Сорго, резистентний до ацетолактатсинтазного гербіциду













Формула / Реферат
1. Гібрид сорго, у якому зародкова плазма зазначеного гібриду сорго дає можливість досягнути резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами на рівнях зазначених одного або кількох гербіцидів, що звичайно б інгібували ріст гібриду сорго, де зародкова плазма зазначеного гібриду сорго дає можливість досягнути резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами, які включають:
(і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або кілька нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного аденіном в позиції 1641, та гуаніну, заміщеного тиміном в позиції 1684; або (іі) мутації в ацетолактатсинтазному гені, як отримано в АТСС NO: PTA-7999.
2. Гібрид сорго за п. 1, де зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, яка складається із сульфонілсечовин, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів.
3. Гібрид сорго за п. 1, де зазначена резистентність до інгібування одним або кількома ацетолактатсинтазними гербіцидами інтродукована у зазначену зародкову плазму гібриду сорго шляхом інтрогресії.
4. Гібрид сорго за п. 1, де насіння від гібриду сорго покривається шаром ацетолактатсинтазною гербіциду.
5. Спосіб контролю бур'янів поблизу гібриду сорго за любим з пп. 1-4, який включає:
a) підготування одного або кількох ацетолактатсинтазних гербіцидів, як визначено в будь-якому з попередніх пунктів,
b) доставлення зазначених одного або кількох ацетолактатсинтазних гербіцидів до лану, який містить гібрид сорго, та
c) контроль бур'янів поблизу зазначеного гібриду сорго, при якому наявність зазначених одного або кількох гербіцидів негативно впливає на ріст бур'янів і не впливає негативно на ріст зазначеного гібриду сорго.
6. Спосіб за п. 5, де зазначений гібрид сорго утворений шляхом уведення гетерологічного гена, котрий містить одну або кілька мутацій для надання резистентності до одного або кількох ацетолактатних гербіцидів, у зазначений гібрид сорго.
7. Спосіб для продукування рослинної лінії гібриду сорго, резистентної до одного або кількох ацетолактатсинтазних гербіцидів, який включає:
a) ідентифікацію зародкової плазми, що надає зазначеної гербіцидної резистентності, де зазначену резистентну до гербіциду зародкову плазму отримують із резистентного до гербіциду гібриду сорго за будь-яким з пп. 1-4, та
b) уведення зазначеної зародкової плазми в елітну рослинну лінію сорго здійснюють шляхом уведення гетерологічного гена.
8. Спосіб за п. 7, де зазначене інтродукування зазначеної зародкової плазми у зазначену елітну рослинну лінію сорго здійснюють шляхом інтрогресії.
9. Спосіб за п. 7, де зазначена гербіцидрезистентна зародкова плазма включає резистентність до одного або кількох ацетолактатсинтазних гербіцидів та резистентність до однієї або кількох сполук із однієї або кількох груп гербіцидів, котрі не є інгібіторами ацетолактатсинтази.
10. Спосіб ідентифікації рослинних ліній сорго, резистентних до ацетолактатсинтазних гербіцидів, який включає:
a) отримання зразка нуклеїнової кислоти від рослини сорго, де зазначений зразок нуклеїнової кислоти включає:
(і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або більше нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного з аденіном в позиції 1641, та гуаніну, заміщеного з тиміном в позиції 1684; або (іі) мутації в ацетолактатсинтазному гені, як отримано в АТСС NO: PTA-7999,
b) підготування ампліфікаційних праймерів для ампліфікації ділянки рослини сорго, яка відповідає ацетолактатсинтазному гену, присутньому у зазначеному зразку нуклеїнової кислоти,
c) доставлення зазначених ампліфікаційних праймерів до зазначеного зразка нуклеїнової кислоти, таким чином, що має місце ампліфікація зазначеної ділянки зазначеного ацетолактатсинтазного гена, та
d) ідентифікацію рослин сорго, резистентних до ацетолактатсинтазних гербіцидів, на основі наявності однієї або кількох мутацій, що надають резистентності до ацетолактатсинтазних гербіцидів, присутньої у зазначеному ампліфікованому зразку нуклеїнової кислоти.
11. Насіння гібриду сорго, яке включає зародкову плазму гібриду сорго, яка дає можливість досягнути резистентність до інгібування одним або кількома ацетолактатсинтазними гербіцидами, який включає:
(і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або кілька нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного з аденіном в позиції 1641, та гуаніну, заміщеного з тиміном в позиції 1684; або (іі) мутації в ацетолактатсинтазному гені, як отримано в АТСС NO: PTA-7999.
12. Насіння за п. 11, де зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, яка складається із сульфонілсечовин, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів.
13. Насіння за п. 11, де зазначена резистентність до інгібування одним або кількома ацетолактатсинтазними гербіцидами інтродукована у зазначену зародкову плазму гібриду сорго шляхом інтрогресії.
14. Насіння за п. 11, де насіння від гібриду сорго покривається шаром ацетолактатсинтазного гербіциду.
15. Гібрид сорго за п. 2, де ацетолактатсинтазний гербіцид є сульфонілсечовинним гербіцидом вибраним із групи, що складається із нікосульфурону, римсульфурону та метсульфурон-метилу.
16. Спосіб за п. 5 або 6, де зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, котра складається із сульфонілсечовинів, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів.
17. Спосіб за п. 16, де ацетолактатсинтазний гербіцид є сульфонілсечовинним гербіцидом, вибраним із групи, що складається із нікосульфурону, римсульфурону та метсульфурон-метилу.
Текст
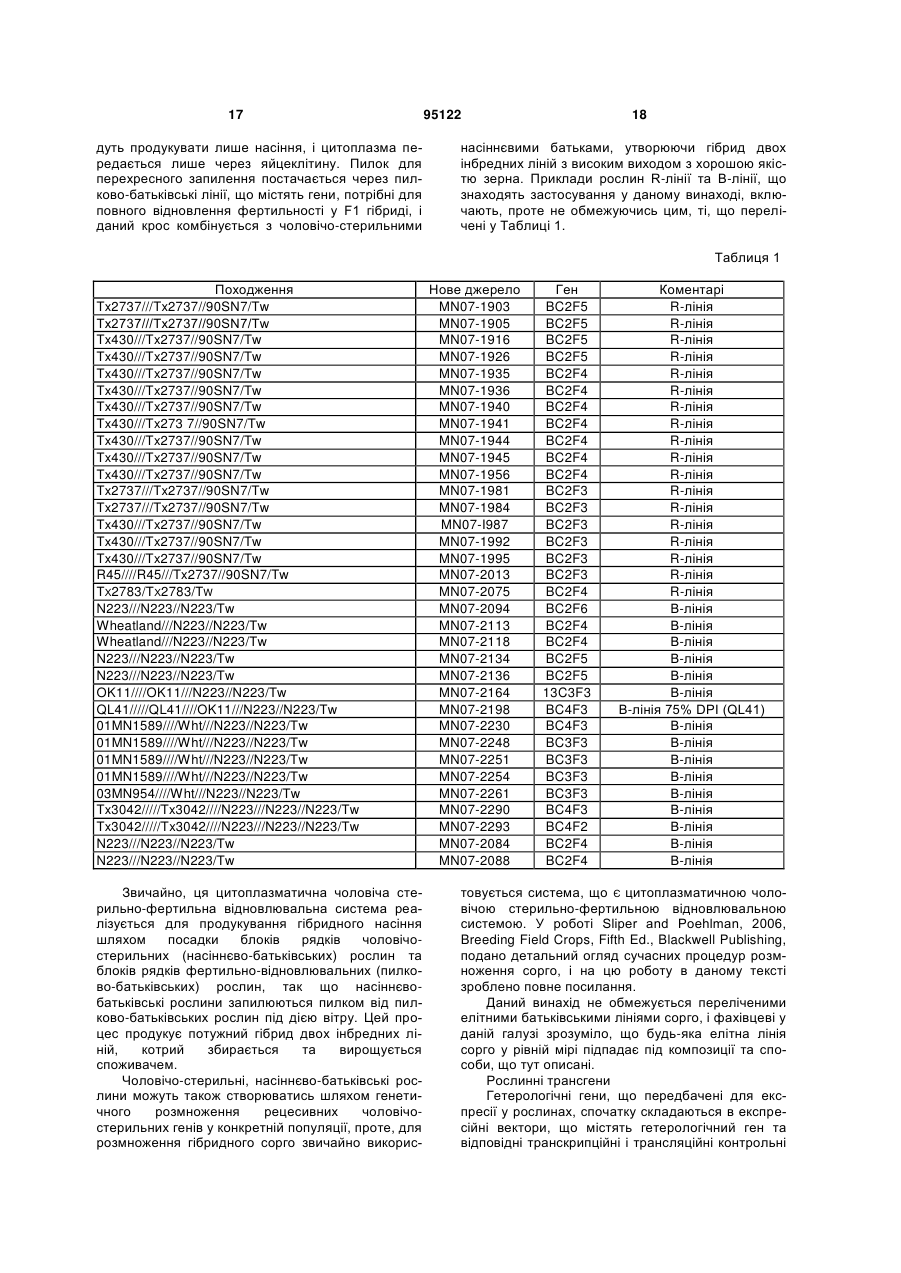
1. Гібрид сорго, у якому зародкова плазма зазначеного гібриду сорго дає можливість досягнути резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами на рівнях зазначених одного або кількох гербіцидів, що звичайно б інгібували ріст гібриду сорго, де зародкова плазма зазначеного гібриду сорго дає можливість досягнути резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами, які включають: (і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або кілька нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного аденіном в позиції 1641, та гуаніну, заміщеного тиміном в позиції 1684; або (іі) 2 (19) 1 3 95122 4 ресії. 9. Спосіб за п. 7, де зазначена гербіцидрезистентна зародкова плазма включає резистентність до одного або кількох ацетолактатсинтазних гербіцидів та резистентність до однієї або кількох сполук із однієї або кількох груп гербіцидів, котрі не є інгібіторами ацетолактатсинтази. 10. Спосіб ідентифікації рослинних ліній сорго, резистентних до ацетолактатсинтазних гербіцидів, який включає: a) отримання зразка нуклеїнової кислоти від рослини сорго, де зазначений зразок нуклеїнової кислоти включає: (і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або більше нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного з аденіном в позиції 1641, та гуаніну, заміщеного з тиміном в позиції 1684; або (іі) мутації в ацетолактатсинтазному гені, як отримано в АТСС NO: PTA-7999, b) підготування ампліфікаційних праймерів для ампліфікації ділянки рослини сорго, яка відповідає ацетолактатсинтазному гену, присутньому у зазначеному зразку нуклеїнової кислоти, c) доставлення зазначених ампліфікаційних праймерів до зазначеного зразка нуклеїнової кислоти, таким чином, що має місце ампліфікація зазначеної ділянки зазначеного ацетолактатсинтазного гена, та d) ідентифікацію рослин сорго, резистентних до ацетолактатсинтазних гербіцидів, на основі наявності однієї або кількох мутацій, що надають резистентності до ацетолактатсинтазних гербіцидів, присутньої у зазначеному ампліфікованому зразку нуклеїнової кислоти. 11. Насіння гібриду сорго, яке включає зародкову плазму гібриду сорго, яка дає можливість досягну ти резистентність до інгібування одним або кількома ацетолактатсинтазними гербіцидами, який включає: (і) послідовність, яка включає SEQ ID NO: 1, яка додатково містить один або кілька нуклеотидних субстетуцій, вибраних з групи, яка складається із гуаніну, заміщеного з аденіном в позиції 1641, та гуаніну, заміщеного з тиміном в позиції 1684; або (іі) мутації в ацетолактатсинтазному гені, як отримано в АТСС NO: PTA-7999. 12. Насіння за п. 11, де зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, яка складається із сульфонілсечовин, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів. 13. Насіння за п. 11, де зазначена резистентність до інгібування одним або кількома ацетолактатсинтазними гербіцидами інтродукована у зазначену зародкову плазму гібриду сорго шляхом інтрогресії. 14. Насіння за п. 11, де насіння від гібриду сорго покривається шаром ацетолактатсинтазного гербіциду. 15. Гібрид сорго за п. 2, де ацетолактатсинтазний гербіцид є сульфонілсечовинним гербіцидом вибраним із групи, що складається із нікосульфурону, римсульфурону та метсульфурон-метилу. 16. Спосіб за п. 5 або 6, де зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, котра складається із сульфонілсечовинів, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів. 17. Спосіб за п. 16, де ацетолактатсинтазний гербіцид є сульфонілсечовинним гербіцидом, вибраним із групи, що складається із нікосульфурону, римсульфурону та метсульфурон-метилу. Дана заявка претендує на пріоритет попередньої патентної заявки CШA за номером 60/873529 від 7 грудня 2006 року, на яку у даному тексті зроблено повне посилання. Даний винахід запроваджує композиції та способи для продукування культурних сортів сорго, що є резистентними до гербіцидів. Зокрема, даний винахід запроваджує рослини сорго, тканини та насіння рослин, що містять змінені ацетолактатсинтазні (ALS) гени та протеїни, резистентні до інгібування гербіцидами, котрі звичайно інгібують активність ALS протеїну. Сорго являє собою другий за важливістю харчовий хлібний злак, що вирощується у Сполучених Штатах. Виробництво є економічно критичним для ферм, котрі працюють у маргінальних щодо кількості атмосферних опадів областях завдяки здатності сорго переносити посуху та спеку. Скотарство і біоенергетична промисловість використовують сорго як енергетичну основу, що робить його універсальною культурою. У світі сорго є п'ятим за значимістю хлібним злаком. Оскільки сорго стійке як до посухи так і спеки, воно являє собою найбільш широко та легко вирощуваний харчовий злак у напівпустельних регіонах еуб-сахельної Африки та у посушливому центральному півострівному регіоні Індії. Як таке, сорго використовується як продукт споживання людиною у більшості найбільш посушливих регіонів світу, що робить його критично важливою харчовою культурою у цих областях. Розробка резистентності до гербіцидів пропонує значні виробничі та економічні переваги, оскільки застосування гербіцидів для контролю бур'янів у даній культурі стало майже універсальною практикою. Проте, застосування таких гербіцидів може також спричинити загибель або послаблений ріст потрібної культурної рослини, роблячи час та спосіб нанесення гербіциду критичним або у деяких випадках нездійсненним. Особливий інтерес для фермерів являє використання гербіцидів з більшою активністю, широким спектром ефективності щодо бур'янів та швидкою деградацією у грунті. Рослини, рослинні тканини та насіння, резистентні до цих сполук, запроваджують привабливий розв'язок шляхом застосування гербіцидів для контролю росту бур'янів з низьким ризиком пошкодження даної рослини. Одним таким класом гербіцидів широкого спектру с такі, шо інгібують активність ацетолакта 5 тсинтазного (ALS) ензиму у рослині. Ацетолактатсинтаза погрібна для продукування незамінних амінокислот, таких як валін, лейцин та ізолейцин, у рослинах (цей біохімічний шлях у людей або інших тварин відсутній). Сорго чутливе до багатьох гербіцидів, що інгібують ALS, котрі вражають однодольні види, і це робить майже неможливим використання таких гербіцидів для контролю трав'янистих бур'янів, оскільки вони будуть також інгібувати ріст даної культурної рослини. Ацетолактатсинтазні гербіциди контролюють широкий спектр трав'янистих та листяних бур'янів при вельми низьких нормах застосування. У теперішній час мається більше 56 різних ALS гербіцидів, виготовлених із різних композицій сульфонілсечовин (SU), імідазолінонів (ІМІ), триазолопіримідинів (TP) та піримідинілтіобензоатів (РТВ). У деяких культурних рослинах, наприклад, тютюні, кукурудзі та соєвих бобах (патент США за номером 5767361) були знайдені мутації, котрі надають ALS гербіцидну резистентність, проте, для сорго таких знахідок у теперішній час не виявлено (щодо огляду дивись роботу Tan et al., 2005, Pest. Manag. Sci. 61:246-257). Через важливість сорго на світовій сцені є потреба у сортах сорго, котрі резистентні до інгібіторних ефектів ALS гербіцидів, що надає можливість отримувати більш високі виходи культур при використанні цих гербіцидів для контролю трав'янистих бур'янів. Даний винахід запроваджує композиції та способи для продукування культурних сортів сорго, що є резистентними до гербіцидів. Зокрема, даний винахід запроваджує рослини сорго, тканини та насіння рослин, що містять змінені ацетолактатсинтазні (ALS) гени та протеїни, резистентні до інгібування гербіцидами, котрі звичайно інгібують активність ALS протеїну. Культурне сорго [Sorghum bicolor (L.) Moench] чутливе до багатьох гербіцидів, що інгібують ALS, котрі вражають однодольні або трав'янисті види. Проте, як тут описано, був знайдений генотип сорго, що виявляє резистентність до ALS гербіцидів. Генетичний аналіз ідентифікував генетичні відмінності у зародковій плазмі сорго, котрі дають ALS гербіцидний резистентний фенотип. В одному варіанті даний винахід запроваджує одну або кілька рослин сорго, зародкова плазма яких включає мутацію, котра надає даній рослині резистентності до ALS гербіцидів. Крім того, у додаткових варіантах даний винахід стосується нащадків (наприклад, F1, F2, F3 і т.д.) кросу зазначеної рослини, де зародкова плазма зазначеного нащадка має ту саму мутацію як і батьківська рослина. Таким чином, варіанти даного винаходу запроваджують гібриди сорго, зародкова плазма яких містить мутацію, таку, що фенотип даних рослин є ALS гербіцид резистентним. В одному варіанті даний винахід запроваджує гібрид сорго, де зародкова плазма зазначеного гібриду сорго надає резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами при рівнях зазначених одного або кількох гербіцидів, котрі звичайно інгібують ріст гібриду сорго. У деяких варіантах зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються 951226 із групи, що складається із сульфонілсечовин, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів. У деяких варіантах зародкова плазма зазначеного гібриду сорго, що надає резистентності до інгібування одним або кількома ацетолактатсинтазними гербіцидами, включає мутації в ацетолактатному гені, як отримано у ATCC No. РТА7999. У деяких варіантах насіння від зазначеного гібриду сорго покривається шаром ацетолактатсинтазного гербіциду. В одному варіанті даний винахід запроваджує спосіб контролю бур'янів поблизу гібриду сорго як тут описано, що включає запровадження одного або кількох ацетолактатсинтазних гербіцидів, застосування зазначених одного або кількох ацетолактатсинтазних гербіцидів до лану, який містить гібрид сорго, як тут описано, та контроль бур'янів поблизу зазначеного гібриду сорго, такий, що застосування зазначених одного або кількох гербіцидів негативно впливає на ріст бур'янів і не впливає негативно на ріст зазначеного гібриду сорго. У деяких варіантах зазначені один або кілька ацетолактатсинтазних гербіцидів вибираються із групи, що складається із сульфонілсечовин, імідазолінонів, триазолопіримідів та піримідинілтіобензоатів. У деяких варіантах зазначений гібрид сорго включає одну або кілька мутацій в ацетолактатсинтазному гені, як отримано у ATCC No. РТА-7999. В одному варіанті даний винахід запроваджує гібрид сорго, де зазначений гібрид сорго включає зародкову плазму, яка містить одну або кілька мутацій в ацетолактатсинтазному гені, таку, що зазначеному гібриду надається резистентність до одного або кількох ацетолактатсинтазних гербіцидів. У деяких варіантах зазначений гібрид сорго створюється шляхом інтрогресії зародкової плазми сорго, що містить зазначені одну або кілька мутацій, для надання резистентності до одного або кількох ацетолактатсинтазних гербіцидів. У деяких варіантах зазначений гібрид сорго створюється шляхом уведення гетерологічного гена, що містить одну або кілька мутацій, для надання резистентності до одного або кількох ацетолактатсинтазних гербіцидів. В одному варіанті даний винахід запроваджує спосіб для продукування рослинної лінії гібриду сорго, резистентної до одного або кількох ацетолактатсинтазних гербіцидів, який включає ідентифікацію зародкової плазми, що надає зазначеної гербіцидної резистентності, де зазначена резистентна до гербіциду зародкова плазма отримується із резистентної до гербіциду рослини сорго, та введення зазначеної зародкової плазми в елітну рослинну лінію сорго. У деяких варіантах зазначене введення зазначеної зародкової плазми у зазначену елітну рослинну лінію сорго здійснюється шляхом інтрогресії. У деяких варіантах зазначене введення зазначеної зародкової плазми у зазначену елітну рослинну лінію сорго здійснюється шляхом уведення гетерологічного гена. В одному варіанті даний винахід запроваджує гібрид сорго, де зародкова плазма зазначеного гібриду включає запроваджену резистентність до одного або кількох ацетолактатсинтазних гербіцидів та резистентність до однієї або кількох сполук 7 із однієї або кількох груп гербіцидів, котрі не є інгібіторами ацетолактатсинтази. В одному варіанті даний винахід запроваджує спосіб ідентифікації рослинних ліній сорго, резистентних до ацетолактатсинтазних гербіцидів, який включає доставляння зразка нуклеїнової кислоти до рослини сорго, запровадження ампліфікаційних праймерів для ампліфікації ділянки рослини сорго, яка відповідає ацетолактатсинтазному гену, присутньому у зазначеному зразку нуклеїнової кислоти, застосування зазначених ампліфікаційних праймерів до зазначеного зразка нуклеїнової кислоти, таке, що має місце ампліфікація зазначеної ділянки зазначеного ацетолактатсинтазного гена, та ідентифікацію рослин сорго, резистентних до ацетолактатсинтазних гербіцидів, основану на наявності однієї або кількох мутацій, то надають ацетолактатсинтазну гербіцидну резистентність, присутню у зазначеному ампліфікованому зразку нуклеїнової кислоти. В одному варіанті даний винахід запроваджує насіння сорго, де зазначена зародкова плазма зазначеного насіння містить мутантний ацетолактатсинтазний ген, так що зазначена мутація надає резистентності до інгібування ацетолактасинтазними гербіцидами. У деяких варіантах зародкова плазма зазначеного насіння сорго містить мутантний ацетолактатсинтазний ген, як отримано у ATCC No. РТА-7999. У деяких варіантах мутантний ацетолактатсинтазний ген являє собою функціональний фрагмент гена, як отримано у ATCC No. РТА-7999, такий, що кодує протеїновий фрагмент, що достатньо для надання рослині сорго резистентності до інгібування ацетолактатсинтазними гербіцидами. У деяких варіантах даний винахід запроваджує рослини сорго, що виростають із зазначеного насіння і, крім того, частини рослин, що включають зазначені рослини сорго, вирощені із зазначеного насіння. В одному варіанті даний винахід також запроваджує рослини гібриду сорго, що мають усі фізіологічні та морфологічні характеристики зазначеної рослини сорго, вирощеної із зазначеного насіння сорго. У додаткових варіантах даний винахід запроваджує тканинні культури та регенеровані тканинні культури, котрі виникають із зазначеного насіння сорго або зазначеної частини рослини сорго, що містять мутацію у зазначеному ацетолактатсинтазному гені, як отримано у ATCC No. РТА7999. У деяких варіантах даний винахід запроваджує гібрид сорго, що містить ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419 згідно з ATCC No. РТА-7999. У деяких варіантах ALS гербіцид резистентний ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, 95122 8 як отримано у зародковій плазмі KSU 06MN8419, містить одну або кілька амінокислотних субституцій Val 531 Ilе та Trp545Leu, наприклад, як отримано у SEQ ID NO:1. В одному варіанті даний винахід запроваджує спосіб продукування насіння сорго, що включає схрещування рослини, котра містить мутантний ацетолактатсинтазний ген, як отримано у ATCC No. РТА-7999, самої з собою або з другою рослиною сорго та збирання зазначеного насіння від зазначеного кросу. У деяких варіантах способи продукування зазначеного насіння сорго включають висівання батьківської насіннєвої лінії сорго, де зазначена батьківська насіннєва лінія містить зародкову плазму, котра надає резистентності до ацетолактатсинтазних гербіцидів, з батьківською запильниковою лінією сорго, де зародкова плазма зазначеної запильникової насіннєвої лінії містить зародкову плазму, котра надає резистентності до ацетолактатсинтазних гербіцидів, вирощування разом зазначеного батьківського насіння та запильникових рослин сорго, надаючи можливість зазначеним батьківсько-насіннєвим рослинам обпилюватись зазначеними батьківськими запильниковими рослинами, та збирання насіння, що є результатом зазначеного обпилювання. Опис фігур Фігура зображує подвійну мутацію Val 531 Ilе та Trp545Leu у сортовому ALS гені, що асоціюється, як було знайдено, з ALS гербіцидною резистентністю. Визначення Як застосовується у даному тексті, термін "резистентний" та "толерантний" використовується щодо рослин, наприклад, рослин сорго, котрі здатні витримувати умови (наприклад, гербіциди, такі як ALS гербіциди), шкідливі для інших штамів тих самих видів. Як застосовується у даному тексті, термін "сорт культурної рослини" (культивар) є синонімом "різновиду" і використовується щодо культурних рослин, які являють собою групу схожих рослин, котрі за структурними властивостями та ефективністю можуть бути ідентифіковані із інших культиварів у межах того самого виду. Як застосовується у даному тексті, термін "гібрид" стосується нащадка або нащадків генетично несхожих рослинних батьків або лінії, утворених в результаті контрольованого перехресного обпилення, на відміну від негібридного насіння, утвореного в результаті природного обпилення. Як застосовується у даному тексті, термін "нащадки" стосується генерацій рослини, де походження даної генерації може бути прослідковано назад до зазначеної рослини. Як застосовується у даному тексті, термін "похідна" резистентної до гербіциду рослини включає як нащадків цієї резистентної до гербіциду рослини, так і будь-яку мутантну, рекомбінантну або розроблену методами генної інженерії похідну цієї рослини, як того самого виду, так і різних видів, де гербіцид резистентна властивість(сті) оригінальної гербіцид резистентної рослини перенесена на похідну рослину. Як застосовується у даному тексті, термін "рослинна тканина" включає диференційовану та не 9 диференційовану тканини рослин, включаючи ті, що присутні у корінні, порослі, листі, пилку, насінні та новоутвореннях, так само як і клітини в культурі (наприклад, окремі клітини, протопласти, зародки, калюс і т.д.). Рослинна тканина може бути у планті, органній культурі, тканинній культурі або клітинній культурі. Як застосовується у даному тексті, термін "частина рослини" стосується рослинної структури або рослинної тканини, наприклад, пилку, сім'язародку, тканини, бобу, насінини та клітини. У деяких варіантах даного винаходу трансгенні рослини є сільськогосподарськими рослинами. Як застосовується у даному тексті, термін "Fгенерація" та "дочірня генерація" стосується будьякої консекутивної генерації рослин, клітин, тканин або організмів після біпарентального схрещування. Генерація, що є результатом спарювання біпарентального кросу (тобто двох батьків), є першою дочірньою генерацією (що позначається як "F1" та "F1"), з посиланням на насіння та його рослину, тоді як та, що є результатом схрещування F1 індивідуумів, є другою дочірньою генерацією (що позначається як "F2" або "F2" з посиланням на насіння та його рослину. Як застосовується у даному тексті, термін "зародкова плазма" стосується будь-якого генетичного матеріалу рослин, що містить функціональні одиниці спадковості. Термін "елітна зародкова плазма", з посиланням на рослину, стосується спадкового матеріалу з доведеною генетичною перевагою. Як застосовується у даному тексті, термін "елітна рослина" стосується будь-якої рослини, що є результатом розмноження та селекції для досягнення найвищої агрономічної продуктивності. Наприклад, елітні рослини сорго, на які посилаються у даному тексті, включають, проте не обмежуючись цим, Тх430, Тх2737, Тх2783, 00MN7645, НР162, Wheatland, Тх3042, OK11, QL41 та Тх643, Bt лінії сорго. Як застосовується у даному тексті, термін "ознака" або "фенотип" стосується спостережної та/або вимірної властивості організму. Наприклад, даний винахід описує рослини, що є ALS гербіцид резистентними. Як застосовується у даному тексті, термін "ALS гербіцид", відомий також як AHAS гербіцид, стосується гербіциду, котрий інгібує активність ацетолактатсинтазного (відомого також як ацетогідроксикислотносинтазного) ензиму у рослині. Приклади ALS гербіцидів, як тут описано, включають, проте не обмежуючись цим, сульфонілсечовини (SU), імідазолінони (ІМІ), триазолопіримідини (TP) та піримідинілтіобензоати (РТВ). Як застосовується уданому тексті, терміни "маркер" та "ДНК маркер" і "молекулярний маркер" з посиланням на "селективний маркер" стосуються фізіологічної або морфологічної ознаки, що може бути визначена як маркер для свого власного вибору або для вибору інших ознак, тісно зв'язаних із цим маркером. Наприклад, таким маркером може бути ген або ознака, що асоціюється з гербіцидною толерантністю, включаючи, проте не обмежуючись цим, повторення простої послідовності 95122 10 (SSR), поліморфізм одного нуклеотиду (SNP), генетичні інсерції та/або делеції і таке подібне. Як застосовується у даному тексті, термін "інтрогрес" та "інтрогресінг" і "інтрогресія" стосується звичайних (тобто класичних) способів запилювального розмноження з уведенням стороннього генетичного матеріалу у лінію розмноження. Наприклад, даний винахід запроваджує культурні рослини сорго, що піддані інтрогресії мутантним ALS геном для забезпечення гербіцидної толерантності шляхом схрещування двох рослинних генерацій. Як застосовується у даному тексті, термін "дикого типу", з посиланням на ген, стосується функціонального гена, спільного по всій популяції рослини. Функціональним геном дикого типу є такий, що найчастіше спостерігається у популяції, і таким чином довільно визначає "нормальну" або "дикого типу" форму даного гена. Як застосовується у даному тексті, терміни "модифікований" або "мутантний", або "функціональний мутант", з посиланням на ген або генний продукт, стосуються, відповідно, гена або генного продукту, котрий виявляє модифікації у послідовності та/або функціональних властивостях (тобто змінені характеристики) при порівнянні з геном або генним продуктом дикого типу. Так, терміни "модифікований" та "мутантний", з посиланням на нуклеотидну послідовність, стосуються нуклеїновокислотної послідовності, що відрізняється одним або кількома нуклеотидами від іншої, звичайно спорідненої нуклеїновокислотної послідовності, і термін "функціональний мутант", з посиланням на поліпептид, що кодується зазначеною "модифікованою" або "мутантною" нуклеїновою кислотою, стосується протеїну або поліпептиду, котрий зберігає активність. У теперішній заявці ALS мутантний протеїн, або його "функціональний мутант" являє собою ALS ген, що зберігає свою природну активність у створенні незамінних амінокислот. Крім того, "модифікована" нуклеотидна послідовність інтерпретується як така, що знайдена у виродженому генетичному коді, як це відомо фахівцям у даній галузі. Наприклад, даний генетичний код є виродженим, оскільки є багато прикладів, в яких різні кодони визначають ту саму амінокислоту; генетичний код, в якому кожна із деяких амінокислот може кодуватись більше ніж одним кодоном. Передбачається, що даний винахід включає таку виродженість (наприклад, де гібрид сорго містить ALS ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до SEQ ID NО:1), як знайдено, наприклад, у зародковій плазмі сорго. Як застосовується у даному тексті, термін "гетерологічний", з посиланням на ген або нуклеїнову кислоту, стосується гена, що був підданий деяким маніпуляціям. Як застосовується у даному тексті, термін "ділянка" або "функціональний фрагмент", з посиланням на протеїн (як у виразах "фрагмент даного протеїну" або "протеїновий фрагмент"), стосується 11 фрагментів цього протеїну. Зазначені фрагменти можуть варіювати за розміром від чотирьох амінокислотних залишків до повної амінокислотної послідовності мінус одна амінокислота. У теперішньому винаході протеїновий фрагмент є, переважно, функціональним, так що цей протеїновий фрагмент надає резистентності даній рослині до інгібування ацетолактатсинтазними гербіцидами. В одному варіанті даний винахід запроваджує гени, що кодують змінені ALS гени та протеїни. У деяких варіантах даний винахід запроваджує використання гербіцидів, котрі не інгібують ALS ензим у сорго, що містить змінений ALS ензим, для зниження кількості однодольних та дводольних бур'янистих рослин, присутніх на культурному лані, де зазначені бур'янисті рослини чутливі до ALS гербіцидів. В одному варіанті даний винахід запроваджує ALS резистентну зародкову плазму рослини сорго, отриману, наприклад, в результаті схрещення двох батьківських рослин, дикого сорго "Tailwind або Tw" та елітної запильникової лінії Тх2783, з отриманням F1 генерації, з наступними двома зворотними кросами (BC2F3:F4) і результуючим насінням, депонованим під ATCC No: РТА-7999, що позначене як KSU 06MN8419. В одному варіанті даний винахід запроваджує зародкову плазму сорго, то надає резистентності до інгібування ALS гербіцидами і також надає резистентність до комах, проти плямистого стеблового свердлувальника Chilo partellus (Girijashankar еі аl., 2005, Plant Cell Rep. 24:513-522, на яку у даному тексті зроблено повне посилання). Наприклад, гібрид сорго, зародкова плазма якого містить синтетичний cryl Ac ген із Bacillus thuringiensis (Bt), піддається інтрогресії у лінію сорго, і зародкова плазма надає резистентності до ALS гербіцидів. Так само як і впровадження ALS гербіцидної резистентності та резистентності до комах здійснюється шляхом рослинного трансгенезу у той самий гібрид сорго. Фахівцям у даній галузі відомі різні способи, як тут описано, котрі придатні для впровадження двох або кількох атрибутів резистентності у той самий гібрид сорго. В одному варіанті ALS гербіцид резистентний ген, як отримано у сорго, що містить ALS зародкову плазму KSU 06MN8419, депоновану під ATCC No: РТА-7999, уводиться в елітні різновиди сорго шляхом розмноження та селекції рослин, запроваджуючи у такий спосіб розробку резистентних до гербіцидів різновидів культур, що будуть толерантними до застосування ALS інгібуючих гербіцидів для контролю бур'янів. У деяких варіантах ALS гербіцид резистентний ген є, на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419. У деяких варіантах ALS гербіцид резистентний ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомоло 95122 12 гічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419, містить одну або кілька амінокислотних субституцій Val 531 Ilе та Trp545Leu. Впровадження цієї ознаки гербіцидної резистентності у вищезгадані різновиди культурних рослин дозволяє використовувати ці гербіциди для контролю однодольних та дводольних бур'янів, що ростуть у присутності цих культур. У деяких варіантах уведення ALS резистентної зародкової плазми в елітні лінії здійснюється шляхом інтрогресії або класичних методів розмноження. У деяких варіантах уведення ALS резистентного гена в елітні лінії здійснюється шляхом трансгенезу гетерологічного гена. У деяких варіантах даний винахід запроваджує гібрид сорго, де принаймні один предок зазначеного гібриду сорго одержаний із ALS резистентної зародкової плазми, позначеної KSU 06MN8419 та депонованої під ATCC No: РТА-7999. У деяких варіантах теперішній винахід запроваджує гібрид сорго, де принаймні один предок зазначеного гібриду сорго містить ген ацетолактатсинтази, котрий надає резистентності до ALS гербіцидів, як отримано у зародковій плазмі під назвою KSU 06MN8419, депонованій під ATCC No: РТА-7999. У деяких варіантах ALS резистентний гербіцидний ген, як отримано у гібриді сорго, є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419. У деяких варіантах ALS гербіцид резистентний ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419, містить одну або кілька амінокислотних субституцій Val 531 Ilе та Trp545Leu. У деяких варіантах ALS гербіцид резистентна зародкова плазма піддається інтрогресії в елітну лінію сорго з використанням класичних способів розмноження. Приклади класичних способів розмноження для сорго можна знайти, наприклад, у роботі Sleper and Pochlman, 2006, Breeding Field Crops, Fifth Edition, Blackwell Publishing, на яку у даному тексті зроблено повне посилання. В одному варіанті ALS гербіцид резистентна зародкова плазма піддається інтрогресії у рослину сорго, що запроваджує їжу для споживання людиною. У деяких варіантах ALS гербіцид резистентна зародкова плазма піддається інтрогресії у рослини сорго, що запроваджують корм для домашніх тварин (наприклад, домашньої птиці, великої рогатої худоби, свиней, овець і т.д.). В одному варіанті ALS гербіцид резистентний ген уводиться у рослинний геном шляхом трансгенезу з використанням векторів та технологій, відомих у даній галузі. В одному варіанті даний винахід запроваджує способи для контролю бур'янів на ланах культур 13 них рослин сорго. У деяких варіантах контроль бур'янів включає застосування ALS гербіциду до зазначеного лану культурних рослин сорго, так що ріст бур'янів інгібується, але на ріст рослин сорго шкідливим чином це не впливає. У деяких варіантах ALS гербіцид, що застосовується, виходить із сімейства сульфонілсечовинних гербіцидів, і включає один або кілька активних інгредієнтів амідосульфурон, азімсульфурон, бензсульфуронметил, хлорімурон-етил, хлорсульфурон, ціносульфурон, циклосульфамурон, етаметсульфуронметил, етоксисульфурон, флазасульфурон, флупірсульфурон-метил-натрій, форамсульфурон, галосульфурон-метил, імазолсульфурон, йодосульфурон-метил-натрій, мезосульфурон-метил, метсульфурон-метил, нікосульфурон, оксасульфурон. примісульфурон-мстил, піраксосульфуронетил, римсульфурон, сульфометурон-метил, сульфосульфурон, тіфенсульфурон-метил, триасульфурон, трибенурон-метил, трифлоксисульфурон-натрій, трифлусульфурон-метил, триофенсульфурон та тригосульфурон. У деяких варіантах ALS гербіцид, що застосовується, виходить із сімейства імідазолінонових гербіцидів, і включає один або кілька активних інгредієнтів імазаметабенз-метил, імазамокс, імазапік, імізапір. імізакін та імазетапір. У деяких варіантах ALS гербіцид, що застосовується, виходить із сімейства піримідинілтіобензоатних гербіцидів, і включає один або кілька активних інгредієнтів - біспірибакнатрій, пірибензоксим, пірифталід, піримінобакметил та піритіобак-натрій. У деяких варіантах ALS гербіцид, що застосовується, виходить із сімейства триазолопіримідинових гербіцидів, і включає один або кілька активних інгредієнтів - хлорансулам-метил, диклузолам, флорасулам, флуметсулам, метосулам та пеноксулам. У деяких варіантах ALS гербіцид, що застосовується, включає комбінацію активних інгредієнтів із одного або кількох ALS гербіцидних сімейств як тут розкрито. Проте, теперішня заявка не обмежується використаним ALS гербіцидом, і фахівцеві у даній галузі зрозуміло, що у будь-який час можуть бути відкриті нові хімікати, що інгібують ALS ензим. В одному варіанті даний винахід запроваджує використання трансгена, що містить гетерологічний ген, такий як ген, що кодує мутантний ALS протеїн, для впровадження вибраної агрономічної ознаки ALS гербіцидної резистентності. В одному варіанті даний трансген включає мутантний ALS ген, як отримано у зародковій плазмі KSU 06MN8419, депонованій під ATCC No: РТА-7999. У деяких варіантах зазначений ALS ген є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до мутантного ALS гена, як отримано у зародковій плазмі KSU 06MN8419. У деяких варіантах зазначений мутантний ALS ген, котрий є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до мутантного ALS 95122 14 гена, як отримано у зародковій плазмі KSU 06MN8419, містить одну або кілька амінокислотних субституцій Val 531 Ilе та Trp545Leu. Striga, звичайно відома як бур'яниста рослина із родини Scrophulariaceae, належить до роду шкідливих паразитних рослин трав та однодольних зернових культур, таких як маїс, пшениця та сорго. Сильна забур'яненість Striga лану культурних рослин завдає шкоди рослинам-хазяям, спричинюючи низькорослість, хлороз, в'янення і, нарешті, загибель рослини-хазяїна, особливо у її природному середовищі, де може бути нестача води. Striga є природною у напівпустельних та жарких пасовищних регіонах Африки та Азії, проте вона може розростатись поза своєї природної зони, і у теперішній час забур'яненість цим шкідником існує на сільськогосподарських землях у Сполучених Штатах. Забур'яненість широко розповсюджена в Африці, тому Striga є актуальною проблемою для фермерів у напівпустельних та суб-сахельних регіонах, де сорго є основною харчовою культурою. Дану паразитну рослину важко винищити, оскільки її насіння може лежати у стані спокою у грунті до десяти років перед тим як зійде. Біологічні сигнали для проростання Striga включають стимулювання виділеннями коренів із їх хазяйської рослини. Насіння Striga розвивається шляхом спрямування заразної структури до хазяїна, що приєднується до кореня хазяїна та вторгається у судинну систему хазяїна, позбавляючи у такий спосіб хазяїна води, мінералів та вуглеводнів. Після здійснення первинного контакту з хазяїном розпочинається ініціювання проростання та утворення додаткових коренів, і обкрадання рослини-хазяїна зростає. У природних місцеположеннях, де води мало, Striga ефективно позбавляє рослину-хазяїна мінералів та води. Паразитизм Striga інгібується ALS гербіцидами. Даний винахід не обмежується конкретним механізмом. Дійсно, розуміння механізму не є необхідним для практичної реалізації даного винаходу. Проте, зараження Striga коренів сорго виключається шляхом покриття насіння сорго ALS гербіцидами до його висівання. В одному варіанті насіння гібриду сорго, що містить зародкову плазму, котра надає ALS гербіцидну резистентність зазначеному гібриду сорго, покривається одним або кількома ALS гербіцидами до його висівання. У деяких варіантах насіння з нанесеним покриттям висівається на сільськогосподарські угіддя, де ростуть паразитні види рослин Striga. У деяких варіантах гібридні рослини сорго, що виростають із зазначеного насіння з покриттям, резистентні до зараження Striga. В одному варіанті даний винахід запроваджує гібрид сорго, зародкова плазма якого надає резистентності до ALS гербіцидів і резистентність щодо одного або кількох додаткових гербіцидів із однієї або кількох різних груп гербіцидів. Наприклад, додаткові групи гербіцидів, що застосовуються для інгібування росту бур'янів, включають, проте не обмежуючись цим, інгібітори ліпідного синтезу (наприклад, арилоксифеноксипропіонати, циклогексанодеіони. бензофурани, хлорокарбонові кислоти, фосфородитіоати, тіокарбамати), інгібітори 15 фотосинтезу у фотосистемі II (наприклад, фенілкарбамати, піридазинони, триазини, триазинони, триазолінони, урацили, аміди, сечовини, бензотіадіазинони, нітрили, феніл піридини), інгібітори фотосинтезу у фотосистемі І (наприклад, біпіридиліуми), інгібітори протопорфіриногеноксидази (наприклад, дифенілефіри, N-фенілфталіміди, оксадіазоли, оксизолідиндіони, фенілпіразоли, піримідиндіони, тіадіазоли), інгібітори каротеноїдного біосинтезу (наприклад, піридазинони, піридинкарбоксаміди, ізоксазолідинони, триазоли), інгібітори 4-гідроксифеніл-піруват-діоксигенази (наприклад, калістемони, ізоксазоли, піразоли, ірикетони), інгібітори EPSP синтази (наприклад, гліцини), інгібітори глутамінсинтази (наприклад, фосфінові кислоти), інгібітори дигідроптероатсинтази (наприклад, карбамати), інгібітори мікротрубчастого складання (наприклад, бензаміди, бензойні кислоти, динітроаніліни, фосфороамідати, піридини), інгібітори поділу клітин (наприклад, ацетаміди, хлороацетаміди, оксиацетаміди), інгібітори синтезу клітинної оболонки (наприклад, нітрили, триазолокарбоксаміди) та інгібітори ауксинового транспорту (наприклад, фталамати. семікарбазони). Класичне розмноження сорго Польові культури класично розводять за допомогою способів, в яких використовується перевага рослинного методу(дів) обпилювання. Рослина розглядається як "самозапилювальна", якщо пилок від однієї квітки може бути перенесений на ту саму або іншу квітку, тоді як рослини розглядаються як "перехресно-опилювані", якщо пилок має переноситись від квітки на іншу рослину для здійснення запилення. Рослини, котрі самозапилюються та селекціонуються протягом багатьох поколінь, стають гомозиготними у більшості, якщо не всіх своїх локусів, створюючи у такий спосіб однорідну популяцію справжніх гомозиготних нащадків. Схрещення двох гомозиготних рослин від відмінних джерел або двох різних гомозиготних ліній приведе до створення однорідної популяції гібридних рослин, котрі, більш ніж ймовірно, будуть гетерозиготними у ряді локусів. Схрещення двох рослин, кожна із яких гетерозиготна у ряді локусів, створить генерацію гібридних рослин, котрі генетично відмінні і неоднорідні. Рослини сорго самозапилювальні, але можуть також розмножуватись шляхом перехресного запилення. Ріст гібридів сорго потребує розвитку запильникових батьків (відновлювачів плодючості) та насіннєвих батьківських інбредів з використанням цитоплазматичної чоловічої стерильнофертильної відновлювальної системи, схрещення насіннєвих батьків та оцінки даних кросів. Програми селекційного розмноження поєднують бажані ознаки, у даній заявці бажаною ознакою є резистентність рослин до ALS гербіцидів. Ця ознака вводиться у розмножувальний пул із однієї або кількох ліній, так що нові інбредні лінії створюються шляхом схрещення, з наступною селекцією рослин з бажаною ознакою, з наступним додатковим схрещенням і т.д. Нові інбреди схрещуються з іншими інбредними лініями (наприклад, елітними 95122 16 рослинними лініями, подібними до описаних у даному тексті). Селекційне розмноження починається зі схрещення двох генотипів, таких як Tailwind, та елітної лінії сорго (наприклад, Тх430, Тх2737, Тх2783, 00MN7645, HP162, Wheatland, Тх3042, OK11, QL41, Тх643 та Bt сорго). Якщо зазначені оригінальні два батьки не запроваджують усі бажані характеристики, тоді в розмножувальну популяцію можуть бути включені інші джерела. Наприклад, якщо потрібен гібрид, такий, що бажаним є як ALS гербіцидна резистентність, так і резистентність до іншого гербіциду, як тут описано, тоді можуть бути схрещені рослини з обома цими атрибутами з використанням класичних способів розмноження. У селекційному методі найкращі рослини самозапилюються та піддаються селекції у послідовних генераціях. У наступних генераціях гетерозиготна умова надає шлях до гомогенних ліній як результат самозапилення та селекції. Типово, у селекційному методі практикуються п'ять або більше генерацій самозапилення та селекції (наприклад, S1, S2, S3, S4, S5 і т.д.). Для поліпшення рослинної лінії використовується зворотне схрещування. Зворотне схрещування переносить специфічну бажану ознаку від одного джерела до іншого, що не має такої ознаки. Це здійснюється, наприклад, шляхом схрещування донора (наприклад, Tailwind) з інбредною лінією (наприклад, Тх2783, елітною запильниковою лінією, як тут описано). Нащадки цього кросу потім схрещуються зворотно (тобто піддаються зворотному схрещуванню) з елітною інбредною лінією, з наступною селекцією у результуючому нащадку бажаної ознаки (наприклад, резистентності до ALS гербіцидів). За п'ять або більше генерацій зворотного схрещування з селекцією бажаної потрібної ознаки нащадки є типово гетерозиготними відносно локусу (локусів). що контролює бажаний фенотип, але будуть схожими з елітними батьками відносно інших генетичних ознак. Останнє зворотне схрещування, типово, самозапилювальне, щоб дати чистих нащадків щодо гена, котрий переноситься. У сучасних гібридних інтрогресивних програмах розведення сорго розробляються нові батьківські лінії, котрі являють собою або насіннєвобатьківські лінії (наприклад, Wheatland, Тх3042, N223, 01MN1569, 03MN7645, OK11, QL41 та Тх643) або пилково-батьківські лінії (наприклад, Тх430, Тх2737, Тх2783, R45, 00MN7645 та НР162), у залежності від того, містять вони чи не містять гени, що відновлюють плодючість: насіннєвобатьківські лінії не мають генів, що відновлюють плодючість, і є чоловічо-стерильниими у деяких цитоплазмах (відомі також як рослини "А-лінії"') та чоловічо-фертильними в інших цитоплазмах (відомі також як рослини "В-лінії"), тоді як пилковобатьківські лінії не є чоловічо-стерильними і все ж містять гени, що відновлюють плодючість (відомі також як рослини "R-лінії"). Насіннєво-батьківські лінії, типово, створюються цитоплазматично чоловічо-стерильними, так що пиляки у цих рослинах мінімальні або відсутні, і, отже, не потребує перехресного запилення. Насіннєво-батьківські лінії бу 17 дуть продукувати лише насіння, і цитоплазма передається лише через яйцеклітину. Пилок для перехресного запилення постачається через пилково-батьківські лінії, що містять гени, потрібні для повного відновлення фертильності у F1 гібриді, і даний крос комбінується з чоловічо-стерильними 95122 18 насіннєвими батьками, утворюючи гібрид двох інбредних ліній з високим виходом з хорошою якістю зерна. Приклади рослин R-лінії та В-лінії, що знаходять застосування у даному винаході, включають, проте не обмежуючись цим, ті, що перелічені у Таблиці 1. Таблиця 1 Походження Tx2737///Tx2737//90SN7/Tw Тх2737///Тх2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx273 7//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx2737///Tx2737//90SN7/Tw Tx2737///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw Tx430///Tx2737//90SN7/Tw R45////R45///Tx2737//90SN7/Tw ТХ2783/ТХ2783/ТW N223///N223//N223/Tw Wheatland///N223//N223/Tw Wheatland///N223//N223/Tw N223///N223//N223/Tw N223///N223//N223/Tw OK11////OK11///N223//N223/Tw QL41/////QL41////OK11///N223//N223/Tw 01MN1589////Wht///N223//N223/Tw 01MN1589////Wht///N223//N223/Tw 01MN1589////Wht///N223//N223/Tw 01MN1589////Wht///N223//N223/Tw 03MN954////Wht///N223//N223/Tw Tx3042/////Tx3042////N223///N223//N223/Tw Tx3042/////Tx3042////N223///N223//N223/Tw N223///N223//N223/Tw N223///N223//N223/Tw Звичайно, ця цитоплазматична чоловіча стерильно-фертильна відновлювальна система реалізується для продукування гібридного насіння шляхом посадки блоків рядків чоловічостерильних (насіннєво-батьківських) рослин та блоків рядків фертильно-відновлювальних (пилково-батьківських) рослин, так що насіннєвобатьківські рослини запилюються пилком від пилково-батьківських рослин під дією вітру. Цей процес продукує потужний гібрид двох інбредних ліній, котрий збирається та вирощується споживачем. Чоловічо-стерильні, насіннєво-батьківські рослини можуть також створюватись шляхом генетичного розмноження рецесивних чоловічостерильних генів у конкретній популяції, проте, для розмноження гібридного сорго звичайно викорис Нове джерело MN07-1903 MN07-1905 MN07-1916 MN07-1926 MN07-1935 MN07-1936 MN07-1940 MN07-1941 MN07-1944 MN07-1945 MN07-1956 MN07-1981 MN07-1984 MN07-I987 MN07-1992 MN07-1995 MN07-2013 MN07-2075 MN07-2094 MN07-2113 MN07-2118 MN07-2134 MN07-2136 MN07-2164 MN07-2198 MN07-2230 MN07-2248 MN07-2251 MN07-2254 MN07-2261 MN07-2290 MN07-2293 MN07-2084 MN07-2088 Ген BC2F5 BC2F5 BC2F5 BC2F5 BC2F4 BC2F4 BC2F4 BC2F4 BC2F4 BC2F4 BC2F4 BC2F3 BC2F3 BC2F3 BC2F3 BC2F3 BC2F3 BC2F4 BC2F6 BC2F4 BC2F4 BC2F5 BC2F5 13C3F3 BC4F3 BC4F3 BC3F3 BC3F3 BC3F3 BC3F3 BC4F3 BC4F2 BC2F4 BC2F4 Коментарі R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія R-лінія В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія 75% DPI (QL41) В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія В-лінія товується система, що Є цитоплазматичною чоловічою стерильно-фертильною відновлювальною системою. У роботі Sliper and Poehlman, 2006, Breeding Field Crops, Fifth Ed., Blackwell Publishing, подано детальний огляд сучасних процедур розмноження сорго, і на цю роботу в даному тексті зроблено повне посилання. Даний винахід не обмежується переліченими елітними батьківськими лініями сорго, і фахівцеві у даній галузі зрозуміло, що будь-яка елітна лінія сорго у рівній мірі підпадає під композиції та способи, що тут описані. Рослинні трансгени Гетерологічні гени, що передбачені для експресії у рослинах, спочатку складаються в експресійні вектори, що містять гетерологічний ген та відповідні транскрипційні і трансляційні контрольні 19 елементи, з використанням способів, що добре відомі фахівцям у даній галузі. Способи включають in vitro рекомбінантні ДНК методи, синтетичні методи та in vivo генетичну рекомбінацію. Характерні методи широко описані у даній галузі (дивись, наприклад, Sambrook at аl. (1989), Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, Plainview, N.Y., and Ausubel, F.M. et al. (1989), Current Protocols in Molecular Biology, John Wiley & Sons, New York, N.Y., на які у даному тексті зроблені посилання). Загалом, ці вектори містять нуклеїновокислотну послідовність, що кодує гетерологічний ген. зчеплений оперативно з промотором та іншими регуляторними послідовностями (наприклад, енхансерами, поліаденілувальними сигнальними послідовностями і т.д.), що потрібні для експресії у рослині. Промотори включають, проте не обмежуючись цим, конститутивні промотори, тканинні-, органні- і специфічні до розвитку промотори, та індуковні промотори. Приклади промоторів включають, проте не обмежуючись цим, конститутивний промотор 35S вірусу мозаїки кольорової капусти; індукований ушкодженням промотор із томатів, лейцин амінопептидаза (Chao et al. 1999, Plant Physiol 120:979-992, на яку у даному тексті зроблено повне посилання); хімічно-індуковний промотор із тютюну, Pathogenesis-Related 1 (індукований Sметиловим ефіром саліцилової кислоти та бензотіадіазол-7-тіокарбонової кислоти); промотор температурного шоку (патент США за номером 5187267, на який у даному тексті зроблено повне посилання); індуковний тетрацикліном промотор (патент CШA за номером 5057422, на який у даному тексті зроблено повне посилання); та специфічні щодо насіння промотори. Експресуючі кластери можуть додатково містити будь-які послідовності, потрібні для експресії мРНК. Такі послідовності включають, проте не обмежуючись цим, транскрипційні термінатори, енхансери, такі як інтрони, вірусні послідовності, та послідовності для цілеспрямування і енного продукту на специфічні органели та клітинні компартменти. Для застосування в експресії послідовностей з використанням промоторів, таких які розкриті у даному тексті, мається різновид транскрипційних термінаторів. Транскрипційні термінатори відповідальні за термінацію транскрипції поза даним транскриптом та коректне поліаденілування. Відповідні транскрипційні термінатори та ті, що, як відомо, функціонують у рослинах, включають, проте не обмежуючись цим, CaMV 35S термінатор, tml термінатор, pea rbcS Е9 термінатор та опалін і октапін синтазний термінатор (Odell et аl., 1985, Nature 313:810; Rosenberg et al., 1987, Gene, 56;125; Guerineau et al., 1991. Моl. Gen. Genet. 262:141; Proudfoot, 1991, Cell; 64:671; Sanfacon et al., 1990, Genes Dev. 5:141; Mogen et al., 1990. Plant Cell, 2:1261; Munroe et al., 1990, Gene, 91:151; Ballas et al., 1989, Nucleic Acids Res. 17:7891; Joshi et al.. 1987, Nucleic Acid Res., 15:9627, на які у даному тексті зроблені посилання у повній цілісності). У деяких варіантах конструктивні елементи 95122 20 для експресії гетерологічного гена, який являє інтерес, включають одну або кілька послідовностей, котрі, як знайдено, підсилюють експресію гена із середини транскрипційної одиниці. Ці послідовності можуть бути використані у сполученні з нуклеїновокислотною послідовністю, що являє інтерес, для підсилення експресії у рослинах. Було показано, що різні інтронні послідовності підсилюють експресію, особливо в однодольних клітинах. Інтронні послідовності звичайно введені у рослинні трансформаційні вектори, типово, у нетрансльовану лідерну послідовність. У деяких варіантах конструктивний елемент для експресії потрібної гетерологічної нуклеїновокислотної послідовності також включає регулятор, такий як сигнал ядерної локалізації (Kalderon et al., 1984, Cell 39:499; Lassner et al., 1991, Plant Molecular Biology 17:229), рослинну трансляційну консенсусну послідовність (Joshi. 1987, Nucleic Acids Research 15:6643), інтрон (Luchrsen and Walbot, 1991, Моl. Gen. Genet. 225:81), і таке подібне, оперативно зчеплений з нуклеїновокислотною послідовністю, що кодує гетерологічний ген. При приготуванні конструкта, що містить нуклеїновокислотну послідовність, котра кодує гетерологічний ген, або кодує послідовність, сконструйовану для зниження експресії серологічного гена, можна маніпулювати різними ДНК фрагментами, щоб запровадити ДНК послідовності у бажаній орієнтації (наприклад, сенсовій або антисенсовій), і, як годиться, у погрібній рамці зчитування. Наприклад, можуть бути використані адаптери або лінкери для з'єднання ДНК фрагментів, або можуть бути застосовані інші маніпуляції для запровадження фучних рестрикційних сайтів, вилучення надлишкових ДНК, вилучення рестрикційцих сайтів і такого подібного. З цією метою застосовується, переважно, in vitro мутагенез, відновлення праймерів, рестрикція, гібридизація, резекція, лігірування і таке подібне, де задіяні інсерції, делеції або субституції (наприклад, транзиції та трансверсії). Для трансформації рослин наявні множинні трансформаційні вектори. Вибір вектора для в ВИкористання буде залежати від способу трансформації, якому віддається перевага, та виду мішені для трансформації. Для деяких видів мішеней перевага віддається різним антибіотичним або гербіцидним селектовним маркерам. Селектовні маркери, що звичайно використовуються у трансформації, включають nptll ген, котрий надає резистентності до канаміцину та споріднених антибіотиків (Messing and Vierra, 1982, Gene 19:259; Bevan et al., 1983, Nature 304:184, на які у даному тексті зроблені посилання), bar ген, котрий надає резистентності до гербіцидного фосфінотрицину (White et al., 1990. Nucl Acids Res. 18:1062; Spencer et al., 1990. Theor. Appl. Genet. 79:625, на які у даному тексті зроблені посилання у всій повноті), hph ген, котрий надає резистентності до антибіотичного гігроміцину (Blochlinger and Diggelmann. 1984, Моl. CeIl. Biol. 4:2929, на яку у даному тексті зроблено посилання у всій повноті), та dhfr ген, що надає резистентності до метотрексату (Bourouis et al., 1983, EMBO J.,2:1099, на яку у даному тексті 21 зроблено повне посилання). У деяких варіантах Ті (Т-ДНК) плазмідний вектор адаптований для застосування в опосередкованому Agrobacterium процесі трансфекції, так як у патенті США за номером 6369298 (сорго) та патентах США за номерами 5981839, 6051757, 5981840, 5824877 та 4940838, на які у даному тексті зроблені повні посилання. Конструювання рекомбінантних Ті та Ri плазмід, загалом, йде за методами, що типово використовуються з більш загальними векторами, такими як pBR322. Додатково можуть використовуватись допоміжні генетичні елементи, котрі інколи знаходяться з природними плазмідами і інколи конструюються із сторонніх послідовностей. Вони можуть включати, проте не обмежуючись цим, структурні гени для антибіотичної резистентності як селективні гени. Тепер використовуються дві системи рекомбінантних Ті та Ri плазмідних векторних систем. Перша система називається "коінтегратною" системою. У цій системі шатл-вектор, що містить потрібний ген, уводиться шляхом генетичної рекомбінації у неонкогенну Ті плазміду, що містить як цисдіючі, так і транс-діючі елементи, потрібні для трансформації рослин, як, наприклад, у pMLJ1 шатлвекторі та неонкогенній Ті плазміді PGV3850. Використання Т-ДНК як фланкуючої області у конструкційному елементі для інтеграції у Ті- або Riплазміду описано у ЕPO No. 116718 та PCT заявках за номерами WO 84/02913, 02919 та 02920; Herrera-Estrella. 1983, Nature 303:209-213; Fraley et al., 1983, Proc. Natl. Acad. Sci., USA 80:4803-4807; Horsch et al., 1984, Science 223:496-498; та DeBlock et al., 1984, ЕМВО J. 3:1681-1689, на які у даному тексті зроблені повні посилання. Друга система називається "бінарною" системою, в якій використовуються дві плазміди, і погрібний ген уводиться у шатл-вектор, що містить цисдіючі елементи, погрібні для і трансформації рослини. Інші потрібні функції запроваджуються у транс неонкогенною Ті плазмідою, як проілюстровано pBIN19 шатл-вектором і неонкогенною Ті плазмідою PAL4404. Деякі із цих векторів маються у продажу. У деяких варіантах потрібна нуклеїновокислотна послідовність націлюється на визначений локус на геномі рослини. Сайт-спрямована інтеграція потрібної нуклеїновокислотної послідовності у геном рослинної клітини може досягатись, наприклад, шляхом гомологічної рекомбінації з використанням послідовностей, отриманих із Agrobacterium. Загалом, рослинні клітини піддаються інкубації зі штамом Agrobacterium, котрий містить цілеспрямовуючий вектор, в якому послідовності, гомологічні до ДНК послідовності всередині локусу-мішені, фланковані Agrobacterium трансфер-ДНК (Т-ДНК) послідовностями, як раніше описано (патент США за номером 5501967, на який у даному тексті зроблено повне посилання). Фахівцеві у даній галузі відомо, що гомологічна рекомбінація може досягатись з використанням цілеспрямовуючих векторів, що містять послідовності, котрі є гомологічними до будь-якої частини рослинного гена-мішені, належать вони до регуляторних елементів даного гена або кодуючих діля 95122 22 нок гена. Гомологічна рекомбінація може досягатись у будь-якій області рослинного гена, оскільки нуклеїновокислотна послідовність областей, що фланкують сайт-мішень, відома. Agrobacterium tumefaciens являє собою загальну ґрунтову бактерію, що спричиняє хворобу корончастий гал шляхом переносу деякої частини своєї ДНК до рослини-хазяїна. Перенесена ДНК (Т-ДНК) стабільно інтегрована у геном рослини, де її експресія приводить до синтезу рослинних гормонів і, таким чином, до пухлинного росту даних клітин. Передбачуваний макромолекулярний комплекс утворюється у процесі Т-ДНК переносу із бактеріальної клітини у рослинну клітину. У деяких варіантах нуклеїнові кислоти, як тут розкрито, використовуються для конструювання векторів, одержаних із рослинних (+) PHK вірусів (наприклад, вірус мозаїки костера, вірус мозаїки тютюну, вірус мозаїки люцерни, вірус мозаїки огірка, вірус мозаїки томату та їх комбінації і гібриди). Загалом, уведений гетерологічний полінуклеотид може бути експресований із цих векторів як злитий протеїн (наприклад, протеїн оболонки злитого протеїну) або із його власного промотору чи іншого промотору. Методи для конструювання та використання таких вірусів описані у патентах США за номерами 5846795, 5500360, 5173410, 5965794, 5977438 та 5866785, на які у даному тексті зроблені посилання у всій повноті. У деяких варіантах потрібна гетерологічна нуклеїновокислотна послідовність, що містить мутантний ALS трансген, як отримано у зародковій плазмі KSU 06MN8419, депонованій під ATCC No. РТА-7999, уводиться безпосередньо у рослину. У деяких варіантах мутантний ALS трансген є на принаймні 70% гомологічним, принаймні 80% гомологічним, принаймні 85% і гомологічним, принаймні 90% гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гена, як отримано у зародковій плазмі KSU 06MN8419. У деяких варіантах мутантний ALS трансген, котрий є на принаймні 70% і гомологічним, принаймні 80% гомологічним, принаймні 85% гомологічним, принаймні 90% і гомологічним, принаймні 95% гомологічним, принаймні 97% гомологічним або принаймні 99% гомологічним до ALS гербіцид резистентного гена, як отримано у зародковій плазмі KSU 06MN8419, містить одну або кілька амінокислотних субституцій Val 531 Ilе та Trp545Leu. Один вектор, корисний для методів прямого переносу генів у комбінації з селекцією гербіцидом Basia (або фосфінотрицин), являє собою модифіковану версію плазміди рСІВ246, з промотором CaMV 35S в оперативному злитті з Е. coli GUS геном та CaMV 35S транскрипційним термінатором (WO 93/07278, на яку у даному тексті зроблено посилання). Коли нуклеїновокислотна послідовність, що кодує гетерологічний ген, оперативно зчеплена з відповідним промотором і введена у придатний вектор для застосованого визначеного способу трансформації (наприклад, один із вищеописаних векторів), рекомбінантна ДНК, описана вище, може бути введена у рослинну клітину з використан 23 ням ряду визнаних у даній галузі шляхів. Фахівцям у даній галузі відомо, що вибір способу залежить від типу рослини-мішені для трансформації. У деяких варіантах даний вектор утримується епісомним чином. У деяких варіантах даний вектор інтегрований у геном. У деяких варіантах для введення вектора у рослинну клітину застосовується пряма трансформація в пластидний геном (наприклад, дивись патенти США за номерами 5451513, 5545817, 5545818; PCT заявку WO 95/16783, на які у даному тексті зроблені повні посилання). Базовий спосіб для хлоропластної трансформації включає введення ділянок клонованої пластидної ДНК, фланкуючої селектовний маркер, разом з нуклеїновою кислотою, що кодує потрібні послідовності, у придатну тканину-мішень (наприклад, з використанням біолістиків або протопластної трансформації з хлоридом кальцію або PEG). 1-1,5 kb фланкуючі ділянки, що називаються цілеспрямовуючими послідовностями, полегшують гомологічну рекомбінацію з пластидним геномом і у такий спосіб дозволяють заміну або модифікацію специфічних ділянок даної пластоми. Спочатку точкові мутації у хлоропластних 16S rRNA та rps 12 генах, що надають резистентності до спектиноміцину та/або стрептоміцину, застосовуються як селектовні маркери для трансформації (Svab et al., 1990, Proc. Natl. Acad. Sci., 87:8526; Staub and Maliga. 1992, Plant Cell, 4:39, на які у даному тексті зроблено посилання). Присутність клонуючих сайтів між цими маркерами дозволяє здійснювати з допомогою пластидного спрямовуючого вектора уведення молекул сторонньої ДНК (Staub and Maliga, 1993, EMBO J., 12:601). Значні підвищення частоти трансформації одержуються шляхом заміни рецесивної рРНК або r-протеїн антибіотик резистентних генів на домінантний селектовний маркер, бактеріальний aadA ген, що кодує спектиноміцин-детоксифікуючий ензим аміноглікозид-3'-аденілтрансферазу (Svab and Maliga, 1993, Proc. Natl. Acad. Sci., 90:913). У даній галузі відомі й інші селектовні маркери, корисні для пластидної трансформації, котрі підпадають під обсяг даного винаходу. Одержуються рослини, гомоплазматичні до пластидних геномів, що містять дві зазначені нуклеїновокислотні послідовності, розділені промотором даного винаходу, і вони, переважно, здатні до високої експресії РНК, кодованих молекулою ДНК. В одному варіанті вектори, корисні у практиці даного винаходу, мікроін'єктуються прямо у рослинні клітини (Crossway, 1985, Моl. Gen. Genet. 202:179). У деяких варіантах даний вектор переноситься у клітину рослини з використанням поліетиленгліколю (Krens et al., 1982, Nature, 2%:72; Crossway et al., 1986, Bio Techniques, 4:320); злиття протопластів з іншими об'єктами, такими як мініклітини, клітини, лізосоми або інші, здатні до злиття тіла з ліпідними поверхнями (Fraley et al., 1982. Proc. Natl. Acad. Sci., USA, 79:1859); та протопласт трансформації (EP 0292435); прямого переносу гена (Paszkowski et al., 1984, EMBO J., 3:2717; Hayashimoto et al., 1990, Plant Physiol. 93:857). У деяких варіантах даний вектор може бути також уведений у рослинні клітини шляхом елект 95122 24 ропорації. (Fromm, et аl.. 1985, Proc. Natl. Acad. Sci. USA 82:5824; Riggs et al., 1986. Proc. Natl. Acad. Sci. USA 83:5602). У цьому способі рослинні протопласти піддаються електропорації у присутності плазмід, що містять генний конструкційний елемент. Електричні імпульси поля високої напруженості зворотним чином порушують проникність біомембран, надаючи можливість уведення плазмід. Піддані електропоруванню рослинні протопласти реформують стінку клітини, поділяють і утворюють рослинний калюс. На додаток до прямої трансформації, у деяких варіантах, вектори, що містять нуклеїновокислотну послідовність, яка кодує гетерологічний ген, переносяться з використанням трансформації, опосередкованої Agrobacterium (Hinchee et al., 1988, Biotechnology, 6:915; Ishida et al., 1996, Nature Biotechnology 14:745, на які у даному тексті зроблені посилання). Agrobacterium є репрезентативним представником роду грам-негативної родини Rhizobiaceae. Його види відповідальні за рослинні пухлини, такі як корончастий гал та хвороба волосатого кореня. У диференційованій тканині, яка характерна для пухлин, утворюються та катаболізуються амінокислотні похідні, відомі як опіни. Бактеріальні гени, відповідальні за експресію опінів, є зручним джерелом контрольних елементів для химеричних експресуючих кластерів. Гетерологічні генетичні послідовності (наприклад, нуклеїновокислотні послідовності, оперативно зчеплені з промотором даного винаходу) можуть бути введені у відповідні рослинні клітини за допомогою Ті плазміди Agrobacterium tumefaciens (описаної раніше). Ті плазміда переноситься у рослинні клітини при зараженні Agrobacterium tumefaciens і стабільно інтегрується у рослинний геном (Schell, 1987, Science, 237:1176). Види, що чутливі до інфікування Agrobacterium. можуть бути трансформовані in vitro. Способи трансформації для продукування трансгенних рослин сорго з використанням трансформації, опосередкованої Agrobacterium, запроваджені у патенті США за номером 6369298. У деяких варіантах зазначений вектор уводиться шляхом балістичного прискорення частинок (патент США за номером 4945050; Casas et al., 1993. Proc. Natl. Acad. Sci. USA 90:11212, на які у даному тексті зроблені повні посилання). У деяких варіантах після селекції трансформованого рослинного матеріалу, що може експресувати гетерологічний ген, який кодує гетерологічний протеїн або його варіант, усі рослини регенеруються. Регенерація рослин із культивованих протопластів описана у роботах Evans et al., Handbook of Plant Cell Cultures, Vol. 1: (MacMillan Publishing Co. New York, (1983); Vasil I.R. (ed.), Cell Culture and Somatic Cell Genetics of Plants, Acad. Press, Orlando, Vol. 1, (1984) та Vol. Ill, (1986), на які у даному тексті зроблені повні посилання. Відомо, що багато рослин можуть бути регенеровані із культурних клітин або тканин, включаючи, проте не обмежуючись цим, усі головні види цукрової тростини, цукрові буряки, бавовну, фруктові та інші дерева, бобові та овочі і однодольні (наприклад, рослини, що описані вище). Засоби для регенерації варіюють від виду до виду рослин, але, 25 загалом, спочатку запроваджується суспензія трансформованих протопластів, що містить копії гетерологічного гена. Утворюється калюсна тканина, і із калюсу можуть бути індуковані пагінці, котрі потім укорінюються. Як альтернатива, формування зародку може бути індуковано із протопластної суспензії. Ці зародки розвиваються і утворюють зрілі рослини. Культуральні середовища містять, загалом, різні амінокислоти та гормони, такі як ауксин та цитокініни. Пагінці та корені звичайно розвиваються одночасно. Ефективність регенерації буде залежати від середовища, генотипу та історії даної культури. Відтворюваність регенерації залежить від контролю цих змінних. Приклади Приклад 1 - Гербіцидна резистентність у генотипі сорго Насіння дикого генотипу сорго, котрий, як було показано, виявляє ALS резистентний фенотип, позначений "Tailwind", та чутливий до гербіциду генотип сорго, позначений 90SN7, висівали у 12 літрові горщики у теплиці. Кілька рослин кожного типу обприскували або 1) Lightning Herbicide (комбінація імазетаніру та імазапіру) при нормі 2,56 -1 унцій акрс (2х обробка гербіцидом), 2) Steadfast® Herbicide (DuPont™; комбінація нікосульфурону та -1 римсульфурону) при нормі 1,50 унцій акрс (2х обробка гербіцидом) або 3) комбінація Lightning -1 Herbicide при нормі 2,56 унцій акрс та Steadfast® ТМ -1 Herbicide (DuPont при нормі 1,50 унцій акрс . Для кожної обробки рослини Tailwind не виявляли, по суті, ушкодження за 12 діб після аплікації гербіциду, тоді як рослини 90SN7 загинули, демонструючи, що Tailwind мав перехресну резистентність до IMI (Lightning Herbicide) та SU (Steadfast® Herbicide) класів ALS інгібуючих гербіцидів. Приклад 2 - Кроси Tailwind з слізними батьківськими лініями сорго Tailwind схрещувався з різними елітними батьками, включаючи Тх430 та Wheatland. F2 популяції, одержані від кросів з цими батьками, оцінювались за методом сегрегаційного аналізу для визначення кількості генів, включених в експресію толерантності. Популяції рослин вирощувались у теплиці і обприскувались 1x та 3х дозами Accent® Herbicide (DuPont™; нікосульфурон), Option® Herbicide (BayerCropScience; форамсульфурон), та Steadfast® Herbicide. Підрахунки кількості живих/загиблих рослин дозволили провести генетичний аналіз. Сегрегаційний аналіз дав єдиний головний, частково домінантний ген у популяції, що була одержана від Тх430 для кожної гербіцидної обробки. Схожі аналізи популяцій, отриманих від кросів з Wheatland, дали єдиний головний, частково домінантний ген, так само як і потенційно два або три гени-модифікатори, що впливали на відносну експресію ознаки толерантності. Спроби розмноження рослин ініціювались шляхом внесення зворотним схрещуванням ознаки толерантності у комерційно важливі елітні батьківські запилювальні рослини сорго, включаючи 95122 26 Тх430, Тх2737, Тх2783, 00MN7645 та HP162, так само як і комерційно важливі елітні батьківські насіннєві рослини сорго, включаючи Wheatland, Тх3042, OK11, QL41 та Тх643, із селекцією гербіцидної толерантності у кожній генерації. Результуюче насіння від кросу Tailwind з Тх2783 (ВС2F3:F4) було депоноване в ATCC для загального доступу. Приклад 3 - Секвенірування генів щодо ALS резистентного гена Спроби секвенірування генів ініціювались для визначення того, чи існувала для гербіцид толерантного фенотипу цільова візуальна мутація. Пошук послідовностей здійснювався з використанням інструмента National Institute of Health's Basic Local Alignment Search Tool (BLAST) в інституті The Institute for Genomic Research (TIGR). З допомогою бази даних по зібранню рослинних транскриптів (Plant Transcript Assemblies) було ідентифіковане зібрання транскриптів (Transcript Assembly), що репрезентує сорго ALS ген (ТА3960 4558; SEQ ID NO:1). Були сконструйовані ампліфікаційні праймери для полімеразної ланцюгової реакції (ПЛР), F4r-CACATCACCCTTGTACCAGCTC (SEQ ID NO: 3) та В5-GATTGTGCACATTGATATTGATCC (SEQ ID NO: 4), для ампліфікації ділянок гена сорго, аналогічних ділянкам Arabidopsis thaliana AHAS гена, що, як вважається, впливає на експресію ALS-гербіцидної толерантності (A. thaliana: Ala122, Рго197, Ala205, Trp574 та Ser653; Tan et al., 2005). I Ампліфікаційні праймери полімеразної ланцюгової реакції були з успіхом використані для ампліфікації ділянки гена у гербіцид резистентному (S1-1, S1-2 та S1-3) та чутливих до гербіциду (Тх623 та Тх430) генотипах сорго з використанням наступних умов термоциклювання: денатурація при 94 C протягом 60 с, відпал при 62 C протягом 45 с та екстензія при 72 C протягом 45 с. ПЛР ампліфікаційні продукти очищали з використанням очисного набору QIAquick PCR Purification Kit (QIAGEN) та піддавали секвеніруванню на установці для секвенірування в університеті штату Канзас. Отримана амінокислотна послідовність (SEQ ID NО:2) виявила мутації у Val531Ile (GTC - АТС), що відповідають А. thaliana Val560lle, та Trp545Leu (TGG - TTG), що відповідають A. thaliana Trp574Leu, у гербіцид резистентних генотипах (Фігура). На всі згадані у даній заявці публікації та патенти зроблені посилання. Різні модифікації та варіації описаних методів та композицій винаходу будуть очевидними фахівцям у даній галузі без відходу від обсягу та суті даного винаходу. Хоча даний винахід описаний у зв'язку зі специфічними варіантами, яким віддається перевага, слід розуміти, що винахід, згідно з формулою винаходу, не має бути надто обмеженим такими специфічними варіантами. Дійсно, різні модифікації описаних методів для реалізації винаходу, котрі зрозумілі фахівцям у споріднених галузях, як мається на думці, підпадають під обсяг наступних пунктів формули винаходу. 27 Комп’ютерна верстка А. Крулевський 95122 28 Підписне Тираж 24 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюSorgho resistant towards acetolactate synthase herbicide
Автори англійськоюTuinstra, Mitchell, R., Al-Khatib, Kassim
Назва патенту російськоюСорго, резистентный к ацетолактатсинтазному гербициду
Автори російськоюТуинстра Митчел Р., Ал-Хатиб Кассим
МПК / Мітки
МПК: A01H 5/00, C12N 15/82
Мітки: гербіциду, резистентний, сорго, ацетолактатсинтазного
Код посилання
<a href="https://ua.patents.su/14-95122-sorgo-rezistentnijj-do-acetolaktatsintaznogo-gerbicidu.html" target="_blank" rel="follow" title="База патентів України">Сорго, резистентний до ацетолактатсинтазного гербіциду</a>
Спосіб вирощування сорго цукрового як енергетичної культури

Номер патенту: 55786
Опубліковано: 27.12.2010
Автори: Сторожик Лариса Іванівна, Мандровська Світлана Миколаївна, Балан Василь Миколайович, Петриченко Сергій Миколайович
МПК: A01B 79/00
Мітки: сорго, культури, цукрового, вирощування, енергетичної, спосіб
Формула / Реферат:
Спосіб вирощування сорго цукрового як енергетичної культури, що включає передпосівну підготовку ґрунту, внесення органічних та мінеральних добрив, посів в першій декаді травня та догляд за рослинами, який відрізняється тим що сівбу сорго цукрового проводять сумісно з маточниками буряків з шириною міжряддя 45 см та нормою висіву 10-12 кг/га.
Спосіб боротьби з бур’янами у посівах сорго

Номер патенту: 5584
Опубліковано: 28.12.1994
Автори: Георг Пассіотас, Вернер Фьорі, Хенрі Щепанські, Хенрі Мартін
МПК: A01N 37/34, C07C 317/44, A01N 43/32, A01N 43/02, C07D 295/185, C07C 311/16, C07C 275/50, C07C 239/00, A01N 41/10, A01N 43/26, A01N 47/02, C07C 251/32, C07D 263/14, A01N 25/32, C07C 313/00, C07D 317/22, A01N 37/50, C07C 323/47, C07C 317/28, C07D 319/00, C07D 317/18, A01N 43/28, C07C 67/00, A01N 47/12, C07C 251/58
Мітки: бур'янами, посівах, сорго, спосіб, боротьби
Формула / Реферат:
Способ борьбы с сорняками в посевах сорго путем предварительной обработки семян антидотом - производным бензацетонитрила и до-всходовой обработки почвы гербицидом N-(1-метил-2-метоксиэтил)-N-хлорацетил-2-этил-6-метиланилином в дозе 1,5-2,5 кг/га, отличающийся тем, что, с целью уменьшения фитоток-сического воздействия гербицида на сорго, в качестве производного бензацетонитрила используют соединение формулыв количестве 0,75-4...
Спосіб підвищення польової стійкості рослин картоплі до гербіциду типу тітус

Номер патенту: 28279
Опубліковано: 10.12.2007
Автори: Шарапа Микола Григорович, Каліцький Петро Францевич, Мороз Іван Харитонович, Кононученко Валерій Васильович
МПК: A01P 13/02, A01N 25/32
Мітки: типу, підвищення, картоплі, гербіциду, спосіб, тітус, рослин, стійкості, польової
Формула / Реферат:
Спосіб підвищення польової стійкості рослин картоплі до гербіциду типу тітус, що включає застосування гербіциду проти бур'янів шляхом обробки посівів сумішшю гербіциду по сходах картоплі та добавки, який відрізняється тим, що використовують як добавку біостимулятор росту рослин емістим С з нормою внесення 5-10 мл на 1 га посадок.
Спосіб отримання харчового сиропу із цукрового сорго

Номер патенту: 26940
Опубліковано: 10.10.2007
Автори: Салавор Оксана Мирославівна, Григоренко Наталія Олександрівна, Штангеєва Надія Іванівна, Штангеєв Валерій Остапович
МПК: C13D 1/00
Мітки: спосіб, сорго, сиропу, цукрового, харчового, отримання
Формула / Реферат:
Спосіб отримання харчового сиропу із цукрового сорго, що передбачає очищення та подрібнення стебел, вилучення соку, відокремлення мезги, коагуляцію нецукрів, клейстеризацію крохмалю, ферментативний гідроліз, інактивацію ферментів, коагуляцію високомолекулярних сполук, фільтрування та концентрування, який відрізняється тим, що для коагуляції високомолекулярних сполук колоїдної дисперсності використовують флокулянт-антисептик, діючою речовиною...
Спосіб реєстрації генотипів сорго

Номер патенту: 48475
Опубліковано: 25.03.2010
Автори: Сиволап Юрій Михайлович, Шевчук Ганна Юріївна, Кожухова Наталія Едуардівна
МПК: A01H 1/00
Мітки: реєстрації, генотипів, спосіб, сорго
Формула / Реферат:
Спосіб реєстрації генотипів сорго, що включає аналіз ідентифікаційних ознак сорту, який відрізняється тим, що опис ідентифікаційних ознак здійснюється у вигляді формули, що відображає певний алельний склад мікросателітних локусів генотипу сорго, які досліджували за допомогою полімеразної ланцюгової реакції.
Попередній патент: Пристрій та спосіб запобігання вибуху електричного трансформатора
Наступний патент: Багатофункціональна іграшка
Випадковий патент: Спосіб оптимізації лікування алкогольного цирозу печінки