Штам escherichia coli bl21(spatr) – продуцент рекомбінантного білка spaa бактерії erysipelothrix rнusiopatнiаe
Номер патенту: 65123
Опубліковано: 25.11.2011
Автори: Ображей Анатолій Федорович, Гільчук Павло Володимирович, Дерябін Олег Миколайович, Дерябіна Олена Григорівна, Скрипник Валерій Григорович, Тарасов Олександр Анатолійович




Формула / Реферат
Штам Escherichia coli BL21(SPAtr) - продуцент рекомбінантного білка SpaA з М.м. 47 кДа бактерії Erysipelothrix rhusiopathie, одержаний в результаті трансформації клітин Escherichia coli штаму BL21(DE3) рекомбінантною плазмідною ДНК pSPAA(tr), яка має довжину 6374 п.н. та містить наступні фрагменти:
фрагмент ДНК довжиною 1085 п.н., який є фрагментом гена SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae та має послідовність:
,
регуляторні елементи експресії цільового білка - Т7/Lас-промотор та Т7-термінатор загальною довжиною 62 п.н.;
оrі - область початку реплікації плазмід СоlЕ1-типу;
селективний маркер - ген стійкості до канаміцину Kan довжиною 812 п.н.;
оrі fl - область початку реплікації бактеріофага fl довжиною 455 п.н.,
використання штаму Escherichia coli BL21(SPAtr), забезпечує синтез рекомбінантного білка SPAA до 1,5 г і більше з 1 дм3 бактеріальної культури.
Текст
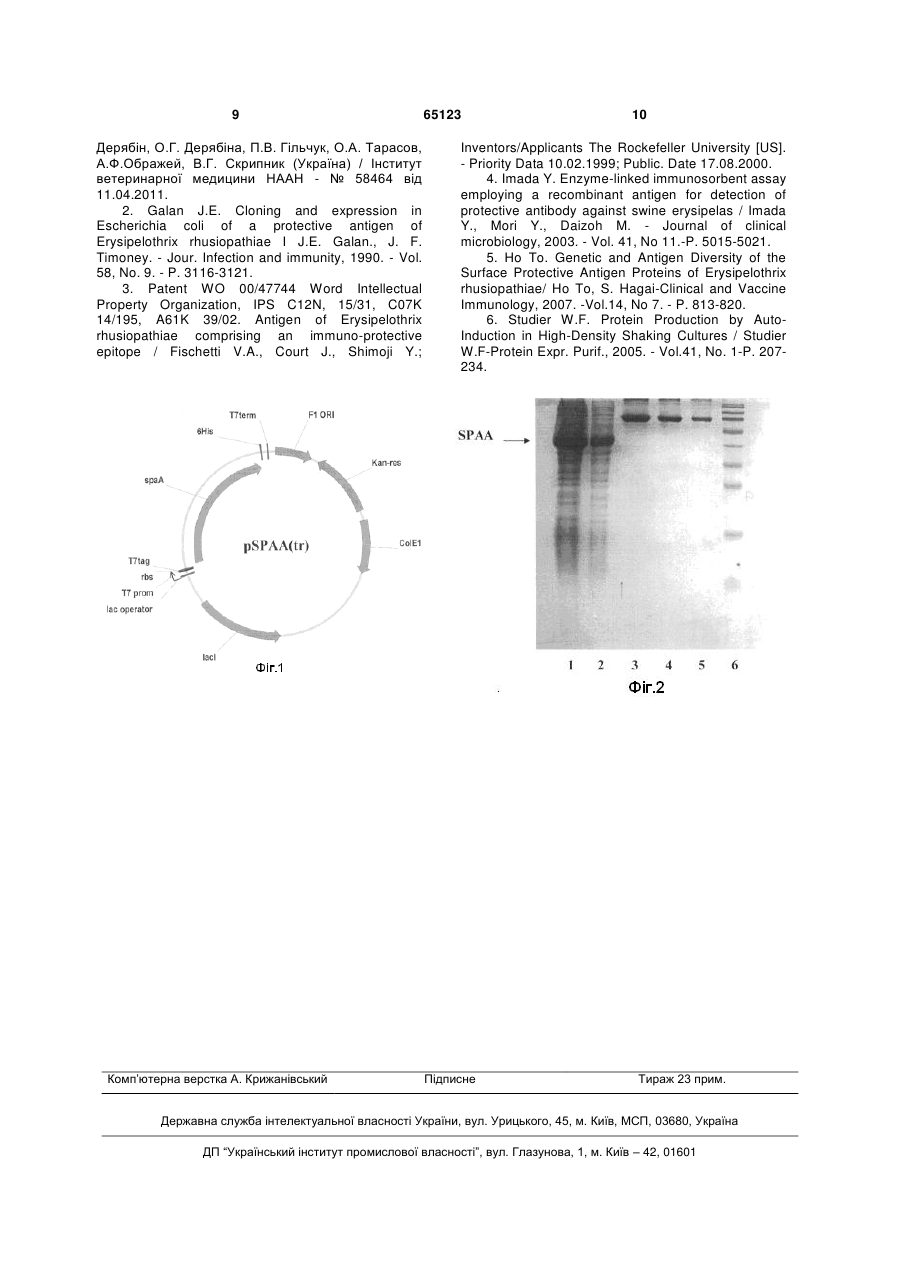
Штам Escherichia coli BL21(SPAtr) - продуцент рекомбінантного білка SpaA з М.м. 47 кДа бактерії Erysipelothrix rhusiopathiae, одержаний в результаті трансформації клітин Escherichia coli штаму BL21(DE3) рекомбінантною плазмідною ДНК pSPAA(tr), яка має довжину 6374 п.н. та містить наступні фрагменти: фрагмент ДНК довжиною 1085 п.н., який є фрагментом гена SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae та має послідовність: регуляторні елементи експресії цільового білка Т7/Lас-промотор та Т7-термінатор загальною довжиною 62 п.н.; оrі - область початку реплікації плазмід СоlЕ1типу; селективний маркер - ген стійкості до канаміцину Kan довжиною 812 п.н.; оrі fl - область початку реплікації бактеріофага fl довжиною 455 п.н., використання штаму Escherichia coli BL21(SPAtr), забезпечує синтез рекомбінантного білка SPAA до 3 1,5 г і більше з 1 дм бактеріальної культури. Корисна модель належить до біотехнології, зокрема до генетичної інженерії, та може використовуватися для одержання рекомбінантного поверхневого антигену SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae, придатного для виробництва вакцин ветеринарного призначення проти бешихи свиней. Так, корисна модель стосується штаму Escherichia coli B121(DE3), який є трансформованим рекомбінантною плазмідною ДНК pSPAA(tr), яка містить N-кінцевий фрагмент гена SPAA Erysipelothrix rhusiopathiae, та представляє собою продуцент рекомбінантного білка SPAA з (19) UA (11) 65123 (13) U (21) u201106054 (22) 16.05.2011 (24) 25.11.2011 (46) 25.11.2011, Бюл.№ 22, 2011 р. (72) ДЕРЯБІН ОЛЕГ МИКОЛАЙОВИЧ, ДЕРЯБІНА ОЛЕНА ГРИГОРІВНА, ГІЛЬЧУК ПАВЛО ВОЛОДИМИРОВИЧ, ТАРАСОВ ОЛЕКСАНДР АНАТОЛІЙОВИЧ, ОБРАЖЕЙ АНАТОЛІЙ ФЕДОРОВИЧ, СКРИПНИК ВАЛЕРІЙ ГРИГОРОВИЧ (73) ІНСТИТУТ ВЕТЕРИНАРНОЇ МЕДИЦИНИ 3 М.м. 47 кДа грампозитивної бактерії Erysipelothrix rhusiopathiae. Бактерія Erysipelothrix rhusiopathiae належить до родини Corynebacteriаceаe і є збудником хвороби - бешихи свиней (Erysipelas suum), яке характеризується запальною еритемою шкіри та септичними явищами, а також ураженням суглобів, верукозним ендокардитом та некротичними ураженням шкіри. Збудник має високу стійкість у зовнішньому середовищі. Джерелом і резервуаром інфекції служать багато видів тварин (свині, вівці, велика рогата худоба, собаки, кури, качки, гризуни, риби, раки й ін.). Основне джерело інфекції свині, у яких захворювання перебігає найчастіше в гострій формі, та гризуни. Основним засобом боротьби з даним захворюванням є імунопрофілактика із застосуванням вакцин на основі живих авірулентних штамів збудника або інактивованих вакцин на основі бактеринів чи лізатів патогенних штамів. Загальним недоліком використання вказаних типів вакцин є необхідність в культивуванні великих об'ємів бактерій Erysipelothrix rhusiopathiae, що може мати негативний вплив на оточуюче середовище, а також створювати загрозу інфікування персоналу. У зв'язку з цим, на сьогоднішній день більш перспективним є отримання рекомбінантного поверхневого антигену SPAA Erysipelothrix rhusiopathiae шляхом мікробіологічного синтезу з використанням штаму Escherichia coli, трансформованого рекомбінантною плазмідною ДНК, що несе ген SPAA Erysipelothrix rhusiopathiae. Такий підхід забезпечує можливість синтезу цільового продукту в значно більшій кількості з використанням недорогої сировини. На сьогодні описано декілька варіантів отримання рекомбінантного білка SPAA Erysipelothrix rhusiopathiae в Escherichia coli з використанням рекомбінантних плазмід, придатного як для використання як антигену для імуноферментного аналізу, так і як компонента субодиничної вакцини. Так, в роботі Galan Е. та Timoney J. [2] описаний спосіб поєднання фрагментів геномної ДНК Erysipelothrix rhusiopathiae, отриманої в результаті часткової переварки ферментом EcoR1, з фрагментами ДНК фага gt11 із подальшим зараженням рекомбінантними фагами культури Escherichia coli і отримання в ній рекомбінантного білка. Недоліком даного методу є його складність і неможливість стандартного відтворення. Існує опис конструкції для отримання рекомбінантного поверхневого антигену Erysipelothrix rhusiopathiae в патенті Fischetti V.A. та ін. [3], які містять різні фрагменти гена SPAA Erysipelothrix 65123 4 rhusiopathiae, отримані в ПЛР та клоновані в вектор серії pQE (Qiagen). Важливим недоліком цієї конструкції є можливість отримання в рекомбінантній формі лише С-термінальної частини білка SPAA Erysipelothrix rhusiopathiae, в інших випадках спостерігалася деградація продукту. Найбільш близьким продуцентом протективного рекомбінантного білка SPAA Erysipelothrix rhusiopathiae до того, що заявляється, є продуцент, описаний Imada Y. et.al [4] та, з незначними модифікаціями, - Но То та Shinya Nagai [5]. Вектором для клонування і експресії рекомбінантних білків була плазміда pQE32 (Qiagen), яка містить сильний промотор фага Т5. Фрагменти гена SPAA Erysipelothrix rhusiopathiae різної довжини отримували в полімеразній ланцюговій реакції зі специфічними олігонуклеотидними праймерами, після чого клонували в плазміду pQE32 і отриманою рекомбінантною плазмідою трансфікували клітини Е. coli. Індукцію синтезу виконували ізопропіл--D-тіогалактопіранозидом (1ПТГ), отримані тільця включення денатурували за допомогою 6М гуанідинхлориду та очищали методом афінної хроматографії. Недоліком описаної системи є використання недостатньо ефективного промотору для даного антигену, що не забезпечує синтез продукту в препаративній кількості; необхідність використання для індукції синтезу рекомбінантного білка ІПТГ, який є токсичною речовиною для клітин Escherichia coli. До недоліків слід віднести і те, що достатня антигенна активність рекомбінантного білка спостерігалась лише при використанні конструкцій, які відповідали повному гену SPAA або його значній частині. Задачею корисної моделі є створення штамупродуценту, який забезпечує високий рівень експресії білка SPAA Erysipelothrix rhusiopathiae. Вказана задача вирішується за рахунок створення штаму Escherichia coli B121(SPAtr), який несе рекомбінантну плазмідну ДНК [1] та забезпечує продукцію білка SPAA бактерії Erysipelothrix rhusiopathiae на високому рівні. Заявлений штам Escherichia coli B121(SPAtr), який являє собою продуцент рекомбінантного білка SPAA Erysipelothrix rhusiopathiae, отримують шляхом трансформації плазмідною ДНК pSPAA(tr) клітин Escherichia coli B121(DE3). При цьому рекомбінантна плазмідна ДНК pSPAA(tr), що кодує синтез рекомбінантного білка SPAA з М.м. 47 кДа бактерії Erysipelothrix rhusiopathie, має довжину 6374 п.н. та містить наступні фрагменти: фрагмент ДНК довжиною 1085 п.н., який є фрагментом гена SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae та має послідовність: 5 - регуляторні елементи експресії цільового білка - Т7/Lас-промотор та Т7-термінатор загальною довжиною 62 п.н.; - оrі - область початку реплікації плазмід СоlEІтипу; - селективний маркер - ген стійкості до канаміцину Kan довжиною 812 п.н.; - оrі fl - область початку реплікації бактеріофага fl довжиною 455 п.н., Використання штаму Escherichia соlі BL21(SPAtr), що пропонується, забезпечує вихід рекомбінантного білка SPAA до 1,5 г і більше з 1 3 дм бактеріальної культури. Штам-продуцент Escherichia coli BL21(SPAtr) характеризується наступними культуральноморфологічними характеристиками: 1. Клітини дрібні, потовщені, паличковидної форми, грамнегативні, не спороносні. 2. Клітини добре ростуть на простих поживних середовищах. На агаризованому середовищі утворюють колонії круглі, гладкі притиснуті, каламутні, блискучі, сірі, з рівними краями. 3. Клітини добре ростуть при температурі 440 °C з оптимальним значенням pH поживного середовища 6,8-7,5. 4. Клітини проявляють стійкість до канаміцину 3 в концентрації 100 мкг/см . 5. Особливою властивістю клітин є здатність до синтезу поліпептиду з амінокислотною послідовністю, що відповідає поверхневому антигену SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae. Рекомбінантний білок SPAA під час ферментації продуцента Escherichia coli BL21(SPAtr) накопичується в клітинах у водонерозчинній формі в вигляді "тілець включення". Ця особливість має ряд переваг, зокрема, рекомбінантний білок легко виділяється з клітин у вигляді нерозчинного осаду, після чого очищається від більшості білків бактерії шляхом відмивання буферним розчином з додаванням детергентів. Залишок розчиняють у концентрованому розчині хаотропних агентів та піддають рефолдингу (ренатурації) з наступним хроматографічним очищенням. Опис корисної моделі супроводжується наступними фігурами: 65123 6 , Фіг.1 Фізична карта рекомбінантної плазміни pSPAA(tr). Фіг. 2 Ефективність експресії рекомбінантного білка SPAA Erysipelothrix rhusiopathiae клітинами продуцента Escherichia coli BL21(SPAtr). Електрофореграма лізатів клітин Е. coli, в яких індукували синтез SPAA з М.м. 47 кДа (за протоколом аутоін3 дукції, об'єм 0,1 см ). Варіанти: 1-2 - лізати клітин 3 3 (нанесено 0,01 см і 0,002 см суспензії бактеріальних клітин); 3-5 - БСА (нанесено - 3, 1,5, 0,75 мкг); 6 - білки-маркери молекулярної маси. Корисна модель ілюструється наведеними нижче прикладами, які стосуються процедури проведення трансформації клітин Escherichia coli рекомбінантною плазмідою pSPAA(tr) та одержання рекомбінантного білка SPAA Erysipelothrix rhusiopathiae при культивуванні клітин Escherichia coli штаму BL21(SPAtr). Приклад 1. Трансформація клітин Escherichia coli BL-21(DE3) рекомбінантною плазмідою pSPAA(tr) Для трансформації клітин Escherichia coli штаму BL-21(DE3) використовують плазміду pSPAA(tr), яку попередньо було виділено з клітин Escherichia coli штам XL-1 після їх трансформації лігазною сумішшю. Виділення плазміди проводили методом лужного лізису. Для цього колонію клітин Escherichia coli штам XL-1 з агаризованого селективного середовища висівають у рідке селективне середовище - бульйон LB. Склад рідкого середовища (на 1 3 дм ): бакто-триптон - 10 г, бакто-дріжджовий екстракт -5 г, хлорид натрію -10 г, канаміцин - 50 мг, 3 вода - до 1 дм , pH 7,0. Колонію переносили мік3 робіологічною петлею в 0,003 см рідкого стерильного середовища та проводили інкубацію при температурі 37 °C протягом 18-24 год. при 100 об./хв 3 0,0015 см суспензії клітин центрифугували, осад 3 ресуспендували в 0,2 см розчину для лізису № 1, що має наступний склад: глюкоза - 50 мМ, ТрисНСl-25 мМ (pH 8,0), ЕДТА - 10 мМ. Суспензію клітини інкубували протягом 5 хв при кімнатній тем3 пературі, після чого додавали 0,4 см розчину для лізису №2, що має наступний склад: гідроксид натрію - 0,2 М, SDS-1 %, обережно ресуспендували, 3 додавали 0,1 см розчину 10 М амонію ацетату (pH 7 8,0), витримували протягом 5 хв на крижаній бані та центрифугували протягом 5 хв в режимі 10-14 3 тис. об./хв при температурі 0 °C. До 0,2 см надо3 саду додавали 0,4 см охолодженого етилового спирту, суміш перемішували, витримували протягом 10 хв при температурі мінус 20 °C та центрифугували протягом 5 хв в режимі 10-14 тис. об./хв при температурі 0 °C. Надосад відкидали, а осад, що являє собою суміш плазмідної ДНК та клітинної 3 РНК, розчиняли у 0,1 см води та використовували для трансформації клітин Escherichia coli штаму BL-21(DE3). Для трансформації клітин Escherichia coli штаму BL-21(DE3) колонію культури, отриману на агаризованому поживному середовищі, переносили в 100 мл рідкого поживного середовища, що має наступний склад (на 1 дм): 20 г пептону, 5,5 г бакто-дріжджового екстракту, 0,01 см розчину 1 М 3 хлориду калію, 0,002 см розчину 5 М хлориду натрію. Культуру клітин вирощували до ОГ=0,6 (довжина хвилі 600 нм). Для отримання компетентних клітин суспензію бактеріальної культури центрифугували протягом 5 хв. в режимі 6 тис об./хв. при 3 температурі 4 °C та ресуспендували в 0,015 см охолодженого до 0 °C розчину 100 мМ хлориду кальцію. Суспензію витримували протягом 30 хв на крижаній бані та повторно центрифугували в 3 аналогічних умовах, ресуспендували в 0,003 см 3 охолодженого до 0 °C розчину 0,1 см мМ хлориду кальцію, отримуючи, таким чином, компетентні 3 клітини. До 0,1 см суспензії компетентних клітин 3 додавали 0,001 см розчину ДНК плазміди pSPAA(tr) та клітинної РИК у воді та інкубували в наступному режимі: протягом 20 хв. на крижаній бані, 30 сек - при температурі 42 °C, 30 сек - на крижаній бані. 3 Після цього до клітин додавали 0,9 см рідкого середовища LB і суспензію клітин в рідкому середовищі інкубували протягом 60 хв з покачуванням при температурі 37 °C та наносили на агаризоване середовище LB на чашці Петрі. Чашки інкубували протягом 18-24 год. при температурі 37 °C. Окремі колонії, утворені на чашках, після тестування на ефективність експресії цільового білка, використовували як продуцент Escherichia coli штам BL21(SPAtr) для отримання рекомбінантного білка SPAA Erysipelothrix rhusiopathiae. Приклад 2. Експресія рекомбінантного білка SPAA Erysipelothrix rhusiopathiae клітинами Escherichia coli штаму BL21(SPAtr). Отримані у Прикладі 1 клітини Escherichia coli штаму BL-21(DE3), трансформовані плазмідою pSPAA(tr), піддавали культивуванню в середовищі з аутоіндукцією [6] з метою визначення найбільш придатного клону для синтезу цільового білка. Для цього 20 різних колоній трансформантів окремо 3 мікробіологічною петлею переносили в 0,1 см рідкого селективного середовища LB, яке має на3 ступний склад (на 1 дм ): бакто-триптон - 10 г, бакто-дріжджовий екстракт - 5 г, хлорид натрію - 10 г, 3 канаміцин - 50 мг, вода - до 1 дм , інкубували при температурі 37 °C протягом 18-24 год. при 100 об./хв. До бактеріальної суспензії кожного з трансформантів додавали гліцерин до 40 % за об'ємом 65123 8 та консервували в спеціальних кріопробірках в 3 об'ємі по 0,1 см шляхом заморожування при температурі мінус 70 °C, отримуючи, таким чином, головний банк культури клітин-продуцента. Головний банк в результаті повинен складатися не менше ніж з 20 кріопробірок, отриманих з окремої колонії трансформантів. Бактеріальні культури головного банку окремого трансформанту, розморожували та відновлювали їх метаболізм шляхом трьох послідовних пасажів на агаризованому селективному середовищі. Окрему колонію з третього пасажу переносили 3 в 0,003 см рідкого селективного середовища з 2 % глюкози та вирощували при температурі 37 °C протягом 18-24 год. при 100 об./хв до оптичної густини бактеріальної суспензії А=0,6 (при довжині хвилі 600 нм). До цього моменту вже готувалось середовище для аутоіндукції - середовище LB з антибіотиком та додаванням розчинів NPS (20X, (1 М Na2HPO4, 1 М КН2РО4, 0,5 M(NH4)2SO4), 5052 (50Х, 25 % glycerol, 2,5 % glucose, 10 % a-lactose monohydrate та 1М MgSO4 (відповідно 1/20, 1/50 і 1/1000 об'єму). Культуру з потрібною ОГ інокулювали в це середовище з розрахунку 1/1000 та культивували до ОГ=4-7 (=600). Культивування тривало при 37 °C 16-18 годин і закінчувалося через годину після припинення зростання ОГ при =600. Відбирали аліквоти бактеріальної суспензії та визначали рівень експресії цільового білка в клітинах бактерій методом електрофорезу в поліакриламідному гелі з додецилсульфатом натрію. Для цього до 0,001 см суспензії додавали 1/4 об'єму денатуруючого буфера наступного складу: Трис-НС1-200 мМ (pH 6,8), 2--меркаптоетанол - 400 мМ, SDS-4 %, бромфеноловий синій - 0,01 %, гліцерин - 40 %, інкубували на киплячій водній бані протягом 5 хв та електрофоретично розділяли в двоступеневій буферній системі в 12 %-вому акриламідному гелі. Виявлення білків в гелі проводили методом фарбування Кумассі G-250. Визначення молекулярної маси білків та їх процентного вмісту проводили денситометрично методом внутрішньої нормалізації. Відповідно до результатів обробки електрофореграм найбільш ефективний штам цільового білка, отриманий з єдиної колонії трансформантів, залишали в головному банку, використовуючи його для отримання робочого банку клітин продуцента, в той час як інші - менш ефективні - видаляли. Таким чином, заявлена корисна модель дозволяє одержати при культивуванні клітинпродуцента Escherichia coli B121(SPAtr) експресію рекомбінантного білка SPAA Erysipelothrix rhusiopathiae з М.м. 47 к Да - не менше ніж 1,5 3 г/дм бактеріальної суспензії. Рекомбінантний білок SPAA з М.м. 47 кДа придатний для створення на його основі ветеринарних препаратів. Джерела інформації: 1. Деклараційний патент України на корисну модель. Україна, МПК А61К39/245 (2006.01). "Рекомбінантна плазмідна ДНК pSPAA(tr), що кодує послідовність гена поверхневого антигену SPAA ERYSIPELOTHRIX RHUSIOPATHIAE". 2011 О.М. 9 Дерябін, О.Г. Дерябіна, П.В. Гільчук, О.А. Тарасов, А.Ф.Ображей, В.Г. Скрипник (Україна) / Інститут ветеринарної медицини НААН - № 58464 від 11.04.2011. 2. Galan J.E. Cloning and expression in Escherichia coli of a protective antigen of Erysipelothrix rhusiopathiae I J.E. Galan., J. F. Timoney. - Jour. Infection and immunity, 1990. - Vol. 58, No. 9. - P. 3116-3121. 3. Patent WO 00/47744 Word Intellectual Property Organization, IPS C12N, 15/31, C07K 14/195, A61K 39/02. Antigen of Erysipelothrix rhusiopathiae comprising an immuno-protective epitope / Fischetti V.A., Court J., Shimoji Y.; Комп’ютерна верстка А. Крижанівський 65123 10 Inventors/Applicants The Rockefeller University [US]. - Priority Data 10.02.1999; Public. Date 17.08.2000. 4. Imada Y. Enzyme-linked immunosorbent assay employing a recombinant antigen for detection of protective antibody against swine erysipelas / Imada Y., Mori Y., Daizoh M. - Journal of clinical microbiology, 2003. - Vol. 41, No 11.-P. 5015-5021. 5. Ho To. Genetic and Antigen Diversity of the Surface Protective Antigen Proteins of Erysipelothrix rhusiopathiae/ Ho To, S. Hagai-Clinical and Vaccine Immunology, 2007. -Vol.14, No 7. - P. 813-820. 6. Studier W.F. Protein Production by AutoInduction in High-Density Shaking Cultures / Studier W.F-Protein Expr. Purif., 2005. - Vol.41, No. 1-P. 207234. Підписне Тираж 23 прим. Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюEscherichia coli bl21(spatr) strain producing recombinant spaa bacterial protein of erysipelothrix rhusiopathie
Автори англійськоюDeriabin Oleh Mykolaiovych, Deriabina Olena Hryhorivna, Hilchyk Pavlo Volodymyrovych, Tarasov Oleksandr Anatoliiovych, Obrazhei Anatoliy Fedorovych, Skrypnyk Valerii Hryhorovych
Назва патенту російськоюШтамм escherichia coli bl21(spatr) - продуцент рекомбинантного белка spaa бактерии erysipelothrix rhusiopathie
Автори російськоюДерябин Олег Николаевич, Дерябина Елена Григорьевна, Гильчук Павел Владимирович, Тарасов Александр Анатольевич, Ображей Анатолий Федорович, Скрипник Валерий Григорьевич
МПК / Мітки
МПК: A61K 39/27
Мітки: бактерії, bl21(spatr, erysipelothrix, рекомбінантного, штам, білка, rнusiopatнiаe, продуцент, escherichia
Код посилання
<a href="https://ua.patents.su/5-65123-shtam-escherichia-coli-bl21spatr-producent-rekombinantnogo-bilka-spaa-bakteri-erysipelothrix-rnusiopatniae.html" target="_blank" rel="follow" title="База патентів України">Штам escherichia coli bl21(spatr) – продуцент рекомбінантного білка spaa бактерії erysipelothrix rнusiopatнiаe</a>
Штам escherichia coli bl21 (inf-a(2b)) – продуцент рекомбінантного людського лейкоцитарного інтерферону альфа-2b

Номер патенту: 35669
Опубліковано: 25.09.2008
Автори: Скринник Максим Михайлович, Самойленко Вадим Анатолійович, Куркіна Оксана Вікторівна, Дерябін Олег Миколайович
МПК: C12P 21/02, A61K 38/19, C12N 15/19
Мітки: escherichia, inf-a(2b, продуцент, людського, лейкоцитарного, рекомбінантного, штам, альфа-2b, інтерферону
Формула / Реферат:
Штам Е. соlі BL21(INF-a(2b)) - продуцент рекомбінантного людського лейкоцитарного інтерферону альфа-2b, одержаний в результаті трансформації клітин Е. соlі штаму BL21(DE3) рекомбінантною плазмідною ДНК pSSK-INF-a(2b), яка має довжину 5814 п.н. та містить наступні фрагменти:фрагмент ДНК довжиною 501 п.н., який є модифікованим геном людського інтерферону альфа-2b та має послідовність:регуляторні елементи експресії цільового...
Штам escherichia coli к12 “inv” sbb – продуцент рекомбінантної неактивної субодиниці в дифтерійного токсину corynebacterium diphtheriae

Номер патенту: 22160
Опубліковано: 10.04.2007
Автори: Олійник Олена Сергіївна, Романюк Світлана Іванівна, Колибо Денис Володимирович, Кабернюк Андрій Аркадійович, Буркальова Дар''я Олександрівна, Редчук Тарас Анатолійович, Комісаренко Сергій Васильович
МПК: A61K 47/48, C07K 7/08, C12N 15/11, A61K 39/44
Мітки: субодиниці, токсину, рекомбінантної, штам, неактивної, diphtheriae, продуцент, inv, дифтерійного, corynebacterium, escherichia
Формула / Реферат:
Штам Escherichia coli K12 “inv” sbB як продуцент рекомбінантної неактивної субодиниці В дифтерійного токсину Corynebacterium diphtheriae, гібридизованої з полігістидиновим тагом.
Рекомбінантна плазмідна днк рspaa(tr), що кодує послідовність гена поверхневого антигену spaa erysipelothrix rhusiopathie

Номер патенту: 58464
Опубліковано: 11.04.2011
Автори: Тарасов Олександр Анатолійович, Скрипник Валерій Григорович, Ображей Анатолій Федорович, Дерябін Олег Миколайович, Гільчук Павло Володимирович, Дерябіна Олена Григорівна
МПК: A61K 39/245
Мітки: днк, рspaa(tr, erysipelothrix, кодує, плазмідна, гена, послідовність, антигену, поверхневого, рекомбінантна, rhusiopathie
Формула / Реферат:
Рекомбінантна плазмідна ДНК pSPAA(tr), що кодує послідовність гена поверхневого антигену SPAA Erysipelothrix rhusiopathiae, яка має довжину 6374 п.н. та містить наступні фрагменти:фрагмент ДНК довжиною 1085 п.н., який є фрагментом гена SPAA грампозитивної бактерії Erysipelothrix rhusiopathiae та має послідовність:- регуляторні елементи експресії цільового білка - Т7/lас-промотор та Т7-термінатор загальною довжиною 62...
Штам escherichia coli к 12 “inv” sbа – продуцент рекомбінантної неактивної субодиниці а дифтерійного токсину corynebacterium diphtheriae

Номер патенту: 23009
Опубліковано: 10.05.2007
Автори: Кабернюк Андрій Аркадійович, Колибо Денис Володимирович, Романюк Світлана Іванівна, Комісаренко Сергій Васильович, Редчук Тарас Анатолійович, Олійник Олена Сергіївна, Буркальова Дар''я Олександрівна
МПК: A61K 39/44
Мітки: corynebacterium, токсину, субодиниці, diphtheriae, продуцент, inv, штам, escherichia, неактивної, рекомбінантної, дифтерійного
Формула / Реферат:
Штам Escherichia coli К 12 "inv" sbA як продуцент рекомбінантної неактивної субодиниці А дифтерійного токсину Corynebacterium diphtheriae, гібридизованої з полігістидиновим тагом.
Штам escherichia coli k12 „inv” sbв – продуцент рекомбінантної неактивної субодиниці b дифтерійного токсину corynebacterium diphtheriae, гібридизованої з полігістидиновим тагом

Номер патенту: 84357
Опубліковано: 10.10.2008
Автори: Кабернюк Андрій Аркадійович, Олійник Олена Сергіївна, Редчук Тарас Анатолійович, Комісаренко Сергій Васильович, Колибо Денис Володимирович, Романюк Світлана Іванівна
МПК: A61K 47/48, A61K 39/44, C12N 15/11, C07K 7/08
Мітки: corynebacterium, escherichia, дифтерійного, штам, гібридизованої, токсину, рекомбінантної, тагом, неактивної, diphtheriae, inv, продуцент, субодиниці, полігістидиновим
Формула / Реферат:
Штам Escherichia coli K12 "inv" sbB як продуцент рекомбінантної неактивної субодиниці В дифтерійного токсину Corynebacterium diphtheriae гібридизованої з полігістидиновим тагом, амінокислотна послідовність якої...
Попередній патент: Спосіб прогнозування ремоделювання міокарда у хворих на акромегалію
Наступний патент: Спосіб формування ілеоасцендоанастомозу
Випадковий патент: Спосіб одержання стероїдних напівпродуктів або їх 22r/s стереоізомерів