Стійка до грибків трансгенна рослина solanaceae
Номер патенту: 102522
Опубліковано: 25.07.2013
Автори: Мюскенс Марина Воутера Марія, Аллефс Йозефус Якобс Хенрікус Марія, ван дер Воссен Едвін Андріс Герард







Формула / Реферат
1. Трансгенна рослина або трансгенна тканина, або їх частина, що є представником родини Solanaceae, яка включає рослинну клітину, яка включає:
(i) полінуклеотид, що кодує білок Rpi-blb2, який включає молекулу нуклеїнової кислоти, вибрану з групи, яка складається з:
(а) молекули нуклеїнової кислоти, що кодує принаймні зрілу форму поліпептиду, представленого у SEQ ID NO: 2 або 4;
(b) молекули нуклеїнової кислоти, яка включає кодувальну послідовність, як представлено у SEQ ID NO: 1 або 3, або 5, або 6, що кодує принаймні зрілу форму поліпептиду;
(с) молекули нуклеїнової кислоти, нуклеотидна послідовність яких є результатом виродження генетичного коду стосовно нуклеотидної послідовності (а) або (b);
(d) молекули нуклеїнової кислоти, що кодує поліпептид, який одержаний з поліпептиду, що кодується полінуклеотидом згідно з будь-яким з підпунктів від (а) до (с), шляхом заміни, делеції та/або додання однієї або декількох амінокислот амінокислотної послідовності поліпептиду, що кодується полінуклеотидом згідно з будь-яким з підпунктів від (а) до (с);
(е) молекули нуклеїнової кислоти, що кодує поліпептид, послідовність якого має ідентичність 85% або більше до амінокислотної послідовності поліпептиду, який кодується молекулою нуклеїнової кислоти згідно з (а) або (b);
(f) молекули нуклеїнової кислоти, що включає фрагмент або частину поліпептиду, яка несе епітоп, що кодується молекулою нуклеїнової кислоти згідно з будь-яким з підпунктів від (а) до (е);
(g) молекули нуклеїнової кислоти, що включає полінуклеотид, який має послідовність молекули нуклеїнової кислоти, що є ампліфікованою з бібліотеки нуклеїнових кислот при використанні праймера, як представлено у Таблиці 3b;
(j) молекули нуклеїнової кислоти, що кодує поліпептид, який впізнається моноклональним антитілом, що утворене проти поліпептиду, який кодується молекулою нуклеїнової кислоти згідно з будь-яким з підпунктів (а) - (g);
(k) молекули нуклеїнової кислоти, одержуваної шляхом скринінгу прийнятної бібліотеки при жорстких умовах з зондом, який має послідовність молекули нуклеїнової кислоти згідно з будь-яким з підпунктів (а) - (j); та
(l) молекули нуклеїнової кислоти, комплементарний ланцюг якої гібридизується у жорстких умовах з молекулою нуклеїнової кислоти згідно з будь-яким з підпунктів (а) або (k);
або комплементарного ланцюга будь-якої з молекул нуклеїнової кислоти згідно з будь-яким з підпунктів (а) - (l);
або такого, що кодує поліпептид, який кодується сегментом хромосоми або групою зчеплення 6 Solanum bulbocastanum або Solanum tuberosum, що косегрегує з маркером, вибраним з Таблиці 3а або 3b, або включає сайт реплікації або сайт гібридизації для вказаного маркера, та який опосередковує резистентність до патогену типу Oomycetes;
або
(іі) вектор, що містить полінуклеотид, визначений в (i), або включає полінуклеотид, визначений в (i), та додатковий ген резистентності, або є одержаним за допомогою інсерцію вказаного полінуклеотиду, визначеного в (i), у вектор або інсерцію вказаного полінуклеотиду, визначеного в (i), та додаткового білка резистентності;
або
вказані трансгенна рослина або трансгенна тканина, або їх частина, що є отримуваними введенням полінуклеотиду, визначеного в (i), або цього полінуклеотиду та полінуклеотиду, який кодує додатковий білок резистентності, або вектора, визначеного в (ii), у геном вказаної рослини, рослинної клітини або рослинної тканини, або у їх частину.
2. Трансгенна рослина або трансгенна тканина, або їх частина згідно з пунктом 1, в якій у полінуклеотидi, визначеному в (i), маркер являє собою Е40М58 СТ119 або СТ216.
3. Трансгенна рослина або трансгенна тканина, або їх частина згідно з пунктом 1 або 2, в якій полінуклеотид, визначений в (i), являє собою ДНК або РНК.
4. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-3, в якій у векторі, визначеному в (ii), полінуклеотид, що кодує білок Rpi-blb2, або такий, що кодує додатковий білок резистентності, є оперативно зв'язаним з послідовностями контролю експресії, та/або є оперативно зв'язаним з послідовністю нуклеїнової кислоти, яка кодує трансгенний сигнал регуляції експресії, що дозволяє здійснювати експресію в прокаріотичних або еукаріотичних хазяйських клітинах.
5. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-4, в якій у векторі, визначеному в (ii), полінуклеотид, що кодує білок Rpi-blb2, або такий, що кодує додатковий білок резистентності, є оперативно зв'язаним з послідовностями контролю експресії, що мають походження від тих самих видів, що й полінуклеотид, який кодує білок Rpi-blb2, або такий, що кодує додатковий білок резистентності.
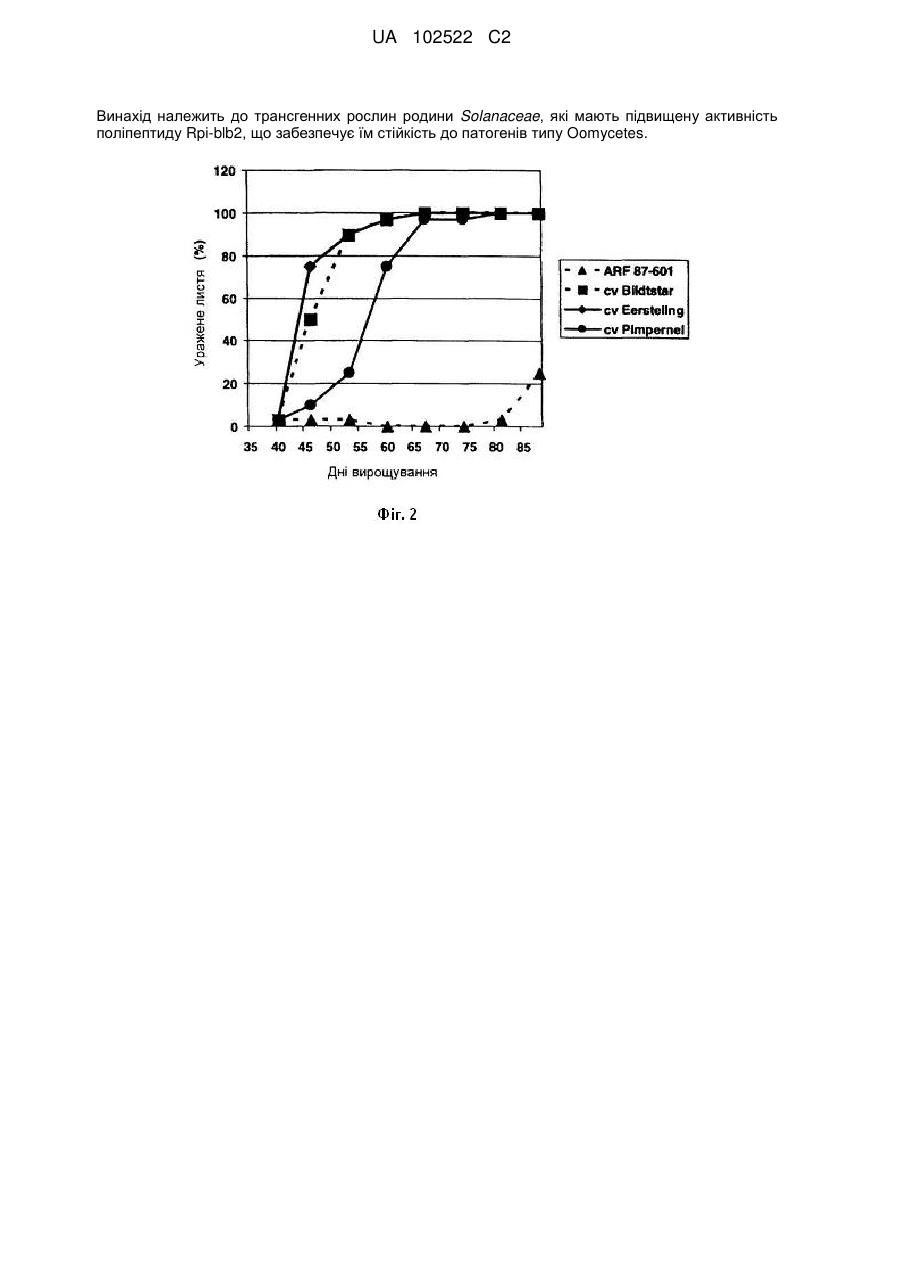
6. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-4, яка при наявності полінуклеотиду, визначеного в (i), або вектора, визначеного в (ii), є резистентною до рослинного патогену типу Оomycetes.
7. Трансгенна рослина або трансгенна тканина, або їх частина згідно з пунктом 6, в якій рослинний патоген являє собою патоген порядку Pythiales або Peronosperales.
8. Трансгенна рослина або трансгенна тканина, або їх частина згідно з пунктом 6 або 7, в якій рослинний патоген являє собою патоген видів P. infestans, Phytophthora erythroseptica, Phytophthora capsici, Phytophthora sojae, Phytophthora parasitica var. nicotianae, Bremia lactuca, Peronospera tabaci або Plasmopara viticola.
9. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-8, в якій білок резистентності характеризується Р-петлею та доменом NBS.
10. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-9, в якій додатковий ген резистентності являє собою ген, який кодує Rpi-blb, R1, R-ber, Rpi1, R2, R3, R4, R5, R6, R7, R8, R9, R10, R11, Ph-1, Ph-2 та/або Ph-3.
11. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-10, в якій додатковий білок резистентності являє собою білок Rpi-blb.
12. Трансгенна рослина або трансгенна тканина, або їх частина згідно з будь-яким з пунктів 1-11, яка походить від родини Solanaceae, переважно S. bulbocastanum, картоплі (S. tuberosum), томатів (S. lycopersicum або Lycopersicon lycopersicum (L.) Karsten ex Farwell), петунії, цифомандри томатне дерево (S. betaceum), динного дерева (S. muricatum) або баклажанів (S. melongena).
13. Матеріал для розмноження трансгенної рослини або рослинної тканини згідно з будь-яким з пунктів 1-12.
Текст
Реферат: UA 102522 C2 (12) UA 102522 C2 Винахід належить до трансгенних рослин родини Solanaceae, які мають підвищену активність поліпептиду Rpi-blb2, що забезпечує їм стійкість до патогенів типу Oomycetes. UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Даний винахід належить до нового способу підвищення стійкості рослини, зокрема Solanaceae, переважно картоплі та томатів, до рослинних патогенів типу Oomycetes, що включає підвищення активності поліпептиду згідно з даним винаходом. Винахід також належить до полінуклеотидів та векторів, що включають такі полінуклеотиди. Винахід також стосується відповідних векторів, клітин, трансгенних рослин та трансгенного матеріалу для розмноження, який походить з них, способів їх одержання та їх застосування для одержання продуктів харчування, кормів, насіння, фармацевтичних препаратів або чистих хімічних речовин. Метою рослинної біотехнологічної роботи є покоління рослин, що має сприятливі нові властивості, наприклад для підвищення сільськогосподарської продуктивності, підвищення якості у випадку продуктів харчування, або для одержання специфічних чистих хімічних речовин або фармацевтичних препаратів (Dunwell JM (2000) J Exp Bot 51 Spec No: 487-96). Механізми природного захисту рослин проти патогенів часто бувають неефективними. Тільки грибкові захворювання приводять до щорічних втрат врожаю, що складають багато більйонів доларів США. Вбудовування сторонніх генів з рослинних. тваринних або мікробних джерел може підвищити рівень цього захисту. Прикладами можуть слугувати захист тютюну від пошкодження, спричиненого поїданням комахами, шляхом експресії ендотоксинів Bacillus thuringiensis під контролем промотору 35S CaMV (Vaeck et al.(1987) Nature 328: 33-37) або захист тютюну від грибкової інфекції шляхом експресії хітинази бобових під контролем промотору CaMV (Broglie et al.(1991) Science 254: 1194-1197). Проте більшість з описаних підходів пропонують стійкість до одного патогену або вузького спектру патогенів. Незважаючи на відоме стихійне лихо, що сталося з ірландською картоплею у середині 19ого століття, фітофтороз ще й зараз продовжує залишатися одним з найбільш спустошливих серед усіх захворювань культурних рослин. Фітофтороз спричиняється грибком з групи ооміцетів Phytophthora infestans, спеціалізованим патогеном, що головним чином викликає захворювання листя та плодів у межах видів Solanaceae, зокрема, у картоплі і томатів. Ці грибки вперше були виявлені у Мексиці і тому з цих причин Мексика вважається центром походження цих грибків. Обидва схрещуваних типа А1 та А2 є постійно присутніми, наприклад, районі Толука. P. infestans також виявлено на природних видах Solanum у віддалених районах Мексики. Крім того, у Мексиці виявлено багато видів Solanum, що забезпечують одержання бульб та мають високу ступінь резистентності до фітофтори. Переважні способи для запобігання загибелі культур або для зниження втрат врожаю передбачають застосування фунгіцидів, які запобігають або лікують інфекцію, спричинену P. infestans. Замість широкого застосування хімічних пестицидів може бути бажаним альтернативний підхід до боротьби з фітофторозом. Для одержання стійкості до фітофторозу селекціонери раніше концентрували свої зусилля на інтрогресії домінантних генів R, одержаних з Solanum demissum, місцевого дикого виду картоплі, що характерний для Мексики. Було ідентифіковано одинадцять таких генів R, деякі з яких було картовано до специфічних локусів на генетичній карті картоплі (розглянуто у Gebhardt and Valkonen, 2001), а нещодавно було клоновано ген R1. R1 та R2 розташовані у хромосомах 5 та 4, відповідно. R3, R6 та R7 розташовані у хромосомі 11. Невідомі гени R, що надають стійкості сорту до фітофтори, було також описано у S. tuberosum ssp. andigena, S. berthaultii та S. pinnatisectum. Стійкість, що індукується цими R-генами, була (майже) повною, але, як виявилося, була нетривалою у будь-якому випадку. Завдяки високому ступеню резистентності та простоті перенесення, багато культиварів мають стійкість, яка має походження від S. demissum. На жаль, специфічна для сорту стійкість, що має походження від S. demissum, хоча і є майже повною, не є тривалою. Оскільки виведені колись нові культивари картоплі вирощувалися у значних масштабах на великих площах, у P. infestans виникли нові вірулентності, які зробили патоген здатним долати інтрогресовану стійкість. Більш тривалу польову резистентність до фітофтори, що часто є кількісною по своїй природі і, як передбачається, не є сортоспецифічною, можна знайти в деяких мексиканських видах Solanum та центральноамериканських і південноамериканських видах. Проте цей тип стійкості є складним для перенесення у культивари картоплі шляхом схрещування та селекції за фенотипічними ознаками. Диплоїдний S. bulbocastanum з Мексики та Гватемали є одним із видів коренеплодів, який давно відомий своєю високою резистентністю до фітофторозу. Однак, класична передача резистентності від диких видів Solanum до культивованої картоплі часто не вдається із-за різниці в плоїдності та значенні ендоспермного балансу (Endosperm Balance Number) (EBN). Незважаючи на ці проблеми, інтрогресія ознаки резистентності S. bulbocastanum була успішною. Нещодавно, було виявлено, що соматичні гібриди S. bulbocastanum та S. tuberosum та обернено схрещені є високо резистентними до фітофторозу, навіть за значного натиску хвороби (Helgeson et 1998). Незважаючи на повідомлення про пригнічення рекомбінації, 1 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 резистентність в обернено схрещеному матеріалі виявляється в 8-ій хромосомі в межах приблизно 6 сМ від RFLP-маркерів СР53 та СТ64. CAPS-маркер, що походить із томатного RFLP-зонду СТ88 відокремлюється разом із резистентністю. Таким чином, останніми роками розробка рослинної резистентності до патогенів типу Oomyceta просувається вперед. Однак, 40 інтенсивних та безперервних досліджень та селекції так і не призвели до виходу на ринок резистентних культиварів. Переважна маса генів, ідентифікованих останніми роками, надає лише расовоспецифічну резистентність. Більше того, досягнута резистентність не є стійкою. До того ж, застосування засобів захисту рослин є загальновизнаним навантаженням на навколишнє середовище. Отже, в деяких західних країнах законодавство стало більше обмежувати, а деколи, частково забороняти застосування певних фунгіцидів, що робить хімічну боротьбу із цим захворюванням більш складною. До того ж, хімічна боротьба є більш дорогою. На завершення, наступним обмеженням є розвинення у грибків резистентності до специфічних фунгіцидів, таких як металаксил, про що повідомлялося з багатьох країн світу. Відповідно, проблема, що лежить в основі даного винаходу - це створення нового засобу та способів для ефективного захисту рослин від фітофторозу та подібних захворювань. Розв'язання технічної проблеми досягнуто шляхом створення рішень, описаних у формулі. Таким чином, даний винахід стосується методу створення або підвищення резистентності рослин до рослинного патогену типу Oomycetes, що включає підвищення активності протеїну Rpi-blb2 в рослині або тканині, органі або клітині рослини, або її частині. Rpi-blb2 є типом LZ-NBS-LRR гену R та має гомологію послідовності до гену томатів Мі-1, який надає резистентності до трьох видів нематод, що утворюють кореневі нарости (Meloidogyne spp.), а також до попелиці картоплі Macrosiphum euphorbiae (Vos et 1998; Rossi et 1998; Milligan et 1998), також до В- та Q-біотипів білої мушки Bemisia tabaci (Nombela et 2003). Як було відкрито для Rpi-blb, Rpi-blb2 також забезпечує повну резистентність до усіх P. infestans ізолятів, що несуть багато факторів вірулентності, та расова специфічність не була доведена. Термін "Rpi-blb2" стосується полінуклеотиду, що кодує поліпептид, який має згадану тут активність протеїну або поліпептид, який має згадану активність протеїну Rpi-blb2. Чи стосується наступний термін "Rpi-blb2" поліпептиду або полінуклеотиду буде зрозуміло з контексту, де його буде використано. Терміном "створення" або "збільшення", або "стимулювання" "резистентності рослини" вказується на те, що резистентність рослини або її частини збільшена або створена або стимульована в порівнянні із згадуваною. "Надання", "виникнення", "створення", "стимулювання" або "збільшення" резистентності до патогену, означає, що захисні механізми певних видів рослин або сортів є в більшій мірі резистентні до одного або більше патогенів завдяки використанню способу згідно з винаходом, у порівнянні з диким типом рослини, до якого спосіб згідно з даним винаходом не застосовувався, при інших ідентичних умовах (наприклад, таких, як кліматичні умови, умови росту, види патогенів та подібні). Збільшена резистентність проявляється переважно в зниженні проявів симптомів захворювання, симптоми захворювання включать, на додачу до вищезгаданих шкідливих ефектів, наприклад, також ефективність проникнення патогену в рослину або рослинні клітини, або проліферацію в, або на ній. В цьому контексті, симптоми захворювання переважно знижені принаймні на 10 % або принаймні на 20 %, особливо бажано, принаймні на 40 % або 60 %, надзвичайно особливо бажано, принаймні на 70 % або 80 % та найбільш бажано, принаймні на 90 % або 95 %. Під терміном "збільшений", як його тут вжито, розуміється, що активність генного продукту є вищою ніж у вихідного. Отже, термін "збільшений" включає той факт, що активність, наприклад, активність генного продукту Rpi-blb2 або іншого генного продукту, створено de novo, якщо ця активність, наприклад, описана тут активність Rpi-blb2, не була виявлена в вихідному. Термін "збільшений" також стосується стимуляції активності генного продукту. Підвищена експресія гену, тобто його активація, може бути стимульована кількома шляхами, наприклад, застосуванням хімічного або біотичного стресу організму. Наприклад, резистентність до інфікуючих паразитів, опосередкована геном, може бути активована інфікуванням паразитом, наприклад, P. infestans, та полягати в збільшеній резистентності до цього та/або інших патогенів. Отже, в подальшому, термін "збільшений" також включає терміни "стимульований" та "створений". "Патогенна резистентність" вказує на зниження або ослаблення симптомів хвороби у рослини внаслідок інфікування патогеном. Симптоми можуть бути численними, але переважно охоплювати ті, що прямо або опосередковано мають небажаний вплив на якість рослин, якість 2 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 врожаю, придатність для використання як кормової добавки або харчової добавки, або які також роблять посів, вирощування або обробляння культури ускладненими. "Патоген" в межах даного винаходу, означає за допомогою прикладів але не обмежуючись ними, віруси або віроїди, бактерії, гриби, паразити тварин такі, як, наприклад, комахи та нематоди. Термін "протеїн Rpi-blb2" вказує на протеїн або поліпептид, експресія якого в рослині або її частині надає резистентність рослинам або частинам рослин до одного з описаних тут патогенів порівняно з нерезистентним штамом. Рослина або тканина, орган або клітина рослини або її частина, що мають підвищену активність протеїну Rpi-bib2 є менш сприйнятливими до інфікування патогеном, зокрема, патогеном типу Oomycetes, переважно P.infestans, ніж рослина або її частина, яка має ідентичну генетичну основу але не має генетичного елементу, необхідного для забезпечення експресії Rpi-blb2 (зазначених тут як "дикий тип" або "порівнюваний"). Тести для перевірки резистентності рослини або її частини є добре відомими кваліфікованим фахівцям. Резистентність до P. infestans може бути визначена як індекс споруляції згідно Flier, 2001. Flier описує індекс 2 2 споруляції як рівень споруляції на 1 cm . Таким чином, зниження споруляції на 1 cm на 20 % порівняно із диким типом тут визначається як резистентність. В прикладах, що ілюструють даний винахід, індекс споруляції був визначений як рівень споруляції на ушкодження. Таким чином, термін "резистентність" також може бути визначений як 20 % зниження споруляції на ушкодження, порівняно із диким типом. Останнє визначення є переважним. В переважних реалізаціях, споруляція в досліді знижена, переважно, на 30 %, більш бажано, знижена на 50 %, значно більш бажано, на 70 %, ще бажаніше, більш ніж на 80 %, ще більш бажано, на 85 % та 90 %. Найбільш бажаним є зниження на 95 % або більше. Згідно з цим, в даному винаході "активність" протеїну Rpi-blb2 означає, що експресія протеїну надає згадане зниження індексу споруляції. Також спостерігалося, що типовою відповіддю для рослин, що містили Rpi-blb2, на інфікування P. infestans є наявність малих ушкоджень, без будь-яких чітких споруляцій, наприкінці періоду росту. Таким чином, в одній з реалізацій, активність Rpi-blb2 визначена як наявність малих ушкоджень, без будь-яких чітких споруляцій, як описано в експериментах. Резистентність Rpi-blb2 проявляється в некротичних місцях, які мають низький рівень споруляції. Експеримент, проведений на відокремлених листках, демонструє активність Rpi-blb2. Експеримент описано в прикладі 17 та на фігурі 18. Різниця між Rpi-blb2 та іншими генами резистентності до P. infestans полягає в тому, що створюється низький рівень споруляції (Фігура 18). Дослід з відокремленим листом, в якому ушкодження присутнє на Rpi-blb2-генотипі (ARD 92-1197-16) демонструє низький рівень спорангіїв, у порівнянні з повною відсутністю спорангіїв на генотипі, що містить ген R2 S. demissum. Індекс споруляції становить лише 1,1 % від сприйнятливого фенотипу (cv.Bintje) (Таблиця 7 та Фігура 18). Польові експерименти також показали, що Rpi-blb2 призводить до низького рівня інфікування. Симптоми фітофторозу розвинулися протягом вегетаційного періоду на низькому рівні (Фігура 3, ARF87-801) або наприкінці вегетаційного періоду (Фігура 2, ARF87-601; Фігура 3, ARF87-507 та ARF87-601). Таким чином, в одній з реалізацій, активність Rpi-blb2 також визначена, як одержана після експресії в некротичних регіонах рослин, які мали низький рівень споруляцій, як описано в експериментах. Таким чином, в одній з реалізацій, спосіб даного винаходу створює рослини, які мають некротичні регіони з низьким рівнем споруляцій або менше. Термін "вихідний" стосується організму або його частини, наприклад, клітини, який є суттєво, на скільки це можливо, подібним у геномі, протеомі та/або метаболомі до релевантного організму або його частини, наприклад, клітини, наприклад, до рослини даного винаходу. Таким чином, термін "вихідний" стосується прикладу організму або його частини, наприклад, клітини, який суттєво генетично, протеомно та/або метаболічно ідентичний до організму даного винаходу або його частини, але в якому не спостерігається активність генетичного продукту, наприклад, Rpi-bib2, що і є основною відмінністю порівнюваного геному, протеому та/або метаболому. Таким чином, порівнюваною може бути рослина або її частина, яка не експресує або експресує занадто мало діючого генетичного продукту. наприклад, вона не кодує Rpi-blb2 або не транскрибує ген, що кодує Rpi-blb2 або не транслює активну Rpi-blb2 мРНК. Таким чином, порівнюваний зразок не пропонує модифікацію, яка створює активний генетичний продукт в достатній кількості для прояву у фенотипі як описано. Чи є дві рослини в основному генетично ідентичні, може бути досліджено методами, відомими досвідченим фахівцям, наприклад, за допомогою аналізу "fingerprint", наприклад, як описано в Roldan-Ruiz, Theor. Appl. 3 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Genet., 2001, 1138-1150. Характер експресії протеїнів може бути досліджено як описано в літературі, наприклад, за допомогою гель-електрофорезу (1D, 2D, 3D), мас-спектральним аналізом та іншими методами наприклад, як описано в www.waters.com, www.proteine.orq.au, www.proteomesci.com, www.sdu.dk/Nat/CPA. Метаболом може бути досліджений фахівцем, як описано в літературі, наприклад, за допомогою HPLC, GC, OPLC, LC-MS, GC-MS, LC-MS-MS, та іншими методами як описано, наприклад, в www.metabolic-explorer.com, www.ki.se/icsb2002/pdf/ICSB_209.pdf, www.qenomics.ruq.nl/technologies.htm, Fiehn etal., Nature Biotech, 18 (2000), 1157, Raamsdonk et al., Nature Biotech, 19 (2991), 45-50, Buchholz, Anal. Biochem, 295 (2001) 129-137, Soga et al., Anal Chem.74 (2002) 2233-2239. Для підвищення резистентності до патогену, вихідний організм або його частина є сприйнятливими до інфікування патогеном, наприклад, патогеном рослин, наприклад, Р. infestans. Переважно, вихідним є клон того організму, в який було введено, наприклад, відповідний полінуклеотид, наприклад, полінуклеотид винаходу, або активатор, наприклад, активатор відповідного генного продукту, що опосередковує активність, наприклад, активатор, що підвищує експресію відповідного полінуклеотиду або похідного згаданого полінуклеотиду, або активатор відповідного поліпептиду, наприклад, поліпептиду даного винаходу, та/або відповідний вектор, що кодує відповідний генетичний продукт. Наприклад, переважним вихідним для способу даного винаходу є організм або його частина, який є клоном організму або його частини, наприклад, клітини, яка була трансфікована або трансформована полінуклеотидом або вектором даного винаходу. Якщо клон, як описано, не може бути ідентифікований, то можна розщепити, вибити або вимкнути відомим способом ті елементи, які в основному опосередковують відповідну активність, наприклад, опосередковуючи підвищення активності Rpi-blb2, наприклад, підвищуючи експресію в організмі, наприклад, рослині. Фахівцеві добре відомо, як знизити або пригнітити відповідний генетичний продукт, наприклад, зниженням або пригніченням експресії, наприклад, Rpi-blb2. Такий клон далі може бути порівняний із організмом, одержаним згідно зі способом даного винаходу, наприклад, генотипом, що експресує Rpi-blb2, резистентним до P. infestans. Термін "рослина", як його використано тут, належить до усіх родів та видів вищих та нижчих рослин царства рослин. Термін включає дорослі рослини, насіння, пагони та саджанці, та їхні похідні частини, розмножувальний матеріал, рослинні органи, тканини, протопласти, калюс та інші культури, наприклад, клітинні культури, та будь-які інші типи угрупування рослинних клітин, які дають функціональні та структурні одиниці. "Дорослі рослини" належить до рослин на будьякій бажаній стадії розвитку поза саджанцем Саджанець належить до молодої незрілої рослини на ранній стадії розвитку. "Рослина" охоплює усі однорічні та багаторічні однодольні та дводольні рослини. Переважними для даного винаходу є рослини, які використовуються як продукти харчування або корми, наприклад, роди однодольних або дводольних, зокрема види, такі як описані вище, наприклад, види злакових рослин, або представники родини Solanaceae, найбільш бажано, картопля або томати. Як є відомим для фахівця, спосіб згідно з даним винаходом включає подальшу селекцію тих рослин, в котрих, на відміну, або порівняно з вихідними рослинами, існує, або збільшена резистентність до принаймні одного згаданого патогену. "Селекція" стосовно рослин, в котрих, на відміну або порівняно із вихідною рослиною, існує або збільшена резистентність до принаймні одного патогену, означає усі методи, які є придатними для визначання існуючої або збільшеної резистентності до патогенів. Це можуть бути симптоми патогенної інфекції, але вони також можуть включати описані тут симптоми, які відноситися до якості рослин, якості врожаю, придатності до використання як кормової добавки або харчової добавки та подібне. Таким чином, в одній з реалізацій способу, згідно з даним винаходом, протеїн Rpi-blb2 кодується полінуклеотидом, який включає молекулу нуклеїнової кислоти, вибрану з групи, що складається з: a) молекул нуклеїнової кислоти, які кодують принаймні готову форму поліпептиду, наведеного в SEQ ID NO: 2 або 4; b) молекул нуклеїнової кислоти, які включають кодуючу послідовність, наведену в SEQ ID NO: 1 або 3, або 5 або 6, що кодують принаймні готову форму поліпептиду; c) молекул нуклеїнової кислоти, чия нуклеотидна послідовність внаслідок виродженості генетичного коду вироджена у нуклеотидну послідовність (а) або (b); d) молекул нуклеїнової кислоти, що кодують поліпептид, який походить з поліпептиду, кодованого полінуклеотидом (а) - (с) шляхом заміщення, делеції та/або додавання однієї або 4 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 більше амінокислот з(до) амінокислотної послідовності поліпептиду, кодованого полінуклеотидом від (а) до (с); e) молекул нуклеїнової кислоти, що кодує поліпептид, послідовність котрого має ідентичність 70 % або більше з амінокислотною послідовністю поліпептиду, кодованого молекулою нуклеїнової кислоти (а) або (b); f) молекул нуклеїнової кислоти, які включають фрагмент або епітоп-вмісну частину поліпептиду, кодованого будь-якою молекулою нуклеїнової кислоти від (а) до (e); g) молекул нуклеїнової кислоти, які включають полінуклеотид, який має послідовність молекули нуклеїнової кислоти, ампліфікованої з бібліотеки нуклеїнових кислот за допомогою праймерів як приведено в табл. 3b, зокрема ARF1F та ARF1R; h) молекул нуклеїнової кислоти, що кодують фрагмент, що починається амінокислотами: 1, 30, 50, 100, 200, 300, 500, або 1000 та закінчується амінокислотами: 1276, 1000, 500, 300, 200, 50, 30, або 1 поліпептиду, кодованого будь-якою з (а) до (g); і) молекул нуклеїнової кислоти, що включають принаймні 20 нуклеотидів будь-якого з полінуклеотидів з (а) або (d); j) молекул нуклеїнової кислоти, що кодують поліпептид, який розпізнається моноклональними антитілами, які одержано до поліпептиду, кодованого будь-якою з молекул нуклеїнової кислоти від (а) до (h); k) молекул нуклеїнової кислоти, одержуваних скринінгом відповідної бібліотеки за жорстких умов, за допомогою зонда, який має послідовність будь-якої з молекул нуклеїнової кислоти від (а) до (j), або її фрагменту із щонайменше 15, переважно 30, 60, 100 або більше нуклеотидів; та l) молекул нуклеїнової кислоти, комплементарна послідовність котрих гібридизується за жорстких умов із будь-якою з молекул нуклеїнової кислоти від (а) до (k); або комплементарного ланцюга будь-якої з молекул нуклеїнової кислоти згідно з (а) - (l); або ті, що експресують поліпептид, кодований сегментом або лінкерною групою 6 Solanum bulbocastanum, який виділяється сумісно із маркером з таблиці 3А та який опосередковує резистентність до патогенів, зокрема, до патогенів, вибраних з групи типу Oomycetes; В одному з втілень, полінуклеотид способу, згідно винаходу, не складається з послідовності, зображеної в Seq. ID NO.: 7 та/або 9 та/або не складається з послідовності нуклеїнової кислоти, що кодує протеїн, зображений в Seq. ID NO.: 8 та/або 10. В одному з втілень, полінуклеотид способу, згідно винаходу, не складається з послідовності молекули нуклеїнової кислоти Мi1.1 або Мi1.2, та/або молекули нуклеїнової кислоти, що кодує протеїн Мi1.1 або Мi1.2. Таким чином, в одному з втілень, полінуклеотид способу, згідно винаходу, може не складатися із послідовностей, приведених Rossi et al. 1998, PNAS USA 95:9750-9754, Milligan et al., 1998. Plant Cell 10:1307-1319; та/або WO98/06750. Порівняння послідовностей Rpi-blb2, Мi1.1 та Мi1.2 показано на фігурах з 15 по 17. Термін "лінкерна група", як його вжито тут, стосується двох або більше рис та/або локусів та/або генів та/або маркерів, які мають тенденцію бути успадкованими разом як результат сполучення між двома згаданими рисами та/або локусами та/або генами та/або маркерами. Чим ближчі риси та/або локуси та/або гени та/або маркери, тим менша ймовірність, що вони будуть розділені під час відновлення ДНК або процесу реплікації, такого як мітоз або мейоз у еукаріотів, й, таким чином, існує більша вірогідність, що вони будуть успадковані разом. Існує стільки ж лінкерних груп, скільки гомологічних пар у хромосомах. Термін "лінкерна група 6" стосується лінкерної групи картоплі або томатів, яка зв'язана із хромосомою 6, цей зв'язок було встановлено визначенням маркерів із відомим розташуванням в хромосомі, ґрунтуючись на роботі, опублікованій Bernatzky та Tanksley (1986) та Tanksley et al.(1992). Лінкерна група має той самий номер, що й хромосома, яка їй відповідає. В томатах, хромосоми нумеруються згідно з їхньою довжиною в пахітені. Ці номери були застосовані Barton (1950); хромосома 1 є найдовшою, хромосома 12 є найкоротшою. Також, щодо довжини, то такі характеристики як позиція центромерів, а також кількість та розподіл гетерохроматину слугують для ідентифікації кожної хромосоми. Коротке плече хромосоми позначається як "S", довге - як "L"; отже "1S" вказує на коротке плече хромосоми 1; як, наприклад, у Barton, D.W. (1950) American Journal of Botany. 37, 639-643, Bernatzky, R. and Tanksley, S.D. (1986) Genetics 112, 887-898, Tanksley, S.D., et al., (1992) Genetics 132, 1141-1160. Термін "сумісне виділення", як його вжито тут, стосується тенденції двох або більше рис та/або локусів та/або генів та/або маркерів бути успадкованими разом. Наприклад, більш конкретно, регіон хромосоми 6, який виділяється сумісно із Rpi-blb2 є коротким плечем, яке в томатах несе морфологічний маркер Мі. 5 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Також, одна з реалізацій способу згідно з даним винаходом, стосується способу даного винаходу, в якому протеїн Rpi-blb2 кодується полінуклеотидом даного винаходу, наприклад, є кодованим полінуклеотидом, наведеним в Seq. ID. 1 або 3 або 5 або 6 або його фрагментом. За допомогою пошуку BLASTX були визначені гени із найбільшою гомологією до послідовностей Rpi-blb2, ними були гени та протеїни МI1.1 та МI1.2; дивись фігури з 15 по 17. Обидва гени, що мали високу подібність до послідовності, приведеної в Seq ID NO.: 1 або 3 або 5 або 6, але не надавали рослинам резистентності до патогенів типу Oomycetes. От чому активність Мi1.1 та Мi1.2 є іншою активністю, ніж активність поліпептиду даного винаходу. Послідовність ORF Мi1.1 та Мi1.2 та кодованих протеїнів приведено в Seq. ID NO.: з 7 по 10. Також заявка ЕР401764.4 стосується генів Мі. Раніше відома послідовність генів Мi1.1 та МI1.2 виключена з полінуклеотидів даного винаходу, зокрема Seq ID NO.: 7 та 9 виключені. Також виключеним може бути полінуклеотид, який може кодувати поліпептид Seq. ID NO.: 8 або 10, Таким чином, з реалізації також виключені послідовності, що кодують протеїни Мi1.1 та Мi1.2. Протеїни із нижчою гомологією до поліпептиду, кодованого полінуклеотидом даного винаходу є протеїни Него Resistance 1 та 2 (Genbank AccNo.: gi26190252 та gi26190254), Протеїни А, В, С, D та Е резистентності до Tospovirus [Genbank AccNos.:gi15418709, gi15418710, gi15418712, gi15418713, gi15418714]; R1 [Genbank AccNo.: gi17432423] та Prf [Genbank AccNo.: gi8547237] послідовності або кодуючі послідовності котрих також виключені із послідовностей даного винаходу. Терміни "ген(и)", "полінуклеотид", "послідовність нуклеїнової кислоти", "нуклеотидна послідовність", або "молекула(и) нуклеїнової кислоти", як вони вжиті тут, відносяться до полімерних форм нуклеотидів будь-якої довжини, та рибонуклеотидів або дезоксирибонуклеотидів. Терміни стосуються лише первинної структури молекули. Таким чином, цей термін включає дволанцюгові та одноланцюгові ДНК, та РНК. Він також включає відомі типи модифікацій, наприклад, метилування, "caps''-заміщення одного або більшої кількості природних нуклеотидів аналогами. Переважно, послідовність ДНК згідно даного винаходу включає кодуючу послідовність, що кодує визначений тут поліпептид. "Кодуюча послідовність" є нуклеотидною послідовністю, яка транскрибована в мРНК та або трансльована в поліпептид, коли поміщена під керування відповідних регуляторних послідовностей. Межі кодуючої послідовності визначені старт-кодоном трансляції на 5'-кінці та стоп-кодоном трансляції на 3'-кінці. Кодуюча послідовність може включати, але не обмежуючись ними, мРНК, кДНК, рекомбінантні нуклеотидні послідовності або геномну ДНК, причому також можуть бути присутні в певних випадках інтрони. За допомогою "гібридизації", означає, що такі молекули нуклеїнової кислоти гібридизуються за звичайних умов гібридизації, переважно за жорстких умов, як це описано, наприклад, в Sambrook (Molecular Cloning; A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989)). Прикладом одних з таких жорстких умов гібридизації є гібридизація в 4XSSC при 65 °C, із наступним відмиванням в 0.1XSSC при 65 °C протягом однієї години. Іншими, характерними жорсткими умовами є гібридизація в 50 % формаміді, 4XSSC при 42 °C. Також, умови на етапі відмивання можуть бути вибрані серед умов, обмежених умовами низької жорсткості (приблизно 2Х SSC при 50 °C) та умовами високої жорсткості (приблизно 0.2Х SSC при 50 °C, переважно при 65 °C) (20Х SSC: 0.3M цитрату натрію, 3М NaCl, pH 7.0). Також, температура на етапі відмивання може зростати від умов низької жорсткості при кімнатній температурі, приблизно при 22 °C, до умов високої жорсткості, приблизно при 65 °C. Обидва параметри, концентрація солі та температура, можуть бути змінені одночасно, або один з двох параметрів може підтримуватися постійним, коли тільки інший змінюється. Денатуруючі засоби, наприклад, формамід або SDS, також можуть використовуватися під час гібридизації. В присутності 50 % формаміду, гібридизація переважно відбувається при 42 °C. Деякі наступні приклади умов для гібридизації та етапу відмивання приведені нижче: (1) Умови гібридизації можуть бути вибрані серед, наприклад, наступних умов: a) 4Х SSC при 65 °C, b) 6X SSC при 45 °C, c) 6Х SSC, 100 mg/ml денатурованої фрагментованої ДНК сперми риби при 68 °C, d) 6X SSC, 0.5 % SDS, 100 mg/ml денатурованої ДНК сперми лосося при 68 °C, e) 6Х SSC, 0.5 % SDS, 100 mg/ml денатурованої фрагментованої ДНК сперми лосося, 50 % формамід при 42 °C, f) 50 % формамід, 4Х SSC при 42 °C, g) 50 % (vol/vol) формамід, 0.1 % коров'ячий сироватковий альбумін, 0.1 % Ficoll, 0.1 % полівінілпіролідон, 50 тМ натрієво-фосфатний буфер рН 6.5, 750 mM NaCl, 75 тМ цитрат натрію при 42 °C, 6 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 h) 2Х або 4Х SSC при 50 °C (умови низької жорсткості), або і) 30 to 40 % формамід, 2Х або 4Х SSC при 42 °C (умови низької жорсткості). (2) Етапи відмивання можуть бути вибрані серед, наприклад, наступних умов: а) 0.015 М NaCl/0.0015 М цитрат натрію/0.1 % SDS при 50 °C. b)0.1XSSC при 65 °C. c) 0.1Х SSC, 0.5 % SDS при 68 °C. d) 0.1X SSC, 0.5 % SDS, 50 % формамід при 42 °C. с) 0.2Х SSC, 0.1 % SDS при 42 °C. f) 2X SSC при 65 °C (умови низької жорсткості). В одній з реалізацій способу даного винаходу, полінуклеотид даного винаходу включає полінуклеотид, який гібридизує до молекули нуклеїнової кислоти, яка включає або складається із молекули нуклеїнової кислоти, яка має послідовність, приведену в Seq ID No. 1 або 3, або 5, або 6, або її фрагмент. Фрагмент включає або складається, переважно, із 15, 20, 30, 40, 70, 100, 300, 500, 700, 1000 або більше залишків Seq ID No.1 або 3, або 5, або 6. В переважній реалізації, полінуклеотид даного винаходу включає полінуклеотид, який гібридизується за "жорстких" умов гібридизації із молекулою нуклеїнової кислоти, яка включає або складається із молекули нуклеїнової кислоти, яка має послідовність, наведену в Seq ID No.1 або 3, або 5, або 6, або її фрагмент. Термін "за жорстких умов гібридизації", як його вжито тут, вказує на будь-які зі згаданих тут жорстких умов гібридизації. В наступній реалізації, термін "за жорстких умов гібридизації" вказує на умови гібридизації, згадані в прикладах або використані в Sambrook (Molecular Cloning; A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989). В одній з переважних реалізацій, термін "за жорстких умов гібридизації", як його вжито тут, вказує на усі зі згаданих тут жорстких умов гібридизації і означає, що полінуклеотид гібридизується за усіх згаданих жорстких умов. Rpi-blb2, одержаний із інших організмів, може бути кодований іншими послідовностями ДНК, які гібридизуються із послідовностями, наведеними в Seq ID No.1 або 3 або 5 або 6 за послаблених умов гібридизації, та які кодують експресію пептидів, що мають активність Rpiblb2. Також, деякі застосування можуть бути виконані за умов низької жорсткості без будь-яких наслідків щодо специфічності гібридизації. Наприклад, аналіз "Southern blot" загальної ДНК може бути проведений із полінуклеотидом даного винаходу та відмитий за низької жорсткості (55 °C в 2xSSPE, 0,1 % SDS). Гібридизаційний аналіз може виявити лише просту структуру генів, що кодують Rpi-blb2. Наступним прикладом таких умов низької жорсткості є 4XSSC при 50 °C або гібридизація із від 30-40 % формаміду при 42 °C. Такі молекули включають фрагменти, аналоги або похідні Rpi-blb2 даного винаходу та відрізняються, наприклад, амінокислотною та/або нуклеотидною делецією(ями), вставкою(ами), заміною(ами), приєднанням(ями) та/або рекомбінацією(ями) або будь-якою(ими) іншою(ими) модифікацією(ями), відомою(ими) з рівня техніки окремо, або в комбінації з вищеописаними амінокислотними послідовностями або нуклеотидною(ими) послідовністю(остями), що їм відповідає(ють) Тим не менш, бажано використовувати жорсткі умови гібридизації. Термін "гомологія" означає, що відповідні молекули нуклеїнової кислоти або кодовані протеїни є функціонально та/або структурно еквівалентними. Молекули нуклеїнової кислоти, які гомологічні до молекул нуклеїнової кислоти, описаних вище, та які є похідними згаданих молекул нуклеїнової кислоти є, наприклад, варіаціями згаданих молекул нуклеїнової кислоти, які представляють модифікації, які мають таку саму біологічну функцію, зокрема, кодування протеїнів із такою ж або в значній мірі такою ж самою біологічною функцією. Це можуть бути природні варіації такі, як послідовності із інших сортів рослин або видів або мутації. Ці мутації можуть бути природними або можуть бути одержані методами мутагенезу. Алельні варіації можуть бути природними алельними варіаціями, так само, як і штучно отриманими або генноінженерними варіантами. Структурні еквіваленти можуть бути, наприклад, визначені дослідженням зв'язування згаданого поліпептиду з антитілами. Структурний еквівалент має подібні імунологічні характеристики, наприклад, включає подібні епітопи. Терміни "фрагмент", "фрагмент послідовності" або "частина послідовності" означають вкорочену послідовність, порівняно із оригінальною послідовністю. Вкорочена послідовність (послідовність нуклеїнової кислоти або протеїну) може значно змінюватися по довжині; мінімальним розміром є послідовність довжини, достатньої для забезпечення послідовності, принаймні порівняно подібної функції та/або активності оригінальної послідовності, тоді як максимум розміру не є критичним. При деяких застосуваннях, максимальний розмір, зазвичай не є значно більшим, ніж того потребує забезпечення бажаної активності та/або функції(й) оригінальної послідовності. 7 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Типово, довжина вкороченої амінокислотної послідовності знаходиться в межах від 5 до більш ніж 1260 амінокислот. Більш типово, однак, довжина складатиме максимум 1000 амінокислот, переважно, максимум від 500 до 100 амінокислот. Зазвичай, бажано вибирати послідовності, із принаймні близько 10, 12 або 15 амінокислот, до максимум близько 20 або 25 амінокислот. Термін "епітоп" вказує на специфічні імунореактивні сайти в середині антигену, також відомі, як антигенні детермінанти. Ці епітопи можуть бути лінійними ділянками мономерів в полімерній композиції, такі як амінокислоти в протеїні, або складатися, або бути представленими більш складними вторинними або третинними структурами. Фахівцеві зрозуміло, що усі імуногени (наприклад, речовини, здатні підвищувати імунну відповідь) є антигенами; однак, деякі антигени, такі як гаптени, не є імуногенами, але можуть бути імуногенними приєднуючись до молекули носія. Термін "антиген" включає посилання на речовину, до якої можуть бути утворені антитіла та/або до котрих антитіла є специфічно імунореактивними. В одній з реалізацій, даний винахід стосується епітопу Rpi-blb2. Термін "одна або декілька амінокислот" стосується принаймні однієї амінокислоти, але не більш ніж такої кількості амінокислот, яка може призвести до гомології менше 70 %. Переважно, подібність є більшою за 75 % або 80 %, більш бажано, більш ніж 85 %, 90 % або 95 %, ще більш бажано, більшою ніж 96 %, 97 %, 98 %, або 99 % подібності. Терміни "полінуклеотид" та "молекула нуклеїнової кислоти" також стосується "виділених" полінуклеотидів та молекул нуклеїнової кислоти. "Виділена" молекула нуклеїнової кислоти є відокремленою від інших молекул нуклеїнової кислоти, які є присутніми в природних джерелах нуклеїнової кислоти. Переважно, "виділена" нуклеїнова кислота не має послідовностей, які зазвичай фланкують нуклеїнову кислоту (наприклад, послідовності розташовані на 5' та 3' кінцях нуклеїнової кислоти) в геномній ДНК організму, з котрого одержано нуклеїнову кислоту. Наприклад, при різних реалізаціях полінуклеотид даного винаходу може мати менш ніж 5 kb, 4kb, 3kb, 2kb, 1 kb, 0.5 kb або 0.1 kb або менше нуклеотидів послідовності, що природно фланкує молекулу нуклеїнової кислоти в геномній ДНК клітини, з якої походить нуклеїнова кислота. Крім того, полінуклеотиди даного винаходу, зокрема, "виділена" молекула нуклеїнової кислоти така, як молекула кДНК, може бути в значній мірі відокремлена від клітинного матеріалу або культурального середовища, у випадку одержання за допомогою методів рекомбінації, або відокремлена від хімічних попередників або інших хімічних речовин, у випадку хімічного синтезу. Також, полінуклеотиди даного винаходу включають молекулу нуклеїнової кислоти, яка є комплементарною до однієї з нуклеотидних послідовностей вищезгаданих полінуклеотидів або їхньої частини. Молекула нуклеїнової кислоти, яка є комплементарною до однієї з нуклеотидних послідовностей наведених в SEQ ID No:1 або 3, або 5, або 6 є суттєво комплементарною до однієї з нуклеотидних послідовностей наведених в SEQ ID No: 1 або 3, або 5, або 6 та здатна гібридизуватися з однієї з нуклеотидних послідовностей наведених в SEQ ID No: 1 або 3, або 5, або 6, утворюючи стабільний дуплекс. Полінуклеотид даного винаходу, являє собою нуклеотидну послідовність, яка принаймні на 70 %, переважно, принаймні на 75 %, більш бажано, принаймні на 80 %, 90 %, або 95 %, та ще більш бажано, принаймні на 96 %, 97 %, 98 %, 99 % або більше гомологічна до нуклеотидної послідовності, наведеної в SEQ ID No: 1 або 3, або 5, або 6, або її частини. Полінуклеотид даного винаходу є нуклеотидною послідовністю, яка гібридизує, переважно гібридизує за жорстких умов, як визначено тут, до однієї з нуклеотидних послідовностей, наведених в SEQ ID No: 1 або 3, або 5, або 6, або її частини. Крім того, полінуклеотид даного винаходу може містити лише частину кодуючого регіону однієї з послідовностей в SEQ ID No: 1 або 3, або 5, або 6, наприклад, фрагменту, який може бути використаний як зонд або праймер, або фрагмент кодуючої послідовності гену, що кодує протеїн Rpi-blb2. Нуклеотидні послідовності, визначені клонуванням гену, що кодує даний протеїн Rpi-blb2, дозволяють створювати зонди та праймери для визначення та/або клонування його гомологів в інших типах клітин та організмах. Зонд/праймер, як правило, містить в значній мірі очищені олігонуклеотиди. Олігонуклеотид, як правило, включає ділянку нуклеотидної послідовності, яка гібридизується за жорстких умов із принаймні майже 12, 15, переважно майже з 20 або 25, більш переважно, майже з 40, 50 або 75 послідовними нуклеотидами смислового ланцюга однієї із послідовностей, наприклад, наведених в SEQ ID No: 1 або 3, або 5, або 6, антисмислової послідовності однієї із послідовностей, наприклад, наведених в SEQ ID No: 1 або 3, або 5, або 6, або її природних мутантів. Праймери, що базуються на нуклеотидах даного винаходу можуть бути використані у реакціях PLR для клонування гомологів Rpi-blb2, наприклад, такі праймери, як описано в прикладах даного винаходу, наприклад, наведені в табл. 3а або 3b, переважно, 8 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 використовують праймери ARF1F та ARF1R. PLR із праймерами univ24R та univT4L давали в результаті фрагмент Rpi-blb2, який може бути використаний як описано тут. Ці набори праймерів є рівноцінними. Фахівцям відомо, як скомбінувати праймери для одержання бажаного продукту, наприклад клону повної довжини або частини послідовності. Зонди, що базуються на нуклеотидних послідовностях Rpi-blb2, можуть бути використані для визначення транскриптів або геномних послідовностей, які їх кодують, або гомологічних протеїнів. Зонди також, можуть включати приєднану мічену групу, наприклад, мічена група може бути радіоізотопом, флуоресцентною сполукою, ферментом або кофактором ферменту. Такі зонди можуть бути використані як частини геномних маркерних тестів для визначання клітин, які експресують Rpiblb2, наприклад, для визначення рівня молекул нуклеїнової кислоти, що кодують Rpi-blb2 в досліджуваних клітинах, наприклад, для визначення рівнів мРНК Rpi-blb2, або для визначення геномної мутації або делеції гену Rpi-blb2. Полінуклеотид даного винаходу кодує поліпептид або його частину, яка включає амінокислотну послідовність, яка достатньо гомологічна до амінокислотної послідовності SEQ ID No: 2 або 4 так, що протеїн або його частина зберігає здатність брати участь в резистентності до патогенів, зокрема, активності протеїну Rpi-blb2 як описано в дослідах на рослинах. Як використано тут, термін "достатньо гомологічний" вказує на протеїн або його частину, яка має амінокислотні послідовності, які включають мінімальну кількість подібних або еквівалентних (наприклад, залишок амінокислоти, який має такий самий бічний ланцюг, як залишок амінокислоти однієї з послідовностей поліпептиду даного винаходу), до амінокислотних залишків амінокислотної послідовності Seq ID No.: 2 або 4 так, що протеїн або його частина здатна брати участь в резистентності рослин до вказаних патогенів. Приклади активності протеїну Rpi-blb2 описані тут. Отже, функція протеїну Rpi-blb2 надає прямо чи опосередковано резистентність до рослинних патогенів, переважно до згадуваних тут патогенів, більш переважно до P.infestans. Протеїн принаймні на 70 %, переважно принаймні на 75 %, більш переважно принаймні на 80 %, 90 %, або 95 %, та ще більш переважно принаймні на 96 %, 97 %, 98 %, 99 % або більше гомологічний до повної амінокислотної послідовності SEQ ID No: 2 або 4. Частини протеїну, що кодуються полінуклеотидом даного винаходу є, переважно, біологічно активними. Як, зазначалося тут, термін "біологічно активна частина" призначений для позначення частини, наприклад, домену/фрагменту, який надає резистентність до ооміцетного патогену рослини та/або Bemisia tabaci та/або афід, або має імунологічну активність, таку, що вони зв'язуються із антитілом специфічним до протеїну Rpi-blb2, або вони мають активність як пояснюється в прикладах, або як описано тут. Інші фрагменти нуклеїнової кислоти, що кодують біологічно активні частини поліпептиду даного винаходу можуть бути отримані виділенням частини однієї з послідовностей SEQ ID No: 1 або 3, або 5, або 6, які експресують кодовану частину протеїну або пептиду Rpi-blb2 (наприклад, рекомбінантною експресією in vitro) та визначенням активності кодованої частини протеїну. Також винахід охоплює полінуклеотиди, які відрізняються від однієї з нуклеотидних послідовностей, приведених в SEQ ID No:1 або 3, або 5, або 6 (або їх частин), завдяки виродженості генетичного коду, й таким чином кодують поліпептид Rpi-blb2, як такий, що кодується послідовностями, приведеними в Seq ID No.: 2 або 4. Полінуклеотид даного винаходу, являє собою нуклеотидну послідовність, яка кодує протеїн, що має амінокислотну послідовність, приведену в SEQ ID No: 2 або 4. В наступній реалізації, полінуклеотид даного винаходу, кодує протеїн повної довжини, який в значній мірі гомологічний до амінокислотної послідовності наведеної в SEQ ID No: 2 або 4. Також, фахівцю буде зрозуміло, що в популяціях може існувати поліморфізм послідовності ДНК, який призводить до змін в амінокислотних послідовностях (наприклад, в популяції S. bulbocastanum). Такий генетичний поліморфізм в гені Rpi-blb2 може мати місце серед особин популяції завдяки природній мінливості. Як використано тут, термін "ген" та "рекомбінантний ген" вказує на молекули нуклеїнової кислоти, які мають відкриті рамки зчитування, що кодують Rpi-blb2, переважно Rpi-blb2i із S. bulbocastanum. Такі природні варіації звичайно можуть давати мінливість 1-5 % в нуклеотидній послідовності гену Rpi-blb2. Будь-яка та всі такі нуклеотидні варіації та одержані амінокислотні поліморфізми в Rpi-blb2, які є результатом природної мінливості та не змінюють функціональну активність Rpi-blb2 необхідно розуміти як такі, що знаходяться в межах даного винаходу. Полінуклеотиди, які відповідають природним варіантам та гомологам кДНК Rpi-blb2 даного винаходу не з S. Bulbocastanum, можуть бути виділені на основі їхньої гомології до 9 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 полінуклеотидів Rpi-blb2 із S. Bulbocastanum, розкритих тут за допомогою полінуклеотиду даного винаходу, або його частини, як гібридизаційного зонда, відповідно до стандартної схеми гібридизації за жорстких умов. При цьому, при іншій реалізації, полінуклеотид винаходу має довжину приблизно 20 нуклеотидів. Переважно, він гібридизується за жорстких умов із молекулою нуклеїнової кислоти, яка включає нуклеотидну послідовність полінуклеотиду даного винаходу, наприклад, SEQ ID No: 1 або 3, або 5, або 6. При інших реалізаціях, нуклеїнова кислота має довжину принаймні 20, 30, 50, 100, 250 або більше нуклеотидів. Термін "гібридизується за жорстких умов" як визначено вище, призначено для опису умов гібридизації та відмивання, за котрих нуклеотидна послідовність принаймні на 65 % подібна до кожної іншої послідовності, та які переважно залишаються гібридизованими одна з одною. Переважно, умови є такими, що послідовності, принаймні на 70 %, більш бажано, принаймні на 75 %, 80 %, ще більш бажано, принаймні на 85 %, 90 %, або 95 %, або більше є ідентичними одна з одною і залишається гібридизованими одна з одною. Переважно, полінуклеотид даного винаходу, який гібридизується за жорстких умов до послідовності приведеної в SEQ ID No: 1 або 3, або 5, або 6, відповідає природним молекулам нуклеїнової кислоти. Як тут вжито, "природна" молекула нуклеїнової кислоти відповідає молекулі РНК або ДНК, яка має нуклеотидну послідовність, що зустрічається в природі (наприклад, кодує природний протеїн). Переважно, вона кодує полінуклеотид природного Rpi-blb2 S. bulbocastanum. На додачу до природних варіантів послідовностей Rpi-blb2, що можуть існувати в популяції, фахівець може також визнати, що зміни можуть бути введені за допомогою мутації в нуклеотидну послідовність полінуклеотиду, що кодує Rpi-blb2, й таким чином призвести до змін в амінокислотній послідовності кодованого Rpi-blb2 без зміни функціональної здатності Rpi-blb2. Наприклад, нуклеотидні заміни, що призводять до амінокислотних замін в "неважливих" амінокислотних залишках, можуть бути зроблені в послідовності полінуклеотиду, що кодує Rpiblb2, наприклад, SEQ ID No: 1 або 3, або 5, або 6. "Неважливі" амінокислотні залишки є залишками, які можуть бути змінені в послідовності дикого типу протеїну Rpi-blb2 без зміни активності згаданого протеїну Rpi-blb2, тоді як "важливі" амінокислотні залишки є залишками,необхідними для активності протеїну Rpi-blb2. Інші амінокислотні залишки, однак, (наприклад, ті, що не є консервативними або є лише напівконсервативними в домені, що несе активність Rpiblb2) можуть не бути важливими для активності, й, таким чином, скоріше відповідати змінам без зміни активності Rpi-blb2. Відповідно, фахівцю є відомими кодони, які можуть бути змінені при переході від організму до організму. От чому ми будемо пристосовувати використання кодонів в полінуклеотиді даного винаходу, до застосування організму, в якому полінуклеотид або поліпептид експресується. Таким чином, винахід стосується полінуклеотидів, що кодують Rpi-blb2, які містять зміни в амінокислотних залишках, які не є важливими для активності Rpi-blb2. Такі Rpi-blb2-и відрізняються амінокислотною послідовністю від послідовності, що міститься в SEQ ID No: 2 або 4 але зберігають описану тут активність Rpi-blb2. Полінуклеотид може містити нуклеотидну послідовність, що кодує поліпептид, причому поліпептид містить амінокислотну послідовність, яка принаймні на 70 % подібна до амінокислотної послідовності, наведеної в SEQ ID No: 2 або 4 та здатна брати участь в резистентності до рослинного патогену. Переважно, протеїн, кодований молекулою нуклеїнової кислоти, є принаймні на 70 % подібний до амінокислотної послідовності SEQ ID No: 2 або 4, більш бажано, принаймні на 75 % подібний до амінокислотної послідовності SEQ ID No: 2 або 4, ще більш бажано, принаймні на 80 %, 90 %, або 95 % подібний до амінокислотної послідовності SEQ ID No: 2 або 4, та ще більш бажано, принаймні на 96 %, 97 %, 98 %, 99 % подібний до амінокислотної послідовності SEQ ID No: 2 або 4. Для визначення відсотку гомології двох амінокислотних послідовностей (наприклад, однієї із Seq ID No.: 2 або 4 та її мутантної форми) або двох нуклеїнових кислот, послідовності суміщаються для досягнення оптимального порівняння (наприклад, можуть бути введені проміжки в послідовності одного протеїну або нуклеїнової кислоти для оптимального суміщення з іншим протеїном або нуклеїновою кислотою). Потім порівнюються амінокислотні залишки або нуклеотиди у відповідних амінокислотних положеннях або нуклеотидних положеннях. Коли положення в одній послідовності (наприклад, однієї з послідовностей SEQ ID No: 2 або 4) зайнято таким самим амінокислотним залишком або нуклеотидом, як у відповідній позиції в іншій послідовності (наприклад, в мутантній формі вибраної послідовності), тоді молекули є гомологічними в цій позиції (наприклад, як використано тут, амінокислотна або нуклеїновокислотна "гомологія" є еквівалентом до амінокислотної або нуклеїновокислотної "ідентичності", "подібності"). Відсоток гомології між двома послідовностями є функцією кількості спільних ідентичних позицій в послідовностях (тобто, % гомології = кількість ідентичних позицій/загальна кількість позицій х 100). 10 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Гомологія може бути обрахована порівнянням за допомогою програмного алгоритму GAP (Wisconsin Package Version 10.0, University of Wisconsin, Genetics Computer Group (GCG), Madison, USA; Altschul et al.(1997) Nucleic Acids Res.25:3389 et seq), при встановленні наступних параметрів: Gap weight: 50 Length weight: 3 Average match: 10 Average mismatch: 0 Наприклад, послідовність, яка має, принаймні 80 % гомології із послідовністю SEQ ID NO: 1 на нуклеїновокислотному рівні означає, що послідовність, яка при порівнянні з SEQ ID NO: 1 при порівнянні вище згаданою програмою із вказаними установками параметрів, має принаймні 80 % гомології. Гомологія між двома поліпептидами означає, що ідентичність амінокислотної послідовності в кожному випадку є на повну довжину послідовності, й може бути обрахована порівнянням за допомогою програмного алгоритму GAP (Wisconsin Package Version 10.0, University of Wisconsin, Genetics Computer Group (GCG), Madison, USA), при встановленні наступних параметрів: Gap weight: 8 Length weight: 2 Average match: 2,912 Average mismatch: - 2,003 Наприклад, послідовність, яка має, принаймні 80 % гомології із послідовністю SEQ ID NO: 2 на протеїновому рівні, розуміється як значення того, що послідовність, яка при порівнянні з SEQ ID NO: 2 при порівнянні вище згаданою програмою із вказаними установками параметрів, має принаймні 80 % гомології. В даній заявці, гомологія може бути визначена за допомогою програми clustalW, яка може бути знайдена на www.ebi.ac.uk/tools, потрібно вибрати аналіз послідовностей та вибрати опцію clustalW (множинне суміщення послідовностей). Усі опції виконувалися за стандартних умов, які є наступними: alignment: full; output format: aln w/numbers; output order: aligned; color alignment: no; ktup (word size): def; window length: def; score type: percent; topdiag: def; pairgap: def; matrix: def; gap open: def; end gaps: def; gap extension: def; gap distances: def; cpu mode: single; tree graph/type: cladogram; tree graph/distances: hide; phyloge-netic tree/tree type: none; phylogenetic tree/correct dist.: off; phylogenetic tree/ignore gaps: off. Однак, бажано гомологію обчислювати за допомогою clustalW. Функціональні еквіваленти, одержані з поліпептидів, наведених в SEQ ID NO: 2 або 4 згідно винаходу, шляхом заміщень, вставок, або делецій мають принаймні 70 %, переважно принаймні 80 %, більш бажано принаймні 90 %, особливо бажано принаймні 95 %, надзвичайно бажано, принаймні 98 % гомології з одним із поліпептидів, наведених в SEQ ID NO: 2 або 4 відповідно до даного винаходу, та відрізняються тими самими властивостями, що й поліпептиди, наведені в SEQ ID NO: 2 або 4. Функціональні еквіваленти, одержані з нуклеотидних послідовностей, приведених в SEQ ID No: 1 або 3, або 5, або 6 згідно винаходу, шляхом заміщень, вставок, або делецій мають принаймні 70 %, переважно принаймні 80 %, більш бажано принаймні 90 %, особливо бажано принаймні 95 %, надзвичайно бажано принаймні 98 % гомології з одним із поліпептидів, наведених в SEQ ID NO: 2 або 4 відповідно до даного винаходу, та мають по суті ті самі властивості, що й поліпептиди, наведені в SEQ ID NO: 2 або 4. "По суті ті самі властивості " функціонального еквіваленту є усі що розуміються в плані внесення у фенотип резистентності до патогену або внесення або збільшення резистентності до, принаймні одного патогену, при збільшенні кількості протеїну, активності або функції згаданого функціонального еквіваленту Rpi-blb2 в рослині або тканині, частині рослини або її клітини. Більш того, споруляція та ушкодження фенотипу після інфікування в поєднанні зі згаданим підвищенням кількості протеїну, активності або функції згаданого функціонального еквіваленту, розуміється як важлива властивість. Молекула нуклеїнової кислоти, що кодує Rpi-blb2 гомологи протеїнової послідовності SEQ ID No: 2 або 4, може бути створена введенням однієї або більше нуклеотидних замін, вставок або делецій до нуклеотидної послідовності полінуклеотиду даного винаходу, зокрема SEQ ID No: 1 або 3, або 5, або 6 таким чином, що одна або більше нуклеотидних замін, вставок або делецій введені в кодований протеїн. Мутації можуть бути введені в послідовності, наприклад, SEQ ID No: 1 або 3, або 5, або 6 за допомогою стандартних методів, таких як сайт-спрямований мутагенез та PLR-опосередкований мутагенез. Переважно, консервативні заміни зроблені в одному або більше розрахованих неосновних амінокислотних залишках. "Консервативна амінокислотна заміна" це заміна, при якій амінокислотний залишок заміщено амінокислотним залишком, який має подібний бічний ланцюг. Сімейства амінокислотних залишків, які мають подібний бічний ланцюг, зазначено в літературі. Ці сімейства включають амінокислоти з 11 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 основними бічними ланцюгами (наприклад, лізин, аргінін, гістидин), кислотним бічним ланцюгом (наприклад, аспарагінова кислота, глютамінова кислота), незарядженими бічними ланцюгами (наприклад, гліцин, аспарагін, глютамін, серин, треонін, тирозин, цистеїн), неполярними бічними ланцюгами (наприклад, аланін, валін, лейцин, ізолейцин, пролін, фенілаланін, метіонін, триптофан), з бета-розгалуженими бічними ланцюгами (наприклад, треонін, валін, ізолейцин) та ароматичними бічними ланцюгами (наприклад, тирозин, фенілаланін, триптофан, гістидин). Отже, розрахований неосновний амінокислотний залишок в Rpi-blb2, переважно заміщений іншим амінокислотним залишком того самого сімейства. Також, при іншій реалізації мутації можуть бути введені випадково, впродовж усієї або в частині кодуючої послідовності Rpi-blb2, як наприклад насичувальним мутагенезом, та одержані мутанти можуть бути перевірені на описану тут активність Rpi-blb2 для визначення мутантів, які зберегли активність Rpi-blb2. За мутагенезом однієї з послідовностей SEQ ID No:1 або 3 або 5 або 6, кодований протеїн може бути експресований рекомбінантно та активність протеїну може бути визначена, за допомогою, наприклад, описаного тут методу (дивись Приклади). В іншій реалізації в способі даного винаходу активність протеїну Rpi-blb2, та іншого протеїну резистентності підвищено. Очікувалося, що в польових умовах присутність більш ніж одного гену резистентності є виграшною, зокрема генів, що несуть резистентність до такого самого патогену. У випадку якщо присутній ізолят патогену, наприклад, раси P.infestans, який здатен подолати резистентність до одного із R-генів, інші один або більше R-reH(n) залишається(ються) функціональними, й робить(лять) неможливим інфікування рослини. Присутні два нездолані R-гени значно знижують шанси цього патогену, зокрема раси P.infestans, яка зможе мутувати в расу, яка здолає два або більше R-генів. В подальшому "поліпептид резистентності" або "протеїн резистентності" стосується поліпептиду, активність котрого (збільшена) несе резистентність до придатного генотипу ("дикого типу" або "вихідного"). Відповідно, Rpi-blb2 є протеїном резистентності, так само як і, наприклад, Rpi-blb (або RB або Sbu1). "Інший протеїн резистентності" стосується іншого протеїну резистентності, який є протеїном даного винаходу, причому термін "протеїн резистентності" включає поліпептид даного винаходу а також один або більше інший(і) протеїн(и) резистентності. Також зрозуміло, що термін "та інший протеїн резистентності" стосується одного або більше інших протеїнів резистентності. Таким чином, може бути підвищена активність одного або більше додаткових протеїнів резистентності. Інші протеїни резистентності описані далі. Однак, в загалі, будь-який протеїн резистентності може бути сумісно експресований із поліпептидом даного винаходу або його активність може бути підвищена будь-яким з описаних тут методів для Rpi-blb2. В переважній реалізації, інший протеїн може включати домен LRR та Р-петлю. Клонування та опис характеристик більш ніж 30 (R) генів резистентності до хвороб рослин, які несуть резистентність до бактерій, грибів, ооміцетів, вірусів, нематод або комах дозволило їх класифікувати по структурних класах залежно від специфіки патогену (огляд за Dangl and Jones, 2001). Найбільшим класом описаних R генів, який містить майже 0.5 процента розрахункових генів в геномі Arabidopsis, передбачається, що вони кодують внутрішньоклітинні протеїни, які несуть багатий на лейцин повтор (LRR) та домени сайту зв'язування нуклеотиду (NBS), фрагменти також знайдені в інших рецепторних протеїнах та протеїнах передачі сигналу. Rпротеїни NBS-LRR переважно відрізняються N-кінцем, який має послідовність подібну до Tollпротеїну Drosophila та рецепторного домену інтерлейкіну-1 ссавців (TIR-NBS-LRR), або кодує супер-спіралізовану структуру (CC-NBS-LRR), іноді у формі лейцинової "застіжки-блискавки" (LZ-NBS-LRR). Також може бути мембранно-зв'язаним, протеїном NBS-LRR, який за розрахунками є цитоплазматичним На відміну від цього, два інші класи R-протеїнів можуть нести LRR-и які за розрахунками є трансмембранними, із зовнішньоклітинним доменом LRR: LRR-трансмембранні (LRR-TM) Cf-протеїни та LRR-TM-кіназний Ха21-протеїн. Описані Rпротеїни які не мають LRRs, є Pto-ген томатів, ге HHs1pro-1 буряків, ген mlo ячменю, гени Rpw8 з Arabidopsis та ген Rpg1 ячменю. Згідно гіпотези "ген-проти-гену", резистентність до захворювання супроводжується сприйняттям рослинними R-протеїнами ефекторних молекул із антивірусною функцією (Avr), й таким чином запуску деяких комплексів розпізнавання еліситору, шляхів передачі сигналу, що ведуть до гіперчутливої відповіді (HR). Вважається, що разом з іншими рецепторами протеїни NBS-LRR R мають модульну структуру із окремими розпізнавальними та сигнальними доменами, згідно із чим LRR є можливим розпізнавальним доменом, а N-кінцевий регіон включає NBS, основний сигнальний домен. Функціональний аналіз рекомбінантних LRRпротеїнів, показує, що розпізнавальна специфічність дійсно знаходиться в LRR. Більш того, LRR 12 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 і є найбільш варіабельним регіоном, щільно пов'язаним із NBS-LRR-протеїнами та підпадає під селекційний відбір. Однак, накопичується свідчення того, що LRR-и також роблять внесок в сигнальну функцію через негативну регуляцію, включаючись у можливі внутрішньоклітинні взаємодії Тепер було клоновано п'ять R-генів з картоплі, включаючи два R-гени, що несуть резистентність до фітофторозу, та усі вони належать до класу CC/LZ-NBS-LRR рослинних Rгенів. Тоді як R1-ген одержаний із S.demissum, несе расову специфічність до фітофторозу, нещодавно клонований ген Rpi-blb (або RB або Sbu1) із S.bulbocastanum несе повну резистентність до спектру ізолятів P.infestans, що мають багато вірулентних факторів, та не проявляв жодної расової специфічності. Також, як описано раніше, покоління рослин із соматичних гібридів, що містять Rpi-blb, були невразливі фітофторою під час польових експериментів у Мексиці, де були знайдені майже усі раси грибів. Було показано, за допомогою комплементації сприйнятливого фенотипу в культивованій картоплі та томатах потенціал міжвидового трансферу резистентності широкого спектру дії до фітофтори в культивованих Solanaceae із статево несумісних специфічних до хазяїна видів за допомогою трансформації єдиними клонованими R-генами US 6,127,607 описує протеїни резистентності із LRR-доменами та Р-петлями. Зміст US 6,127,607 повністю включено через посилання. Зокрема, колонки з 6 по 8 та 11 описує LRR-домени та Р-петлі. Крім того в Song, 2003, PNAS 100 (16), 9128-9133 приведено порівняння Rpi-blb LRR фрагментів на Fig.4 приведено на сторінці 9132 огляд LRRдоменів. Домени поліпептиду даного винаходу приведено на Фіг. 14 а також на Фіг. 15. Переважно активність одного або більше протеїну(ів) резистентності вибраних із групи І, що включає Rpi-blb (синонім RB або Sbu1), Rpi-ABPT1, Rpi-blb3, Rpi-mcd, R1, R-ber (синонім R12), Rpi1, R2, R3a, R3b, R4, R5, R6, R7, R8, R9, R10, R11, Ph-1, Ph-2 та Ph-3 є підвищеною. Переважно, щоб додатково до Rpi-blb2 також принаймні збільшеною є активність Rpi-blb. В іншій реалізації способу даного винаходу, експресія, наприклад трансгенного, протеїну Rpi-blb2 є збільшеною, та іншого трансгенного гену резистентності є збільшеною Протеїн резистентності експресується разом із Rpi-blb2 (або RB або Sbu1), який переважно є згадуваним тут протеїном, зокрема Rpi-blb, Rpi-ABPT1, Rpi-blb3, R1, Rpi1, R-ber, Rpi-mcd, R2, R3a, R3b, R6, R7, Ph-1, Ph-2 або Phi 3 але може також бути один з інших протеїнів, що несуть резистентність до рослинних патогенів, відомих фахівцям. Як вжито тут, термін "підвищена експресія" згідно даного винаходу також включає de novoекспресію полінуклеотиду або поліпептиду. Найбільш переважним, є підвищення резистентності через сумісну експресію поліпептиду даного винаходу із Rpi-blb, Rpi-blb та Rpi-blb2 разом надають повну резистентність в дослідах на відокремленому листі з ізолятами P.infestans як описано в прикладах, та в Song 2003, PNAS 100 (16), 9128. Згадані гени, що несуть резистентність, наприклад, описані в RB або Sbu1 (синоним Rpi-blb): AY336128 [gi: 32693280], (Song et al., 2003). ВАС клони 177 013 та СВ3А14 які мають ген Rpi-blb депоновані в GenBank із вхідними номерами AY303171 та AY303170. R1: AF447489 [gi: 9117432422], (Ballvora et al., 2002) Rpi1: Kuhl, J.C., Hanneman, R.E., and Havey, M.J., (2201) Characterization and mapping of Rpi1, a late blight resistance locus from diploid (1EBN) Mexican Solanum pinnatisectum.Molecular genet.Genomics 265: 977-985.R7ber: Ewing, E.E., Simko, I., Smart, CD., Bonierbale, M.W., Mizubuti, E.S.G., May, G.D., and Fry, W.E., (2000) Genetic mapping from field tests of qualitative and quantitative resistance to Phytophthora infestans in a population derived from Solanum tuberosum and Solanum berthaultii.Molecular breeding 6:25-36.R2: Li, X., vanEck, H.J., vanderVoort, J.NAM., Huigen, D.J., Stam, P., and Jacobsen, E.(1998) Autotetraploids and genetic mapping using common AFLP markers: the R2 allele conferring resistance to Phytophthora infestans mapped on potato chromosome 4.Theoretical and Applied Genetics 96 (8): 1121-112. R3, R6, R7: Elkharbotly, A., Palominosanchez, C, Salamini, F., Jacobsen, E., and Gebhardt, C.(1996) R6 and R7 alleles of potato conferring race-specific resistance to Phytophthora infestans (Mont) de Вагу identified genetic loci clustering with the R3 locus on chromosome XI.Theoretical and Applied.Genetics 92 (7): 880-884.Ph-1: Bonde and Murphy (1952) Main Agric.Exp.Stn.Bull.No 497 Ph-2: Moreau, P., Thoquet, P., Olivier, J., Laterrot, H., and Grimsley, N.H. (1998) Genetic mapping of Ph-2, a single locus controlling partial resistance to Phytophthora infestans in tomato. Molecular Plant Microbe Interactions 11 (4): 259-269. Ph-3: Chunwongse, J., Chunwongse, C, Black, L, and Hanson, P.(2002) Molecular mapping of the Ph-3 gene for late blight resistance in tomato. Journal of Horticultural Science & Biotechnology 77 (3): 281-286. Rpi-blb3, Rpi-ABPT1 and Rpi-mcd: Park, Т.Н., Van der Vossen, E., Vleeshouwers, V.G.A.A., Tan, A., Visser, R.G.F.and Van Eck, H.J.2004. Major resistance genes for tuber and leaf resistance to 13 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Phytophthora infestans in potato: An outline of a PhD project.Crop Functional Genomics 2004, July 2004, Jeju, Korea, page 93. R3a and R3b: Huang, S., Vleeshouwers, V.G.A.A., Werij, J.S., Hutten, R.C.B., Van Eck, H.J., Visser, R.G.F, and Jacobsen, E.(2004). The R3 resistance to Phytophthora infestans in potato is conferred by two closely linked R-genes with distinct specificities. MPMI 17 (4), 428-435. В одній з реалізацій, активність Rpi-blb2 збільшені, згідно даного винаходу, наприклад, збільшена експресія полінуклеотиду, даного винаходу та збільшено експресію, принаймні однієї нуклеїнової кислоти, що кодує Rpi-blb, Rpi-ABPT1, Rpi-blb3, Rpi-mcd R1, R-ber, Rpi1, R2, R3a, R3b, R6, R7, Ph-1, Ph-2 та/або Ph-3 причому молекула нуклеїнової кислоти, вибрана з групи, що складається з: (а) молекули нуклеїнової кислоти, що кодує принаймні зрілу форму поліпептиду Rpi-blb (або RB- або Sbu1-), переважно такого, що кодується послідовністю, приведеною за вхідним номером GenBank: AY336128 [gi: 32693280]; поліпептиду R1, переважно такого, що кодується послідовністю, приведеною за вхідним номером GenBank: AF447489 [gi 9117432422]; полінуклеотид Rpi-blb3, Rpi-ABPT1 ат/або Rpi-mcd, переважно такий, що кодується послідовністю або похідною з неї за інформацією приведеною в Park, Т.Н., Van der Vossen, E., Vleeshouwers, V.G.A.A., Tan, A., Visser, R.G.F.and Van Eck, H.J.2004. Major resistance genes for tuber and leaf resistance to Phytophthora infestans in potato: An outline of a PhD project. Crop Functional Genomics 2004, July 2004, Jeju, Korea, page 93. поліпептид R3a та/або R3b, переважно такий, що кодується послідовністю або похідною з неї за інформацією приведеною в Huang, S., Vleeshouwers, V.G.A.A., Werij, J.S., Hutten, R.C.B., Van Eck, H.J., Visser, R.G.F, and Jacobsen, E.(2004). R3-резистентність до Phytophthora infestans в картоплі, що несуть два тісно зв'язані R-гени із значною специфічністю. МРМІ 17 (4), 428-435. Резистентність протеїн, що несе резистентність до патогену, переважно P. infestans, картований та описаний як приведені, як, наприклад, для для Rpi1 в Kuhl, J.C., Hanneman, R.E., and Havey, M.J., (2001) Characterization and mapping of Rpi1, a late blight resistance locus from diploid (1EBN) Mexican Solanum pinnatisectum.Molecular genet.Genomics 265: 977-985; для R-ber в Ewing, E.E., Simko, I., Smart, CD., Bonierbale, M.W., Mizubuti, E.S.G., May, G.D., and Fry, W.E., (2000) Genetic mapping from field tests of qualitative and quantitative resistance to Phytophthora infestans in a population derived from Solanum tuberosum and Solanum berthaultii. Molecular breeding 6:25-36; для R2 в Li, X., vanEck, H.J., vanderVoort, J.NAM., Huigen, D.J., Stanri, P., and Jacobsen, E.(1998) Autotetraploids and genetic mapping using common AFLP markers: the R2 allele conferring resistance to Phytophthora infestans mapped on potato chromosome 4 Theoretical and Applied Genetics 96 (8): 1121-1128; для R3, R6, R7 в Elkharbotly, A., Palominosanchez, C, Salamini, F., Jacobsen, E., and Gebhardt, C. (1996) R6 and R7 alleles of potato conferring race-specific resistance to Phytophthora infestans (Mont) de Bary identified genetic loci clustering with the R3 locus on chromosome XI.Theoretical and Applied.Genetics 92 (7): 880-884; для Ph-1 в Bonde and Murphy (1952) Main Agric.Exp.Stn.Bull.No 497; або для Ph-2 в Moreau, P., Thoquet, P., Olivier, J., Laterrot, H., and Grimsley, N.H. (1998) Genetic mapping of Ph-2, a single locus controlling partial resistance to Phytophthora infestans in tomato.Molecular Plant Microbe Interactions 11 (4): 259-269; та/або для Ph-3 в Chunwongse, J., Chunwongse, C, Black, L, and Hanson, P. (2002) Molecular mapping of the Ph-3 gene for late blight resistance in tomato. Journal of Horticultural Science & Biotechnology 77 (3): 281-286; або поліпептид, що несе резистентність до патогену, переважно P.infestans отриманий зі згаданих публікацій; b) молекули нуклеїнової кислоти, нуклеотидна послідовність яких є результатом виродження генетичного нуклеотидної послідовності (а); c) молекули нуклеїнової кислоти, що кодує поліпептид, який одержаний з поліпептиду, що кодується полінуклеотидом (а) або (b), шляхом заміни, делеції та/або додання однієї або декількох амінокислот амінокислотної послідовності поліпептиду, що кодується полінуклеотидом (а) або (b); d) молекули нуклеїнової кислоти, що кодує поліпептид, послідовність якого має ідентичність 70 % або більше до амінокислотної послідовності поліпептиду, який кодується молекулою нуклеїнової кислоти (а); 14 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 e) молекул нуклеїнової кислоти, які включають фрагмент або епітоп-несучу частину поліпептиду, кодованого будь-якою молекулою нуклеїнової кислоти від (а) по (d); f) молекула нуклеїнової кислоти, що кодує фрагмент, який починається з амінокислоти 1, 30, 50, 100, 200, 300, 500 або 1000, та закінчується амінокислотою 1276, 1000, 500, 300, 200, 50, або 1 поліпептиду, що кодується будь-якою молекулою нуклеїнової кислоти згідно з (а) - (e) та однією зі згаданих активностей; g) молекули нуклеїнової кислоти, що включає принаймні 20 нуклеотидів будь-якого з полінуклеотидів (а) або (b); h) молекули нуклеїнової кислоти, що кодує поліпептид, який упізнається моноклональним антитілом, що утворене проти поліпептиду, який кодується молекулою нуклеїнової кислоти згідно з будь-яким з підпунктів (а) - (h); і) молекули нуклеїнової кислоти, одержуваної шляхом скринінгу прийнятної бібліотеки при жорстких умовах з зондом, який має послідовність молекули нуклеїнової кислоти згідно з будь-яким з підпунктів (а) - (h) або її фрагменту розміром принаймні 20; та, переважно 30 або більше нуклеотидів; та j) молекули нуклеїнової кислоти, комплементарний ланцюг якої гібридизується у жорстких умовах з молекулою нуклеїнової кислоти згідно з будь-яким з (а) або (і); або комплементарного ланцюга будь-якої з молекул нуклеїнової кислоти згідно (а) - (j). Відповідно, спосіб даного винаходу, несе резистентність однієї із згаданих рослин, рослинної тканини або рослинної клітини даного винаходу до рослинного патогену типу Oomycetes, переважно патогену порядку Pythiales або Peronosperales, більш переважно сімейства Pythiaceae або Peronosporaceae, більш переважно роду Phytophthora або Bremia або Peronospera або Plasmopara, найбільш переважно де патогеном є один з видів Phytophthora parasitica var.nicotianae (яка спричиняє крім іншого "чорної ніжки" тютюну), Phytophthora sojae (яка спричиняє Phytophthora-ну кореневу гниль в бобах сої), Phytophthora capsici (яка спричиняє гниль у перцю та кабачку та томатів), Phytophthora erythroseptica (яка спричиняє рожеву гниль картоплі), Plasmopara viticola (яка спричиняє несправжню мучнисту росу винограду), Bremia lactuca (яка спричиняє несправжню мучнисту росу салату-латук) або Peronospora tabaci (яка спричиняє синю плісень тютюну). Активність Rpi-blb2 в рослинах, рослинних клітинах, рослинних тканинах, рослинних органах або їхніх частинах, згідно даного винаходу, можу бути підвищена, створена або стимульована за допомогою методу, які є відомими фахівцям та описані, наприклад, в Sambrooket al., Cold Spring Harbor Laboratory Press, NY, 1989. Отже, в переважній реалізації, даний винахід стосується способу даного винаходу, в якому експресія є "de novow-експресією. Термін "de novo-експресія" в клітині, тканині або в організмі або його частині як вона розуміється тут, вказує на експресію генетичного продукту після невизначуваності згаданого генетичного продукту або активності згаданого генетичного продукту, зокрема відповідного поліпептиду або полінуклеотиду в клітині, тканині або в організмі або його частині. Переважно, щоб ген, що кодує поліпептид або полінуклеотид в клітині, тканині або в організмі або його частині та який має бути de novo-експресований, не був присутнім в геномі клітини, тканини або організму або його частин. Якщо експресія генетичного продукту не визначається в клітині, тканині або в організмі або його частинах, звичайно робиться висновок про те, що експресія не відбувається в клітині, тканині або в організмі або його частині. Відповідно, якщо активність не можу бути виявлена, звичайно робиться висновок про те, що відповідна активність не існує, фахівцю є відомими способи виявлення та засоби досягнення більшої чутливості. Отже, в переважній реалізації, термін "de novo-експресія", вказує на нову або додаткову експресію в системах, де рівень активності, наприклад, із-за низького рівня експресії або експресії (майже) неактивного генетичного продукту є надто малою щоб нести резистентність до рослинного патогену, зокрема P.infestans. Порівняння "knock out" - штамів із низько та/або високоекспресуючими фенотипами можу показати, чи є різниця в резистентності до будь-якого згадуваного тут патогену є оглядовим. Відповідно, в іншій реалізації даного винаходу, є збільшеною ендогенна активність Rpi-blb2 та/або іншого протеїну резистентності. Рівень експресії в клітині можу бути підвищений способами відомими фахівцям. Декілька методик описані тут, наприклад, трансгенна експресія полінуклеотиду або поліпептиду даного винаходу. Полінуклеотид або поліпептид можуть бути зовнішнього походження Переважно, щоб полінуклеотид, що вводиться, був того самого генетичного походження, що і клітина хазяїн, рослинна клітина, рослинна тканина або рослина. Активність, зокрема, ендогенна активність, а також активність трансгенно експресованого Rpi-blb2 може бути підвищена декількома способами. Відповідно, в переважній реалізації, 15 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 активність описаних тут протеїнів резистентності є підвищеною одним або більше наступними етапами: a) стабілізація білка резистентності; b) стабілізація кодуючої мРНК білка резистентності; c) підвищення специфічної активності білка резистентності; d) експресію або підвищення експресії гомологічного або штучного фактора транскрипції для експресії білка резистентності; e) стимуляцію активності білка резистентності за допомогою екзогенних факторів індукції; f) експресію гена, що кодує трансгенний білок резистентності; та/або g) збільшення числа копій гена, що кодує білок резистентності. В загальному випадку, активність організму, зокрема рослинної клітини, рослини, або рослинної тканини можу бути збільшена збільшенням кількості специфічного протеїну, наприклад, протеїну резистентності у вказаному організмі. "Кількість протеїну" розуміється у значення кількості поліпептиду, переважно Rpi-blb2, в організмі, тканині, клітині, або клітинному компартменті. "Збільшення" кількості протеїну означає кількісне збільшення маси протеїну в організмі, тканині, клітині, або клітинному компартменті, наприклад, одним із описаних нижче способів, порівняно із диким типом цього самого роду або виду, до котрого цей метод не був застосований, за однакових усіх інших умов (таких як, наприклад, культуральних умов, віку рослин, та інше). Збільшення кількості, принаймні на 10 %, переважно принаймні 20 %, або принаймні 50 % особливо переважно принаймні 70 % або 90 % дуже особливо переважно принаймні 100 %, найбільш переважно принаймні 200 % та більше. "Збільшення" активності означає збільшення загальної активності протеїну в організмі, тканині, клітині, або клітинному компартменті, порівняно із диким типом цього самого роду або виду, до котрого цей метод не був застосований, за однакових усіх інших умов (таких як, наприклад, культуральних умов, віку рослин, та інше). Збільшення кількості, принаймні на 10 %, переважно принаймні 20 %, або принаймні 50 % особливо переважно принаймні 70 % або 90 % дуже особливо переважно принаймні 100 %, найбільш переважно принаймні 200 % та більше. В контексті цього, ефективність резистентності до патогену може коливатися я більший або менший бік порівняно із значенням отриманим, коли збільшувався один із протеїнів Rpi-blb2 приведених на SEQ ID NO: 2 або 4. Переважні функціональні еквіваленти є такими, в котрих ефективність патогенної резистентності, вимірювана, наприклад, ефективністю пенетрації патогену або як описано тут, відрізняється але не більш ніж на 50 %, переважно на 25 %, особливо переважно на 10 % порівняно зі значеннями отриманими відновленням протеїну Rpiblb2 приведеного на SEQ ID NO: 2 або 4. Особливо переважними є такі послідовності, де підвищення підвищує ефективність патогенної резистентності, більш ніж на 50 %, переважно на 100 %, особливо переважно на 500 % дуже особливо переважно на 1000 % порівняно зі значеннями отриманими відновленням одного із протеїну Rpi-blb2 приведеного на SEQ ID NO: 2 або 4. Будь-які порівняння переважно проводяться за аналогічних умов. "Аналогічні умови" означає, що всі умови, такі як наприклад, умови культивування або вирощування, умови аналізів (такі як, буфер, температура, субстрати, концентрація патогену та подібне) підтримуються ідентичними між експериментами, для порівняння та відрізняються лише послідовністю порівнюваних поліпептидів Rpi-blb2, їхнім організмом походження та якщо можливо, патогеном. При виборі патогену береться до уваги видова специфічність, оскільки кожне порівняння потребує того щоб патоген, який є найбільш подібний до іншого еквіваленту. Завдяки збільшеній активності Rpi-blb2, зростає резистентність рослини або її частини. В переважній реалізації, спосіб даного винаходу призводить до зниження індексу споруляції принаймні на 30 %, після інфікування P.infestans, порівняно із диким типом, більш переважно зниження на 50 %, значно переважно на 70 %, більш значно переважно більш ніж на 80 %, більш переважно на 85 % та 90 %. Най більш переважне на 95 % або більше. Відповідно, даний винахід також стосується згаданого полінуклеотиду даного винаходу, як визначено вище, що кодує білок Rpi-blb2, який включає молекулу нуклеїнової кислоти, вибрану з групи, яка складається з: a) молекул нуклеїнової кислоти, які кодують, принаймні готову форму поліпептиду, приведеного на SEQ ID NO: 2 або 4; b) молекул нуклеїнової кислоти, які включають кодуючу послідовність приведену на SEQ ID NO: 1 або 3, або 5 або 6, що кодують, принаймні готову форму поліпептиду; c) молекул нуклеїнової кислоти, чия нуклеотидна послідовність внаслідок виродженості генетичного коду вироджена у нуклеотидну послідовність (а) або (b); 16 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 d) молекул нуклеїнової кислоти, які кодують поліпептид, що походить з поліпептиду, кодованого полінуклеотидом шляхом заміщення, делеції та/або додавання однієї або більше амінокислот з(до) амінокислотної послідовності поліпептиду, кодованого полінуклеотидом від (а) до (с); e) молекул нуклеїнової кислоти, що кодує поліпептид, послідовність котрого має ідентичність 70 % або більше до амінокислотної послідовності поліпептиду, кодованого молекулою нуклеїнової кислоти (а) або (b); f) молекул нуклеїнової кислоти, які включають фрагмент або епітоп-несучу частину поліпептиду, кодованого будь-якою молекулою нуклеїнової кислоти від (а) по (e); (g) молекули нуклеїнової кислоти, що включає полінуклеотид, який має послідовність молекули нуклеїнової кислоти, що є ампліфікованою з бібліотеки нуклеїнової кислоти при використанні праймера, як представлено у Таблиці 3b; 3d, переважно ARF1F або ARF1R; h) молекул нуклеїнової кислоти, що кодують фрагмент, що починається амінокислотами: 1, 30, 50, 100, 200, 300, 500 або 1000 та закінчується амінокислотами: 1276, 1000, 500, 300, 200, 50, 30 або 1 поліпептиду, кодованого будь-якою із від (а) до (g); і) молекул нуклеїнової кислоти, що включають принаймні 20 нуклеотидів будь-якого з полінуклеотидів від (а) до (d); j) молекул нуклеїнової кислоти, що кодує поліпептид, який розпізнається моноклональними антитілами, які одержано до поліпептиду кодованого будь-якою з молекул нуклеїнової кислоти від (а) до (h); k) молекул нуклеїнової кислоти, одержуваних скринінгом відповідної бібліотеки за жорстких умов, за допомогою зонда, який має послідовність будь-якої з молекул нуклеїнової кислоти від (а) до (j) або його фрагменту із 15, переважно 30, 60, 90 або більше нуклеотидів; та l) молекул нуклеїнової кислоти, комплементарна послідовність котрих гібридизується за жорстких умов із будь-якою з молекул нуклеїнової кислоти від (а) до (k); або комплементарного ланцюга будь-якої з молекул нуклеїнової кислоти згідно з (а) - (l); або ті, що кодують поліпептид кодований сегментом або лінкерною групою 6 Solanum bulbocastanum який виділяється сумісно із маркером з таблиці 3а або 3b та який опосередковує резистентність до патогенів, зокрема, до патогенів вибраних з групи типу Oomycetes; В одному з втілень, полінуклеотид способу, згідно винаходу не складається з послідовності зображеної в Seq ID NO.: 7 та/або 9 та/або не складається з послідовності нуклеїнової кислоти, що кодує протеїн зображений в Seq ID NO.: 8 та/або 10. В одному з втілень, полінуклеотид способу, згідно винаходу не складається з послідовності молекули нуклеїнової кислоти Міl.1 або Mil.2 та/або молекули нуклеїнової кислоти, що кодує протеїн Міl.1 або Mil.2. Таким чином, в одному з втілень, полінуклеотид способу, згідно винаходу може не складатися із послідовностей приведених Rossi et al. 1998, PNAS USA 95:9750-9754, Milligan et al., 1998. Plant Cell 10:1307-1319; та/або WO98/06750. В наступній реалізації, полінуклеотид даного винаходу походить або виділений із геному організму обраного із групи, що складається із, Menyanthaceae, Solanaceae, Sclerophylacaceae, Duckeodendraceae, Goetzeaceae, Convolvulaceae, Cuscutaceae, Polemoniaceae, та Hydrophyllaceae згідно Systema Naturae 2000, Brands, S.J., Amsterdam або має походження з них, більш переважно вони вибрані з групи, що складається з, Atropa, Browallia, Brunfelsia, Capsicum, Cestrum, Cyphomandra, Datura, Fabiana, Franciscea, Hyoscyamus, Lycium, Mandragora, Nicandra, Nicotiana, Petunia, Physalis, Schizanthus та Solanum згідно Systema Naturae 2000, Brands, S.J., Amsterdam або має походження з них, ще більш переважно вони вибрані з групи, що складається з сімейства Solanaceae, переважно S.bulbocastanum, картоплі (S.tuberosum), помідорів (S. lycopersicum), петунії, томатного дерева (S.betaceum), динна груша (S. muricatum) та баклажан (S. melongena). Ще більш переважними є помідор або картопля або S.bulbocastanum як джерело полінуклеотидів для даного винаходу. Найбільш переважним джерелом є S.bulbocastanum. Rpi-blb2 був виділений із матеріалу S.tuberosum одержаного в АВРТ. Таким чином, з точки зору таксономії описаний Rpi-blb2 має також походження із S.tuberosum. Однак, ген був присутнім на інтрогресійному фрагменті, який ймовірно походить із S.bulbocastanum. Багато сортів S.tuberosum мають інтрогресійні фрагменти із видів Solanum, а не лише S.tuberosum. Тому, S.tuberosum може, згідно таксономічної системи також бути джерелом полінуклеотидів даного винаходу, зокрема тих, що походять з АВРТ S.tuberosum, а також інших різновидів видів Solanum одержаних подібним чином. Відповідно, в іншій реалізації, полінуклеотид винаходу одержаний з S.tuberosum. 17 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 Полінуклеотид даного винаходу, наприклад, молекула нуклеїнової кислоти, яка має нуклеотидну послідовність SEQ ID No: 1 або 3, або 5, або 6, або її частину, можу бути виділена стандартними молекулярно-біологічними методиками та наведена тут інформація про послідовність. Наприклад, Rpi-blb2 кДНК може бути виділена з бібліотеки, за допомогою всієї або частини однієї із послідовностей полінуклеотиду даного винаходу, в якості гібридизаційного зонду та стандартними методиками гібридизації (наприклад, як описано в Sambrook et al., Molecular Cloning: A Laboratory Manual. 2nd, ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). Більш того, полінуклеотид охоплює цілу або частину послідовностей полінуклеотиду даного винаходу, які можуть бути виділені за допомогою реакції полімеразного ланцюга з використанням праймерів виготовлених на основі цієї послідовності (наприклад, молекула нуклеїнової кислоти, які включає цілу послідовностей полінуклеотиду даного винаходу) Наприклад, мРНК може бути виділена з клітин, наприклад, S.bulbocastanum або іншої рослини (наприклад, методом гуанідін-тіоцианатної екстракції за Chirgwin et al.(1979) Biochemistry 18: 5294-5299) та кДНК може бути одержана з використанням реверсної транскриптази (наприклад, Moloney MLV реверсна транскриптаза, виробництва Gibco/BRL, Bethesda, MD; або AMV реверсна транскриптаза, виробництва Seikagaku America, Inc., St. Petersburg, FL). Синтетичний олінуклеотид даного винаходу, являє собою нуклеотидну послідовність, яка гібридизує, переважно гібридизує за жорстких умов, як визначено тут, до однієї з нуклеотидних послідовностей приведених в SEQ ID No: 1 або 3, або 5, або 6, або її частини. Полінуклеотид даного винаходу може бути ампліфікований з використанням кДНК, або геномної ДНК як шаблону та відповідних олігонуклеотидних праймерів згідно стандартної ампліфікаційної методики PLR. Ампліфікований таким чином нуклеотид може бути клонований у відповідний вектор та описаний за допомогою аналізу послідовності ДНК. Крім того, олігонуклеотиди, що відповідають нуклеотидній послідовності Rpi-blb2 можуть бути одержані стандартною методикою синтезу, наприклад, з використанням автоматизованого синтезатора ДНК. В реалізації даного винаходу, протеїн Rpi-blb2 кодується сегментом хромосоми 6 або лінкерної групи 6 Solanum bulbocastanum або S.tuberosum: Також даний винахід включає сегмент хромосоми 6 або лінкерної групи 6 Solanum bulbocastanum або S.tuberosum. В одній з переважних реалізацій способу даного винаходу, експресований протеїн Rpi-blb2 кодується полінуклеотидом, який включає сегмент хромосоми 6 або лінкерної групи 6 Solanum bulbocastanum. Переважно, згаданий сегмент є групою, яка включає інші cis-активний елемент, наприклад, промотори, енхансери, сайти зв'язування, та ін. або trans-активні елементи, такі як кофактори або інші протеїни резистентності, які несуть підвищену резистентність. Геномні фрагменти, які включають ген Rpi-blb2 та інші регуляторні елементи приведено на ID NO.: 5 та 6. Фахівцю відомо, як можна одержати хромосомний сегмент, наприклад, клонуванням хромосомних фрагментів у BACs, як наприклад, у Song, 2003, PNAS 100 (16), 9128 або описано тут та в цитованих тут посиланнях. Відповідно, до наступної реалізації, полінуклеотид даного винаходу, або полінуклеотид, який кодує протеїн Rpi-blb2 косегрегує змаркером, вибраним з Таблиці 3а, або включає сайт реплікації або сайт гібридизації для вказаного маркера. Як докладно описано в прикладах, резистентність до P.infestans може бути картована за допомогою маркерів приведених в Таблиці 3а або ЗЬ. Чим ближче розташований маркер до гену, тим вищий відсоток ліній, наприклад, похідних клонів, в котрих ген косегрегує із вказаним маркером. Відповідно, в переважній реалізації, полінуклеотид даного винаходу косегрегує з маркером Е40М58, СТ119 та/або СТ216. В іншій реалізації даний винахід стосується способу одержання рекомбінантного вектора, що включає інсерцію полінуклеотиду згідно даного винаходу у вектор або інсерцію вказаного полінуклеотиду та додаткового білка резистентності у вектор. Відповідно, в іншій реалізації даного винаходу, даний винахід стосується вектора, що включає полінуклеотид даного винаходу або вказаний полінуклеотиду та додатковий ген резистентності одержаний за способом даного винаходу. Я вжито тут, термін "вектор" вказує на молекулу нуклеїнової кислоти, здатну транспортувати полінуклеотид до якої він був приєднаний. Одним з типів векторів є "плазміда", яка є дволанцюговою кільцевою ДНК в котру може бути ліговано додатковий сегмент ДНК. Іншим типом вектору є вірусний вектор, в якому додаткові сегменти ДНК або РНК можуть бути ліговані у вірусний геном. 18 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Певні вектори є здатними до самостійної реплікації в клітині хазяїні в котру вони введені (наприклад, бактеріальні вектори, які мають бактеріальне походження реплікації та епісомальні вектори ссавців). Інші вектори (такі як, неепісомальні вектори ссавців) інтегровані в геном клітини хазяїна при введені до геному хазяїна, й таким чином реплікуються в усьому геномі хазяїна. Більш того, певні вектори здатні спрямовувати експресію генів, з котрими вони функціонально зв'язані. Такі вектори тут називаються "експресійні вектори". В загальному випадку, "експресійні вектори", що використовуються в методах рекомбінантної ДНК часто є плазмідами. В даному описі, "плазміда" та "вектор" можуть використовуватися взаємозамінно, оскільки плазміда, є найбільш застосовною формою вектора. Однак, винахід включає й наступні форми експресійних векторів, такі як вірусні вектори, (наприклад, реплікація дефектних ретровірусів, аденовіруси та адено-асоційовані віруси), які слугують еквіваклентним функціям. Даний винахід, також стосується космід, вірусів, бактеріофагів та інших векторів, які звичайно використовуються в генетичній інженерії, які містять молекулу нуклеїнової кислоти, згідно даного винаходу. Методи, які відомі фахівцям можуть бути використані для конструювання різноманітних плазмід та векторів; дивись наприклад, методики описані в Sambrook, Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory (1989) N.Y. та Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1989). Також, молекули нуклеїнової кислоти та вектори винаходу можуть бути введені в ліпосоми для доставки в клітини мішені. В іншій реалізації, вектор даного винаходу або спосіб даного винаходу відрізняється тим, що полінуклеотид, що кодує білок Rpi-blb2, або додатковий білок резистентності, є функціонально зв'язаним з послідовностями контролю експресії, та/або є функціонально зв'язаним з послідовністю нуклеїнової кислоти, яка кодує трансгенний сигнал регуляції експресії, що дозволяє здійснювати експресію в прокаріотичних або еукаріотичних хазяйських клітинах. В переважній реалізації, даний винахід стосується вектору даного винаходу або способу даного винаходу, в якому полінуклеотид, що кодує протеїн Rpi-blb2, та/або такий, що кодує додатковий білок резистентності, є функціонально зв'язаним з послідовностями контролю експресії, що мають походження від тих самих видів, що й полінуклеотид, який кодує протеїн Rpi-blb2, та/або додатковий білок резистентності. В цьому випадку, молекула нуклеїнової кислоти, згідно винаходу експресується в клітині, й принципово можливо таким чином модифікувати кодуючу послідовність, що протеїн знаходитиметься в будь-якому компартменті рослинної клітини. Вони включають ядро, ендоплазматичний ретикулум, вакуоль, мітохондрії, пластиди, такі як амілопласти, хлоропласти, хромопласти, апопласти, позаклітинне середовище, олійні тільця, пероксисоми та інші компартменти рослинної клітини (для довідки дивись Kermode, Crit. Rev. Plant Sci. 15, 4 (1996), 285-423 та цитовані там посилання). Полінуклеотид може бути функціонально злитий із придатним полінуклеотидом, наприклад, вектором, що кодує сигнал для транспорту в бажаний компартмент. В іншій переважній реалізації, даний винахід стосується вектору в якому полінуклеотид, даного винаходу є функціонально зв'язаним з послідовностями контролю експресії, що дозволяє здійснювати експресію в прокаріотичних або еукаріотичних хазяйських клітинах. Природа цих регуляторних послідовностей різниться залежно від організму хазяїна. В прокаріотах, регуляторні послідовності звичайно включають промотор, рибосомальний сайт зв'язування та термінатори. В прокаріотах, регуляторні послідовності звичайно включають промотори, термінатори, та в деяких випадках енхансери, трансактиватори або фактори транскрипції. Термін "регуляторна послідовність" повинен включати, як мінімум компоненти, присутність котрих є необхідною для експресії, та можуть також включати додаткові корисні компоненти. Термін "функціонально зв'язаний" стосується безпосередньої близькості розташування, при якій компоненти, як вони описані знаходяться в зв'язку, що дозволяє їм функціонувати певним чином. Регуляторна послідовність "функціонально зв'язана" із кодуючою послідовністю є приєднаною до неї таким чином, що експресія кодуючої послідовності досягається за умов сумісних із регуляторною послідовністю. Якщо регуляторна послідовність є промотором, є очевидним для фахівця, що використовується дволанцюгова нуклеїнова кислота. Функціональне зв'язування розуміється в значенні, наприклад, послідовного розташування промотору послідовності нуклеїнової кислоти для експресування, якщо можливо, наступного регуляторного елементу, як наприклад, термінатору таким чином, що кожні регуляторні елементи можуть задовольняти свою функцію коли рекомбінантно експресується послідовність нуклеїнової кислоти, залежно від розташування послідовності нуклеїнової кислоти, по 19 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 відношенню до смислової та антисмислової РНК. В цьому випадку, прямий зв'язок в хімічному сенсі обов'язково не потрібен. Генетичні регуляторні послідовності, такі як, наприклад, енхансерні послідовності, також спрямовують свою функцію на цільову послідовність з місця, що знаходиться досить далеко або спрямовується з інших молекул ДНК. Переважне розташування є таким, якщо послідовність нуклеїнової кислоти, яка повинна експресуватися рекомбінантно, розташована після послідовності, що діє як промотор, таким чином, що дві послідовності зв'язані ковалентно між собою. Відстань між промоторною послідовністю та послідовністю нуклеїнової кислоти, що має експресуватися рекомбінантно, переважно менша за 200 пар основ, особливо переважно менша за 100 пар основ, дуже особливо переважно менша за 50 пар основ. Функціональний зв'язок, та експресійна касета, можуть бути одержані звичайною рекомбінацією та клонувальними методиками, як описано, наприклад, в Maniatis T, Fritsch EF and Sambrook J (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY), in Silhavy TJ, Berman ML and Enquist LW (1984) Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor (NY), in Ausubel FM et al.(1987) Current Protocols in Molecular Biology, Greene Publishing Assoc, and Wiley Interscience and in Gelvin et al.(1990). In: Plant Molecular Biology Manual. Однак, інші послідовності які, наприклад, діють як лінкер із специфічними сайтами розщеплення ферментів рестрикції, або сигнальні пептиди, також можуть бути розташовані між двома послідовностями. Вставка послідовностей може вести також до експресії злитих протеїнів. Переважно, експресійна касета, що складається із зв'язаних промотору та послідовності нуклеїнової кислоти, що має експресуватися, може існувати у вормі інтегрованій у вектор та бути вставденою в частину рослинного геному, наприклад, трансформацією. Такі регуляторні послідовності описані, наприклад, в Goeddel; Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990) or see: Gruber and Crosby, in: Methods in Plant Molecular Biology and Biotechnology, CRC Press, Boca Raton, Florida, eds.:Glick and Thompson, Chapter 7, 89-108 включаючи посилання в них. Регуляторні послідовності, включають ті що є напряму визначальними для експресії нуклеотидної послідовності в багатьох типах клітин хазяїнів та ті, що прямо експресують нуклеотидну послідовність лише в окремих типах клітин за певних умов. Фахівцями може бути визначено, що створення експресійного вектору може залежати від таких факторів як вибір для трансформації клітини хазяїна, рівень експресії бажаного протеїну та інше. Експресійні вектори винаходу можуть бути введені в клітину хазяїна, й таким чином продукувати протеїни або пептиди, включаючи злиті протеїни або пептиди, кодовані полінуклеотидами, як тут описано. Рекомбінантні вектори експресії даного винаходу можуть бути сконструйовані для експресії згаданих протеїнів резистентності, переважно Rpi-blb2, в прокаріотичних та еукаріотичних клітинах. Наприклад, гени, що кодують полінуклеотид даного винаходу можу бути експресований в бактеріальних клітинах, таких як E.coli, C.glutamicum, Agrobacterium tumefaciens, клітинах комах (із застосуванням бакуловірусного вектору експресії), клітинах дріжджів або інших грибів (дивись Romanos, (1992), Yeast 8: 423-488; van den Hondel, (1991) J.W.Bennet & LLLasure, eds., p.396-428: Academic Press: San Diego; and van den Hondel, (1991) in: Applied Molecular Genetics of Fungi, Peberdy, eds., p.1-28, Cambridge University Press: Cambridge), algae (Falciatore et al., 1999, Marine Biotechnology. 1, 3:239-251), та кліфтинах багатоклітинних рослин (дивись Schmidt, R.(1988), Plant Cell Rep.: 583-586); Plant Molecular Biology and Biotechnology, С Press, Boca Raton, Florida, chapter 6/7, S.71-119 (1993); F.F.White, B.Jenes et al., Techniques for Gene Transfer, in: Transgenic Plants, Vol. 1, Engineering and Utilization, eds.:Kung und RWu, Academic Press (1993), 128-43; Potrykus, Annu. Rev. Plant Physiol. Plant Molec. Biol. 42 (1991), 205-225 (та посилання на процитовані в них) або кітинах ссавців. Також, придатні клітини хазяїни обговорені в Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Також, рекомбінантний вектор експресії може бути тринскрибований та трансльований in vitro, наприклад, з використанням регуляторних послідовностей промотору Т7 та полімерази 17. Експресія протеїнів в прокаріотах часто проводиться із векторами, що містять визначальні або ідуковні промотори, які спрямовують експресію як злитих так незлитих протеїнів. Злиті вектори додають число амінокислот до протеїну, що ними кодуються, звичайно до амінного кінця рекомбінантного протеїну але також і до С-кінця або злиті із придатною ланкою в протеїні. Такі злиті вектори звичайно слугують трьом цілям: 1) збільшення експресії рекомбінантного протеїну; 2) збільшення розчинності рекомбінантного протеїну; та 3) сприяти очищенню рекомбінантного протеїну, діючи як ліганд у афінній очистці. Також, злитий вектор може 20 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 кодувати додаткові протеїни, які які експресують підтримку та збільшення активності Rpi-blb2 або резистентності рослини до рослинних патогенів, наприклад, інших протеїнів резистентності. Часто, у злитих експресійних векторах, сайт каталітичного розщеплення введено до злитої частини та рекомбінантного протеїну для можливості розділення рекомбінантного протеїну від злитої частини, наступним очищенням злитого протеїну. Такі ферменти, та їхні сдочірні розпізнавальні послідовності, включають Фактор Ха, тромбін та ентерокіназу. Тапові злиті експресійні вектори включають pGEX (Pharmacia Biotech Inc; Smith, D.B.and Johnson, K.S.(1988) Gene 67:31-40), pMAL (New England Biolabs, Beverly, MA) та pRIT5 (Pharmacia, Piscataway, NJ). Приклади, придатних індуковних незлитих експресійних векторів Е.соlі. включають pTrc (Amann (1988) Gene 69:301-315) and pET 11d (Studier et al., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, California (1990) 60-89 Одним з підходів до максималізації експресії рекомбінантного протеїну є експресування протеїну в бактерії хазяїні із неповною здатністю до протеолітичного розщеплення рекомбінантного протеїну (Gottesman, S., Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, California (1990) 119-128). Іншим підходом є зміна послідовності нуклеїнової кислоти, нуклеїнової кислоти, що буде вставлена в експресійний вектор, так щоб окремі кодони кожної амінокислоти, тули такими як переважно застосовуються у вибраній для експресії бактерії, тікій як Е.соlі або C.glutamicum (Wada et al.(1992) Nucleic Acids Res.20:21112118). Така зміна послідовностей нуклеїнової кислоти даного винаходу може бути виконана стандартною методикою синтезу ДНК. Також, вектор може бути дріжджовим експресійним вектором. Приклади векторів для експресії у дріжджах S.cerivisae включають pYepSed (Baldari, et al., (1987) Embo J.6:229-234), pMFa (Kurjan and Herskowitz, (1982) Cell 30:933-943), pJRY88 (Schultz et al., (1987) Gene 54:113123), та pYES2 (Invitrogen Corporation, San Diego, CA). Переважно, полінуклеотид даного винаходу або описаний тут є функціонально зв'язаний із рослинною послідовністю керування експресією, наприклад, експресійними касетами. Рослинні експресійні касети, переважно, містять регуляторні послідовності здатні керувати генною експресією в рослинних клітинах, та функціонально зв'язані таким чином, що кожна послідовність задовольняє своїй функції, такій як термінація транскрипції, наприклад, сигнали поліаденілування. Переважні сигнали поліаденілування мають походження із тДНК Agrobacterium tumefaciens такої як ген 3, відомий як октопін-синтаза Ті-плазміди рТіАСН5 (Gielen et al., EMBO J.3 (1984), 835 ff) або її функціональні еквіваленти але також є придатними усі функціонально активні в рослинах термінатори. Так як експресія гену рослини дуже часто не обмежена рівнями транскрипції то й рослинна експресійна касета переважно містить інші функціонально зв'язані послідовності, такі як ' енхансери трансляції, такі як overdrive-послідовність, яка має 5 -нетрансльовну лідерну послідовність з вірусу мозаїки тютюну, яка посилює співвідношення протеїну до РНК (Gal-lie et al 1987, Nucl. Acids Research 15:8693-8711). Також, описаний тут полінуклеотид може бути функціонально зв'язаний із відповідним промотором, що несе ген експресії специфічного за часовою, клітинною або тканинною ознакою. Переважними є промотори, що керують конститутивною експресією (Benfey et al., EMBO J.8 (1989) 2195-2202) як в одержаних із рослинних вірусів, таких як 35S CAMV (Franck et al., Cell 21(1980) 285-294), 19S CaMV (також дивись US5352605 та WO8402913) або рослинні промотори такі як малі субодиниці Rubisco описані в US4962028. Термін рослинно-специфічні промотори розуміється в значенні, в принципі, як будь-який промотор, здатний спрямовувати експресію генів, зокрема чужорідних генів, в рослинах або частинах рослин, рослинних клітинах, рослинних тканинах або рослинних культурах. В цьому контексті, експресія може бути, наприклад, конститутивною, індуковною або залежною від розвитку. Наступне є переважним: a) Конститутивні промотори Переважні вектори, які роблять можливим конститутивну експресію в рослинах (Benfey et al.(1989) EMBO J 8:2195-2202). "Конститутивний" промотор розуміється в значенні, що цей промотор забезпечує експресію у великій кількості, переважно, в усіх тканинах протягом значного періоду розвитку рослини, переважно на всіх стадіях розвитку рослини. Зокрема, переважно застосовується, рослинний промотор або промотор одержаний із рослинного вірусу. Зокрема, переважним є промотор 35S-транскрипт вірусу мозаїки цвітної капусти CaMV (Franck et al.(1980) Cell 21:285-294; Odell et al.(1985) Nature 313:810-812; 21 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Shewmaker et al.(1985) Virology 140:281-288; Gardner et al.(1986) Plant Моl Biol 6:221-228) або 19S CaMV промотор (US 5,352,605; WO 84/02913; Benfey et al.(1989) EMBO J 8:2195-2202). Іншим придатним конститутивним промотором є промотор "мала субодиниця Rubisco (SSU)" (US 4,962,028), промотор легумін В (GenBank Ace. No.X03677), промотор нопалін синтази Agrobacterium, подвійний промотор TR, Agrobacterium OCS (котопін-синтази) промотор, промотор убіквітину (Holtorf S et al.(1995) Plant Моl Biol 29:637-649), промотор убіквітину-1 (Christensen et al.(1992) Plant Моl Biol 18:675-689; Bruce et al.(1989) Proc Natl Acad Sci USA 86:9692-9696), промотор Smas, промотор циннаміл-алкоголь-дегідрогенази (US5683439), промотори вакуолярних субодиниць АТФ-ази або промотор багатого на пролін білку пшениці (WO91/13991), та інші промотори генів, конститутивна експресія котрих відома фахівцям. b) Тканинно-специфічні промотори Переважними також є промотори зі специфічністю до пильників, зав'язі, квітів, листя, пагонів, коренів та насіння. Насіннєво-специфічні промотори, такі як, наприклад, промотор фазеоліну (US 5,504,200; Bustos MM et al.(1989) Plant Cell 1(9):839-53), промотор гену 2S альбуміну (Joseffson LG et al.(1987) J.Biol.Chem 262:12196-12201), промотор легуміну (Shirsat A et al.(1989) Моl Gen.Genet 215(2): 326-331), промотор USP (невідомий промотор насіння) (Baumiein H et al.(1991) Моl Gen Genet 225(3):459-67), промотор гену напіну (US 5,608,152; Stalberg К et al.(1996) L Planta 199:515-519), промотор протеїну зв'язування сахарози (WO 00/26388) або промотор легуміну В4 (LeB4; BSumlein H et al.(1991) Моl Gen.Genet 225: 121-128; Baeumlein et al.(1992) Plant Journal 2(2):233-9; Fiedler U et al.(1995) Biotechnology (NY) 13(10):1090f), промотор олеозину Arabidopsis (WO 98/45461), промотор Все4 Brassica (WO 91/13980). Інші придатні насіннєво-специфічні промотори, які є промоторами генів, які кодують глютенін високої молекулярної маси (HMWG), гліадін, галуджуючий фермент, АДФ глюкоза-пірофосфатаза (АГФ-аза) або крохмаль-синтаза. Іншими переважними є промотори, які дозволяють насіннєво-специфічну експресію в однодольних, таких як кукурудза, ячмінь, пшениця, жито, рис та подібні. Наступне може бути переважним: промотор гену Ipt2 або Ipt1 (WO 95/15389, WO 95/23230) або промотор описаний в WO 99/16890 (промотори гордеїну, гену глютеїну, гену оризину, гену проламіну, гену гліадіну, гену глютеліну, гену зеїну, гену казіріну або гену секаліну). Клубеневі-, коренеплодні-, або корене-специфічні промотори, наприклад, палатиновий промотор класу І (В33), промотор інгібітору кутепсину D картоплі. Промотори специфічні до листя такий як цитозольний промотор FBPase картоплі (WO 97/05900), промотор Rubisco (ribulose1,5-bisphosphate carboxylase) SSU (small subunit) або промотор ST-LSI із картоплі (Stockhaus et al.(1989) EMBO J 8:2445-2451). Дуже особливо переважними є епідерміс-специфічні промотори, наприклад, промотор гену OXLP (оксалат-оксилаза-подібний протеїн) (Wei et al.(1998) Plant Моl. Biol. 36:101-112). Промотори специфічні до квітів такі як, наприклад, промотор фітоен-синтази (WO 92/16635) або промотор гену Р-rr (WO 98/22593). Інші специфічні промотори такий як промотор 5126 (US 5,689,049, US 5,689,051), промотор glob-l та промотор у-зеїну. с) Хімічно індуковні промотори Експресійна касета також може включати хімічно індуковні промотори (дивись статтю: Gatz et al.(1997) Annu Rev Plant Physiol Plant Моl Biol 48:89-108), за допомогою котрих може керуватися експресія ендогенного гену в рослині в певний момент часу. Такі промотори як, наприклад, промотор PRP1 (Ward et al.(1993) Plant Моl Biol 22:361-366), промотор індуковний саліциловою кислотою (WO 95/19443), промотор індуковний бензолсульфонамідом (ЕР 0 388 186), промотор індуковний тетрацикліном (Gatz et al.(1992) Plant J 2:397-404), промотор індуковний абсцизовою кислотою (ЕР 0 335 528) або промотор індуковний етанол- або циклогексаноном (WO 93/21334) можуть використовуватися й подібнії. d) Промотори індуковні стресом або патогеном Наступні переважні промотори, це індуковні біотичним або абіотичним стресом, такий як, наприклад, індуковний патогеном промотор гену PRP1 (Ward et al.(1993) Plant Моl Biol 22:361366), промотор індуковний високою температурою hsp70 або hsp80 (US 5,187,267), альфаамілазний промотор індуковний низькою температурою (WO 96/12814), промотор індуковний світлом PPDK, або промотор індуковний пораненням pinll (ЕР375091). Промотори індуковні патогеном охоплюють ті, що індукуються внаслідок інфікування патогеном, такі як, наприклад, гени протеїнів PR, SAR протеїнів, р-1,3-глюканази, хітінази та 22 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 подібні (for example Redolfi et al.(1983) Neth J Plant Pathol 89:245-254; Uknes, et al.(1992) The Plant Cell 4:645-656; Van Loon (1985) Plant Моl Virol 4:111-116; Marineau et al.(1987) Plant Моl Biol 9:335-342; Mat-ton et al.(1987) Molecular Plant-Microbe Interactions 2:325-342; Somssich et al.(1986) Proc Natl Acad Sci USA 83:2427-2430; Somssich et al.(1988) Моl Gen Genetics 2:93-98; Chen et al.(1996) Plant J 10:955-966; Zhang and Sing (1994) Proc Natl Acad Sci USA 91:2507-2511; Warner, etal.(1993) Plant J 3:191-201; Siebertz et al.(1989) Plant Cell 1:961-968(1989). Також охоплюються промотори індуковний пораненням такияк гену pinll (Ryan (1990) Ann Rev Phytopath 28:425-449; Duan et al.(1996) Nat Biotech 14:494-498), °F the wunl and wun2 gene (US 5,428,148), або гену winl та win2 (Stanford et al.(1989) Моl Gen Genet 215:200-208), системіну (McGurl et al.(1992) Science 225:1570-1573), або гену WIP1 (Rohmeieret al.(1993) Plant Моl Biol 22:783-792; Eckelkamp et al.(1993) FEBS Letters 323:73-76), гену MPI (Corderok et al.(1994) The Plant J 6(2): 141-150) та подібні. Промотори залежні від розвитку Інші придатні промоторами є, наприклад, промотори специфічні до дозрівання плодів, такі як наприклад, промотор помідорів специфічний до дозрівання плодів (WO 94/21794, ЕР 409625). Промотори залежні від розвитку включають частково тканинно-специфічні промотори, оскільки певні тканини за природою є залежними від розвитку типами. Може бути бажаним, щоб поліпептид даного винаходу був лише активним або мав лише підвищену активність в тканинах, які є трансфікованими або пенетрованими згадуваним тут патогеном. Особливо переважними є конститутивні промотори та листо- та/або пагоноспецифічні та епідерміс-специфічні промотори, найбільш переважними є індуковні патогеном та епідерміс-специфічні промотори. Також переважними є природні промотори, які наприклад включають геномні фрагменти, приведені на ID NO.: 5 та 6 Також, інші промотори можуть бути функціонально зв'язані з послідовністю нуклеїнової кислоти, що має експресуватися, які промотори роблять можливою експресію в інших тканинах або в інших організмах, таких як бактерія Е.соlі. Придатними рослинними промоторами є, в принципі, усі описані вище промотори. Термін "послідовності генетичного контролю" розуміється в широкому значенні та вказує також на всі ті послідовності, які мають вплив на матеріалізацію або функцію експресійної касети, згідно даного винаходу. Наприклад, послідовності генетичного контролю модифікують транскрипцію та трансляцію в прокаріотичних або еукаріотичних організмах. Переважно, експресійні касети, згідно даного винаходу, охоплюють промотори зі специфічністю до ембріонального епідермісу та/або квітів, 5'-вище послідовності нуклеїнової кислоти для рекомбінантної експресії та 3'-нижче термінальної послідовності, як додаткової послідовності генетичного контролю, якщо можливо, інших додаткових регуляторних елементів, в кожному випадку функціонально зв'язаному до послідовності нуклеїнової кислоти, що має рекомбінантно експресуватися. Послідовності генетичного контролю також охоплюють інші промотори, промоторні елементи або мінімальні промотори, усі з яких можуть модифікувати властивості, що спрямовують експресію. Таким чином, наприклад, тканинно-специфічна експресія може додатково залежати від певних стресорних чинників, що належать до послідовності генетичного контролю. Такі елементи було описано, як наприклад, водний стрес, абсцизова кислота (Lam E and Chua NH, J Biol Chem 1991; 266(26): 17131-17135) та тепловий стрес (Schoffl F et al., Molecular & General Genetics 217(2-3):246-53, 1989). Іншими переважними керуючими послідовностями є, наприклад, Грам-позитивні промотори ату та SP02, та дріжджові або грибові промотори ADC1, MFa, АС, Р-60, CYC1, GAPDH, TEF, rp28, ADH. В принципі, усі природні промотори зі своїми регуляторними послідовностями, такими як згадані вище, можуть використовуватися в способі згідно даного винаходу Також, переважно можуть використовуватися синтетичні промотори. Послідовності генетичного контролю крім того можуть охоплювати нетрансльовні 5'-регіони, інтрони або некодуючі 3'-регіон генів, такий як, наприклад, інтрон актину-1, або інтрони 1, 2, та 6 Adh1-S (для довідки: The Maize Handbook, Chapter 116, Freeling and Walbot, Eds., Springer, New York (1994)). Також було показано, що вони можуть грати значну роль в регуляції генної експресії. Відповідно, було показано, що нетрансльовні 5'-послідовності можуть посилювати тимчасову експресію гетерологічних генів. Прикладами енхансерів трансляції які можуть згаданий є 5'-лідерна послідовність вірусу мозаїки тютюну (Gallie et al.(1987) Nucl Acids Res 15:8693-8711) та подібні. Також вони можуть сприяти тканинній специфічності (Rouster J et al.(1998) Plant J 15:435-440). 23 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 Експресійна касета переважно може включати одну або більше відомих енхансерних послідовностей, функціонально зв'язаних із промотором, який робить можливою підвищену рекомбінантну експресію послідовності нуклеїнової кислоти. Інші переважні послідовності, такі як додаткові регуляторні елементи або термінатори, можуть бути вставлені до 3' кінця послідовності нуклеїнової кислоти, що має рекомбінантно експресуватися. Одна або більше копій послідовності нуклеїнової кислоти можуть бути представлені в генетичному конструкті для рекомбінантної експресії. В іншій реалізації природна термінаторна послідовність представлена геномним фрагментом, приведеним на ID NO.: 5 та/або 6. Сигнали поліаденілування, які є придатними на роль керуючих послідовностей є рослинними сигналами поліаденілування, переважно ті, які в основному відносяться до сигналів поліаденілування Т-ДНК із Agrobacterium tumefaciens, зокрема 3' гену Т-ДНК (октопінсинтази) плазміди Ті pTiACHS (Gielen et al.(1984) EMBO J 3:835 et seq.) або його функціонального еквівалентів. Приклади термінаторних послідовностей, які є особливо придатними є термінатор OCS (октопінсинтази) та термінатор NOS (нопалін-синтази). Керуючі послідовності, також можуть розумітися як такі, що роблять можливою гомологічну рекомбінацію або інсерцію в геном організму хазяїна або який дозволяє видалення з геному. У випадку гомологічної рекомбінації, наприклад, природний промотор певного гену може бути замінений на промотор зі специфічністю до ембріонального епідермісу та/або квітів. Методи, такі як технологія сге/lox дозволяють тканево-специфічне, якщо можливо, індуцибельне, видалення експресійної касети з геному організму хазяїна (Sauer В (1998) Methods. 14(4):38192). В цьому методі, специфічні фланкуючі послідовності (Іох послідовності), які пізніше дозволяють видалення за допомогою cre-рекомбінази, приєднані до цільового гену. Експресійна касета та вектори, що походять з неї можуть включати інші функціональні елементи. Термін "функціональний елемент" повинен розумітися в широкому сенсі й вказувати на всі елементи, які мають вплив на генерацію, ампліфікацію або функцію експресійних касет, векторів або трансгенних організмів, згідно даного винаходу. Наступне можу бути прийняте, як приклади але не обмеження: а) Селекційні маркери, які несуть резистентність до інгібітору метаболізму, такого як 2дезоксиглюкоза-6-фосфат (WO 98/45456), антибіотики або біоциди, переважно гербіциди, такі як, наприклад, канаміцин, G 418, блеоміцин або гігроміцин, або фосфінотріцин та подібні. Особливо переважними селекційними маркерами є ті, що несуть резистентність до гербіцидів. Як приклади можуть бути приведені наступні: послідовності ДНК, що кодують фосфінотріцинові ацетил-трансферази (PAT) та які інактивують інгібітори глютамін-трансферази (гени bar та pat), гени 5-енол-пірувілшикімат-3-фосфат синтази (гени EPSP-синтази), які несуть резистентність до Glyphosater (N-(фосфонометил)гліцину), ген дох, який кодує ферменти розщеплення Glyphosater (гліфозат-оксиредуктаза), ген deh (який кодує деалогеназу, яка інактивує далапон), сульфонілсечовина- та імідазолінон-інактивуючі ацетолактат-синтази, та гени bxn, які кодують бромоксиніл-розщеплюючий нітриладний фермент, ген aasa, який несе резистентність до антибіотику апектіноміцину, ген стрептоміцинової фосфотрансферази (SPT), який робить можливою резистентність до стрептоміцину, ген неоміцинової фосфотрансферази (NPTII), який несе резистентність до канаміцину або генецетидіну, ген гігроміцинової фосфотрансферази (НРТ), який опосередковує резистентність до гіроміцину, ген ацетолактазної синтази (ALS), який несе резистентність до сульфонілсечовинних гербіцидів (наприклад мутованих варіантів ALS із, наприклад, мутацією S4 та/або Hra). Репортерні гени, які кодують добре визначувані протеїни та, завдяки своїй кольоровій або ферментативній активності, роблять можливим визначення ефективності трансформації, сайту експресії або часу експресії. Дуже особливо переважними в цьому контексті є гени, що кодують репортерні протеїни (Schenborn Е, Groskreutz D.Mol Bio-technol.1999; 13(1):29-44) такі як зелений флуоресцентний протеїн (GFP) (Sheen et al.(1995) Plant Journal 8(5)777-784; Haseloff et al.(1997) Proc Natl Acad Sci USA 94(6):2122-2127; Reichel et al.(1996) Proc Natl Acad Sci USA 93(12):5888-5893; Tian et al.(1997) Plant Cell Rep 16:267-271; WO 97/41228; Chui WL et al.(1996) Curr Biol 6:325-330; Leffel SM et al.(1997) Biotechniques.23(5):912-8), хлорамфеніколтрансферазу, люциферазу (Ow et al.(1986) Science 234:856-859; Millar et al.(1992) Plant Моl Biol Rep 10:324414), ген екворину (Prasheret al.(1985) Biochem Biophys Res Commun 126(3): 1259-1268), Bгалактозидазу, ген локусу R (що кодує протеїн, який регулює вироблення пігментів антроцианіну (червоного забарвлення) в рослинних тканинах, й таким чином робить можливим прямий аналіз промоторної активності без додавання інших додаткових речовин або хромогенних субстратів) Dellaporta et al., In: Chromosome Structure and Function: Impact of New Concepts, 18th Stadler 24 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 60 Genetics Symposium, 11:263-282, 1988), (3-глюкоуронідаза є дуже особливо переважною (Jefferson et al., EMBO J. 1987, 6, 3901-3907). c) Походження реплікації, яке забезпечує ампліфікацію експресійних касет або векторів, згідно винаходу в, наприклад, Е.соІІ. Прикладами, які беруться до уваги є ORI (походження реплікації ДНК), pBR322 ori або Р15А ori (Sambrook et al.: Molecular Cloning. A Laboratory Manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989). d) Елементи, які є необхідними для рослинної трансформації, опосередкованої Agrobacterium, такі як наприклад, права та ліва границя Т-ДНК або vir-регіону. Для селектування клітин, які успішно пройшли гомологічну рекомбінацію, або іншими словами для селектування трансформованих клітин, як правило, необхідно додатково ввести селективний маркер, який несе резистентність до біоциду (наприклад, гербіциду), інгібітор метаболізму, такий як, 2-дезоксиглюкоза-6-фосфат (WO 98/45456) або антибіотик, для клітин, які успішно пройшли рекомбінацію. Селективні маркери дозволяють провести відокремлення трансформованих клітин від нетрансформованих (McCormick et al.(1986) Plant Ceil Reports 5:8184). Введення в експресійну касету, згідно даного винаходу, в організм або клітини, органи, тканини, частини або їхнє насіння (переважно в рослини або рослинні клітини, органи, тканини, частини або насіння) може переважно бути виконане за допомогою векторів які включають експресійні касети. Експресійна касета, може бути введена у вектор (наприклад, плазміду) за допомогою прийнятного сайту розщеплення. Отримана плазміда, спочатку вводиться у Е.соlі. Коректно трансформовані Е.соlі. селектують, вирощують та одержують рекомбінантні плазміди способами відомими досвідченому фахівцеві. Рестрикційний аналіз та секвенування може служити перевіркою етапу клонування. Іншими промоторами для експресії у специфічних рослинах є, наприклад, промотор гену напіну із насіння рапсу (US5608152), USP-промотор із Vicia faba (Baeumlein et al., Моl Gen Genet, 1991, 225 (3):459-67), олеозиновий промотор із Arabidopsis (W09845461), фазеоліновий промотор із Phaseolus vulgaris (US5504200), Все4-промотор із Brassica (W09113980) або легуміновий промотор В4 (LeB4; Baeumlein et al., 1992, Plant Journal, 2 (2):233-9) а також інша промотори, що несуть насінє-специфічну експресію в однодольних рослинах, таких як, кукурудза, ячмінь, пшениця, рис та інші. Придатними промоторами є: промотор гену Ipt2 або Ipt1 із ячменю (WO 95/15389 та WO 95/23230) або промотор описаний в WO 99/16890 (промотори із ячменю гену гордеїну, рисового гену глютеїну, рисового гену оризину, рисового гену проламіну, пшеничного гену гліадіну, пшеничного гену глютеліну, кукурудзяного гену зеїну, вівсяного гену глютеліну, гену казіріну із сорго або житнього гену секаліну). Також, полінуклеотид даного винаходу може бути клонований в експресійний вектор в антисмисловій орієнтації. Тобто, молекула ДНК є функціонально зв'язана з регуляторною послідовністю, таким чином, який забезпечує експресію (через транскрипцію молекули ДНК) молекули РНК, яка є антисмисловою до мРНК, кодованої полінуклеотидом даного винаходу. Регуляторні послідовності, функціонально зв'язані з нуклеїновою кислотою, клонованою в антисмисловій орієнтації, може бути вибраний, як такий, що забезпечує постійну експресію молекули антисмислової РНК в різних типах клітин, наприклад, вірусні промотори та/або енхансери або регуляторні послідовності можуть бути вибрані які спрямовують конститутивну, тканинно-специфічну або клітинно-специфічну експресію антисмислової РНК. Вектор антисмислової експресії може бути у вигляді рекомбінантної плазміди, плазміди або ослабленого вірусу, в котрих продукуються молекули антисмислової нуклеїнової кислоти під контролем високоефективного регуляторного регіону, активність котрого може бути визначена типом клітини в який введено вектор. Обговорення регуляції генетичної експресії за допомогою антисмислових генів дивись, Weintraub, H.et al., Antisense RNA as a molecular tool for genetic analysis, Reviews-Trends in Genetics, Vol. 1(1) 1986 and Моl et al., 1990, FEBS Letters 268:427430. В іншій реалізації даний винахід стосується способу одержання рекомбінантної клітини хазяїна, що включає введення вектору або полінуклеотиду даного винаходу або вказаного вектору або вказаного полінуклеотиду та вектору для експресії додаткового протеїну резистентності в клітину хазяїна. Векторна ДНК може бути введена в прокаріотичні або еукаріотичні клітини звичайними методами трансформації або трансфекції. Як вжито тут, терміни "трансформація" та "трансфекція", "кон'югація" та "трансдукція" повинні вказувати на різноманітні визнані методики введення чужорідної нуклеїнової кислоти (наприклад, ДНК), в клітину хазяїна, включаючи сумісне осадження фосфатом кальцію або хлоридом кальцію, DEAE-декстран-опосередкована трансфекція, ліпофекція, природна компетенція, хімічно опосередкована передача або 25 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 електропорація. Придатні способи трансформації або трансфекції клітин хазяїнів, включаючи рослинні клітини можуть бути знайдені в Sambrook, et al.(Molecular Cloning: A Laboratory Manual. 2nd, ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989) та інших лабораторних довідниках, таких як Methods in Molecular Biology, 1995, Vol. 44, Agrobacterium protocols, ed: Gartland and Davey, Humana Press, Totowa, New Jersey. Для стабільної трансфекції еукаріотичних клітин, відомо, що залежно від використаних вектору експресії та методики трансфекції, лише маленькі фракції клітин можуть інтегрувати чужорідну ДНК у свій геном. Для ідентифікації та селектування цих інтегрантів, ген, що кодує селективний маркер (наприклад, резистентність до антибіотиків) зазвичай вводиться в клітину хазяїна поряд із бажаним геном. Преважні селективні маркери включають ті, що несуть резистентність до ліків, таких як G418, гігроміцин та метотрексат або в рослини які несуть резистентність до такого гербіциду як гліфозат глуфозинат. Нуклеїнова кислота, що кодує селективний маркер може бути введена в клітину хазяїна в тому ж самому векторі, що кодує поліпептид даного винаходу або може бути введений в окремому векторі. Клітини, стабільно трансфіковані введеною нуклеїновою кислотою можуть бути ідентифіковані за допомогою, наприклад, хімічною селекцією (тобто, клітини, які мають включений селективний маркерний ген виживуть, тоді як інші клітини загинуть). Також можуть одержані клітини хазяїни, які мають селективні системи, які дозволяють регулювати експресію введеного гену. Наприклад, включення полінуклеотиду даного винаходу у вектор, розміщенням його під керування Іас-оперону дозволяє експресувати полінуклеотид лише в присутності IPTG. Такі регуляторні системи є добре відомими на практиці. Переважно, введена молекула нуклеїнової кислоти є чужорідною для даної клітини хазяїна. Під "чужорідною" розуміють, що ця молекула нуклеїнової кислоти є або гетерологічною по відношенню до клітини хазяїна, це означає, що вона одержана з клітини або організму із відмінною генетичною основою або є гомологічною по відношенню до клітини хазяїна, але розташована в іншому геномному середовищі, від того яке природно оточує таку молекулу нуклеїнової кислоти. Це означає, що якщо молекула нуклеїнової кислоти є гомологічною по відношенню до клітини хазяїна, вона не розташована в своєму природному місці в геномі вказаної клітини хазяїна, зокрема, вона є оточена іншими генами. В цьому випадку, молекула нуклеїнової кислоти може знаходитися або під керуванням свого власного промотору або під керуванням гетерологічного промотору. Вектор або молекула нуклеїнової кислоти, згідно даного винаходу, яка знаходиться в клітині хазяїні може бути або інтегрована в геном клітини хазяїна або знаходитися в якійсь позахромосомній формі. Відповідно до цього, також має бути зрозумілим, що молекула нуклеїнової кислоти даного винаходу може використовуватися для відновлення або створення мутантного гену шляхом гомологічної рекомбінації (Paszkowski (ed.), Homologous Recombination and Gene Silencing in Plants. Kluwer Academic Publishers (1994)). Відповідно, в іншій реалізації даний винахід стосується клітини хазяїна одержаної методом генетичної інженерії, за допомогою полінуклеотиду даного винаходу або вектору даного винаходу або вказаного вектору або вказаного полінуклеотиду та вектору або полінуклеотиду для експресії додаткового протеїну резистентності. Терміни "клітина хазяїн" та "рекомбінантна клітина хазяїн" тут є взаємозамінними. Є зрозумілим, що ці терміни вказують не лише на крему клітину але й на її потомство або потенціальне потомство такої клітини. Завдяки певним модифікаціям можуть з'являтися успішні покоління, завдяки або мутаціям або впливам зовнішнього середовища, такі потомства, фактично, можуть не бути ідентичними до батьківських клітин, але включатися в осяг та умови викладеного тут. Наприклад, полінуклеотид даного винаходу може бути введений в бактеріальні клітини, клітини комах, клітини грибів або клітини ссавців (такі як, клітини яєчників китайського ховрахи (СНО) або клітини COS), водоростей, інфузорій, рослинних клітин або грибів. Придатні клітини є добре відомими кваліфікованим фахівцям. Переважними є E.coli, baculovirus, Agrobacterium, або рослинні клітини. Також, клітина хазяїн може бути трансформована таким чином, що додаткові ферменти та протеїни, що підтримує експресію є (наg)експресованими, підвищують резистентність до рослинних патогенів. Переважно, додаткові гени резистентності також експресуються, переважно один або більше генів резистентності, переважно гени, згадані тут, та/або також експресовані. Найбільш переважною є сумісна експресія Rpi-blb2 та Rpi-blb. Наступними переважними є клітини однієї зі згаданих тут рослин, зокрема, одного з вище згаданих Solanaceae, найбільш переважними є картопля, помідор, петунія, томатне дерево, баклажан або динна груша. 26 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 В іншій реалізації, даний винахід стосується способу одержання поліпептиду даного винаходу, зокрема протеїну, що має активність Rpi-blb2, що включає культивування хазяйської клітини згідно даного винаходу та виділення поліпептиду, який кодується вказаним полінуклеотидом та експресується хазяйською клітиною, з культури або клітин. Термін "експресія" означає продукування протеїну або нуклеотидної послідовності клітини. Однак, згаданий термін також включає експресію протеїну в безклітинній системі. Він включає транскрипцію в РНК-продукт, посттранскрипційну модифікацію та/або трансляцію в протеїновий продукт або поліпептид із ДНК, що кодує цей продукт, також можливі посттрансляційні модифікації. Залежно від застосованих специфічних конструктів та умов, протеїн може бути виділений з клітин, культурального середовища або з обох. Для досвідченого спеціаліста, є добре відомим те, що можливо не лише експресувати нативний протеїн але й експресувати протеїн який утворений злиттям поліпептидів або додавати сигнальні послідовності, які спрямовують протеїн до певних компартментів клітини хазяїна, як такі, що забезпечують секрецію протеїну в культуральне середовище. Також, такий протеїн та його фрагменти можуть бути синтезовані хімічно та/або модифіковані згідно стандартних методів, описаних, наприклад нижче. Клітина хазяїн даного винаходу, така як прокаріотична або еукаріотична клітина в культурі, може бути використана для вироблення (наприклад, експресування) поліпептиду, кодованого полінуклеотиду винаходу, переважно полінуклеотиду, який має активність Rpi-blb2. Іншим методом, який може бути застосований додатково в рослинах є прямий трансфер ДНК в квітку, що розвивається або генетичний трансфер, опосередкований Agrobacterium. Відповідно, винахід також пропонує способи продукування Rpi-blb2 за допомогою клітин даного винаходу. В одній з реалізацій, спосіб включає культивування клітини хазяїна даного винаходу в придатному середовищі, й таким чином виробляння поліпептиду даного винаходу. Також, спосіб включає виділення та/або відновлення вказаного поліпептиду з культурального середовища або клітини хазяїна. Поліпептид даного винаходу переважно виробляється методами рекомбінантної ДНК. Наприклад, молекула нуклеїнової кислоти, що кодує протеїн клонується в експресійний вектор (як описано вище), експресійний вектор вводиться в клітину хазяїна (як описано вище) та вказаний поліпептид експресується в клітині хазяїні. Вказаний поліпептид може бути виділений з клітини за відповідною схемою очищення, з використанням стандартних методик очищування протеїнів. Також для рекомбінантного експресування, поліпептид або пептид даного винаходу можуть бути синтезовані з використанням стандартних методик синтезу пептидів. Більш того, нативні Rpi-blb2 можуть бути виділені з клітин (таких як, ендотеліальні клітини), наприклад, з використанням антитіл даного винаходу, як описано нижче, зокрема антитіл до Rpi-blb2, які можуть одержані стандартною методикою із застосуванням поліпептид даного винаходу або його фрагменту, наприклад, поліпептиду цього винаходу. В одній з реалізацій, даний винахід стосується протеїну Rpi-blb2 або протеїну, що має активність Rpi-blb2. В іншій реалізації, даний винахід стосується поліпептиду, що має амінокислоту послідовність, кодовану полінуклеотидом даного винаходу або одержуваний за способом даного винаходу. В одному з втілень, полінуклеотид не складається з послідовності, послідовності зображеної в Seq ID NO.: 8 та/або 10 та/або не складається з послідовності кодованої молекулою нуклеїнової кислоти, приведеної в Seq ID NO.: 7 та/або 9. В одному з втілень, поліпептид даного винаходу не складається з послідовності протеїну Мi1.1 або Мi1.2 та/або протеїну кодованого молекулою нуклеїнової кислоти, що кодує протеїн Мі1.1 або Mil.2. Таким чином, в одному з втілень, поліпептид даного винаходу, може не складатися із послідовностей приведених в Rossi et al. 1998, PNAS USA 95:9750-9754, Milligan et al., 1998. Plant Cell 10:1307-1319; та/або WO98/06750 Терміни "протеїн" а "поліпептид" використані в цій заявці є рівноцінними. "Поліпептид" вказує на полімер амінокислот (амінокислотну послідовність) та не вказує на певну довжину молекули Отже пептиди та олігопептиди включені в межі визначення поліпептиду Термін також вказує на або включає посттранскрипційну модифікацію поліпептиду, наприклад, глікозилування, ацетилування, фосфорилування а подібне. Включеними до визначення є, наприклад, поліпептиди, що містять один або більше аналогів амінокислот (включаючи, наприклад, неприродні амінокислоти, та інше), поліпептиди із заміщеними зв'язками, а також інші, відомі в практиці модифікації, як природного так і неприродного походження. 27 UA 102522 C2 5 10 15 20 25 30 35 40 45 50 55 Переважно поліпептиди є виділеними. "Виділений" або "чищений" протеїн або його біологічно активна частина є в значній мірі вільною від клітинного матеріалу при одержуванні за допомогою методів рекомбінації ДНК, або хімічних попередників при синтезуванні хімічно. Термін "в значній мірі вільний від клітинного матеріалу" включає одержування поліпептиду даного винаходу в якому протеїн є відокремленим від клітинних компонентів клітин, в котрих він продукувався природним або рекомбінантним шляхом. В одному з втілень, термін "в значній мірі вільний від клітинного матеріалу" включає одержування, яке має менш ніж приблизно 30 % (за сухою масою) "забруднюючого протеїну", більш переважно менш ніж приблизно 20 % "забруднюючого протеїну", ще більш переважно менш ніж приблизно 10 % "забруднюючого протеїну", та найбільш переважно менш ніж приблизно 5 % "забруднюючого протеїну". Термін "забруднюючий протеїн", вказує на поліпептиди, які не є поліпептидами даного винаходу. Коли рекомбінантно продукується поліпептид даного винаходу або його біологічно активна частина, також бажано щоб протеїновий препарат був "в значній мірі вільний від культурального середовища", тобто, культуральне середовище представляло менш ніж приблизно 20 %, ще більш переважно менш ніж приблизно 10 %, та найбільш переважно менш ніж приблизно 5 % об'єму. Термін "в значній мірі вільний від хімічних попередників або інших хімікатів" включає препарати, які є суб'єктом даного винаходу, наприклад, поліпептиди даного винаходу, які відокремлені від хімічних попередників або інших хімікатів, які залучені до синтезу протеїну. Термін "в значній мірі вільний від хімічних попередників або інших хімікатів" включає препарати, які мають менш ніж приблизно 30 % (за сухою масою) хімічних попередників або хімікатів не Rpi-blb2, більш переважно менш ніж приблизно 20 % хімічних попередників або хімікатів не Rpiblb2, ще більш переважно менш ніж приблизно 10 % хімічних попередників або хімікатів не Rpiblb2, та найбільш переважно менш ніж приблизно 5 % хімічних попередників або хімікатів не Rpi-blb2. В переважних реалізаціях, виділені протеїни або їхні біологічно активні частини, що не мають забруднюючих протеїнів того ж організму з котрого одержано поліпептид даного винаходу. Звичайно, ці протеїни виробляються методами рекомбінантної ДНК. Полінуклеотид даного винаходу, можуть брати участь в поліпептиді або його частину, яка включає амінокислотну послідовність, яка достатньо гомологічна до амінокислотної послідовності SEQ ID No: 2 або 4 так, що протеїн або його частина зберігає здатність брати участь в резистентності до патогенів даного винаходу. Частина протеїну, є переважно активною частиною як описано тут. Переважно, полінуклеотид даного винаходу, має амінокислотну послідовність приведену в SEQ ID No: 2 або 4. Також поліпептид може мати амінокислотну послідовність, яка кодується нуклеотидною послідовністю, яка гібридизується, переважно гібридизується за жорстких умов, як описано вище, із нуклеотидною послідовністю полінуклеотиду даного винаходу. Відповідно, поліпептид має амінокислотну послідовність, яка кодується нуклеотидною послідовністю, яка принаймні на 70 %, переважно принаймні на 75 %, більш переважно принаймні на 80 %, 90 %, або 95 %, та ще більш переважно принаймні на 96 %, 97 %, 98 %, 99 % або більше гомологічна до повної амінокислотної послідовності SEQ ID No: 2 або 4. Переважний поліпептид даного винаходу, переважно має принаймні одну активність протеїну Rpi-blb2, описану тут, наприклад, його резистентність або імунологічні активності. Переважний поліпептид даного винаходу включає амінокислотну послідовність, яка кодується нуклеотидною послідовністю, яка гібридизується, переважно гібридизується за жорстких умов, із нуклеотидною послідовністю полінуклеотиду SEQ ID No: 1 або 3, або 5, або 6 або таку, що гомологічні їм, як описано вище. Також поліпептид даного винаходу може відрізнятися від SEQ ID No: 2, або 4 за амінокислотною послідовністю, із-за природної мінливості або мутагенезу, як докладніше тут описано. Також, поліпептид має амінокислотну послідовність, яка принаймні на 70 %, переважно принаймні на 75 %, більш переважно принаймні на 80 %, 90 %, або 95 %, та ще більш переважно принаймні на 96 %, 97 %, 98 %, 99 % або більше гомологічний до повної амінокислотної послідовності SEQ ID No: 1 або 3, або 5, або 6. Біологічно активна частина поліпептиду даного винаходу має амінокислотну послідовність, яка має походження із амінокислотної послідовності протеїну Rpi-blb2, наприклад, амінокислотної послідовності в SEQ ID No: 2 або 4 або амінокислотної послідовності гомологічного до неї протеїну, який включає менше амінокислот, ніж протеїн повної довжини Rpi-blb2 або протеїн повної довжини гомологічний до протеїну Rpi-blb2 приведеного тут, та проявляє принаймні одну активність протеїну Rpi-blb2. Звичайно, біологічно (або імунологічна) активні частини, такі як, пептиди, які, наприклад, 5, 10, 15, 20, 30, 35, 36, 37, 38, 39, 40, 50, 100 або більше амінокислот в довжину мають домен або ділянку, із принаймні однією активністю або епітопом протеїну Rpi-blb2. Також, інші біологічно активні частини, в котрих інші видалені 28
ДивитисяДодаткова інформація
Назва патенту англійськоюFungus-resistant transgenic solanaceae plant
Автори англійськоюVan der Vossen Edvin Andries Gerard, Allefs Josephus Jacoubs Henricus Maria, Muskens Marina Woutera Maria
Назва патенту російськоюСтойкое к грибкам трансгенное растение solanaceae
Автори російськоюван дер Воссен Эдвин Андрис Герард, Аллефс Йозефус Якобс Хенрикус Мария, Мюскенс Марина Воутера Мария
МПК / Мітки
МПК: C12N 15/29, A01H 5/00, C12N 15/82
Мітки: трансгенна, рослина, solanaceae, стійка, грибків
Код посилання
<a href="https://ua.patents.su/186-102522-stijjka-do-gribkiv-transgenna-roslina-solanaceae.html" target="_blank" rel="follow" title="База патентів України">Стійка до грибків трансгенна рослина solanaceae</a>
Спосіб індукування стійкості до вірусу, що містить послідовність потрійного блока генів 3, за умови, що це не х вірус картоплі, у клітині рослини або рослині, трансгенна рослина, стійка до вірусу

Номер патенту: 70915
Опубліковано: 15.11.2004
Автори: Жонар Жерар, Блейкастен-Гроссханс Клодін, Гюйіє Убер, Лефебвр Марк, Бузубаа Салах, Річардс Кен, Вейєн Гю
МПК: C07K 14/08, C12N 15/82
Мітки: індукування, містить, спосіб, блока, картоплі, вірусу, клітині, рослини, стійкості, рослина, трансгенна, генів, умови, вірус, стійка, потрійного, послідовність
Формула / Реферат:
1. Спосіб індукування стійкості до вірусу, що містить послідовність потрійного блока генів 3, за умови, що це не X вірус картоплі, у клітині рослини або рослині, який включає стадії:- одержання конструкції нуклеїнової кислоти, яка містить послідовність нуклеїнової кислоти, що відповідає щонайменше на 70% послідовності нуклеїнової кислоти потрійного блока генів 3 зазначеного вірусу або його відповідної кДНК, функціонально злиту з однією...
Стійкі до грибків рослини родини solanaceae

Номер патенту: 90849
Опубліковано: 10.06.2010
Автори: Мюскенс Марина Воутера Марія, Аллефс Йозефус Якобс Хенрікус Марія, ван дер Воссен Едвін Андріс Герард
МПК: C12N 15/29, A01H 5/00, C12N 15/82, C07K 14/415
Мітки: стійкі, родини, грибків, рослини, solanaceae
Формула / Реферат:
1. Спосіб формування або підвищення стійкості рослин, що є представником родини Solanaceae, до рослинного патогену типу Oomycetes, що включає підвищення активності білка Rpi-blb2 у рослині або тканині, органі або клітині рослини або в їх частині, шляхом експресії молекули нуклеїнової кислоти, що кодує трансгенний білок Rpi-blb2, та/або підвищення числа копій молекули нуклеїнової кислоти, що кодує білок Rpi-blb2, в якому вказаний білок...
Трансгенна рослина кукурудзи mir604

Номер патенту: 94893
Опубліковано: 25.06.2011
Автори: Мегхіджі Мьоз, Штейнер Генрі-Йорк, Чен Ерік
МПК: C12Q 1/68, C12N 15/11, G01N 33/53, A01H 5/00
Мітки: трансгенна, рослина, кукурудзи, mir604
Формула / Реферат:
1. Виділена молекула нуклеїнової кислоти, що містить SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3 або SEQ ID NO: 4 або комплементи SEQ ID NO: 1-4.2. Амплікон, який містить молекулу нуклеїнової кислоти за п. 1.3. Рослина кукурудзи, яка містить молекулу нуклеїнової кислоти за п. 1.4. Рослина кукурудзи за п. 3, у якій після розщеплення геномної ДНК рослини рестриктазою KрnІ при гібридизації в строгих умовах з використанням...
Стрес-толерантна трансгенна рослина пшениці

Номер патенту: 90279
Опубліковано: 26.04.2010
Автори: Чемберлейн Дуглас Елан, Макнейл Скотт Девід, Боувер Роберт Сіндікомб
МПК: C12N 15/29, A01H 5/00, C12N 15/82
Мітки: стрес-толерантна, рослина, пшениці, трансгенна
Формула / Реферат:
1. Стрес-толерантна рослина пшениці, яка відрізняється тим, що дана рослина пшениці була трансформована молекулою нуклеїнової кислоти, що кодує орнітин-аміно-трансферазу (ОАТ).2. Стрес-толерантна рослина пшениці за п. 1, яка відрізняється тим, що молекулою нуклеїнової кислоти є молекула кДНК, яка має нуклеотидну послідовність, в основному, відповідну до тієї, що показана на фіг. 2 (SEQ ID N0:1), або її біологічно активний...
Модифікований фермент dmo та трансгенна рослина, що його продукує

Номер патенту: 94613
Опубліковано: 25.05.2011
Автори: Клементе Томас Е., Фен Пол К.К., Вікс Дональд П., Фласінскій Станіслав, Думітру Разван
МПК: A01H 5/00, C12N 15/82
Мітки: трансгенна, рослина, фермент, продукує, модифікований
Формула / Реферат:
1. Молекула нуклеїнової кислоти, вибрана з групи, що складається з:a) молекули нуклеїнової кислоти, що кодує поліпептид з SEQ ID NO:1;b) молекули нуклеїнової кислоти, що включає послідовність з SEQ ID NO:2; іc) молекули нуклеїнової кислоти, що кодує поліпептид, який має щонайменше 90 % ідентичності послідовності з поліпептидом з SEQ ID NO:1, де поліпептид має дикамба-монооксигеназну активність і включає цистеїн у...
Попередній патент: Спосіб одержання меламіну
Наступний патент: Застосування інгібіторів мієлопероксидази для лікування множинної системної атрофії
Випадковий патент: Спосіб визначення орієнтації осі візира та нашоломна система цілевказування для його реалізації