Спосіб відтворення гетерозиготних форм вищих рослин
Номер патенту: 82013
Опубліковано: 25.02.2008
Автори: Парій Юлія Олександрівна, Якушко Юрій Євгенович, Спиридонов Владислав Геннадійович, Мельничук Максим Дмитрович, Ситник Катерина Сергіївна, Антіпов Ігор Олександрович, Парій Мирослав Федорович




Формула / Реферат
Спосіб відтворення гетерозиготних форм вищих рослин, що включає отримання гомозиготних ліній із гетерозиготних рослин, схрещування гомозиготних ліній та оцінку отриманих гетерозиготних рослин, розмноження ліній та відтворення гетерозиготної форми шляхом схрещування, який відрізняється тим, що гомозиготні лінії отримують в культурі in vitro, індукуючи соматичну редукцію хромосом в мітотичних поділах, підбирають пари ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сукупності відповідають генотипу вихідної гетерозиготної рослини та будуть відновлювати його при схрещуванні.
Текст
Спосіб відтворення гетерозиготних форм вищи х рослин, що включає отримання гомозиготних ліній із гетерозиготних рослин, схрещування гомозиготних ліній та оцінку отриманих гетерозиготних рослин, розмноження ліній та відтворення гетерозиготної форми шляхом схрещування, який відрізняється тим, що гомозиготні лінії отримують в культурі in vitro, індукуючи соматичну редукцію хромосом в мітотичних поділах, підбирають пари ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сукупності відповідають генотипу вихідної гетерозиготної рослини та будуть відновлювати його при схрещуванні. (19) (21) a200610714 (22) 10.10.2006 (24) 25.02.2008 (72) ПАРІЙ МИРОСЛАВ ФЕДОРОВИЧ, U A, СИТНИК КАТЕРИНА СЕРГІЇВНА, UA, ПАРІЙ ЮЛІЯ ОЛЕКСАНДРІВН А, UA, ЯКУШКО ЮРІЙ ЄВГЕНОВИЧ, UA, АНТІПОВ ІГОР ОЛЕКСАНДРОВИЧ, U A, СПИРИДОНОВ ВЛАДИСЛАВ ГЕННАДІЙОВИЧ, U A, МЕЛЬНИЧУК МАКСИМ ДМИТРОВИЧ, U A (73) НАЦІОНАЛЬНИЙ АГРАРНИЙ УНІВЕРСИТЕТ, UA (56) RU C1 2150821, 20.06.2000. RU C1 2150822, 20.06.2000. RU C1 2035134, 20.05.1995. 3 82013 гомозиготних ліній із гетерозиготних рослин, схрещування гомозиготних ліній та оцінку отриманих гетерозиготних рослин, розмноження ліній та відтворення гетерозиготної форми шляхом схрещування, згідно винаходу гомозиготні лінії отримують шляхом використання культури мікроспор чи макроспор in vitro, або шляхом використання механізмів апоміксису in vivo, попередньо блокуючи обмін ділянками між гомологічними хромосомами у мейозі гетерозиготної рослини, підбирають пари ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сукупності відповідають генотипу вихідної гетерозиготної рослини та будуть відновлювати його при схрещуванні. Запропонований спосіб характеризується такими суттєвими ознаками: отримання гомозиготних ліній із гетерозиготних рослин, - або використовуючи культуру мікроспор чи макроспор in vitro, попередньо блокуючи обмін ділянками між гомологічними хромосомами у мейозі вихідної гетерозиготної рослини, -або використовуючи механізми апоміксису in vi vo, попередньо блокуючи обмін ділянками між гомологічними хромосомами у мейозі вихідної гетерозиготної рослини, підбір пар ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сукупності відповідають генотипу вихідної гетерозиготної рослини та відновлюють його при схрещуванні, схрещування гомозиготних ліній та проведення оцінки отриманих гетерозиготних рослин, розмноження ліній, які є батьківськими формами відібраних на основі оцінки гетерозиготних рослин, відтворення гетерозиготної форми шляхом схрещування. Нові, відмінні від прототипу, ознаки забезпечують відтворення гетерозиготних форм незалежно від способу їх отримання - як тих, що отримані внаслідок схрещування гомозиготних ліній так і тих, які отримані в інший спосіб. Відмінні від прототипу ознаки забезпечують також значне скорочення часу на отримання та відтворення гетерозиготної форми, оскільки відсутня необхідність проводити інбридинг протягом 8-10 поколінь для отримання гомозиготних ліній. Методика культивування мікро- та макроспор в умовах in vitro є одним із сучасних способів вирішення ряду питань практичної селекції, розмноження та підтримання певних рослинних форм [Vratislav Kucera, Miroslava Vyvadilova and Miroslav Klima (2002) Utilisation of doubled haploids in winter oilseed rape (Brassica napus L.) Breeding. Czech J. Genet. Plant Breed., 38: 50-54]. Сутність даного підходу полягає у використанні явища тотіпотентності та високої регенераційної здатності, що притаманні рослинним мікро та макроспорам [Dunwell, J.M. (1985) Anther and ovary culture. In S.W.J. Bright and M.G.K. Jones (ed.) Cereal tissue and cell culture. Matinus Nijhoff/Dr. W. Junk Publishers, Dordrecht, the Netherlands. P. 144]. Мікро та макроспори рослинних організмів утворюються у процесі мейозу і регенеровані з них рослини є гаплоїдними, що відображається на їх 4 фенотипі та фертильності. Для більшості практичних цілей необхідно подвоювати гаплоїдний набір хромосом створюючи дигаплоїдні рослини (Weiguo Liu, Ming Y. Zheng, Enrique A. Polle and Calvin F. Konzak (2002) Highly efficient doubled-haploid production in wheat (Triticum aestivum L.) via Induced microspore embryogenesis. Crop Science, 42: 686-692). Кількість пилкових зерен у пиляку досягає 100000 (). Це дозволяє у способі відтворення гетерозиготних форм із однієї гетерозиготної рослини отримати достатню кількість диплоїдних ліній для подальшого підбору пар ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини. Імовірність підібрати комплементарну лінію у рослин, які мають 10 хромосом у галоїдному наборі, при. блокуванні обміну ділянками між хромосомами, складає 1/2048. Кросинговер - одне із ключових явищ, яке лежить в основі рекомбінації генетичного матеріалу. Це явище включає ряд послідовних процесів: кон'югація хромосом, утворення бівалентів, утворення хіазм, гомологічна рекомбінація. У нормі кросинговер забезпечує рекомбінацію генетичного матеріалу в межах гомологічних хромосом. Репресія експресії генів, які контролюють ці процеси на різних рівнях та активності відповідних ферментів призведе до відсутності обміну ділянками між гомологічними хромосомами. Наприклад, на сьогодні розроблені методи репресії експресії генів [Patent No: US 6,573,099 В2 Genetic construction for delaying or repressing expression of a target gene]. Існують і більш прості шляхи попередження обміну ділянками між гомологічними хромосомами. Так, при пригніченні функціонування клітин тапетуму пшениці, жита, кінських бобів та цибулі 0,04% НСІ спостерігався повний асинапсис хромосом. Індукований асинапсис спостерігали і інші дослідники при дії низькими та високими температурами та хімічними речовинами [цит. по Константинов «Мейоз» Мн., Изд-во. БГУ 1971]. При дії різними фізичними та хімічними факторами на мітоз значно збільшується частота соматичної редукції, наприклад, це спостерігалося при дії розчину хлоралгідрату на соматичні клітини гороху, ефіру на паростки кукурудзи, після опромінення коренів бобів ультракороткими радіохвилями, в корінцях Rhoeo соматична редукція була індукована дією індоліл-3-оцтової кислоти. Зміна температурних умов також може викликати соматичну редукцію [цит. по Константинов «Мейоз» Мн., Изд-во. БГУ 1971]. Це дозволяє отримати дигаплоїдні форми, хромосоми яких успадковані цілісно без змін. Рекомбінація відбувається лише на рівні незалежної сегрегації хромосом. Гомологічні хромосоми диплоїдного організму, за виключенням дигаплоїдних ліній, різняться одна від одної послідовністю нуклеотидів у молекулі ДНК. На сьогодні розроблено велику кількість молекулярно-генетичних методик, які дозволяють розрізнити відмінні за нуклеотидним складом 5 82013 молекули ДНК. Наприклад SSR-PCR придатний для ідентифікації молекули ДНК хромосоми, оскільки ДНК еукаріотів містять прості повтори нуклеотидів, а хромосоми мають поліморфізм за кількість мотивів у повторах (наприклад мотив (GGT) може повторюватись 22 рази в одній хромосомі та 29 разів в гомологічній). Це дозволяє, використавши олігонуклеотидні праймери, ампліфікувати ділянки повторів та методом гель-електрофорезу визначити їх довжину [А.Е. Солоденько, А.В. Саналатий, Ю.М. Сиволап Идентификация генотипов подсолнечника с помощью микросателитных маркеров // Цитология и генетика № 2.- 2004]. Найбільш придатним для ідентифікації гомологічних хромосом є метод SNP (single nucleotide polymorphism) аналізу, що дозволяє ідентифікувати поліморфізм на рівні одного нуклеотиду, який властивий гомологічним молекулам ДНК [Giancola S, McKhann HI, Berard A, Camilleri C, Durand S, Libeau P, Roux F, Reboud X, Gut IG, Brunei D. Utilization of the three highthroughput SNP genotyping methods, the GOOD assay, Amplifluor and TaqMan, in diploid and polyploid plants //Theor Appl Genet. 2006 Apr; 112(6): 1115-24. Epub 2006 Feb 2.]. Методи SSRPCR та/або SNP-аналізу дозволяють розрізнити яка із гомологічних хромосом вихідної гетерозиготної рослини потрапила до конкретної дигаплоїдної рослини і на основі цього підібрати пари ліній, кожна з яких буде нести одн у із гомологічних хромосом вихідного гетерозиготного організму. Винахід здійснюють таким чином. Гетерозиготну рослину кукурудзи, яку необхідно відтворити, вирощують в ґрунті. В період цвітіння кукурудзи волоть занурюють у розчин 0,04% НСІ. Проводять цитологічне дослідження мейозу в пиляках і відмічають ті квітки, де спостерігається асинапсис. Потім волоті з такими квітками збирають на одноядерній або ранній двоядерній стадії формування мікроспор, загортають у вологий паперовий рушник та алюмінієву фольгу і витримують при температурі +4°С протягом 14 днів. Колоски відділяють від волоті та стерилізують 15 хвилин у 2,5% розчині гіпохлориду натрію з додаванням детергенту. Матеріал 3 рази відмивають у стерильній дистильованій воді. Для, виділення мікроспор з пиляків, останні обробляють 0,44Μ розчином сахарози та фільтрують отриману суміш через фільтр з розміром пор 100мкм. Фільтрат центрифугують у градієнті щільності перкола (10, 15 та 30% у 0,44% розчині сахарози) та відбирають життєздатні мікроспори, культивують та отримують дигаплоїдні рослини згідно методик, що описані [Ingrid Ε. Aulinger Combination of in vitro androgenesis and biolistic transformation: an approach for breeding transgenic maize (Zea mays L.) lines. A dissertation for the degree of Doctor of Natural Sciences, Zurich, 2002 P. 32-33) та (Guichang Zhang, Connie M. Williams, Mary K. Campenot, Locksley E. McGann and David D. Cass (1992) Improvement of longevity and viability of 6 sperm cells isolated from pollen of Zea mays L. Plant Physiol. 100, 47-53]. Із отриманих дигаплоїдних рослин та вихідної гетерозиготної рослини екстрагують ДНК. Проводять полімеразну ланцюгову реакцію із олігонуклеотидними праймерами до SSR л оку сів і розподіляють продукти ампліфікації методом гельелектрофорезу. На основі отриманих даних визначають за якими SSR локусами вихідна рослина була гетерозиготною і відбирають пари ліній, які містять різні алелі даного локусу. При цьому SSR локуси повинні бути локалізовані у всіх 10 хромосомах кукурудзи не менше, ніж по 2 локуси на кожному плечі хромосоми. На основі отриманих даних визначають групи зчеплення алелів SSR локусів і відбирають дигаплоїдні рослини, хромосоми яких були сформовані без кросинговеру у мейозі вихідної рослини. Підбирають пари ліній так, щоб кожна з ліній в парі несла одну із гомологічних хромосом вихідного гетерозиготного організму. Рослини адаптують до умов in vivo і пересаджують у грунт. Проводять самозапилення рослин і в наступному поколінні схрещують їх у межах підібраних пар. У наступному поколінні проводять вивчення гібридів та ліній і відбирають ті, які , найбільш придатні для відтворення вихідного гетерозиготного генотипу в польових умовах. Відібрані лінії розмножують в польових умовах до необхідних об'ємів і схрещують на ділянках гібридизації. Отримують гібридне насіння, генотип зародків якого відповідає генотипу вихідної гетерозиготної форми. Або гетерозиготну рослину кукурудзи, яку необхідно відтворити, вирощують в гр унті. В період цвітіння кукурудзи волоть занурюють у розчин 0,04% НСІ. Проводять цитологічне дослідження мейозу в пиляках і відмічають ті квітки, в яких спостерігається асинапсис. Потім із цих квіток збирають пилок, наносять його на попередньо ізольовані жіночі суцвіття рослин кукурудзи, гомозиготних за рецесивним геном ig\ (indeterminate gametophytel), який обумовлює андрогенез та гомозиготні за домінантним геном Вх (benzoxa zinlessl), який обумовлює фіолетове забарвлення сходів FeCl3. Гомозиготність за геном ig\ обумовлює утворення на даній рослині насіння шляхом андрогенезу, а маркерний ген дозволяє розрізнити рослини, сформовані внаслідок нормального статевого розмноження (мають прояв домінантного гена - забарвлюються) і гаплоїдні андрогенні рослини (рослини дикого типу - не забарвлюються). Збирають насіння і пророщують на фільтрувальному папері. Відбирають форми за маркерним геном, і проводять додатково цитологічні дослідження кінчиків коренів для визначення рівня плоїдності рослин. Проводять диплоїдизацію. Із отриманих дигаплоїдних рослин та вихідної гетерозиготної рослини екстрагують ДНК. Проводять полімеразну ланцюгову реакцію із олігонуклеотидними праймерами до SSR локусів і розподіл продуктів ампліфікації методом гель-електрофорезу. На основі отриманих даних визначають за якими SSR локусом вихідна рослина була гетерозиготною і відбирають пари ліній, які містять різні алелі 7 82013 8 даного локусу. При цьому SSR локуси повинні бути локалізовані у всіх 10 хромосомах кукурудзи не менш, ніж по 2 локуси на кожному плечі хромосоми. На основі цих же даних визначають групи зчеплення алелів SSR локусів і відкидають дигаплоїдні рослини, хромосоми яких були сформовані внаслідок кросинговеру у мейозі вихідної рослини. Підбирають пари ліній так, щоб кожна з ліній в парі несла одну із гомологічних хромосом вихідного гетерозиготного організму. Рослини адаптують до умов in vivo і пересаджують у гр унт. Проводять самозапилення рослин і в наступному поколінні схрещують їх у межах підібраних пар. У наступному поколінні проводять вивчення гібридів та ліній і відбирають ті, які найбільш придатні для відтворення вихідного гетерозиготного генотипу в польових умовах. Відібрані лінії розмножують у польових умовах до необхідних об'ємів і схрещують на ділянках гібридизації. Отримують гібридне насіння, генотип зародків якого відповідає генотипу вихідної гетерозиготної форми. Порівняльна характеристика варіантів здійснення винаходу: vi vo, попередньо блокуючи обмін ділянками між гомологічними хромосомами у мейозі гетерозиготної рослини, або індукуючи соматичну редукцію з подальшим підбором пар ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сукупності відповідають генотипу вихідної гетерозиготної рослини та будуть відновлювати його при схрещуванні використовуються для відтворення гетерозиготних форм вищих рослин незалежно від способу їх отримання. Отримання гомозиготних ліній з використанням культури мікроспор чи макроспор in vitro Матеріальні затрати . більші, в зв'язку із використанням культури мікроспор та/або макроспор, однак цей варіант забезпечує таку кількість дигаплоїдів, яка значно збільшує імовірність підбору комплементарних пар дигаплоїдних ліній, кількість отриманих ліній обмежується кількістю пилкових зерен, що продукує рослина і ефективністю методу культури мікро та/або макроспор. Отримання гомозиготних ліній з використанням механізмів апоміксису in vivo Порівняльний аналіз заявленого винаходу з прототипом показав, що заявлений спосіб характеризується новими суттєвими ознаками, які полягають у тому, що отримують гомозиготні лінії: -або використовуючи культур у мікроспор чи макроспор in vitro, -або використовуючи механізми апоміксису in vivo, попередньо блокуючи обмін ділянками між гомологічними хромосомами у мейозі вихідної гетерозиготної рослини, з отриманих гомозиготних ліній підбирають пари ліній так, що кожна з ліній в парі несе одну із гомологічних хромосом вихідної гетерозиготної рослини, а їх генотипи у сук упності відповідають генотипу ви хідної гетерозиготної рослини і відновлюють його при схрещуванні. Заявлений винахід забезпечує відтворення гетерозиготних форм незалежно від способу їх отримання - як тих, що отримані внаслідок схрещування гомозиготних ліній так і тих, які отримані в інший спосіб. Досягнення такого технічного результату не випливає явним чином із рівня розвитку те хніки. В загально доступних джерелах відсутні відомості про те, що гомозиготні лінії із гетерозиготної рослини, які отримують або шляхом використання культури мікроспор чи макроспор in vitro, або шляхом використання механізмів апоміксису in Матеріальні затрати значно менші, у зв'язку Із відсутністю використання методів культури in vitro, однак цей варіант здійснення винаходу може бути застосований тільки для культур, у яких вдалося індукувати андрогенез in vivo. Кількість отриманих ліній обмежується кількістю пилкових зерен, що продукує рослина і ефективністю індукованого андрогенезу.
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for reproducing heterozygous forms of higher plants
Автори англійськоюParii Myroslav Fedorovych, Sytnyk Kateryna Serhiivna, Parii Yulia Oleksandrivna, Yakushko Yurii Yevhenovych, Antipov Ihor Oleksandrovych, Spyrydonov Vladyslav Henndiiovych, Spyrydonov Vladyslav Hennadiiovych, Melnychuk Maksym Dmytrovych
Назва патенту російськоюСпособ воспроизведения гетерозиготных форм высших растений
Автори російськоюПарий Мирослав Федорович, Сытник Екатерина Сергеевна, Парий Юлия Александровна, Якушко Юрий Евгеньевич, Антипов Игорь Александрович, Спиридонов Владислав Геннадиевич, Спиридонов Владислав Геннадьевич, Мельничук Максим Дмитриевич
МПК / Мітки
МПК: A01H 1/04
Мітки: рослин, відтворення, гетерозиготних, спосіб, вищих, форм
Код посилання
<a href="https://ua.patents.su/4-82013-sposib-vidtvorennya-geterozigotnikh-form-vishhikh-roslin.html" target="_blank" rel="follow" title="База патентів України">Спосіб відтворення гетерозиготних форм вищих рослин</a>
Спосіб визначення толерантності вищих рослин до техногенного хімічного забруднення навколишнього середовища

Номер патенту: 21281
Опубліковано: 15.03.2007
Автори: Балічиєва Ділярам Валіївна, Ібрагімова Евеліна Енверівна
МПК: G01N 1/00, G01N 17/00
Мітки: середовища, хімічного, спосіб, забруднення, навколишнього, толерантності, рослин, вищих, визначення, техногенного
Формула / Реферат:
Спосіб визначення толерантності вищих рослин до техногенного хімічного забруднення, що включає збирання репродуктивного матеріалу в екологічно сприятливій і техногенній зонах, фіксацію репродуктивного матеріалу в оцтовокислому алкоголі (3:1) при 12-ти годинній експозиції, промивання його різними концентраціями спиртів, збереження в 80 % етиловому спирті в холодильнику, готування тимчасових давлених препаратів, підрахунок кількості фертильних...
Спосіб отримання рослин винограду від вихідних форм з низькою фертильністю

Номер патенту: 14365
Опубліковано: 15.05.2006
Автори: Павлова Ірина Олександрівна, Клименко Віктор Павлович
Мітки: рослин, фертильністю, спосіб, вихідних, отримання, низькою, винограду, форм
Формула / Реферат:
Спосіб отримання рослин винограду від вихідних форм з низькою фертильністю, що включає культивування зародків з частиною оболонки насіння in vitro з висаджуванням їх на живильне середовище з додаванням регуляторів росту, який відрізняється тим, що спочатку диференціюють вихідні форми за класом безнасінності і категорією експресії в культурі насіння, культивування in vitro здійснюють в дві або три стадії: пророщення насіння, формування рослин...
Споруда для очищення стічних вод з використанням вищих водяних рослин

Номер патенту: 3377
Опубліковано: 15.11.2004
Автори: Шаповал Олександр Євгенович, Кравець Валентин Васильович, Грищенко Наталія Валентинівна, Петрук Володимир Володимирович, Прушанський Максим Йосипович, Маслов Володимир Михайлович, Лівшиць Марк Ноєвич
Мітки: вищих, очищення, водяних, споруда, використанням, стічних, рослин, вод
Формула / Реферат:
1. Споруда для очищення стічних вод з використанням вищих водяних рослин, що складається з послідовно з'єднаних між собою біоставків, в яких утримуються на плаву ділянки вищих водяних рослин, яка відрізняється тим, що споруда додатково має біоставок для первинного відстоювання стічної води, поміж біоставками з ділянками рослин на плаву розміщений біоставок з ділянкою вищих водяних рослин, висаджених на біоплато, що складається з шару піску та...
Спосіб створення чоловічостерильних форм для одержання гібридного насіння буряків

Номер патенту: 37708
Опубліковано: 15.05.2001
Автори: Парій Федір Микитович, Мельничук Юрій Вікторович, Малієнко Вадим Анатолійович
МПК: A01H 1/04
Мітки: чоловічостерильних, спосіб, насіння, створення, гібридного, форм, одержання, буряків
Текст:
...рослин використовують закріплювачі стерильності П-типу на S-плазмі (ідиотип S xx ZZ Rfpar Rfpar). Винахід характеризується такими суттєвими ознаками: розмноження ЧС-рослин за допомогою закріплювачів стерильності 0-типу, схрещування з закріплювачем П-типу на S-плазмі (ідиотип S xx ZZ Rfpar Rfpar), використання одержаних ЧС-форм як материнського компоненту гібриду. Із цих суттєвих ознак використання при створенні ЧС-форм...
Спосіб підвищення виходу гаплоїдних рослин цукрових буряків
Номер патенту: 24323
Опубліковано: 25.06.2007
Автори: Парій Федір Микитович, Рябовол Людмила Олегівна
МПК: A01H 1/00
Мітки: рослин, підвищення, буряків, виходу, цукрових, гаплоїдних, спосіб
Формула / Реферат:
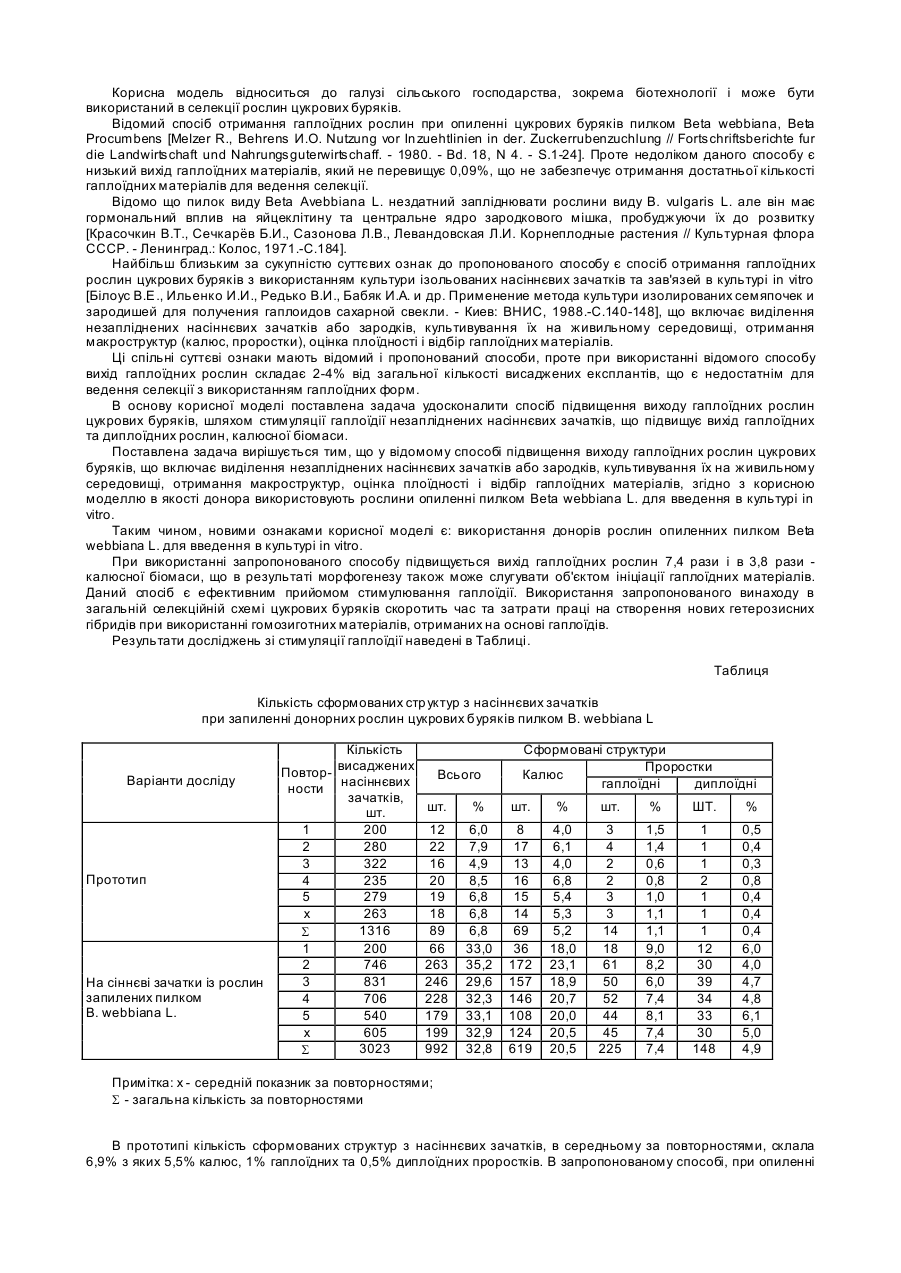
Спосіб підвищення виходу гаплоїдних рослин цукрових буряків, що включає виділення незапліднених насіннєвих зачатків або зародків, культивування їх на живильному середовищі, отримання макроструктур, оцінку плоїдності і відбір гаплоїдних матеріалів, який відрізняється тим, що як донор використовують рослини, обпилені пилком Beta webbiana L., для введення в культуру in vitro.
Попередній патент: Радіолокаційна система
Наступний патент: Пристрій для транспортування та очистки коренебульбоплодів
Випадковий патент: Пристрій для контролю та реконфігурації резервованої системи