Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів
Номер патенту: 45942
Опубліковано: 25.11.2009
Автори: Романенко Олександр Вікторович, Борисова Тетяна Олександрівна, Крисанова Наталія Валеріївна, Борисов Арсеній Андрійович, Сівко Роман Віталійович, Вовк Андрій Іванович



Формула / Реферат
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів, загальної формули:
 .
.
Текст
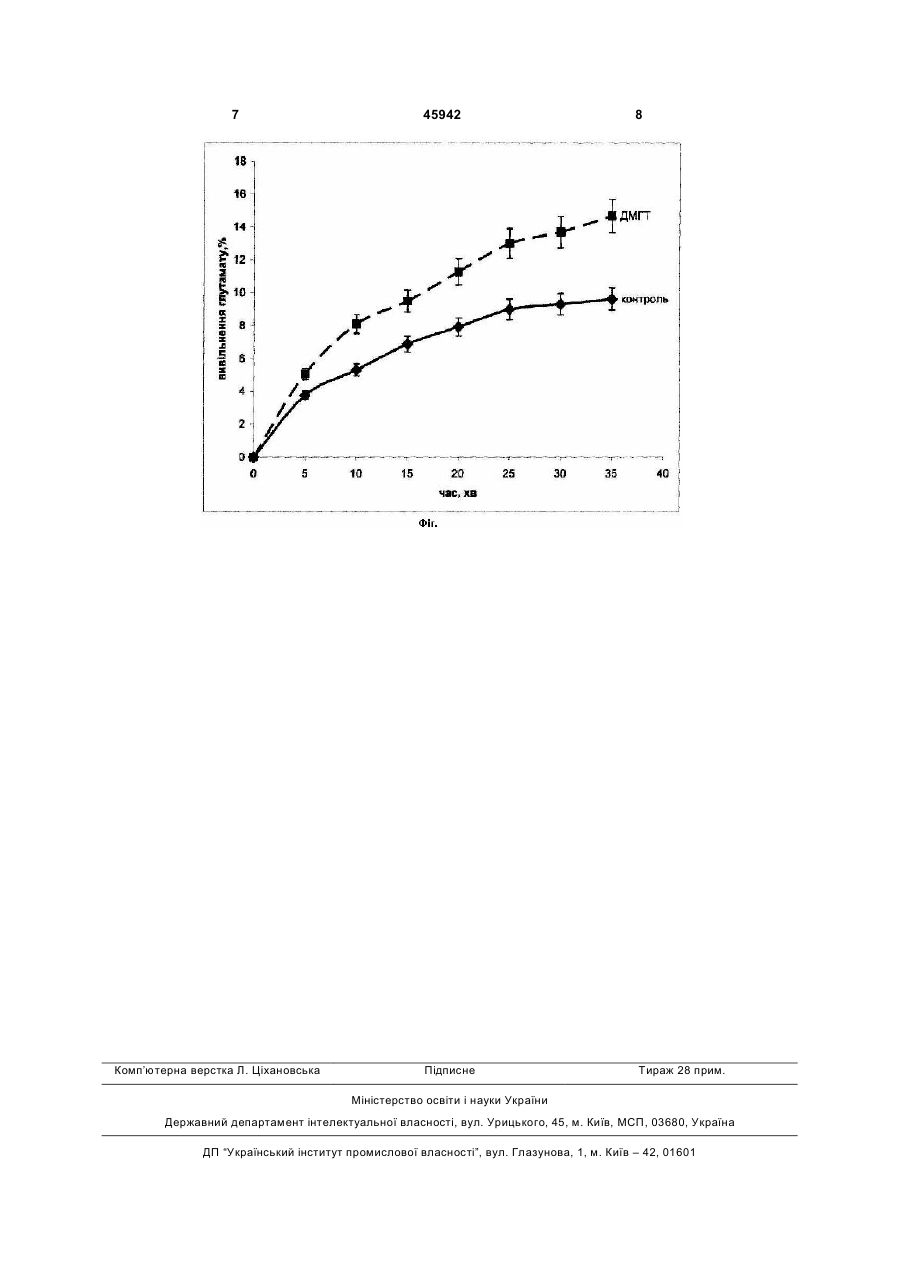
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів, загальної формули: H21C10CO2CH2 CH3 + N Cl Корисна модель належить до медицини, а саме до фармакології. Завданням корисної моделі є дослідження дії сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксиетил)тіазолій хлориду (ДМГТ) H21C10CO2CH2 CH3 + N Cl неративні процеси [1]. Тіамін залучений до контролю процесів квантової секреції нейромедіатора у синапсах різних типів [2-4]. До складу молекули тіаміну входять тіазолієвий та піримідиновий цикли, які поєднані метиленовим містком. Серед біологічно активних речовин в організмі, наявність тіазолієвого циклу є характерною особливістю структури тіаміну та його похідних. Синтезований аналог вітаміну В1 - ДМГТ має непорушений тіазолієвий цикл [5]. Довгий вуглеводневий хвіст у складі ДМГТ сприяє вбудовуванню молекули у фосфоліпідний бішар біологічних мембрани. Ряд властивостей ДМГТ дозволяє розглядати цю сполуку як потенційний лікарський засіб [5-9]. При уведенні в організм лабораторних тварин ДМГТ знижує больову чутливість, пригнічує орієнтувальну та пошукову поведінку, викликає седативний, центральний міорелаксуючий, протисудомний, снодійний ефекти [5]. Було показано, що ця сполука здатна знижувати нейротоксичний вплив білкового токсину a-латротоксину з отрути павука каракурта (Latrodectus mactans tradecimguttatus) [6]. ДМГТ викликає блокування квантової секреції медіатора в міоневральних синапсах у відповідь на подразнення нервових волокон або прикладання a-латротоксину. В дослідах на штучних фосфоліпідних мембранах було встановлено, що ДМГТ U (13) 45942 (11) на нестимульоване вивільнення глутамату з ізольованих нервових закінчень (синаптосом) головного мозку щурів. Вивчення дії ДМГТ на нестимульоване вивільнення глутамату з синаптосом, виділених з головного мозку щурів, проводилося у відділі нейрохімії Інституту біохімії ім. О.В. Палладіна НАН України і на кафедрі біології Національного медичного університету імені О.О. Богомольця. ДМГТ синтезовано у відділі механізмів біоорганічних реакцій Інституту біоорганічної хімії та нафтохімії НАН України. Вітамін B1 (тіамін) є необхідним для функціонування центральної та периферичної нервової системи ссавців. При недостатності вітаміну В1 в організмі виникають розлади рефлекторної діяльності, координації рухів, розвиваються нейродеге . UA CH2CH2OH CH2CH2OH (19) S S 3 знижує провідності іонних каналів, сформованих в них a-латротоксином, амфотерицином В або ністатином [6, 8, 9]. Було також продемонстровано, що в ізольованих нервових закінченнях головного мозку щурів ДМГТ суттєво пригнічує процес натрій-залежного накопичення глутамату - нейромедіатору, що обумовлює процесі збудження у центральній нервовій системі ссавців [10]. Стимульоване деполяризацією нервових закінчень Са2+-залежне вивільнення з них нейромедіаторів, яке відбувається під час злиття синаптичних везикул з плазматичною мембраною нервових закінчень, є одним з ключових етапів синаптичної передачі. Вивільнений при цьому в синаптичну щілину глутамат зв'язується з мембранними рецепторами та активує відповідні сигнальні шляхи, що забезпечує подальшу передачу сигналу. За відсутності стимуляції нервових закінчень концентрація позаклітинного глутамату є однією з основних характеристик, що визначає спонтанну активність мозку. За нормальних умов концентрація глутамату в синаптичній щілині підтримується на достатньо низькому рівні, що становить менше ніж 1мкМ. Завдяки цьому унеможливлюється надмірна активація іонотропних та метаботропних глутаматних пост- та пресинптичних рецепторів. В той же час надзвичайно важливим для нормального функціонування мозку є процес тонічної активації цих рецепторів позаклітинним глутаматом. Підвищена концентрація глутамату у синаптичній щілині порушує синаптичну передачу та створює підґрунтя для розвитку неврологічних симптомів. Базальне вивільнення глутамату з нервових закінчень, яке відбувається за відсутності їх подразнення, є однім з процесів, що визначає позаклітинну концентрацію глутамату в стані спокою. Беручи до уваги наведені вище дані, є доцільним проаналізувати вплив ДМГТ на нестимульоване вивільнення глутамату з нервових закінчень головного мозку щурів. В основу корисної моделі поставлено те, що ДМГТ викликає збільшення нестимульованого вивільнення радіоактивно міченого L-[14C]глутамату з попередньо навантажених ним препаратів ізольованих нервових терміналей (синаптосом) головного мозку щурів. Нестимульоване вивільнення L[14С]глутамату з синаптосом збільшувалося в присутності ДМГТ у концентрації 100мкМ у середовищі інкубації на 30% у порівнянні з контролем. Дія ДМГТ не залежить від присутності іонів кальцію в середовищі інкубації. Позаклітинний рівень L[14C]глутамату становив в кальційвмісному середовищі за відсутності ДМГТ - 0,235±0,013нмоль/мг білка, в присутності ДМГТ в концентрації 100мкМ в середовищі інкубації упродовж 40хв. 0,290±0,013нмоль/мг білка; а в безкальцієвому середовищі за відсутності ДМГТ 0,231±0,013нмоль/мг білка, в присутності ДМГТ в концентрації 100мкМ - 0,280±0,013нмоль/мг білка. Ознаки способу. Методика виділення нервових закінчень (синаптосом) з головного мозку щурів та визначення вивільнення L-[14C]-глутамату з синаптосом 45942 4 Виділення синаптосом з головного мозку щу рів. Щурів-самців лінії Wistar масою 100-120г декапітували, великі півкулі головного мозку швидко переносили в розчин, що містив 0,32М сахарози, 5мМ Hepes-NaOH (pH 7,4) та 0,2мМ етилендіамінтетраоцтової кислоти (ЕДТА). Усі операції проводилися при 4°С. Синаптосоми виділяли з гомогенату мозку диференційним центрифугуванням і центрифугуванням в градієнті щільності фіколлу, застосовуючи метод Котмана [11] у такій модифікації: розчин сахарози для приготування градієнту фіколлу містив 5мМ Hepes-NaOH (pH 7,4) і 0,2мМ ЕДТА [12]. Синаптосомальну фракцію, отриману при центрифугуванні гомогенату головного мозку в градієнті щільності фіколлу, розводили 10 об’ємами 0,32М сахарози, 5мМ Hepes-NaOH (pH 7,4) і 0,2мМ ЕДТА та центрифугували при 20000g упродовж 20хв. Отриманий осад повільно суспендували в 4мл оксигенованого холодного середовища, що містило (в мМ): NaCl - 126, КСl - 5, MgCl2 - 1,4, NaH2PO4 - 1,0, HEPES - 20, СаСl2 - 2, dглюкозу - 10 (pH 7,4). При цьому кінцева концентрація білка становила 4 мг/мл. Синаптосоми використовували в експериментах упродовж 2-4 годин після отримання. Концентрацію білка визначали за методом Ларсона [13]. Визначення вивільнення L-[14С]глутамату з синаптосом. Суспензія синаптосом розводилася стандартним сольовим розчином так, що містила 1мг білка/мл, і після 10хв преінкубації при 37°С навантажувалася L-[14С]глутаматом (500нМ, 238мСі/ммол) (Amersham, Великобританія) в кальцієвому стандартному сольовому розчині упродовж 10хв. Після цього суспензія синаптосом відмивалася 10 об'ємами льодяного стандартного сольового розчину і розводилася до концентрації 1мг білка/мл і відразу використовувалася для визначення вивільнення L-[14С]глутамату з синаптосом. Аліквоти (120мкл; 25-30мкг навантажених L[14С]глутаматом синаптосом), преінкубували 10хв при 37°С. Нестимульоване вивільнення L[14С]глутамату з синаптосом у кальційвмісному середовищі та у безкальцієвому середовищі відбувалося 1-40хв. Через визначений проміжок часу суспензію синаптосом швидко осаджували в мікроцентрифузі, 10000g протягом 20с. Аліквоти надосаду (90мкл) та солюбілізованого додецилсульфатом натрію осаду (90мкл) змішували з синтиляційною рідиною ACS (Amersham, Великобританія) (1,5мл) та визначали радіоактивність за допомогою синтиляційного лічильника Tracor Analytic Delta 300 (США). Загальний вміст радіоактивності визначали як суму радіоактивності у аліквоті надосаду та у аліквоті солюбілізованого осаду. Згідно протоколу дослідження нестимульоване вивільнення глутамату з ізольованих нервових закінчень головного мозку щурів вивчалося з використанням радіоактивно міченого L-[14С]глутамату, яким попередньо були навантажені синаптосоми (див. методику). В наших експериментах спостерігається постійне нестимульоване (базальне) вивільнення L-[14С]глутамату, що є однією з основних 5 характеристик препарату синаптосом (Фіг., суцільна лінія). На теперішній час механізм, завдяки якому L-[14С]глутамат спонтанно вивільнюється з нервових закінчень у синаптичну щілину, остаточно не з'ясовано, але передбачається можливість залучення до цього процесу спонтанного екзоцитозу, аніонних каналів, цистеїн-глутаматного обмінника та трансмембранної дифузії. Було продемонстровано, що прикладання ДМГТ до нервових закінчень призводило до збільшення нестимульованого вивільнення L-[14С]глутамату з синаптосом. На Фіг. (переривиста лінія) представлено динаміку нестимульованого вивільнення L-[14С]глутамату з синаптосом за умови їх інкубації у присутності ДМГТ в концентрації 100мкМ. Показано, що ДМГТ суттєво посилює нестимульоване вивільнення L[14С]глутамату з ізольованих нервових закінчень упродовж 1-35хв (n=5; Р≤0,05). Було показано, що в присутності іонів Са2+ 100мкМ ДМГТ викликає ефект подібний такому, що було зафіксовано у безкальцієвому середовищі. За умови інкубації синаптосом у кальційвмісному середовищі без ДМГТ позаклітинний рівень L-[14С]глутамату становив 0,235±0,013нмоль/мг білка, а з ДМГТ у концентрації 100мкМ упродовж 40хв - 0,290±0,013нмоль/мг білка; а у безкальцієвому середовищі без ДМГТ позаклітинний рівень L-[14С]глутамату становив 0,231±0,013нмоль/мг білка, а з ДМГТ в концентрації 100мкМ упродовж 40хв - 0,280±0,013нмоль/мг білка. Таким чином, дія ДМГТ не залежить від присутності іонів кальцію в середовищі інкубації. Таким чином, слід підкреслити наступне: 1. ДМГТ збільшує нестимульоване вивільнення L-[14С]глутамату з ізольованих нервових закінчень (синаптосом) головного мозку щурів. 2. Внесення ДМГТ у концентрації 100мкМ у середовище інкубації синаптосом обумовлює збільшення нестимульованого вивільнення з них L[14С]глутамату на 30%. 3. Обумовлене ДМГТ посилення нестимульованого вивільнення L-[14С]глутамату з нервових закінчень не залежить від присутності іонів кальцію в середовищі інкубації. Джерела інформації: 1. Bettendorff L. Thiamine in excitable tissues: reflections on a non-cofactor role // Metab. Brain Dis. - 1994. - 9, №3. - P.183-209. 2. Романенко А.В. Действие тиамина на нервно-мышечную передачу у лягушки // Нейрофизиология. - 1985. - 17, №6. - С.794-800. 3. Романенко А.В. Действие тиамина на различные типы синаптических соединений // Нейрофизиология. - 1986. - 18, № 5. - С.621-629. 4. Романенко А.В., Гнатенко В.М., Владимирова И.А.. Действие тиамина на нервно-мышечную передачу в гладких мышцах // Нейрофизиология. 1994. - 26, №6. - С.449-457. 45942 6 5. Вовк А.И., Романенко А.В., Муравьева И.В., Зайцев Л.М. 3-децоксикарбонилметил-4-метил-5β-гидроксиэтилтиазолий хлорид или децоксикарбонилметил-4-метилтиазолий хлорид, угнетающие нервно-мышечную передачу и обладающие транквилизирующей активностью // Авторское свидетельство 1547267 СССР, МКИ4 С07D277/24, А61К31/425. Заявлено 18.07.88. Зарегистрировано 01.11.89. 6. Романенко А.В., Вовк А.И., Шатурский О.Я. Действие тиазолевых аналогов витамина В1 на нервно-мышечную передачу и вызванную αлатротоксином секрецию медиатора в скелетной мышце // Нейрофизиология. - 1995. - 27, № 5/6. - с. 368-374. 7. Романенко А.В., Гнатенко В.М., Владимирова И.А., Вовк А.И. Пре- и постсинаптическая модуляция нервно-мышечной передачи в гладких мышцах тиазолевыми аналогами витамина B1 // Нейрофизиология. - 1995. - 27, №5/6. - С.375-386. 8. Романенко О.В., Вовк А.І., Гіммельрейх Н.Г. Шатурський О.Я. Сполука 3децилоксикарбонілметил-4-метил-5-(2гідроксіетил)тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином В. // Патент на корисну модель №22875 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 квітня 2007р. 8с. 9. Шатурський О.Я. Романенко О.В., Вовк А.І., Гіммельрейх Н.Г. Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином. // Патент на корисну модель №27417 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 жовтня 2007р. - 8с. 10. Борисова Т.О., Крисанова Н.В., Сівко Р.В., Романенко О.В., Вовк А.І. Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрій-залежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів // Патент на корисну модель №38155 зареєстровано в Державному реєстрі патентів України на корисні моделі 25 грудня 2008р. - 8с. 11. Cotman C. W. Isolation of synaptosomal and synaptic plasma membrane fractions // Meth. Enzymol. - 1974. - 31. - P.445-452. 12. Borisova T.A., Krisanova N.V. Presynaptic transporter-mediated release of glutamate evoked by the protonophore FCCP increases under altered gravity conditions // Adv. Space Res. - 2008. - 42, N12. - P.1971-1979. 13. Larson E., Howlett В., Jagendorf A. Artificial reductant enhancement of the Lowry method for protein determination // Anal. Biochem. - 1986. - 155. - P.243-248. 7 Комп’ютерна верстка Л. Ціхановська 45942 8 Підписне Тираж 28 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of compound 3-decyloxycarbonylmethyl-4-methyl-5-(2-hydroxyethyl)thiazolium chloride as compound promoting increase in unstimulated release of glutamate from nerve terminals of rat cerebral cortex
Автори англійськоюBorysova Tetiana Oleksandrivna, Romanenko Oleksandr Viktorovych, Krysanova Natalia Valeriivna, Sivko Roman Vitaliovych, Borysov Arsenii Andriiovych, Vovk Andrii Ivanovych
Назва патенту російськоюПрименение соединения 3-децилоксикарбонилметил-4-метил-5-(2-гидроксиэтил)тиазолий хлорид как соединения, которое вызывает увеличение нестимулированного высвобождения глутамата из нервных окончаний головного мозга крыс
Автори російськоюБорисова Татьяна Александровна, Романенко Александр Викторович, Крысанова Наталия Валериевна, Сивко Роман Витальевич, Борисов Арсений Андреевич, Вовк Андрей Иванович
МПК / Мітки
МПК: C07D 277/00
Мітки: збільшення, глутамату, викликає, вивільнення, яка, головного, хлорид, закінчень, нестимульованого, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, щурів, сполуки, нервових, мозку, застосування
Код посилання
<a href="https://ua.patents.su/4-45942-zastosuvannya-spoluki-3-deciloksikarbonilmetil-4-metil-5-2-gidroksietiltiazolijj-khlorid-yak-spoluki-yaka-viklikaeh-zbilshennya-nestimulovanogo-vivilnennya-glutamatu-z-nervovikh-za.html" target="_blank" rel="follow" title="База патентів України">Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає збільшення нестимульованого вивільнення глутамату з нервових закінчень головного мозку щурів</a>
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів

Номер патенту: 45941
Опубліковано: 25.11.2009
Автори: Касаткіна Людмила Олександрівна, Остапченко Людмила Іванівна, Вовк Андрій Іванович, Крисанова Наталія Валеріївна, Борисова Тетяна Олександрівна, Борисов Арсеній Андрійович, Романенко Олександр Вікторович
МПК: C07D 277/00
Мітки: деполяризацію, головного, закінчень, застосування, щурів, сполуки, мембрани, ізольованих, мозку, плазматичної, яка, нервових, хлорид, викликає, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій
Формула / Реферат:
Застосування сполуки 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид як сполуки, яка викликає деполяризацію плазматичної мембрани ізольованих нервових закінчень головного мозку щурів, загальної формули:.
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрієзалежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів

Номер патенту: 38155
Опубліковано: 25.12.2008
Автори: Крисанова Наталія Валеріївна, Борисова Тетяна Олександрівна, Сівко Роман Віталійович, Вовк Андрій Іванович, Романенко Олександр Вікторович
МПК: C07D 277/00
Мітки: процес, мозку, сполука, накопичення, ізольованими, нервовими, має, закінченнями, головного, яка, щурів, глутамату, натрієзалежного, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, ефект, хлорид, інгібуючий
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має інгібуючий ефект на процес натрієзалежного накопичення глутамату ізольованими нервовими закінченнями головного мозку щурів, загальної формули:.
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином

Номер патенту: 27417
Опубліковано: 25.10.2007
Автори: Шатурський Олег Ярославович, Гіммельрейх Ніна Германівна, Вовк Андрій Іванович, Романенко Олександр Вікторович
МПК: C07D 277/00
Мітки: каналів, іонну, сполука, хлорид, провідність, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій, блокуючий, ефект, має, яка, утворених, ністатином
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил)тіазолій хлорид, яка має блокуючий ефект на іонну провідність каналів, утворених ністатином, загальної формули:.
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил) тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином в

Номер патенту: 22875
Опубліковано: 25.04.2007
Автори: Романенко Олександр Вікторович, Гіммельрейх Ніна Германівна, Вовк Андрій Іванович, Шатурський Олег Ярославович
МПК: A61K 47/12, C07C 255/00
Мітки: амфотерицином, хлорид, 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил, сполука, має, іонну, яка, блокувальний, утворених, тіазолій, провідність, ефект, каналів
Формула / Реферат:
Сполука 3-децилоксикарбонілметил-4-метил-5-(2-гідроксіетил) тіазолій хлорид, яка має блокувальний ефект на іонну провідність каналів, утворених амфотерицином В, загальної формули:.
Бетатирон – інгібітор вивільнення медіатору норадреналіну з адренергічних нервових терміналей

Номер патенту: 41727
Опубліковано: 17.09.2001
Автор: Дужак Володимир Григорович
МПК: A61K 31/155
Мітки: медіатору, бетатирон, інгібітор, вивільнення, нервових, терміналей, адренергічних, норадреналіну
Формула / Реферат:
Застосування бетатирону як інгібітора вивільнення медіатору норадреналіну з адренергічних нервових терміналей.
Випадковий патент: Засіб для лікування запальних захворювань пародонта