Спосіб трансформації рослин і лінія клітин ембріогенного калюсу zea mays






Формула / Реферат
1. Спосіб трансформації рослинної клітини або тканини геном, що представляє інтерес, який включає обробку рослинної клітини або тканини Agrobacterium в умовах, які інгібують індукований Agrobacterium некроз (АІН), причому Agrobacterium включає вектор, який містить ген, що представляє інтерес, де умови, які інгібують АІН, включають: (а) контактування рослинної клітини або тканини з Agrobacterium після обробки температурним шоком; або (б) контактування рослинної клітини або тканини з Agrobacterium у присутності агента, який інгібує АІН, де цей агент включає (і) хімічний інгібітор, де хімічний інгібітор являє собою сполуку, що вибрана з групи, яка включає інгібітори етилену, за виключенням нітрату срібла, інгібітори синтезу етилену, антагоністи гібереліну і інгібітори фосфатази; або (іі) нуклеотидну послідовність, яка кодує мРНК або протеїн, що інгібує АІН.
2. Спосіб за п. 1, де умови, при яких інгібується АІН, включають контактування рослинної клітини з Agrobacterium після обробки рослинної клітини або тканини температурним шоком.
3. Спосіб за п. 2, де обробка температурним шоком триває 2-10 хв.
4. Спосіб за п. 3, де обробка температурним шоком триває 4-8 хв.
5. Спосіб за п. 2, де обробку температурним шоком здійснюють при температурі 40-50°С.
6. Спосіб за п. 5, де обробку температурним шоком здійснюють при температурі 42-48°С.
7. Спосіб за п. 1, де умови, при яких інгібується АІН, включають контактування рослинної клітини або тканини з Agrobacterium у присутності хімічного інгібітора, де хімічний інгібітор являє собою сполуку, що вибрана з групи, яка включає інгібітори етилену, за виключенням нітрату срібла, інгібітори синтезу етилену, антагоністи гібереліну і інгібітори фосфатази.
8. Спосіб за п. 1, де умови, при яких інгібується АІН, включають контактування рослинної клітини або тканини з Agrobacterium у присутності нуклеотидної послідовності, яка кодує мРНК або протеїн, що інгібує АІН.
9. Спосіб за п. 8, де нуклеотидна послідовність доставляється або експресується в рослинній клітині або тканині.
10. Спосіб за п. 8, де нуклеотидна послідовність включає кодуючу послідовність, яка вибрана з групи, що включає ген р35, ген іар, ген dad-1, ген bcl та ген, який здатний пригнічувати хворобливу реакцію у рослин.
11. Спосіб за п. 10, де ген, який здатний пригнічувати хворобливу реакцію у рослин, є nahG або mlо.
12. Спосіб за п. 8, де нуклеотидна послідовність включає кодуючу послідовність, яка вибрана з групи, що включає ген р35, ген іар та ген dad-1.
13. Спосіб за п. 1, де рослинна клітина або тканина являє собою рослинну клітину або тканину злакових.
14. Спосіб за п. 13, де клітина рослини або тканина є клітиною або тканиною кукурудзи.
15. Спосіб за п. 13, де клітина рослини або тканина є клітиною або тканиною пшениці.
16. Спосіб за п. 1, де рослинна тканина є ембріогенним калюсом.
17. Спосіб за п. 1, де рослинна клітина є незрілим ембріоном.
18. Спосіб за п. 1, який додатково включає одержання регенерату зрілої трансгенної рослини з трансформованих рослинних клітин або тканин.
19. Спосіб за п. 18, де рослинна клітина або тканина являє собою рослинну клітину або тканину злакових.
20. Спосіб за п. 19, де клітина рослини або тканина є клітиною або тканиною кукурудзи.
21. Спосіб за п. 19, де клітина рослини або тканина є клітиною або тканиною пшениці.
22. Спосіб за п. 18, де рослинна тканина є ембріогенним калюсом.
23. Спосіб за п. 18, де рослинна клітина є незрілим ембріоном.
24. Лінія клітин НЕ/89 ембріогенного калюсу, Zea mays L., що включають кодуючу послідовність гена р35.
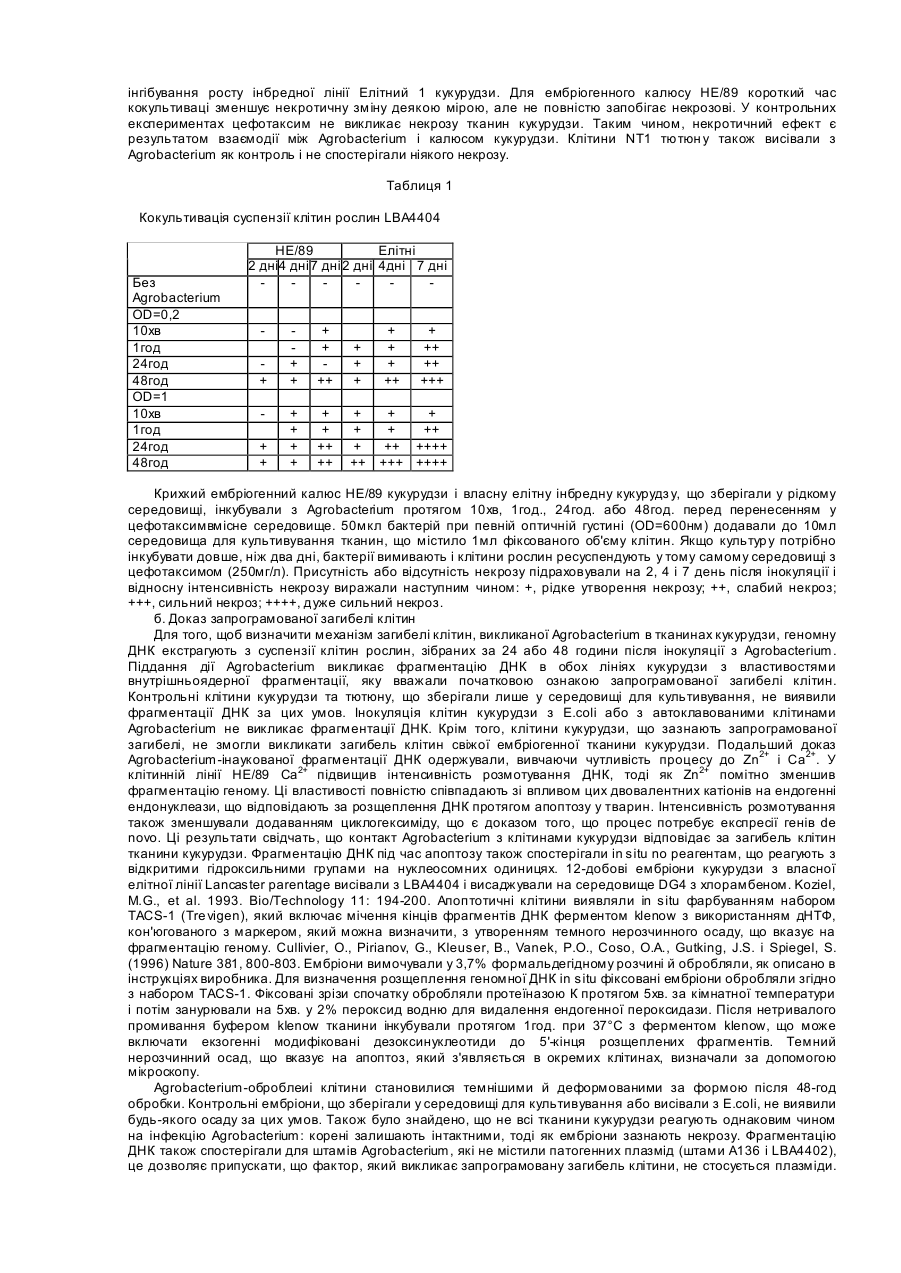
Текст
Цей винахід стосується способу трансформації рослин з використанням Agrobacterium здійсненням трансформації в присутності агента, який інгібує Agrobacterium-індукований некроз, і/або з використанням штаму Agrobacterium, який не викликає некрозу. Перший спосіб і один з найбільш широко використовуваних способів уведення чужорідного генетичного матеріалу в рослини оснований на природній трансформаційній системі Agrobacterium spp. з використанням рекомбінантних Ті (що викликає пухлини) або Ri (що викликає ріст коренів) плазмід, де Т-ДНК містить потрібний ген. Потрібний ген включають у генетичний матеріал рослини за таким самим механізмом, що використовується в природі для включення онкогенів або генів біосинтезу опінів. Трансформація Agrobacterium придатна для рослин, які інфікуються й трансформуються Agrobacterium в природі з утворенням пухлин і/або волосатих коренів, але не придатна для інших рослин. Наприклад, невідомо, чи члени родини трав'янистих рослин (Gramineae), такі як кукурудза, утворюють пухлини під впливом Agrobactehum і є зовсім непридатними для трансформації Agrobacterium. Ishida, et al. Nature Biotechnology (1996) 14: 745-750, здійснив трансформацію ембріонів певної лінії кукурудзи (А188) та її гібридів з використанням Agrobacterium, що несли "супер-бінарні" вектори, але повторюваність цих результатів і застосовуваність цієї системи до інших ліній кукурудзи і/або з використанням Agrobacterium, що несли інші вектори, є нез'ясованим. Також було повідомлено про трансформацію Agrobacterium кукурудзи з використанням апікальної меристеми як тканини-мішені, але низька продуктивність апікальної меристеми як тканини-мішені порівняно з, наприклад, ембріогенним калюсом, робить цей підхід гіршим за оптимальний. Навіть деякі дводольні рослини, такі як виноград, соя або перець, які є чутливими до інфекції Agrobacterium і утворення пухлин в природі, незважаючи на це, важко піддаються трансформації за лабораторних умов, тому що кращі для трансформації та регенерації тканини-мішені виявили слабку відповідь на вплив Agrobacterium. Недостатнє розуміння механізмів опору інфекції Agrobacterium і утворення пухлин стало перешкодою для вирішення проблеми або навіть належного оцінювання проблеми трансформації Agrobacterium у рослин, що не піддаються трансформації Agrobacterium, таких як Gramineae. Було виявлено, що Agrobacterium здатний викликати апоптотичний некроз в клітинах рослин, зокрема родини Graminieae, наприклад, кукурудзи. Були досліджені способи селекції і створення Agrobacterium, які меншою мірою здатні викликати некроз, також як і способи інгібування некрозу в клітинах, що піддаються трансформації. Некроз, який спостерігається в культурі клітин Gramineae при впливі Agrobacterium, - це запрограмована загибель клітин, за допомогою якої клітина, очевидно, через механізм передачі сигналу, керує своєю власною загибеллю. Це видно з пасивної загибелі клітини, тобто некрозу, що складається з порушення цілісності мембрани й наступного розбухання, що призводить до лізису, наприклад, під дією різних токсинів. Клітини, що зазнають запрограмованої загибелі, виявляють характерні морфологічні зміни і фрагментацію ДНК, які разом визначають процес апоптозу і здійснюються за механізмом, що включає експресію гена de novo. Стереотипні властивості апоптозу, описані для клітин тварин, включають зменшення фізичних розмірів, втрату міжклітинного контакту в організованій тканині, конденсацію ядра й хроматину, фрагментацію ДНК (ДНК "розмотування"), пухирчастість ядра й мембрани. Автори показали, що кокультивація Agrobacterium з тканинами кукурудзи або пшениці приводить до процесу, дуже аналогічному до апоптозу клітин тварин, де загибель клітин характеризується розщепленням ДНК на олігонуклеосомальні фрагменти й визначає морфологічні зміни. У клітин кукурудзи, які піддавали дії Agrobacterium, Ca2+ підвищував інтенсивність фрагментації ДНК, але Zn2+ мав протилежний ефект, який був аналогічний ефекту цих двовалентних катіонів на ендогенні ендонуклеази, що відповідають за розщеплення ДНК під час апоптозу клітин тварин. Ця фрагментація також зменшується додаванням циклогексиміду, що є доказом того, що процес потребує експресії гена de novo. Запрограмована загибель клітини у відповідь на дію Agrobacterium ще не була виявлена. Також було виявлено, що цей Agrobacterium-індукований некроз (АІН), що спостерігали у Gramineae, може бути інгібований використанням АІН-інгібуючих агентів або хімічних сполук, таких як нітрат срібла, або АІНінгібуючих нуклеотидних послідовностей, стабільно інтегрованих або тимчасово функціональних в клітинах, що підлягають трансформації. Також було виявлено за допомогою процедури скринінгу Agrobacterium, що Agrobacterium штами відрізняються за їх здатністю індукувати некроз, так що можуть бути вибрані штами, які є придатними для трансформації непридатних рослин, таких як Gramineae. Таким чином, декілька втілень винаходу включають наступне: 1. Спосіб трансформації рослинної клітини потрібним геном, що полягає у піддаванні вищезгаданої клітини рослини дії Agrobacterium за умов, які інгібують АІН, наприклад, присутність АІН-інгібуючого агента або застосування теплового шоку, де вищезгадана Agrobactenum містить одну або кілька плазмід (векторів), що містять один або кілька потрібних генів. 1.1. В одному втіленні вищезгаданого способу АІН-інгібуючим агентом є хімічний інгібітор. Краще, якщо хімічним інгібітором є сполука, вибрана з групи, що складається з етиленових інгібіторів (наприклад, 2,5норборнадієну, норборнену, тіосульфату срібла і нітрату срібла), інгібіторів синтезу етилену (наприклад, аміноетоксивінілгліцину (АВГ), солей кобальту, ацетилсаліцилової кислоти або саліцилової кислоти), антагоністів гібереліну (наприклад, абсцизова кислота (АК)) і інгібіторів фосфа тази (наприклад, окадова кислота). Найкраще, якщо хімічним інгібітором є етиленовий інгібітор, краще, якщо нітрат срібла. Протеїни та пептиди можуть діяти також як і хімічні інгібітори. Прикладами протеїнів, які зустрічаються в природі, є DAD-1, бакуловірусні інгібітори апоптозу (БІА), бакуловірус р35 або синтетичні пептидні аналоги каспаз, здатних давати початок апоптозові. Краще, якщо хімічний інгібітор присутній в ефективній концентрації, наприклад, для нітрату срібла в концентрації від 0,1 до 20мг/л, краще 1-10мг/л. 1.2.1. В альтернативному втіленні цього способу АІН-інгібуючим агентом є нуклеотидна послідовність. АІНінгібуюча нуклеотидна послідовність може інгібувати АІН безпосередньо або кодуванням АІН-інгібуючих мРНК, що кодують АІН-інгібуючий білок. Наприклад, це може бути антизмістовний олігонуклеотид або ген, що кодує антизмістовну мРНК, яка є антизмістовною генові, що кодує фермент, який бере участь у некрозі (наприклад, протеаза, кіназа або фосфатаза), або регуляторний білок. В альтернативному варіанті він може містити кодуючу ділянку гена, здатного інгібувати апоптоз під контролем промотору, здатного експресуватися в рослинах, наприклад, кодуюча ділянка гена bcl-1 ссавців під контролем промотору, здатного експресуватися в рослинах, кодуюча ділянка гена, інгібуючого апоптоз з бакуловірусу, такого як р35 або рІАР або гена, здатного пригнічувати відповідь на хворобу в рослинах, наприклад, nahG, dad-1 або mlo. АІН-інгібуюча нуклеотидна послідовність, що експресує АІН-інгібуючий білок, може, за бажанням, бути пристосована для експресії в рослині-хазяїні створенням синтетичної нуклеотидної послідовності, що кодує той самий білок, але використовуючи кодони, які є кращими для рослини-хазяїна, і уникаючи нуклеотидних послідовностей, наприклад, сигналів проліаденілювання або сайтів сплайсингу всередині кодуючої ділянки, які можуть впливати на оптимальну експресію в рослині-хазяїні, наприклад, аналогічно способам, описаним у патентах США №5380831 або 5610042. 1.2.2. АІН-інгібуюча н уклеотидна послідовність може бути стабільно включена в геном рослини, що підлягає трансформації, або може бути лише короткочасно присутньою й функціональною, наприклад, під час або вже безпосередньо перед тим, як клітину піддають дії Agrobacterium. Короткочасну експресію можна одержати, наприклад, використовуючи агроінфекційну систему, де Т-ДНК несе два однакові віруси у тандемі так, що вірусний реплікон, який несе АІН-інгібуючу нуклеотидну послідовність, може реплікуватися в клітині. У цій системі вірус типово не інтегрується в рослинний геном, але реплікуватиметься великою кількістю і створюватиме високій рівень короткочасної експресії. Клітини, створені таким чином, щоб набути резистентності до некрозу, можуть тоді бути трансформовані з використанням Agrobacterium, що має Тіплазміду, що містить потрібний ген, який буде включений у геном, тоді як вірус буде зникати через регенерацію і не передаватиметься насінню. Таким чином, нащадки інфікованої рослини стабільно трансформовані потрібним геном, але не АІН-інгібуючою нуклеотидною послідовністю. Короткочасна експресія може бути одержана, як альтернатива, уведенням короткої АІН-інгібуючої олігонуклеотидної послідовності в рослину, наприклад, антизмістовної послідовності. 1.3. У подальшому альтернативному втіленні АІН зменшений або інгібований тепловою обробкою тканини рослини, що підлягає трансформації, перед кокультивацією Agrobacterium. Теплову обробку виконували при 40-50°С, краще при 42-48°С, 45°С - це краща температура для кукурудзи. Обробка триває протягом 2-10 хвилин, краще 4-8 хвилин. 2. Спосіб створення фертильної, трансгенної рослини, що складається з трансформування тканини рослини вищезгаданим способом 1 і регенерації таким чином трансформованої тканини. Краще, якщо тканина вибрана з незрілих ембріонів або ембріогенного калюсу, наприклад, калюс кукурудзи типу І. Там, де тканиною є незрілий ембріон, спосіб складається з перенесення ембріонів на середовище ініціювання калюсу перед або після піддавання дії Agrobacterium і регенерації рослини з таким чином одержаного ембріогенного калюсу. Краще, якщо потрібний ген (або один з них) є селектабельним або здатним до виявлення маркером, що дозволяє селекцію або ідентифікацію трансформованої клітини, трансформованої тканини і/або регенерованої трансформованої рослини. 3. Використання АІН-інгібуючого агента, наприклад, хімічного інгібітору АІН (наприклад, етиленового інгібітору, або, наприклад, нітрату срібла) або АІН-інгібуючої нуклеотидної послідовності (наприклад, послідовності, що кодує АІН-інгібуючу мРНК або білок) у способі трансформації Agrobactenum рослини або тканини або їх клітини, або способі створення фертильної трансгенної рослини, наприклад, способі згідно з 1 або 2, згаданими вище. 4. Трансгенна рослина, одержана вищезгаданим способом 2, або насіння, або їх нащадки. 5. Рослина, тканина рослини або клітина рослини, що містить нуклеотидну послідовність гетерологічного походження, яка інгібує АІН, наприклад, синтетична нуклеотидна послідовність або нуклеотидна послідовність з геному іншого виду організму. 6. Культура рослинних клітин або тканин, наприклад, для використання при трансформації Gramineae, що містить а) хімічний інгібітор АІН, б) Agrobactenum, що містить плазміду, яка містить потрібний ген, і в) воду й необхідні солі. Краще, якщо хімічний інгібітор присутній у некроз-інгібуючій концентрації. Краще, якщо хімічним інгібітором є сполука, вибрана з групи, що складається з етиленових інгібіторів (наприклад, 2,5-норборнадієн, норборнен, тіосульфат срібла і нітрат срібла), інгібіторів синтезу етилену (наприклад, аміноетоксивінілгліцин (АВГ), солей кобальту, ацетилсаліцилової кислоти або саліцилової кислоти), антагоністів гібереліну (наприклад, абсцизової кислоти (АК)) і інгібіторів фосфатаз (наприклад, окадової кислота). Найкраще, якщо хімічним інгібітором є нітрат срібла, наприклад, в концентрації 0,1-20мг/л, краще 1-10мг/л. Оптимальний склад і концентрація необхідних солей відомі у галузі й можуть дещо відрізнятися залежно від виду, тип у і стадії розвитку клітини або тканини, але загалом є такими, що концентрації іонів і поживних речовин у середовищі (наприклад, концентрації нітратів, іонів калію, фосфатів, іонів кальцію, іонів магнію, натрію іонів і іонів хлору) підтримуються в кількості, яка добре переноситься клітинами рослин і Agrobactenum. Середовище для культивування може за бажанням містити придатні поживні речовини (наприклад, цукри, такий як сахароза), вітаміни, амінокислоти, гормони та інші компоненти (наприклад, 2,4-D), як відомо у галузі культивування клітин рослин або тканин. Краще, якщо рослинами є рослини з родини Gramineae, наприклад, кукурудзи. Краще, якщо клітини або тканини містять ембріогенний калюс, наприклад, Типу І або Типу II. 7. Спосіб трансформації тотипотентної клітини рослини з родини Gramineae, що складається з піддання популяції тотипотентних клітин або тканин, що містять тотипотентні клітини, Agrobacterium, що містить одну або кілька плазмід, які містять один або кілька потрібних генів, де Agrobacterium є зі штаму, який не викликає значного рівня некрозу у ви щезгаданій популяції при тривалості дії та концентрації, достатній для досягнення трансформації вищезгаданої клітини. Краще, якщо тканинами є тканини, інші, ніж апікальна меристема, наприклад, краще, якщо недиференційовані тканини, наприклад, висаджені зиготичні ембріони або ембріогенний калюс, краще, якщо ембріогенний калюс під час фази ініціації (наприклад, до 28 днів, краще, якщо до 14 днів від висадження на середовище для ініціювання) або серійно пророщуваний ембріогений калюс, наприклад, калюс Типу І або Типу II. 8. Спосіб визначення придатності штаму Agrobacterium для використання при трансформації здатних до регенерації клітин рослини з родини Gramineae, що складається з піддання вищезгаданої популяції здатних до регенерації клітин рослини штамові Agrobacterium і спостерігання або визначення кількості некрозу у вищезгаданій популяції клітин. 9. Штам Agrobacterium, який був генетично модифікований для зменшення або видалення експресії фактора некрозу з Agrobacterium або похідної з такого модифікованого штаму. Фактор некрозу з Agrobacterium - це термолабільний фактор, що знаходиться у концентрованому супернатанті, здатному викликати некроз, наприклад, запрограмовану загибель клітин в ембріонах кукурудзи. Несумісність між Agrobacterium і клітинами кукурудзи ймовірно включає деякі генетичні компоненти. Цей винахід описує три гени з Agrobacterium, що приймають участь у загибелі клітин тканин кукурудзи. Вони визначаються скринінгом бібліотеки BAG Agrobacterium. Було знайдено 3 незалежних BAG, що викликають загибель клітин і виявляють гомологію з xyl A-xylB, virВ1 і acvB. Можливо, що ви щевказані гени є лише підсімейством генів, що відповідають за несумісність Agrobactenum з тканинами кукурудзи. Краще, якщо клітина рослини, використана в способах винаходу, є з виду рослини або сорту, який не утворює галлів або волосяних коренів при трансформації Agrobacterium. Краще, якщо видом рослини є член трав'янистої родини (Gramineae), найкраще кукурудза або пшениця, зокрема, кукурудза. Краще, якщо клітина рослини є тотипотентною клітиною, тобто клітиною, здатною до регенерації у рослину, краще фертильну рослину. Тотипотентні клітини присутні, наприклад, у незрілих ембріонах і ембріогенному калюсі. Краще, якщо тканиною-мішенню є інша, ніж апікальна меристема. Ще краще, якщо тканина-мішень містить недиференційовані тканини, наприклад, висаджені зиготичні ембріони або ембріогенний калюс. Ембріогенний калюс може бути з ембріону, взятого протягом фази ініціації калюсу (наприклад, періоду аж до 28 днів, краще аж до 14 днів від висадження незрілого ембріону на середовище для ініціювання) або може бути серійно розмножуваний ембріогенний калюс, наприклад, калюс Типу І або Типу II. Краще, якщо калюсом кукурудзи Типу І є тканина, що містить тотипотентні клітини для використання в цьому винаході. Краще, якщо Agrobacterium, вибрана з A.tumefaciens і A. rhizogenes. Краще, якщо штам Agrobactenum - це штам A.tumefaciens, найкраще штам, що використовує нопалін. Якщо штам Agrobacterium - це штам A.rhizogenes, то краще штам, що використовує агропіни або манопіни. Найкраще, якщо Agrobacterium - це Agrobacterium, яка не викликає некрозу у Gramineae, наприклад, Agrobactenum, вибраний зі штамів A. tumefaciens А і В. Штами Agrobactenum А і В були депоновані у Американське зібрання типових культур (АТСС) 12301 Parklawn Drive, Роквілль, штат Меріленд 20852/США, під АТСС номерами 55964 і 55965, відповідно, 2 травня, 1997, згідно з Будапештською угодою. Краще, якщо потрібний ген є геном, що відповідає за резистентність до гербіциду, резистентність до хвороби або резистентність до комах, або є селективним, або здатен бути підрахованим маркером, і містить промотор, функціональний у рослин, кодуючу ділянку і 3'-кінцеву ділянку. До генів резистентності до гербіцидів належать ген AHAS резистентності до імідазолінону або гербіцидів ряду сульфонілсечовини, ген резистентності pat або bar до біалафосу або глюфозинату, ген синтази EPSP, що надає резистентність до гліфозату, тощо. До генів резистентності до хвороб належать гени ферментів синтетичних антибіотиків, наприклад, ферменти синтетичних піролнітринів, гени резистентності з рослин й под. До генів резистентності до комах належать гени інсектицидних білків з Bacillus thuringiensls. До селективних маркерів належать гени резистентності до гербіцидів і гени резистентності до антибіотиків (наприклад, гігроміцину або канаміцину), також як і позитивні селективні маркери, такі як ген манозофосфатізомерази. До маркерів, які можна визначити кількісно, належать гени ферментів, активність котрих легко піддається підрахунку, такі як ген gus і ген cat. Плазміда може містити більш, ніж один потрібний ген і/або Agrobactenum може містити різні плазміди, що несуть різні потрібні гени. Термін Agrobacterium-індукований некроз (АІН), що використовується тут, стосується будь-якої викликаної Agrobacterium загибелі клітини рослини, але, зокрема, стосується викликаної Agrobacterium запрограмованої загибелі клітини. ПРИКЛАДИ Приклад 1 - Ме ханізм дії некрозу у кукур удзи, яку піддавали дії Agrobacterium а. Виявлення некрозу кукурудзи, але не тютюну в присутності Agrobacterium. Ембріогенний калюс кукурудзи й ембріони виявились особливо складними тканинами для трансформації Agrobacterium. У наступних прикладах використовуються інбредні лінії кукурудзи (Zea mays L.) НЕ/89 і лінію, що має назву Елітний 1. Крихкий ембріогенний калюс лінії кукурудзи НЕ/89 був наданий Gunter Dorm. НЕ/89 описаний у Morocz, S., Dorm, G., Nemeth, J. і Dudits, D. (1990) Theor. Appl. Genet 80: 721-726). Елітний 1 - це запатентована елітна лінія Новартіс (Novartis), споріднена В73. Суспензія культури Елітний 1 бере початок з крихкого ембріогенного калюсу, вибраного з незрілих ембріонів. НЕ/89 лінія вирощується у модифікованій рідкому середовищі N6 з додаванням 500мг/л Bacto tryptone, 30г/л сахарози і 0,5мг/л 2,4-дихлорфеноксиоцтової кислоти (2,4-D) (N6M). Елітна 1 лінія вирощується у рідкому середовищі N6 (Chu et al., 1975) з 30г/л сахарози і 2мг/л 2,4-D (2N63S). Лінія клітин Nicotiana tabacum NT-1, вирощена на середовищі Мурашиге та Скуга (Murashige and Skoog) з 2мг/л 2,4-D і сахарози (30г/л) (MS3S), використовується як Agrobacterium-Myrnmm контроль. Суспензії культур зберігаються у рідкому середовищі на роторному шейкері (120об/хв) і субк ультивуються кожні 7 днів. Бактерії вирощують на YP середовищі (5г/л екстракту дріжджів, 10г/л пептону, 5г/л NaCI, pH6,8) протягом 24 годин при 28°С. Бактерії центрифугують і ресуспендують у придатному рослинному середовищі. Для інокуляції Елітного 1, НЕ/89 і NT-1 клітин рослин, бактерії ресуспендують у 2N63SM, N6 M і MS3S рідких середовищах, відповідно. Невдовзі після інокуляції ембріогенного калюсу кукурудзи, багато з клітин стали некротичними (Таблиця 1). Некроз спостерігається навіть після перенесення ембріогенного калюсу на середовище, що не містить бактерій, і цефотаксимвмісне середовище. Кокультивації клітин кукурудзи з дуже розведеними культурами Agrobacterium протягом дуже короткого терміну часу достатньо, щоб викликати некротичну відповідь або інгібування росту інбредної лінії Елітний 1 кукурудзи. Для ембріогенного калюсу НЕ/89 короткий час кокультиваці зменшує некротичну зміну деякою мірою, але не повністю запобігає некрозові. У контрольних експериментах цефотаксим не викликає некрозу тканин кукурудзи. Таким чином, некротичний ефект є результатом взаємодії між Agrobacterium і калюсом кукурудзи. Клітини NT1 тютюн у також висівали з Agrobacterium як контроль і не спостерігали ніякого некрозу. Таблиця 1 Кокультивація суспензії клітин рослин LBA4404 Без Agrobacterium OD=0,2 10хв 1год 24год 48год OD=1 10хв 1год 24год 48год НЕ/89 Елітні 2 дні4 дні 7 дні 2 дні 4дні 7 дні + + + + + + + ++ + + + + + + ++ + ++ ++ +++ + + + + + + ++ ++ + + + + + ++ + ++ ++++ ++ +++ ++++ Крихкий ембріогенний калюс НЕ/89 кукурудзи і власну елітну інбредну кукурудз у, що зберігали у рідкому середовищі, інкубували з Agrobacterium протягом 10хв, 1год., 24год. або 48год. перед перенесенням у цефотаксимвмісне середовище. 50мкл бактерій при певній оптичній густині (OD=600нм) додавали до 10мл середовища для культивування тканин, що містило 1мл фіксованого об'єму клітин. Якщо культур у потрібно інкубувати довше, ніж два дні, бактерії вимивають і клітини рослин ресуспендують у тому самому середовищі з цефотаксимом (250мг/л). Присутність або відсутність некрозу підраховували на 2, 4 і 7 день після інокуляції і відносну інтенсивність некрозу виражали наступним чином: +, рідке утворення некрозу; ++, слабий некроз; +++, сильний некроз; ++++, дуже сильний некроз. б. Доказ запрограмованої загибелі клітин Для того, щоб визначити механізм загибелі клітин, викликаної Agrobacterium в тканинах кукурудзи, геномну ДНК екстрагують з суспензії клітин рослин, зібраних за 24 або 48 години після інокуляції з Agrobacterium. Піддання дії Agrobacterium викликає фрагментацію ДНК в обох лініях кукурудзи з властивостями внутрішньоядерної фрагментації, яку вважали початковою ознакою запрограмованої загибелі клітин. Контрольні клітини кукурудзи та тютюну, що зберігали лише у середовищі для культивування, не виявили фрагментації ДНК за цих умов. Інокуляція клітин кукурудзи з Е.соlі або з автоклавованими клітинами Agrobacterium не викликає фрагментації ДНК. Крім того, клітини кукурудзи, що зазнають запрограмованої загибелі, не змогли викликати загибель клітин свіжої ембріогенної тканини кукурудзи. Подальший доказ Agrobacterium-інаукованої фрагментації ДНК одержували, вивчаючи чутливість процесу до Zn2+ і Са2+. У клітинній лінії НЕ/89 Са2+ підвищив інтенсивність розмотування ДНК, тоді як Zn2+ помітно зменшив фрагментацію геному. Ці властивості повністю співпадають зі впливом цих двовалентних катіонів на ендогенні ендонуклеази, що відповідають за розщеплення ДНК протягом апоптозу у тварин. Інтенсивність розмотування також зменшували додаванням циклогексиміду, що є доказом того, що процес потребує експресії генів de novo. Ці результати свідчать, що контакт Agrobacterium з клітинами кукурудзи відповідає за загибель клітин тканини кукурудзи. Фрагментацію ДНК під час апоптозу також спостерігали in situ no реагентам, що реагують з відкритими гідроксильними групами на нуклеосомних одиницях. 12-добові ембріони кукурудзи з власної елітної лінії Lancaster parentage висівали з LBA4404 і висаджували на середовище DG4 з хлорамбеном. Koziel, M.G., et al. 1993. Bio/Technology 11: 194-200. Апоптотичні клітини виявляли in situ фарбуванням набором TACS-1 (Tre vigen), який включає мічення кінців фрагментів ДНК ферментом klenow з використанням дНТФ, кон'югованого з маркером, який можна визначити, з утворенням темного нерозчинного осаду, що вказує на фрагментацію геному. Cullivier, О., Pirianov, G., Kleuser, В., Vanek, P.O., Coso, O.A., Gutking, J.S. і Spiegel, S. (1996) Nature 381, 800-803. Ембріони вимочували у 3,7% формальдегідному розчині й обробляли, як описано в інструкціях виробника. Для визначення розщеплення геномної ДНК in situ фіксовані ембріони обробляли згідно з набором TACS-1. Фіксовані зрізи спочатку обробляли протеїназою К протягом 5хв. за кімнатної температури і потім занурювали на 5хв. у 2% пероксид водню для видалення ендогенної пероксидази. Після нетривалого промивання буфером klenow тканини інкубували протягом 1год. при 37°С з ферментом klenow, що може включати екзогенні модифіковані дезоксинуклеотиди до 5'-кінця розщеплених фрагментів. Темний нерозчинний осад, що вказує на апоптоз, який з'являється в окремих клітинах, визначали за допомогою мікроскопу. Agrobacterium-оброблеиі клітини становилися темнішими й деформованими за формою після 48-год обробки. Контрольні ембріони, що зберігали у середовищі для культивування або висівали з Е.соlі, не виявили будь-якого осаду за цих умов. Також було знайдено, що не всі тканини кукурудзи реагують однаковим чином на інфекцію Agrobacterium: корені залишають інтактними, тоді як ембріони зазнають некрозу. Фрагментацію ДНК також спостерігали для штамів Agrobacterium, які не містили патогенних плазмід (штами А136 і LBA4402), це дозволяє припускати, що фактор, який викликає запрограмовану загибель клітини, не стосується плазміди. Запрограмовану загибель клітини також спостерігали, коли ембріони кукурудзи інкубували протягом 20 хвилин з супернатантом, 20-кратно концентрованим зі штаму Agrobacterium (LBA 4404), що викликає загибель клітин, але не тоді, коли ембріони інкубували з супернатантом, який готували тим самим шляхом з Ε.coli. Запрограмована загибель клітин також не відбувається, якщо концентрований супернатант Agrobacterium нагрівали протягом 20 хвилин при 100°С перед інкубацією. Ембріогенний калюс кукурудзи часто містить зони клітин, ще очевидно живих, та інші зони, які є некротичними. Той факт, що лише обмежені зони відмирають, навіть у ситуаціях, в яких клітини за межами чутливої зони піддають одночасно дії стимулу, дозволяє припускати, що тимчасово або постійно існує різнорідність у елементах, що контролюють відповідь окремих клітин на різні стимули в організованій тканині. Кількість клітин в організованій тканині, що зазнають запрограмованої загибелі, може бути малою порівняно з загальною масою, і процес може бути асинхронним. Індукція PCD в одній або кількох клітинах у певній зоні або тканині може передати швидку загибель оточуючи х клітин, що спостерігали для клітин тварин, і останні можуть не виявляти фенотипу запрограмованої загибелі, але все рівно гинуть. Приклад 2: Інгібування Agrobacterium-індуковано'і запрограмованої загибелі клітин Приготування тканин для інокуляції з Agrobacterium: Незрілі ембріони (довжиною 1,2-2,2мм) асептично вирізали через 14-15 днів після запилення зі стерилізованих, вирощених у теплицях пестиків, і висаджували скутелумом догори на середовище для ініціювання калюсу, 2DG4+5мг/л хлорамбену (2DG4-5CN). Середовище 2DG4 - це середовище Дункана (Duncan), модифіковане таким чином, щоб містило 20мг/л сахарози. Ембріони, що дали калюс: Незрілі ембріони стерилізували, як описано вище, висаджували на середовище для ініціювання калюсу (2DG4-5CN) і культивували там протягом 1-7 днів. Тканини, що утворилися, використовують як зразки для інокуляції. Молодий калюс типу І: Ембріони висаджували на середовище для ініціювання калюсу (2DG+5 Хлорамбену) протягом 7-20 днів для утворення калюсу. Цей калюс є типовим калюсом типу І для використаних інбредів кукурудзи, які є компактними і відносно добре організованими кластерами клітин. Утворений ембріогенний калюс вирізали з ембріонів і переносили на середовище для зберігання калюсу 2DG4+0,5мг/л (2,4-дихлорфенокси)-оцтової кислоти (2,4-D) і використовували як зразки для інокуляції. Калюс типу І: Калюс типу І, одержаний способом, описаним вище, може зберігатися на середовищі для зберігання (2DG4+2,4 D 0,5мг/л) і субк ультивуватися кожні 2 тижні. Agrobacterium: У цих експериментах використовували штам A. tumefaciens LBA4404 (pAL 4404, pSB1). pAL4404 - це допоміжна плазміда. pSB1 - це плазміда, що може функціонувати у бага тьох хазяїв, яка містить ділянку гомології з pGIGUP і 15,2т.п.н. Kpnl фрагменту з вір улентної ділянки рТіВо542 (Ishida et al., 1996; High efficiency transformation of maize (Zea mays L) mediated Agrobacterium tumefaciens, Nature Biotechnology 14, 745-750). Уведення плазміди pGIGUP за допомогою електропорації у LBA4404 (pAL4404, pSB1) приводить до коінтеграції pGIGUP і pSB1. Т-ДНК цієї плазміди містить ген PAT, що експресується в рослинах, під контролем убіквітинового промотору для надання резистентності до фосфінотрицину, і ген GUS з інтроном у N-кінцевому кодоні кодуючої послідовності під контролем промотору Gelvin. Цей інтрон-GUS ген експресує активність GUS в клітинах рослин, але не у Agrobacterium. Ріст бактерій: Agrobacterium вирощували протягом 3 днів на середовищі YP (5г/л екстракту дріжджів, 10г/л пептону, 5г/л NaCI, 15г/л агару, рН6,8) з 50мг/л спектиноміцину і 10мг/л тетрацикліну. Бактерії збирали петлею і суспендували в рідкому середовищі N6 при густині 109-5´109клітин/мл. Клітини Agrobacterium також можна збирати з нічних культур на середовищі YP і ресуспендувати у рідкому середовищі N6. Кокультивація в присутності або без нітрату срібла: Тканини кукурудзи, одержані як описано вище, висівали з Agrobacterium. Тканини кукурудзи інкубували з суспензією бактерій протягом 510хв. і потім висаджували на середовище з або без нітрату срібла (1-10мг/л). Тканини також висівали 5мкл краплинами 109-5´109клітин/мл суспензії Agrobacterium, розташованими на верхньому шарі тканини. Після інокуляції тканини кукурудзи культивували в темряві при 25°С з або без нітрату срібла в середовищі (1-10мг/л). Через 2 або 3 дні після інокуляції тканини переносили на те саме середовище з цефотаксимом (250мг/л) з або без нітрату срібла. Результати: Чутливість ембріонів кукурудзи і ембріогенного калюсу кукур удзи до Agrobacterium. Ембріогенний калюс може розвиватися з ембріонів кукурудзи при висадженні на середовище для ініціювання калюсу. Клітинні лінії, що розвилися з ембріонів, залишатимуться високо ембріогенними при зберіганні на середовищі, що містить 2,4-D. Визначали чутливість цього ембріогенного калюсу і ембріонів з інбредної лінії кукурудзи до Agrobacterium LBA4404. Помітне коричневіння тканини спостерігали після кокультивації Agrobactenum і тканин кукурудзи. Цю реакцію також спостерігали з LBA4402, що не містив своїх патогенних плазмід. Контрольні ембріони й ембріогенний калюс, що кокультивували з Escherichia coli і піддавали тим самим процедурам, не виявив загибелі клітин або значного коричневіння. Таким чином, загибель клітин - це результат взаємодії між клітинами кукурудзи і Agrobactenum. Було виявлено, що ембріони є більш чутливими до Agrobactenum, ніж ембріогенний калюс. Концентрація 10 9 клітин/мл є достатньою, щоб викликати загибель клітин. Крім того, коротка обробка (5хв.) є достатньою, щоб викликати цю реакцію. Визначали ефект додавання нітрату срібла до середовища кокультивації і середовища для ініціювання калюсу. Додавання нітрату срібла створює умови для утворення здорових тканин (Таблиці 2 і 3). Приблизно 50% ембріонів, висаджених на середовище, що містило нітрат срібла, здатні утворювати ембріогенний калюс. Оптимальний ефект одержують з ембріонів, що висаджували на середовище для ініціювання калюсу протягом принаймні 2 днів перед висадженням з Agrobacterium. Було знайдено, що комбінація цих двох факторів занадто запобігає некрозові ембріогенного калюсу кукур удзи після кокультивації Agrobactenum. Зменшене коричневіння ембріогенного калюсу корелює з ініціюванням калюсу і виживаністю тканини після інокуляції Agrobactenum. Приблизно 80% контрольних ембріонів, які не обробляли Agrobacterium, дали початок ембріогенному калюсу. Ефект нітрату срібла також спостерігали з калюсом типу І, що висівали з Agrobacterium (Таблиця 3). Було виявлено, що молоді лінії більш резистентні до інокуляції Agrobacterium і попередня обробка нітратом срібла покращує відновлення здорової тканини. Для того, щоб довести, що присутність нітрату срібла протягом кокультивації не зменшує вірулентності Agrobacterium, листкові диски тютюну висівали з LBA4404(GIGUP) при концентрації 109клітин/мл. Ніякого значного зменшення активності GUS не спостерігали в листках тютюну. Таблиця 2 Інокуляція ембріонів кукурудзи з LBA4404 Ембріони й ембріони з ембріогенним калюсом висівали з Agrobacterium LBA4404 (GIGUP) (109клітин/мл). Основним середовищем було 2DG4+5 Хлорамбен (2DG4). Усі обробки були здійснені на приблизно 80-100 ембріонах і повторені двічі. Ініціювання калюсу підраховували за два тижні після інокуляції Попередня обробка (час) Без попередньої обробки Без попередньої обробки Без попередньої обробки 2DG4+AgNO3 (1 день) 2DG4+AgNO3 (4 днів) 2DG4 (6 днів) 2DG4 (6 днів) +1 день на 2DG4+AgNO3 Коінокуляційне середовище 2DG4 2DG4 2DG4+AgNO3 2DG4+AgNO3 2DG4+AgNO3 2DG4+AgNO3 2DG4+AgNO3 Перенесення Результат 2DG4+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 1% 30% 55% 43% 27% 65% 88% Таблиця 3 Інокуляція калюсу кукурудзи тип у І Калюс типу І висівали з Agrobacterium LBA4404 (GIGUP) (10 9клітин/мл). Основним середовищем, яке використовували в цих експериментах, було 2DG4+0,5 2,4D (2DG4) з або без нітрату срібла (AgNO3). Усі експерименти виконували на приблизно 150 шматках калюсу типу І. Виживаність тканин підраховували за 2 тижні після інокуляції Калюс типу 1 ½-місячні 10-місячні Попередня обробка 2DG4 2DG4+AgNO3 2DG4 2DG4 2DG4+AgNO3 2DG4 Коінокуляція 2DG4+AgNO3 2DG4+AgNO3 2DG4 2DG4+AgNO3 2DG4+AgNO3 2DG4 Перенесення 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+Cef 2DG4+AgNO3+Cef 2DG4+AgNO3+Cef 2DG4+Cef Результат 75% 95% 22% 55% 66% 12% Приклад 3: Трансформація Agrobactehum незрілих зиготичних ембріонів і виділення трансформованого калюсу з використанням фосфінотрицину, гізроміцину або манози як селекційного агента. Незрілі ембріони одержували приблизно за 10-14 днів після самозапилення. Незрілі зиготичні ембріони розподіляли серед різних планшетів, що містили середовище, здатне викликати і підтримували утворення ембріогенного калюсу при приблизно 25 незрілих ембріонах на планшет. Незрілі ембріони висівали або на планшет, або у рідину, як показано у прикладі 2 з Agrobacterium, що несли Ті-плазміду, яка містить селективний ген-маркер. Проведенням серій експериментів виявляються оптимальні умови для незрілих ембріонів. В одному оптимальному стані незрілі ембріони висаджували на середовище для ініціювання калюсу, що містило нітрат срібла (10мг/л), або перед або відразу після інокуляції з Agrobacterium. Приблизно 25 незрілих ембріонів розташовували на кожному планшеті. За 16-72 години після інокуляції незрілі ембріони переносили у середовище для ініціювання калюсу з нітратом срібла і цефотаксимом. Селекцію трансформованих клітин здійснювали наступним чином: а. маркер резистентності до РРТ: Трансформацію здійснювали з використанням штаму Agrobacterium, що несла плазміду з геном, яка кодує резистентність до фосфінотрицину на ділянці Т-ДНК. Трансформовані клітини вибирали in vitro застосуванням фосфінотрицину при концентрації 3мг/л 2-20 днів після інокуляції і підтримували загалом протягом 2-12 тижнів. Ембріогенний калюс, одержаний таким чином, піддають регенерації в присутності або без фосфінотрицину на стандартному середовищі для регенерації. Усі рослини перевіряються у тесті з хлорфенолом червоним (ХЧ) на резистентність до РРТ. Цей тест використовує рНчутливий індикаторний барвник для виявлення того, які клітини ростуть в присутності РРТ. Клітини, що ростуть, змінюють рН у середовищі й змінюють колір індикатора хлорфенолу червоного на жовтий. Рослини, що експресують ген резистентності до РРТ, легко визначали в цьому тесті. Рослини, позитивні в цьому ХЧтесті, перевіряються полімеразною ланцюговою реакцією ПЛР на присутність гена PAT. Рослини, які є позитивними у ПЛР-тесті, перевіряються Саузерн-блотингом. б. Маркер резистентності до гігроміцину: Трансформацію здійснювали з використанням штаму Agrobacterium, що несе плазміду з геном (hpt, гігроміцин В-фосфотрансфераза), який кодує резистентність до гігроміцину на ділянках Т-ДНК. Трансформовані клітини вибирають з використанням гігроміцину при концентрації 3 мг/л 2-20 днів після інокуляції і зберігають протягом 2-12 тижнів. Ембріогенний калюс, одержаний таким чином, піддають регенерації в присутності або без селективного агента на стандартному середовищі для регенерації. Усі рослини перевіряються на резистентність до гігроміцину. У цьому тесті легко визначали рослини, що експресують ген резистентності до гігроміцину. Рослини, позитивні у цьому тесті, перевіряються ПЛР на присутність гена hpt Рослини, які є позитивними у ПЛР-тесті, перевіряються Саузернблотингом. в. Позитивна селекція з манозою: Трансформацію здійснювали з використанням штаму Agrobacterium, який несе плазміду з геном (манозофосфатізомераза), що кодує толерантність до манози на ділянці Т-ДНК, манозу використовують для селекції трансформованих клітин in vitro. Ця селекція може бути застосована при концентрації 1г/л 2-20 днів після інокуляції і здійснюватись протягом 2-12 тижнів. Ембріогенний калюс, одержаний таким чином, може бути регенерований в присутності або без манози на стандартному середовищі для регенерації. Усі рослини перевіряються на тест з хлорфенолом червоним (ХЧ) на толерантність до манози. Цей тест використовує рН-чутливий індикаторний барвник для виявлення того, які клітини ростуть у присутності манози. Клітини, що ростуть, змінюють рН у середовищі та колір індикатору хлорфенолу червоного на жовтий. У цьому тесті легко визначали рослини, що експресують толерантність до манози. Рослини, позитивні по ХЧ-тесту, перевіряються ПЛР на присутність гена манози. Рослини, які є позитивними на ПЛР-тест, перевіряються Саузерн-блотингом. Приклад 4: Трансформація Agrobacterium калюсу з незрілих зиготичних ембріонів і виділення трансформованого калюсу з використанням фосфінотрицину, гігроміцину і манози як селективних агентів. Калюс типу І одержують з незрілих зиготичних ембріонів з використанням стандартних способів культивування. Приблизно 25 шматочків калюсу типу І переносили на середовище для зберігання, що містило нітрат срібла, або перед, або після інокуляції з Agrobacterium. Інокуляція може бути здійснена, як описано у прикладі 2. Приблизно за 16-72 годин після інокуляції калюс переносили у стандартне культуральне середовище, що містило нітрат срібла і цефотаксим. Селекція може бути застосована відразу після перенесення на це середовище або після 1-20 днів. Калюс потім субкультивували для селекції протягом приблизно 2-12 тижнів, після чого той калюс, що вижив і почав рости, переносили у стандартне регенераційне середовище для одержання рослин. Селекцію здійснювали як у попередньому прикладі для клітин, трансформованих генами резистентності до фосфінотрицину, резистентності до гігроміцину або манозофосфатізомерази. Приклад 5: Трансформація калюсу типу І кук урудзи інокуляцією з Agrobacterium і виділення трансформованого калюсу з використанням фосфінотрицину, гігроміцину і манози як селективних агентів. Калюс беруть з висаджених незрілих ембріонів елітного генотипу. Культури субкультивуються кожні два місяці на середовищі для зберігання (2DG4+0,5мг/л 2,4-D) і групи клітин, які збирали за 2-3 дні після субкультивування, переносили на середовище 2DG4. Після або перед інокуляцією з Agrobacterium групи клітин висаджували на середовище 2DG4, що містило нітрат срібла. Інокуляція Agrobactrium може бути здійснена, як описано у прикладі 2. За 16-72 години після інкубації калюс переносили у свіже середовище для зберігання, що містило цефотаксим і нітрат срібла. Калюс субкультивували на селекційному середовищі протягом приблизно 2-12 тижнів з селективним агентом, після чого калюс, який вижив і почав рости, переносили у стандартне регенераційне середовище для одержання рослин. Селекцію здійснювали яку попередньому прикладі для клітин, трансформованих геном резистентності до фосфінотрицину, резистентності до гігроміцину або манозофосфатізомерази. Приклад 6: Обробка тепловим шоком запобігає апоптозові, що викликається Agrobacterium Незрілі ембріони: Інбредні лінії кукурудзи вирощували в теплиці. Незрілі ембріони (0,8-2,2мм довжиною) асептично вирізали з поверхнево-стерилізованих колосків між 8 і 15 днями після запилення, залежно від факторів навколишнього середовища. Ембріони висаджували скутелумом догори на середовищі для ініціювання калюсу. Для більшості інбредних ліній ембріони висаджували на LS середовище (Linsmaier і Skoog, Physiol. Plants. 18: 100-127, 1965). Ембріони CG00526 висаджували на 2DG4+5мг/л хпорамбену (2DG4-5CN). Середовище 2DG4 - це середовище Дункана, модифіковане таким чином, що містить 20мг/л сахарози (Koziel et al., Bio/Technology 11: 194200,1993). Калюс типу І: Незрілі ембріони стерилізували, як описано вище, висаджували на середовище для ініціювання калюсу (2DG4-5Chl) і культивували там протягом 1-7 днів. Ембріони висаджували на середовище для ініціювання калюсу (2DG4+5 Chl) протягом 7-20 днів для утворення калюсу. Цей калюс є типовим калюсом типу І. Калюс типу І був як компактні кластери відносно добре організованих клітин. Створений ембріогенний калюс вирізали з ембріонів і переносили у середовище для зберігання калюсу, 2DG4+0,5мг/л 2,4-дихлорфеноксиоцтової кислоти (2,4-D) і використовували для інокуляції. Калюс типу І, одержаний способом, описаним вище, може зберігатися на середовищі для зберігання (2DG4+2,4 D 0,5мг/л) і субкультивуватися приблизно кожні 2 тижні. Agrobacterium; Штам, що використовували, був A.tumefaciens LBA4404 (pAL4404, p8B1). pAI-4404 - це допоміжна плазміда (Ooms et al., 7: 15-29, 1982). pSB1 - це плазміда, що може функціонувати у багатьох хазяїв, що містить ділянку гомології з pGIGUP і 15,2т.п.н. Kpnl фрагмент з вірулентної ділянки рТіВо542 (Ishida et al., 1996; Nature Biotechnology 14, 745-750). Уведення плазміди pGIGUP за допомогою електропорації у LBA4404 (pAL4404, pSB1) приводить до коінтеграції pGIGUP і pSBl pGIGUP містить ген фосфінотрицинацетилтрансферази (PAT), що експресується у рослин, під контролем убіквітинового промотору кукурудзи для надання резистентності до фосфінотрицину (Christensen et al., Plant Моl. Biol. 18: 675-689,1992). Вона також містить ген β-глюкуронідази (GUS) з інтроном у N-кінцевому кодоні кодуючої послідовності під контролем складеного промотору, що походить з генів октопін- і манопінсинтази (тример промотору октопінсинтази вище активуючої послідовності з доменом промотору манопінсинтази, Ni et al., Plant J., 7: 661-676, 1995). Цей інтрон-ген GUS експресує активність GUS в клітинах рослин, але не у Agrobacterium. Agrobacterium вирощували протягом 3 днів на середовищі YP (5г/л екстракту дріжджів, 10г/л пептону, 5г/л NaCI, 15г/л агару, рН6,8) з 50мг/л спектиноміцину і 10мг/л тетрацикліну. Бактерії збирали петлею й суспендували в рідкому середовищі N6 при густині 109-5´109клітин/мл. Agrobacterium може, як альтернатива, бути зібрана з нічних культур на середовищі YP і ресуспендована у рідкому середовищі N6. Він може також бути преіндукований протягом 4-6год у Agrobacterium індукційному середовищі (AIM; K2HPO4, 10,5г; КН2РО 4, 4,5г; (NH 4)2SO4 , 1,0г; NаЦитрат2Н2О, 0,5г; MgSO4H2 O (1M), 1,0мл; Глюкоза, 2,0г; Гліцерин, 5,0мл; MES (10мМ); Аце тосирингон (50100мМ); рН5,6). Теплова обробка тканин: Перед кокультивацією тканини кукурудзи переносили у посудину Еппендорфа у рідкому середовищі N6 й інкубували протягом 4хв. при 45°C у водяній бані. Середовище потім заміщували приготовленою суспензією Agrobacterium, як описано вище. Через 5хв. за кімнатної температури тканини висаджували на придатне тверде середовище. Через 3 дні після коінокуляції тканини або фарбували X-Glu для визначення активності GUS, або висаджували на середовище з цефотаксимом (250мг/л). Тканини кукурудзи потім перевіряли для визначення відповіді калюсу, що вказує на відсоток тканини, яка вижила після інокуляції Agrobactenum. Кокультивація: Тканини кукурудзи, які готували, як описано вище, висівали з Agrobactenum. Їх намочували у суспензії бактерій протягом 5-10хв. і потім висаджували на тверде середовище. Після інокуляції тканини кукурудзи культивували у темряві при 25°С. Через 2 або 3 дні після інокуляції тканини переносили на те саме середовище з цефотаксимом (250 мг/л). Визначення in situ фрагментації ДНК: Ембріони намочували у 3,7% формальдегідному розчині й обробляли, як описано виробником (Trevigen). Для визначення розщеплення геномної ДНК in situ фіксовані ембріони обробляли набором TACS-1 (Cullivier et al., Nature 381: 800-803, 1996). Фіксовані зрізи обробляли протеїназою К протягом 5хв. за кімнатної температури і потім занурювали протягом 5хв. у 2% пероксид водню для видалення ендогенної пероксидази. Після нетривалого промивання буфером klenow тканини інкубували протягом 1год. при 37°C з ферментом klenow, який здатний включати екзогенні модифіковані дезоксинуклеотиди до 5'-кінця розщеплених фрагментів. Темний нерозчинний осад, що вказує на апоптоз, що з'являється в окремих клітинах, визначали за допомогою мікроскопу. Результати: - Головний феномен коричневіння спостерігали після кокультивації Agrobactenum і тканин кукурудзи, а концентрація 109клітин/мл могла викликати загибель клітин. Ембріони виявились більш чутливими до Agrobactenum, ніж ембріогенний калюс. - Ембріони і калюс типу І піддавали тепловій обробці й потім висівали з Agrobactenum при концентрації 109клітин/мл. Тканини перевіряли через три дні кокультивації й культивування протягом тижня. Рівень захисту, що був наданий завдяки попередній обробці, підраховували по кількості тканин, що вижили після інокуляції. Усі ембріони, що висівали з Agrobacterium після попередньої теплової обробки, виявили ініціювання калюсу, тоді як жодного калюсу не утворилося з ембріонів, які не піддавали тепловій обробці (Таблиця 4). Таблиця 4 Ембріони висівали з 109клітинами/мл Agrobactenum LBA4404 (pGIGUP) і потім висаджували на середовище для ініціювання калюсу. Основне середовище - це 2DG4+5CN (2DG4). Усю обробку проводили на приблизно 80-100 ембріонах і повторювали двічі. Ініціювання калюсу підраховували за два тижні після інокуляції Попередня обробка Контроль Без теплової обробки + Agro Теплова обробка + Agro Ембріони з утвореним калюсом / Ембріони, що висівали 137/150(91%) 3/150(2%) 112/150(75%) Той самий корисний ефект теплової обробки спостерігали для калюсу (Таблиця 5). Таблиця 5 Калюс типу І висівали з 109клітинами/мл Agrobacterium LBA4404 (pGIGUP). Основне середовище, що використовували, 2DG4+0,5 2,4D (2DG4). Усі експерименти здійснювали на приблизно 1000 шматочках калюсу типу І. Виживаність тканин підраховували за 2 тижні після інокуляції Попередня обробка 3-місячна лінія калюсу типу 1 Agrobacterium Agro + теплова обробка 10-місячна лінія калюсу типу І Agrobacterium Agro + теплова обробка Виживаність 0/1000 »1000/1000 0/1000 »1000/1000 Захист, що спостерігали після попередньої теплової обробки, виявився зв'язаним з тепловою обробкою, тому що головні білки теплового шоку можуть бути ампліфіковані RT-ПЛР. - Ніякої фрагментації ДНК не визначали у ембріонів, що піддавали попередній тепловій обробці. - Ембріони з різних ліній кукурудзи висівали зі штамом Agrobacterium LBA4404 (GIGUP), вирощеним у розчині AIM з або без попередньої теплової обробки. Через три дні кокультивації ембріони фарбували на активність GUS. Результати наведені у Таблиці 6. Було виявлено, що попередня теплова обробка має корисний ефект на короткочасну експресію. Таблиця 6 Штам OF502 FNU007 HAF031 2N217A 1NJ20 JEF091 Обробка GUS поз. % від загального* HS 15 74 HS 90 96 HS 0 67 HS 13 46 HS 5 38 HS 59 81 Рейтинг GUS ** 0,3 1,8 2,0 3,4 0,0 0,9 0,1 0,4 0,1 0,4 1,2 1,4 -: Без теплової обробки. HS: тепловий шок. * відсотки GUS, основані на розмірі зразка з приблизно 50 незрілих ембріонів. ** Рейтинг GUS, оснований на 0-5 шкалі (0= без фарбування GUS, 1=1-5 плям/ембріон, 2=5-15 плям, 3=15-30 плям, 4=~25% поверхні ембріону фарбовано на GUS, і 5=~50% поверхні ембріону фарбовано на GUS). Приклад 7: Трансформація пшениці Agrobacterium Незрілі зиготичні ембріони (0,75-1,25мм) вирізали й висаджували на основане на MS середовище з 3мг/л 2,4-D, 300мг/л глутаміну, 150мг/л аспарагіну і 3% сахарозою (3MS3S) протягом 0, 3, 4, 5, 6, 7, 8 або 21 днів перед інокуляцією з Agrobacterium. До 21 дня експлантати утворили ембріогенну масу, що була 3-тижневим калюсом. Кокультивація: Тканини пшениці висівали з Agrobacterium, описаним у Прикладі 6. Тканини пшениці намочували у суспензії бактерій протягом 5-10хв. і потім висаджували на тверде середовище. Після інокуляції тканини пшениці культивували в темряві при 25°С. 2 або 3 дні після інокуляції тканини переносили на те саме середовище з цефотаксимом (250мг/л). Експлантати висівали без попередньої обробки або вони піддавались тепловому шоку при 42-48°С протягом 4-8 хвилин. Теплову обробку здійснювали перед інокуляцією. Ембріони можуть бути піддані тепловій обробці у 3MS inf або AIM з 100мМ AS рідині, тоді як калюс обробляли "сухим". Інокуляція: Експлантати висівали або у посудини Еппендорфа або на планшети. При висадженні у посудини 1 мл розчину Agrobacterium вносили в посудину, перемішували вручну або струшували і залишали на 5-15 хвилин. Розчин Agrobacterium і експлантатний матеріал потім виливали на чашку з 3MS3S з 100мМ AS і рідину видаляли одноразовою піпеткою для перенесення з вузьким наконечником. При завершенні інокуляції на планшети Agrobacterium переносили піпеткою безпосередньо на експлантати і видаляли за 5-15 хвилин. Ембріони розташовані таким чином, що осі ембріонів контактують з середовищем. Ініціювання калюсу, селекція і регенерація: Ембріони вирощували на середовищі для індукції калюсу (3MS3S) з антибіотиком протягом 3 тижнів. Ембріогенний калюс розрізали й розташовували на MS3S (без 2,4-D) з 5мг/л GA3 і 1мг/л NAA з селекційним агентом і антибіотиком протягом 2 тижнів, потім видаляли у MS3S з антибіотиком і вищою концентрацією селекційного агента на приблизно 4 тижні. Кільчики потім переносили у бокси Магента з 1/2 солями MS і 0,5мг/л NAA, підтримуючи концентрацію селекційного агента тією самою, як і на останньому етапі. Для калюсу селекція і регенерація та сама, за винятком того, що калюс вирощували лише протягом максимально 6 тижнів з часу висадження ембріонів, таким чином, висівали 3-тижневий калюс, кокультивували і вирощували протягом максимум ще 3 тижнів перед тим, як почнеться селекція і регенерація. Результати: - Фрагментацію ДНК вивчали у ембріонів, інокульованих Agrobacterium, як описано у Прикладі 6. Ніякої фрагментації ДНК не визначали у ембріонів, які піддавали попередній тепловій обробці. - Попередня теплова обробка перед інокуляцією Agrobacterium може запобігати початку апоптозу. Приклад 8: Пригнічення Agrobacterium-індукованого апоптозу в клітинах кукурудзи за допомогою р35 та іар Вектори для трансформації biolistic: Вектори, що використовували для трансформації кукурудзи пристроєм biolistic, всі є похідними pUC. pUbiPAT містить ген bar, що експресується в рослинах і кодує фосфінотрицин-ацетилтрансферазу (PAT) під контролем убіквітинового промотору кукур удзи (Christensen et al., 1992) для надання резистентності до фосфінотрицину (РРТ). Кодуючу ділянку р35, іар і cfacM клонували під контролем промотору металотіонеїнподібного гена кукурудзи (MTL) (de Framond, 1991). р35 і іар були надані Lois Miller (University of Georgia, Athens, Georgia). Фрагмент Pstl-EcoRI p35, що містить кодуючу ділянку, вирізали з pHSPSSVI* (Clem і Miller, 1994), клонували у відповідні сайти pBluescript (Stratagene, La Jolia, California) і потім клонували як фрагмент Pstl-Asp718 у відповідні сайти pMTL. pMTL містить промотор MTL і термінатор CaMV35S. Фрагмент Sall-Spel, що містить кодуючу ділянку іар з pHSCplAPVI* (Clem і Miller, 1994) клонували у Xhol-Spel сайти pMTL. Кодуючу послідовність dad-1 клонували як Sall-Xbal з кДНК клону 12Т7 Arabidopsis (Мічіганський університет). Вектори для трансформації Aarobacterium: р35, іар і dad-1 кодуючі ділянки під контролем промотору MTL клонували між граничними послідовностями у супербінарний вектор pSBH. Ген β-глюкуронідази (GUS) з інтроном у N-кінцевому кодоні кодуючої послідовності знаходиться під контролем складеного промотору з генів октопін- і манопінсинтази (mas; тример промотору октопінсинтази вище активуючої послідовності з доменом промотору манопінсинтази; Ni et al., 1995). Mas-GUS, що експресує активність GUS в клітинах рослин, але не у Agrobacterium клонували у pSB11 для одержання pMasGUS. Ці вектори потім уводили у LBA4404 (pAL4404, pSB1) за допомогою електропорації з 0,2см Bio-Rad кюветами при напруженості поля 2кВ/см, опорі 600Ом і ємності 25мкФ. pSB1 - це плазміда, що містить ділянку гомології з pSB11 і 15,2т.п.н. Kpnl фрагмент з вірулентної ділянки рТіВо542 (Ishida et al., 1996). Уведення плазміди pSB11 за допомогою електропорації у LBA4404 (pAL4404, p8B1) приводить до коінтеграції pSB11 і pSB1. Інокуляція ембріонів кукурудзи з Aprobactenum: Незрілі ембріони (0,8-2,5мм) асептично вирізали через 12-15 днів після запилення і висаджували скутелумом догори на середовищі для ініціювання калюсу (2DG4+5 Хлорамбен; Duncan et al., 1985). Ембріони висівали з Agrobactenum при густині 109клітин/мл протягом 5хв. і потім культивували на середовищі для ініціювання калюсу протягом 3 днів. Тканини потім переносили на те саме середовище з цефотаксимом (250мг/л). Виживаність тканин підраховували за 2 тижні після інокуляції. Ініціація калюсутипу І: Ембріони висаджували на середовище для ініціювання калюсу протягом 7-20 днів для утворення калюсу. Ці калюси є типовими калюсами типу І, які є компактними кластерами відносно добре організованих клітин (Suttie et al., 1994). Ембріогенний калюс, що утворився, вирізали з ембріонів і переносили у середовище для зберігання калюсу, 2DG4+0,5мг/л (2,4-дихлорфенокси)оцтової кислоти (2,4-D). Калюс типу І, одержаний у спосіб, описаний вище, може зберігатися на середовищі для зберігання (2DG4+2,4 D 0,5мг/л) і субкультивува тися приблизно кожні 2 тижні. Калюс типу І використовували в експериментах з інокуляцією Agrobactenum і для трансформації пристроєм biolistic. Експерименти з трансформації: Плазмідну ДНК осаджували на 0,3-1мм золотий мікроносій, як описано в інструкції DuPont Biolistic. 2мкг антиапоптотичного гена і 2мкг pUbiPAT використовували на 50мкл мікроносія. 16 шматочків калюсу типу І на кожен планшет бомбардували з використанням пристрою PDS-1000/He biolistic (DuPont). Тканини розташовували на полиці 8см нижче зупиняючого екрану і 10´10мкм екран з нержавіючої сталі використовували з розривними дисками 4,48МПа. Після бомбардування тканини культивували у темряві протягом одного дня при 25°С, потім переносили у середовище для зберігання калюсу 2DG4+0,5мг/л 2,4-D. Через 10 днів тканини переносили на те саме середовище з 100мг/л РРТ. Через 6-8 тижнів тканини переносили на 40мг/л РРТ. Деякі тканини потім використовували для експериментів з інокуляцією і деякі з них переносили на середовище для регенерації (Мурашиге і Скуга, що містило 3% сахарозу, 0,25мг/л ансимітолу і 5мг/л кінетину) з 16-годинною світловою добою. Трансформовані рослини визначали з використанням тесту з хлорфенолом червоним (ХЧ) для перевірки на резистентність до РРТ (Cramer et al.,1993) і потім підтверджували ПЛР. Інокуляція трансгенного калюсу типу і з Aarobacterium: Трансгенні тканини типу І одержували, як описано вище, висіванням з Agrobacterium. Тканини кукурудзи намочували у суспензії бактерій протягом 5-10хв. Після інокуляції тканини кукурудзи культивували в темряві при 25°С на 2DG4+0,5мг/л 2,4-D. За два або три дні після інокуляції тканини переносили на те саме середовище з цефотаксимом (250мг/л). Аналіз трансгенного матеріалу: Геномну ДНК екстрагують зі 100мг калюсу за допомогою набору Isoquick (Microprobre, CA) і ресуспендують у 20мкл води. 1мкл використовують для реакції ПЛР. Реакції ПЛР здійснюють у Perkin-Elmer термічних пристроях для підтримки циклів у 25мкл реакції з використанням 1´ПЛР буфер у, 0,5 одиниці AmpliTaq, 200мкМ кожного дНТФ, 0,2мкМ кожного праймеру. Для визначення присутності гену pat у тканинах, ПЛР реакції здійснювали з PAT праймерами при температурі гібридизації 55°С. Для lap, р35 і dad-1 визначення генів у трансгенних тканинах використовували наступні праймери для кожної реакції Ρ і І, Ρ і 35, Ρ і D при температурі гібридизації 55°С, 55°С, 48°С відповідно. РАТ1: РАТ2 MTL(P): lap (I): p35(35): dad-1(D): 5'-TG TCTCCGGA GAGG AGACC-3' 5'-ССААСАТСАТСССА ТССАСС-3' 5'-AGG TG TCCATGG TGC TCAAG- 3' 5'-ACAATCGA ACCGCACACGTC-3' 5'-CCAGGTA GCAGTCG TTG TG T-3' 5'-CCTTGmCCTTTG TTCAC T-3' (SEQ (SEQ (SEQ (SEQ (SEQ (SEQ ID ID ID ID ID ID NO: NO: NO: NO: NO: NO: 1) 2) 3) 4) 5) 6) p35 і lap RT-ПЛР: Загальну РНК екстрагували з трансгенного калюсу з використанням Tripure Isolation Reagent (Boehringer Mannheim), обробляли ДНКазою, що не містила домішок РНКази, і 0,5мкг брали для синтезу кДНК з використанням RT-ПЛР (Clontech) і праймер оліго-dT. Після синтезу другого ланцюга 1/20 цієї реакції використовували як матрицю для реакції ПЛР з Taq ДНК-полімеразою. р35 праймери, що використовували для RT-ПЛР, наступні: 5'-GGTCCTATACGAAGCGTTAC-3' (ПОСЛІДОВНІСТЬ №: 7) і 5'CCACGTAGC AGTCGTTGTGT-3' (ПОСЛІДОВНІСТЬ №:8), що ампліфікують 300п.н. транскрипту. праймери іар, що використовували для RT-ПЛР, це: 5'-CATGTCTGACTTGCGATTGG-3' (ПОСЛІДОВНІСТЬ №: 9) і 5'-ACAATCGAACCGC AC ACGTC-3' (ПОСЛІДОВНІСТЬ №: 10), що ампліфікують 248п.н. транскрипту. Для того, щоб виключити забруднення геномної ДНК, паралельно готували контрольні реакції кДНК, в яких не використовували зворотну транскриптазу. Результати: - Більшість ембріонів і ембріогенних калюсів не вижили після інокуляції з Agrobactehum. Коротка обробка (5хв.) є достатньою, щоб викликати цю реакцію. Тканини, що кокультивували з Eschehchia coli, піддавали тій самій процедурі, не виявляли будь-якого пошкодження. - р35, lap і dad-1 клонували у рослинну експресійну касету між граничними послідовностями Т-ДНК. Оскільки ці антиапоптотичні гени несуться Т-ДНК, вони повинні переноситися в клітини кукурудзи під час інокуляції. Тканини кукурудзи висівали з Agrobacterium LBA4404 (109клітин/мл) з або без генів-пригнічувачів загибелі клітин (у дужках). Після кокультивації ембріони кукурудзи або калюсу типу І перевіряють на виживаність як показник рівня захисту, що передається через експресію антиапоптотичних генів. Усі процедури проводили на приблизно 50-80 експлантатах кукурудзи і повторювали двічі. Тканини переносили через 3 дні після інокуляції на те саме середовище, що містило цефотаксим. Вплив Agrobactenum підраховували за два тижні після інокуляції. Результати наведені у Таблицях 7 і 8, відповідно. Таблиця 7 (ембріони) Без бактерій LBA4404 LBA4404 (р35) LBA4404 (lap) LBA4404 (dad-1) Кількість тканин, що виявили відповідь/Кількість перевірених тканин (%) Експеримент №1 Експеримент №2 52/63 (82%) 25/31 (80%) 2/55 (3%) 19/75 (25%) 3/89 (3%) 18/74 (24%) 22/89 (24%) 21/75 (28%) 4/48 (8%) 10/66 (15%) Таблиця 8 (калюс типу І) Без бактерій LBA4404 LBA4404 (р35) LBA4404 (іар) LBA4404 (dad1) Кількість тканин, що виявили відповідь/Кількість перевірених тканин (%) Експеримент Експеримент №1 №2 76/80 (95%) 72/80 (90%) 8/80 (10%) 6/80 (7.5%) 23/80 (29%) 27/80 (33%) 25/80 (31%) 30/80 (37%) 15/80 (18%) 13/80 (16%) Приблизно 80% ембріонів утворювали ембріогенний калюс, коли їх не обробляли Agrobacterium. Приблизно 25% ембріонів, які обробляли Agrobacterium, що несе або р35, або іар, утворювали ембріогенний калюс, тоді як лише 3% ембріонів, що висівали з Agrobacterium, виявили ініціювання калюсу. Подібний ефект спостерігали з калюсом типу І, який висівали з Agrobactenum, що несе р35 або іар (Таблиця2). dad-1 створював лише низький рівень захисту у будь-який системі. - Було виявлено, що короткочасна експресія р35 і іар певною мірою зменшує коричневіння тканин, але не повністю інгібує цей феномен. Це можна пояснити низькою ефективністю перенесення Т-ДНК у клітини кукурудзи. Ви ходячи з короткочасної експресії GUS, було виявлено, що тканина-мішень не однорідно трансформована. Це може бути показником того, що ли ше кілька клітин є реципієнтами Т-ДНК. - Структура ембріогенного калюсу, що виник з ембріонів, які висівали з Agrobacterium, що містив р35 або іар, не була такою компактною як контроль. Це можна пояснити тим фактом, що лише клітини, які одержували Т-ДНК, що несла антиапоптотичні гені, вижили, і що окремі трансформовані клітини не обов'язково передають сигнал до сусідньої клітини. Цей тип росту тканини може бути доказом того, що гени-пригнічувачі загибелі клітин рятують приречені клітини дуже ефективно, але їх короткотривала експресія не надає повного захисту проти впливу Agrobactenum. - Ембріогенні калюси, трансгенні по р35, іар і dad-1, утворилися внаслідок перенесення вищезгаданих генів під контролем промотору, що експресується в рослинах, у калюс типу І з використанням бомбардування мікрочастками. Конструкти кобомбардують геном bar, тканини піддають селекції на РРТ і трансформацію цього калюсу спочатку підтверджують ПЛР (Таблиця 9). Таблиця 9 Трансформаційний експеримент UbiPAT MTLp35, UbiPAT MTLiap, UbiPAT MTLdad, UbiPAT #ПЛР+ 5PAT+ 9PAT+, 3p35+ 7 PAT+, 5iap+ 11 PAT+, 4dad-1+ Чутливість до інокуляції Agrobacterium потім перевіряли на незалежні події. Калюс висівали з Agrobacterium LBA4404 (109клітин/мл). Перевіряли дві незалежні лінії трансгенного калюсу. Усі процедури повторювали двічі. Тканини переносили через 3 дні після інокуляції на те саме середовище, що містило цефотаксим. Виживаність тканин кукурудзи підраховували за два тижні після інокуляції з Agrobacterium (Таблиця 10). Таблиця 10 Експеримент з трансформації UbiPAT MTLp35, UbiPAT MTLiap, UbiPAT MTLdad, UbiPAT Кількість резистентного до LBA4404 калюсу Лінія 1 Лінія 2 1/24 (4%) 1/32 (3%) 22/25 (88%) 25/28 (89%) 20/22 (90%) 19/25 (76%) 12/27 (44%) 9/22 (41%) Ембріогенний калюс, трансгенний по р35 і іар, не виявляв загибелі клітин при інокуляції з Agrobacterium, тоді як головний феномен коричневіння спостерігали на контрольних тканинах після кокультивації. Присутність dad-1 гену затримувало початок апоптозу, але не блокувало загибелі клітин, також як і іар та р35. Результати повторювали у кількох незалежних експериментах. - Захисний ефект р35 і іар був також очевидний при вимірюванні за допомогою електрофорезу олігонуклеосомальних фрагментів ДНК. - Було виявлено, що ефект dad-1 більш очевидний, коли він стабільно експресується в тканинах - Існує чітка кореляція між експресією антиапоптотичних генів і відсутністю загибелі клітин. Гени р35 та lap експресуються в трансгенному калюсі, про що свідчить зворотна транскрипція-ПЛР (RT-ПЛР). Контроль був однорідно негативним. Приклад 9: Скринінг й ідентифікація штамів Agrobacterium, які не викликають загибель клітин кукурудзи Чутливість ембріогенного калюсу та ембріонів добре вивчених ліній кукурудзи (А188) і власної елітної ліні (Елітна 2) до різних штамів Agrobacterium перевіряли з використанням панелі з 40 штамами дикого типу Agrobacterium. Ці штами спочатку перевіряли на їх сумісність з ембріогенним калюсом і ембріонами А188 і Елітним 2. Серед них 6 з 40 штамів не повністю запобігають росту тканин кукурудзи і надалі перевіряються на Елітних 2 ембріонах, висаджених на середовище для ініціювання калюсу і висіяних при концентрації 10 9 клітин/мл. Ініціювання калюсу підраховували за 2 тижні після інокуляції й експресували як відсоток ембріонів, що мали відповідь. Ці штами також перевіряли на їх вірулентність стосовно листкових дисків тютюну, висаджених на середовище, що не містило гормонів, і варто уваги те, що потенціал штамів, який викликає запрограмовану загибель клітини у кукур удзи, не корелює з вірулентністю. Відсоток у Таблиці 11 описує кількість листкових дисків з пухлинами. Таблиця 11 Порівняння вірулентності ненекрогенних штамів Штами 1 2 3 4 А В Джерело Вірулентність, що 2DG4+5Chl Опін перевіряли (%) на тютюні (%) ? 100 37 нопалін 82 25 Роза Персик Срібляста тополя аргопін хризантема L,L Sap роза нопалін salix нопалін 66 100 92 76 50 63 66** 58** ** Хороший кал юс типу І. Штами Agrobacterium А і В були депоновані в АТСС 2 Травня, 1997 згідно з Будапештською угодою під АТСС номерами 55964 і 55965, відповідно. Приклад 10: Гени Agrobacterium, що передають апоптоз у клітини кукурудзи Бібліотеки BAG (штучні хромосоми бактерій) створювали з геномних фрагментів ДНК Agrobacterium приблизно 100-200т.п.н. з метою визначення генетичних елементів, що відповідають за загибель клітин у ембріогенній тканині кукурудзи. Бібліотеки вводили у Escherichia coli. Для того, щоб перевірити, чи такий фон придатний для скринінгу бібліотеки ВАС, Е.соlі, що містила лише вектор ВІВАС, висівали на ембріони кукурудзи. Було знайдено, що Е.coli не викликає характерної апоптотичної відповіді. Було підраховано, що приблизно 25-30 клонів ВАС з 150-200т.п.н. вставками є достатніми, щоб "покрити" цілий геном Agrobacterium, припускаючи, що Agrobacterium має розмір геному, доволі еквівалентний Е.coli (4,6´106п.н.). Відносно велику кількість клонів (200) гібридизували з Agrobacterium хромосомним зондом (chvD) для того, щоб перевірити, чи бібліотеки ВАС, зроблені у Ε. coli, є чіткою копією геному Agrobacterium. Серед 200 клонів, що перевіряли, 5 засвітилися з зондом chvD (дані не наведені). Однією з проблем є те, що генетичні компоненти з Agrobacterium можуть "мовчати" у Е.coli. Крім того, додаткові фактори з рослин і/або Agrobacterium можуть бути необхідними для експресії такого клону. Таким чином, вектор ВАС також містить початок реплікації, функціональний у Agrobacterium, і може бути легко трансформований у Agrobacterium таким чином. Ріст бактерій: Agrobacterium вирощували протягом 3 днів на середовищі YP (5г/л екстракту дріжджів, 10г/л пептону, 5г/л NaCI, 15г/л агару, рН6,8) з 100мг/л канаміцину при необхідності. Бактерії збирали петлею і суспендували в рідкому середовищі N6 за густини 109-5´109клітин/мл. Клітини Agrobacterium можуть також бути зібрані з нічних культур на середовищі YP і ресуспендовані у рідкому середовищі N6. Е.соlі DH10B вирощували у LB середовищі (1% бакто-триптону, 0,5% екстракту бакто-дріжджів, 170мМ NaCI, рН7,0). Після трансформації бактерій вирощували у розчині SOC (2% бакто-триптону, 0,5% екстракту бакто-дріжджів, 10мМ NaCI, 2,5мМ KСІ, 10мМ МgСІ2, 10мМ MgSO 4, 2мМ глюкоза, рН7,0). Приготування геномної ДНК з Aprobactenum: Штам LBA4404 використовували для створення бібліотеки. Протокол, матеріали і реагенти, що використовували, - усі з набору Imbed (Bioiabs, USA). 4 ´10мл культур Agrobacterium вирощували вночі при 28°С. За годину перед збиранням додавали 180мг/мл хпорамфеніколу для вирівнювання хромосом. Клітини центрифугували при 800´g при 4°С протягом 15хв. і осад висушували на сонці. Клітини попередньо нагрівали у 0,5мл розчину суспензії клітин (10мМ Tris-HCI, 20мМ NaCI, 100мМ EDTA, рН7,2) до 42°С перед зануренням у агарозу. Клітини Agrobactenum занурювали у агарозу для здійснення деградації клітинної стінки і депротеїнізації, уникаючи пошкодження ДНК. Для того, щоб занурити клітини у агарозні стрижні, клітини перемішують з однаковим об'ємом 1% агарози, що має низьку точк у плавлення, у ddН2О, о холоджену до 42°С, і потім залишають для затвердіння у формі Gel Syringe. При затвердінні агарози шматочки її переносили у 3 об'єми розчину лізозиму (7мг/мл лізозиму, 10мМ Tris-HCI, 50мМ NaCI, 100мМ EDTA, 0,2% дезоксихолату натрію, 0,5% N-лаурилсаркозину, рН7,2) й інкубували протягом 2 години при 37°С з обережним перемішуванням. Розчин лізозиму видаляли і стрижні промивали двічі 3 мл розчину для промивання протягом 15хв. одночасно. Стрижні потім інкубували у 3 мл розчину протеїнази K (1мг/мл протеїнази К, 100мМ EDTA, 1% N-лаурилсаркозину, 0,2% натрію дезоксихолату, рН8,0) протягом 20 годин при 42°С з обережним перемішуванням. Розчин протеїнази K видаляли відсмоктуванням і стрижні діалізували двічі проти 5мл розчину для промивання (20мМ Tris-HCI, 50мМ EDTA, рН8,0) протягом 15хв., потім 3мл розчину PMSF (20мМ Tris-HCI, 50мМ EDTA, 1мМ фториду фенілметилсульфонілу, рН8,0) протягом однієї години, після чого проводили два промивання розчином для промивання. Стрижні потім промивали двічі 5мл розчину для зберігання (2мМ Tris-HCI, 5мМ EDTA, рН8,0) протягом 15хв. і потім знову завантажували у GelSyringes. Зразки 1мм довжиною (10-20мл) нарізають лезом і діалізують проти буферу ендонуклеази рестрикції Notl (Biolabs, USA) протягом 30хв. при 4°С. Буфер ендонуклеази рестрикції заміщують на свіжий буфер й інкубацію здійснюють вночі при 37°С з 20 одиницями ферменту Notl (Biolabs, USA). Часткове розщеплення ДНК Agrobactenum досягається використанням Notl, що є рідким рестрикційним ферментом. Notl теоретично утворює фрагменти довжиною 18т.п.н. після загального розщеплення, припускаючи, що геном Agrobactenum має вміст G/C 60% (результати екстрапольовані з різних генів Agrobacterium). Аліквоти потім завантажували піпеткою, після чого кінчик відрізали. ДНК відокремлювали електрофорезом з пульсуючим полем у гелі (PFG) на 1% агарозного геля у 0,5´ТВЕ при 14°С з використанням Bio-Rad CHEF Mapper при 6В/см протягом 32год. Гель фарбували протягом 20хв. у 1мг/мл EtBr ddH2O бані й агарозні зрізи, які містили фрагменти, що відповідали 100-200т.п.н., вирізали з гелю. Агарозні зрізи діалізували двічі проти ТЕ протягом однієї години при 4°С і двічі з агаразним буфером (Biolabs, USA) протягом однієї години при 4°С, розплавляли при 65°С протягом 10хв. і розщепляли однією одиницею агарази (Biolabs, USA) на 100мг агарози. Концентрацію ДНК визначали флуориметрично з використанням DyNA Quant 200 (Hoefer, USA). Одержання вектору ВІВАС: Вектор ВІВАС був люб'язно наданий доктором Carol M.Hamilton (Cornell University, Ithaca) (Hamilton et al., Proc. NatL Acad. Sci. (USA) 93, 9975-9979, 1996). ВІВАС призначений для реплікації як у E.coli, так і Agrobacterium, і також містить ген sacBII, що дозволяє пряму селекцію для клонованих вставок на сахарозному середовищі. Вектор ВІВАС також несе F-фактор епісомного походження, який дозволяє стабільне зберігання однієї або двох копій на клітину з аж до 300 т.п.н. вставками (Shizuya et al., Proc. Natl. Acad. Sci. (США) 89, 8794-8797, 1992). Плазміду виділяли з використанням набору Wizard® Plus Maxiprep (Promega, USA). Фенолхлороформну екстракцію здійснювали перед осадженням ізопропанолом. Плазміду розщепляли Notl і дефосфорилювали 1 одиницею лужної фосфатази креветки (Boerhinger Mannheim) при 37°С протягом однієї години. З'єднування здійснювали у 40мл, в яких 150нг ДНК Agrobacterium інкубували з 17нг обробленого вектору (підраховане молярне співвідношення 10:1 з вектором у надлишку) з 400 одиницями Т4 ДНК лігази (Boerhinger) при 17°С вночі. Перед трансформацією з'єднування діалізували проти ТЕ і 1/10 ТЕ у MicroCollodion мішечках (Sartorius, Germany) при 4°С протягом 2 годин. Трансформація DH10B бібліотекою рВАС Трансформацію компетентних клітин DH10B Е.coli (F, recA1; ElectroMax, Gibco-BRL, США) здійснювали за допомогою електропорації у попередньо охолоджених Gene Pulser кюветах (Bio-Rad, USA) з Gene Pulser™ (Bio-Rad, США) з використанням наступних показників: напруга 2,5кВ, ємність 25мФ і опір 200Вт. 25мл компетентних клітин перемішують з 1мл розчину для з'єднання для кожної електропорації. Після електропорації клітини переносили у 0,5мл розчину SOC й інкубували при 37°С у колесі, що обертається навколо своєї осі, протягом 45хв. 150мл аліквоти клітин висаджували на середовище LB, що містило канаміцин (40мг/мл) і сахарозу (5%). Виділення ДНК ВАС і визначення розміру вставки ДНК клонів ВАС ВАС клони вирощували протягом ночі за тривалого перемішування у 10мл LB з канаміцином (50мг/мл). ДНК екстрагували згідно зі способом лужного гідролізу (Sambrook et al., 1989) з наступними модифікаціями: Рнказу 30мг/мл додавали до ресуспендованого розчину і здійснювали одне очищення фенол-хлорформом перед осадженням ізопропанолу. Осад н уклеїнової кислоти розчиняли у 50мл ddH2O. ДНК відокремлювали PFG електрофорезом на 1% агарозному гелі у 0,5 ´ТВЕ при 14°С з використанням Bio-Rad CHEF Mapper при 6В/см протягом 32 годин. Інокуляція ембріонів кукурудзи з Acirobacterium: Незрілі ембріони (0,8-2,5мм) асептично вирізали 12-15 днів після запилення й висаджували скутелумом догори на середовище для ініціювання калюсу (2DG4+5 Хлорамбен) (Duncan "D" середовище з глюкозою: N6ocновa, В5-мінор, MS залізо, 20г/л сахарози, 5мг/л хлорамбену, 8г/л очи щеного агару, G4 додатки і 10мг/л глюкози, рН5,8) (Duncan et al., 1985). Бактеріальні клітини ресуспендують у 2N63SM (3 ,97г/л N6 солей, N6 вітамінів, 30г/л са харози, 2мг/л 2,4-D, 8г/л очи щеного агару, рН5,8). Ембріони висівали з Agrobacterium або з Е.соlі при густині 109клітин/мл протягом 5хв. і потім культивували на середовищі для ініціювання калюсу протягом 3 днів. Тканини потім переносили на те саме середовище з цефотаксимом (250мг/л). Виживаність тканин підраховували через 2 тижні після інокуляції. Трансформація Agrobacterium Трансформацію Agrobacterium здійснювали, як описано вище (Wen-Jun і Forde, Nucleic. Acids Research 20: 8385,1989), у 0,2см кюветі для електропорації (Віо-Rad Laboratories Ltd., США) з використанням Gene Pulser™ (Bio-Rad, США) при напруженості поля 10кВ/см (2,00кВ), ємності 25мФ і опорі 600Ом. Вектори для трансформації biolistic: Вектори, що використовували для трансформації кукурудзи пристроєм biolistic, є всі похідними pUC. pMTL - це трансформаційний вектор рослин, що містить промотор металотіонеїнподібний ген кукурудзи (MT-L) (de Framond, 1991). Кодуючу ділянку wB1 ампліфікували ПЛР з клоном BAG як матрицею і зі штамом А з наступними праймерами: 5'-GGAGAGGCGGTGTTAGTT-3' (ПОСЛІДОВНІСТЬ №: 11); 5'-CATC ATCGC ATTCGAGAG-3' (SEQ ID NO: 12). ПЛР реакції здійснювали у Perkin-Elmer термічних пристроях для проведення циклічних реакцій у 25мкл реакційного середовища з 1´ ПЛР буфером, 0,5 одиниці AmpliTaq, 200мкМ кожного дНТФ, 0,2мкМ кожного праймера за температури гібридизації 55°С. Продукт ПЛР спочатку клонували у вектор pCR2.1 для ТА клонування (Invitrogen, San Diego, США). Потім вирізали Xbal-Spel подвійним розщепленням і потім клонували у Xbal сайт вектора pMTL у змістовному і антизмістовному положенні з утворенням pMTL-virBt і pMTL-virBlas, відповідно. Так само, продукт ПЛР, ампліфікований зі штаму А, клонували у pMTL у змістовному і антизмістовному положенні з утворенням pMTL-virBIA і p MTL-virBIAas, відповідно. acvB клонували у Xbal-Xhol сайт pMTL-змістовної і рМТІ_антизмістовної з утворенням pMTL-acvB і pMTLacvBas. pGUS містить ген β-глюкуронідази (GUS) з інтроном у N-кінцевий кодон кодуючої послідовності й під контролем складеного промотору з генів октопіну і манопінсинтази (тример промотору октопінсинтази вище активуючої послідовності з доменом промотору манопінсинтази) (Ni et al., 1995). Суспензії клітин кукурудзи Суспензії культур Zea Ma ys сорту Black Me xican Sweet (BMS) зберігали у N6 середовищі (Chu et al., 1975) з додаванням 30г/л сахарози і 2 мг/л 2,4-дихлорфеноксиоцтової кислоти (2,4-D) (2N63S). Суспензії клітин кукурудзи, що використовували для бомбардування, одержують з 3-добових культур, що швидко ростуть. Перед бомбардуванням приблизно 0,5мл фіксованого об'єму клітин фільтрують у вакуумі на 7-см фільтрах (Whatman, №4). Висаджені клітини зберігали протягом 4 годин при 25°С на середовищі 2N6 з фітагаром як затверджувачем, що містило 120г/л сахарози перед бомбардуванням. Бомбардування клітин рослин Тканини бомбардували золотими мікрочастками, на які осаджували суміш плазмід. ДНК плазміди pGUS використовували як внутрішній контроль при дослідженнях на короткочасну експресію. Для котрансформаційних експериментів золоті частки несли рівну масу усіх ДНК плазмід (0,5мкг кожної ДНК плазміди на мішень). Придатну кількість кожної ДНК перемішують у загальному об'ємі 10мкл і осаджували 50мкл 2,5Μ СаСl2 і 20мкл 0,1Μ основи без спермідину на 50мкл 0,3мкм золоті мікроносіїв (60мг/мл). Бомбардування мікрочастками здійснювали пристроєм PDS-1000 He biolistic (DuPont) з використанням 7,58МПа розривних дисків зі зразком, розташованим на 8см нижче зупиняючого екрану. Тести короткочасної експресії β-глюкуронідазну активність визначали хемолюмінесцентним тестом з набору GUS-Light (Tropix). βглюкуронідазну активність виражали у одиницях світла, які визначали люмінометром Analytical Luminescence model 2001 протягом 10 секунд при 25°С. Гібридизація колоній Однодобові BAG клони переносили на нейлонові фільтри (Hybond-N, Amersham Life Sciences, Великобританія) для гібридизації з радіоактивно міченими Agrobacterium зондами. ДНК-зонди мітили [а32 Р]дЦТФ з використанням набору для мічення Pharmacia. chvG зонд і w7D1 зонд відповідали ПЛР ампліфікованому фрагменту. Фільтри бібліотеки попередньо гібридизували у гібридизаційному розчині при 65°С протягом двох годин (5´SSPE, 5´ Denhardt's розчин, 0,5% ДСН 0,1мг/мл ДНК зі сперми лосося). Гібридизацію здійснювали у тому самому буфері при 65°С протягом ночі. Фільтри промивали у 1´SSC, 0,1% ДСН протягом 15хв. при 65°С, після чого інкубували у 0,1´SSC, 0,1% ДСН протягом 15хв. при 65°С. З фільтрів знімали сухі відбитки і плівку залишали протягом ночі при -70°С з використанням інтенсифікуючого екрану. Для ампліфікації ПЛР петлю LBA4404 ресуспендували у ddH2O, нагрівали до 95°С протягом 15хв., центрифугували і 1мл використовували для ПЛР реакції за наступних умов: 5хв. при 95°С і потім 30 циклів при 95°С протягом 45 секунд, при 55°С протягом 45 секунд і при 72°С протягом 45 секунд. Результати - Приблизно 20 ембріонів кукурудзи висівали з одним клоном з ВАС бібліотеки і висаджували на середовище для ініціювання калюсу. Після кокультивації ембріони переносили на те саме середовище з 250мг/мл цефотаксиму. Ембріони потім перевіряли на пошкодження, викликані рекомбінантними бактеріями. Серед 160 клонів, які піддавали скринінгу, визначали 4 незалежні ВАС клони (ВАС1, ВАС2, ВАС3, ВАС4). Для подальшого визначення ділянки на клоні ВАС, що відповідає за загибель клітин тканин кукурудзи, фрагменти Hindlll з клонів ВАС субклонували у вектор bluescript Кожний з цих клонів перевіряються на тканинах кукурудзи для того, щоб визначити, який з них зберіг здатність викликати загибель клітин. - Послідовності ДНК визначали, використовуючи субклони й олігонуклеотиди. Послідовності ВАС1-2, ВАС2-2 і ВАС3-2 виявили гомологію з virB1, xyl A-xylB і acvB, відповідно. ВАС-4 такий самий, як і ВАС3-2. - ВАС клони, що несуть acvQ (ВАС3 і ВАС4), virB1 (BAC1) і xyl A-xylB (BAC2) уводили у штам А, що дало штами A(acv/B), A(virB1) і А(хуІА). Ці штами використовували для інокуляції тканин кукурудзи. Тканини, що висівали з LBA4404, виявили типовий зразок загибелі клітин, тоді як інфекція штамом А не змогла створити ці симптоми. A(acvB), A(virB1) і А(хуl А) викликали загибель клітин на різних рівнях, A(acvB) - більше всіх. Присутність цих генів таким чином перетворює взаємодію штаму А з к укур удзи і робить тканини кукурудзи більш чутливими до інфекції (Таблиця 12). Таблиця 12 Штами Без бактерій LBA4404 Е.соlі Штам А Е.соlі (ВАС1) Е.соlі (ВАС2) Е.соlі (ВАС3) Е.соli (ВАС4) Штам А (ВАС1) Штам А (ВАС2) Штам А (ВАС3) Штам А (ВАС4) Експ.1 19/22 (86%) 2/20 (10%) 17/20 (85%) 19/20 (95%) 5/20 (25%) 10/19 (52%) 3/18 (16%) 4/19 (21%) 5/19 (26%) 9/22 (41%) 3/20 (15%) 2/20 (10%) Експ.2 15/18 (83%) 1/17 (6%) 16/19 (84%) 17/21 (81%) 5/17 (30%) 10/20 (50%) 4/17 (24%) 3/19 (16%) 6/17 (35%) 10/23 (43%) 2/18 (11%) 3/19 (16%) Експ.3 16/18 (88%) 3/20 (15%) 17/21 (81%) 17/20 (85%) 4/18 (22%) 9/19 (47%) 3/20 (15%) 2/21 (9%) 8/25 (32%) 8/19 (42%) 3/21 (14%) 4/20 (20%) - У Саузерн аналізі Hindlll-розщеплену геномну ДНК з штамів Agrobactenum А136, LBA4404, LBA4402, А і В зондували за помірних умов з кожним фрагментом, що несе ху/А або acvB. Фрагменти, гомологічні ху/А, присутні у всі х перевірених штамах. Послідовності, що гібридизувалися з acvB, були знайдені у LBA4404, LBA4402 і А136 і викликали загибель клітин кукурудзи. Цікаво те, що acvB немає у штамах А і В Agrobactenum. acv/B таким чином обмежений у своєму розповсюдженні. Як вже було згадано, acvB присутній у відносно малій кількості штамів Agrobactenum (Wirawan et al., Biosci. Biotech. Biochem. 60: 44-49, 1993). ху/А і acvB не входять у Ті-плазміду, тому що вони присутні в штамі LBA4402, що не несе Ті-плазміди. - Для того, щоб визначити, чи експресія acvB і virB1 всередині клітин кукурудзи є летальною, ці гени клонували у рослинний експресійний вектор- і котрансформували pGUS вектором, що експресує βглюкуронідазою (GUS). У випадку, якщо ген, що перевіряють, є летальним, активність GUS зменшена або зовсім немає (Mindrinos et at., 1994). Кодуючу ділянку двох генів субклонували у змістовному й антизмістовному розташуванні у рослинний експресійний вектор, що містив промотор MTL, і за допомогою бомбардування клітин кукурудзи разом з вектором pGUS. За 30 годин активність GUS оцінювали у клітин кукурудзи. Результати показані у Таблиці 13. Активність GUS зменшена приблизно вдвічі, коли virB1 кобомбардували з pGUS. Клітини кукурудзи, що кобомбардували acvB і pGUS, узгоджено виявили 35-кратне зменшену активність GUS. Це свідчить, що експресія acvB у великій кількості всередині клітин кукурудзи є шкідливою. Таблиця 13 Конструкти плазмід переносяться в клітини кукурудзи пристроєм biolistic. Після інкубації протягом 30год. тканини гомогенізують і визначають GUS активність. Активність виражали в одиницях/мг білка. Незалежні бомбардування аналізуються і дані як середні значення п'яти повторів ±SD. Плазміни +pGUS PUC pMTL-virB-l pMTL-virB-las pMTL-virB1K6 pMTL-virB1K6as pMTL-acvB pMTL-acvBas Середнє ±SD (одиниці/мкг білка) 2082±199 1014±106 2543+176 1738±330 2168 ±282 61 ±19 2378 ±292 ОПИС ПОСЛІДОВНОСТЕЙ (1) ЗАГ АЛЬНІ ДАНІ: (і) ЗАЯВНИК: (A) ІМ'Я: NOVARTIS AG (B) ВУЛИЦЯ: Schwarzwaldallee 215 (C) МІСТО: Базель (Ε) COUNTRY: Швейцарія (F) ПОШТОВИЙ КОД: 4058 (G) ТЕЛЕФОН: +41 61 324 11 11 (Η) ТЕЛЕФАКС: + 41 61 322 75 32 (іі) НАЗВА ВИНАХОДУ: Способи трансформації рослин (ііі) КІЛЬКІСТЬ ПОСЛІДОВНОСТЕЙ: 10 (iv) КОМП'ЮТЕРНА ФОРМА: (A) ТИП НОСІЯ: Гнучкий диск (B) КОМП'ЮТЕР: ІВМ-сумісний (C) ОПЕРАЦІЙН А СИСТЕМА: PC-DOS/MS-DOS (D) ПРОГРАМНЕ ЗАБЕЗПЕЧЕННЯ: Patentin Release №1.0, Версія №1.25 (ЕРО) (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:1: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 19п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВЮТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: РАТ1 (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ №:1: TGTCTCCGGA GAGGAGACC (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:2: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: РАТ2 (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ №:2 ССААС АТС АТ GCCATCCACC (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:3: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: MTL (Р) (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ №:3: AGGTGTCCAT GGTGCTC AAG (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:4: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: lap (I) (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ AC AATCGAAC CGC ACACGTC (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:5: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (А) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: р35 (35) (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ CCAGGTAGCA GTCGTTGTGT (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:6: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (vii) ОПОСЕРЕДКОВАНЕ ДЖЕРЕЛО: (В) КЛОН: dad-1 (D) (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ CCTTSTTTCC TTTGTTCACT (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:7: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ GGTCCTATAC GAAGCGTTAC (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:8: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ CCACGTAGC A GTCGTTGTGT (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:9: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (А) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ №:4: №:5 №:6 №:7: №:8: (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ CATGTCTGAC TTGCGATTGG (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:10: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 20п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ AC AATCGAAC CGC ACACGTC (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:11: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 18п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ііі) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ GGAGAGGCGG TGTTAGTT (2) ДАНІ ЩОДО ПОСЛІДОВНОСТІ №:12: (і) ВЛАСТИВОСТІ ПОСЛІДОВНОСТІ: (A) ДОВЖИНА: 18 п.н. (B) ТИП: нуклеїнова кислота (C) ЛАНЦЮГОВІСТЬ: одинична (D) ТОПОЛОГІЯ: лінійна (іі) ТИП МОЛЕКУЛИ: ДНК (геномна) (ІІІ) ГІПОТЕТИЧНІСТЬ: НЕМАЄ (ііі) АНТИЗМІСТОВНІСТЬ: НЕМАЄ (хі) ОПИС ПОСЛІДОВНОСТІ: ПОСЛІДОВНІСТЬ CATC ATCGC A TTCGAGAG №:9: №:10: №:11: №:12
ДивитисяДодаткова інформація
Назва патенту англійськоюA method for transformation of plants
Назва патенту російськоюСпособ трансформации растений
МПК / Мітки
МПК: C07K 14/01, C12N 5/10, C12N 15/82, A01H 1/00, C12N 15/09, C07K 14/415
Мітки: трансформації, рослин, калюсу, лінія, ембріогенного, клітин, спосіб
Код посилання
<a href="https://ua.patents.su/18-72443-sposib-transformaci-roslin-i-liniya-klitin-embriogennogo-kalyusu-zea-mays.html" target="_blank" rel="follow" title="База патентів України">Спосіб трансформації рослин і лінія клітин ембріогенного калюсу zea mays</a>
Спосіб оцінки реакції бластної трансформації лімфоцитів у культурі клітин

Номер патенту: 48845
Опубліковано: 15.08.2002
Автори: Копійка Віра Вікторівна, Фролов Олександр Кирилович, Федотов Євген Рудольфович
МПК: C12N 5/08, C12R 1/91, G01N 33/48, G01N 21/00
Мітки: культури, лімфоцитів, спосіб, клітин, реакції, бластної, трансформації, оцінки
Формула / Реферат:
Спосіб оцінки реакції бластної трансформації лімфоцитів (РБТЛ) у культурі клітин, що включає забір крові, вилучення лімфоцитів, постановку культури, розподіл отриманої культури на два зразки, стимулювання одного зразка антигенами або мітогенами, інкубацію зразків, визначення ступеня РБТЛ, який відрізняється тим, що інкубацію проводять протягом доби, готують адгезивні відбитки з наступною їх фіксацією, флуорохромуванням акридиновим оранжевим,...
Лінія перещеплюваних клітин тестикулів поросят для вірусологічних досліджень

Номер патенту: 40865
Опубліковано: 15.08.2001
Автори: Піотрович Віталій Анатолійович, Жуковський Анатолій Миколайович, Кучерявенко Олексій Олександрович
МПК: C12N 5/06
Мітки: тестикулів, досліджень, поросят, вірусологічних, лінія, клітин, перещеплюваних
Формула / Реферат:
Лінія перещеплюваних клітин тестикулів поросят №103 (Депозитарій Державного наукового-контрольного інституту біотехнології і штамів мікроорганізмів) для вірусологічних досліджень.
Кодуюча послідовність днк гена гідроксифенілпіруватдіоксигенази, химерний ген, вектор, рослинна клітина, рослина, які містять таку послідовність, спосіб трансформації рослин, спосіб селективної гербіцидної обро

Номер патенту: 72422
Опубліковано: 15.03.2005
Автори: Ролланд Анне, Саілланд Алайн, Паллетт Кен, Матріндж Мішель
МПК: C12N 15/53, C12N 15/09, C12N 5/10, C12N 15/82, C12N 9/02, A01H 5/00, C12N 15/00, C12N 1/21
Мітки: спосіб, рослин, рослина, химерний, обро, таку, гена, рослинна, ген, селективної, вектор, гербіцидної, містять, кодуюча, гідроксифенілпіруватдіоксигенази, трансформації, послідовність, днк, клітина
Формула / Реферат:
1. Кодуюча послідовність гена гідроксифенілпіруватдіоксигенази (ГФПД), яка відповідає послідовностям SEQ ID №1, SEQ ID №2 або SEQ ID №3.2. Кодуюча послідовність за п. 1, яка відрізняється тим, що вона походить із Pseudomonas sp.3. Кодуюча послідовність за п. 2, яка відрізняється тим, що вона походить із Pseudomonas fluorescens. 4. Кодуюча послідовність за п. 1, яка відрізняється тим, що вона походить із...
Спосіб одержання та генетичної трансформації рослин beta vulgaris

Номер патенту: 54369
Опубліковано: 17.03.2003
Автори: Холл Роберт Девід, Колійн-Хоойманс Катеріна Марія, Кренс Франціскус Андріес, Верхоевен Хендрікус Адріанус, Вейєнс Гай, Данвел Джеймс Мартін
МПК: C12N 15/09, A01H 4/00, C12N 5/10
Мітки: рослин, спосіб, vulgaris, трансформації, генетично, одержання
Формула / Реферат:
1. Способ получения растения Beta vulgaris, включающий культивирование устьичных клеток Beta vulgaris в регенерационной среде и регенерацию целого растения из указанных клеток.2. Способ по п. 1, в котором растение является сахарной свеклой.3. Способ по любому из предшествующих пунктов, в котором устьичную клетку подвергают регенерации в интактном органе.4. Способ по любому из пунктов 1 или 2, в котором указанную устьичную...
Спосіб одержання клітин різних типів біологічних тканин зі стромальних клітин кісткового мозку ссавців

Номер патенту: 67602
Опубліковано: 15.06.2004
Автори: Щегельська Олена Анатоліївна, Кульшин Володимир Євгенович, Микулинський Юрій Юхимович, Омельченко Олена Анатоліївна
МПК: C12N 5/08, A61K 35/28
Мітки: тканин, мозку, біологічних, різних, спосіб, одержання, стромальних, ссавців, клітин, кісткового, типів
Формула / Реферат:
Спосіб одержання клітин різних типів біологічних тканин з стромальних клітин кісткового мозку ссавців шляхом культивування в специфічних кондиційних середовищах з додаванням у середовище специфічних індукторів, який відрізняється тим, що як специфічні індуктори використовують фактори, що виділяють з відповідних біологічним тканинам клітин ембріонального походження.
Попередній патент: Спосіб і пристрій для пейджерного виклику радіотерміналу у системі радіозв’язку
Наступний патент: Спосіб обмеження водоприпливу у газових та газоконденсатних свердловинах
Випадковий патент: Мобільний телефон з функцією сповіщення