Генетична модифікація mon89788 сої і способи її визначення
Номер патенту: 94582
Опубліковано: 25.05.2011
Автори: Тейлор Ненсі, Райнхарт Дженніфер, Дікінсон Еллен, Малвен Маріанне




















Формула / Реферат
1. Послідовність нуклеїнових кислот, яка включає послідовність SEQ ID NO:1, 2, 3 або 4.
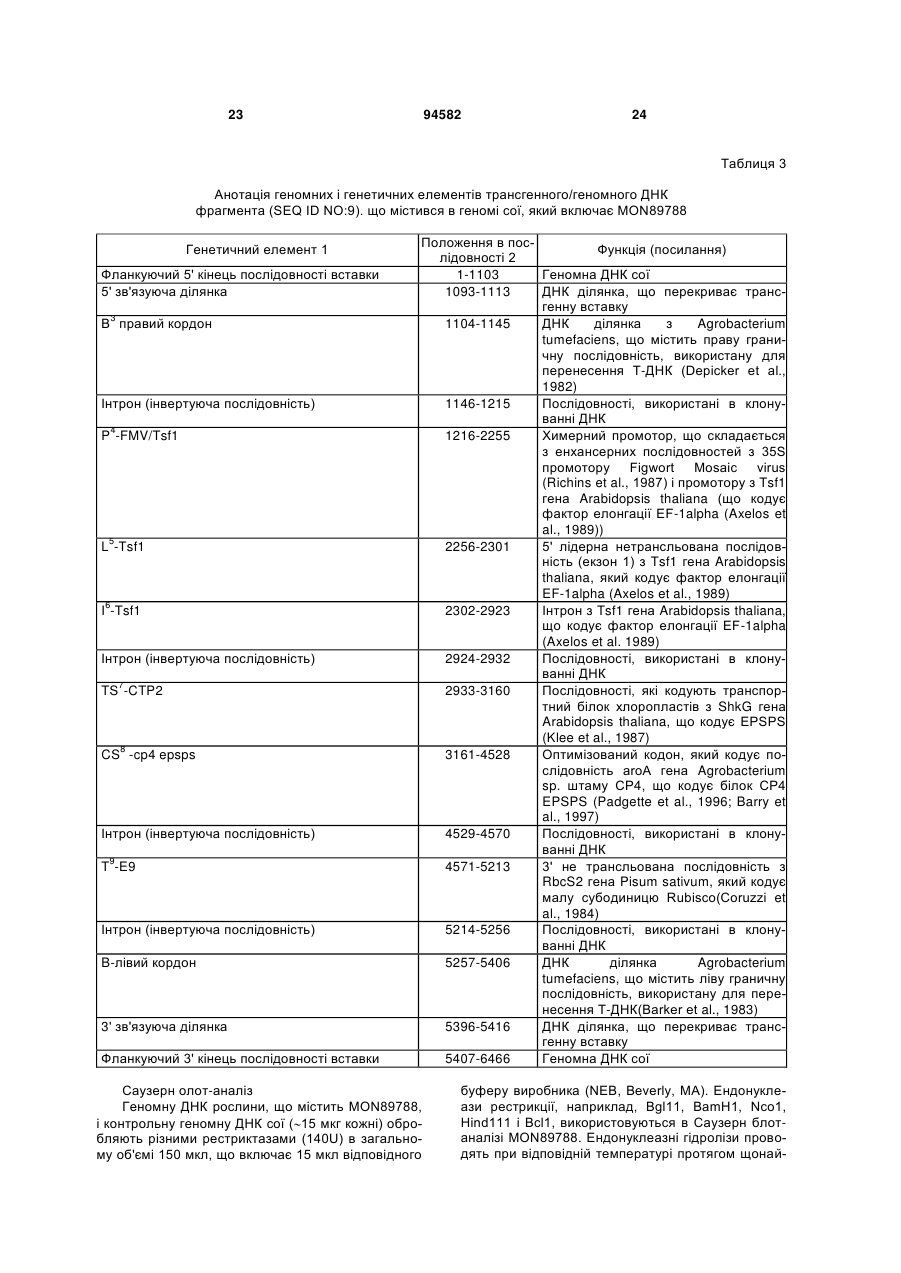
2. Рослина сої або її частина, яка включає генну модифікацію MON89788, де насіння сої, що включає генну модифікацію MON89788, депоноване в ATCC під номером PTA-6708, де геном вказаної рослини включає щонайменше одну молекулу ДНК, вибрану з групи, що складається з SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:21 і SEQ ID NO:22.
3. Частина рослини сої за п. 2, яка являє собою клітину, пилок, насінний зачаток, квітку, пагін, корінь або лист.
4. Рослина сої за п. 2, яка являє собою потомство будь-якого покоління рослини сої, що включає вказану генну модифікацію MON89788.
5. Рослина сої за п. 2, в якій геном вказаної рослини здатний виробляти діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, причому вищевказаний амплікон включає SEQ ID NO:1, 2, 3 або 4.
6. Насіння рослини за п. 2, де насіння містить генетичну модифікацію MON89788.
7. Насіння рослини за п. 6, в якому ДНК насіння здатна виробляти діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, де вказаний амплікон включає SEQ ID NO:1, 2, 3 або 4.
8. Товарний продукт з сої, отриманої з насіння за п. 6, що включає нуклеїнову кислоту, яка виробляє діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, де вищевказаний амплікон включає SEQ ID NO:1, 2, 3 або 4.
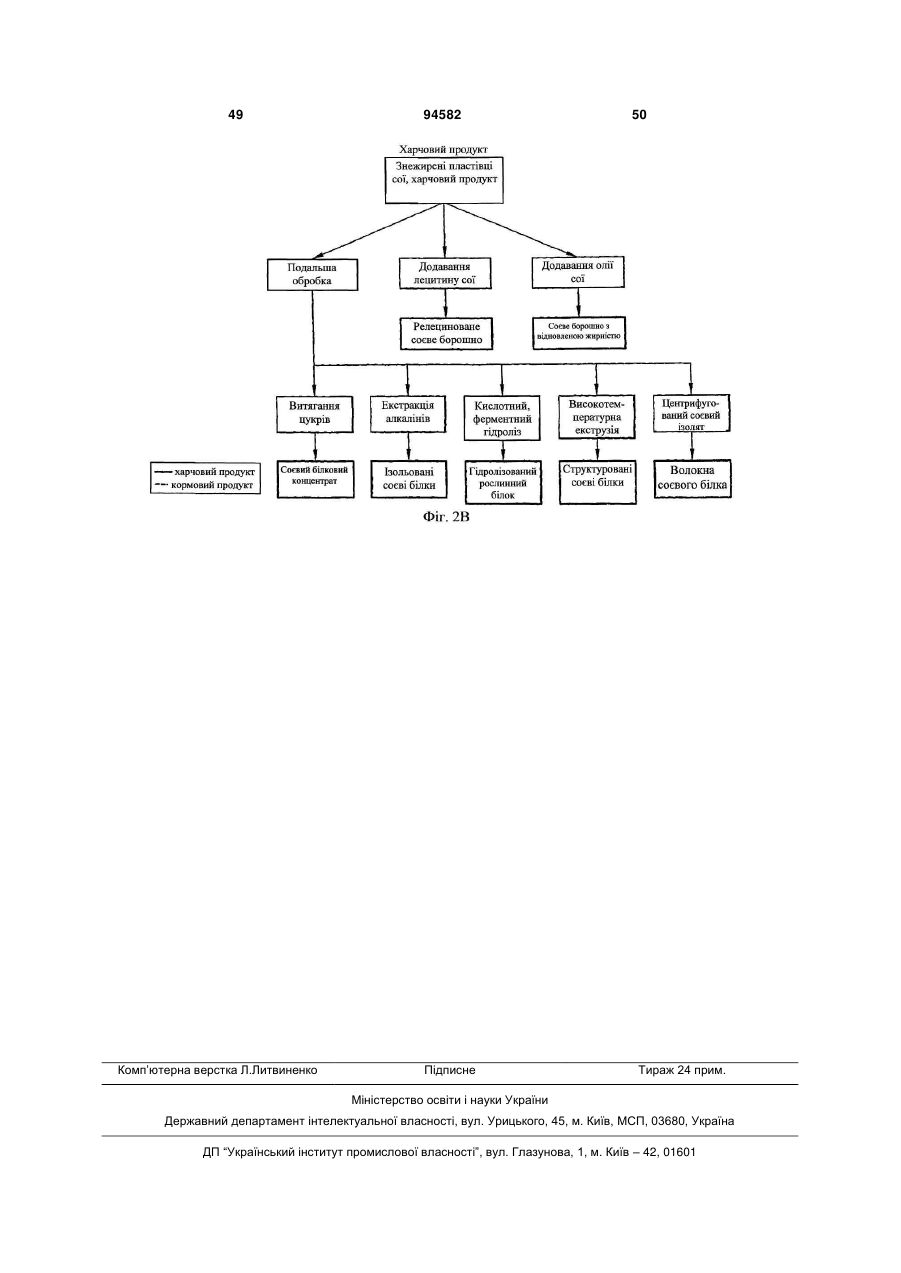
9. Товарний продукт з сої за п. 8, який являє собою крупу, борошно, пластівці або олію.
10. Полінуклеотидна молекула ДНК праймера, що включає щонайменше 11 суміжних нуклеотидів з SEQ ID NO:3, або її комплемент, яка використовується в способі ампліфікації ДНК для отримання діагностичного амплікону для генної модифікації MON89788.
11. Виділена полінуклеотидна молекула ДНК праймера, що включає щонайменше 11 суміжних нуклеотидів з SEQ ID NO:4, або її комплемент, яка використовується в способі ампліфікації ДНК для отримання діагностичного амплікону для випадку MON89788.
12. Набір для виявлення ДНК, специфічний для генної модифікації MON89788, що включає щонайменше нуклеїнову кислоту, яка включає 11 або більше суміжних нуклеотидів гомологічних або комплементарних до SEQ ID NO:3 або SEQ ID NO:4.
13. Спосіб отримання стійкої до гербіциду гліфосату рослини сої, що включає введення в геном вказаної рослини генної модифікації MON89788.
14. Спосіб за п. 13, що включає стадії:
(a) схрещування першої рослини сої, що включає генну модифікацію MON89788, з другою рослиною сої, яка не включає генну модифікацію MON89788, для отримання рослин потомства; і
(b) селекція щонайменше першої рослини потомства, яка включає вказану генну модифікацію MON89788 і є стійкою до гліфосату.
15. Спосіб за п. 14, що додатково включає самозапилення вказаної першої рослини потомства, для отримання другого покоління рослин потомства, і селекцію щонайменше першої рослини, гомозиготної для вказаної генної модифікації MON89788.
16. Спосіб визначення присутності ДНК, що відповідає генній модифікації сої MON89788 в зразку, де спосіб включає:
(a) контактування зразка, що включає ДНК сої, з набором праймерів, які при використанні в реакції ампліфікації нуклеїнових кислот з геномною ДНК з сої з генною модифікацією MON89788 виробляють діагностичний амплікон для генної модифікації MON89788 сої; і
(b) проведення реакції ампліфікації нуклеїнових кислот, отримуючи таким чином діагностичний амплікон; і
(с) визначення діагностичного амплікону.
17. Спосіб визначення присутності нуклеїнової кислоти, що відповідає генній модифікації MON89788, в зразку, де спосіб включає:
(a) отримання зразка ДНК сої; і
(b) дослідження зразка на присутність послідовності ДНК генної модифікації MON89788.
18. Спосіб за п. 17, в якому дослідження зразка ДНК включає визначення присутності послідовності нуклеїнової кислоти щонайменше однієї з SEQ ID NO:1, SEQ ID NO:2 або їх комплементів.
19. Рослина сої, що має стійкість до гліфосату, в якій вказана стійкість генетично пов'язана з молекулою нуклеїнової кислоти, що включає SEQ ID NO:1, 2, 3 або 4.
20. Спосіб отримання товарного продукту з сої, що включає:
(a) отримання рослини сої або її частини за п. 2; і
(b) виробництво товарного продукту сої з рослини сої або її частини.
21. Спосіб за п. 20, в якому товарний продукт з сої являє собою крупу, борошно, пластівці, виділений білок або олію.
22. Спосіб контролю зростання бур'янів на полі з рослинами сої, що включають генну модифікацію MON89788, де спосіб включає обробку поля кількістю гліфосату, ефективного для контролю росту бур'янів, в якому рослини сої виявляють стійкість до гліфосату.
23. Спосіб за п. 22, в якому обробка поля здійснюється на стадії росту від V1 до R4.
Текст
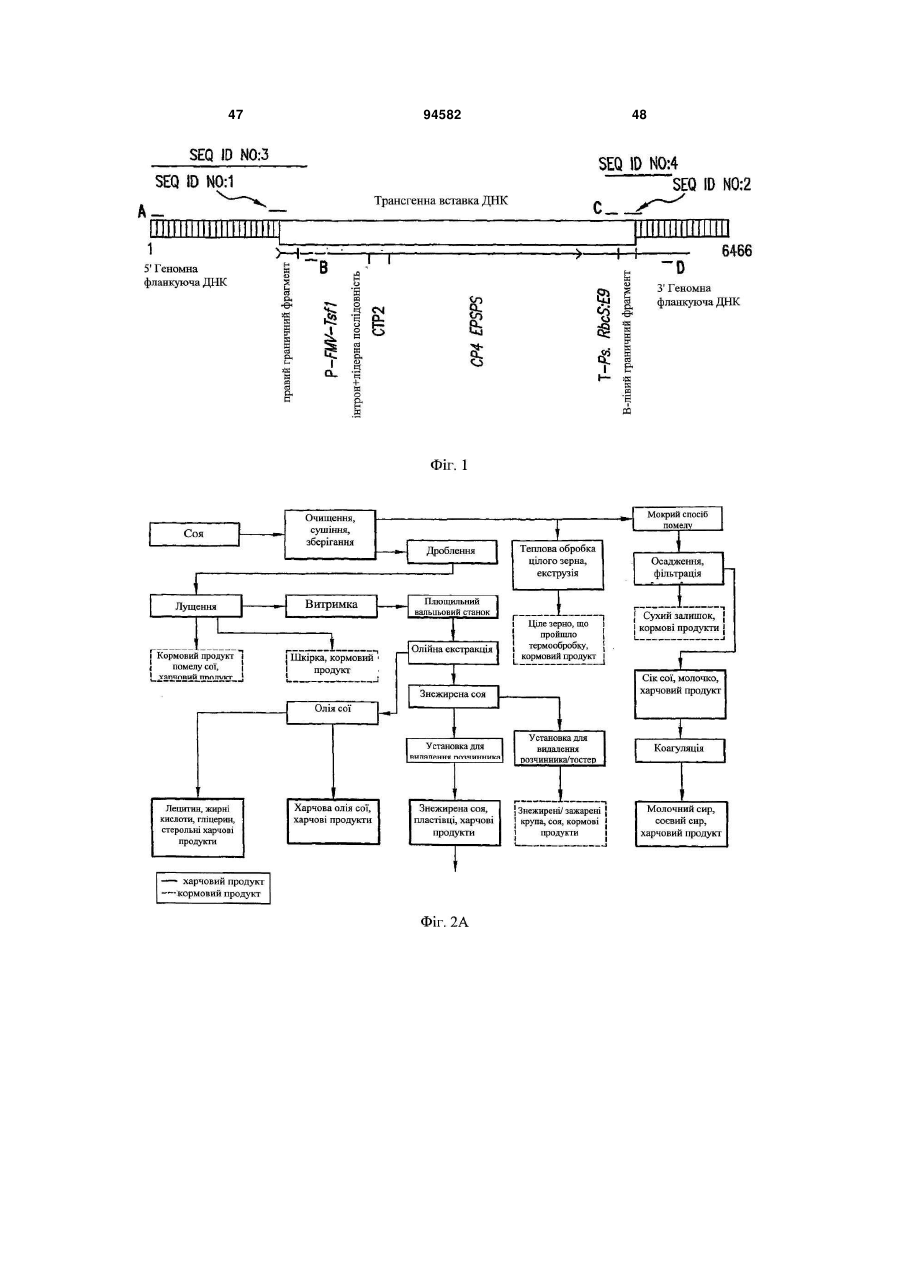
1. Послідовність нуклеїнових кислот, яка включає послідовність SEQ ID NO:1, 2, 3 або 4. 2. Рослина сої або її частина, яка включає генну модифікацію MON89788, де насіння сої, що включає генну модифікацію MON89788, депоноване в ATCC під номером PTA-6708, де геном вказаної рослини включає щонайменше одну молекулу ДНК, вибрану з групи, що складається з SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:21 і SEQ ID NO:22. 3. Частина рослини сої за п. 2, яка являє собою клітину, пилок, насінний зачаток, квітку, пагін, корінь або лист. 4. Рослина сої за п. 2, яка являє собою потомство будь-якого покоління рослини сої, що включає вказану генну модифікацію MON89788. 5. Рослина сої за п. 2, в якій геном вказаної рослини здатний виробляти діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, причому вищевказаний амплікон включає SEQ ID NO:1, 2, 3 або 4. 6. Насіння рослини за п. 2, де насіння містить генетичну модифікацію MON89788. 7. Насіння рослини за п. 6, в якому ДНК насіння здатна виробляти діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, де вказаний амплікон включає SEQ ID NO:1, 2, 3 або 4. 8. Товарний продукт з сої, отриманої з насіння за п. 6, що включає нуклеїнову кислоту, яка виробляє діагностичний амплікон для генної модифікації MON89788 при дослідженні способом ампліфікації ДНК, де вищевказаний амплікон включає SEQ ID NO:1, 2, 3 або 4. 9. Товарний продукт з сої за п. 8, який являє собою крупу, борошно, пластівці або олію. 10. Полінуклеотидна молекула ДНК праймера, що включає щонайменше 11 суміжних нуклеотидів з SEQ ID NO:3, або її комплемент, яка використовується в способі ампліфікації ДНК для отримання діагностичного амплікону для генної модифікації MON89788. 11. Виділена полінуклеотидна молекула ДНК праймера, що включає щонайменше 11 суміжних нуклеотидів з SEQ ID NO:4, або її комплемент, яка використовується в способі ампліфікації ДНК для UA (21) a200714839 (22) 26.05.2006 (24) 25.05.2011 (86) PCT/US2006/020323, 26.05.2006 (31) 60/685,584 (32) 27.05.2005 (33) US (46) 25.05.2011, Бюл.№ 10, 2011 р. (72) МАЛВЕН МАРІАННЕ, US, РАЙНХАРТ ДЖЕННІФЕР, US, ТЕЙЛОР НЕНСІ, US, ДІКІНСОН ЕЛЛЕН, US (73) МОНСАНТО ТЕКНОЛОДЖИ ЛЛС, US (56) TERRY C F ET AL: "EVENT-SPECIFIC DETECTION AND ROUNDUP READY SOYA USING TWO DIFFERENT REAL TIME PCR DETECTION CHEMISTRIES" ZEITSCHRIFT FUER LEBENSMITTEL-UNTERSUCHUNG UND FORSCHUNG. A, EUROPEAN FOOD RESEARCH AND TECHNOLOGY, SPRINGER, HEIDELBERG, DE, vol. 213, no. 6, November 2001 (2001-11), pages 425-431 WO A 0244407, 06.06.2002 WO A 2004072235, 26.08.2004 WINDELS P ET AL: "CHARACTERISATION OF THE ROUNDUP READY SOYBEAN INSERT" ZEITSCHRIFT FUER LEBENSMITTELUNTERSUCHUNG UND -FORSCHUNG. A, EUROPEAN FOOD RESEARCH AND TECHNOLOGY, SPRINGER, HEIDELBERG, DE, vol. 213, no. 2, August 2001 (2001-08), pages 107-112 US A 2003199681, 23.10.2003 HERNANDEZ M ET AL: "Development of melting temperature-based SYBR Green I polymerase chain reaction methods for multiplex genetically modified organism detection" ANALYTICAL BIOCHEMISTRY, ACADEMIC PRESS, SAN DIEGO, CA, US, vol. 323, no. 2, 15 December 2003 , Abstract ROTT M E ET AL: "Detection and quantification of roundup ready soy in foods by conventional and realtime polymerase chain reaction" JOURNAL OF AGRICULTURAL AND FOOD CHEMISTRY, AMERICAN CHEMICAL SOCIETY. WASHINGTON, US, vol. 52, no. 16, 20 July 2004 (2004-07-20), Abstract US A 6462258, 08.10.2002 2 (19) 1 3 94582 4 отримання діагностичного амплікону для випадку MON89788. 12. Набір для виявлення ДНК, специфічний для генної модифікації MON89788, що включає щонайменше нуклеїнову кислоту, яка включає 11 або більше суміжних нуклеотидів гомологічних або комплементарних до SEQ ID NO:3 або SEQ ID NO:4. 13. Спосіб отримання стійкої до гербіциду гліфосату рослини сої, що включає введення в геном вказаної рослини генної модифікації MON89788. 14. Спосіб за п. 13, що включає стадії: (a) схрещування першої рослини сої, що включає генну модифікацію MON89788, з другою рослиною сої, яка не включає генну модифікацію MON89788, для отримання рослин потомства; і (b) селекція щонайменше першої рослини потомства, яка включає вказану генну модифікацію MON89788 і є стійкою до гліфосату. 15. Спосіб за п. 14, що додатково включає самозапилення вказаної першої рослини потомства, для отримання другого покоління рослин потомства, і селекцію щонайменше першої рослини, гомозиготної для вказаної генної модифікації MON89788. 16. Спосіб визначення присутності ДНК, що відповідає генній модифікації сої MON89788 в зразку, де спосіб включає: (a) контактування зразка, що включає ДНК сої, з набором праймерів, які при використанні в реакції ампліфікації нуклеїнових кислот з геномною ДНК з сої з генною модифікацією MON89788 виробляють діагностичний амплікон для генної модифікації MON89788 сої; і (b) проведення реакції ампліфікації нуклеїнових кислот, отримуючи таким чином діагностичний амплікон; і (с) визначення діагностичного амплікону. 17. Спосіб визначення присутності нуклеїнової кислоти, що відповідає генній модифікації MON89788, в зразку, де спосіб включає: (a) отримання зразка ДНК сої; і (b) дослідження зразка на присутність послідовності ДНК генної модифікації MON89788. 18. Спосіб за п. 17, в якому дослідження зразка ДНК включає визначення присутності послідовності нуклеїнової кислоти щонайменше однієї з SEQ ID NO:1, SEQ ID NO:2 або їх комплементів. 19. Рослина сої, що має стійкість до гліфосату, в якій вказана стійкість генетично пов'язана з молекулою нуклеїнової кислоти, що включає SEQ ID NO:1, 2, 3 або 4. 20. Спосіб отримання товарного продукту з сої, що включає: (a) отримання рослини сої або її частини за п. 2; і (b) виробництво товарного продукту сої з рослини сої або її частини. 21. Спосіб за п. 20, в якому товарний продукт з сої являє собою крупу, борошно, пластівці, виділений білок або олію. 22. Спосіб контролю зростання бур'янів на полі з рослинами сої, що включають генну модифікацію MON89788, де спосіб включає обробку поля кількістю гліфосату, ефективного для контролю росту бур'янів, в якому рослини сої виявляють стійкість до гліфосату. 23. Спосіб за п. 22, в якому обробка поля здійснюється на стадії росту від V1 до R4. Передумови винаходу Ця заявка претендує на пріоритет попередньої заявки США № 60/685,584, зареєстрованої 27 травня, 2005, зміст якої включений в даний опис за допомогою посилання. 1. Галузь винаходу Даний винахід стосується нового і відмітного зразка трансгенної трансформації сої, позначеного як генна модифікація MON89788, отриманого з нього культурного сорту сої, частин рослин і продуктів. До винаходу також відносяться способи визначення присутності молекули ДНК, специфічної для MON89788, в екстракті частин рослини або екстракті насіння. 2. Опис пов'язаної галузі Соя (Glycine max) являє собою важливу сільськогосподарську культуру в багатьох областях світу. Для вдосконалення агрономічних властивостей і якості продукту до сої були застосовані способи біотехнології. Одна така агрономічна властивість, важлива в отриманні сої, являє собою стійкість до гербіцидів, зокрема, стійкість до гербіциду гліфосату. Лінія сої, стійка до гербіцидів, була б корисною властивістю при боротьбі з бур'янами. N-фосфонометилгліцин, також відомий як гліфосат, являє собою добре відомий гербіцид, який має активність до широкого спектра видів рослин. Гліфосат являє собою активний інгредієнт Roundup® (Monsanto Co., St. Louis, MO), безпечний гербіцид, який має придатний короткий період напівжиття в навколишньому середовищі. При нанесенні на поверхню рослини, гліфосат системно переміщується по рослині. Гліфосат є фітотоксичним внаслідок його пригнічення шляхом шкімової кислоти, який забезпечує прекурсор в синтезі ароматичних амінокислот. Гліфосат пригнічує фермент 5-енолпірувил-3-фосфошикіматсинтазу (EPSPS), виявлену в рослинах. Стійкість до гліфосату може досягатися експресією варіантів EPSPS, які мають більш низьку спорідненість до гліфосату і тому зберігають свою каталітичну активність в присутності гліфосату (патенти США № 5633435; 5094945; 4535060 і 6040497). Ферменти, які руйнують гліфосат в тканинах рослин (патент США № 5463175), також здатні додавати клітинну стійкість до гліфосату. Такі гени використовують для отримання трансгенних сільськогосподарських культур, які стійкі до гліфосату, тим самим, дозволяючи використовувати гліфосат для ефективної боротьби з бур'янами з мінімальним побоюванням пошкодження сільськогосподарської культури. Наприклад, стійкість до гліфосату була генетично сконструйована в кукурудзі (патент США № 5554798), пшениці (патент США 6689880), бавовні (патент США 6740488), сої (WO 9200377) і рапсі (патентна заявка США 5 20040018518). Трансгени стійкості до гліфосату, і трансгени стійкості до інших гербіцидів, наприклад, ген bar, (Toki et al., 1992; Thompson et al., 1987; фосфінотрицин ацетилтрансфераза (DeBlock et al., 1987), для стійкості до гербіциду глюфозинату) також корисні як селектовані маркери або оцінюючі маркери, і можуть забезпечити корисний фенотип для відбору рослин, пов'язаних з іншими агрономічно корисними властивостями. Відомо, що експресія чужорідних генів в рослинах перебуває під впливом їх хромосомного положення, можливо внаслідок структури хроматину (наприклад, гетерохроматин) або близькості елементів регуляції транскрипції (наприклад, енхансери) поблизу сайта інтеграції (Weising et al., 1988). З цієї причини, це часто є необхідним для скринінга великої кількості випадків для того, щоб ідентифікувати випадок, який характеризується оптимальною експресією гена, який представляє інтерес. Наприклад, було виявлено, що в рослинах і в інших організмах може існувати значна різниця в рівнях експресії вбудованого гена серед оцінюваних випадків. Також можуть існувати відмінності в просторових і часових структурах експресії, наприклад, відмінності у відносній експресії трансгена в різних тканинах рослин, які не можуть відповідати структурам, очікуваним з елементів регуляції транскрипції, представлених у вбудованому генному конструкті. З цієї причини, прийнято отримувати від сотень до тисяч різних генетично модифікованих зразків і проводити скринінг таких зразків до одиничного варіанту, який має бажані в комерційних цілях рівні експресії трансгенів і структури. Зразок, який має бажані рівні і структури експресії трансгенів, є корисним для інтрогрессії трансгена всередину інших генетичних фонів шляхом випадкового статевого схрещування з використанням традиційних способів селекції. Потомство таких схрещувань підтримує характеристики експресії трансгенів вихідного трансформанту. Ця стратегія використовується, щоб забезпечити надійну експресію генів в ряді різновидів, які добре пристосовуються до місцевих умов зростання. Було б вигідно вміти визначати присутність конкретного зразка для того, щоб визначити, чи містить потомство статевого схрещування цікавлячий трансген. Крім того, спосіб визначення конкретного зразка був би корисний для відповідності нормам, які вимагають передринкового дозволу і, зокрема, для маркування харчових продуктів, отриманих з рослин сільськогосподарських культур. Можливо визначити присутність трансгена будь-яким добре відомим способом виявлення полінуклеїнових кислот, таким як полімеразна ланцюгова реакція (PCR) або гібридизація ДНК з використанням полінуклеїнових кислот як зондів. Ці способи визначення зазвичай фокусуються на часто вживаних генетичних елементах, таких як промотори, термінатори, маркерні гени і так далі. У результаті, такі способи не можуть бути корисними для проведення відмінностей між різними генетично модифікованими зразками (лініями), зокрема такими, які отримані з використанням того ж самого ДНК конструкта за винятком послідовності хромосомної ДНК ("фланкуюча ДНК"), розташованої 94582 6 поруч з включеною трансгенною ДНК. Специфічний PCR аналіз для ліній обговорюється, наприклад, Windels et al. (1999), який ідентифікував лінію стійких до гліфосату сої 40-3-2 шляхом PCR з використанням набору праймерів, які перекривають ділянку з'єднання між вставленим трансгеном і фланкуючою ДНК, зокрема, використовуючи один праймер, який включав в себе послідовність з вставки, і другого праймера, який включав в себе послідовність з фланкуючою ДНК. Способи виявлення специфічної ДНК генетично модифікованої трансгенної рослини також були описані в патентах США № 6893826; 6825400; 6740488; 6733974 і 6689880; 6900014 і 6818807, зміст кожної з яких включений повністю в даний опис за допомогою посилання. Цей винахід стосується генетично модифікованої гліфосат-стійкої сої ΜΟΝ89788 (також названої як ΜΟΝ19788 або GM_A19788) і молекул ДНК, що містяться в цих рослинах сої, які є корисними в способах визначення рослини і її потомства, і тканин рослини, отриманих з MON89788. Суть винаходу Даний винахід стосується трансгенної генетично модифікованої сої, позначеної як MON89788 (також названої як MON19788), і її потомства, що має показове насіння, депоноване в American Type Culture Collection (ATCC) під номером РТА-6708. Іншим аспектом винаходу є клітини рослин або регенеровані частини рослини або насіння сої з генною модифікацією MON89788. Також винахід включає в себе частини рослин сої з генною модифікацією MON89788, які включають, але не обмежуються перерахованим, клітини, пилок, насінний зачаток, квіти, пагони, коріння, листя і продукти, отримані з MON89788, наприклад соєві крупи, борошно і олію. Один аспект винаходу стосується композицій і способів для визначення присутності трансгенної/геномної з'єднаної ділянки ДНК рослини або насіння або продуктів, отриманих з частин рослин або насіння сої з генною модифікацією MON89788. Представлені молекули ДНК, які включають щонайменше одну молекулу ДНК з трансгенною/геномною ділянкою сполуки, вибрану з групи, що складається з SEQ ID NO:1 і SEQ ID NO:2, і їх комплементи, де сполучена молекула перекриває вставлений сайт, який включає гетерологічну ДНК, вставлену в геном клітини сої, і геномну ДНК з клітини сої, яка фланкує вставлений сайт сої з генною модифікацією MON89788. Такі сполучені послідовності можуть, в одному аспекті винаходу, бути визначені як такі, що включають нуклеотиди 1093-1113 або 5396-5416 з SEQ ID NO:9, відповідно. В інших аспектах винаходу, сполуки можуть бути визначені як такі, що включають додаткові ділянки фланкуючого геному або трансгена, наприклад, і можуть бути визначені як такі, що включають одну або більше послідовностей, представлену нуклеотидами 1073-1113, 1043-1113, 10931133, 1093-1163, 1043-1163, 5376-5416, 5346-5416, 5396-5436, 5396-5416, 5396-5466 або 5346-5466 з SEQ ID NO:9. Тому такі послідовності і рослини, і насіння, що включає ці послідовності, є одним з аспектів винаходу. 7 Представлена нова молекула ДНК, яка являє собою трансгенну/геномну ділянку ДНК SEQ ID NO:3 або її комплемент, з сої з генною модифікацією MON89788. Рослина сої і насіння, яке включає SEQ ID NO:3 в своєму геномі, являють собою один з аспектів даного винаходу. SEQ ID NO:3 додатково включає SEQ ID NO:1 повністю. Згідно з іншим аспектом винаходу, представлена молекула ДНК, яка являє собою трансгенну/геномну ділянку ДНК SEQ ID NO:4, або її комплемент, в якому ця молекула ДНК є новою для сої з генною модифікацією MON89788. Рослина сої і насіння, яке включає SEQ ID NO:4 в своєму геномі, являє собою один з аспектів даного винаходу. SEQ ID NO:4 додатково включає SEQ ID NO:2 повністю. Згідно з іншим аспектом винаходу, представлені дві молекули нуклеїнових кислот для використання в способі виявлення ДНК, в якому перша молекула нуклеїнової кислоти включає щонайменше 11 або більше суміжних полінуклеотидів будьякої області трансгенної ділянки молекули ДНК з SEQ ID NO:3, і друга нуклеїнова кислота являє собою молекулу схожої довжини будь-якої області 5' фланкуючої ділянки геномної ДНК сої з SEQ ID NO:3, в якій ці молекули нуклеїнових кислот при спільному використанні є корисними як праймери в способі ампліфікації ДНК, в якому проводять амплікон. Амплікон, отриманий з використанням цих праймерів в способі ампліфікації ДНК, є діагностичним для ДНК генної модифікації MON89788. Амплікон, отриманий з описаними праймерами, які гомологічні або комплементарні ділянці з SEQ ID NO:3, що включає SEQ ID NO: 1, є одним з аспектів винаходу. Згідно з іншим аспектом винаходу, дві молекули нуклеїнових кислот надані для використання в способі виявлення ДНК, в якому перша молекула нуклеїнової кислоти включає щонайменше 11 або більше суміжних полінуклеотидів будь-якої області трансгенної ділянки молекули ДНК з SEQ ID NO:4, і друга молекула нуклеїнової кислоти аналогічної довжини будь-якої ділянки 3' фланкуючої геномної ДНК сої з SEQ ID NO:4, в якій ці молекули нуклеїнових кислот при спільному використанні є корисними як праймери в способі ампліфікації ДНК, в якому проводять амплікон. Амплікон, отриманий з використанням цих праймерів в способі ампліфікації ДНК, є діагностичним для ДНК генної модифікації MON89788. Амплікон, отриманий з описаними праймерами, які гомологічні або комплементарні ділянці з SEQ ID NO:4, що включає SEQ ID NO:2, є одним з аспектів винаходу. Будь-яка пара праймерів нуклеїнових кислот, отримана з SEQ ID NO:3 або SEQ ID NO:4, або SEQ ID NO:9 або їх комлементів, які при використанні в реакції ампліфікації ДНК, проводять діагностичний амплікон для тканини з сої з генною модифікацією MON89788, такою як амплікон, який включає SEQ ID NO:1 або SEQ ID NO:2 або будьяка ділянка з SEQ ID NO:9, відповідно, являє собою інший варіант здійснення винаходу. У конкретному варіанті здійснення, пара праймерів може містити праймер A (SEQ ID NO:5) і праймер D (SEQ ID NO:8). 94582 8 Інший аспект винаходу представляє рослину сої, або насіння, або продукт, отриманий з рослини або насіння, що включає генну модифікацію MON89788, в якої геномна ДНК при виділенні з рослини сої, або насіння, або продуктів, проводить амплікон в способі ампліфікації ДНК, який включає SEQ ID NO:1 або SEQ ID NO:2. Ще один аспект винаходу представляє рослину сої, або насіння, або продукт, отриманий з рослини або насіння, що включає MON89788, в яких геномна ДНК при виділенні з рослини сої, або насіння, або продуктів, проводить амплікон в способі ампліфікації ДНК, де в способі ампліфікації ДНК використовуються молекули ДНК праймерів SEQ ID NO:5 і SEQ ID NO:6. Ще один аспект винаходу являє собою рослину сої, насіння, продукт або товарний продукт (продукт споживання), отримані з рослини або насіння, що включає генну модифікацію MON89788, в якій геномна ДНК при виділенні з рослини сої, або насіння, або продуктів, проводить амплікон в способі ампліфікації ДНК, де в способі ампліфікації ДНК використовуються молекули ДНК праймерів SEQ ID NO:7 і SEQ ID NO:8. Продукт або товарний продукт може включати в себе, без обмеження, харчовий продукт або продукт харчування, що включає або отриманий з одного або більше наступних продуктів рослин сої, які включають генну модифікацію MON89788: лецитин, жирні кислоти, гліцерин, стерол, харчове масло, знежирені соєві пластівці, соєві крупи, що включає в себе знежирені і висушені соєві крупи, соєве молоко, яке згорнулося, соєвий сир, соєве борошно, концентрат соєвого білка, виділений соєвий білок, розчинений у воді рослинний білок, текстурат сої і соєве білкове волокно. Згідно з іншим аспектом винаходу, представлений спосіб визначення в зразку присутності ДНК, відповідної, зокрема, ДНК сої з генною модифікацією MON89788. Такий спосіб включає: (а) контактування зразка, що включає ДНК, з парою ДНК праймерів; (b) здійснення реакції ампліфікації нуклеїнової кислоти, яка виробляє при цьому амплікон; і (с) визначення амплікону, в якому вищевказаний амплікон включає в себе SEQ ID NO:1 або SEQ ID NO:2. Набір, що включає молекули ДНК праймерів, які при використанні в способі ампліфікації ДНК проводять амплікон, що включає SEQ ID NO:1 або SEQ ID NO:2, являє собою додатковий аспект винаходу. Згідно з іншим аспектом винаходу, представлений спосіб визначення в зразку присутності ДНК, відповідної, зокрема, ДНК сої з генною модифікацією MON89788. Такий спосіб включає: (а) контактування зразка, що включає ДНК із зондом, яка гібридизується в жорстких умовах гібридизації з геномною ДНК з сої з генною модифікацією MON89788 і не гібридизується в жорстких умовах гібридизації з ДНК контрольної рослини сої; (b) піддавання зразка і зонда жорстким умовам гібридизації; і (с) визначення гібридизації зонда з ДНК сої з генною модифікацією MON89788, в якому вказаний зонд включає SEQ ID NO:1 або SEQ ID NO:2. Зразок може включати в себе насіння, рослини потомства, або частину рослини, що вклю 9 чають генну модифікацію сої MON89788, або будьякі з наступних продуктів, отриманих з рослини, які включають MON89788: лецитин, жирні кислоти, гліцерин, стерол, харчове масло, знежирені соєві пластівці, соєві крупи, які включають в себе знежирені і висушені соєві крупи, соєве молоко, що згорнулося, соєвий сир, соєве борошно, концентрат соєвого білка, виділений соєвий білок, розчинений у воді рослинний білок, текстурат сої і соєве білкове волокно. Набір, який включає ДНК зонд, що містить молекулу ДНК, яка гомологічна або комплементарна до SEQ ID NO:1 або SEQ ID NO:2, являє собою один з аспектів винаходу. Набір, який містить молекулу ДНК, що включає SEQ ID NO:18, SEQ ID NO:19, або SEQ ID NO:20, або їх комплемента, також являють собою один з аспектів винаходу. Згідно з іншим аспектом винаходу, представлений спосіб отримання рослини сої, яка стійка до застосування гліфосату, що включає в себе стадії: (а) статеве схрещування першої батьківської стійкої до гліфосату рослини сої, що включає генну модифікацію MON89788, і другої батьківської рослини сої, які не мають стійкості до гліфосату, отримуючи при цьому множину рослин потомства; і (b) виділення рослини потомства, яка стійка до застосування гліфосату. Способи селекції можуть додатково включати в себе стадії схрещування батьківської рослини, що включає генну модифікацію сої MON89788, з другою батьківською рослиною сої, яка також стійка до гліфосату, і виділення стійкого до гліфосату потомства за допомогою молекулярної маркерної ДНК, генетично зв'язаної з стійким до гліфосату фенотипом, виявленим у кожного батька. Інший аспект винаходу являє собою спосіб боротьби з бур'янами на полі рослин сої, що включають генну модифікацію MON89788, в якому вказаний спосіб включає засівання поля насінням сої, що містить генну модифікацію MON89788, де вказане показове насіння депоноване в АТСС під інвентарним № РТА-6708, забезпечення росту вказаного насіння і обробку вказаних рослин ефективною дозою гліфосату для боротьби з бур'янами на вказаних полях. Викладені вище й інші аспекти винаходу стануть більш зрозумілі з наступного докладного опису і прикладених фігур. Короткий опис фігур Фіг. 1. Організація трансгенної вставки в геном рослини сої, що включає генну модифікацію MON89788. Фіг. 2А-2В. Обробка товарних продуктів з сої. Докладний опис Даний винахід стосується нової трансформованої сої з генетичною модифікацією, позначеної як MON89788, яка забезпечує стійкість до гліфосату, і частин рослин, і насіння, і продуктів, вироблених з рослин, частин рослин, насіння, і продуктів, які включають вказану генетичну модифікацію. Винахід представляє молекули ДНК, які є новими в геномі клітин сої, що включає генетичну модифікацію MON89788 і молекули ДНК, які можуть бути використані в різних способах виявлення ДНК для ідентифікації ДНК MON89788 в зразку. Винахід 94582 10 представляє спосіб боротьби з бур'янами в полі рослин, що містять MON89788, шляхом обробки бур'янів в полі, яке містить рослини, що містять генетичну модифікацію MON89788, гербіцидом гліфосатом. Наступні визначення і способи представлені для кращого опису даного винаходу і для інструктування фахівців в даній галузі в практиці даного винаходу. Якщо не вказано особливо, терміни представлені, щоб бути зрозумілими фахівцям, компетентним в даній галузі. Визначення загальних термінів в молекулярній біології також можна знайти в Rieger et al. (1991) і Lewin (1994). Використовується номенклатура для основ ДНК, як представлено в з 1.822 37 CFR. Для використання в даному описі, термін "соя" означає Glycine max і включає в себе всі модифікації рослини, які можуть бути виведені з сої. Для використання в даному описі, термін "який включає" означає "такий, що включає, але не обмежується". "Гліфосат" стосується N-фосфонометилгліцину і його солей. Гліфосат являє собою активний компонент Roundup® гербіциду (Monsanto Co.). Обробка "гербіцидом гліфосатом" стосується обробок з використанням гербіцидів Roundup®, Roundup Ultra®, Roundup Pro® або будь-яких інших гербіцидних сполук, що містять гліфосат. Приклади комерційних препаратів гліфосату включають, без обмежень, препарати, які продаються Monsanto Company як гербіциди ROUNDUP®, ROUNDUP® ULTRA, ROUNDUP® ULTRAMAX, ROUNDUP® CT, ROUNDUP® EXTRA, ROUNDUP® BIACTIVE, ROUNDUP® BIOFORCE, RODEO®, POLARIS®, SPARK® і ACCORD®, всі з яких містять гліфосат у вигляді його солі ізопропіламонію; ROUNDUP® WEATHERMAX (калієва сіль гліфосату), препарати, що продаються Monsanto Company як гербіциди ROUNDUP® DRY і RIVAL®, які містять гліфосат у вигляді його амонійної солі; препарати, що продаються Monsanto Company як ROUNDUP® GEOFORCE, які містять гліфосат у вигляді його натрієвої солі; і препарати, що продаються Syngenta Crop Protection як гербіцид TOUCHDOWN®, який містить гліфосат у вигляді його триметилсульфонієвої солі. Обробка поля, що містить рослини сої, стійкі до гліфосату, які включають лінію MON89788, будь-якою з цих сполук гербіциду гліфосату буде контролювати ріст бур'янів на полі і не буде впливати на ріст або врожайність рослин сої, що включають генетичну модифікацію MON89788. Трансгенну "генетичну модифікацію" отримують трансформацією клітин рослини гетерологічною ДНК, наприклад, нуклеїновокислотним конструктом, який включає в себе цікавлячий трансген, відновленням популяції рослин, отриманих внаслідок вставки трансгена в геном рослини, і селекцією окремої рослини, що характеризується вставкою в окрему ділянку геному. Термін "генетично модифікований" стосується початкової трансформації і потомства трансформанту, які включають в себе гетерологічну ДНК. Термін "генетично модифікований" також стосується потомства, отриманого шляхом статевого схрещування між трансфо 11 рмантом і іншою модифікацією, який включає в себе гетерологічну трансгенну ДНК і фланкуючу геномну ДНК. Термін "генетично модифікований" також стосується ДНК початкового трансформанту, що включає вставлену ДНК і фланкуючу геномну послідовність, яка безпосередньо примикає до вставленої ДНК, для якої можна було б чекати її переміщення в потомство, яке отримує вставлену ДНК, що включає в себе цікавлячий трансген як результат статевого схрещування однієї батьківської клітинної лінії, яка включає в себе вставлену ДНК (наприклад, вихідний трансформант і потомство, отримане в результаті самозапилення), і батьківської лінії, яка не містить вставлену ДНК. Рослина сої, стійка до гліфосату, може бути вирощена першим статевим схрещуванням першої батьківської рослини сої, що складається з рослини сої, вирощеної з трансгенної гліфосатстійкої рослини сої, що включає MON89788, або рослину сої, яка є потомством схрещування такої рослини, яка експресує гліфосат-стійкий фенотип, і другої батьківської рослини сої, яка не має стійкості до гліфосату, отримуючи, таким чином, множину рослин першого потомства; і потім відбираючи рослину потомства, яка стійка до застосування гербіциду гліфосату. Ці стадії додатково можуть включати зворотне схрещування гліфосат-стійкої рослини потомства з другою батьківською рослиною сої або третьою батьківською рослиною сої з подальшим відбором потомства шляхом застосування гліфосату або ідентифікацією молекулярних маркерів, пов'язаних з властивістю, отримуючи, таким чином, рослину сої, яка стійка до застосування гербіциду гліфосату. Можна використовувати молекулярні маркери, які включають сполучені молекули ДНК, ідентифіковані в 5' і 3' ділянках вставки трансгена в генетичну модифікацію MON89788. Також зрозуміло, що дві різні трансгенні рослини можуть також бути схрещені з отриманням потомства, яке містить два незалежно виділених екзогенних трансгени. Також розглянуте зворотне схрещування з батьківською рослиною і перехресне схрещування з нетрансгенною рослиною, як описано раніше для вегетативного розмноження. Описи інших способів селекції, які поширені для різних властивостей і сільськогосподарських культур, можна виявити в одному з декількох посилань, наприклад, Fehr, (1987). "Зонд" - це виділена нуклеїнова кислота, до якої прикріпляють звичайну детектовану мітку або репортерну молекулу, наприклад, радіоактивний ізотоп, ліганд, хемілюмінесцентний агент або фермент. Такий зонд комплементарний до ланцюга мішенної нуклеїнової кислоти, і у випадку даного винаходу - до ланцюга геномної ДНК рослини сої, що включає генетичну модифікацію MON89788, в будь-якій з рослини сої або насіння, або із зразка або екстракту рослини або насіння, яке включає вказану генетичну модифікацію ДНК. Зонди згідно з даним винаходом включають в себе не тільки дезоксирибонуклеїнову або рибонуклеїнову кислоти, але також поліаміди і інші матеріали зондів, які зв'язуються специфічно з мішенною послідовністю 94582 12 ДНК і можуть використовуватися для виявлення присутності цієї мішенної послідовності ДНК. "Праймери" являють собою виділені полінуклеїнові кислоти, які випалюються в комплементарні ланцюги мішенної полінуклеїнової кислоти шляхом гібридизації нуклеїнової кислоти з утворенням гібриду між праймером і мішенним ланцюгом полінуклеїнової кислоти, з подальшим подовженням вздовж мішенного ланцюга полінуклеїнової кислоти за допомогою полімерази, наприклад, ДНК полімерази. Пари праймерів даного винаходу пов'язані з використанням їх для ампліфікації молекули мішенної полінуклеїнової кислоти, наприклад, полімеразною ланцюговою реакцією (PCR) або іншими звичайними способами ампліфікації нуклеїнових кислот. Зонди і праймери зазвичай мають 11 полінуклеотидів або більше в довжину, переважно 18 полінуклеотидів або більше, більш переважно 24 полінуклеотидів або 30 полінуклеотидів або більше. Такі зонди і праймери гібридизуються специфічно з мішенною молекулою при гібридизації в умовах високої жорсткості. Переважно, зонди і праймери згідно з даним винаходом мають повну ідентичність послідовностей з мішенною молекулою, хоча зонди, які відрізняють від послідовності мішені і які зберігають здатність до гібридизації до послідовності мішені в умовах високої жорсткості, можуть бути сконструйовані стандартними способами. Способи отримання і використання зондів і праймерів описані, наприклад, в Sambrook et al. (1989); Ausubel et al. (1992); і Innis et al. (1990). Пари PCR-праймерів (набір праймерів) можуть бути виведені з відомої послідовності, наприклад, використовуючи комп'ютерні програми, спеціально призначені для цього, такі як Primer (Version 0.5, й 1991, Whitehead Institute for Biomedical Research, Cambridge, ΜΑ). Праймери і зонди, засновані на фланкуючій геномній ДНК і послідовностях вставки, розкриті в даному описі (SEQ ID NO:1-4 і 9), можуть використовуватися для підтвердження і, якщо необхідно, для виправлення розкритих послідовностей стандартними способами, наприклад, шляхом виділення відповідної молекули ДНК з депонованого насіння, що включає MON89788, і для визначення послідовності нуклеїнових кислот таких молекул. Додатково зв'язані молекули ДНК можна виділити з геному клітини, що включає MON89788, який включає трансгенну вставку і геномну фланкуючу ділянку, і фрагменти цих молекул можна використовувати як праймери або зонди. Зонди і праймери нуклеїнових кислот даного винаходу гібридизуються при жорстких умовах з мішенною послідовністю ДНК. Будь-який звичайний спосіб гібридизації або ампліфікації нуклеїнових кислот можна використовувати для ідентифікації присутності ДНК з лінії MON89788 в зразку. Молекули нуклеїнових кислот або їх фрагменти піддаються специфічній гібридизації з іншими молекулами нуклеїнових кислот при деяких обставинах. Як використано в даному описі, вважається, що дві молекули нуклеїнових кислот піддаються специфічній гібридизації одна з одною, якщо дві 13 молекули здатні до утворення антипаралельної, дволанцюгової структури нуклеїнових кислот і є досить довгими для збереження цієї структури в умовах високої жорсткості. Вважається, що молекула нуклеїнової кислоти є "комплементарною" іншій молекулі нуклеїнової кислоти, якщо вони виявляють повну комплементарність. Як використано в даному описі, вважається, що молекули виявляють "повну комплементарність", коли кожний нуклеотид однієї з молекул комплементарний нуклеотиду іншої. Вважається, що дві молекули є "мінімально комплементарними", якщо вони можуть гібридизуватися одна з одною досить стабільно, щоб дозволити їм залишитися відпаленими одна з одною при щонайменше звичайних умовах "низької жорсткості". Аналогічно, вважається, що молекули є "комплементарними", якщо вони можуть гібридизуватися одна з одною з достатньою стабільністю, щоб дозволити їм залишитися відпаленими одна з одною при звичайних умовах "високої жорсткості". Звичайні жорсткі умови описані Sambrook et al., 1989, і Haymes et al. (1985). Таким чином відступи від повної комплементарності дозволені, оскільки такі відступи ніколи не запобігають повністю здатності молекул утворювати дволанцюгову структуру. Щоб молекула нуклеїнових кислот виконувала функцію праймера або зонда, необхідна тільки достатня комплементарність послідовності, щоб бути здатною до утворення стабільної дволанцюгової структури у використаних конкретному розчиннику і концентраціях солі. Як використано в даному описі, значно гомологічна послідовність являє собою послідовністю нуклеїнових кислот, яка специфічно гібридизується з комплементарною послідовністю нуклеїнових кислот, з якою вона порівняна в умовах високої жорсткості. Придатні жорсткі умови, які сприяють гібридизації ДНК, наприклад, 6,0 X хлорид натрію/цитрат натрію (SSC) при приблизно 45°С, з подальшим відмиванням 2,0 SSC при 50°С, відомі фахівцеві в даній галузі або можуть бути знайдені в Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989), 6.3.1-6.3.6. Наприклад, концентрацію солі на стадії відмивання можна вибрати від низької жорсткості, приблизно 2,0 SSC при 50°С, до високої жорсткості, приблизно 0,2 SSC при 50°С. Крім того, температуру на стадії відмивання можна збільшити від умов низької жорсткості при кімнатній температурі, приблизно 22°С, до умов високої жорсткості при приблизно 65°С. Можна змінювати як температуру, так і сіль, або можна залишати постійними або температуру, або концентрацію солі, тоді як параметр, що залишився, зміниться. У переважному варіанті здійснення, нуклеїнова кислота даного винаходу специфічно гібридизується з однією або більше молекулами нуклеїнових кислот, представленими в SEQ ID NO:1-4 і 9, їх комплементами або фрагментами, при помірно жорстких умовах, наприклад, при приблизно 2,0 SSC і приблизно 65°С. Зокрема, в переважному варіанті здійснення, нуклеїнова кислота даного винаходу специфічно гібридизувалася з однією або більше молекулами нуклеїнових кислот, представленими в SEQ ID NO:1-4 і 9, їх комплементах або фрагментах, при 94582 14 умовах високої жорсткості. В одному аспекті даного винаходу, переважна маркерна молекула нуклеїнової кислоти даного винаходу включає послідовність нуклеїнової кислоти, як представлена в SEQ ID NO:1 або SEQ ID NO:2 або їх комплементах або фрагментах. В іншому аспекті даного винаходу, переважна маркерна молекула нуклеїнової кислоти даного винаходу має від 80% до 100% або від 90% до 100% ідентичності послідовності з послідовністю нуклеїнових кислот, представлених в SEQ ID NO:1 або SEQ ID NO:2 або їх комплементах або фрагментах. Молекулярні маркерні молекули ДНК, які включають SEQ ID NO:1 або SEQ ID NO:2, або їх комплементи або фрагменти, можна використовувати як маркери в способах селекції рослин для ідентифікації потомства генетичного схрещування, схожого зі способами, описаними для простого маркерного аналізу повторів послідовності ДНК, в Cregan et al. (1997); кожне з яких повністю включене в даний опис за допомогою посилання. Гібридизація зонда з мішенною молекулою ДНК може бути визначена будь-якими способами, відомими фахівцям в даній галузі, і вони можуть включати, але ними не обмежуватися, флуоресцентні мітки, радіоактивні мітки, мітки, засновані на антитілах, і хемілюмінісцентні мітки. Розглядаючи ампліфікацію мішенної послідовності нуклеїнових кислот (наприклад, PCR) з використанням специфічної ампліфікаційної пари праймерів, "жорсткі умови" являють собою умови, які дозволяють парі праймерів гібридизуватися тільки з мішенною послідовністю нуклеїнових кислот, з якою праймер, що має відповідну послідовність дикого типу (або її комплемент), може зв'язуватися і переважно виробляти унікальний продукт ампліфікації, амплікон, в термальній реакції ампліфікації ДНК. Термін "специфічний для (мішенної послідовності)" показує, що зонд або праймер гібридизується в суворих умовах гібридизації тільки з мішенною послідовністю в зразку, що включає мішенну послідовність. Як використано в даному описі, "ампліфікована ДНК" або "амплікон" стосуються продукту ампліфікації нуклеїнових кислот мішенної послідовності нуклеїнових кислот, яка є частиною матриці нуклеїнових кислот. Наприклад, для визначення, чи містить рослина сої, отримана статевим схрещуванням, трансген з генетичною модифікацією ΜΟΝ89788, або чи включає зразок сої, зібраний з поля, ΜΟΝ89788, або екстракт сої, такий як крупа, борошно або олія, чи включає ΜΟΝ89788. ДНК, виділена із зразка тканини рослини сої або екстракту, може бути піддана способу ампліфікації нуклеїнових кислот з використанням пари праймерів, які включають в себе праймер, отриманий з прилеглої геномної ділянки інсерційного сайта вставленої гетерологічної трансгенної ДНК, і другого праймера, отриманого з вставленої гетерологічної трансгенної ДНК для виробництва амплікону, який є діагностичним для випадку присутності ДНК. Амплікон має довжину і має послідовність, яка також є діагностичною для випадку. Амплікон може змінювати довжину від загальної довжини пари праймерів, плюс одна нуклеотидна пара основи, або плюс 15 приблизно п'ятдесят пар нуклеотидних основ, або плюс приблизно двісті п'ятдесят пар нуклеотидних основ, або плюс приблизно триста п'ятдесят пар нуклеотидних основ або більше. Альтернативно, пару праймерів можна отримати з фланкуючої геномної послідовності на обох боках вставленої ДНК, з тим, щоб отримати амплікон, який включає в себе повну вставлену нуклеотидну послідовність. Член пари праймерів, отриманої з геномної послідовності рослини, може розташовуватися на відстані від вставленої трансгенної молекули ДНК, ця відстань може змінюватися від однієї нуклеотидної пари основи вище до приблизно двадцяти тисяч пар нуклеотидних основ. Зокрема, використання терміну "амплікон" виключає димери праймерів, які можуть утворюватися в термальній реакції ампліфікації ДНК. Ампліфікація нуклеїнових кислот може проводитися будь-яким з різних способів реакції ампліфікації нуклеїнових кислот, відомих в даній галузі, що включають в себе полімеразну ланцюгову реакцію (PCR). Множина способів ампліфікації відома в даній галузі і описана, inter alia, в патентах США № 4683195 і 4683202 і в Innis et al. (1990). Способи ампліфікації PCR були розроблені для збільшення до 22 тис. п.о. геномної ДНК і до 42 тис. п.о. ДНК бактеріофагів (Cheng et al., 1994). Ці способи, а також інші способи, відомі в даній галузі ампліфікації ДНК, можуть використовуватися в практиці даного винаходу. Послідовність гетерологічної вставки ДНК або фланкуючої послідовності з лінії сої MON89788 може бути верифікована і виправлена, якщо необхідно, шляхом ампліфікування таких молекул з геному лінії, використовуючи праймери, отримані з послідовностей, представлених в даному описі, подальшими способами стандартного секвенування ДНК, застосованих до PCR амплікону або до виділеної клонованої трансгенної/геномної ДНК. Амплікон, отриманий цими способами, можна виявити множиною способів. Один такий спосіб представляє Genetic Bit Analysis (Nikiforov et al., 1994), де олігонуклеотид ДНК призначений для перекриття як примикаючої фланкуючої геномної послідовності ДНК, так і вставленої трансгенної послідовності ДНК. Олігонуклеотид імобілізовний на стінках мікроямкового планшета. Після PCR цікавлячої ділянки (використовуючи один праймер у вставленій послідовності і один в примикаючій фланкуючій геномній послідовності) одноланцюжковий продукт PCR може гібридизуватися з імобілізованим олігонуклеотидом і служити як матриця для реакції подовження одиничної основи з використанням ДНК полімерази і міченими ddNTPs, специфічними для очікуваної наступної основи. Зчитування даних може бути флуоресцентним або заснованим на ELISA. Сигнал показує присутність вставленої/фланкуючої геномної послідовності, внаслідок успішної ампліфікації, гібридизації і одноосновного подовження. Інший спосіб являє собою спосіб піросеквенування, як описано Winge (2000). У цьому способі олігонуклеотид призначений для перекриття з'єднання геномної ДНК, яка перекривається, і ДНК вставки. Олігонуклеотид гібридизується з однола 94582 16 нцюжковим продуктом PCR з цікавлячої ділянки (один праймер у вставленій послідовності і один у фланкуючій геномній послідовності) і інкубується в присутності ДНК-полімерази, АТР, сульфурилази, люциферази, апірази, аденозин 5' фосфосульфату і люциферину. DNTPs додають окремо, і включення призводить до світлового сигналу, який вимірюється. Світловий сигнал показує присутність трансгенної вставленої/фланкуючої послідовності внаслідок успішної ампліфікації, гібридизації і одно- або багатоосновного подовження. Поляризація флуоресценції, як описано Chen et al. (1999), являє собою спосіб, який можна використовувати для визначення амплікону даного винаходу. При використанні цього способу олігонуклеотид призначається для перекриття з'єднання геномної фланкуючої і вставленої ДНК. Олігонуклеотид гібридизується з одноланцюжковим продуктом PCR з цікавлячої ділянки (один праймер у вставленої ДНК і один у фланкуючої геномної послідовності ДНК) і інкубується в присутності ДНК полімерази і флуоресцентно-міченого ddNTP. Одноосновне подовження призводить до включення ddNTP. Включення можна виміряти як зміна поляризації з використанням флуориметра. Зміна поляризації показує присутність трансгенної вставленої/фланкуючої послідовності внаслідок успішної ампліфікації, гібридизації і одноосновного подовження. Taqman® (PE Applied Biosystems, Foster City, CA) описується як спосіб визначення і кількісного вимірювання присутності послідовності ДНК і повністю зрозумілий в інструкціях, представлених виробником. Коротко, FRET олігонуклеотидний зонд призначається для перекриття з'єднання геномної фланкуючої і вставленої ДНК. FRET зонд і PCR праймери (один праймер у вставленій послідовності ДНК і один у фланкуючій геномній послідовності) беруть участь в циклі в присутності термостабільної полімерази і dNTPs. Гібридизація FRET зонда призводить до розщеплення і вивільнення флуоресцентної частинки з частинки гасіння у FRET зонді. Флуоресцентний сигнал показує присутність фланкуючої геномної/трансгенної вставленої послідовності внаслідок успішної ампліфікації і гібридизації. Molecular Beacons був описаний для використання у виявленні послідовностей, як представлено в Tyangi et al. (1996). Коротко, FRET олігонуклеотидний зонд призначається для перекриття з'єднання фланкуючої геномної і вставленої ДНК. Специфічна структура FRET зонда призводить до цієї вторинної структури, яка зберігає флуоресцентні і гасильні частинки в безпосередній близькості. FRET зонд і PCR праймери (один праймер у вставленій послідовності ДНК і один у фланкуючій геномній послідовності) беруть участь в циклі в присутності термостабільної полімерази і dNTPs. Після успішної PCR ампліфікації, гібридизація FRET зонда з мішенною послідовністю призводить до видалення в зонді вторинної структури і просторового розділення флуоресцентних і гасильних частинок. Отримують флуоресцентний сигнал. Флуоресцентний сигнал показує присутність флан 17 куючої геномної/трансгенної вставленої послідовності внаслідок успішної ампліфікації і гібридизації. Інші описані способи, такі як мікрофлюїдики (патентна публікація США 2006068398, патент США № 6544734), представляють способи і пристрої розділення і ампліфікування зразків ДНК. Оптичні барвники використовувалися для визначення і кількісного вимірювання специфічних молекул ДНК (WO/05017181). Пристрої з нанотрубок (WO/06024023), які включають електронний прилад для молекул ДНК або наногранули, які зв'язують специфічні молекули ДНК і потім можуть визначатися, є корисними для визначення молекул ДНК даного винаходу. Набори для виявлення ДНК можуть розроблятися з використанням композицій, розкритих в даному описі, і способів, описаних або відомих в даній галузі виявлення ДНК. Набори корисні для встановлення ДНК лінії сої в зразку і можуть застосовуватися в способах селекції рослин сої, що містять ДНК. Набори можуть містити ДНК праймери або зонди, які гомологічні або комплементарні SEQ ID NO: 1-4 і 9, або ДНК праймери або зонди, гомологічні або комплементарні ДНК, що міститься в трансгенних генетичних елементах ДНК, де ці ДНК послідовності можуть використовуватися в реакціях ампліфікації ДНК або як зонди в способі гібридизації ДНК. Структура ДНК з трансгенних генетичних елементів, що містяться в геномі сої і проілюстровані на фіг. 1, включає 5' геномну ділянку геному A3244 сої, фланкуючу трансгенну вставку, вставку, яка включає ділянку правої граничної області (RB) з Agrobacterium tumefaciens, химерний промотор FMV/Tsf1 і споріднені зв'язані елементи (патент США 6660911; також званий як FMV/E1F1), функціонально зв'язаний з кодуючою послідовністю хлоропластного трансгенного білка EPSPS з Arabidopsis (названу в даному описі як СТР2 або TS-AtEPSPS CTP2, патент США 5633435, функціонально зв'язаний зі стійким до глікофосату EPSPS (названий в даному описі як СР4 EPSPS або аrоА:СР4, виділений зі штаму СР4 Agrobacterium tumefaciens, і кодуючу послідовність, модифіковану для збільшеної експресії в клітинах рослин, патент США 5633435), функціонально зв'язаний з 3' термінальною ділянкою з рибулозо-1,5-бісфосфаткарбоксилази гороху (названу в даному описі як Е9 3' або T-Ps.RbcS:E9, Coruzzi et al., (1984), ділянка лівої граничної області (LB) з Agrobacterium tumefaciens, і 3' геномною областю геному A3244 сої, яка фланкує трансгенну вставку. Молекули ДНК, корисні як праймери в способах ДНК ампліфікації, можна отримувати з послідовностей генетичних елементів трансгенної вставки, що містить трансгенну модифікацію MON89788. Ці праймерні молекули можуть використовуватися як частина праймерного набору, який також включає в себе молекулу ДНК праймера, отриману з геному сої, що фланкуючу трансгенну вставку. Соя з генетичною модифікацією MON89788 була отримана трансформацією лінії сої A3244 (патент США 5659114) способом з використанням Agrobacterium, наприклад, способами, описаними в патентах США 6384301 і 7002058 94582 18 (включені в даний опис повністю за допомогою посилання). Автори даного винаходу виявили, що лінія сої, що містить геномну ділянку Т-типу MON89788 (Ттип являє собою з'єднання трансгена і зв'язаної ділянки гаплотипу геному рослини) в своєму геномі, має поліпшену врожайність в порівнянні з лінією, що містить колишню 40-3-2 геномну ділянку Ттипу. Це було показано в відтворюваних польових випробуваннях, що включають дані з врожайності, зібрані в багатьох областях США (патентна заявка США 60/685584). Наступні приклади включають в себе приклади для демонстрації певних переважних варіантів здійснення винаходу. Фахівцям в даній галузі потрібно враховувати, що способи, розкриті в нижченаведених прикладах, які представляють підходи, що добре і функціонально обгрунтовані авторами в практиці винаходу, можуть розглядатися з метою створення окремих переважних способів здійснення на практиці. Однак фахівцям в даній галузі потрібно розуміти в світлі даного розкриття, що багато змін можна проводити в розкритих специфічних варіантах здійснення і отримувати подібний або схожий результат без відхилення від суті і обсягу винаходу. Приклади Приклад 1 Отримання оцінки амплікону для MON89788 геномної ДНК ДНК з трансгенної лінії сої MON89788 виділяють з тканини, що включає насіння сої, тканину рослинних волокон, або крупу. ДНК виділяють з тканини, використовуючи Qiagen's DNeasy Plant Miniprep Kit, згідно з інструкціями виробника (Qiagen Corp. Valencia, CA). Отримують PCR продукт, який містить ділянку геномної ДНК, яка фланкує 5' кінець T-DNA (перенесення ДНК, що містить трансген) вставки в геном рослини, що містить MON89788. Цей ДНК продукт містить SEQ ID NO:3. PCR можна проводити, використовуючи один праймер, розроблений для гібридизації з геномними послідовностями ДНК, які фланкують 5' кінець трансгенної вставки (ДНК праймер A, SEQ ID NO:5; див. фіг. 1), спарений з другим праймером (ДНК праймер В, SEQ ID NO:6), розташованим в промоторній ділянці трансгена (патент США 6660911, SEQ ID NO:28, включений в даний опис за допомогою посилання і представлений в SEQ ID NO:9). Продукт PCR отримують від 3' кінця вставки трансгена, який містить ділянку геномної ДНК, фланкуючої 3' кінець Т-ДНК вставки в геном рослини, що містить MON89788. Цей ДНК продукт містить SEQ ID NO:4. PCR можна проводити, використовуючи один праймер, розроблений для гібридизації з геномними послідовностями ДНК, які фланкують 3' кінець вставки кожного випадку (ДНК праймер D, SEQ ID NO:8) і спарений з другим праймером (ДНК праймер 3, SEQ ID NO:7), розташованим в T-Ps.RbcS:E9 3' транскрипційної термінальних послідовності на 3' кінці вставки. Матриця PCR включає в себе 50 нг геномної ДНК. Як негативний контроль використовується 50 нг геномної ДНК нетрансгенного сорту сої. 19 94582 Кожна проба PCR містить 5 мкл 10 X буферу для TM REDAccuTaq LA DNA Polymerase Mix (SigmaAldrich, St Louis, MO), 200 мкМ кожного dNTP (Sigma-Aldrich), 0,4 мкМ кожного праймера, і 2,5 TM TM одиниць JumpStart REDTaq DNA Polymerase (Sigma-Aldrich) в 50 мкл загального об'єму проби. PCR проби виготовляють в наступних циклічних умовах: 1 цикл при 94°С протягом 3 хвилин (хв); 32 або 35 циклів при 94°С протягом 30 секунд (с), 58°С протягом 30 сек, 72°С протягом 30 сек або 1 хв.; 1 цикл при 72°С протягом 10 хв. Пари праймерів ДНК лінії використовують для отримання оцінки амплікону для геномної ДНК MON89788. Ці пари праймерів лінії включають, але ними не обмежуються, праймери А і В (SEQ ID NO:5 і 6) і пари праймерів лінії С і D (SEQ ID NO:7 і 8), які використовуються в описаному способі ампліфікації ДНК. Додатково до цих пар праймерів, будь-яка пара, отримана з SEQ ID NO:3 або SEQ ID NO:4, або їх комплементи, які при використанні в реакції ампліфікації ДНК проводять амплікон, який містить SEQ ID NO:1 або SEQ ID NO:2, діагностичний для тканини, отриманої для лінії сої MON89788, відповідно, можуть використовуватися. Умови ампліфікації ДНК, показані в таблиці 1 і 20 таблиці 2, можна використовувати для отримання діагностичного амплікону MON89788, використовуючи відповідні випадки пари праймерів. Будьяка модифікація цих способів, використана для отримання діагностичного амплікону для MON89788, знаходиться в межах знань фахівця даної галузі. Приблизно екстракт, що містить ДНК рослини або насіння сої MON89788, або продукту, отриманого з рослини, що містить MON89788, який при дослідженні способом ампліфікації ДНК проводив діагностичний амплікон для сої з генною модифікацією MON89788, можна використовувати як матрицю ампліфікації для визначення, чи присутній MON89788. Амплікон утворюється використанням щонайменше однієї праймерної послідовності, отриманої з SEQ ID NO:3 або SEQ ID NO:4, які при використанні в способі PCR утворюють діагностичний амплікон для лінії з генною модифікацією MON89788. Наприклад, отримання ампліконів MON89788 може проводитися з використанням Stratagene Robocycler, MJ Engine, Perkin-Elmer 9700, або Eppendorf Mastercycler Gradient thermocycler, як показано в таблиці 2, або способами і приладами, відомими фахівцеві в даній галузі. Таблиця 1 Процедура PCR і умови реакційної суміші для встановлення трансгенної 5' вставленої/геномної ділянки з'єднання сої MON89788 Стадія Реагент 1 Вільна від нуклеаз вода 2 10Х реакційний буфер (з MgCl2) 3 4 5 6 7 8 10 мМ розчин dATP, dCTP, dGTF і dTTP Праймер A (SEQ ID NO:5 ресуспендували в 1Х ТЕ буфері або вільній від нуклеаз воді до концентрації 10 мкМ) Праймер В (SEQ ID NO:6 ресуспендували в 1Х ТЕ буфері або вільній від нуклеаз воді до концентрації 10 мкМ) RNase, DNase free (500 мкг/мл) REDTaq ДНК полімераза (1 одиниця/мкл) Виділена ДНК (матриця): - Зразки для аналізу: окреме листя сумарне листя (максимальне 10 листя/пул) - Негативний контроль - Негативний контроль - Позитивний контроль Кількість Додати до кінцевого об'єму 20 мкл 2,0 мкл 0,4 мкл 0,2 мкл 0,2 мкл Коментарі 1Х кінцева концентрація буфера, 1,5 мМ кінцева концентрація MgCl2 200 мкМ кінцева концентрація кожного dNTP 0,1 мкМ кінцева концентрація 0,1 мкМ кінцева концентрація 0,1 мкл 50 нг на реакцію 1,0 мкл (рекомендовано скидати на- 1 одиниця на реакцію конечники до наступного етапу) - 10-200 нг геномної ДНК - 200 нг геномної ДНК -50нг нетрансгенної геномної ДНК сої - ніякої матричної ДНК (розчин, в якому ресуспендували ДНК) - 50 нг геномної ДНК сої, що містить MON89788 Обережно змішати і, якщо необхідно (ніякого нагрівання поверхні термоциклера), додати 1-2 краплі мінерального масла на поверхню кожної проби. Приступити до PCR в термоциклері Stratagene Robocycler (Stratagene, La Jolla, CA), MJ Engine (MJR-Biorad, Hercules, CA), Perkin-Elrner 21 94582 9700 (Perkin Elmer, Boston, MA), або Eppendorf Mastercycler Gradient (Eppendorf, Hamburg, Germany), використовуючи наступні циклічні параметри (таблиця 2). Термоциклер MJ Engine або Eppendorf Mastercycler Gradient повинен бути 22 встановлений в розрахованому режимі. Запустити термоциклер Perkin-Elrner 9700 зі швидкістю, яка швидко збільшується, встановленою на максимумі. Таблиця 2 Умови термоцикл ера Цикл № 1 34 1 Цикл № 1 34 1 Цикл № 1 34 1 94°С 94°С 64°С 72°С 72°С Параметри: Stratagene Robocycler 3 хвилини 1 хвилина 1 хвилина 1 хвилина і 30 секунд 10 хвилин 94°С 94°С 64°С 72°С 72°С Параметри: MJ Engine або Perkin-Elrner 9700 3 хвилини 30 секунд 30 секунд 1 хвилина 10 хвилин 94°С 94°С 64°С 72°C 72°C Параметри: Eppendorf Mastercycler Gradient 3 хвилини 15 секунд 1 5 секунд 1 хвилина 10 хвилин Приклад 2 Визначення послідовності трансгенної/геномної ділянки і метод дослідження за Саузерном ДНК секвенування продуктів PCR забезпечує ДНК, яку можна використовувати для конструювання додаткових молекул ДНК в якості праймерів і зондів для ідентифікації рослин і насіння сої, що включає генетичну модифікацію MON89788. Продукти PCR очікуваних розмірів, представлені 5' і 3' трансгенними/геномними послідовностями, були виділені шляхом розділення продуктів PCR в 2,0% агарозному гелі електрофорезом. Виділяють продукти PCR, які включають 5' і 3' ДНК ділянки, які перекривають вставлене з'єднання між трансгенною вставкою всередині геному сої. 5' і 3' продукти PCR для MON89788 очищають електрофорезом в агарозному гелі, з подальшим виділенням з агарозного матрикса з використанням набору QIAquick Gel Extraction Kit (catalog # 28704, Qiagen Inc., Valencia, CA). Очищені продукти PCR потім секвенують (наприклад, ABI Prism™ 377, ΡΕ Biosystems, Foster City, CA) і аналізують (наприклад, DNASTAR програма секвенування, DNASTAR Inc., Madison, WI). Визначали послідовність ДНК для сегмента нуклеотидних пар основ, що представляє собою трансгенну/геномну ділянку генетичної модифікації MON89788, зображена на фіг. 1 і ідентифікована як SEQ ID NO:9. Геномні і трансгенні елементи, які містять SEQ ID NO:9, описані в таблиці 3. 5' і 3' фланкуючі ділянки включені в SEQ ID NO:9 і наведені в SEQ ID NO:21 і 22. Сполучені послідовності являють собою порівняно короткі полінуклеотидні молекули, які є новими послідовностями ДНК і є діагностичними для ДНК MON89788 при виявленні в аналізі визначення полінуклеїнової кислоти. Сполучені послідовності в SEQ ID NO:1 і SEQ ID NO:2 являють собою 10 полінуклеотидів на кожній стороні ділянки вставки трансгенного фрагмента і геномної ДНК сої в MON89788. Більш довгі або короткі полінуклеотидні сполучені послідовності можна вибрати з SEQ ID NO:3 або SEQ ID NO:4. Сполучені молекули (5' зв'язувальна ділянка SEQ ID NO:1 і 3' зв'язувальна ділянка SEQ ID NO:2) корисні як ДНК зонди або як праймерні молекули ДНК в способах виявлення ДНК. Праймери і зонди, використані в Taqman® способі (Roche Molecular Systems, Inc., Pleasanton, СА) для виявлення випадку специфічної молекули ДНК, були розроблені для генної модифікації MON89788. Праймерні молекули позначені як SQ2824 (SEQ ID NO:10), SQ2826 (SEQ ID NO:11), SQ1141 (SEQ ID NO:12), SQ1142 (SEQ ID NO:13), SQ5543 (SEQ ID NO:14), і молекули зондів позначені як РВ871-6FAM (SEQ ID NO:15), PB2191-VIC (SEQ ID NO:16) і PB57-VIC (SEQ ID NO:17). Праймери і зонди були використані в способі Taqman® згідно з інструкціями виробника, щоб забезпечити діагностичний амплікон для ДНК, що включає MON89788. Тканини сої, що включають перероблені продукти, наприклад крупу, є корисними джерелами ДНК для цього способу. Додаткові праймери, використані для отримання амплікону з соєвої крупи, включають в себе SEQ ID NO:18-20. 23 94582 24 Таблиця 3 Анотація геномних і генетичних елементів трансгенного/геномного ДНК фрагмента (SEQ ID NO:9). що містився в геномі сої, який включає MON89788 Генетичний елемент 1 Фланкуючий 5' кінець послідовності вставки 5' зв'язуюча ділянка 3 В правий кордон Інтрон (інвертуюча послідовність) 4 P -FMV/Tsf1 5 L -Tsf1 6 I -Tsf1 Інтрон (інвертуюча послідовність) 7 TS -CTP2 8 CS -cp4 epsps Інтрон (інвертуюча послідовність) 9 T -E9 Інтрон (інвертуюча послідовність) B-лівий кордон 3' зв'язуюча ділянка Фланкуючий 3' кінець послідовності вставки Положення в посФункція (посилання) лідовності 2 1-1103 Геномна ДНК сої 1093-1113 ДНК ділянка, що перекриває трансгенну вставку 1104-1145 ДНК ділянка з Agrobacterium tumefaciens, що містить праву граничну послідовність, використану для перенесення Т-ДНК (Depicker et al., 1982) 1146-1215 Послідовності, використані в клонуванні ДНК 1216-2255 Химерний промотор, що складається з енхансерних послідовностей з 35S промотору Figwort Mosaic virus (Richins et al., 1987) і промотору з Tsf1 гена Arabidopsis thaliana (що кодує фактор елонгації EF-1alpha (Axelos et al., 1989)) 2256-2301 5' лідерна нетрансльована послідовність (екзон 1) з Tsf1 гена Arabidopsis thaliana, який кодує фактор елонгації EF-1alpha (Axelos et al., 1989) 2302-2923 Інтрон з Tsf1 гена Arabidopsis thaliana, що кодує фактор елонгації EF-1alpha (Axelos et al. 1989) 2924-2932 Послідовності, використані в клонуванні ДНК 2933-3160 Послідовності, які кодують транспортний білок хлоропластів з ShkG гена Arabidopsis thaliana, що кодує EPSPS (Klee et al., 1987) 3161-4528 Оптимізований кодон, який кодує послідовність аrоА гена Agrobacterium sp. штаму СР4, що кодує білок СР4 EPSPS (Padgette et al., 1996; Barry et al., 1997) 4529-4570 Послідовності, використані в клонуванні ДНК 4571-5213 3' не трансльована послідовність з RbcS2 гена Pisum sativum, який кодує малу субодиницю Rubisco(Coruzzi et al., 1984) 5214-5256 Послідовності, використані в клонуванні ДНК 5257-5406 ДНК ділянка Agrobacterium tumefaciens, що містить ліву граничну послідовність, використану для перенесення Т-ДНК(Barker et al., 1983) 5396-5416 ДНК ділянка, що перекриває трансгенну вставку 5407-6466 Геномна ДНК сої Саузерн олот-аналіз Геномну ДНК рослини, що містить MON89788, і контрольну геномну ДНК сої (15 мкг кожні) обробляють різними рестриктазами (140U) в загальному об'ємі 150 мкл, що включає 15 мкл відповідного буферу виробника (NEB, Beverly, ΜΑ). Ендонуклеази рестрикції, наприклад, Bgl11, BamH1, Nco1, Hind111 і Bcl1, використовуються в Саузерн блотаналізі MON89788. Ендонуклеазні гідролізи проводять при відповідній температурі протягом щонай 25 менше 6 годин. Після інкубації, ДНК осаджують 3М ацетатом натрію і 2,5 об'ємами етанолу. Далі ДНК відмивають 70% етанолом, висушують і ресуспендують в 40 мкл ТВЕ. Буфер для зразків (0,2 X) додають до зразків і потім піддають електрофорезу в агарозних гелях (0,8%) протягом 16-18 годин при 30 вольтах. Гелі забарвлюють бромистим етидієм, потім обробляють депуринізуючим розчином (0,125 N HCl) протягом 10 хвилин, денатуруючим розчином (0,5 Μ гідроксиду натрію, 1,5 Μ хлориду натрію) протягом 30 хвилин, і, нарешті нейтралізуючим розчином (0,5 Μ Trizma base, 1,5 Μ хлориду натрію) протягом 30 хвилин. ДНК переносять на мембрану Hybond-N (Amersham Pharmacia Biotech, Buckingamshire, England), використовуючи Turboblotter (Schleicher and Schuell, Dassel, Germany) протягом 4-6 годин і потім фіксували мембрану з використанням UV світла. Мембрани попередньо гібридизували 20 мл розчином DIG Easy Hyb (Roche Molecular Biochemicals, Indianapolis, IN; cat. #1603558) протягом 2-4 годин при 45°С. Радіоактивні ДНК зонди 32 ( Р dCTP), гомологічні або комплементарні до SEQ ID NO:1, або SEQ ID NO:2, або SEQ ID NO:3, або SEQ ID NO:4, або їх ділянку отримують, використовуючи Radprime DNA Labeling kit (Invitrogen, Carlsbad, CA; cat. #18428-011). Невключені нуклеотиди видаляють, використовуючи колонки SEPHADEX G-50 (Invitrogen). Розчин для попередньої гібридизації замінюють на 10 мл заздалегідь нагрітого розчину DIG Easy Hyb, що містить денатурований зонд до кінцевої концентрації 1 мільйон одиниць на мл. Блоти гібридизують при 45°С протягом 16-18 годин. Блоти відмивають розчином зниженої жорсткості (5Х SSC, 0,1 X SDS) при 45°С і потім повторно відмивають розчином високої жорсткості (0,1 X SSC, 0,1% SDS) при 65°С. Блоти піддають впливу люмінесцентного екрана (Amersham Biosciences, Piscataway, NJ) протягом >2 годин, і експонування оцінюють з використанням апарату Data Storm 860 (Amersham Biosciences). Ці наведені способи і умови може змінювати фахівець в даній галузі визначення ДНК в зразку. Приклад 3 Боротьба з бур'янами Контролювання росту бур'янів на полі сої, що містить MON89788. Поле засаджують насінням сої, що містить MON89788, насіння росте в рослини, і поле рослин обробляють гербіцидним препаратом, що містить гліфосат. На полі застосовують ефективну дозу препарату гліфосату в дозах обробки від приблизно 0,25 1b ае/А (фунтів еквівалента кислоти гліфосату/акр) до 3 або більше 1b ае/А. Часто дози, що застосовуються, варіюють від приблизно 0,75 1b ае/А до 1,5 1b ае/А при частоті однієї або більше обробок протягом сезону росту у міру необхідності контролю росту бур'янів на полі. З оброблених рослин збирають врожай насіння рослин, що містить MON89788. Депозит Monsanto Technology LLC, насіння сої, що представляє генну модифікацію MON89788, розкриту вище і вказану в формулі винаходу, був виконаний за умовами Будапештського договору в American Type Culture Collection (АТСС), 10801 94582 26 University Boulevard, Manassas, Va. 20110. Депозитарний номер АТСС для депозиту, що містить лінію MON89788 (також відомий як MON19788 або GM_A19788), визначений як РТА-6708 від 11 травня, 2005. Депозит буде зберігатися в депозитарії протягом 30 років, або протягом 5 років після останнього запиту, або протягом терміну дії патенту, в залежності від того, що більш тривало, і при необхідності буде замінений протягом цього періоду. Маючи продемонстровані і описані принципи даного винаходу, фахівцеві в даній галузі повинно бути очевидно, що винахід може бути модифікований за своєю структурою і деталями без відхилення від таких принципів. Автори заявляють, що всі модифікації знаходяться в межах суті і обсягу прикладеної формули винаходу. Всі публікації і опубліковані патентні документи, викладені в цьому описі, включені в даний опис за допомогою посилання в однаковій мірі, як якби включення кожної окремої публікації або патентної заявки було обумовлене особливо і індивідуально. Посилання Наступні посилання, за умови, що вони представляють зразкові методичні або інші деталі, додаткові до викладених в цьому документі, включені в даний опис за допомогою конкретного посилання. Патент США 4535060 Патент США 4683195 Патент США 4683202 Патент США 5094945 Патент США 5463175 Патент США 5554798 Патент США 5633435 Патент США 5633435 Патент США 5659114 Патент США 6040497 Патент США 6384301 Патент США 6544734 Патент США 6660911 Патент США 6660911 Патент США 6689880 Патент США 6689880 Патент США 6733974 Патент США 6740488 Патент США 6740488 Патент США 6818807 Патент США 6825400 Патент США 6893826 Патент США 6900014 Патент США 7002058 Патентна заявка США 60/685584 Публікація США 20040018518 Публікація США 2006068398 Ausubel et al., В: Current Protocols in Molecular Biology, John, Wiley & Sons, Inc, New York, 1992. Chen et al., Genome Res., 9:492-498, 1999. Cheng et al., Proc. Natl Acad. Sci USA, 91:56955699,1994. Coruzzi et al., ΕΜΒΟ J., 3:1671-1679, 1984. Cregan et al., B: DNA markers: Protocols, applications, and overviews, Wiley-Liss NY, 173-185, 1997. DeBlock et al.,EMBO J., 6:2513-2522, 1987. 27 Fehr, В: Breeding Methods for Cultivar Development, Wilcox (Ed.), Amer. Soc. of Agronomy, Madison WI, 1987. Haymes et al., B: Nucleic acid hybridization, a practical approach, IRL Press, Washington, DC, 1985. Innis, et al., B: PCR Protocols. A guide to Methods and Application, Academic Press, Inc. San Diego, 1990. Lewin, В: Genes V, Oxford University Press, NY, 1994. Nikiforov, et al. Nucleic Acid Res., 22:4167-4175, 1994. Публікація РСТ WO 9200377 94582 28 Публікація РСТ WO/05017181 Rieger et al., Β: Glossary of Genetics: Classical th and Molecular, 5 Ed., Springer-Verlag: NY, 1991. Sambrook et al., Β: Molecular cloning: a th laboratory manual, 2 Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989. Thompson et al., ΕΜΒΟ J. 6:2519-2523, 1987. Toki et al., Plant Physiol, 100:1503-1507, 1992. Туangi et al., Nature Biotech., 14:303-308, 1996. Weising et al., Ann. Rev. Genet., 22:421-477, 1988. Windeis et al., Med. Fac. Landbouww, 64/5b:459462, 1999. Wingem, Innov. Pharma. Tech., 00:18-24, 2000. 29 94582 30 31 94582 32 33 94582 34 35 94582 36 37 94582 38 39 94582 40 41 94582 42 43 94582 44 45 94582 46 47 94582 48 49 Комп’ютерна верстка Л.Литвиненко 94582 Підписне 50 Тираж 24 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюGenetic modification mon89788 of soybean and methods for its determination
Автори англійськоюMalven Marianne, Rinehardt Jennifer, Tailor Nancy, Dickinson Ellen
Назва патенту російськоюГенетическая модификация mon89788 сои и способы ее определения
Автори російськоюМалвен Марианне, Райнхарт Дженнифер, Тейлор Ненси, Дикинсон Эллен
МПК / Мітки
МПК: C12N 15/82, A01H 5/00, C12Q 1/68
Мітки: визначення, mon89788, сої, модифікація, генетична, способи
Код посилання
<a href="https://ua.patents.su/25-94582-genetichna-modifikaciya-mon89788-so-i-sposobi-viznachennya.html" target="_blank" rel="follow" title="База патентів України">Генетична модифікація mon89788 сої і способи її визначення</a>
Кристалічна модифікація iv піраклостробіну, способи її одержання, композиція для захисту рослин та застосування

Номер патенту: 89830
Опубліковано: 10.03.2010
Автори: Фогельбахер Уве Йозеф, Вукельт Йорг, Шнайдер Карль-Хайнріх, Крьоль Томас, Ноак Райнер, Раульс Маттіас, Гьотц Роланд, Майєр Вінфрід, Ерк Петер, Ціглер Ханс, Кокс Герхард
МПК: C07D 231/22, A01N 47/24
Мітки: кристалічна, композиція, піраклостробіну, застосування, захисту, одержання, модифікація, рослин, способи
Формула / Реферат:
1. Кристалічна модифікація IV піраклостробіну, яка у рентгенівській порошковій дифрактограмі при 25 °С показує щонайменше три з наступних рефлексів:d = 6,02 ± 0,01 Å, d = 4,78 ± 0,01 Å, d = 4,01 ±0,01 Å, d = 3,55 ±0,01 Å, d = 3,01 ± 0,01 Å.2. Кристалічна модифікація IV за п. 1 з температурою плавлення в межах від 62 до 72 °С.3. Кристалічна модифікація IV за п. 1 з...
Генетична матриця

Номер патенту: 58368
Опубліковано: 15.07.2003
Автор: Задорожніков Костянтин Георгійович
МПК: G06G 7/66, C07H 21/04, C12Q 1/68
Формула / Реферат:
Генетична матриця, складена з амінокислот, яка відрізняється тим, що вона скомпонована у відповідності до універсального гармонійного модуля.
Біциклічна сполука, способи її одержання, фармацевтичні композиції, що її містять,способи лікування та попередження різних захворювань, проміжні сполуки та способи їх одержання

Номер патенту: 72883
Опубліковано: 16.05.2005
Автори: Баджі Ашок Чаннавеераппа, Чакрабарті Ранджан, Лохрей Брадж Бхушан, Лохрей Відіа Бхушан, Рамануджам Раджагопалан, Калчар Шіварамаййа
МПК: A61K 9/14, A61P 19/10, A61P 13/12, A61K 31/538, A61K 9/20, A61P 9/10, A61P 25/28, A61P 3/10, A61K 9/08, C07D 413/06, A61P 3/04, A61K 9/10, A61K 31/5415, A61K 9/48, A61P 17/06, A61P 3/06, C07D 279/00, C07D 265/36
Мітки: композиції, біциклічна, проміжні, сполука, лікування, захворювань, містять,способи, способи, різних, фармацевтичні, одержання, попередження, сполуки
Формула / Реферат:
1. Сполука формули (І), (I)де групи R1, R2, R3, R4 і групи R5 і R6, коли вони приєднані до атома вуглецю, можуть бути однаковими або різними і означають водень, галоген, гідрокси або необов'язково заміщену групу, вибрану з алкілу, алкокси, фенілу, карбонової кислоти або сульфонової кислоти ; один або обидва замісники R5 і R6 можуть також означати оксогрупу,...
Трансформатор високої напруги і великої потужності з розподілюючим екраном (ре) долюка і способи виготовлення, визначення параметрів і випробування ре (варіанти)

Номер патенту: 73849
Опубліковано: 15.09.2005
Автори: Шевкунова Олена Романівна, Долюк Роман Петрович, Долюк Іван Петрович, Остащенко Ольга Романівна
МПК: H01F 27/34, H01F 41/06
Мітки: розподілюючим, екраном, способи, параметрів, потужності, долюка, великої, напруги, варіанти, випробування, визначення, високої, трансформатор, ре, виготовлення
Формула / Реферат:
1. Трансформатор високої напруги (ВН) і великої потужності з приєднаним до лінійного виводу обмотки ВН розподілюючим екраном (РЕ) спірального типу, що містить послідовні ємності,сформовані шарами концентричних між собою і витками обмотки ізольованих дугових обкладок, що мають у перерізі форму співвісних овалів, і з'єднані по колу з розміщенням точок входу і виходу кожної ємності на різнойменних краях її обкладок з утворенням суцільної...
Способи визначення несучої здатності мастильної плівки та придатності мастильного матеріалу до використання

Номер патенту: 93616
Опубліковано: 25.02.2011
Автори: Літовка Сергій Володимирович, Лисіков Євген Миколайович, Косолапов Віктор Борисович, Воронін Сергій Володимирович
МПК: G01N 33/26, G01N 27/02
Мітки: мастильної, несучої, використання, визначення, здатності, плівки, мастильного, придатності, матеріалу, способи
Формула / Реферат:
1. Спосіб визначення несучої здатності мастильної плівки, за яким послідовно навантажують трибосистему з точковим контактом, яка реалізована у вигляді контакту куль, у середовищі мастильного матеріалу, який відрізняється тим, що реєструють навантаження руйнування мастильної плівки по досягненню металевого контакту, який визначають шляхом порівняння електричного опору змащеного контакту з попередньо визначеною величиною електричного опору...
Попередній патент: Пристрій для демонстрації інформації з растровою структурою освітлювальних елементів
Наступний патент: Композиція для внесення в ґрунти
Випадковий патент: Спосіб лікування кишкової непрохідності