіткових меристем. Як тільки вони з'являються, рослина входить у пізню фазу бутонізації, коли формуються органи квітки. Якщо присутні відповідні сигнали навколишнього середовища й сигнали стадії розвитку, рослина переходить до росту квітки, або репродуктивного росту. Якщо такі сигнали порушуються, рослина не буде здатною розпочинати репродуктивний ріст, а отже, підтримуватиме вегетативний ріст. [0108] Термін "ідіоплазма" стосується генетичного матеріалу особини (наприклад, рослини), групи особин (наприклад, лінії, сорту або родини рослин), або клону, отриманого з лінії, сорту, виду або культури. Ідіоплазма може бути частиною організму або клітини, або може бути відділена від організму або клітини. Як правило, ідіоплазма забезпечує генетичний матеріал зі специфічним молекулярним складом, який забезпечує фізичну основу для деяких або всіх спадкових якостей організму або культури клітин. Як застосовується в даному документі, "ідіоплазма" включає клітини, насіння або тканини, з яких можуть вирости нові рослини або частини рослин, такі як листки, стебла, пилок або клітини, з яких можна виростити цілу рослину. [0109] Антитіла за даним винаходом включають поліклональні й моноклональні антитіла та їхні суміші, які можуть являти собою будь-які з IgG, IgA, IgM, IgE, IgD і будь-який їхній ізотип, наприклад, IgG1, IgG2, IgG3 або IgG4. У випадку моноклонального антитіла ілюстративним класом антитіл є IgG. Підкласи IgG включають, наприклад, IgG1, IgG2, IgG3 і IgG4. Антитіла включають інтактні й химерні молекули імуноглобулінів з двома важкими ланцюгами повної довжини й двома легкими ланцюгами повної довжини (наприклад, зріла частина послідовностей варіабельної ділянки важкого й легкого ланцюгів), а також підпослідовності/фрагменти важкого або легкого ланцюга, які зберігають щонайменше частково функцію (наприклад, специфічність зв'язування T6PP або афінність зв'язування T6PP) батьківського інтактного антитіла, що специфічно зв'язує T6PP і, зокрема, модифіковану T6PP. Підпослідовності можуть мати таку ж або практично таку саму специфічність зв'язування, афінність зв'язування, як і батьківські інтактні й химерні антитіла. 18 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 [0110] Одержання моноклональних або поліклональних антитіл може здійснюватися за допомогою методик, які добре відомі у даній галузі техніки. "Поліклональні" антитіла являють собою антитіла, отримані з різних B-клітинних джерел. Вони є комбінацією молекул імуноглобулінів, які секретують специфічний антиген, кожний з яких ідентифікує окремий епітоп. Поліклональні антитіла типово одержують за допомогою інокуляції придатного ссавця, такого як миша, кролик або коза. Наприклад, композицію, що містить білок T6PP, який здатний продукувати антиген, ін'єкційно вводять ссавцю. Присутність нового білка індукує B-лімфоцити продукувати імуноглобуліни IgG, специфічні до антигену T6PP. Поліклональні імуноглобуліни IgG потім очищають з сироватки ссавця. [0111] Вираз "моноклональное антитіло", як застосовується в даному документі, стосується препарату з молекул антитіл з одним молекулярним складом. Моноклональне антитіло проявляє одну специфічність і афінність зв'язування до конкретного епітопу. Спосіб одержання моноклональних антитіл включає одержання імунних соматичних клітин з потенціалом до продукування антитіл, зокрема, B-лімфоцитів, які були попередньо імунізовані за допомогою антигену, що становить інтерес, in vivo або in vitro і які є придатними для злиття з лінією Bклітинної мієломи. [0112] Лімфоцити ссавців типово імунізують шляхом in vivo імунізації тварини (наприклад, миші) за допомогою необхідного білка або поліпептиду, наприклад, за допомогою T6PP за даним винаходом. Такі імунізації повторюють при необхідності з інтервалами до декількох тижнів для одержання достатнього титру антитіл. Після імунізації тварини можна застосовувати в якості джерела лімфоцитів, що продукують антитіла. Після останньої бустер-ін'єкції антигену тварин умертвляють і видаляють клітини селезінки. Лімфоцити мишей забезпечують більш високий відсоток стабільного злиття з лініями мієломи миші, які описані в даному документі. З них миша BALB/c є переважною. Проте, інші лінії мишей, кролика, хом'яка, вівцю й жабу також можна застосовуватися в якості хазяїв для одержання клітин, що продукують антитіла. Див.; Goding (в Monoclonal Antibodies: Principles and Practice, 2d ed., pp. 60-61, Orlando, Fla., Academic Press, 1986). [0113] Переважно зливаються такі клітини, що продукують антитіла, які перебувають на стадії плазмобласта, що ділиться. Соматичні клітини можна отримувати з лімфатичних вузлів, селезінок і периферичної крові тварин, які підлягали первинному впливу антигена, а переважні лімфатичні клітини залежать великою мірою від їхньої емпіричної ефективності в конкретній системі злиття. Лімфоцити, що секретують антитіла, потім піддають злиттю з (мишачими) клітинами B-клітинної мієломи або трансформованими клітинами, які здатні до необмеженого розмноження в культурі клітин, з одержанням, тим самим, безсмертної клітинної лінії, що секретує імуноглобулін. Отримані злиті клітини, або гібридоми, культивують і отримані колонії піддають скринінгу щодо продукування необхідних моноклональних антитіл. Колонії, що продукують такі антитіла, клонують і вирощують in vivo або in vitro для одержання великих кількостей антитіла. Опис теоретичної основи й практичної методології злиття таких клітин викладений в Kohler and Milstein, Nature 256:495 (1975), яка включена в даний документ за допомогою посилання. [0114] Збільшена врожайність сільськогосподарської культури являє собою ознаку, що представляє значний економічний інтерес в усьому світі. Врожайність зазвичай визначають як вимірюваний підсумок економічної цінності сільськогосподарської культури. Вона може визначатися у формі кількості та/або якості. Врожайність безпосередньо залежить від декількох факторів, наприклад, числа й розміру органів, структури рослини (наприклад, числа гілок), утворення насіння, старіння листків та іншого. Розвиток кореня, поглинання поживних речовин, стресостійкість і рання міць також можуть бути важливими факторами при визначенні урожайності. Крім того, у сільському господарстві дуже бажаною є розробка сільськогосподарських культур, які можуть демонструвати збільшену врожайність за оптимальних умов росту, а також за неоптимальних умов росту (наприклад, при посусі, в умовах абіотичного стресу). Таким чином, оптимізація вищезгаданих факторів може вносити вклад у підвищення врожайності сільськогосподарських культур. [0115] Урожайність насіння є особливо важливою ознакою, оскільки насіння багатьох рослин важливе для харчування людей і тварин. Сільськогосподарські культури, такі як кукурудза, рис, пшениця, канола і соя, забезпечують більше половини загальної кількості калорій, які споживає людина, або через пряме споживання насіння як такого, або через споживання сільськогосподарських тварин, вигодуваних переробленим насінням. Насіння рослин також є джерелом цукрів, олій і багатьох видів метаболітів, які застосовуються у різних промислових процесах. Насіння складається з зародка (джерела нових пагонів і коренів) і ендосперму (джерела поживних речовин для росту зародка під час проростання та під час раннього росту 19 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 сходів). До розвитку насіння залучені багато генів, і він потребує перенесення метаболітів від кореня, листків і стебел до насіння, що розвивається. Ендосперм асимілює метаболічні попередники вуглеводів, олій і білків і синтезує з них запасні макромолекули для наливання зерна. [0116] Рання міць являє собою інший важливий аспект урожайності. Поліпшення ранньої міці являє собою важливу мету сучасних програм селекції рису в культиварів рису як помірного, так і тропічного пояса. Довгі корені важливі для надійного закріплення в ґрунті рису, що висаджується у воду. Рис, що висаджується у воду, висівають безпосередньо на затоплені поля, де рослини повинні швидко з'явитися з води, тому більш довгі пагони забезпечують збільшене виживання молодих сходів. Якщо практикують посів рядками, для задовільної появи сходів важливі більш довгі мезокотилі та колеоптилі. Можливість розробки у рослин ранньої міці матиме велике значення в сільському господарстві. Наприклад, слабка рання міць стала обмеженням для введення гібридів маїсу (Zea mays L.), які основані на ідіоплазмі з Кукурудзяного пояса США в Атлантичній частині Європи. В іншому аспекті рання міць може також забезпечувати більш ранні періоди збирання деяких сільськогосподарських культур, що може служити перевагою в багатьох сільськогосподарських застосуваннях. [0117] Рослини, розроблені для поліпшеної врожайності за умов різних біотичних і абіотичних стресів, становлять особливий інтерес в галузі сільського господарства. Наприклад, абіотичний стрес є основною причиною втрати врожаю в усьому світі, знижуючи середні рівні врожайності найбільш важливих культурних рослин більш ніж на 50 % (Wang et al., Planta (2003) 218: 1-14). Абіотичні стреси можуть спричинятися посухою, повенями, засоленістю, екстремальними значеннями температури, хімічною токсичністю й окисним стресом. Можливість поліпшення стійкості рослин до абіотичного стресу буде мати велику економічну перевагу для фермерів в усьому світі та буде забезпечувати культивування сільськогосподарських культур під час несприятливих умов, а також на територіях, де культивування сільськогосподарських культур іншим способом неможливе. [0118] У деяких випадках урожайність рослини визначають відносно кількості рослинної біомаси, яку може продукувати конкретна рослина. Більша рослина з більшою площею листків, як правило, може поглинати більше світла, поживних речовин і вуглекислого газу, ніж менша рослина, а отже, ймовірно, буде прибавляти більшу вагу протягом такого самого періоду (Fasoula & Tollenaar 2005 Maydica 50:39). Збільшена рослинна біомаса може також бути дуже бажаною в таких процесах, як перетворення біомаси (наприклад, кукурудзи, трав, сорго, очерету) на паливо, таке як, наприклад, етанол або бутанол. [0119] Можливість збільшення врожайності рослин матиме багато застосувань у таких галузях, як сільське господарство, одержання декоративних рослин, деревництво, садівництво, одержання біопалива, фармацевтика, ферментна промисловість, де використовують рослини як фабрики для цих молекул, і лісівництво. Збільшення врожайності може також знаходити застосування в одержанні мікробів або водоростей для застосування в біореакторах (для біотехнологічного одержання речовин, таких як фармацевтичні препарати, антитіла, вакцини, паливо, або для біологічного перетворення органічних відходів) та інших подібних галузях. [0120] Селекціонери рослин часто зацікавлені у поліпшенні специфічних аспектів урожайності залежно від досліджуваної сільскогосподарської культури або рослини, а також частини цієї рослини або сільскогосподарської культури, які мають відносну економічну цінність. Наприклад, селекціонер рослин може очікувати конкретно на поліпшення рослинної біомаси (ваги) однієї або декількох частин рослини, які можуть включати надземні (придатні для збирання) частини та/або придатні для збирання частини під землею. Це особливо доречно там, де споживче значення мають надземні частини або підземні частини рослини. Для багатьох сільськогосподарських культур, особливо злаків, дуже бажане поліпшення врожайності насіння. Збільшена врожайність насіння може проявлятися багатьма шляхами, причому кожний окремий аспект урожаності насіння має різну важливість для селекціонера рослин залежно від досліджуваної сільськогосподарської культури або рослини та її кінцевого застосування. [0121] Для селекціонера рослин була б великою перевагою можливість підібрати та вибрати аспекти врожайності, які підлягають зміні. Також може бути дуже бажаною можливість підібрати ген, що підходить для зміни конкретного аспекту врожайності (наприклад, урожайності насіння, ваги біомаси, коефіцієнта використання води, врожайності при стресових умовах). Наприклад, збільшення швидкості наливу у комбінації зі збільшеною вагою тисячі зерен буде дуже бажаною для сільськогосподарської культури, такої як кукурудза. Для рису й пшениці буде дуже бажаною комбінація збільшеної швидкості наливу, процентного відношення маси врожаю до загальної маси рослин і збільшеної ваги тисячі зерен. 20 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 [0122] Трегалоза являє собою дуже стабільний дисахарид, що складається з двох молекул глюкози. Вона не розпадається при нагріванні до 100˚C протягом 24 годин у широкому діапазоні pH. Її низька реакційна здатність, фізіохімічні властивості й стабільність роблять її винятковим осмопротектором. За відсутності води вона стабілізує білки, мембрани та інші клітинні структури, заміщуючи воду шляхом утворення водневих зв'язків з полярними залишками. Після висушування трегалоза утворює сітчасту склоподібну структуру, яка обмежує рух молекул, що запобігає агрегації білків і дифузії вільних радикалів (Brumfield G (2004) Nature 428:14-15, Paul MJ (2004) Plant Biotechnol J, 2 pp. 71–82). [0123] Раніше вважалося, що лише певні рослини, такі як стійкі до висушування види, наприклад, рослини, що відроджуються, накопичують трегалозу, оскільки вони роблять це в значних кількостях, які легко виміряти. Проте, нещодавні генетичні й геномні дані показують, що всі рослини, досліджені до теперішнього часу, мають здатність синтезувати трегалозу, але, як правило, в дуже малих кількостях (Paul MJ, et al. (2008) Annu Rev Plant Biol 59:417-441). Більше того, виявилося, що метаболізм трегалози є вкрай важливим, тому що позбавлення рослини здатності продукувати трегалозу, наприклад, за допомогою мутації, є летальним (Eastmond PJ, et al. (2002) Plant J 29:225-235). Більшість видів сільськогосподарських культур містять слідові кількості трегалози (які вимірюються на низькому мікромолярному або навіть наномолярному рівні), хоча характерне для маїсу накопичення є невідомим. [0124] "Трегалозний шлях", як застосовується в даному документі, складається із двох біосинтетичних ферментів, трегалоза-6-фосфатсинтази (TPS) і трегалоза-6-фосфатфосфатази (T6PP), та одного гідролізуючого ферменту, трегалази, залученого у розпад, див. Фіг. 3. На даний час невідомо, скільки ізоформ (TPS) або (T6PP) існує у маїсі. У модельній рослині Arabidopsis загалом налічується 11 генів TPS (Paul et al. (2008) Annu Rev Plant Biol 59:417-441), проте, каталітична активність була показана лише для AtTPS1 (Vandesteene L, et al. (2010) Mol Plant 3:406-419). Функція інших 10 гомологів TPS Arabidopsis залишається незрозумілою. В Arabidopsis існує 10 генів T6PP, і хоча більшість не були добре охарактеризовані, усі вони, ймовірно, мають T6PP активність (Schluepmann H, et al. (2012) J Exp Bot:1-12; Paul et al., (2008) Annu Rev Plant Biol 59:417-441). Навпаки, Arabidopsis містить лише один ген, що кодує трегалазу (Müller J, et al. (2001) Plant Physiol 125:1086-1093; Frison M, et al. (2007) FEBS Letters 581:40104016; Paul et al. 2008). Аналогічна ситуація спостерігається у рису (Shima S, et al. (2007) FEBS J 274:1192-1201; Zang et al, (2011) Plant Mol Biol 76:507-522). Дані визначення профілів експресії показують, що члени сімейства генів диференційно експресуються в ході розвитку рослини (Scheulpmann et al., (2004) Plant Physiol 135:879-890; Zang et al., (2011) Plant Mol Biol 76:507522). Різноманітність тканиноспецифічної експресії й трансляції серед деяких членів сімейств генів T6PP і TPS представляє явний доказ того, що ці ферменти здійснюють важливі й, можливо, різні функції в рослині (Satoh-Nagasawa et al., (2006) Nature 441:227-230; Mustroph et al., (2009) Proc Natl Acad Sci USA 106:18843-18848; Ramon et al., (2009) Plant Cell Environ 32:1015-1032). [0125] Ферменти трегалозного шляху також регулюються на посттрансляційному рівні. Білки TPS являють собою субстрати для споріднених з неферментуючими сахарозу (SNF1) протеїнкіназами (SnRK1) (Zhang et al., (2009) Plant Physiol 149:1860-1871). Деякі фосфопротеїни TPS (AtTPS5, 6 & 7) зв'язують 14-3-3 білки (Harthill et al., (2006) Plant J 47:211-223). Координаційне фосфорилювання й кепування сайту фосфорилювання за допомогою 14-3-3 білків можуть являти собою можливий механізм, за допомогою якого регулюється активність TPS. Нітратредуктаза регулюється за допомогою аналогічного механізму, який базується на переході світло/темрява (Huber et al., (2002) Plant Mol Biol 50:1053-1063). Harthill et al. припускають, що усі з нітратредуктази, 6-фосфофрукто-2-кіназа/фруктоза-2,6-бісфосфатази (F2KP) і TPS модифікуються за допомогою однієї кінази (імовірно, SnRK1), зв'язують 14-3-3 білки та у деякій мері можуть регулюватися циклом світло/темрява (Harthill et al. (2006) Plant J 47:211-223). [0126] Складність в експресії й регуляції дає вагомі підстави припускати, що білки в шляху метаболізму трегалози мають бути згруповані з ферментами, важливими для метаболізму рослини, які виконують як каталітичні, так і регуляторні функції. Їхня точна роль повністю не зрозуміла, але перспективну модель можна асоціювати з трьома головними функціями: передача сигналу за участю цукрів, розвиток рослин і захист від стресу. [0127] Ґрунтуючись на опублікованих повідомленнях стосовно мікробіологічних систем (Wiemken, A. (1990) J Gen Microbiol 58:209-217), в яких стабілізація мембран та інші переваги корелюють із високими рівнями трегалози (1-10 % сухої ваги, г/г), ранні роботи з генетичної модифікації фокусувались на застосуванні трегалози в якості осмопротектора. Деякі опубліковані дослідження припускали, що трегалоза захищає рослини від посухи та інших 21 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 осмотичних стресів (Garg et al., (2002) PNAS 99:15898-15903; Paul et al., (2008) Annu Rev Plant Biol 59:417-441; Ge et al., 2008; Li et al., 2011). Проте, коли рівні трегалози вимірювали у трансгенних рослинах, розроблених для надмірного накопичення трегалози, було виявлено, що вони були дуже низькими, близько мікрограмів на грам сирої ваги. Філогенетична інформація також показує, що незважаючи на поширення в рослинах генів трегалозного шляху, лише невелике число рослин накопичує метаболіти трегалози на рівнях, необхідних для безпосередньої дії в якості осмопротектора або стабілізатора клітинних структур. [0128] Діяльність у середині 1990-их з одержання трегалози в рослинах (для харчової промисловості) відрізнялася від діяльності з одержання у рослинах молекул інших "чужорідних" цукрів (Eastmand and Graham, (2003) Current Opin Plant Biol 6:231-235; Pellny et al. (2004) Plant Biotechnol J, 2 pp. 71–82; Paul M.J. (2007) Current Opinion in Plant Biology Volume 10, Issue 3, June, Pages 303–309). Зміна метаболізму трегалози в трансгенних рослинах часто призводила до неочікуваних плейотропних дефектів росту (Lordachescu and Imai, (2008) J Integrative Plant Biol 50:1223-1229). [0129] P.J. Eastmond et al. першими показали незаперечну потребу в активному гені TPS рослини, TPS1 Arabidopsis, для розвитку зародка (Eastmond P.J. et al., (2002) Plant J 29:225235). Гомозиготні tps1 мутанти не розвивалися до зрілого насіння. Schluepmann et al. показали, що tps1 мутант може бути врятований завдяки експресії гена otsA E. coli, що кодує трегалоза-6фосфатсинтазу, що дозволяє припустити, що T6P є активним компонентом трегалозного шляху, який регулює метаболізм рослини. Концентрація T6P у рослинних клітинах є дуже низькою, що доволяє припустити, що вона функціонує в якості сигнальної молекули (Scheulpmann et al., (2004) Plant Physiol 135:879-890). Невелика зміна в її концентрації може мати істотний "гормоноподібний" ефект на регуляцію метаболічних процесів у рослині (Rolland et al. (2002) Plant Cell:S185-S205). [0130] У нинішній час існує зростаюча кількість доказів для підтвердження залучення трегалозного шляху в передачу сигналів за участю цукрів, що є необхідним для всіх організмів. Це забезпечує регуляцію рівнів цукрів між крайніми значеннями великої кількості й нестачі (Eveland and Jackson, (2012) J Exp Bot:1-11). Наприклад, у ссавців регуляція рівнів цукру в крові за допомогою інсуліну є необхідною для підтримання оптимального стану здоров'я. [0131] Концепція, згідно якої трегалозні цукри залучені в передачу сигналу цукрами й енергетичний статус, з'ясована у мікробів, зокрема дріжджів, і в цей час підтверджується у рослин (Schluepmann et al., (2012) J Exp Bot:1-12). У рослин T6P синтезується з G6P і UDPG, подальших продуктів розпаду сахарози. UDPG одержують безпосередньо за допомогою сахарозосинтази; G6P одержують з глюкози за допомогою інвертази й гексокінази або за допомогою фруктози й фруктоза-6-фосфату (F6P) за допомогою сахарозосинтази, фруктокінази та фосфоглюкозаізомерази. UDPG і G6P також являють собою два головні активовані попередники, з яких, зрештою, можуть бути отримані багато клітинних функціональних сполук (фігура 4). UDPG являється попередником для основних полісахаридів клітинної стінки та для гліколіпідів. G6P являється попередником для синтезу крохмалю, NADPH шляхом окисної частини пентозофосфатного шляху (Masakapalli S.K. et al., (2010) Plant Physiol 152:602-619), і ATP шляхом гліколізу, циклу Кребса й окисного фосфорилювання. Крім того, G6P може бути перетворений на фруктоза-6-фосфат і, разом з UDPG, може застосовуватися для синтезу сахароза-6-фосфату, а потім сахарози. Будучи зробленим з UDPG і G6P, T6P перебуває на перетинах основних вуглеводних потоків у рослинах. Таким чином, рівні T6P у рослинних клітинах можуть відображати доступність гексозофосфатів, UDPG і сахарози. [0132] Важливе відкриття показує, що T6P зв'язується з SnRK1 з інгібуванням її активності (Zhang et al., 2009) Plant Physiol 149:1860-1871), див. Фіг. 5. SnRK1 являє собою гетеротримерний білок та є рослинним гомологом протеїнкінази тварин, що активується AMP, та протеїнкінази дріжджів, що не ферментує сахарозу (SnF1) (Paul et (2008) Annu Rev Plant Biol 59:417-441; Schluepmann et al., (2012) J Exp Bot:1-12; Polge and Thomas (2007) Trends in Plant Science Volume 12, Issue 1, January 2007, Pages 20–28). Глобальна генна експресія в рослинах, які надекспресують ген субодиниці SnRK1, нагадує реакцію рослини на стрес, викликаний нестачею вуглецевої поживної речовини, або на енергетичний стрес, спричинені тривалим темновим періодом, інгібуванням фотосинтеза або зануренням у воду (Baena-González et al., (2007) Nature 448:938-942). Таким чином, збільшений рівень SnRK1 опосередковує реакції на стрес, викликаний низькою енергією. У той же час інактивація SnRK1 стимулює процеси, необхідні для утилізації та анаболізму вуглецю (фігура 6). [0133] В Arabidopsis T6P був опосередковано асоційований NADPH-залежними процесами (Kolbe et al., (2005) PNAS 102:11118-11123). Рівні T6P в Arabidopsis мають тенденцію до обернено-пропорційної кореляції з рівнями G6P (Schleupmann et al.,(2003) Proc Natl Acad Sci 22 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 USA 100:6849-6854) і прямої кореляції з рівнями сахарози (Martinez-Barajas et al., 2011). В Arabidopsis сахароза або трегалоза, що застосовуються для живлення, збільшують рівні T6P і активують включення вуглецю в крохмаль, що являє собою використання вуглецю для запасання. T6P залучений в окисно-відновну активацію AGPази в гетеротрофних тканинах (Schleupmann et al., (2012) J Exp Bot:1-12). Крім того, AGPаза істотно активується за допомогою опосередкованого тіоредоксином відновлення, якщо рівні T6P збільшені за допомогою експресії трансгена. Фермент NAPDH-тіоредоксинредуктаза C (NTRC) використовує NADPH для окисновідновної активації AGPази. Мутанти за NTRC не реагують на живлення трегалозою, що дозволяє припустити, що T6P діє на фермент NTRC або попередній елемент у шляху окисновідновної активації AGPази. Розглядаючи разом, T6P функція у рослинах впливає на енергетичну передачу сигналу, а також окисно-відновну передачу сигналу за допомогою NADPH; T6P необхідний, оскільки він контролює доступність NADPH – субстрату для відновного біосинтезу. Проте механізми, за допомогою яких T6P проявляє свій вплив на залежну від NADPH окисно-відновну передачу сигналу, у рослин невідомі. Природа залучення SnRK1 в цьому випадку невідома (Schleupmann et al., (2012) J Exp Bot:1-12). SnRK1 необхідна для активації AGPази (Kolbe et al., (2005) PNAS 102:11118-11123) і, очевидно, є активною як вище, так і нижче T6P в окисно-відновній активації AGPази в гетеротрофних тканинах. [0134] T6P є необхідним метаболітом, тому що він контролює доступність субстрату для росту. Ріст сходів Arabidopsis інгібується, якщо їх вирощують на середовищах з трегалозою, і ефект послаблюється при живленні сахарозою та трегалозою. Було показано, що зупинка росту на трегалозі викликається накопиченням T6P; інгібування трегалози корелює з 10-кратним збільшенням T6P (Schluepmann et al.,(2004) Plant Physiol 135:879-890). Інгібування росту сходів Arabidopsis пов'язане зі значним накопиченням крохмалю в сім'ядолях. Навпаки, у кореневих кінчиках не показане типове накопичення крохмалю. Живлення сходів трегалозою порушує розподіл вуглецю, при цьому весь вуглець залишається в його джерелі - сім'ядолях, та спостерігається помітне вуглецеве голодування у верхівках. Проте, активація за допомогою T6P AGPази та включення вуглецю в крохмаль не є причиною інгібування росту. Навпаки,інгібування активності SnRK1за допомогою T6P, як було показано, викликає інгібування росту на трегалозі, оскільки сходи, які надекспресували каталітичну субодиницю (KIN10) SnRK1, росли на трегалозі (Delatte et al., (2011) Plant Physiol 157:160-174). Таким чином, очевидно, сходи перестають рости на трегалозі, оскільки накопичення T6P інгібує активність SnRK1. [0135] Це може здаватися парадоксальним, що як украй мала кількість T6P, як у tps1 мутанта, так і вкрай велика кількість T6P за відсутності вуглецю, що надається екзогенно, зупиняє ріст рослини. Механізм, за допомогою якого T6P/SnRK1 здійснює контроль за метаболізмом у рослині, далекий від розуміння. Проте, не обмежуючись теорією, у гетеротрофних тканинах кількість T6P збільшується, коли доступна сахароза. Збільшення кількості T6P інгібує активність SnRK1 таким чином, що клітини можуть переключатися на анаболічні процеси, необхідні для росту. Коли знижуються рівні сахарози або доступність вуглецю, то знижується рівень T6P, знімаючи контроль з активності SnRK1; при цьому анаболічні процеси інгібуються. Також відомо, що активність SnRK1 активується під час реакцій на вуглецевий і енергетичний стрес (фігура 6). SnRK1 сприймає сигнал про низький рівень енергії або вуглецю та опосередковує процеси, необхідні для того, щоб зробити вуглець доступним у клітинах-реципієнтах для росту. Активність SnRK1 необхідна, доки вуглець не стане знову доступним у гетеротрофних тканинах. Потім синтезується сахароза, а рівень T6P збільшується достатньою мірою, щоб інгібувати активність SnRK1. [0136] Даний шлях передачі сигналу за допомогою T6P/SnRK1 жодним чином не є універсальним. В Arabidopsis зв'язування T6P з SnRK1 потребує усе ще невловимої розчинної білкової молекули, фактора 1 (Zhang et al., (2009) Plant Physiol 149:1860-1871), яку не можна виявити в зрілому листку. Крім того, субодиниці комплексу SnRK1 гетерогенно розподілені в рослинних тканинах (Bitrián et al., (2011) Plant J 65:829-842). Гетерогенність, таким чином, очікується при інгібуванні SnRK1 за допомогою T6P у гетеротрофних тканинах і фотоавтотрофних тканинах (Bitrian et al., (2011) Plant J 65:829-842). Також T6P є зарядженим метаболітом і, тому, навряд чи вільно експортується в інші клітини (Schleupmann et al., (2012) J Exp Bot:1-12). [0137] Генетичні та біохімічні дані показують, що трегалозний шлях є необхідним для розвитку рослин і контролює клітинний морфогенез. Втрата ферментативної активності TPS1 в Arabidopsis є летальною для зародків (Eastmond et al., (2002) Plant J 29:225-235; Paul et al., (2008) Annu Rev Plant Biol 59:417-441). Був встановлений зв'язок експресії AtTPS1 зі специфічними характерними для стадії розвитку фенотипами, такими як уповільнений розвиток зародка, змінений ріст кореня і пагона і змінений перехід до цвітіння. Дані показують, що 23 UA 115768 C2 5 10 15 20 25 30 35 40 45 50 55 60 відсутність T6P послабляє здатність зародка до переходу у фазу розвитку з поділом клітин (Paul et al., (2008) Annu Rev Plant Biol 59:417-441). [0138] У маїсу RAMOSA3 (RA3), який є важливим для розгалуження суцвіття, кодує білок T6PP, що складається з одного домену (Satoh-Nagasawa et al. (2006) Nature 441:227-230). Уважається, що експресія T6PP в ділянках, що оточують пазушні меристеми, може сприяти передачі рухливого сигналу, що має короткий діапазон, для регуляції ідентичності та детермінованості меристеми. Втрата RA3 активності викликає важкі аномалії качанів, які включають довгі пазушні пагони в основі качана на ранньому розвитку (Eveland & Jackson (2012) J Exp Bot:1-11). [0139] Chary et al. описує ідентифікацію мутанта з фенотипом-1 форми клітини (csp-1) Arabidopsis thaliana. CSP-1 демонструє велику кількість дефектів у процесах розвитку рослини, наприклад, довжині стебла рослини, морфології листка, розвитку кореня, зменшеному розгалуженні пагонів і вповільненому цвітінні (Chary et al. (2008) Plant Physiol146: 97-107). Мутація була виявлена в гені AtTPS6. Це забезпечує істотний доказ, вказуючий на те, що ген TPS є важливим модифікатором клітинної морфології та розвитку всієї рослини. Незрозуміло, який механізм дії лежить в основі цих серйозних впливів на розвиток. Можливо, що вони, щонайменше частково, опосередковані шляхом передачі сигналу за допомогою T6P/SnRK1. [0140] T6P у більшості зразків маїсу присутній у концентрації
ДивитисяДодаткова інформація
Назва патенту англійськоюMethods of increasing yield and stress tolerance in a plant
Автори англійськоюBasu, Shib, Cohn, Jonathan, Nuccio, Michael
Автори російськоюБасу Шиб, Кон Джонатан, Нуччио Майкл
МПК / Мітки
МПК: C12N 15/82, C12N 9/16, A01H 1/06
Мітки: спосіб, стресу, підвищення, абіотичного, рослини, стійкості
Код посилання
<a href="https://ua.patents.su/101-115768-sposib-pidvishhennya-stijjkosti-roslini-do-abiotichnogo-stresu.html" target="_blank" rel="follow" title="База патентів України">Спосіб підвищення стійкості рослини до абіотичного стресу</a>
Спосіб підвищення врожайності і/або збільшення сили рослин, і/або підвищення стійкості або опірності рослини до абіотичних факторів стресу і застосування аміду за зазначеним способом

Номер патенту: 105001
Опубліковано: 10.04.2014
Автори: Праде Алешандер Гуттенкунст, Таварес-Родрігес Марко-Антоніо, Бегліоміні Едсон, Фьосте Дірк, Вільгельм Рональд, Мерк Міхаель, Екко Марлон
МПК: A01N 47/24, A01N 43/56, A01P 21/00
Мітки: сили, абіотичних, способом, стійкості, зазначеним, застосування, рослин, врожайності, рослини, аміду, стресу, опірності, спосіб, підвищення, факторів, збільшення
Формула / Реферат:
1. Спосіб підвищення врожайності і/або збільшення сили, і/або підвищення стійкості або опірності сільськогосподарських рослин до абіотичних факторів стресу, який включає обробку паростків рослин амідом, який має формулу І (сполука І), (I)у якійR4 означає дифторметил або трифторметил;R5 означає водень;М означає фенільне кільце, де...
Спосіб діагностики стійкості винограду до абіотичного стресу методом індукції флуоресценції хлорофілу
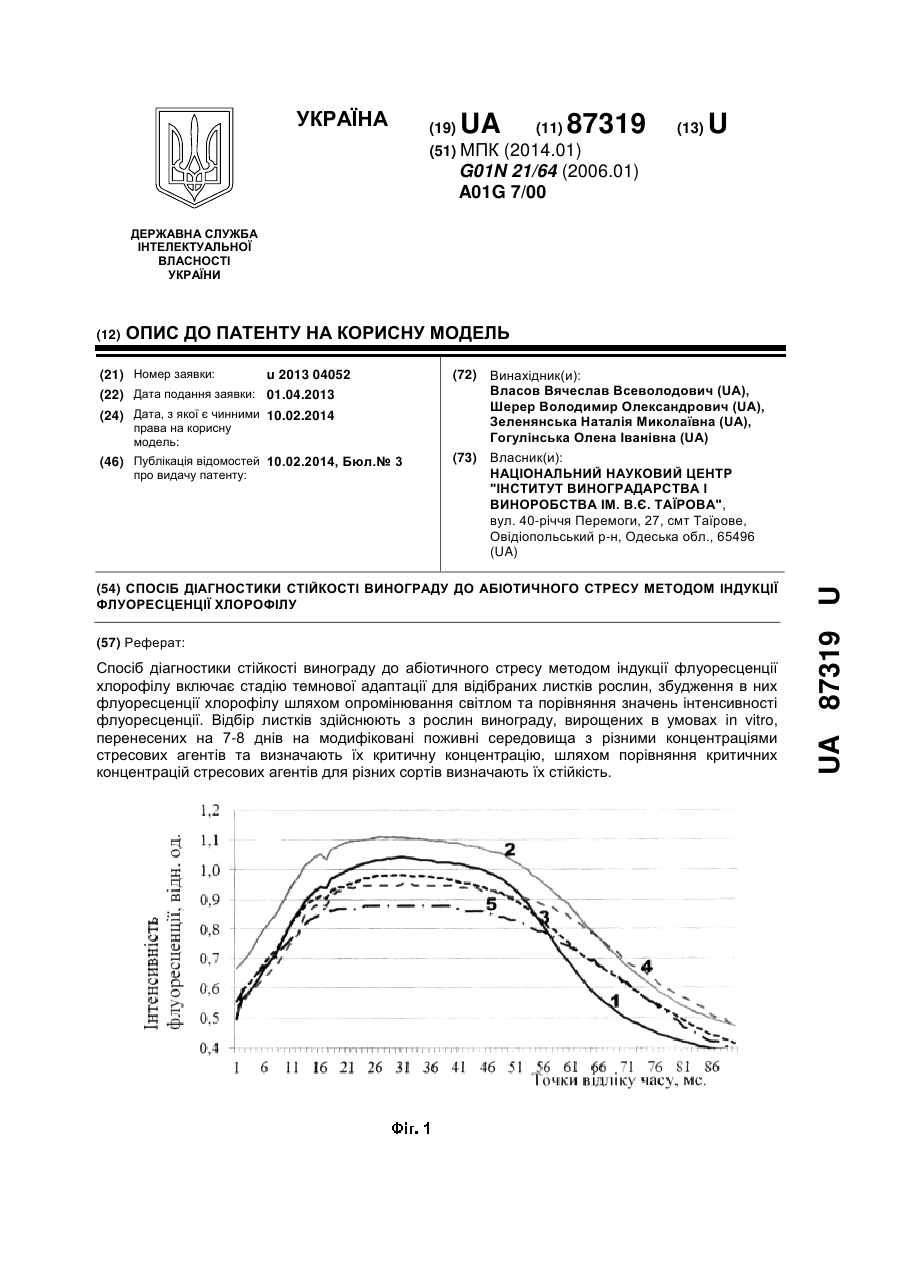
Номер патенту: 87319
Опубліковано: 10.02.2014
Автори: Власов В'ячеслав Всеволодович, Шерер Володимир Олександрович, Гогулінська Олена Іванівна, Зеленянська Наталія Миколаївна
МПК: A01G 7/00, G01N 21/64
Мітки: абіотичного, флуоресценції, діагностики, методом, стресу, стійкості, хлорофілу, спосіб, винограду, індукції
Формула / Реферат:
Спосіб діагностики стійкості винограду до абіотичного стресу методом індукції флуоресценції хлорофілу, що включає стадію темнової адаптації для відібраних листків рослин, збудження в них флуоресценції хлорофілу шляхом опромінювання світлом та порівняння значень інтенсивності флуоресценції, який відрізняється тим, що відбір листків здійснюють з рослин винограду, вирощених в умовах in vitro, перенесених на 7-8 днів на модифіковані поживні...
Застосування азолів для підвищення стійкості рослин або частин рослин проти абіотичного стресу

Номер патенту: 102856
Опубліковано: 27.08.2013
Автори: Кене Хайнц, Діттген Ян, Лєр Штефан, Тібес Йорг, Буш Марко, Хойзер-Хан Ізольде
МПК: A01N 37/42, A01P 21/00, A01N 49/00, A01N 43/653, C05G 3/02
Мітки: рослин, абіотичного, підвищення, частин, стійкості, азолів, застосування, стресу
Формула / Реферат:
1. Застосування принаймні однієї сполуки, вибраної з групи, що включає тебуконазол, метконазол та протіоконазол у комбінації з абсцизовою кислотою для підвищення стійкості рослин до абіотичних стресорів.2. Застосування за п. 1, яке відрізняється тим, що принаймні одну сполуку, вибрану з групи, що включає тебуконазол, метконазол та протіоконазол, застосовують із дотриманням норми витрати від 0,01 до 3 кг/га. 3. Застосування за...
Застосування інгібіторів сукцинатдегідрогенази для підвищення стійкості рослин або частин рослин проти абіотичного стресу

Номер патенту: 108613
Опубліковано: 25.05.2015
Автори: Хойзер-Хан Ізольде, Еббінгхаус Дірк, Діттген Ян
МПК: A01N 43/56, A01P 15/00
Мітки: інгібіторів, рослин, абіотичного, стійкості, сукцинатдегідрогенази, частин, стресу, застосування, підвищення
Формула / Реферат:
1. Застосування біксафену для підвищення стійкості рослин проти абіотичних стресорів, причому біксафен наносять на рослини ще до початку дії стресорів, причому біксафен застосовують із дотриманням норми витрати від 0,01 до 3 кг/га у перерахунку на чистий біксафен, абіотичним стресором є посуха, а рослиною є ячмінь.2. Застосування за п. 1, яке відрізняється тим, що оброблена рослина є трансгенною.3. Застосування за п. 1 або 2,...
Спосіб поліпшення стійкості рослин до абіотичного стресу з використанням карбоксамідних або тіокарбоксамідних похідних

Номер патенту: 114647
Опубліковано: 10.07.2017
Автори: Діттген Ян, Крісто П'єр, Шмутцлер Дірк
МПК: A01N 43/56
Мітки: абіотичного, використанням, тіокарбоксамідних, стресу, похідних, карбоксамідних, поліпшення, стійкості, рослин, спосіб
Формула / Реферат:
1. Спосіб обробляння рослин для підвищення стійкості до посухи, що включає нанесення вказаним рослинам, насінню, з якого вони проростають, або до ділянки, де вони ростуть, нефітотоксичної, ефективної для поліпшення стійкості рослин до посухи кількості сполуки, яку вибирають із групи, яка складається...
Попередній патент: Способи отримання ізохінолінонів і тверді форми ізохінолінонів
Наступний патент: Пристрій для відведення текучого середовища пристроєм центрифугування
Випадковий патент: Лікувально-профілактичний засіб "біопшениця"