Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів
Номер патенту: 45941
Опубліковано: 25.11.2009
Автори: Романенко Олександр Вікторович, Касаткіна Людмила Олександрівна, Борисов Арсеній Андрійович, Вовк Андрій Іванович, Борисова Тетяна Олександрівна, Крисанова Наталія Валеріївна, Остапченко Людмила Іванівна



Формула / Реферат
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів, загальної формули:
 .
.
Текст
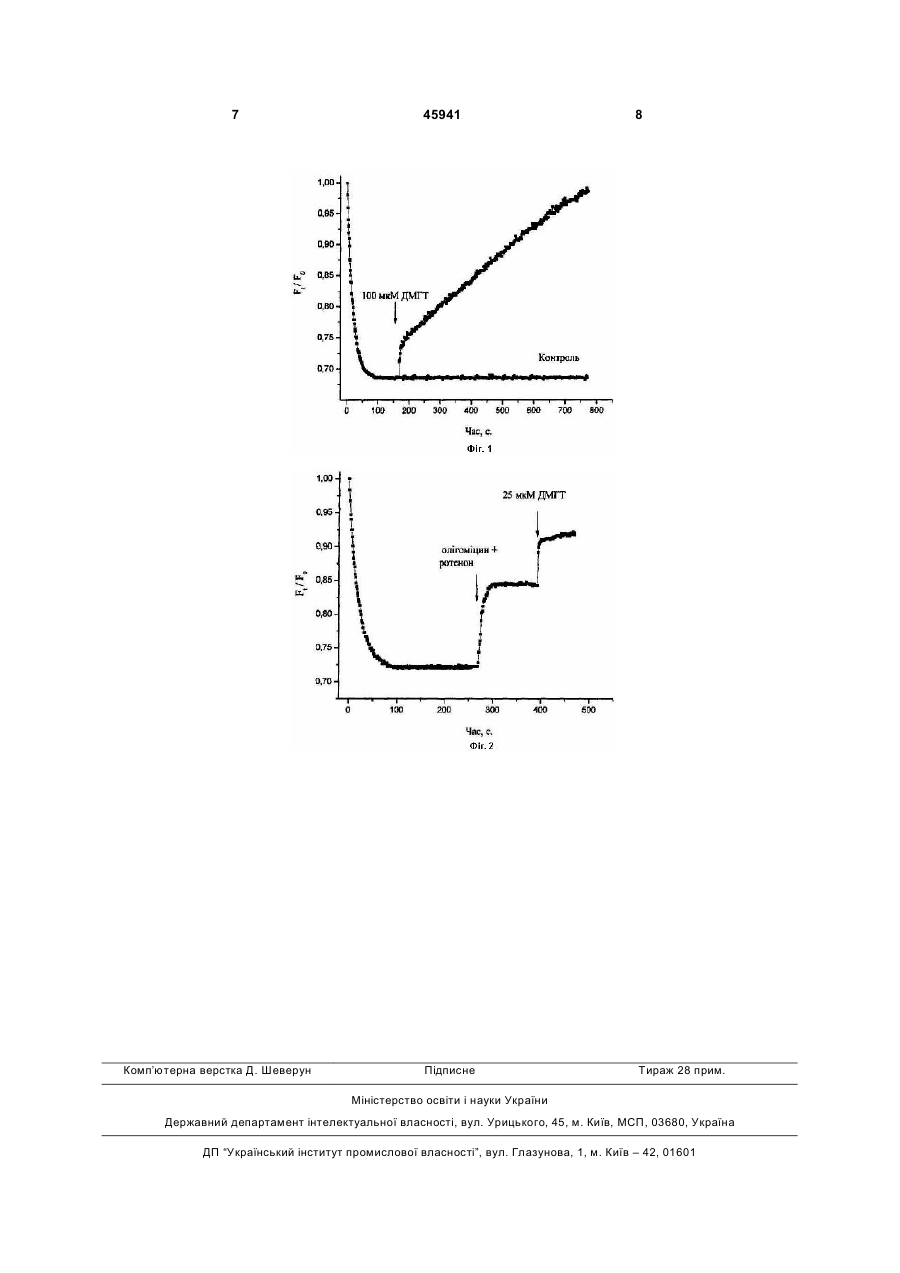
Застосування сполуки 3децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів, загальної формули: H21C10CO2CH2 CH3 + N Cl Корисна модель належить до медицини, а саме до фармакології. Завданням корисної моделі є дослідження дії сполуки 3-децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлориду (ДМГТ) рухів, розвиваються нейродегенеративні процеси [1]. Експериментально встановлено активуючий вплив вітаміну В1 на процеси квантової секреції нейромедіатора у синапсах різних типів [2-4]. Молекула тіаміну містить у своєму складі тіазолієвий та піримідиновий цикли, які поєднані метиленовим містком. Наявність тіазолієвого циклу є характерною особливістю тіаміну та його похідних. Серед усіх ендогенних біологічно активних речовин в організмі лише тіамін та його похідні містять у своєму складі тіазолієвий цикл. Раніше було синтезовано структурний аналог вітаміну В1 з непорушеним тіазолієвим циклом - ДМГТ [5]. У складі молекули ДМГТ присутній довгий вуглеводневий хвіст, що, очевидно, сприяє вбудовуванню ДМГТ у біомембрани. За багатьма властивостями ДМГТ може розглядатись як потенційний лікарський засіб [5-9]. Уведений лабораторним тваринам ДМГТ чинить на них седативну, центральну міорелаксуючу, протисудомну, снодійну дію. При цьому також спостерігається зниження больової чутливості, пригнічення орієнтувальної пошукової поведінки тварин. ДМГТ також здатен знижувати нейротоксичний вплив на тварин білкового токсину α-латротоксину, виділеного з отрути павука каракурта (Latrodectus mactans tradecimguttatus). В міоневральних синапсах ДМГТ викликає блокування (11) 45941 (13) U . UA CH2CH2OH (19) на мембранний потенціал ізольованих нервових закінчень головного мозку щурів. Вивчення дії ДМГТ на мембранний потенціал ізольованих нервових закінчень головного мозку щурів проводилось у відділі нейрохімії Інституту біохімії ім. О.В. Палладіна НАН України і на кафедрі біології Національного медичного університету імені О.О. Богомольця. ДМГТ синтезовано у відділі механізмів біоорганічних реакцій Інституту біоорганічної хімії та нафтохімії НАН України. Вітамін В1 (тіамін) є незамінним компонентом процесів обміну речовин в центральній нервовій системі. При В1-гіпо- та В1-авітамінозах виникають порушення рефлекторної діяльності, координації S 3 викликаної квантової секреції медіатора у відповідь на подразнення нервових волокон або дію αлатротоксину. У модельних штучних фосфоліпідних мембранах ДМГТ знижує провідність іонних каналів, сформованих α-латротоксином, амфотерицином В або ністатином. Глутамат є нейромедіатором у центральній нервовій системі ссавців, який обумовлює збудження нервових клітин. Він бере участь у здійсненні таких важливих функцій головного мозку, як розпізнавання, пам'ять та навчання. Було показано, що ДМГТ викликає інгібування процесу натрійзалежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів [10]. Потенціал плазматичної мембрани нервових закінчень є одним з ключових факторів, який визначає ефективність синаптичної передачі. Його зниження призводить до модуляції роботи потенціал-залежних іонних каналів, натрій-залежних транспортерів, що може обумовлювати зміни у вивільненні нейромедіаторів шляхом екзоцитозу з нервових закінчень та у процесі синаптичної передачі в цілому. Беручи до уваги дані, що наведені вище, доцільним є аналіз впливу ДМГТ на потенціал плазматичної мембрани нервових закінчень головного мозку щурів, як потенційного шляху модуляції цією сполукою синаптичної передачі. В основу корисної моделі поставлено те, що ДМГТ викликає деполяризацію плазматичної мембрани нервових закінчень (синаптосом) ізольованих з головного мозку щурів. Додавання ДМГТ (25100 мкМ) до синаптосом, попередньо навантажених потенціал-чутливим флуоресцентним зондом родаміном 6G, призводить до вивільнення флуоресцентного зонду з синаптосом. Повне вивільнення флуоресцентного зонду родаміну 6G відбувається упродовж 10 хв при додаванні до синаптосом ДМГТ у концентрації 100 мкМ. Дія ДМГТ є дозо-залежною та не залежить від присутності іонів кальцію в середовищі інкубації. Ознаки способу. Методика виділення пресинаптичних нервових закінчень (синаптосом) головного мозку щурів та вимірювання потенціалу плазматичної мембрани синаптосом. Виділення синаптосом з головного мозку щурів. Щурів-самців лінії Wistar масою 100-120 г декапітували, великі півкулі головного мозку швидко переносили в розчин, що містив 0,32 М сахарози, 5 мМ Hepes-NaOH (рН 7,4) та 0,2 мМ етилендіамінтетраоцтової кислоти (ЕДТА). Усі операції проводилися при 4°С. Синаптосоми виділяли з гомогенату мозку диференційним центрифугуванням і центрифугуванням в градієнті щільності фіколлу, застосовуючи метод Котмана [11] у такій модифікації: розчин сахарози для приготування градієнту фіколлу містив 5 мМ Hepes-NaOH (рН 7,4) і 0,2 мМ ЕДТА [12]. Синаптосомальну фракцію, отриману при центрифугуванні гомогенату головного мозку в градієнті щільності фіколлу, розводили 10 об'ємами 0,32 М сахарози, 5 мМ Hepes-NaOH (рН 7,4) і 0,2 мМ ЕДТА та центрифугували при 20000 g 45941 4 упродовж 20 хв. Отриманий осад повільно суспендували в 4 мл оксигенованого холодного середовища, що містило (в мМ): NaCl - 126, КСl - 5, MgCl2 - 1,4, NaH2PO4 - 1,0, HEPES - 20, СаСl2 - 2, dглюкозу - 10 (рН 7,4). При цьому кінцева концентрація білка становила 4 мг/мл. Синаптосоми використовували в експериментах упродовж 2-4 годин після отримання. Концентрацію білка визначали за методом Ларсона [13]. Вимірювання потенціалу плазматичної мембрани синаптосом. Дослідження змін потенціалу плазматичної мембрани синаптосом проводили з використанням потенціал-чутливого флуоресцентного зонду родаміну 6G (Molecular Probes, США). Він зв'язується з плазмалемою синаптосом та з мембраною мітохондрій залежно від їх мембранних потенціалів. Щоб виключити внесок мітохондрій у зміни загального рівня флуоресценції синаптосом, дослідження проводили в присутності блокатору дихального ланцюга мітохондрій - ротенону (4 мкМ), та інгібітору мітохондріальної аденозинтрифосфатази (АТФази) - олігоміцину (4 мг/мл). Суспензію синаптосом преінкубували в кальцієвмісному (2 мМ СаСl2) та безкальцієвому середовищі (2 мМ ЕГТА) середовищі, що містило (в мМ): NaCl - 126, КСl - 5, MgCl2 - 1,4, NaH2PO4 - 1,0, HEPES - 20, d-глюкозу 10 (рН 7,4) упродовж 10 хв при 37°С, а потім вносили в спеціальну кварцеву кювету для вимірювання флуоресценції, що містила родамін 6G (0,5 мкМ). Флуоресценцію реєстрували при постійному перемішуванні суспензії синаптосом при 37°С (кінцева концентрація білка - 0,15 мг/мл). Потенціал плазматичної мембрани синаптосом оцінювали по інтенсивності флуоресценції родаміну 6G згідно формули: F=Ft/F0 де F0 - інтенсивність флуоресценції родаміну 6G за відсутності синаптосом у кюветі, Ft - інтенсивність флуоресценції родаміну 6G у присутності синаптосом у кюветі. Інтенсивність флуоресценції родаміну 6G визначалась на спектрофлуориметрі Hitachi MPF-4 (Японія). При цьому довжина хвилі збудження становила 528 нм, а довжина хвилі емісії - 551 нм (спектральна ширина щілин - 5 нм). В наших експериментах накопичення родаміну 6G синаптосомами відбувалося упродовж однієї хвилини (Фіг. 1, 2). У безкальцієвому середовищі (Фіг.1) додавання 100 мкМ ДМГТ призводило до швидкого вивільнення з синаптосом акумульованого родаміну 6G, що свідчило про миттєву деполяризацію мембранних структур нервових закінчень викликану ДМГТ. В наших дослідженнях мінімальна концентрація ДМГТ при якій була зафіксована його дія на мембранний потенціал синаптосом становила 25 мкМ. Ступінь впливу ДМГТ на нього зростав при збільшенні концентрацій ДМГТ в інкубаційному середовищі від 25 до 100 мкМ, що вказувало на дозо-залежність зазначеного ефекту ДМГТ. Було також показано, що у присутності 2 мМ Са2+ у середовищі інкубації синаптосом, ДМГТ в концентрації 100 мкМ викликав ефект подібний тому, що було зафіксовано у безкальцієвому середовищі. Таким чином, дія ДМГТ на мембранний 5 потенціал синаптосом не залежить від присутності іонів кальцію у середовищі інкубації. Оскільки родамін 6G зв'язується з плазматичною мембраною нервових закінчень та з мембраною мітохондрій відповідно до їх потенціалів, для визначення впливу ДМГТ саме на плазматичну мембрану, виключивши при цьому можливий внесок мітохондрій у реєструємі зміни загального рівня флуоресценції, було використано блокатор дихального ланцюга мітохондрій - ротенон (4 мкМ) та інгібітор мітохондріальної АТФази - олігоміцин (4 мг/мл). Як свідчать наведені на Фіг. 2 дані, ДМГТ в концентрації 25 мкМ викликає в присутності ротенону та олігоміцину в середовищі інкубації вивільнення попередньо акумульованого синаптосомами флуоресцентного зонду родаміну 6G, в наслідок чого суттєво (на 30 %) збільшується інтенсивність флюоресцентного сигналу. Це свідчить про обумовлену ДМГТ деполяризацію безпосередньо плазматичної мембрани нервових закінчень. Таким чином, слід підкреслити наступне: 1. ДМГТ в концентрації 100 мкМ викликає повне вивільнення попередньо акумульованого синаптосомами потенціал-чутливого флуоресцентного зонду родаміну 6G упродовж 10 хв, що свідчить про швидку деполяризацію мембранних структур нервових закінчень. Ефект ДМГТ є дозозалежним. Мінімальна концентрація ДМГТ при якій була зафіксована його дія на мембранний потенціал синаптосом становила 25 мкМ. 2. В присутності в інкубаційному середовищі блокатору дихального ланцюга мітохондрій - ротенону та інгібітору мітохондріальної АТФази олігоміцину (умов за яких виключено внесок мітохондрій у зміни загального рівня флуоресценції синаптосом), було виявлено зменшення мембранного потенціалу плазматичної мембрани нервових закінчень на 30 % при додаванні 25 мкМ ДМГТ в середовище інкубації. 3. Дія ДМГТ на мембранний потенціал плазматичної мембрани нервових закінчень не залежить від присутності іонів кальцію в середовищі інкубації. Література: 1. Bettendorff L. Thiamine in excitable tissues: reflections on a non-cofactor role // Metab. Brain Dis. 1994. - 9, № 3. - P. 183-209. 2. Романенко А.В. Действие тиамина на нервно-мышечную передачу у лягушки // Нейрофизиология. - 1985. - 17, № 6. - с. 794-800. 3. Романенко А.В. Действие тиамина на различные типы синаптических соединений // Нейрофизиология. - 1986. - 18, № 5. - с. 621-629. 4. Романенко А.В., Гнатенко В.М., Владимирова И.А.. Действие тиамина на нервно-мышечную передачу в гладких мышцах // Нейрофизиология. 1994. - 26, № 6. - С. 449-457. 5. Вовк А.И., Романенко А.В., Муравьева И.В., Зайцев Л.М. 3-децоксикарбонилметил-4-метил-5 45941 6 β-гидроксиэтилтиазолий хлорид или децоксикарбонилметил-4-метилтиазолий хлорид, угнетающие нервно-мышечную передачу и обладающие транквилизирующей активностью // Авторское свидетельство 1547267 СССР, МКИ4 С07D 277/24, А61К 31/425. Заявлено 18.07.88. Зарегистрировано 01.11.89. 6. Романенко А.В., Вовк А.И., Шатурский О.Я. Действие тиазолевых аналогов витамина В1 на нервно-мышечную передачу и вызванную αлатротоксином секрецию медиатора в скелетной мышце // Нейрофизиология. - 1995. - 27, № 5/6. - с. 368-374. 7. Романенко А.В., Гнатенко В.М., Владимирова И.А., Вовк А.И. Пре- и постсинаптическая модуляция нервно-мышечной передачи в гладких мышцах тиазолевыми аналогами витамина В1 // Нейрофизиология. - 1995. - 27, № 5/6. - с. 375-386. 8. Романенко О.В., Вовк А.І., Гіммельрейх Н.Г. Шатурський О.Я. Сполука 3децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином В. // Патент на корисну модель № 22875 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 квітня 2007 р. - 8 с. 9. Шатурський О.Я. Романенко О.В., Вовк А.І., Гіммельрейх Н.Г. Сполука 3децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином. // Патент на корисну модель № 27417 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 жовтня 2007 р. - 8 с. 10. Борисова Т.О., Крисанова Н.В., Сівко Р.В., Романенко О.В., Вовк А.І. Сполука 3децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрій-залежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів // Патент на корисну модель № 38155 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 грудня 2008 р. - 8 с. 11. Cotman C. W. Isolation of synaptosomal and synaptic plasma membrane fractions // Meth. Enzymol. - 1974. - 31. - P. 445-452. 12. Borisova T.A., Krisanova N.V. Presynaptic transporter-mediated release of glutamate evoked by the protonophore FCCP increases under altered gravity conditions // Adv. Space Res.- 2008. - 42, N 12. P. 1971-1979. 13. Larson E., Howlett В., Jagendorf A. Artificial reductant enhancement of the Lowry method for protein determination // Anal. Biochem. - 1986. - 155. - P. 243-248. 7 Комп’ютерна верстка Д. Шеверун 45941 8 Підписне Тираж 28 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of compound 3-decyloxycarbonylmethyl-4-methyl-5-(2-hydroxyethyl)thiazolium chloride as compound promoting depolarization of cytolemma of nerve terminals isolated from rat cerebral cortex
Автори англійськоюBorysova Tetiana Oleksandrivna, Romanenko Oleksandr Viktorovych, Ostapchenko Liudmyla Ivanivna, Kasatkina Liudmyla Oleksandrivna, Krysanova Natalia Valeriivna, Borysov Arsenii Andriiovych, Vovk Andrii Ivanovych
Назва патенту російськоюПрименение соединения 3-децилоксикарбонилметил-4-метил-5-(2-гидроксиэтил)тиазолий хлорид как соединения, которое вызывает деполяризацию плазматической мембраны изолированных нервных окончаний головного мозга крыс
Автори російськоюБорисова Татьяна Александровна, Романенко Александр Викторович, Остапченко Людмила Ивановна, Касаткина Людмила Александровна, Крысанова Наталия Валериевна, Борисов Арсений Андреевич, Вовк Андрей Иванович
МПК / Мітки
МПК: C07D 277/00
Мітки: нервових, яка, деполяризацію, викликає, головного, закінчень, сполуки, щурів, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, мозку, плазматичної, хлорид, мембрани, застосування, ізольованих
Код посилання
<a href="https://ua.patents.su/4-45941-zastosuvannya-spoluki-3-deciloksikarbonilmetil-4-metil-5-2-gidroksietiltiazolijj-khlorid-yak-spoluki-yaka-viklikaeh-depolyarizaciyu-plazmatichno-membrani-izolovanikh-nervovikh-zaki.html" target="_blank" rel="follow" title="База патентів України">Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів</a>
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрієзалежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів

Номер патенту: 38155
Опубліковано: 25.12.2008
Автори: Вовк Андрій Іванович, Сівко Роман Віталійович, Романенко Олександр Вікторович, Борисова Тетяна Олександрівна, Крисанова Наталія Валеріївна
МПК: C07D 277/00
Мітки: закінченнями, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, глутамату, натрієзалежного, яка, процес, головного, має, хлорид, нервовими, сполука, ефект, накопичення, ізольованими, щурів, інгібуючий, мозку
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрієзалежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів, загальної формули:.
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином

Номер патенту: 27417
Опубліковано: 25.10.2007
Автори: Романенко Олександр Вікторович, Вовк Андрій Іванович, Шатурський Олег Ярославович, Гіммельрейх Ніна Германівна
МПК: C07D 277/00
Мітки: сполука, ністатином, має, ефект, яка, провідність, утворених, іонну, каналів, хлорид, блокуючий, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином, загальної формули:.
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил) тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином в

Номер патенту: 22875
Опубліковано: 25.04.2007
Автори: Вовк Андрій Іванович, Шатурський Олег Ярославович, Гіммельрейх Ніна Германівна, Романенко Олександр Вікторович
МПК: C07C 255/00, A61K 47/12
Мітки: ефект, сполука, яка, тіазолій, утворених, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил, блокувальний, має, амфотерицином, хлорид, провідність, каналів, іонну
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил) тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином В, загальної формули:.
Спосіб визначення реактивності головного мозку

Номер патенту: 36755
Опубліковано: 16.04.2001
Автори: Березовчук Людмила Василівна, Костилев Михайло Володимирович
МПК: A61B 5/04
Мітки: визначення, головного, реактивності, мозку, спосіб
Текст:
...дезорганізована активність. Було встановлено діагноз: реактивність мозку знижена. Запропонований спосіб визначення реактивності головного мозку було застосовано на 42 хворих на цироз печінки, з яких за фоновою ЕЕГ у 19 осіб було встановлено високу реактивність головного мозку і у 23 осіб - знижену. Для визначення реактивності головного мозку за прототипом обстежуваним було необхідне додаткове проведення функціональної проби з...
Застосування гліального фібрилярного кислого білка структур головного мозку як критерію оцінки функціонального стану щитоподібної залози

Номер патенту: 12548
Опубліковано: 15.02.2006
Автори: Неруш Петро Опанасович, Демченко Олена Михайлівна, Дроздов Олексій Леонідович
МПК: A61B 5/0484
Мітки: головного, функціонального, застосування, кислого, оцінки, щитоподібної, стану, структур, білка, критерію, фібрилярного, мозку, гліального, залози
Формула / Реферат:
Застосування гліального фібрилярного кислого білка структур головного мозку як критерію оцінки функціонального стану щитоподібної залози.
Попередній патент: Нитконапрямник ткацького верстата
Випадковий патент: Табурет