Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в об’ємному зразку нуклеїнової кислоти зі щонайменше 100 організмів






Формула / Реферат
1. Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в об′ємному зразку нуклеїнової кислоти зі щонайменше 100 організмів, причому спосіб містить:
приведення зразка нуклеїнової кислоти в контакт з:
прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайта вставки; і
першим зворотним праймером, здатним зв′язуватися зі вставленою нуклеотидною послідовністю, і
другим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайта вставки;
застосування праймерів для ампліфікації нуклеїнових кислот між праймерами; і
аналіз ампліфікованих нуклеїнових кислот для визначення, присутності або відсутності вставленої нуклеотидної послідовності в зразку нуклеїнової кислоти, де репродуковані нуклеїнові кислоти включають послідовність, яка відповідає послідовності між прямим праймером і першим зворотним праймером в разі присутності вставленої нуклеотидної послідовності, і де репродуковані нуклеїнові кислоти включають послідовність, яка відповідає послідовності між прямим праймером і другим зворотним праймером в разі відсутності вставленої нуклеотидної послідовності.
2. Спосіб за п. 1, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого при наявності вставленої нуклеотидної послідовності.
3. Спосіб за п. 1, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого за відсутності вставленої нуклеотидної послідовності.
4. Спосіб за п. 2, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого за відсутності вставленої нуклеотидної послідовності.
5. Спосіб за п. 1, в якому нуклеїнові кислоти зразка містять більше ніж один повтор визначеного сайта вставки.
6. Спосіб за п. 1, в якому вставлена нуклеотидна послідовність присутня менше ніж в 1 % конкретних сайтів вставки в нуклеїновій кислоті.
7. Спосіб за п. 1, в якому вставлена нуклеотидна послідовність присутня більше ніж в 99 % конкретних сайтів вставки в нуклеїновій кислоті.
8. Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності у визначеному сайті вставки, в об'ємному зразку нуклеїнової кислоти з множини щонайменше 100 організмів, причому спосіб включає:
розділення зразка нуклеїнової кислоти на щонайменше дві частини;
здійснення першого аналізу на основі реакції ампліфікації, що включає
приведення першої частини зразка нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїнової кислотою вище сайта вставки, і першим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою в межі вставленої нуклеотидної послідовності;
застосування прямого праймера і першого зворотного праймера для отримання першого продукту ампліфікації, і
визначення кількості першого продукту ампліфікації;
здійснення другого аналізу на основі реакції ампліфікації, що включає
приведення другої частини зразка нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайта вставки; і другим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайта вставки;
застосування прямого праймера і другого зворотного праймера для одержання другого продукту ампліфікації, і
визначення кількості другого продукту ампліфікації; і
порівняння кількості першого продукту ампліфікації з кількістю другого продукту ампліфікації в зразку нуклеїнової кислоти, де кількість першого продукту ампліфікації вказує на кількість вставленої нуклеотидної послідовності, що присутня в зразку нуклеїнової кислоти.
9. Спосіб за п. 8, де здійснення першої реакції ампліфікації додатково містить приведення першої частини зразка нуклеїнової кислоти в контакт під час або після ампліфікації із зондом, специфічним для першого продукту ампліфікації.
10. Спосіб за п. 8, де здійснення другої реакції ампліфікації додатково містить приведення другої частини зразка нуклеїнової кислоти в контакт під час або після ампліфікації із зондом, специфічним для другого продукту ампліфікації.
11. Спосіб за п. 8, в якому нуклеотидні кислоти зразка містять більше ніж один повтор визначеного сайта вставки.
12. Спосіб за п. 8, в якому вставлена нуклеотидна послідовність присутня менше ніж в 1 % конкретних сайтів вставки в зразку нуклеїнової кислоти.
13. Спосіб за п. 8, в якому вставлена нуклеотидна послідовність присутня більше ніж в 99 % конкретних сайтів в зразку нуклеїнової кислоти.
14. Спосіб за п. 11, в якому спосіб використовують для визначення зиготності.
15. Спосіб за п. 11, в якому спосіб використовують для визначення домішки події вставки в зразку.
16. Спосіб за п. 11, в якому спосіб використовують як індикатор наявності/відсутності в об'ємному зразку додаткової події вставки.
Текст
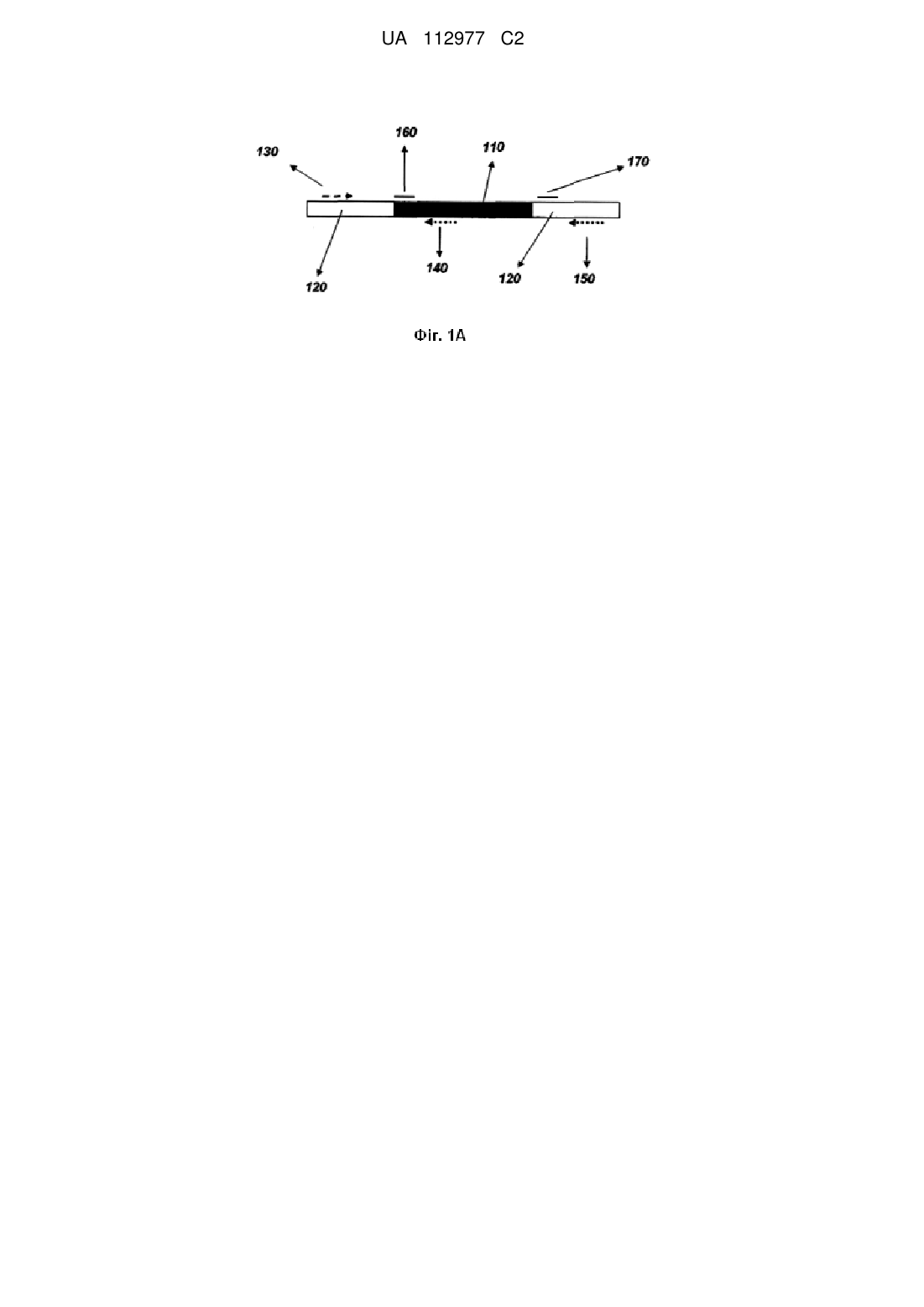
Реферат: Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в об’ємному зразку нуклеїнової кислоти зі щонайменше 100 організмів включає приведення зразка нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки, з першим зворотним праймером, здатним зв′язуватися з вставленою нуклеотидною послідовністю і другим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайту вставки; праймери застосовують для ампліфікації нуклеїнових кислот між праймерами; здійснюють аналіз ампліфікованих нуклеїнових кислот для визначення присутності або відсутності вставленої нуклеотидної послідовності в зразку нуклеїнової кислоти. Спосіб може бути здійснено в межах однієї реакції або з розділенням досліджуваного зразка на щонайменше 2 частини. Спосіб використовують для визначення зиготності, для визначення домішки події вставки в зразку, як індикатор наявності/відсутності в об'ємному зразку додаткової події вставки. UA 112977 C2 (12) UA 112977 C2 UA 112977 C2 5 10 15 20 25 30 35 40 45 50 55 60 ДОМАГАННЯ НА ПРІОРИТЕТ За даною заявкою вимагається пріоритет по даті подачі попередньої патентної заявки Сполучених Штатів серії № 61/428142, поданій 29 грудня 2010 року, відносно "Способів визначення зиготності в об'ємній пробі". РІВЕНЬ ТЕХНІКИ Проведення контролю якості відносно якої-небудь домішки в остаточній лінії надто важливе для успішної продукції гібридного насіння і збереження і зміцнення хороших ділових відносин з покупцями. Забруднення при сходженні насіння остаточної лінії може відбуватися через запилення не передбачених трансгенних або нетрансгенних рослин, що виросли поблизу ділянки продукції, або в процесі обробки насіння і, найважливіше, домішка може виникати внаслідок розсіювання пилка стерильних рослин. Для досягнення повної гомозиготності остаточної лінії і звільнення від будь-яких гемізиготних, нульових або непередбачених трансгенних ліній на сьогоднішній день пропонується спосіб tester-row. У способі tester-row для оцінки статусу зиготності на основі рівнів білків окремої рослини використовується технологія ELISA. Оскільки аналіз базується на основі однієї рослини, він займає дуже багато часу, а також є таким, що дорого коштує. Крім того, здійснення способу tester-row в польових умовах створює додаткові витрати. Спосіб ELISA використовується для визначення сайленсингу експресії трансгена шляхом визначення рівня білка. Він є нечутливим і ненадійним для ідентифікації будь-якої гемізиготної, нульової або будь-якої іншої непередбаченої домішки в остаточній партії насіння. Крім того, при використанні способу ELISA для випробування на наявність малих домішок необхідна роздільна обробка тканин. ОПИС Конкретні варіанти здійснення винаходу включають способи визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в нуклеїновій кислоті. Варіанти здійснення можуть містити: виділення нуклеїнової кислоти з об'ємної проби; приведення нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки, і зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайту вставки. Праймери можуть бути використані для реплікації нуклеїнових кислот між праймерами. Репліковані нуклеїнові кислоти можуть бути проаналізовані для ідентифікації наявності або відсутності в об'ємній пробі вставленої нуклеотидної послідовності. Варіанти здійснення можуть містити: виділення нуклеїнової кислоти із зразка; приведення нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки, і зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайту вставки. Праймери можуть бути використані для реплікації нуклеїнових кислот між праймерами в першій порції і другій порції. Репліковані нуклеїнові кислоти можуть бути проаналізовані для ідентифікації наявності або відсутності в зразку вставленої нуклеотидної послідовності. Друга реакція, або об'єднана з вищезгаданою реакцією, або самостійна, може бути здійснена, використовуючи прямий праймер і зворотний праймер, ця реакція ідентифікує ендогенний ген або послідовність. Ця друга реакція може бути використана як внутрішній контроль для визначення якості і кількості використаних ДНК і/або умов проведення ПЛР. КОРОТКИЙ ОПИС КРЕСЛЕНЬ На фіг. 1А представлене схематичне зображення компонентів способу визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки згідно з варіантом здійснення винаходу. На цій фігурі ймовірно вставлена нуклеотидна послідовність (110) показана темним блоком, в той час як оточуючий геном (120) вказаний відкритими сегментами. Також показані прямий праймер (130), перший зворотний праймер (140) і другий зворотний праймер (150). Додатково представлені необов'язкові специфічний зонд вставки (160) і специфічний зонд дикого типу (170). На фіг. 1В ілюструється модифікований аналіз, який включає створення стандартного протоколу визначення зиготності в двох окремих реакціях: реакції 1, що включає загальний праймер і специфічний праймер дикого типу і специфічний зонд дикого типу (FAM); і реакції 2, що включає внутрішній контроль (ген інвертази 1) із зондом VIC. На фіг. 2А представлене схематичне зображення першого реплікованого продукту (200) згідно з варіантом здійснення винаходу. Також представлені прямий праймер (130), перший зворотний праймер (140) і необов'язковий специфічний зонд вставки (160). На фіг. 2В представлене схематичне зображення першого реплікованого продукту (200) згідно з варіантом здійснення винаходу. Також представлені прямий праймер (130), другий зворотний праймер (150) і необов'язковий специфічний зонд дикого типу (170). 1 UA 112977 C2 5 10 15 20 25 30 35 40 45 50 55 60 На фіг. 3 представлене схематичне зображення компонентів способу визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки згідно з варіантом здійснення винаходу. Перша реакція (400) включає ймовірно вставлену нуклеотидну послідовність (110), яка представлена темним блоком, в той час як оточуючий геном (120) вказаний відкритими сегментами. Також показані прямий праймер (130), перший зворотний праймер (140) і необов'язковий специфічний зонд вставки (160). Друга реакція (500) включає ймовірно вставлену нуклеотидну послідовність (110), яка представлена темним блоком, в той час як оточуючий геном (120) вказаний відкритими сегментами. Також показані прямий праймер (130), другий зворотний праймер (150) і необов'язковий специфічний зонд дикого типу (170). На фіг. 4 представлене графічне відображення результатів флуоресценції FAM в системі LightCycler 480 фірми Roche. На фіг. 5 представлене графічне відображення результатів флуоресценції VIC в системі LightCycler 480 фірми Roche. ДОКЛАДНИЙ ОПИС ВИНАХОДУ Варіанти здійснення винаходу включають способи визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в зразку нуклеїнових кислот. У деяких варіантах здійснення нуклеїнові кислоти можуть бути виділені і/або очищені з одного джерела або сукупності джерел, причому сукупність може включати одне або декілька окремих джерел, кожне з яких може або не може мати відмінні нуклеїнові кислоти. У інших варіантах здійснення джерело нуклеїнових кислот може являти собою, але ними не обмежуючись, тварину, рослину, бактерій, архебактерій, найпростіших, гриби, хромистів, еукаріот, прокаріот, in vivo, in vitro, клітину, насіння, гамету, кукурудзу, сою, пшеницю, рапс, рис і створені джерела. У визначених варіантах здійснення спосіб може містити отримання, виділення, очищення і/або часткове очищення нуклеїнової кислоти. Згідно фіг. 1, виділені нуклеїнові кислоти можуть бути приведені в контакт з прямим праймером (130), здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки (120), і першим зворотним праймером (140), здатним специфічно зв'язуватися з послідовністю всередині вставленої нуклеотидної послідовності (110) (при наявності), і другим зворотним праймером (150), здатним специфічно зв'язуватися з послідовністю нижче (120) сайту вставки, залишаючи праймери для відпалу з виділеними нуклеїновими кислотами. Послідовності, що знаходяться між праймерами, потім по можливості можуть бути репліковані, використовуючи праймери для реплікації між праймерами за допомогою техніки, добре відомої в даній галузі, таких як, але нею не обмежуючись, полімеразна ланцюгова реакція (ПЛР). Відносно сайтів вставки, де вставлена нуклеотидна послідовність (110) присутня і більша ніж фрагмент, що реплікується стандартними способами (наприклад, >5 т. н.), продукти реплікації можуть включати перший реплікований продукт (фіг. 2А (200)), що містить такі послідовності між прямим праймером (130) і першим зворотним праймером (140), але, як правило, може бути відсутнім другий реплікований продукт (фіг. 2В (300)), праймований з другого зворотного праймера (150). Відносно сайтів вставки, де вставлена нуклеотидна послідовність відсутня, продукти репродукції можуть включати другий реплікований продукт (фіг. 2В (300)), що містить такі послідовності між прямим праймером (130) і другим зворотним праймером (150). При наявності вставленої нуклеотидної послідовності (110) в деяких, але не у всіх сайтах вставки в нуклеїновій кислоті, буде виходити суміш двох продуктів. Результати реплікації потім аналізують для визначення наявності і/або відносних рівнів першого реплікованого продукту (200) і/або другого реплікованого продукту (300). Відносно сайтів вставки при наявності вставленої нуклеотидної послідовності, продукти реплікації нуклеїнової кислоти можуть включати перший реплікований продукт (фіг. 2А (200)), що містить такі послідовності між прямим праймером (130) і першим зворотним праймером (150). Відносно сайтів вставки при відсутності вставленої нуклеотидної послідовності, продукти реплікації можуть включати другий реплікований продукт (фіг. 2В (300)), що містить такі послідовності між прямим праймером (130) і другим зворотним праймером (150). При наявності вставленої нуклеотидної послідовності (110) в деяких, але не у всіх сайтах вставки в нуклеїновій кислоті, буде виходити суміш двох продуктів. Результати реплікації потім аналізують для визначення наявності і/або відносних рівнів першого реплікованого продукту (200) і/або другого реплікованого продукту (300). У деяких варіантах здійснення при наявності вставленої нуклеотидної послідовності в сайті вставки прямий праймер і перший зворотний праймер повинні відстояти один від одного менше ніж приблизно на 5 т. н., і прямий праймер і другий зворотний праймер повинні відстояти один від одного більше ніж приблизно на 5 т. н. В додаткових варіантах здійснення, в яких в сайті 2 UA 112977 C2 5 10 15 20 25 30 35 40 45 50 55 60 вставки відсутня вставлена нуклеотидна послідовність, прямий праймер і другий зворотний праймер повинні відстояти один від одного менше ніж приблизно на 5 т. н. У визначених варіантах здійснення згідно з фіг. 3 виділена нуклеїнова кислота може бути розділена на декілька порцій. Перша порція нуклеїнової кислоти може бути приведена в контакт з прямим праймером (130), здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки (120), і першим зворотним праймером (140), здатним специфічно зв'язуватися з послідовністю всередині вставленої нуклеотидної послідовності (110) (при наявності), залишаючи праймери для відпалу з виділеними нуклеїновими кислотами. Послідовності, розташовані між праймерами, потім по можливості можуть бути репліковані, використовуючи праймери для реплікації між праймерами за допомогою техніки, добре відомої в даній галузі, таких як, але нею не обмежуючись, ПЛР. Відносно сайтів вставки, коли вставлена нуклеотидна послідовність (110) присутня, продукти реплікації можуть включати перший реплікований продукт (фіг. 2А (200)), що містить такі послідовності між прямим праймером (130) і першим зворотним праймером (140). Друга порція нуклеїнової кислоти може бути приведена в контакт з прямим праймером (130), здатним зв'язуватися з нуклеїновою кислотою вище сайту вставки (120), і другим зворотним праймером (150), здатним специфічно зв'язуватися з послідовністю нижче (120) сайту вставки, залишаючи праймери для відпалу з виділеною нуклеїновою кислотою. Послідовності, розташовані між праймерами, потім по можливості можуть бути репліковані, використовуючи праймери для реплікації між праймерами за допомогою техніки, добре відомої в даній галузі, таких як, але нею не обмежуючись, ПЛР. Відносно сайтів вставки, коли вставлена нуклеотидна послідовність (110) відсутня, продукти реплікації можуть включати другий реплікований продукт (фіг. 2В (300)), що містить такі послідовності між прямим праймером (130) і другим зворотним праймером (150). Відносно сайтів вставки, коли вставлена нуклеотидна послідовність присутня, продукти реплікації першої порції нуклеїнової кислоти можуть включати перший реплікований продукт (фіг. 2А (200)), що містить такі послідовності між прямим праймером (130) і першим зворотним праймером (150). Відносно сайтів вставки, коли вставлена нуклеотидна послідовність (110) відсутня, продукти реплікації другої порції нуклеїнової кислоти можуть включати другий реплікований продукт (фіг. 2В (300)), що містить такі послідовності між прямим праймером (130) і другим зворотним праймером (150). У інших варіантах здійснення способи можуть бути використані для визначення наявності вставки в нуклеїновій кислоті при наявності вставки менше, ніж в 10 %, 9 %, 8 %, 7 %, 6 %, 5 %, 4 %, 3 %, 2 % або 1 % сайтів вставки в нуклеїнових кислотах. У варіантах здійснення способи можуть бути використані для визначення відсутності вставки в нуклеїновій кислоті при відсутності вставки менше ніж в 10 %, 9 %, 8 %, 7 %, 6 %, 5 %, 4 %, 3 %, 2 % або 1 % сайтів вставки в нуклеїнових кислотах. У деяких варіантах здійснення нуклеїнова кислота, що містить сайт вставки, може являти собою будь-який вид нуклеїнової кислоти, до яких належать, але ними не обмежуючись, ДНК, РНК, PNA або інші модифіковані форми нуклеїнових кислот. У додаткових варіантах здійснення наявність і/або кількість першого і/або другого продукту реплікації може бути визначена будь-якими засобами, відомими в даній галузі, такими як, але ними не обмежуючись, специфічні зонди вставки (160) і специфічні зонди дикого типу (170), відповідно. У визначених варіантах здійснення зонд може являти собою нуклеотидну послідовність, здатну зв'язуватися в специфічному сайті в першому і/або другому продукті реплікації. Відпал зонда може відбуватися в процесі або після реплікації. За допомогою прикладів, що не мають обмежувального характеру, наявність і/або кількість першого і/або другого продукту реплікації може бути визначена, використовуючи хроматографію, гелі, мітки, фрагменти, саузерн-блот і нозерн-блот. У деяких варіантах здійснення для полегшення детекції до одного або декількох зондів може бути приєднаний флуорофор. Крім того, до зонда також може бути приєднана молекула, що гасить флуорофор. До прикладів таких зондів, що містять флуорофор і молекулу, що гасить ® флуорофор, належать система TaqMan і реагенти, доступні у фірм Roche Molecular Diagnostics і/або Applied Biosystems. У інших варіантах здійснення продукція і рівні першого і/або другого продуктів реплікації можуть відстежуватися в режимі реального часу. У деяких варіантах здійснення способи, описані в цьому документі, можуть бути використані для визначення зиготності нуклеїнових кислот (наприклад, генома(ів)) у визначеному сайті вставки. Результати аналізів при наявності першого реплікованого продукту (200) і відсутності другого реплікованого продукту (300) вказують на те, що нуклеїнові кислоти гомозиготні відносно наявності вставки. Результати аналізів з відсутністю першого реплікованого продукту 3 UA 112977 C2 5 10 15 20 25 30 35 40 45 50 55 (200) і наявністю другого реплікованого продукту (300) вказують на те, що нуклеїнові кислоти гомозиготні відносно відсутності вставки. Результати таких аналізів, в яких присутні і перший реплікований продукт (200) і другий реплікований продукт (300), вказують на те, що нуклеїнові кислоти гетерозиготні відносно вставки (наприклад, щонайменше один сайт вставки містить вставку і щонайменше один сайт вставки не містить вставку). У визначених варіантах здійснення набори праймерів і/або зондів можуть бути об'єднані з одним або декількома іншими наборами праймерів і зондів таким чином, щоб можливо було визначити наявність або відсутність однієї або декількох вставок в одному або декількох визначених сайтах вставки в нуклеїновій кислоті зразка. Використовуваний в цьому документі вираз "набір праймерів і/або зондів" включає щонайменше один прямий праймер, здатний зв'язуватися з сайтом вище визначеного сайту вставки, і щонайменше один зворотний праймер, здатний зв'язуватися з сайтом нижче визначеного сайту вставки або всередині визначеної вставки. У інших варіантах здійснення для ідентифікації конкретної вставки в одному або декількох визначених сайтах вставки і/або більшого числа вставок у більшому числі визначених сайтів вставки може бути використане більше число наборів праймерів і/або зондів. У деяких варіантах здійснення способи, описані в цьому документі, можуть бути використані для скринінгу популяції на наявність або відсутність вставки в визначеному сайті вставки. У додаткових варіантах здійснення наявність визначеного сайту вставки може бути визначена для кожного члена популяції. Під виразом "визначений сайт вставки", що використовується в цьому документі, розуміють відоме розташування або консервативну послідовність в нуклеїновій кислоті, куди вставка може бути вставлена відтворюваним чином. У деяких варіантах здійснення наявність конкретного сайту вставки може бути визначена для кожної нуклеїнової кислоти в зразку, як приклад, що не має обмежувального характеру, шляхом продукції першого (200) або другого (300) реплікованого продукту способами, описаними в цьому документі. У інших варіантах здійснення послідовності, фланкуючі конкретний сайт вставки, або послідовності всередині вставки можуть бути консервативними. У додаткових варіантах здійснення такий консерватизм послідовностей, фланкуючих визначений сайт вставки, або послідовностей у вставці може бути обмежений сайтами зв'язування праймерів і/або зондів. Під терміном "консервативний", що використовується в даному контексті, розуміють, що специфічний праймер і/або зонд здатний специфічно зв'язуватися з областю, яка є "консервативною". У певних варіантах здійснення специфічний праймер і/або зонд залишиться пов'язаним з "консервативною" областю при умовах підвищеної жорсткості. Використовувані в цьому документі терміни "вище" і "нижче" є відносними і означають протилежні сторони сайту вставки в нуклеїновій кислоті. Терміни не означають, який з напрямків розташовується "вище" і "нижче" сайту вставки, а лише те, що вони лежать на протилежних сторонах сайту вставки. Використовувані в цьому документі "прямий праймер" і "зворотний праймер" являють собою відносні терміни, що означають праймери, які зв'язуються з різними місцеположеннями на нуклеїновій кислоті таким чином, щоб дати можливість реплікації нуклеїнових кислот між ними способами, доступними в даній галузі, такими як, але нею не обмежуючись, ПЛР. Терміни "прямий" і "зворотний" не означають, де саме конкретний праймер зв'яжеться з сайтом послідовності нуклеїнової кислоти, а лише те, що вони лежать на протилежних сторонах реплікованої послідовності і можуть діяти при реплікації як праймери для полімерази. Використовувані в цьому документі "що містить", "що включає", "що відрізняється" і їх граматичні еквіваленти являють собою охоплюючі і необмежуючі терміни, які не виключають додаткових, неперерахованих елементів або стадій способу, але також включають вужчі терміни "що складається з" і "що по суті складається з". Даний винахід додатково описується в наступних прикладах, які пропонуються як ілюстрація і не призначені для того, щоб яким-небудь чином обмежити винахід. ПРИКЛАДИ ПРИКЛАД 1: Матеріал рослин Скринінг батьківських ліній: Для визначення, чи є крайова послідовність в сайті вставки трансгена високо консервативною і чи можуть трансформант-специфічні праймери бути використані для різного генетичного оточення, скринували в загальному 92 різні інбредні лінії, які представляли різні гетерозисні групи і мали різне походження, такі як північноамериканські, південноамериканські, європейські, stiff stalk, відмінні від stiff stalk, з публічних і особистих джерел (Таблиця 1). 4 UA 112977 C2 Таблиця 1 Перелік матеріалів, використаних для скринінгу крайової послідовності в сайті вставки трансгена Тип матеріалу Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Публічний Особистий Особистий Особистий Особистий Особистий Особистий Особистий Публічний Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Публічний Публічний Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Гетерозисна група Lancaster Flint Stiff stalk Змішана Lancaster Lancaster Stiff stalk Stiff stalk Stiff stalk Lodent Lodent Stiff stalk Stiff stalk Stiff stalk Lodent відмінна від Stiff stalk Lancaster Stiff stalk Stiff stalk Змішана Lodent Flint Lancaster Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Змішана Змішана Змішана Змішана Змішана Змішана Stiff stalk Stiff stalk Lodent Lodent Stiff stalk Stiff stalk Stiff stalk Stiff stalk Lodent Lodent Тропічна Тропічна Тропічна Тропічна Lancaster Lodent Stiff stalk відмінна від Stiff stalk Lodent Lancaster 5 Джерело Північна Америка Європа Північна Америка Південна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Південна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Південна Америка Південна Америка Південна Америка Північна Америка Південна Америка Південна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Південна Америка Південна Америка Південна Америка Південна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка UA 112977 C2 Таблиця 1 Перелік матеріалів, використаних для скринінгу крайової послідовності в сайті вставки трансгена Тип матеріалу Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Публічний Особистий Особистий Особистий Особистий Особистий Публічний Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий Особистий 5 10 Гетерозисна група Lancaster Lancaster Lancaster Lancaster Lancaster Lancaster Lancaster Stiff stalk Stiff stalk Lancaster Lodent Тропічна відмінна від Stiff stalk Stiff stalk відмінна від Stiff stalk відмінна від Stiff stalk Lancaster Lancaster Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Stiff stalk Змішана Suwan Тропічна Тропічна відмінна від Stiff stalk Змішана Змішана Змішана Тропічна Змішана Змішана Lodent Lodent Джерело Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Південна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Північна Америка Південна Америка Південна Америка Південна Америка Північна Америка Північна Америка Південна Америка Південна Америка Південна Америка Південна Америка Північна Америка Північна Америка Північна Америка Скринінг об'ємних проб насіння: Для демонстрації застосування тестування на основі ДНК на об'ємних пробах насіння і для визначення чутливості детекції пули насіння, вказані нижче, створювали, використовуючи відомі суворо гомозиготні DAS-59122, гемізиготні DAS-59122 і нульові (звичайні) насінини. Їх відлічували в шість різних пробірок типу фалькон об'ємом 50 мл: 1. 100 гомозиготних насінин DAS-59122, реплікація-1; 2. 100 гомозиготних насінин DAS-59122, реплікація-2; 3. 99 гомозиготних насінин DAS-59122 і одна гемізиготна насінина DAS-59122, реплікація-1; 4. 99 гомозиготних насінин DAS-59122 і одна гемізиготна насінина DAS-59122, реплікація-2; 5. 99 гомозиготних насінин DAS-59122 і одна нульова (звичайна) насінина, реплікація-1; 6. 99 гомозиготних насінин DAS-59122 і одна нульова (звичайна) насінина, реплікація-2. ПРИКЛАД 2: Обмолот насіння і екстракція ДНК 6 UA 112977 C2 5 10 15 20 25 30 35 40 45 50 55 Насіння дрібно подрібнювали і геномну ДНК виділяли, використовуючи набір DNEasy фірми Qiagen (Valencia, CA). З кожної партії насіння здійснювали п'ять окремих екстракцій геномної ДНК. Очищену геномну ДНК кількісно оцінювали, використовуючи набір з барвником ДНК Picogreen фірми QuantIt, і розводили до стандартизованих концентрацій. ПРИКЛАД 3: Аналізи зиготності на основі TaqMan Аналіз зиготності здійснювали, використовуючи різні набори реагентів, які складалися з відмінних послідовностей праймерів і зондів. Спосіб і реагенти створювали конкретно для трансформанта DAS-59122. Схема аналізу зиготності запропонована на фіг. 1. У способі крім двох зондів використовувалися ген-специфічний праймер, праймер дикого типу і генспецифічний/дикого типу (загальний) праймер. Зонди складалися з специфічного зонда дикого типу і трансгенного специфічного зонда. У першому способі всі праймери і зонди об'єднувалися в одній реакції ("спосіб в одну реакцію"). Для підвищення чутливості визначення зиготності також випробовували додатковий спосіб, в якому здійснювали дві окремі незалежні реакції (фіг. 3) ("спосіб з великим числом реакцій"). Один ряд ямок містив специфічний праймер дикого типу, загальний праймер і специфічний зонд дикого типу. Інший ряд ямок містив специфічний праймер трансгена, загальний праймер і специфічний зонд трансгена. Наприклад, в цьому способі використовували лише два праймера і один зонд в форматі 384-ямкового планшета, в якому один квадрат містив специфічний праймер дикого типу + загальний праймер + специфічний зонд дикого типу, інший квадрат містив специфічний праймер трансгена + загальний праймер + специфічний зонд трансгена. Модифікований аналіз Taqman з детекцією по кінцевій точці: Отримували реакційну суміш, що містила наступні компоненти: вода, 15,35 мкл; 10-кратний буфер для проведення ПЛР, 2,50 мкл; MgCl2 в концентрації 25 мМ, 1,50 мкл; dNTP в концентрації 10 мМ (по 2,5 мМ кожні), 2,0 мкл; загальний прямий праймер в концентрації 20 мкМ (SEQ ID NO:1), 0,25 мкл; зворотний праймер дикого типу в концентрації 20 мкМ (SEQ ID NO:2), 0,25 мкл; двічі мічений зонд дикого типу (SEQ ID NO:3) в концентрації 10 мкМ, на 3' кінці мічений за допомогою VIC і на 5' кінці мічений за допомогою BHQ2, 0,20 мкл; Taq-полімераза HotStar (5 Од/мкл), 0,20 мкл; геномна ДНК в концентрації 10 нг/мкл, 3,0 мкл. Отримували другу реакційну суміш, що містила наступні компоненти: вода, 15,35 мкл; 10кратний буфер для проведення ПЛР, 2,50 мкл; MgCl2 в концентрації 25 мМ, 1,50 мкл; dNTP в концентрації 10 мМ (по 2,5 мМ кожні), 2,0 мкл; загальний прямий праймер в концентрації 20 мкМ (SEQ ID NO:1), 0,25 мкл; зворотний праймер 591227 в концентрації 20 мкМ (SEQ ID NO:4), 0,25 мкл; двічі мічений зонд 591227 (SEQ ID NO:5) в концентрації 10 мкМ, мічений на 3' кінці за допомогою FAM і на 5' кінці за допомогою BHQ1, 0,20 мкл; Taq-полімераза HotStar (5 Од/мкл), 0,20 мкл; геномна ДНК в концентрації 10 нг/мкл, 3,0 мкл. Обидві суміші розкопували в окрему ямку і ампліфікували, використовуючи систему для проведення ПЛР GenAmp 9700 при наступних умовах: 95 °C протягом 15 хвилин (1 цикл); 95 °C протягом 15 секунд, 60 °C протягом 60 секунд (35 циклів). Зчитування флуоресцентного сигналу аналізували і зиготність визначали, виходячи із збудження флуорофору VIC або FAM. Результати: Після визначення консервативності крайових послідовностей в сайті зв'язування праймерів праймери створювали і випробовували на об'ємних пулах насіння, використовуючи стандартний аналіз визначення зиготності Taqman. Результати указали на те, що спосіб з однією реакцією виявився чутливим при визначенні домішки нульової насінини серед суворо гомозиготних насінин, взятих у великій кількості, з чутливістю детекції 1 %. Проте, спосіб з однією реакцією виявився незадовільним у визначенні домішки якого-небудь гемізиготного насіння при рівні наявності 1 %. Низька чутливість могла бути наслідком переважної ампліфікації реакції-1 (див. фіг. 1) через надмірну доступність матриці. Для розв'язання цієї проблеми конкуренції за ресурси в процесі реакції ПЛР спосіб з однією реакцією модифікували у велике число окремих реакцій (фіг. 3). Ця модифікація привела до вибіркової ампліфікації лише націленого регіону за відсутності якої-небудь конкуренції за ресурси. Застосування способу з великим числом реакцій (тестування на зиготність спорідненого насіння) на батьківських "пулах насіння" показало достовірність визначення домішки як гемізиготних, так і звичайного (нульових) насіння у великій кількості суворо гомозиготних насінин з чутливістю детекції 1 % (Таблиця 2). Результати також підтверджували за допомогою ПЛР в режимі реального часу, використовуючи прилад Light Cycler 480 фірми Roche, для забезпечення відсутності артефактів в реакції ПЛР або встановлення їх. 7 UA 112977 C2 Таблиця 2 Чутливість визначення зиготності різними способами Спосіб Спосіб з однією реакцією Спосіб з великим числом реакцій 5 10 15 20 25 30 35 Достовірність визначення при 1 % домішці гемізиготної насінини Достовірність визначення при контамінації 0 % Достовірність визначення при 1 % домішці нульової насінини Так Так Недостовірно Так Так Так Скринінг батьківських ліній: Оскільки трансформант-специфічні праймери можуть бути використані для різного генетичного оточення, для випробування способу з великим числом реакцій використовували 92 відмінні інбредні лінії (Таблиця 1), які представляють різні гетерозисні групи, що виростають в місцевостях, таких як Північна Америка, Південна Америка і Європа. Ці лінії випробовували і підтверджували, що вони вільні від контамінації трансгеном, використовуючи протокол, описаний вище. Аналіз, використовуючи термоциклер в режимі реального часу Roche 480 (фіг. 4 і 5), а також аналізи на основі TaqMan по кінцевій точці показали, що всі протестовані інбредні лінії мають консервативну крайову послідовність, фланкуючу вставку трансгена. Їх успішно ампліфікували з участю специфічного праймера/зонд WT, підтверджуючи, що праймери і зонди для аналізу трансформанта DAS-59122 можуть бути використані для програм інтрогресії при випробуванні зиготності батьківського насіння на остаточних лініях. До деяких з переваг способу з великим числом реакцій належать всі переваги простоти і надійності тестування ДНК над тестом ELISA. Найважливіше, це дозволяє тестувати зиготність, використовуючи об'ємні пули насіння, на відміну від тестування ELISA, яке може ідентифікувати лише окремі рослини. Крім того, цей спосіб може знизити вартість процедури tester-row і ELISA в десять разів. Цей спосіб також може підвищити чутливість аналізу і буде ідентифікувати і інші домішки. Спосіб з великим числом реакцій також може бути використаний як "індикатор" при випробуванні на наявність малих домішок (АР), що може бути використано для тестування на присутність непередбаченого трансформанта, а також повинно показати статус суворої гомозиготності об'ємної проби. Тестування способом з великим числом реакцій довело високу чутливість у визначенні наявності домішки якого-небудь гемізиготного або нульового насіння в партії насіння суворо гомозиготних остаточних ліній. Було показано, що спосіб з великим числом реакцій може ідентифікувати домішку при рівні контамінації 1 % (1 в 100 насінинах). Ця нова методологія привела до встановлення нового напрямку застосування високопродуктивного молекулярного аналізу (НТМА), що краще тестує ступінь чистоти у остаточних ліній, а також може забезпечити десятиразову економію при проведенні процедур в польових умовах. Незважаючи на те, що даний винахід був описаний в декількох варіантах здійснення, даний винахід може бути додатково модифікований в межах суті і об'єму даного опису. Ця заявка, тому, охоплює будь-які варіанти, застосування або адаптацію винаходу, що використовують його основні принципи. Крім того, дана заявка охоплює такі відхилення від даного опису, які узгоджуються з відомою або звичайною практикою в даній галузі, до якої даний винахід належить і яка знаходиться в рамках прикладеної формули винаходу. 8 UA 112977 C2 9 UA 112977 C2 ФОРМУЛА ВИНАХОДУ 5 10 15 20 25 30 35 40 45 50 55 1. Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в об'ємному зразку нуклеїнової кислоти зі щонайменше 100 організмів, причому спосіб містить: приведення зразка нуклеїнової кислоти в контакт з: прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайта вставки; і першим зворотним праймером, здатним зв'язуватися зі вставленою нуклеотидною послідовністю, і другим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайта вставки; застосування праймерів для ампліфікації нуклеїнових кислот між праймерами; і аналіз ампліфікованих нуклеїнових кислот для визначення, присутності або відсутності вставленої нуклеотидної послідовності в зразку нуклеїнової кислоти, де репродуковані нуклеїнові кислоти включають послідовність, яка відповідає послідовності між прямим праймером і першим зворотним праймером в разі присутності вставленої нуклеотидної послідовності, і де репродуковані нуклеїнові кислоти включають послідовність, яка відповідає послідовності між прямим праймером і другим зворотним праймером в разі відсутності вставленої нуклеотидної послідовності. 2. Спосіб за п. 1, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого при наявності вставленої нуклеотидної послідовності. 3. Спосіб за п. 1, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого за відсутності вставленої нуклеотидної послідовності. 4. Спосіб за п. 2, який додатково містить приведення ампліфікованих нуклеїнових кислот в контакт під час або після амплікації із зондом, специфічним для фрагмента, ампліфікованого за відсутності вставленої нуклеотидної послідовності. 5. Спосіб за п. 1, в якому нуклеїнові кислоти зразка містять більше ніж один повтор визначеного сайта вставки. 6. Спосіб за п. 1, в якому вставлена нуклеотидна послідовність присутня менше ніж в 1 % конкретних сайтів вставки в нуклеїновій кислоті. 7. Спосіб за п. 1, в якому вставлена нуклеотидна послідовність присутня більше ніж в 99 % конкретних сайтів вставки в нуклеїновій кислоті. 8. Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності у визначеному сайті вставки, в об'ємному зразку нуклеїнової кислоти з множини щонайменше 100 організмів, причому спосіб включає: розділення зразка нуклеїнової кислоти на щонайменше дві частини; здійснення першого аналізу на основі реакції ампліфікації, що включає приведення першої частини зразка нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїнової кислотою вище сайта вставки, і першим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою в межі вставленої нуклеотидної послідовності; застосування прямого праймера і першого зворотного праймера для отримання першого продукту ампліфікації, і визначення кількості першого продукту ампліфікації; здійснення другого аналізу на основі реакції ампліфікації, що включає приведення другої частини зразка нуклеїнової кислоти в контакт з прямим праймером, здатним зв'язуватися з нуклеїновою кислотою вище сайта вставки; і другим зворотним праймером, здатним зв'язуватися з нуклеїновою кислотою нижче сайта вставки; застосування прямого праймера і другого зворотного праймера для одержання другого продукту ампліфікації, і визначення кількості другого продукту ампліфікації; і порівняння кількості першого продукту ампліфікації з кількістю другого продукту ампліфікації в зразку нуклеїнової кислоти, де кількість першого продукту ампліфікації вказує на кількість вставленої нуклеотидної послідовності, що присутня в зразку нуклеїнової кислоти. 9. Спосіб за п. 8, де здійснення першої реакції ампліфікації додатково містить приведення першої частини зразка нуклеїнової кислоти в контакт під час або після ампліфікації із зондом, специфічним для першого продукту ампліфікації. 10 UA 112977 C2 5 10 10. Спосіб за п. 8, де здійснення другої реакції ампліфікації додатково містить приведення другої частини зразка нуклеїнової кислоти в контакт під час або після ампліфікації із зондом, специфічним для другого продукту ампліфікації. 11. Спосіб за п. 8, в якому нуклеотидні кислоти зразка містять більше ніж один повтор визначеного сайта вставки. 12. Спосіб за п. 8, в якому вставлена нуклеотидна послідовність присутня менше ніж в 1 % конкретних сайтів вставки в зразку нуклеїнової кислоти. 13. Спосіб за п. 8, в якому вставлена нуклеотидна послідовність присутня більше ніж в 99 % конкретних сайтів в зразку нуклеїнової кислоти. 14. Спосіб за п. 11, в якому спосіб використовують для визначення зиготності. 15. Спосіб за п. 11, в якому спосіб використовують для визначення домішки події вставки в зразку. 16. Спосіб за п. 11, в якому спосіб використовують як індикатор наявності/відсутності в об'ємному зразку додаткової події вставки. 11 UA 112977 C2 12 UA 112977 C2 Комп’ютерна верстка Т. Вахричева Державна служба інтелектуальної власності України, вул. Василя Липківського, 45, м. Київ, МСП, 03680, Україна ДП "Український інститут інтелектуальної власності", вул. Глазунова, 1, м. Київ – 42, 01601 13
ДивитисяДодаткова інформація
Назва патенту англійськоюMethods to determine zygosity in a bulked sample
Автори англійськоюChannabasavaradhya, Chandra-Shekara
Автори російськоюЧаннабасаварадхя Чандра-Шекара
МПК / Мітки
МПК: C12N 15/09, C12Q 1/68
Мітки: щонайменше, нуклеїнової, спосіб, послідовності, визначеному, наявності, вставленої, організмів, сайті, нуклеотидної, відсутності, зразку, вставки, об'ємному, визначення, кислоти
Код посилання
<a href="https://ua.patents.su/15-112977-sposib-viznachennya-nayavnosti-abo-vidsutnosti-vstavleno-nukleotidno-poslidovnosti-v-viznachenomu-sajjti-vstavki-v-obehmnomu-zrazku-nuklenovo-kisloti-zi-shhonajjmenshe-100-organizm.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення наявності або відсутності вставленої нуклеотидної послідовності в визначеному сайті вставки в об’ємному зразку нуклеїнової кислоти зі щонайменше 100 організмів</a>
Спосіб визначення послідовності молекули нуклеїнової кислоти (варіанти) та набір для використання при визначенні послідовності нуклеїнової кислоти

Номер патенту: 48119
Опубліковано: 15.08.2002
Автор: Дрманач Радоє
МПК: C12Q 1/68, G01N 37/00, G01N 33/53, C12N 15/09
Мітки: послідовності, нуклеїнової, визначення, визначенні, молекули, варіанти, спосіб, кислоти, використання, набір
Формула / Реферат:
1. Способ определения последовательности молекулы нуклеиновой кислоты, отличающийся тем, что проводят гибридизацию молекулы с комплементарными последовательностями олигонуклеотидов из двух наборов малых олигонуклеотидных зондов известной последовательности, причем первый набор зондов прикреплен к твердой подложке, а второй набор зондов представляет собой меченые зонды, находящиеся в растворе, создают ковалентные связи между гибридизированным...
Олігонуклеотид для ампліфікації послідовностей нуклеїнових кислот, спосіб ампліфікації цільової послідовності нуклеїнової кислоти, набір для здійснення реакції ампліфікації нуклеїнової кислоти

Номер патенту: 49843
Опубліковано: 15.10.2002
Автор: Уілл Стівен Гордон
МПК: C07H 21/00, C12N 15/09, C12Q 1/68
Мітки: нуклеїнової, олігонуклеотид, набір, реакції, здійснення, ампліфікації, нуклеїнових, кислот, послідовностей, послідовності, спосіб, кислоти, цільової
Формула / Реферат:
1. Олігонуклеотид для ампліфікації послідовностей нуклеїнових кислот, який має загальну структуру:або,де: S1 - перша послідовність нуклеотидів довжиною, приблизно, від 5 до 50 нуклеотидів;S2 - друга послідовність довжиною від одного до трьох нуклеотидів;Nu - нуклеотид, до складу якого входить пуринова або піримідинова основа, яка включає в себе екзоциклічний амін;R - модифікаторна...
Рекомбінантна термостабільна днк-полімераза, фрагмент нуклеїнової кислоти, композиція для застосування в реакції секвенування днк, спосіб секвенування нуклеїнової кислоти-мішені, набір для секвенування нуклеїно

Номер патенту: 47423
Опубліковано: 15.07.2002
Автори: Рейчерт Фред Лоуренс, Кальман Ліса Вівіан, Гельфанд Девід Харроу
МПК: C12N 9/12, C12Q 1/68, C12N 15/09, C12N 15/54, C12R 1/19
Мітки: кислоти-мішені, застосування, рекомбінантна, композиція, нуклеїно, днк-полімераза, секвенування, спосіб, набір, нуклеїнової, термостабільна, реакції, фрагмент, кислоти, днк
Формула / Реферат:
1. Рекомбінантна термостабільна ДНК-полімераза, яка являє собою мутовану форму нативної термостабільної ДНК-полімерази, що має амінокислотну послідовність, яка містить амінокислотний фрагмент Ser Gln Ile Glu Leu Arg Xaa, в якому Xaa в положенні 7 вказаного фрагменту є залишком валіну Val або ізолейцину Ilе, причому вказана мутована форма модифікована таким чином, що вона містить відмінну від Glu амінокислоту в положенні 4 послідовності...
Спосіб визначення наявності небезпечних паразитичних організмів у водоймі

Номер патенту: 83037
Опубліковано: 27.08.2013
Автор: Юришинець Володимир Іванович
МПК: G01N 33/18
Мітки: організмів, паразитичних, небезпечних, спосіб, водойми, визначення, наявності
Формула / Реферат:
Спосіб визначення наявності небезпечних паразитичних організмів у водоймі, який включає виявлення у водоймах збудників небезпечних паразитарних хвороб людини і тварин шляхом паразитологічного дослідження, який відрізняється тим, що дослідження проводять по декількох основних блоках, які включають пошук цист патогенних найпростіших та яєць і личинок гельмінтів у воді, дослідження молюсків на наявність партеногенетичних поколінь та церкарій...
Невірусний вектор для введення нуклеїнової кислоти та спосіб його одержання, конденсований комплекс поліпептид/нуклеїнова кислота та спосіб вбудовування нуклеїнової кислоти, що представляє інтерес, в еукаріотич

Номер патенту: 77393
Опубліковано: 15.12.2006
Автори: Алтон Ерік, Менвелл Мішель, Міллєр Ендрю Девід, Меттьюз Девід, Руссель Віллі, Пєрузель Ерік, Мюррей Карл, Тагава Тошіакі
МПК: A61P 43/00, C12N 15/34, A61P 25/00, A61P 35/00, A61K 38/00, A61K 9/127, C12N 15/88, A61K 47/48, A61K 48/00, C07K 14/075, A61P 35/02, C12N 15/09, A61P 25/28
Мітки: введення, представляє, конденсований, комплекс, кислоти, вбудовування, нуклеїнової, інтерес, еукаріотич, невірусний, кислота, спосіб, одержання, вектор
Формула / Реферат:
1. Невірусний вектор для введення нуклеїнової кислоти, який містить конденсований комплекс поліпептид/нуклеїнова кислота і катіонний ліпід, де комплекс включає(а) нуклеотидну послідовність (NOI), що представляє інтерес; і(б) один або декілька аденовірусних поліпептидів або їх похідних, які упаковують нуклеїнову кислоту, де поліпептиди або їх похідні (І) мають здатність зв'язуватися з NOI; і (II) мають здатність конденсувати NOI;...
Попередній патент: З’єднувач для забезпечення з’єднання одного з одним двох електричних кабелів
Випадковий патент: Пристрій для виявлення магнітних міток на сталевих канатах