Плазміда pslc1-1/prd400, штам gv3101 agrobacterium tumefaciens, спосіб одержання трансгенної рослини з масловмісним насінням, спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини
Номер патенту: 49833
Опубліковано: 15.10.2002
Автори: Тейлор Девід С., Зоу Джітао, Катавік Весна, Маккензі Самюел Л., Келлер Вілфред А.







Формула / Реферат
1. Плазміда pSLC1-1/pRD400 (ATCC 97545).
2. Штам GV3101 Agrobacterium tumefaciens, який відрізняється тим, що вищезгаданий штам модифікований введенням дріжджового гена SLC1-1, що має нуклеотидну послідовність SEQ ID NO: 1.
3. Спосіб одержання трансгенної рослини з масловмісним насінням, який відрізняється тим, що в геном вищезгаданої рослини вводять дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну послідовність SEQ ID NO: 1, або дріжджовий ген SLC1, що здатен до експресії і який має нуклеотидну послідовність SEQ ID NO: 3.
4. Спосіб згідно з пунктом 3, в якому вищевказана рослина містить ендогенний ген, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, який відрізняється тим, що додатково здійснюють супресію експресії вищевказаного ендогенного гена, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, що вже присутня у геномі трансгенної рослини з масловмісним насінням.
5. Спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини, що полягає в тому, що вирощують масловмісну рослину, збирають насіння вищевказаної рослини і екстрагують олію з вищезгаданого масловмісного насіння, який відрізняються тим, що вищевказана рослина з масловмісним насінням є трансгенною рослиною з масловмісним насінням, у геном якої введений дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну послідовність SEQ ID NO: 1, або дріжджовий ген SLC1, що здатен до експресії і який має нуклеотидну послідовність SEQ ID NO: 3.
6. Спосіб згідно з пунктом 5, в якому вищевказана рослина містить ендогенний ген, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, який відрізняється тим, що вищевказаний ендогенний ген, що кодує ацилтрансферазу лізо-фосфатидилової кислоти є даун-регульованим.
Текст
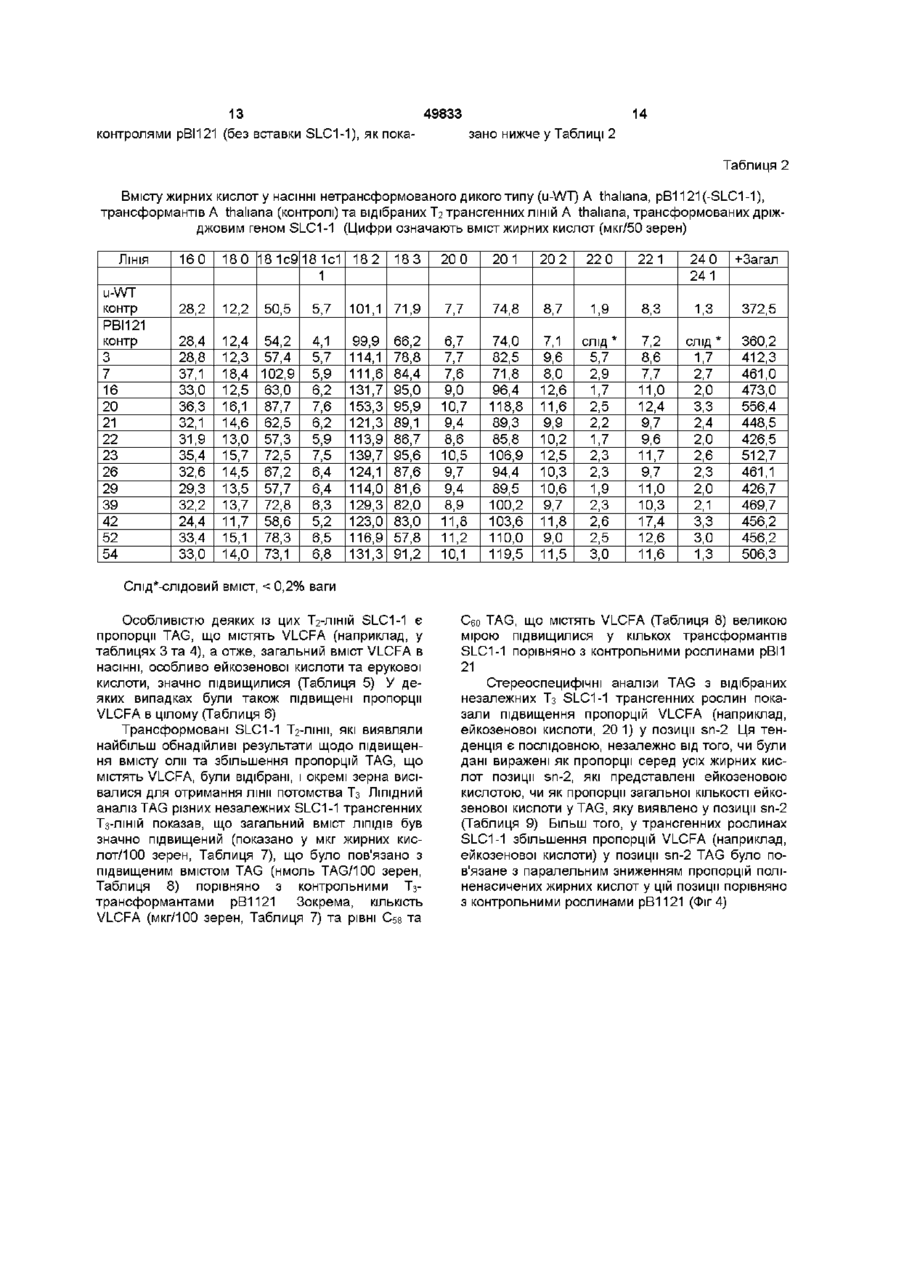
1 Плазміда pSLC1-1/pRD400 (ATCC 97545) 2 Штам GV3101 Agrobactenum tumefaciens, який відрізняється тим, що вищезгаданий штам модифікований введенням дріжджового гена SLC1-1, що має нуклеотидну ПОСЛІДОВНІСТЬ SEQ ID NO 1 3 Спосіб одержання трансгенної рослини з масловмісним насінням, який відрізняється тим, що в геном вищезгаданої рослини вводять дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну ПОСЛІДОВНІСТЬ SEQ ID NO 1, або дріжджовий ген SLC1, що здатен до експресії і який має нуклеотидну ПОСЛІДОВНІСТЬ SEQ ID NO З рослина містить ендогенний ген, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, який відрізняється тим, що додатково здійснюють супресію експресії вищевказаного ендогенного гена, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, що вже присутня у геномі трансгенної рослини з масловмісним насінням 5 Спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини, що полягає в тому, що вирощують масловмісну рослину, збирають насіння вищевказаної рослини і екстрагують олію з вищезгаданого масловмісного насіння, який відрізняються тим, що вищевказана рослина з масловмісним насінням є трансгенною рослиною з масловмісним насінням, у геном якої введений дріжджовий ген SLC1-1, що здатен до експресії і який має нуклеотидну ПОСЛІДОВНІСТЬ SEQ ID NO 1, або дріжджовий ген SLC1, що здатен до експресії і який має нуклеотидну ПОСЛІДОВНІСТЬ SEQ ID NO З 6 Спосіб згідно з пунктом 5, в якому вищевказана рослина містить ендогенний ген, що кодує ацилтрансферазу лізо-фосфатидилової кислоти, який відрізняється тим, що вищевказаний ендогенний ген, що кодує ацилтрансферазу лізофосфатидилової кислоти є даун-регульованим 4 Спосіб згідно з пунктом 3, в якому вищевказана О со го 00 Даний винахід стосується модифікації рослинних ЛІПІДІВ та насінних олій методами генної інженери Якщо конкретніше, винахід стосується способу генетичного модифікування рослин, насіння яких містить олії, для отримання олійного насіння або цілих рослин підвищеної комерційної ЦІННОСТІ Винахід також стосується модифікованих рослин та насіння, генетичних матеріалів та векторів, які використовуються для отримання таких рослин, та подальшої модифікації рослин Останнім часом існує значна зацікавленість у модифікуванні композиції жирних кислот насінних олій та вмісту насіння, що містить олії, молекулярно-генетичними способами для забезпечення надійного ресурсу рапсового насіння з надвисо ким вмістом ерукової кислоти (SHEAR) для використання як промислового харчового запасу Подібна зацікавленість існує і у виробництві інших стратегічних нехарчових олій (наприклад, насінних олій, багатих на гідрокси-, епокси-, короткота середньоланцюгові жирні кислоти і т п) з насіння традиційних олійних культур (рапсу, льону, соняшника, сої) Що стосується харчових олій, то існує значна зацікавленість у ЗМІНІ КОМПОЗИЦІЇ жирних кислот (наприклад, вищі олеїнові/нижчі поліненасичені, нижчі насичені вищі насичені), а також у підвищенні вмісту в насінні олійних культур таких олій, як канола та харчова льняна олія (лінола), соєва та соняшникова олія 49833 На даний час не існує задокументованих прикладів збільшення вмісту олії за допомогою трансгенних засобів, хоча и продукування підвищують шляхом традиційного схрещування та селекції, що дозволяє досягти поліпшення Збільшення ж пропорцій деяких стратегічних жирних кислот, навпаки, було досягнуте через впровадження біосинтезу різних рослинних жирних кислот та генів ацилтрансфераз у насіння, що містить олію Серед прикладів подібних процесів можна назвати такі 1 Експресія серед ньоланцюгової жирної ацил-АСР-тюестерази з Каліфорнія-Бей у Brassiсасеае для збільшення вмісту лаурилової кислоти (12 0) (Calgene, Voelker et al , 1995, 1996 - див Посилання 35 та 36 у додатку "Посилання, що стосуються даного винаходу") 2 Експресія (і-кетоацил-СоА-синтази хохоби у культиварах Brassica napus (Канола) з низьким вмістом ерукової кислоти для підвищення рівня ерукової кислоти, ефект, що спостерігався після експресії у культиварах з високим вмістом ерукової кислоти, був незначним (Calgene, Lassner et al , 1996 - див Посилання 20) 3 Експресія анти-смислової ПОСЛІДОВНОСТІ ДО стеароіл-АСР А9-дезатурази у Brassicaceae для підвищення вмісту стеаринової кислоти (Calgene, Knutzon et al , 1992 - див Посилання 16) 4 Збільшення пропорцій олеїнової кислоти у В napus шляхом косупресм з використанням смислової ПОСЛІДОВНОСТІ, що кодує рослинну мікросомну FAD2 (Д12)-денатуразу (duPont/lnterMountam Canola, Hitz et al , 1995 див Посилання 12) 5 Збільшення пропорцій до 12 0 або 20 1 в 5л-2-позиціі триацилгліцеролів (ТАГ) у насінні рапсу шляхом експресії кокосових або тіарелових ацилтрансфераз лізофосфатидної кислоти ( L P A T s , E C 2 3 1 5 1 ) , В І Д П О В І Д Н О ( C a l g e n e , Knutzon et al , 1995 a & b, - див Посилання 17 та 18, Lassner et al , 1995 - див Посилання 21) Незважаючи на використання рослинних трансгенів, в результаті яких змінювалися пропорції sn-2-лауриновоі та ерукової кислот, у лавровій канолі та насінні рапсу, багатому на ерукову кислоту, ВІДПОВІДНО, загальні пропорції лауринових та ерукових кислот у насінній олії не підвищились і не спостерігалося підвищення загального вмісту жирної кислоти або підвищення продукування олії у цих трансгенах Таким чином, постала потреба у нових шляхах підвищення продукування й поліпшення складу олії в насінні олійних рослин через застосування методів генної інженери Метою даного винаходу є генетична модифікація олійних рослин для поліпшення комерційних якостей таких рослин, їхнього насіння та олій, які продукують такі рослини Ще однією метою винаходу є забезпечення способу модифікації складу та збільшення виходу олій, що продукуються олійними культурами Даний винахід спирається на відкриття того, що, sn-2-генІл ацилгліцеринової жирної ацилтрансферази (SLC1-1 та и алель SLC1) із дріжджів (Saccharomyces cereviseae) можуть бути використані для зміни вмісту олії та и складу у насінні рослин та ліпідах листя Таким чином, згідно з одним аспектом даного винаходу, забезпечується наявність трансгенної олійної рослини, яка має геном, що містить експресибельний дріжджовий SLC1-1 або SLCI-ген Згідно З другим аспектом винаходу, забезпечується наявність насіння трансгенної олійної рослини, яке має геном, що містить експресибельний дріжджовий SLC1-1 або SLCI-ген Згідно З іще одним аспектом винаходу, забезпечується метод отримання трансгенної олійної рослини, який охоплює введення до геному цієї рослини експресибельного дріжджового SLC1-1 або SLC1-гену Винахід також стосується різноманітних плазмід та векторів, застосовуваних у методиці даного винаходу, та паралельного введення інших генів у модифіковані рослини для включення генів SLC1-1 a6oSLC1 Переваги даного винаходу полягають і в тому, що дріжджові гени SLC1-1 або SLC1 можуть використовуватися для збільшення вмісту олії та для зміни загальної композиції жирних кислот, а також для змінення ацильної композиції TAG, включаючи позицію sn-2, та для змінення пропорцій видів TAG у різних олійних рослинах, наприклад, Arabidopsis thahana, у культиварах каноли Brassica napus та Brassica carmata з високим вмістом ерукової кислоти Більш ТОГО, дріжджова sn-2ацилтрансфераза (гени SLC1-1 та SLC1) може бути використана у Brassicaceae з високим вмістом ерукової кислоти для підвищення вмісту олії та для продукування насінних олій з підвищеним вмістом жирних кислот з дуже довгими ланцюгами (VLCFA) та TAG зі зміненою стереоспецифічною композицією по відношенню до жирних кислот з дуже довгими ланцюгами Таким чином, на відміну від попередніх результатів, використання рослинних трансгенів (як було зазначено вище), даний винахід з використанням дріжджового трансгену дозволяє досягти одночасного підвищення вмісту насінної олії, вмісту насінної ерукової кислоти та загальних пропорцій ерукової кислоти у насінній олії Дріжджова sn-2 ацилтрансфераза (гени SLC1-1 та SLC1) також може бути використана в культиварах харчових олій (культивари якості Каноли) Brassicaceae для збільшення вмісту олії та для продукування насінних олій зі зміненими пропорціями олеїнової кислоти, поліненасичених жирних кислот та насичених жирних кислот з дуже довгими ланцюгами ВІДПОВІДНІ SLC1-1 та SLC1 дріжджові алелі можуть бути використані таким же чином Обидві алелі кодують зя-2-ацилтрансферазу, SLC1 відрізняється від SLC1-1 лише в амінокислотній позиції 44 (Глутамш, Q) порівняно з SLC1-1, де в амінокислотній позиції 44 знаходиться Лейцин (L) SLC1-1 та SLC1 трансгенні рослини можуть бути використані як гермплазм-хазяї для подальшої знижуючої регуляції власних ацилтрансфераз рослин Для досягнення потрібної впорядкованої структури TAG-бюсинтезу для продукування TAG за стереоспецифічною схемою, необхідна коор 49833 динована експресія багатьох біохімічних реакцій, включаючи ті, в яких посередником виступає LPAT Одним з конкретних можливих варіантів по відношенню до оптимізацм трансгенної експресії чужорідних LPAT для синтезу TAG з новими ациловими композиціями (наприклад, збільшення дуже довгих ланцюгів жирних кислот у позиції sn2), є можливість виникнення необхідності в одночасній знижуючій регуляції власних LPAT, які вже присутні у трансгенній рослині-хазяїн) (наприклад, LPAT, яка зазвичай ставить поліненасичені Сіє жирні ацильні групи в sn-2-позицію) Загальні гомології між дріжджовою sn-2 ацилтрансферазою та відомими рослинними sn-2ацилтрансферазами (LPAT) є незначними і здебільшого обмежуються С-кінцями білків На відміну від них, рослинні ацилтрансферази мають між собою значно вищу гомологію, і ГОМОЛОГІЧНІ ДІЛЯНКИ поширюються на всю ПОСЛІДОВНІСТЬ Тому використання дріжджових SLC-генів для досягнення описаних нами ефектів надає унікальну можливість подальшого поліпшення цих ознак способом, який був би неможливим у разі здійснення з рослинною ацилтрансферазою первісної трансформації Як наслідок, обмежена гомологія між рослинною й дріжджовою sn-2ацилтрансферазами є досить низькою для того, щоб дозволити здійснювати знижуючу регуляцію LPAT рослини-хазяїна традиційними засобами (наприклад, метод анти-смислової РНК або феномен косупресм, (Мої e t a / , 1990, Van Blokland et al , 1993, De Lange et a/, 1995) без супутніх негативних впливів на експресію дріжджового трансгену , або на розвиток насіння рослини Таким чином, використання дріжджового трансгену має конкретну перевагу над введенням до рослинихазяїна трансгену іншої рослини, що має власну високогомолопчну LPAT Дріжджова sn-2-ацилтрансфераза (гени SLC1-1 та SLC1) може бути використана для збільшення вмісту олії й змінення ацильної композиції TAG в усіх інших зразках насіння, що містить олію, включаючи бурачник (види Вогадо), рицина (Ricmus commums), какао (Theobroma cacao), кукурудзу (Zea mays), бавовну (види Gossypium), різні види Crambe, Cuphea, льон (види Linum), різні види Lesquerella та Limnanthes, настурцію (види Tropaeolum), різні види Oenothera, оливи (види ОІеа), олійну пальму (види Elaeis), арахіс (види Arachis), сафлор (види Carthamus), сою (види Glycme та Soya), соняшник (види Hehanthus), тютюн (види Nicotiana) та різні види Vernoma Для продукування насінної олії з підвищеним вмістом жирних кислот поліпшеної якості (тобто, з підвищеним вмістом пдрокси-жирних кислот і ЗМІНЄННЯМ КОМПОЗИЦІЙ TAG ВІДНОСНО ТИХ, ЩО МІСТЯТЬ гідрокси- жирні кислоти) можуть використовуватися трансформанти насіння, що містить олію, з дріжджовою sn-2-ацилтрансферазою (SLC1-1 та SLCL-гени), через їхню повторну трансформацію, з бюсинтетичними генами усіх інших жирних кислот поліпшеної якості (наприклад, геном пдроксилази рицини або різних видів Lesquerella), або через схрещування зі спорідненими трансформантами олійного насіння, що вже містять такі гени поліпшеної якості Ген SLC1-1 та відповідна алель SLC1 можуть бути використані для модифікації жирних кислот та ЛІПІДНИХ профілів у вегетативних тканинах для поліпшення СТІЙКОСТІ до біотичних та абіотичних стресів рослин (наприклад, підвищена плинність мембран у тканинах коріння та листя для поліпшення морозостійкості) Використання дріжджового гену SLC1-1 та алелі SLC1 у рослинах для викликання змін у загальному ВМІСТІ та композиції ЛІПІДІВ раніше не було ні описано, ані продемонстровано (доведено до практики) як засіб маніпулювання відносними пропорціями або кількостями жирних кислот (наприклад, жирних кислот з дуже довгими ланцюгами), а також для підвищення вмісту олії у культурах, що продукують харчові та промислові олії Раніше не було продемонстровано збільшення продуктивності олії, викликаного трансгенними засобами Якщо точніше то до цього час у не існувало свідчення того, що дріжджові ацилтрансферази - ферменти, відповідальні за синтезування триацилгліцеролів, - були експресовані у рослинах для змінення композиції або вмісту олії Однак, зниження активності діацилгліцеролацилтрансферази у мутанта Arabidopsis thahana призводило до зменшення виходу продуктивності олії та змін ацильної композиції (Katavic et a/, (1995) Plant Physiology, 108 399-409 - див Посилання 15) На Фіг 1 показано нуклеотидну [SEQ ID № 1] та комплементарну до неї амінокислотну [SEQ ID № 2] ПОСЛІДОВНОСТІ кодуючої ділянки дріжджового гену SLC1-1, використаного в даному винаході, стоп-кодон, позначений як "@", і підкреслено висококонсервативну консенсусну ПОСЛІДОВНІСТЬ між бактеріальною та дріжджовою sn-2ацилтрансферазами, На Фіг 2 показано нуклеотидну [SEQ ID № 3] та комплементарну до неї амінокислотну [SEQ ID № 4] ПОСЛІДОВНОСТІ кодуючої ділянки дріжджового гену SLC1, використаного в даному винаході, стоп-кодон, позначений як "@", і підкреслено висококонсервативну консенсусну ПОСЛІДОВНІСТЬ між бактеріальною та дріжджовою sn-2ацилтрансферазами, На Фіг 3 показано напрямки побудови SLC1-1 вектору трансформації рослин, які пояснюються нижче у Деталях Експерименту, рельєфні частини показано не у масштабі, та На Фіг З 4 ПО 7, ЯК І на Таблицях 1 - 20 нижче показано результати тестів, які пояснюються нижче у Деталях Експерименту ПОСЛІДОВНОСТІ гену SLC1-1 [SEQ ID № 1] та алелі SLC1 [SEQ ID № 3] і ПОХІДНІ ВІД НИХ пептичні структури [SEQ ID №№ 2 та 4] є такими, як показано на Фіг 1 і 2, ВІДПОВІДНО Дріжджової ген SLC1 (та пов'язаний з ним суп ресорний алельний ген SLC1-1) були охарактеризовані у двох нижчевказаних публікаціях (опис яких включено нами як посилання) 1 Lester, R L, Wells, G В , Oxford, G and Dickson Я С (1993) Mutant strains of Saccharomyces cerevisiae lacking sphingohpids synthesize novel mositol glycerohpids that mimic sphmgohpid 49833 structures J Biol Chem 268 845-856 - Посилання 22, та 2 Nagiec, M M , Wells, G В , Lester, R L, and Dickson R C , (1993) A suppressor gene that enables Saccharomyces cerevisiae to grow without making sphmgohpids encodes a protein that resembles an Eschenchia coh fatty acyltransferase J Biol Chem 268 22156-22163-Посилання 25 АМІНОКИСЛОТНІ та ДНК-ПОСЛІДОВНОСТІ кодуючої ділянки гену SLC1-1 депонуються у GenBank/EMBL під номером доступу No L13282 (депонована ПОСЛІДОВНІСТЬ, що включає 5'нетрансльовану ділянку, у даній заявці не описується) Ген SLC1 первісно був тонований з дріжджового мутанту, що втратив здатність продукувати сфінголіпіди Було показано, що мутантна алель SLC1 кодує білок, який пригнічує генетичний дефект біосинтезу основи довгого ланцюга сфінголіпіду ПОСЛІДОВНІСТЬ гену SLC1 є гомологічною до гену Е coh PLSC, який, як було заявлено, кодує ацилтрансферазу лізо-фосфатидної кислоти (LPAT, ацилтрансфераза, яка ацилює sn-2позицію лізо-фосфатидної кислоти (LPA) для утворення фосфатидної кислоти (PA) (Coleman, 1990, Coleman 1992) Ген SLC1 був спроможний комплементувати дефект росту у JC201 (штам Е coh, мутований у PLSC) (Coleman, 1990) На підставі спостереження, що штами SLC, які зростають за відсутності довголанцюгової основи, продукують нові ПОХІДНІ фосфатид илінозитолу (Lester e t a l , (1993) J Biol Chem 268 845-856), єдиним можливим висновком авторів було те, що SLC1 кодує білок, спроможний ацилювати sn-2позицію гліцероліпідів, що містять інозитол (можливо, ацилтрансферазу л ізо-фосфатид илінозитолу, LPIT) На підставі цих спостережень було заявлено, що SLC1 кодує дріжджову sn-2 ацилтрансферазу Щоправда, автори статті (Dickson, Lester et al) були не в змозі визначити активність LPAT у комплементуючому мутанті Е coh JC201 У праці Nagiec зі співавторами повідомляється також повідомляється про ПОСЛІДОВНІСТЬ гену для супресорної алелі, що вказує на SLC1-1, у якій нуклеотид 131 має Т замість А, внаслідок чого в амінокислотній позиції 44 глутамш змінюється на лейцин Робочою гіпотезою є та, згідно з якою супресорна алель SLC1-1 кодує варіант ацилтрансферази зі зміненою субстратною специфічністю, яка дозволяє їй використовувати жирну кислоту з дуже довгим ланцюгом (26 0) для ацилювання sn-2-позицм гліцероліпідів, що містять інозитол На даний момент автори не забезпечили переконливого доказу активності, яка кодується SLC1-1 або SLC1 На підставі інтересу авторів даного винаходу до модифікування вмісту жирної кислоти з дуже довгим ланцюгом (VLCFA) у Brassicaceae, винахідники отримали плазміду р 411 Д В/С, що містить SLC1-1 ген-супресор алелі від доктора Діксона з Університету Кентукі, Лексингтон, Кентукі, США Винахідники також вважають, що експресія чужорідного гену у рослинах може дати більше інформації про природу того, що кодують SLC1-1 та SLC1 Спочатку робота, проведена винахідни 8 ками, ідентифікувала використання моделі олійного насіння Arabidopsis thahana, трансформантів з підвищеним вмістом олії у насінні та підвищеними пропорціями TAG, що містить жирні кислоти з дуже довгими ланцюгами (VLCFA = > С-is) Крім того, існує підвищення пропорцій VLCFA у sn-2 позиції TAG, і, разом з цим, зниження пропорцій поліненасичених жирних кислот, естерифікованих у цій позиції SLCI-1-трансформанти В napus cv Него та В carmata (обидва є культиварами, багатими на ерукову кислоту) виявляють підвищений вміст олії й підвищений вміст ерукової кислоти на мг сухої ваги (DW) насіння SLC1-1 трансформанти В napus cv Westar (культивар якості Каноли) виявляють підвищення пропорцій олеїнової кислоти (18 1) і зниження пропорцій поліненасичених жирних кислот (18 2 та 18 3) Гени SLC1-1 та SLC1 можуть бути введені у геном рослин з насінням, що містить олію, й експресовані з застосуванням традиційних методів генної інженери Наприклад, трансформація може включати застосування трансформації Agrobacterium Ті за допомогою плазміди (наприклад, in planta, вакуумною інфільтрацією, інфікуванням прорізів СІМ'ЯДОЛІ або черешка ппокотилю, або бомбардуванням частинками і т п ) ПОСЛІДОВНОСТІ можуть керуватися конститутивними або тканинно-специфічними промоторами - це безпомилково можуть визначити спеціалісти Є підстави розраховувати на широке застосування винаходу щодо різних олійних рослин, оскільки синтез олії здійснюється однаково чи майже однаково в усіх подібних рослинах (див Посилання 29, ЗО, 37, 38, 39 та 40) Даний винахід буде описано більш детально з посиланням на подані нижче деталі експерименту, де подаються спеціальні ілюстрації Однак, слід мати на увазі, що даний винахід не обмежується цими деталями ДЕТАЛІ ЕКСПЕРИМЕНТУ ПОБУДОВА ВЕКТОРІВ ДЛЯ ТРАНСФОРМАЦІЇ CLS-1 Згідно З напрямком клонування, який проілюстровано на Фіг 3, у Полімеразній Ланцюговій Реакції (PCR) з плазмідою р411ДВ/С (отриманою від д-ра Діксона з Університету Кентукі у Лексингтоні, Кентукі, США), яка включає супресорну алель SLC-гену (SLC-1) як зразок, для вироблення SLC1 PRC-фрагменту з BamHI-сайтом по обох кінцях були використані два праймери з 5' ВатНІсайтами рестрикції на ділянках ОМ087 (AGAGAGAGGGATCCATGAGTGTGATAGGTAGG) [SEQ Ю № 5] та ОМ088 (GAGGAAGAAGGATCCGGGTCTATATACTACTCT) [SEQ ID № 6], складених згідно з 5' та 3' послідовностями ВІДПОВІДНО Таким чином, фрагмент (SLC1-1) PSR являє собою супресорну алель гену SLC1 з нуклеотидом Т, який заміщує нуклеотид А у позиції 131, що призводить до того, що амінокислотний залишок змінюється з глутамшу на лейцин у позиції залишку 44 Фрагмент вирізався за допомогою ВатНІ і вставлявся у сайт для клонування ВатНІ, що розміщувався між тандемним промотором 35S та термінатором NOS у векторі рВ1524 (отримано від д-ра Радж С С Датла, з NRC Ін 49833 ституту біотехнологм рослин, 110 Gymnasium Place, Saskatoon, Saskatchewan, Canada, S7N OW9, опубліковано Datla et al , 1993 - див Посилання 9) для отримання вектору SLC1-1-pBI-524 Орієнтацію SLC1-1 у векторі SLC1-1-pBI-524 було підтверджено рестрикцією за допомогою ферменту Bg1ll, який відрізає SLC1-1 у нуклеотиді 377 від кінця 5' й безпосередньо справа від промотора 35S у векторі рВ1524 Підтримується кодон ініціації трансляції SLC1-1 і, таким чином, ПОСЛІДОВНІСТЬ є транскрипційно злитою Фрагмент Hind IN-EcoRI, що містить тандемний промотор 35S, енхансер AMV, кодуючу послідовнсть SLC1-1 та термінатор NOS, було вивільнено з SLC1 -1-рВІ524 й т о н о в а н о у сайт EcoRI-Hmd III вектору RD400 (також отриманого від д-ра Датла, опубліковано Datla et al , 1992 - див Посилання 8) Остаточний вектор pSLC1-1/pRD400 (депонований 9 травня 1996 року згідно з будапештською Угодою в Американське Зібрання Культур Мікроорганізмів (American Type Culture Collection), 12301 Parklawn Drive, Rockville, MD 20852, USA, під депозитарним номером АТСС 97545) вводили у штам GV 3101 Agrobactenum tumefaciens (що несе хелпер-плазміду рМР90, Koncz and Schell, 1986) шляхом електропорацм МОЛЕКУЛЯРНО-БЮЛОПЧНІ МЕТОДИ Усі молекулярно-біологічні методи застосовувалися згідно з методиками, вказаними в Ausubel et al , (1995), якщо немає спеціального зауваження УМОВИ ВИРОЩУВАННЯ РОСЛИН Усі контрольні та трансгенні рослини А thahana вирощували одночасно у камерах з контролем росту при постійному флуоресцентному освітленні (150 - 200 мкЕ м сек ) при 22°С, як описано у Katavic et al , (1995) Усі ІНШІ контрольні та - трансгенні рослини Brassicaceae (В napus, В carmata) вирощували одночасно у теплиці Центру Трансгенних Рослин Р В І, при натуральному освітленні, доповненому лампами з натрієм під високим тиском (HPS-лампи), з фотоперіодом 16 годин (16 год світла/8 год темряви) при 22°С та ВІДНОСНІЙ вологості 25 - 30% ТРАНСФОРМАЦІЯ РОСЛИН ПОСЛІДОВНІСТЬ 5-LC1-1/RD400 аналізували у A thahana методом трансформації in planta, і у культиварах з високим та низьким вмістом еруковоі кислоти В napus, та В carmata (шляхом кокультиваційної трансформації черешків СІМ'ЯДОЛІ та експлантатів ппокотилю з A-tumefaciens, що несе ПОСЛІДОВНІСТЬ SLC1 -1) Аналіз ПОСЛІДОВНОСТІ SLC1-1 у A thahana Рослина дикого типу (WT) A thahana екотипу Columbia вирощувалася у грунті Трансформацію in planta здійснювали шляхом інокуляції у надрізі (Katavic et al, 1994) або вакуумної інфільтрації (Bechtold et al, 1993,) з суспензією штаму GV3101 нічної бактеріальної культури A tumefaciens, який несе нопалінову плазміду-хелпер рМР90 (неактивна Ті-плазміда з інтактною vir-ділянкою, що діє in trans, маркерами гентаміцинної та канаміцинної селекції, (Koncz and Scheil (1986)) та бінарним вектором pSLC1-1/pRD400 Після інокуляції або інфільтрації рослини вирощували до стадії закладки насіння (Ті) Сухе 10 насіння (Ті) збирали у великій КІЛЬКОСТІ Й робили скринінг на селективному середовищі з 50мг/л канаміцину Після перебування протягом двохтрьох тижнів на селективному середовищі проростки переносили у грунт ДНК листя виділяли з стійких до канаміцину Ті-рослин і аналізували шляхом ПЦР-ампліфікацм SLC1-1 фрагменту Молоде листя з Ті рослин, а також дозріле насіння І2 з трансгенних ЛІНІЙ SLC1-1 використовували для ЛІПІДНИХ та біохімічних аналізів Молоде листя та дозріле насіння нетрансформованих рослин дикого типу (WT) екотипу Columbia та рВ1121 трансгенних рослин (бінарний вектор рВ1121, що містить лише селекційний маркер канаміцину та ВІДПОВІДНИЙ ген GUS, Jefferson et al, 1987) використовувалися як контролі під час аналізу ЛІПІДІВ насіння На підставі цих аналізів насіння І2 з ЛІНІЙ, що виявляють зміни у композиції ацилів та/або ВМІСТІ ЛІПІДІВ, вирощували на селекційному середовищі (для видалення гомозиготних сегрегантів WT), а потім переносили у грунт для отримання популяції насіння Тз Аналіз SLC1 -1 послідовностей у Brassica napus та Brassica carmata Експерименти з трансформації також були здійснені на В napus cv Westar (сорти каноли, бідні на ерукову кислоту), В napus , види Hero, Reston та Argentine (усі сорти є багатими на ерукову кислоту) та В carmata (генеалогічна ЛІНІЯ С90-1163, ЛІНІЯ з високим вмістом ерукової кислоти) шляхом кокультивування черешків сім'ядолей та експлантатів ппокотилю з A tumefaciens, що несе ПОСЛІДОВНІСТЬ SLC1-1/RD400 Методи трансформації, за Moloney et al (1989) та DeBlock et al (1989), були змінені для оптимізацм умов трансформації Зміни методу трансформації черешоксім'ядоля (Moloney et al , 1989) включали впровадження 7-денного періоду відновлення експлантату з подальшою кокультивацією на середовищі MS з гормоном бензиладеніном (ВА) та антибіотиком тиментином для видалення Agrobactenum Зміни методу трансформації ппокотильексплантат (DeBlock et al , 1989) включали (1) попередню культивацію експлантату на середовищі MS, загущеному агаром, з гормонами - 2,4дихлорофеноксиоцтовою кислотою (2,4-D) та кінетином (К), (2) кокультивацію експлантатів ппокотилю з Agrobactenum у чашках Петрі з тим самим середовищем, що й при попередній культивації, на стерильних паперових фільтрах, (3) подальша кокультивація, 7-денний період відновлення експлантату на середовищі з гормонами (2,4-D та К) та з тиментином для видалення Agrobactenum, (4) регенерація трансгенних паростків на середовищі MS з гормонами - бензиладеніном (ВА) та зеатином (Z), інгібітором етилену нітратом срібла (АдІЧОз) та антибіотиками тиментином ( Д Л Я видалення Agrobactenum) та канаміцином (для селекції трансформованої клітини/паростка) Зелені паростки переносили у грунт і укорінювали Геномну ДНК виділяли з молодого листя й здійснювали PCR-аналіз та аналіз Саузерна (Southern, 1975) Насіння (Т-і) трансгенних рослин збирали й вирощували у грунті по десять Ті рос 11 лин з кожної трансгенної лінії Достигле насіння (Тг) з цих рослин збирали й піддавали ліпідному та біохімічному аналізам ЛІПІДНІ АНАЛІЗИ ТА ПРОБИ АЦИЛТРАНСФЕРАЗИ (LPAT) Аналізи ЛІПІДІВ листя та насіння з SLC1-1 та \ЛЛ7рВІ121-трансгенних та нетрансформованих рослин дикого типу (WT) Л І П І Д И ВИДІЛЯЛИ З ДОСТИГЛОГО насіння та мо лодого листя, як описано вище (Taylor et al , 1992, Katavic et al , 1995) і піддавали аналізові з застосуванням GC на загальний вміст жирних кислот та композицію жирних кислот Триацилгліцеринові зразки аналізували з застосуванням високотемпературних GC, як описано у Katavic et al , 1995 Стереоспецифічні аналізи TAG виконували на ліпідах інтактного насіння (головним чином TAG), як описано у Taylor et al , 1994, 1993 а &b Проби LPAT Для проб листя з контрольних та SLC1-1трансгенних рослин відбирали листя середнього розміру і забираються зразки тканини кількох листів Для проб насіння збирали 25 - ЗО стручків A thahana на середній стадії розвитку насіння (15 - 18 днів), ДЛЯ отримання зразків насіння Тз, що розвивається, з обох контролів (нетрансформований W T та трансформований рВИ 21), і відбирали трансгеніки SLC1-1 Тг-ембрюни В napus та В carmata на середній стадії розвитку СІМ'ЯДОЛІ збирали з трьох стручків контрольних та відібраних SLCI-1-трансгенних рослин Усі рослинні матеріали одразу ж заморожували у рідкому азоті й зберігали при-70°С до гомогенізації Готували гомогенати з листя рослин та тканин насіння, що розвивається, і проводили проби LPAT, як описано у Taylor et al , (1995 b) Усі процедури з дріжджовими штамами здійснювалися, як описано у Ausubel et al , (1995, Unit 13 1 Basic Techniques of Yeast Genetics) Штами дикого типу S cerevisiae та S pombe культивували у середовищі YPD при 28°С при 270 обертах на хвилину протягом ночі У середній лог-фазі забирали зразки клітин, осаджували їх шляхом центрифугування при 5000 об/хв протягом 5 хвилин і ресуспендували в ЮОмМ Hepes-NaOH, pH 7,4 Лізати клітин готували, використовуючи промиті кислотою скляні кульки, як описано у Ausubel et al , 1995 (Unit 13 1, Section 13 13 4) Проби LPAT проводили при рН 7,4, при збовтуванні на швидкості 100об/хв у водяній бані з температурою 30°С протягом 1 0 - 3 0 хвилин Суміші проб (остаточний об'єм 0,5 мл) містили білок (10-200 мкг, залежно від тканини/екстракту), 90мМ Hepes-NaOH, 0,5мМ АТР, 0,5мМ CoASH, 2мМ спермідину, 45мкМ 18 1 - LPA та на вибір 18мкМ [1- 1 4 С]-18 1-СоА, [1- 1 4 С]-20 1-СоА або [ 1 14 С]-22 1-СоА (кожної зі специфічною активністю 10 пСі/нмоль) як ацильний донор Усі ІНШІ умови вимірювання активності LPAT докладно описані у Taylor et al, (1995b) 1 Н-ЯМР достиглого насіння Аналізи 1 Н-ЯМР для відносного продукування олії (Alexander et al , 1967, Rutar, 1989) проводили на інтактному насінні контролів та трансформованих SLC1-1 В napus cv Hero та В carmata, 49833 12 використовуючи потужний спектрометр Bruker AM, що працює при ЗбОмГц Для зниження анізотропного ЛІНІЙНОГО розсіювання насіння (35/зразок) обертали при 1 кГц у цирконієвому роторі, орієнованому під кутом 54,7° до магнітного поля (обертання зразків під магічним кутом, MASS) РЕЗУЛЬТАТИ Ацил-СоА-специфічність дріжджової (S cereviseae, S pombe) sn-2 ацилтрансферази (LPAT) Лізати дріжджових клітин S cereviseae та S pombe брали на проби відносної активності sn-2ацилтрансферази, з використанням 18 1 LPA як ацилового акцептора та різних радіоактивне мічених ацил-СоА Специфічність ацил-СоА дріжджових LPAT in vitro була досить широкою, і LPAT була спроможна вносити як власну (16 0, 18 1), так і чужорідну (18 2, 18 3, 20 1, 22 1 та рицинолеоїл) ацильні групи в sn-2-позицію 1 8 1 LPA, як показано нижче у Таблиці 1 Таблиця 1 ВІДНОСНІ ацил-СоА S cerevisiae та S pombe активність LPAT з використання м 45мкМ 18 1-LPA як ацилового акцептора Активність Активність LPAT Запас 14С-Ацил-СоА _РАТнмоль/хв/мг відносно 18 1(18мкМ ) білок СоА(%) S cerevisiae 18 1-СоА 3 75 100 18 2-СоА 3 54 94 5 18 1 Д12-ОН-СоА 1 90 50 7 20 1-СоА 1 92 51 3 22 1-СоА 0 33 89 S pombe 18 1-СоА 18 2-СоА 18 1 АІ2-ОН-СоА 20 1-СоА 22 1-СоА 1 50 1 27 0 85 0 38 0 60 100 84 7 56 7 25 3 40 0 Через те, що LPAT (згг-2-ацилтрансфераза) має відносно широку специфічність, трансформація олійного насіння, багатого на жирні кислоти з дуже довгими ланцюгами (A thahana, В napus), дріжджовим геном SLC1-1 може бути заздалегідь розрахована так, щоб збільшити вміст VLCFA, включаючи позицію sn-2 Крім того, можна передбачити, щоб дріжджові трансформанти SLC1 та SLC1-1 були добрими хазяями для трансформації генами пдроксилази рицини (R commums) та різних видів Lesquerella для продукування насінних олій, збагачених гідрокси жирними кислотами Або ж, трансформанти пдроксилази можуть бути статево схрещені з трансформантами SLC11 SLC1 Аналіз ЛІПІДІВ насіння SLC1-1 трансформантів A thahana Дані з трансформації Arabidopsis thahana свідчать, що ген має суттєвий вплив на загальний вміст ЛІПІДІВ та sn-2-композицію TAG Велика КІЛЬКІСТЬ І2 трансгенних ЛІНІЙ SLC1-1 (21 з 48) показали значне збільшення продукування олії порівняно з нетрансформованими контролями та 13 49833 контролями рВИ 21 (без вставки SLC1-1), як показано нижче у Таблиці 2 14 Таблиця 2 Вмісту жирних кислот у насінні нетрансформованого дикого типу (u-WT) A thahana, pB1121 (-SLC1 -1), трансформантів A thahana (контролі) та відібраних Тг трансгенних ЛІНІЙ A thahana, трансформованих дріжджовим геном SLC1-1 (Цифри означають вміст жирних кислот (мкг/50 зерен) ЛІНІЯ u-WT контр РВИ 21 контр 3 7 16 20 21 22 23 26 29 39 42 52 54 16 0 18 0 18 1с9 18 1с1 18 2 1 183 20 0 20 1 20 2 22 0 22 1 24 0 +За гал 24 1 28 2 12 2 50,5 5,7 101 1 71,9 7,7 74, 8 8,7 1,9 8, 3 1,3 372,5 28 28 37 33 36 32 31 35 32 29 32 24 33 33 12 4 54,2 12 3 57,4 18 4 102,9 12 5 63,0 16 1 87,7 14 6 62,5 13 0 57,3 15 7 72,5 14 5 67,2 13 5 57,7 13 7 72,8 11 7 58,6 15 1 78,3 14 0 73,1 4,1 5,7 5,9 6,2 7,6 6,2 5,9 7,5 6,4 6,4 6,3 5,2 6,5 6,8 99, 9 114 1 111 6 131 7 153 3 121 3 113 9 139 7 124 1 114 0 129 3 123 0 116 9 131 3 6,7 7,7 7,6 9,0 10,7 9,4 8,6 10,5 9,7 9,4 8,9 11,8 11,2 10,1 74, 0 82, 5 71, 8 96, 4 118 8 89, 3 85, 8 106 9 94, 4 89, 5 100 2 103 6 110 0 119 5 7,1 9,6 8,0 12,6 11,6 9,9 10,2 12,5 10,3 10,6 9,7 11,8 9,0 11,5 СЛІД* 7, 2 СЛІД* 5,7 8, 6 360,2 412,3 461,0 473,0 556,4 448,5 426,5 512,7 461,1 426,7 469,7 456,2 456,2 506,3 4 8 1 0 3 1 9 4 6 3 2 4 4 0 66,2 78,8 84,4 95,0 95,9 89,1 86,7 95,6 87,6 81,6 82,0 83,0 57,8 91,2 2,9 7,7 1,7 2,7 1,7 2,5 11 0 2,0 12 4 3,3 2,2 9, 7 2,4 1,7 2,3 9,6 2,0 11 7 2,6 2,3 9, 7 2,3 1,9 2,3 11 0 2,0 10 3 2,6 17 4 2,1 3,3 2,5 12 6 3,0 3,0 11 6 1,3 СЛІД*-СЛІДОВИЙ вміст, < 0,2% ваги Особливістю деяких із цих Тг-ліній SLC1-1 є пропорції TAG, що містять VLCFA (наприклад, у таблицях 3 та 4), а отже, загальний вміст VLCFA в насінні, особливо ейкозенової кислоти та ерукової кислоти, значно підвищилися (Таблиця 5) У деяких випадках були також підвищені пропорції VLCFA в цілому (Таблиця 6) Трансформовані SLC1-1 Тг-лінм, які виявляли найбільш обнадійливі результати щодо підвищення вмісту олії та збільшення пропорцій TAG, що містять VLCFA, були відібрані, і окремі зерна висівалися для отримання лінії потомства Тз ЛІПІДНИЙ аналіз TAG різних незалежних SLC1 -1 трансгенних ТЗ-ЛІНІЙ показав, що загальний вміст ЛІПІДІВ був значно підвищений (показано у мкг жирних кислот/100 зерен, Таблиця 7), що було пов'язано з підвищеним вмістом TAG (нмоль TAG/100 зерен, Таблиця 8) порівняно з контрольними Тзтрансформантами рВ1121 Зокрема, КІЛЬКІСТЬ VLCFA (мкг/100 зерен, Таблиця 7) та рівні Css та Сєо TAG, що містять VLCFA (Таблиця 8) великою мірою підвищилися у кількох трансформантів SLC1-1 порівняно з контрольними рослинами рВИ 21 Стереоспецифічні аналізи TAG з відібраних незалежних Тз SLC1-1 трансгенних рослин показали підвищення пропорцій VLCFA (наприклад, ейкозенової кислоти, 20 1) у позиції sn-2 Ця тенденція є послідовною, незалежно від того, чи були дані виражені як пропорції серед усіх жирних кислот позиції sn-2, які представлені ейкозеновою кислотою, чи як пропорції загальної КІЛЬКОСТІ ейкозенової кислоти у TAG, яку виявлено у позиції sn-2 (Таблиця 9) Більш ТОГО, у трансгенних рослинах SLC1-1 збільшення пропорцій VLCFA (наприклад, ейкозенової кислоти) у позиції sn-2 TAG було пов'язане з паралельним зниженням пропорцій поліненасичених жирних кислот у цій позиції порівняно з контрольними рослинами рВ1121 (Фіг 4) 15 49833 16 Таблиця З Види TAG, акумульовані у Тг насінні нетрансформованого контролю дикого типу A thahana та трансформанту SLC1-1 # 42 (нмоль/50 зерен ± SD) ЛІНІЯ TAG С#-^ С5П С5? 49833 17 18 Таблиця 7 Загальний вміст ЛІПІДІВ (МКГ загальних ЖК/100 зерен) та вміст VLCFA (мкг/100 зерен) у достиглому ТЗ насінні контролів рВИ 21 та відібраних SLCI-1 -трансгенних ЛІНІЯХ A thahana (мкг/100 зерен) Загальний вміст ЛІПІДІВ 483 5 568 5 519 7 511 3 520 7 153 11379 851 7 984 6 1056 1 11092 870 0 1039 1 1955 3 ЛІНІЯ рВИ 21 контроль а рВИ 21 контроль б рВИ 21 контроль в рВИ 21 контроль г рВ1121 контроль Avg ± SE (п=4) 42-1 42-4 42-5 23-8 52-2 52-5 52-6 16-5 Вміст VLCFA 1197 127 2 125 1 122 3 123 6 14 315 5 218 6 268 0 287 7 307 5 253 3 281 6 227 0 Таблиця 8 Загальний вміст TAG та вміст TAG Css і Сєо У достиглому насінні Тз насіння контролів рВИ 21 та відібраних трансгенних ЛІНІЯХ SLC1-1 A thahana (нмоль/100 зразків насіння) TAG С # -» С50 Контр рВИ 21 8,5 0,4 ± SE (n=6) 16-5 12,4 23-8 17,7 42-4 11,4 52-6 15,2 н в - не визначено с52 55,3 2,6 88,2 130,8 90,7 106,1 С54 130,9 Сбб 145,3 Сб8 30,9 7,8 9,0 2,7 214,7 251,6 70,5 333,6 409,0 106,8 259,6 366,4 127,7 252,1 322,7 85,5 Сєо нв* 5,6 8,0 14,3 6,0 Загал ьн 371,0 21,6 642,9 1005,9 870,0 787,7 Таблиця 9 Пропорції 20 1 в sn-2 позиції TAG (% ваги sn-2 20 1) та пропорції усіх 20 1, виявлених в sn-2 позиції TAG (% ваги від усіх 20 1 в sn-2 позиції") у достиглому Тз насінні контролів рВИ 21 та відібраних трансгенних ЛІНІЯХ SLC1-1 A thahana (% ваги/100 зразків насіння) ЛІНІЯ % ваги sn-2 20 1 контроль рВИ 21 а 1,7 контроль рВИ 21 б 0,6 контроль рВИ 21 в 0,5 контроль рВИ 21 г 1,6 16-5 4,2 42-1 5,1 42-4 7,9 42-5 5,3 23-8 7,5 52-2 6,2 52-5 5,8 52-6 7,5 % від усіх 20 1 в sn-2 позиції = (% в [sn-2/[3 х % усіх 20 1]] х 100) Аналізи ЛІПІДІВ насіння трансформантів SLC11 В париєтаВ cannata Деякі з SLC1-1 Тг-трансформантних ЛІНІЙ насіння сортів В napus - Hero, Reston та насіння В cannata виявили підвищення вмісту олії (Таблиця 10) та підвищення вмісту ерукової кислоти, виражене у мкг/мг DW або у мкг/зернину (Таблиця 11) % ваги усіх 20 1 в sn-2 поз * 3,6 1,1 0,9 3,0 16,3 8,5 12,8 8,7 12,0 10,0 9,7 12,0 У сортів В napus Hero та Reston насіння кількох трансгенних ЛІНІЙ SLC1-1 виявляло підвищення пропорцій ерукової кислоти (Таблиця 12) порівняно з ВІДПОВІДНИМИ рівнями нетрансформованих контрольних рослин Аналізи окремих зернин пересічних нетрансформованих рослин сорту Hero (рослина 4) та трансформованих ЛІНІЙ SLC1-1 з 19 49833 перспективно високим продукуванням олії та високим фенотипом ерукової кислоти (ЛІНІЯ 8, рослина 6) продемонстрували розподіл цих ознак, що вказують на популяцію насіння, що сегрегує типовим методом Менделя для однієї інсерцм (Таблиця 13) Деякі зразки насіння Hero Ліни 8, рослини 6 демонстрували, можливо, гомозиготний фенотип дикого типу (наприклад, насіння 8-61) або гомози 20 готний фенотип SLC1-1 (наприклад, насіння 8-6К та 8-6Н) для всіх трьох ознак (високе продукування олії, підвищення вмісту ерукової кислоти, підвищення пропорцій ерукової кислоти), тоді, як ІНШІ виявляли скоріше гетерозиготні профілі Дикий тип/SLCI-i з проміжними значеннями для цих трьох характерних ознак (наприклад, насіння 86В) Таблиця 10 Продукування олії (% сухої ваги) в насінні Тг нетрансформованого контролю та відібраних трансгенних ЛІНІЙ SLC1-1 сортів В napus HERO та RESTON та у селекційній лінії В carmata C90-1163 (± SE, де потрібно) Продукування олії (% сухої ваги) ЛІНІЯ В napus, сорт HERO Контроль 5-1 5-4 7-3 7-6 7-9 8-4 8-6 8-7 8-Ю В napus, сорт RESTON Контроль 1 -7 1 -8 2-8 2-9 ЛІНІЯ Brassica carmata C90-1163 Контроль В carmata 10-1-7 В carmata 2-3-6 40,1 ±1,7 46,7 48,7 45,3 46,4 44,9 45,9 50,9 44,9 45,1 33 4 ± 2 2 41,9 40,5 42,1 42,2 35,9 ±1,1 42,8 39,9 Таблиця 11 Вміст ерукової кислоти (виражений у мкг/мг сух ваги або мкг/зернину) у достиглому насінні Тг нетрансформованого контролю та відібраних трансгенних ЛІНІЙ SLC1-1 В napus, сорт HERO, та селекційної лінії В сапnata С90-1163 (± SE для контролів) ЛІНІЯ ЛІНІЯ Brassica carmata C90-1163 Контроль 10-1-7 В napus, сорт HERO Контроль 5-1 5-4 7-3 7-6 7-9 8-4 8-6 8-7 8-Ю 3-1 22 1 (мкг/мг сух ваги ваги ) 22 1 (мкг/зернину) 156,4 ±5,6 180,4 195,5 ±11,7 247,9 249,4 236,1 244,8 229,2 235,7 270,9 238,5 232,7 - 597,7 ± 40,6 900,6 818,8 912 857,6 923,2 1020,3 888,3 900,4 - — не визначено Таблиця 12 49833 21 22 Пропорції ерукової кислоти (виражені у % ваги) у достиглому Т3 насінні нетрансформованого контролю та відібраних трансгенних ЛІНІЯХ В napus, сорти HERO та RESTON (± SE для контролів) % ваги 22 1 ЛІНІЯ В napus, сорт HERO Контроль 5-1 5-4 7-3 7-6 7-9 8-1 8-6 8-7 8-10 3-1 В napus, сорт RESTON Контроль 1 -10 1 -7 1 -8 2-3 2-7 - не визначено 48,6 ±0,6 53,1 -52,1 52,8 -31,4 53,3 51,8 53,6 58,3 34,7 4- 0,2 36,4 35,8 37,4 36,6 41,1 Таблиця 13 Варіанти вмісту ЛІПІДІВ (вираженого у мкг усіх жирних кислот/зернину) та ерукової кислоти (вираженого у мкг 22 1/зернину або у % ваги 22 1) в окремих зернах достиглого насіння Тг нетрансформованої контрольної рослини 4 та SLC1-1 трансгенної рослини 6 лінії 8 В napus, сорт HERO (± SE для середніх значень (AVG)) ЛІНІЯ / насіння AVG Контроль 4 AVG-8 6 8 6G 8 6Н 8 6І 8 6J 8 6К 8 6А 8 6В 8 6С 8 6D 8 6Е 8 6F мкгЖК/ зернину 1076,7 ±61,5 1441,7 ±67,3 1324,8 1704,3 1175,4 1206,8 1694,7 1351,6 1304,5 1221,11449,0 1678,2 1748,0 У деяких трансформованих ЛІНІЯХ Hero спостерігалося помірне збільшення пропорцій ерукової кислоти й загальних пропорцій VLCFA в позиції sn-2 (Таблиця 14) Вплив дріжджового трансгену на підвищення вмісту sn-2 ерукової кислоти у В napus був дещо меншим, за його мкг 22 1 /зернину 507,1 ±33,7 735,4 ± 36,5 710,8 877,1 557,3 629,4 911,1 658,6 670,6 639,1 714,3 844,6 876,8 % ваги 22 1 46,9 ±0,8 51,0 ±0,6 54,1 52,5 47,4 52,2 53,8 48,7 51,4 52,3 49,3 50,3 50,2 здатність змінювати вміст sn-2 ейкозенової кислоти в A thahana (Таблиця 9) Втім, цього можна було очікувати через відносну специфічність sn-2 ацилтрансферази S cerevisiae до різновиду ейкозеноілу - ерукоілу-СоА (Таблиця 1) Таблиця 14 Вміст sn-2 ерукової кислоти та VLCFA у достиглому Тг насінні нетрансформованого контролю та відібраних трансгенних ЛІНІЙ SLC1-1 В napus, сорт HERO Лінія/насіння Контроль HERO HERO 8 - 6 HERO 8 - 6 G (одна зерн ) HERO 3 -1 HERO 8-10 *детектовано лише ерукову кислоту як sn-2 VLCFA sn-2 22 1 1,5 2,8 3,6 4,12 2,22 sn-2 VLCFA 3 4,6 4,44 4,12* 3,7 Аналізи композицій зразків TAG з використанням GC показали, що деякі лінії трансформантів SLC1-1 Hero мали підвищені пропорції С42 TAG та, меншою мірою, С54 та Сбб TAG (Таблиця 15) Пропорції Сб2-Сб8 TAG, що містять 2 або більше С22-жирних кислот, були значно підвищене у трансгенних рослинах Hero SLC1-1 (Таблиця 15), насамперед, за рахунок TAG, які містять дві (С54) або три (С54) Сю жирні кислоти (дані не показано) Подібне підвищення пропорцій С52 TAG спостерігалося у деяких трансгенних ЛІНІЯХ SLC11 В napus, сорт Reston SLC1-1 (Таблиця 15) Таблиця 15 Пропорції С42, С54 та С4 TAG (моль %) у достиглому Т3 насінні нетрансформованого контролю та відібраних трансгенних ЛІНІЙ SLC1-1 В napus, сорти HERO та RESTON (± SE для контролів) C42 36,72 ± 1 42 51,44 46,92 58,48 57,25 55,61 44,78 53,35 С 54 1,32 ± 0 , 0 2 1,81 1,95 1,46 2,19 1,98 2,14 2,22 C44 0 , 1 0 + 0,01 0,12 0,25 0,08 0,14 0,09 0,25 0,22 Загалом С42-С62 38,14 ±1,45 53,37 51,2 58,02 59,58 57,68 47,16 55,79 18,32 23,88 31,67 ЛІНІЯ Контроль Hero 5 - 2 Hero 5 - 4 Hero 5- 10 Hero 7 - 1 Него 7 - 5 Него 8 - 4 Hero 8 - 6 Reston Контроль 1 -8 2-7 0,94 1,06 1,42 0,06 0,07 0,11 19,32 25,01 33,20 Аналізи типових контролів та SLC1-1 трансгенних рослин В napus, сорт Hero щодо варіювання пропорцій Сб2 TAG між різними зразками насіння показали, що популяція насіння SLC1-1 Тг сегрегувала, але багато окремих зерен мали значно вищі пропорції Сб2 TAG, ніж будь-який з нетрансформованих контролів (Таблиця 16) Таблиця 16 Аналізи пропорцій С 62 TAG (моль %) в окремих зразках достиглого Т2 насіння нетрансформованого контролю та трансгенних ЛІНІЙ SLC1-1 В napus, сорт HERO ( ± SE для середніх значень, AVG) C 62 TAG ЛІНІЯ /насіння Контроль Hero 4d 4е 4Ь 4f 4g 4] 4h 4і 4k Контроль Hero AVG Hero 8-6 8-6d 8-oa 8-6b 8-6c 8-6f 8-6g 8-6 h 8-61 8-6] 8-6 k Hero 8-6 AVG Оцінки підвищення 38,54 40,29 36,88 38,81 30,05 35,95 42,84 40,81 43,28 38 6 ± 1 35 36,36 47,63 54,06 54,81 44,4 56,27 53,11 42,19 51,44 58,4 51,35 ±1,82 продуктивності олії у трансгенних ЛІНІЯХ SLC1-1 порівняно з контроля ми, прямо залежали від того, чи вони виражалися в одиницях "на мг сухої ваги", чи "на зернину" (Фіг 5), як це було з оцінками відносного вмісту олії недеструктивним методом 1Н-ЯМР (Фіг 6) Дійсно, результати ЯМР щодо підвищення продукування олії також прямо залежали від зростання ваги зерен у SLC1-1 трансгенних рослин (Фіг 7) і показу вали, що підвищення сухої ваги насіння прямо залежить від підвищення вмісту олії, долею води у насінні можна знехтувати (відсутність широкого резонансу води між ХІМІЧНИМИ зсувами СН2 OCO та СНОСО-) Типові реакції 1Н-ЯМР зразків 35 зернин насіння контролю та "високоолійних" трансгенних ЛІНІЙ SLC1-1 сорту В napus - Hero та В carmata показано у Таблиці 17 Таблиця 17 Інтегральна Н-ЯМР-реакція на резонанси, що визначаються рідкою олією (як описано у Rutar, 1989) у достиглому насінні Т2 нетрансформованих контролів та відібраних трансгенних ЛІНІЙ SLC1-1 В napus, сорт HERO, та селекційної лінії С901163 В cannata (35 зразків насіння, Реакції, пов'язані з інтеграцією контролю при 1 000) інтегральна ВІДПОВІДЬ ЯМР ЛІНІЯ В napus, сорт HERO Контроль Hero 5 - 1 Hero 7 - 3 Hero 7 - 6 Hero 7 - 9 Hero 8 - 4 Hero 8 - 6 Hero 8 - 7 Hero 8- 10 В cannata, сорт С90-1163 Контроль В саг 10-1-7 В саг 2-3-6 1,0000 1,5175 1,2721 1,3875 1,3245 1,5667 1,5297 1,4825 1,6302 1,0000 1,5977 1,7548 Деякі лінії Ї2 насіння SLC1-1трансформованих В napus, сорт Westar (канола) SLC1-1 демонстрували зростання пропорцій олеїнової кислоти та паралельне зменшення пропорцій поліненасичених жирних кислот (18 2 та 18 3) (Таблиця 18) Це йде врозріз з прогнозованим ефектом, на який було вказано у патентній заявці в Університеті Кентукі Таким чином, пропорції моно-ненасичених жирних кислот можуть бути збільшені у харчових оліях шляхом експресії SLC1-1 Крім того, пропорції насичених жирних кислот з дуже довгими ланцюгами у цих ЛІНІЯХ каноли значно підвищилися (Таблиця 18) Таблиця 18 Композиції олеїнових, лінолевих, ліноленових та насичених VLCFA у нетрансформованих контролях та відібраних трансгенних ЛІНІЯХ SLC1-1 В napus, сорт WESTAR (n=2 або 3) ЛІНІЯ Олеїнова Лінолева Ліноленова Ейкозенова Бегенова Лігноцер В napus, с WESTAR Контроль WS-13 WS-15 WS-16 WS-15a 61,03 70,03 71,92 71,06 72,71 17,55 14,80 12,33 12,29 9,69 11,07 3,41 3,71 3,87 3,09 0,55 0,76 0,78 0,97 0,94 0,31 0,49 0,53 0,59 0,65 0,27 0,56 0,48 0,56 0,68 Аналізи LPAT трансформованих ЛІНІЙ Зразки SLC1-1 Ті трансформованих ЛІНІЙ В napus, сорт Westar та В napus, сорт Argentine виявляли підвищення активності 18 1-СоА 1LPAT листя у швидкопоширюваних препаратах гомогенатів листя порівняно з гомогенатами з нетрансформованих контрольних рослин (Таблиця 19) Аналізи LPAT недозрілого насіння нетрансформованих контролівта SLC1-1 трансгенних рослин В napus, сорт Него та В cannata показали, що обидві специфічні активності 181-СоА LPAT та 22 1-СоА LPAT (таблиця 19) були значно підвище ні у трансгенних SLC1 -1 -рослинах Аналізи LPAT недозрілого насіння нетрансформованих контролівта SLC1-1 трансгенних рослин A thahana показали, що активність 20 1-СоА LPAT зросла у кількох трансгенних SLC1 -1 -рослинах (Таблиця 19) Таким чином, у цій заявці ми вперше даємо пряме свідчення того, що продукт дріжджового гену SLC1-1 кодує фермент, який має активність sn-2 ацилтрансферази і який може виявляти активність LPAT in vitro Таблиця 19 Відносна активність LPAT у гомогенатах, приготованих з листя Ті та недозрілого насіння Тг або Тз нетрансформованих контролівта відібраних трансгенних ЛІНІЙ SLC1-1 сортів В napus -WESTAR, ARGENTINE та 27 49833 28 HERO, сортів В cannata - С90-1163 та сорту A thahana - COLUMBIA Усі аналізи проведено, як описано в експериментальному розділі В napusWestar Контроль WS2-5 WS3-8 WS6-7 В napus Arg Контроль Arg 2-8 Arg 3-3 В napus Hero Контроль Hero 3-1 Hero 7-6 Hero 8-6 В car C90-1163 Контроль В car 10-1-7 В napus Hero Контроль Hero 3-1 Hero 7-6 Hero 8-6 A thahana WT u -Контроль 42-1 42-4 42-5 Ті-ЛИСТЯ 18 1-СоА 307 1008 617 1428 Ті-ЛИСТЯ 18 1-СоА 350 996 1557 Тг-недозр насіння 18 1-СоА 580 3470 2035 1370 Тг недозр насіння 18 1-СоА 720 1125 Тг недозр насіння 22 1-СоА 6,4 68,3 53,4 20,2 Тз недозр насіння 20 1-СоА 238 270 380 503 Генетичні аналізи трансформантів SLC1-1 Дані аналізів PSR та Саузерна для ЛІНІЙ трансгенних рослин, подані у даній заявці, зведено у Таблиці 20 Таблиця 20 Результати аналізів PSR та Саузерна ЛІНІЙ трансгенних рослин SLC1-1 (н в = не визначено) Олійне насіння A thahana COLUMBIA В napus WESTAR В napus ARGENTINE В napus HERO В cannata C90-1163 Трансформант # (ЛІНІЯ Т2) 16 20 23 42 52 54 2 3 6 13 15 16 2 3 5 7 8 3 10 2 PSR + + + + + + + + + H В H В H В + + + + + + + + Саузерн Інсерція # одна одна кілька кілька кілька кілька кілька кілька кілька одна кілька кілька кілька кілька одна одна одна одна одна кілька 29 49833 Щоб простежити хараісгер сегрегації у Тггенераціях SLC1 -1 -трансформантів A thahana, насіння з трансгенних ЛІНІЙ (наприклад, ЛІНІЙ 16, 20), яке виявляло підвищення вмісту олії та КІЛЬКОСТІ жирних кислот з довгими (С-із) та дуже (С20 та С22) стерилізували й пророщували на селективному середовищі (50мг/л канаміцину) Характер сегрегації обох ЛІНІЙ був однаковий 3 1 (канаміцинСТІЙКІСТЬ канаміцин-чутливість), що свідчить про те, що маркер сегрегує як один локус Менделя Аналізи гібридизації Саузерна (Southern, 1975) підтвердили наявність однієї інсерцм Т-ДНК на геном У ЛІНІЯХ 23, 42, 52 та 54 аналіз гібридизації за Саузерном передбачає, що усе лінії мають більше однієї інсерцм Т-ДНК на геном Саузерн-аналіз гібридизації насіння у середній фазі розвитку, взятого зі стручків A thahana, ЛІНІЙ 16, 20, 23, 42, 52 та 54, підтвердив експресію гену SLC1-1 в усіх випробуваних ЛІНІЯХ З найвищим рівнем експресії у лінії 42 Саузерн-аналіз геномної ДНК, яка була виділена з трансгенних ЛІНІЙ В napus сорту Westar (2, З, 6, 13, 15, 16) виявив, що лише ЛІНІЯ 13 має одну інсерцію Обидві трансгенні лінії SLC1-1 (2, 3) В napus, сорт Argentine мали кілька інсерцій Трансгенні лінії В napus, сорт Hero (3, 5, 7, 8) та трансгенна ЛІНІЯ 10 В carmata мали по одній інсерцм, тоді, як ЛІНІЯ 2 В carmata мала кілька інсерцій ТДНК на геном ПЕРЕЛІК ПОСЛІДОВНОСТЕЙ (1) ЗАГАЛЬНА ІНФОРМАЦІЯ (і) ЗАЯВНИК (A) ІМ'Я National Research Council of Canada (Б) ВУЛИЦЯ 1200 Montreal Road (B) МІСТО Ottawa (Г) ПРОВІНЦІЯ Ontario (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС К1А OR6 (A) ІМ'Я Zou, Jitao (Б) ВУЛИЦЯ # ЗЕ-1800 Mam Street (B) МІСТО Saskatoon (Г) ПРОВІНЦІЯ Saskatchewan (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС S7H 2Z6 (A) ІМ'Я Taylor, David С (Б) ВУЛИЦЯ 6 2 2 W o l l a s t o n B a y (B) МІСТО Saskatoon ЗО (Г) ПРОВІНЦІЯ Saskatchewan (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС S7J 4СЗ (A) ІМ'Я Katavic, Vesna (Б) ВУЛИЦЯ 301 1121 С McKercherDnve (B) МІСТО Saskatoon (Г) ПРОВІНЦІЯ Saskatchewan (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС S7H 5В8 (A) ИМЯ MacKenzie, Samuel L (Б) ВУЛИЦЯ 17 Cambridge Crescent (B) МІСТО Saskatoon (Г) ПРОВІНЦІЯ Saskatchewan (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС S7H ЗР9 (A) ІМ'Я Keller, Wilfred A (Б) ВУЛИЦЯ 234 Emmeline Road (B) МІСТО Saskatoon (Г) ПРОВІНЦІЯ Saskatchewan (Д) КРАЇНА Canada (Е) ПОШТОВИЙ ІНДЕКС S7J 5В6 00 НАЗВА ВИНАХОДУ МОДИФІКАЦІЯ ЛІПІДІВ РОСЛИН ТА НАСІННИХ ОЛІЙ ШЛЯХОМ ВИКОРИСТАННЯ ДРІЖДЖОВИХ ГЕНІВ SLC (їм) ЧИСЛО ПОСЛІДОВНОСТЕЙ б (iv) ФОРМА КОМП'ЮТЕРНОГО ЧИТАННЯ (A) СПОСІБ ЗБЕРІГАННЯ ІНФОРМАЦІЇ дискета (Б) КОМП'ЮТЕР сумісний з IBM PC (B) ОПЕРАЦІЙНА СИСТЕМА PC-DOS/MSDOS (Г) ПРОГРАМНЕ ЗАБЕЗПЕЧЕННЯ Patentm Release #1 0, Версія # 1 ЗО (ЕРО) (2) ІНФОРМАЦІЯ ПРО SEQ ID N0 1 (і) ХАРАКТЕРИСТИКИ ПОСЛІДОВНОСТЕЙ (A) ДОВЖИНА 947 основних пар (Б) ТИП нуклеїнова кислота (B) ЛАНЦЮГОВІСТЬ одиночна (Г) ТОПОЛОГІЯ лінійна (м) ТИП МОЛЕКУЛИ ДНК(геномна) (vi) ПЕРВІСНЕ ДЖЕРЕЛО (А) ОРГАНІЗМ Saccharomyces cerevisiae (їх) ОСОБЛИВОСТІ (A) НАЗВА/ КЛЮЧ CDS (B) ЛОКАЛІЗАЦІЯ 1 909 (хі) ОПИС ПОСЛІДОВНОСТІ SEQ ID № 1 31 АТС AGT Ma; Ser X 32 49833 CTG Vai ATA lie CTT Leu TGC Cys CGT Arq TIC Phe TTC Leu ТА? TAC TTG T\: їуї Leu 10 AGG TCC GTG Arc Set: Val * GGC Gly TGT Cys GGC Giy TTC TAC Рпз Tyr 25 GGT Gly GTA Val АТС lie АТС lie G3T Gly AAG Lys CAA CAT TTG Gin his Leu 40 „ GCT Ala CTG Leu TGG ATT Trp H e 45 TAC Туг " СТА CTG GCG Mai. L E U Ala GGT AGG Gly Arg 5 CAT Ніз GTC Val ATG AAA Nat: Lys 55 TTG h~Q Leu Kec CTT Leu GGC CTT Gly Leu 60 CCA TAT Pro Tyr 75 ТТЛ Leo TTG Leu CTT GCA Leu Ala 20 ACG ТТЛ ТГЇГ Lsu 35 TCT TTT Cys Pha SO TTG Let. 15 CTC Vil GCC TCT Ala Ser 30 АТС lie ACT The GCG Ala GAC Asp GTC Val AAG Lys ATT lie ATG Met ATT lie GCC Ala BO GGT GXy AGG Arg ATT lie TTC Phe CCC Pro 2B8 AAA Lys TAC Tyr GTC CCC Vai fro 110 TTT Phe 33$ CTG CGT TGG TTC ATG GCT TTG »GT GGT АСА TAT T T C T T A GAC АСА TCT U u C l y T r p P h e M e t A l a L e u S e t G l y ThK Т у г P h e L e u R s p Я г д S e c 115 120 135 384 GTC GTT Val Vai 65 GGC Gly GAG Glu GAG Giu ЛАТ TTG Asn Leu 70 GCC Ala AAG Lys ЛАТ Asn САС His САД Gli ТСС Ser ACC Thr TT.G GAT Leu Asp АТС lie TTC ATG She Met CCT Pro GGT Gly TGC Cys АСА Thr 100 GTT Val ACT Thr AAG Lys AAG TC7 L/s Sec 103 as AAA Lys AAG Lyi эо GCC Ala 35 AGG CAA GAA CCC ATT GAC ЛСС TTC AAT AAA A c g С і л G l u A l a l i e A s p T h r Lew ftsn tys 130 135 AAG AAA Lys Lys 14S CGT TTA GAA AAT GTT G l y L e u QiM ftsn V a l 140 AAC AAG CGT GCT СТА TGG G?T TTT CCT GAG GGT ACC AG.C TCT A a n Lys A r g Ala Leu T c p Val Phe P r o G l u G l y T h e Arg S e r 150 155 160
ДивитисяДодаткова інформація
Назва патенту англійськоюPlasmid pslc1-1/prd400, strain gv3101 agrobacterium tumefaciens, method for obtaining transgenic plant with oilseeds, method for separation of eatable or non-eatable oil from plant oilseed
Назва патенту російськоюПлазмида pslc1-1/prd400, штамм gv3101 agrobacterium tumefaciens, способ получения трансгенного растения с маслосодержащими семенами, способ выделения пищевого или непищевого растительного масла из маслосодержащих семян растения
МПК / Мітки
МПК: C12N 15/09, C11B 1/00, C12N 15/00, A01H 5/00, C11B 1/10, C12N 9/10, C12N 5/10, C12N 15/82, C12N 1/21
Мітки: рослини, олії, спосіб, трансгенної, agrobacterium, tumefaciens, насінням, плазміда, неїстівної, масловмісного, одержання, виділення, їстівної, штам, насіння, gv3101, масловмісним
Код посилання
<a href="https://ua.patents.su/25-49833-plazmida-pslc1-1-prd400-shtam-gv3101-agrobacterium-tumefaciens-sposib-oderzhannya-transgenno-roslini-z-maslovmisnim-nasinnyam-sposib-vidilennya-stivno-abo-nestivno-oli-z-maslovmisn.html" target="_blank" rel="follow" title="База патентів України">Плазміда pslc1-1/prd400, штам gv3101 agrobacterium tumefaciens, спосіб одержання трансгенної рослини з масловмісним насінням, спосіб виділення їстівної або неїстівної олії з масловмісного насіння рослини</a>
Спосіб отримання рослини зі зниженою сприйнятливістю до рослинних паразитичних нематод (варіанти), рекомбінантна днк (варіанти), вектор трансформації рослин, штам agrobacterium та спосіб зменшення збитків урожаю

Номер патенту: 27418
Опубліковано: 15.09.2000
Автори: Годдейн Оскар Йоханнес Марія, Сеймонс Петер Крістіан, Ван Ден Елзен Петрус Йосефус Марія, ван дер Лі Фредерік Маріанна
МПК: A01H 5/00, C12N 9/14, C12N 1/21, C12N 9/10, C12N 5/10, C07K 14/415, C12N 15/82, C12N 15/09
Мітки: трансформації, сприйнятливістю, урожаю, рослини, спосіб, днк, паразитичних, рекомбінантна, отримання, збитків, вектор, штам, зменшення, рослин, agrobacterium, зниженою, рослинних, нематод, варіанти
Текст:
...характер экспрессии, определенный в данном изобретении. Предпочтительно, чтобы промотор был проMOTOPOM-35S мозаичного вируса цветной капусты или его производными, которые, как считают, обычно являются сильными образующими цепь промоторами в тканях растений (Odell и др. 1985, Nature 313, 810-812). Другим предпочтительным примером промотора-В является сильный корневой промотор rot D (Leach и Royagi 1991, Plant. Sci. 79, 69-76) из плазмиды...
Спосіб одержання трансгенної рослини, послідовність днк, яка кодує поліфруктан (леван) сахаразу, рослинна клітина

Номер патенту: 40598
Опубліковано: 15.08.2001
Автори: Рьобер Мануела, Вілльмітцер Лотар, Гайдер Клаус, Гайєр Гебхардт
МПК: C12N 9/10, C12N 15/82
Мітки: яка, рослини, послідовність, поліфруктан, кодує, клітина, трансгенної, спосіб, рослинна, днк, леван, одержання, сахаразу
Формула / Реферат:
1. Способ получения трансгенного растения, характеризующегося модифицированным образованием полифруктана в хлоропластах, амилопластах, митохондриях, цитозоле и/или вакуолях, который включает следующие стадии:а) введение выделенной молекулы ДНК в растительную клетку таким образом, что она способна встраиваться в геном указанной клетки, причем указанная молекула ДНК включает(I) промотор, способный регулировать транскрипцию гена...
Спосіб виробництва олії “гарбіол” із насіння гарбуза”

Номер патенту: 41721
Опубліковано: 17.09.2001
Автори: Мисюк Олександр Тимофійович, Колісник Олексій Федорович
МПК: C11B 1/10
Мітки: спосіб, насіння, виробництва, гарбуза, олії, гарбіол
Формула / Реферат:
1. Спосіб виробництва олії із насіння гарбуза, що включає очистку, сортування, сушіння, пресування насіння, фільтрування та розфасування олії, який відрізняється тим, що сушіння насіння ведуть до вологості 3-5%, після чого додатково проводять провіювання просушеного насіння та його знезаражування кварцовим опромінюванням, а температурні режими технологічної обробки гарбузового насіння знаходяться у межах 40-50°С.2. Спосіб за п.1, який...
Вектор для введення бажаного гена у рослину (варіанти), спосіб одержання трансгенної рослини, спосіб введення, принаймні, двох бажаних генів у рослину

Номер патенту: 40661
Опубліковано: 15.08.2001
Автори: Уєсугі Мікіко, Ебінума Хіроясу, Сугіта Коіті, Мацунага Етсуко
МПК: C07H 21/04, A01H 1/00, C12N 15/29, C12N 15/82, C12N 15/09, C12N 5/10, C12N 5/00, C12N 1/21
Мітки: одержання, варіанти, двох, вектор, бажаних, генів, рослину, спосіб, гена, трансгенної, введення, принаймні, бажаного, рослини
Формула / Реферат:
1. Вектор для введения желаемого гена в растение, включающий указанный желаемый ген и, по крайней мере, один ген с индукцией морфологической аномальности в качестве селективного гена-маркера, причем указанный индуцирующий морфологическую аномальностъ ген представляет собой ген синтеза цитокинина.2. Вектор по п.1, в котором указанный ген синтеза цитокинина представляет собой ген ipt, изопентилтрансферазы, присутствующий в Т-ДНК...
Спосіб отримання резистентної до вірусу трансгенної рослини

Номер патенту: 29438
Опубліковано: 15.11.2000
Автори: Труве Ерккі, Теері Теему, Келве Мерікке, Саарма Март
МПК: C12N 9/12, A01H 5/00, C12N 15/09, C12N 15/82, C12N 5/10, C12N 15/54
Мітки: трансгенної, рослини, спосіб, вірусу, отримання, резистентної
Текст:
...29438 Пробы листьев анализируют методом EUSA с использованием иммунодиагшстачесшго набора для PVX (Boehringer Mannheim). Концентрацию вирусов определяют по оптической плотности цветной реакции при 405 нм. Контр. = контрольный клон; R101, R102, R104, R106, R107, и R2 =трансгенные клоны. Следующие примеры иллюстрируют изобретение. Дополнительные пояснения в отношении методов молекулярной биологии, применяемых в данном изобретении,...
Попередній патент: Спосіб виробництва пресервів з розібраної риби
Випадковий патент: Спосіб стимуляції інтенсивності сомітогенезу птиці монохроматичним світлом