Композиція, що містить виділений фактор лелістада, вакцинна композиція для вакцинації тварин (варіанти), діагностичний набір для виявлення антитіла
Номер патенту: 27788
Опубліковано: 16.10.2000
Автори: Меуленберг Йоханна Йакоба Марія, Терпстра Катарінус, Пол Йоаннес Марія Антоніс, Венсворт Герт, Морманн Робертус Йокобус Марія
















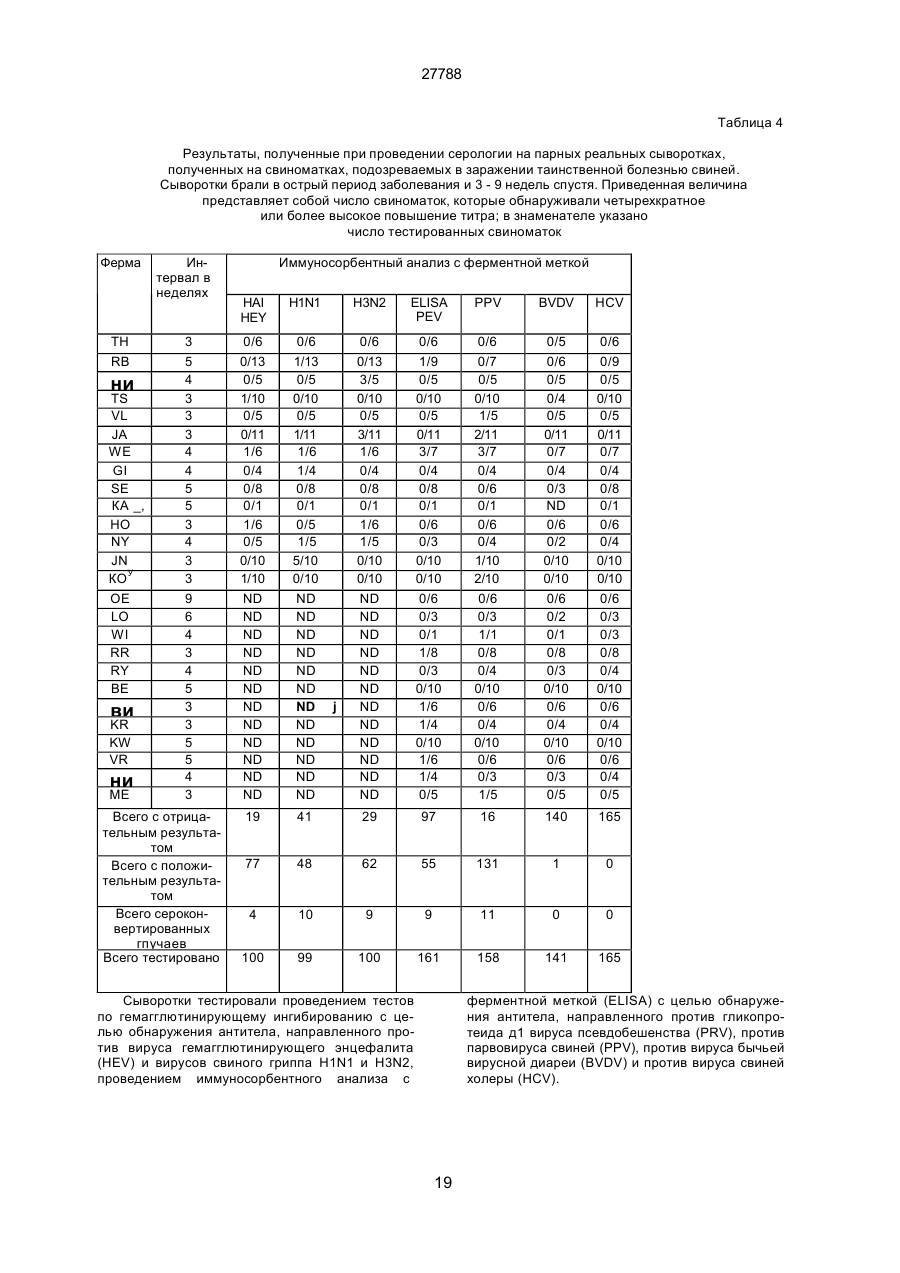
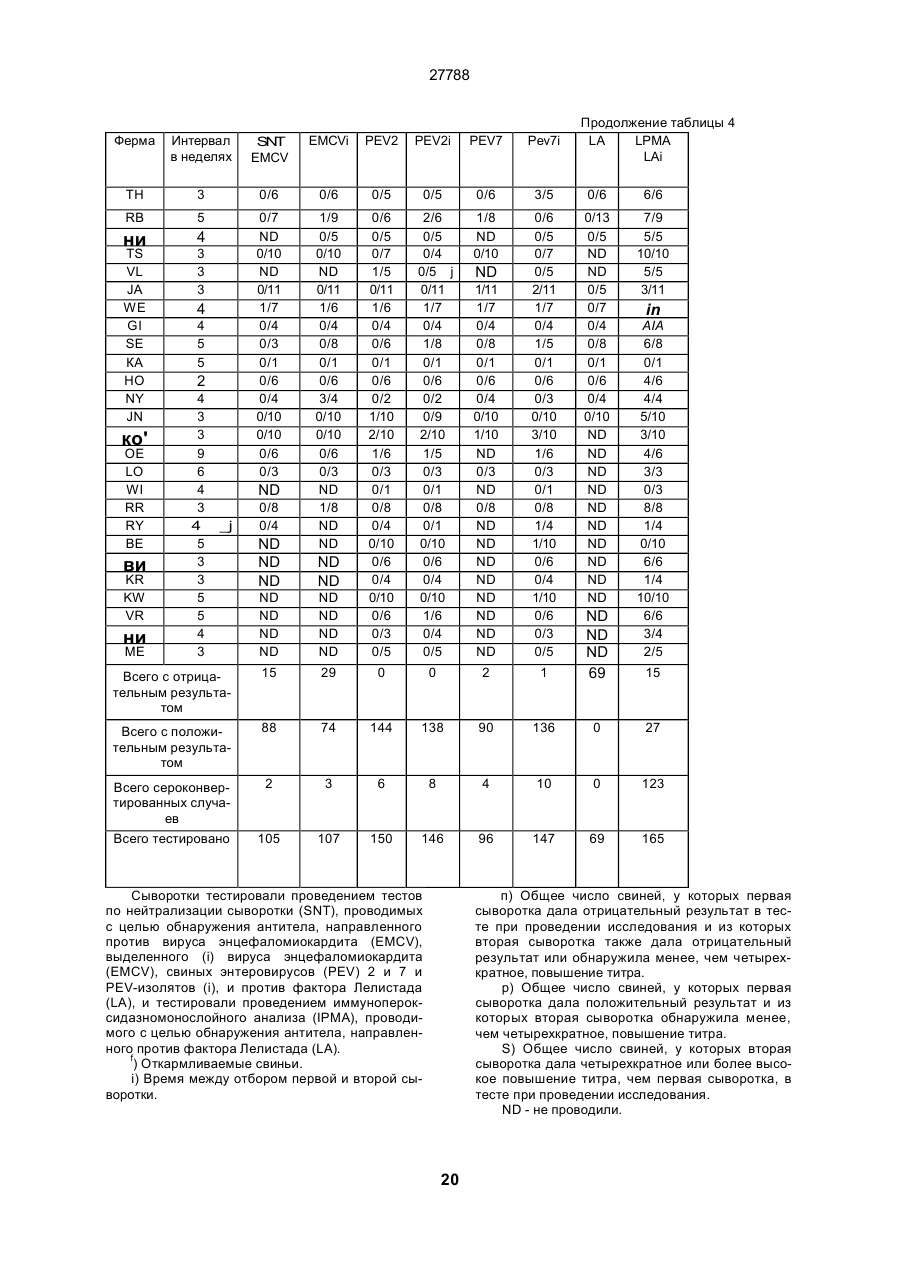


























Текст
1. Композиция, содержащая выделенный фак тор Лелистада (Lelistad Agent), который является этиологическим агентом Таинственного заболева ния свиней (Mystery Swine Desease), причем упо мянутый фактор Лелистада по существу соответ ствует изоляту фактора Лелистада (CD1-NL-2.91), депонированному под номером 1 - 1102, в Инсти туте Пастера. 2. Композиция по п. 1, отличающаяся тем, что содержит убитый выделенный фактор Лелистада. 3. Композиция по п.1, отличающаяся тем, что содержит ослабленный выделенный фактор Ле листада. 4 Вакцинная композиция для вакцинации животных, в частности млекопитающих, в особенности поросят или свиней, для защиты их от Таинствен ного заболевания свиней, содержащая фактор Лелистада по п. 1 и подходящий носитель или адъювант. 5. Вакцинная композиция по п. 4, отличающаяся тем, что содержит убитый фактор Лелистада 6. Вакцинная композиция по п. 4, отличающаяся тем, что содержит ослабленный фактор Лели стада. 7. Вакцинная композиция для вакцинации живот ных, в частности млекопитающих, в особенности поросят или свиней, для защиты их от Таинствен ного заболевания свиней, включающая антиген ную часть или компонент фактора Лелистада, ко торый является этиологическим фактором Таин ственного заболевания свиней, где упомянутый фактор Лелистада по существу соответствует изоляту фактора Лелистада (CD1-NL-2.91), депо нированному под номером 1-1102, в Институте Пастера, и подходящий носитель и адъювант 8. Диагностический набор для обнаружения анти тела, которое специфически распознает фактор Лелистада в образце, в частности, в биологиче ском образце, таком как кровь или сыворотка кро ви, мокрота, слюна или ткань, взятые у поросенка или свиньи, содержащий антигенную часть или компонент фактора Лелистада и подходящие средства обнаружения при проведении анализа на обнаружение антитела. 9. Диагностический набор по п. 8, отличающийся тем, что содержит белок или антигенный полипептид, происходящий из фактора Лелистада, или пептид, мимикрирующий антигенный компонент фактора Лелистада. 10. Диагностический набор по п.8, отличающийся тем, что включает убитый, живой или ослаблен ный фактор Лелистада. Изобретение касается выделения, описания и использования возбудителя таинственного заболевания свиней В изобретении используется открытие фактора, вызывающего заболевание, и проводится определение его геномной организации, геномной нуклеотидной последовательности и белков, закодированных геномом, что делается с целью защиты от инфекции и диагностики инфекций, в частности, защиты от инфекций таинственного заболевания свиней и диагностики таких инфекций, и с целью получения вакцинных составов и диагностических наборов, предназначенных для использования либо в случае таинственного заболевании свиней, либо в случае заболеваний, см О СО 00 см 27788 вызванных другими патогенными микроорганизмами или факторами. Зимой и ранней весной 1991 г. свиная индустрия Дании понесла урон от внезапного появления нового заболевания среди племенных свиноматок. Большинство свиноматок проявляло анорексию, у некоторых происходил выкидыш на поздней стадии беременности (примерно на 110 день), происходила мертворожденность или рождались мумифицированные плоды, и некоторые свиноматки характеризовались проявлением лихорадочного состояния. Иногда обнаруживались свиноматки с синеватыми ушами; по этой причине заболевание получило название "Абортус блаув (синеватость плода)" ("Abortus Blauw"). Заболевание у свиноматок часто сопровождалось появлением респираторного расстройства и смертью их поросят и часто появлением респираторного заболевания и замедлением роста у подросших поросят и откармливаемых свиней. Причина этой эпизоотии была не известна; однако симптомы напоминали симптомы аналогичного заболевания, возникшего в Германии с конца 1990 г., и напоминали симптомы так называемой "Таинственной болезни свиней" ("Mystery Swine Disease", как она наблюдалась с 1987 г. на среднем западе Соединенных Штатов Америки и в Канаде (Hill, 1990). Для обозначения этой болезни использовались различные другие названия: в Германии она известна как "Эпизоотический поздний аборт у свиньи" ("Seuchenhaften Spatabort der Schweine"), а в Северной Америке она также известна как "Таинственная свиная болезнь" ("Mysterious Pig Disease"), "Таинственный репродуктивный синдром" ("Mysterious Reproductive Syndrome") и "Свиной бесплодный и респираторный синдром" ("Swine Infertility and Respiratory Syndrome"). В Северной Америке Лула (Loula, 1990) описал общие клинические признаки болезни следующим образом: 1) отказ от пищи, слабые животные всех возрас тов; 2) выкидыши, рождения мертвого плода, слабые поросята, появления мумифицированного плода; 3) послеопоросные респираторные проблемы; 4) проблемы, связанные с размножением. Не был еще идентифицирован этиологический фактор, но вирус энцефаломиокардита, свиной парвовирус, вирус псевдобешенства, вирус свиного гриппа, вирус бычьей вирусной диареи, вирус свиной холеры, свиные энтеровирусы, вирус заболевания, подобного гриппу, хламиды, лептоспиры - все они были уже названы в качестве возможной причины (Loula, 1990; Mengeling and Lager, 1990, среди других). Изобретение дает состав вещества, включающего в себе выделенный Фактор Лилистада (Lelystad), который является причинным фактором таинственной болезни свиней, причем упомянутый фактор Лелистада является в существенной мере соответствующим изоляту фактора Лелистада (CDI-NL-2.91), внесенному на хранение 5 июня 1991 г. Институтом Пастера, Париж, Франция, депозитный номер 1 - 1102. Слова "в существенной мере соответствующим" относятся к вариациям, которые встречаются в природе, и к искусственным вариациями фактора Лелистада, особенно к тем, которые еще допускают обнаружение способами, подобными гибридизации, способу полимеризной цепной реакции и способу иммуносорбентного анализа с ферментной меткой, проводимым с использованием специфических в отношении фактора Лелистада материалов, таких как специфическая в отношении фактора Лелистада ДНК или специфические антитела. В состав вещества могут входить живой, убитый или ослабленный выделенный фактор Лелистада; рекомбинантный вектор, выделенный из фактора Лелистада; выделенная часть или компонента фактора Лелистада; выделенный или синтетический белок, (поли)пептид или нуклеиновая кислота, выделенные из фактора Лелистада; рекомбинантная нуклеиновая кислота, которая включает в себя нуклеотидную последовательность, выделенную из генома фактора Лелистада; (поли)пептид с аминокислотной последовательностью, выделенной из белка фактора Лелистада, в случае чего (поли)пептид продуцируется клеткой, способной продуцировать его в силу своего генетического сочетания с надлежащей рекомбинантной ДНК; выделенное или синтетическое антитело, которое специфически распознает часть или компоненту фактора Лелистада; или рекомбинантный вектор, который содержит нуклеиновую кислоту, включающую в себя нуклеотидную последовательность, кодирующую белок или антигенный пептид, выделенный из фактора Лелистада. На ДНК-уровне изобретение, в частности, дает рекомбинантную нуклеиновую кислоту, точнее, рекомбинантную ДНК, которая включает в себя специфическую в отношении фактора Лелистада нуклеотидную последовательность, показанную на фиг. 1. Желательно, чтобы упомянутая специфическая в отношение фактора Лелистада нуклеотидная последовательность выбиралась из какойлибо последовательности, ограниченной рамками открытого считывания, как это показано на фиг.1. На пептидно-белковом уровне изобретение, в частности, дает пептид, включающий в себя специфическую в отношении фактора Лелистада аминокислотную последовательность, показанную на фиг. 1. Изобретение, далее, дает вакцинный состав, предназначенный для вакцинации животных, в частности, млекопитающих, точнее, поросят или свиней, с целью защиты их от таинственной болезни свиней, включающий в, себя фактор Лелистада, либо живой, либо убитый, либо ослабленный; или рембинантній вектор, который содержит нуклеиновую кислоту, включающую в себя нуклеотидную последовательность, кодирующую белковый или антигенный пептид, выделенный из фактора Лелистада; антигенную часть или компоненту фактора Лелистада; белок или антигенный полипептид, выделенный из фактора Лелистада, или пептид, мимикрирующий антигенную компоненту фактора Лелистада, и надлежащий носитель или адъювант. Изобретение также дает вакцинный состав, предназначенный для вакцинации животных, в частности, млекопитающих, точнее, поросят и свиней, с целью защиты их от заболевания, вызванного патогенным микроорганизмом, вклю 27788 чающим в себя рекомбинантный вектор, выделенный из фактора Лелистада, нуклеиновую кислоту рекомбинантного вектора, включающего в себя нуклеотидную последовательность, кодирующую белок или антигенный пептид, выделенный из патогенного микроорганизма, и надлежащий носитель или адъювант. Изобретение, далее, дает диагностический набор, предназначенный для обнаружения нуклеиновой кислоты из фактора Лелистада в образце, в частности, в биологическом образце, таком как кровь или сыворотка крови, мокрота, слюна или ткань, взятые у животного, в частности,у млекопитающего, точнее, у поросенка или свиньи, включающий в себя нуклеиновокислотную пробу или пример, которые включают в себя нуклеотидную последовательность, выделенную из генома фактора Лелистада, и надлежащие средства обнаружения, позволяющие обнаруживать нуклеиновую кислоту. Изобретение также дает диагностический набор, предназначенный для обнаружения антигена из фактора Лелистада в образце, в частности, в биологическом образце, таком как кровь или сыворотка крови, мокрота, слюна или ткань, взятые у животного, в частности, у млекопитающего, точнее, у поросенка или свиньи, включающий в себя антитело, которое специфически распознает часть или компоненту фактора Лелистада, и надлежащие средства обнаружения, позволяющие обнаруживать антиген. Изобретение также дает диагностический набор, предназначенный для обнаружения антитела, которое специфически распознает фактор Лелистада в образце, в частности, в биологическом образце, таком как кровь или сыворотка крови, мокрота, слюна или ткань, взятые у животного, в частности, у млекопитающего, точнее, у поросенка или свиньи, включающий в себя фактор Лелистада, антигенную часть или компоненту фактора Лелистада, белок или антигенній полипептпд, выделенный из фактора Лелистада, или пептид, мимикрирующий антигенную компоненту фактора Лелистада, и надлежащие средства обнаружения, позволяющие обнаруживать антитело. Изобретение также касается способа установления наличия у животного, в частности, у млекопитающего, точнее, у поросенка или свиньи, возбудителя таинственной болезни свиней, включающего в себя приготовление образца, в частности, биологического образца, такого как кровь или сыворотка крови, мокрота, слюна или ткань, взятые у животного, и выявление наличия нуклеиновой кислоты, отвечающей фактору Лелистада, антигена, отвечающего фактору Лелистада, или антитела, специфически распознающего фактор Лелистада, где упомянутый фактор Лелистада является этиологическим фактором таинственного заболевания свиней и в основном соответствующим изоляту фактора Лелистада (CDI-NL-2.91), внесенного на хранение 5 июня 1991 г. Институтом Пастера, Париж, Франция, депозитный номер 1-1102. Настоящее изобретение является результатом совместных усилий Центрального ветеринарного института и Региональных служб охраны здоровья животных Нидерландов в их стремлении найти причину нового заболевания в виде "Таинственной болезни свиней". Фермы, на которых свиньи оказались подверженными новому заболеванию, посещали участковые ветеринары Региональных служб охраны здоровья животных. Больных свиней, образцы, взятые у больных свиней, и сыворотки свиноматок, взятые в острый период заболевания и в период выздоровления, направляли для выделения вируса в Региональные службы охраны здоровья животных и в Центральный ветеринарный институт. Парные сыворотки заболевших свиноматок тестировали на наличие антител, направленных против десяти известных свиных вирусов. Были выделены три различных вируса, вирус энцефаломиокардита, свиной энетровирус типа 2 свиной энтеровирус типа 7 и неизвестный фактор, фактор Лелистада. Свиноматки, которые, были зарегистрированы зараженными этой болезнью, характеризовались в основном сероконверсией в фактор Лелистада и редко в какой-либо из иных вирусных изолятов или известных вирусных патогенных микрообоазований. С целью экспериментального воспроизведения таинственной болезни свиней восемь беременных свиноматок были инокулированы интраназально фактором Лелистада на 84-й день беременности. Одна свиноматка родила семь мертвых и четырех живых, но очень слабых поросят на 109-й день беременности; четверо живых поросят умерли через день после рождения. Еще одна свиноматка родила на 116-й день три мумифицированных плода, шесть мертвых поросят и трех живых поросят; два живых поросенка умерли в течение дня. Третья свиноматка родила на 117-й день два мумифицированных плода, восемь мертвых и семь живых поросят. Другие свиноматки опоросились примерно на 115-й день и характеризовались менее тяжелыми репродуктивными потерями. Среднее число живых поросят от всех восьми свиноматок при рождении составило 7,3, а среднее число мертвых поросят при рождении было 4,6. Антитела, направленные против фактора Лелистада, были обнаружены в 10 из 42 образцов сыворотки, взятых до того, как поросята начали сосать. Фактор Лелистада был выделен из трех поросят, умерших вскоре после рождения. Эти результаты свидетельствуют в пользу вывода, что фактор Лелистада является причинным фактором появления таинственной болезни свиней. Фактор Лелистада развивается с цитопатическим эффектом в легочных макрофагах свиней, и он может быть идентифицирован по окрашиванию при проведении иммунопероксидазномонослойного анализа с использованием постинфекционных сывороток свиней с номерами с 829 и Ь822 или с использованием любых иных постинфекционных сывороток SPF - свиней, перечисленных в табл. 5. Антитела, направленные против фактора Лелистада, могут быть идентифицированы по методикам косвенного окрашивания при проведении иммунопероксидазномонослойного анализа. Фактор Лелистада не развивается в любой иной клеточной системе, подвергнутой тестированию. Фактор Лелистада не нейтрализуется гомологическими сыворотками или сыворотками, направленными против совокупности известных вирусов (табл. 5). Фактор Лелистада не гемглютинирует с эритроци 27788 тами, подвергнутыми тестированию. Фактор Лелистада характеризуется размером менее 200 нм, поскольку он проходит через фильтр с порами этого размера. Фактор Лелистада обнаруживает чувствительность к хлороформу. Приведенные выше результаты свидетельствуют о том, что фактор Лелистада не является пока еще идентифицированным как принадлежащий к определенной вирусной группе или к иному микробиологическому виду. Он был внесен на хранение 5 июня 1991 г. под номером 1 -1102 в Институте Пастера, Франция. Теперь установлены геномная организация, нуклеотидные последовательности и полипептиды, установленные из таковых, у фактора Лелистада. Эти данные совместно с данными других исследователей (см. ниже) оправдывают классификацию фактора Лелистада (здесь далее называемого также вирусом Лелистада, ЛВ) как члена новой семьи вирусов Артеривиридов. 6 качестве вируса-прототипа этой новой семьи предлагается взять вирус лошадиного артериита (Eguine Arteritis Virus (EAV)), который станет первым членом новой семьи, для которого стали доступными данные по стратегии репликации генома и его организации (de Vries et al.,1990, и имеющиеся там ссылки). На основании сопоставления полученных данных о последовательностях с данными, имеющимися для вируса элевации лактатдегидрогеназы (Lactate Dehydrogenase-blevating Virus (LDV; Jodeny et al., 1990)), может быть высказано предложение считать вирус элевации лактатдегидрогеназы также членом Артеривиридов. Приведенные геномная организация и трансляционная стратегия Артеривиридов представляется достаточными для помещения этой новой семьи вирусов в суперсемью коронавирусов (Snijderetal. ,1990a). Общим у артеривирусов является то, что их главными целевыми клетками у соответствующих хозяев являются макрофаги. Было показано, что репликация вируса элевации лактатдегидрогеназы ограничивается макрофагами их хозяина, мыши, тода как эта определенная склонность в отношении макрофагов не была еще пока выявлена у вируса лошадиного артериита и вируса Лелистада. Артеривирусы представляют собой сферические заключенные в оболочку частицы, имеющие диаметр 45 - 60 нм и содержащие иносаэдрический нуклеокапсид (Brinton-Darnell and Plagemann, 1975; Horginek et al., 1971; Hyllseth, 1973). Геном Артеривирид состоит из положительной спиральной полиаденилированной РНК-молекулы с размером примерно в 12000 - 13000 оснований (Brinton-Darnell and Plagemann, 1975; van der Zeijst et al., 1975). Лошадиный артериитный вирус репликатирует через находящийся в З'-месте набор из шести субгеномных кислот мРНК с размерами, находящимися в области от 800 до 3600 оснований, которые составляют лидерную последовательность, идущую от 5'-конца геномной РНК, которая соединяется с З1 -концевыми основными последовательностями (de Vries et al., 1990). Здесь было показано, что геномная организация и репликационная стратегия у вируса Лелистада являются схожими с таковыми у вируса ло шадиного артериита, коронавирусов и торовирусов, тогда как геномные размеры у последних вирусов являются совершенно отличными от таковых у вируса Лелистада и вируса лошадиного артериита. Геном у вируса Лелистада состоит из геномной РНК-молекулы с длиной примерно от 14500 до 15500 оснований (оценка произведена на нейтральном агарозном геле), которая репликатирует через находящийся в З'-месте набор из субгеномных кислот РНК. Субгеномные кислоты РНК состоят из лидерной последовательности, длина которой пока еще остается неизвестной, которая исходит из 5'-конца геномной РНК и которая сливается с основными последовательностями, исходящими из З'-конца геномной РНК (фиг. 2). Нуклеотидная последовательность у геномной РНК вируса Лелистада была определена из анализа перекрывающихся кДНК-клонов. Консеквентная последовательность из 15088 пар оснований была получена покрытием почти полного генома вируса Лелистада (фиг. 1). У этой последовательности было идентифицировано 8 кадров открытого считывания: кадр открытого считывания 1А, кадр открытого считывания 1В и кадры открытого считывания от 2 до 7. Считают, что кадры открытого считывания 1А и 1В кодируют вирусную репликазу или полимеразу, тогда как кадры открытого считывания от 2 до 6 кодируют структурную вирусную мембрану (оболочку), т.е. связанные с ней белки. Считают, что кадр открытого считывания 7 кодирует структурный вирусный нуклеокапсидный белок. Поскольку продукты, отвечающие кадрам открытого считывания 6 и 7 у вируса Лелистада, обнаруживают значительную схожесть соответственно с продуктами из областей VpX и Vpl у вируса эливации лактатдегидрогеназы, можно полагать, что последовательности, отвечающие кадрам открытого считывания 6 и 7 должны быть также сильно консервативными среди антигенных вариантов вируса Лелистада. Полная нуклеотидная последовательность, показанная на фиг. 1, и все последовательности и белковые продукты, кодированные кадрами открытого считывания от 1 по 7 и, возможно, другие кадры открытого считывания, находящиеся на последовательности, показанной на фиг. 1, являются особенно пригодными для разработки вакцины, независимо от вкладываемого здесь смысла, и для разработки диагностических инструментов, независимо от вкладываемого здесь смысла. Все возможные варианты и модификации хорошо известны специалистам, работающим в этой области техники. Поскольку теперь представляется возможным определенно идентифицировать фактор Лелистада, т.е. причинный фактор таинственной болезни свиней, можно теперь провести тестирование на заряженность свиней фактором Лелистада. Такие диагностические тесты пока еще никто не проводил. Тест может быть проведен посредством выделения вируса в макоофагах или в иных системах клеточной культуры, в которых может развиваться фактор Лелистада, и окрашивания инфектированньх культур антителами, направленными 27788 против фактора Лелистада (такого как постинфекционные сыворотки с829 или Ь822), однако могут быть также разработаны и применены другие типы диагностических тестов Например, можно использовать методики прямого или косвенного иммуногистологического окрашивания, т е проводить анализ с использованием антител, направленных против фактора Лелистада, который подвергается воздействию метки с флюоресцентными соединениями, такими как изотиоцианат, или воздействию метки с ферментами, такими как пероксидаза хрена обыкновенного Эти методики могут быть использованы для обнаружения антигена фактора Лелистада в тканевых срезах или в иных образцах, взятых у свиней, подозреваемых в заболевании таинственной болезнью свиней Антителами, необходимыми для этих тестов, могут быть антитела с829 пли Ь822 или иные поликлональные антитела, направленные против фактора Лелистада, но могут быть также использованы и моноклональные антитела, направленные против фактора Лелистада Далее, поскольку определена природа и организация генома у фактора Лелистада и нуклеотидная последовательность у этого генома, могут быть идентифицированы специфические нуклеотидные последовательности фактора Лелистада и они могут быть использованы для создания олигонуклеотидных последовательностей, которые могут быть использованы в качестве проб или примеров в диагностических методиках, таких как методика с гибридизацией, методика с полимеразной цепной реакцией или любые иные методики, разработанные для специфического обнаружения нуклеотидных кислотных последовательностей Можно также провести тестирование на антитела, направленное против фактора Лелистада Из табл 5 видно, что у экспериментально зараженных свиней быстро появляются антитела, направленные против фактора Лелистада, и из табл 4 видно, что у свиней, относящихся к этой области, также быстро проявляются антительные реакции, направленные против фактора Лелистада Таким образом, теперь также можно установить, были ли свиньи заражены ранее фактором Лелистада Такое тестирование представляется исключительно важным при определении отсутствия фактора Лелистада у свиней или свиных стад, или свиных популяций, или у свиней целых регионов или стран Тест может быть проведен с использованием иммунопероксидазномонослойного анализа, как это было описано ранее, но при этом могут быть также разработаны и применены другие типы диагностических тестов, направленных на обнаружение антител, направленньх против фактора Лелистада Специфические для фактора Лелистада белки, полипептиды и пептиды или пептидные последовательности, мимикрирующие антигенные компоненты фактора Лелистада, могут быть использованы в таких тестах Такие белки могут быть выделены из самого фактора Лелистада, однако такие белки могут быть также приготовлены способами с использованием рекомбинантной ДНК или с использованием пептидного синтеза При проведении этих тестов могут быть использованы специфические поликлональные и (одновременно или по отдельности) моноклональные антитела, направленные против фактора Лелистада, или специфические компоненты фактора Лелистада и (одновременно или по отдельности) использованы клеточные системы, зараженные фактором Лелистада, или клеточные системы, экспрессирующие антиген фактора Лелистада Антитела могут быть, например, использованы в качестве средства иммобилизации антигена фактора Лелистада (твердую поверхность покрывают антитела, после чего антиген фактора Лелистада связывают антителом), что сопровождается достижением повышенной специфичности у теста, или могут быть использованы в конкурентном анализе (меченое антитело и неизвестное антитело, находящиеся в образце, конкурируют в отношении имевшегося антигена фактора Лелистада) Далее, описанные выше диагностические возможности могут быть применены для тестирования других животных, таких как млекопитающие, птицы, насекомые или рыбы, или растений, или иных живых созданий с целью установления возможности их заражения фактором Лелистада или родственными факторами, с целью установления у них наличия такой инфекции в момент тестирования или с целью выявления их возможой зараженности в прошлом Поскольку теперь фактор Лелистада идентифицирован как причинный фактор таинственной болезни свиней, может быть приготовлена вакцина, поздназначенная для защиты свиней от этого заболевания Такая вакцина может быть поосто приготовлена выращиванием фактора Лелистада в свиных легочных макрофаговых культурах или в иных клеточных системах, в которых развивается фактор Лелистада Фактор Лелистада может быть подвергнут или не подвергнут очистке и убит установленными способами, такими как инактивация формалином или ультрафиолетовым светом Инактивированный фактор Лелистада может быть затем соединен с адъювантами, такими как стимуляторы Фрейнда (Freund), или гидроксидом алюминия, или иными веществами, и этот состав может быть затем введен свиньям Мертвые вакцины могут быть также приготовлены с использованием белковых препаратов фактора Лелистада, выделенных из зараженных фактором Лелистада культур или выделенных из клеточных систем, специфически экспрессирующих белок фактора Лелистада, что достигается применением ДНК-рекомбинантных методик Такие субъединицы фактора Лелистада должны быть затем обработаны так, как это указывалось выше, и это приведет к получению субъединичной вакцины Вакцины, содержащие даже еще меньшие компоненты фактора Лелистада, такие как полипептиды, пептиды или пептиды, мимикрирующие антигенные компоненты фактора Лелистада, являются также приемлемыми для использования в качестве мертвой вакцины Мертвые вакцины, направленные против таинственной болезни свиней, могут быть также приготовлены способами с применением рекомбинантной ДНК, в случае которых геном фактора 27788 Лелистада или части такового встраивают в векторные системы, такие как вирус коровьей оспы, вирус герпеса, вирус псевдо-бешенства, аденовирус, бакуловирус, или в иные приемлемые векторные системы, которые, тем самым, могут экспрессировать антиген фактора Лелистада в надлежащих клеточных системах. Антиген фактора Лелистада, взятый из этих систем, может быть затем использован для создания вакцины, как это говорили выше; и свиньи, вакцинированные такими продуктами, будут проявлять защитные иммунные реакции, направленные против фактора Лелистада. Вакцины, направленные против таинственной болезни свиней, могут также основываться на живых препаратах фактора Лелистада. Поскольку, как это представляется, серьезно страдают от заражения фактором Лелистада только молодые поросята и беременные свиноматки, может использоваться ноослабленный фактор Лелистада, выращенный в свиных легочных макрофагах, в качестве вакцины для более старших поросят или племенных подсвинок. Этим способом свиноматки могут быть защищены от таинственной болезни свиней до начала их беременности, в результате чего они окажутся защищенными от выкидышей и мертворождений и от появления врожденных инфекций у поросят. Кроме того, материнское антитело, которое эти вакцинированные свиноматки передадут своему потомству, будет защищать их потомство от этой болезни. Ослабленные вакцины (модифицированные живые вакцины), направленные против таинственной болезни свиней, могут быть приготовлены серийно переносом фактора Лелистада в свиные легочные макрофаги, затем в легочные макрофаги иных видов или в иные клеточные системы, или в иных животных, таких как кролики, что делается до тех пор, пока не теряется их патогенность. Живые вакцины, направленные против таинственной болезни свиней, могут быть также получены способами с применением рекомбинантной ДНК в случае чего геном фактора Лелистада или части такового встраивают в векторные системы, такие как вирус коровьей оспы, вирус герпеса, вирус псевдобешенства, аденовирус, или в иные приемлемые векторные системы, которые, тем самым, могут экспрессировать антиген фактора Лелистада. Свиньи, вакцинированные такими живыми векторными системами, будут затем проявлять защитные иммунные реакции, направленные против фактора Лелистада. Сам фактор Лелистада подходит, в частности, для использования в качестве живой векторной системы. Инородные гены могут быть вставлены в геном фактора Лелистада и могут быть сделаны экспрессирущими соответствующий белок при заражении макрофагов. Эта клетка, которая является клеткой, представляющей антиген, будет воздействовать на инородный антиген и представлять его в 8-лимфоцитах и Т-лимфоцитах, которые станут выступать с надлежащими иммунными реакциями. Поскольку фактор Лелистада является, повидимому, очень клеточноспецифическим и, возможно, также очень видоспецифическим, эта кле точная система может оказаться очень безопасной системой, которая не причиняет вреда другим клеткам или видам. Фиг.1 изображает нуклеотидную последовательность генома вируса Лелистада. Показана выведенная аминокислотная последовательность, отвечающая идентифицированным кадрам открытого считывания. Метионины, закодированные (предполагаемыми) ATG-стартовыми сайтами, указываются, жирным шрифтом, а предполагаемые сайты N-гликозилирования даются подчеркиванием. Показаны различия у нуклеотидной и аминокислотной последовательности, как это идентифицировано посредством определения последовательности расположения аминокислотных остатков у различных кДНК-клонов. Подчеркиванием выделена нуклеотидная последовательность у примера 25 который был использован в гибридизационных экспериментах (см. фиг. 2 и раздел "Результаты"). Фиг. 3 изображает организацию генома у вируса Лелистада. Клоны кДНК, которые были использованы для определения нуклеотидной последовательности, указаны в верхней части рисунка. Части клонов, которые были идентифицированы посредством определения последовательности расположения аминокислотных остатков указываются черным. В нижней части рисунка показаны кадры открытого считывания, идентифицированные у нуклеотидной последовательности, и показан субгеномный набор кислот мРНК, кодирующих эти кадры. Пунктирными линиями на этих кадрах открытого считывания представлены иные возможные сайты инициирования (ATGсайты), отвечающие этим кадрам. Лидерная последовательность у геномных и субгеномных кислот РНК дается зачерненным прямоугольником. Фиг. 3 изображает характеристики роста фактора Лелистада: пустые квадратики - титр у свободного от клетки вируса; зачерненные квадратики - титр у вируса, ассоциированного с клеткой; сплошная линия - выраженный в процентах цитопатический эффект. Материалы и методы Сбор образцов Образцы и свиней собирали на фермах, где в стадах, по-видимому, имели место эпизоотии таинственной болезни свиней. Важными критериями при выборе фермы с отнесением ее к разряду зараженных таинственной болезнью свиней были следующие критерии, наличие свиноматок, отказывающихся от пищи, наличие мертворожденных плодов и выкидышей, слабое потомство, респираторное заболевание и смерть среди поросят. Исследованию были подвергнуты образцы четырех групп, взятые у свиней: 1) тканевые образцы и мазок со слизистой обо лочки рта, взятые у поросят с этим заболеванием (табл. 1А); 2) образцы крови и мазки со слизистой оболочки рта, взятые у свиноматок с этим заболеванием (табл. 1Ви4); 27788 3) тканевые образцы, назальные мазки и образ цы крови, собранные у свободных от специфиче ского патогенного микроорганизма свиней, экспе риментально зараженных посредством контакта со свиноматками с этим заболеванием,4) тканевые образцы, назальные мазки и образ цы крови, собранные у свободных от специфиче ского патогенного микроорганизма свиней, экспе риментально зараженных посредством инокуля ции образцами крови, взятыми у свиноматок с этим заболеванием (табл. 2 и 5) Приготовление образцов Образцы, необходимые для выделения вируса, получали от поросят и свиноматок, которые по клиническим данным подозревались в заболевании таинственной болезнью свиней, и от экспериментально зараженных свиней, свиноматок и их поросят, не содержавших этого специфического патогенного микроорганизма. Тканевые образцы срезали на криостатном микротоме, и срезы подвергали прямому иммунофлюоресцентному тестированию с конъюгатами, направленными против различных свиных патогенных микроорганизмов. Образцы тканей в виде 10%-ной суспензии готовили в буферном солевом растворе Хэнка (Hank's BSS) с добавленім антибиотиков, и оральные и назальные мазки помещали в буферный солевой раствор Хэнка с добавлением антибиотиков. После выдержки в течение одного часа при комнатной температуре суспензии осветляли центрифугированием в течение 10 мин при 6000 G, и надосадочные жидкости хранили при -70°С для дальнейшего использования. Лейкоцитные фракции отделяли от этилендиаминтетрауксусной кислоты или от крови гепарина, как это описано ранее (Wensvoort and Terpstra, 1988), и хранили при - 70°С. Плазму и сыворотку, предназначенные для выделения вируса, хранили при - 70°С. Сыворотку для проведения серологии брали у свиноматок, которых подозревали, что они находятся в острой фазе протекания таинственной болезни свиней; парную сыворотку брали через 39 недель. Кроме того, сыворотки брали у экспериментально зараженных свиней, не имевших ранее этого специфического патогенного микроорганизма, что делали через определенные промежутки времени; и молозиво и сыворотку брали у экспериментально зараженных свиноматок и их поросят. Сыворотки, необходимые для проведения серологии, хранили при - 20сС. Клетки Свиные легочные макрофаги получали из легких свободных от этого специфического патогенного микроорганизма свиней в возрасте 5-6 недель и из легких взрослых свободных от этого специфического патогенного микроорганизма свиноматок, взятых из собственного стада Центрального ветеринарного института. Легкие промывали от пяти до восьми раз фосфатно-солевым буферным раствором. Каждую аликвоту промывной воды собирали и центрифугировали в течение 10 мин при 300 G. Результирующий клеточный оста ток от центрифугирования снова промывали фосфатно-солевым буферным раствором и вновь суспендировали в среде, предназначенной для содержания клеточной культуры (160 мл среды 199 с добавлением 20 мл 2,95%-го триптозофосфата, 20 мл сыворотки плода коровы и 4,5 мл 1,4%-го бикарбоната натрия), доводя концентрацию до 4x10 клеток в 1 мл. Клеточную суспензию затем медленно смешивали с равным объемом диметилсульфоксидной смеси (6,7 мл указанной выше среды, 1,3 мл сыворотки плода коровы, 2 мл 97%-го диметилсульфоксида), фасовали в ампулы на аликвоты по 2 мл и хранили в жидком азоте. Макрофаги, взятые из одной ампулы, готовили для клеточной культуры двухкратной промывкой раствором MEM Эрла (Earle's MEM) и вновь суспендировали в 30 мл среды роста (среда MEM Эрла с добавлением 10%-ной сыворотки плода коровы, пенициллина в количестве 200 ед/мл, 0,2 мг/мл стрептомицина, 100 ед/мл микостатина и 0,3 мг/мл глутамина). Клетки линии РК-15 (Американская коллекция типовых культур микроорганизмов, CCL 33/ и клетки линии SK-6 (Kasga et ai., 1972) выращивали так, как это описано у Уэусвурта и др. (Wensvoort et al. 1989). Второстепенные свиные почечные (РК2) клетки выращивали в среде MEM Эрла при добавлении 10%-ной сыворотки плода коровы и указанных выше антибиотиков Все клетки выращивали в камере, предназначенной для выращивания клеточных культур, что делали при 37°С и содержании диоксида углерода в 5% МЄТОДЖІЛ выделления вирусов Выделение вирусов проводили согласно установленным методикам при использовании клеток РК2, РК-15 и SK-6 и свиных легочных макрофагов. Первые три разновидности клеток выращивали в склянках на 25 мл (Грейнера (Greiner)) и инокулировали опытным образцом, когда захват монослойным покрытием достигал 70 - 80%. Макрофаги засевали в аликвотах на 10 мкл, находящихся на микротитраторных пластинах на 96 углублений (Грейнер), или при больших объемах в надлежащих склянках и инокулировали опытным образцом в пределах одного часа после засевания. Культуры просматривали ежедневно на наличие цитопатических эффектов и замораживали при - 70°С, когда цитопатический эффект доходил до 50 70°С или после 5 - 1 0 дней развития культуры. Дальнейшие пассажи производили на замороженном-оттаявшем материале пассажного уровня 1 и 2 или выше. Некоторые образцы инокулировали также на девятый-двенадцатый день куриными яйцами с развивающимися эмбрионом. Аллантоиновую среду субинокулровали два раза при использовании инкубационного интервала в три дня; и собранные клетки третьего пассажа, выросшие в культуре, обследовали проведением гемагглютинации при 4°С с использованием эритроцитов цыпленка и проведением иммуносорбентного анализа с ферментной меткой для специального обнаружения нуклеопротеида вирусов гриппа A(De Boer et al., 1990). 27788 Серология Сыворотки тестировали проведением тестов по гемагглютинирующему ингибированию с целью изучения развития антитела, направленного против вируса гемагглютинирующего энцефалита и вирусов свиного гриппа H1N1 и H3N2, что делали согласно прописи Мазурела (Masurel, 1976). Начальное разбавления сывороток в тестах по гемагглютинирующему ингибированию составляли 1 9, после чего сыворотки разбавляли вдвое. Сыворотки тестировали в установленных иммуносорбентных анализах с ферментной меткой на антитела, направленные против гликопротеидов д1 вируса псевдобешенства (PRV; Van Oirschot et al., 1983), свиного парвовируса (PPV; Westenbrink et al., 1989), вируса бычьей вирусной диареи (BVDV; VVfestenbrmk et al, 1986) и вируса свиной холеры (HGV; V\fensvoort et al., 1988). Начальные разбавления в тестах с иммуносорбентным анализом с ферментной меткой составляли 1 5, после чего сыворотки разбавляли вдвое. Сыворотки тестировали на нейтрализацию антител, направленных против доз 30 - 300 TClDso вирусов энцефаломиокардита, свиных эктеровирусов и фактора Лелистада, что делали согласно прописи Терпстра (Terpstra, 1978) Начальные разбавления сывороток в тестах по нейтрализации сыворотки составляли 1:5, после чего сыворотки разбавляли вдвое. Сыворотки тестировали на связывание с фактором Лелистада, проводя иммунопероксидазномонослойный анализ. Фактор Лелистала (LA; код CDI-NL-2.91) засевали на микротитраторных пластинах при добавлении 50 мл среды для выращивания, содержащей 100 ТСЮ5о фактора Лелистада, в углубления микротитраторной пластины, содержащей свежезасеянные легочные макрофаги Клетки выращивали в течение двух дней и затем фиксировали, поступая так, как это описано в литературе (Wensvoort, 1986). Тестируемые сыворотки разбавляли в соотношении 1:10 смесью, содержащей 0,15 М хлорида натрия, 0,05% твина 80, 4% лошадиной сыворотки, или разбавляли еще сильнее, производя последовательные четырехкратные разбавления с добавлением в углубления, и затем инкубировали в течение одного часа при 37°С. Овечьи-анти-свиные импуноглобулины, конъюгированные с пероксидазой хрена обыкновенного (фирма DAKO), разбавляли тем же самым буферным раствором и использовали при втором инкубировании, которое проводили в течение одного часа при 37°С, после чего пластины окрашивали, поступая так, как это описано в литературе (Wensvoort et al., 1986). Интенсивное красное окрашивание цитоплазмы у инфектированных макрофагов указывало на наличие связывания сывороток с фактором Лелистада. Методики идентификации вирусов Идентичность цитопатических изолятов устанавливали определением пловучей плотности в хлориде цезия, оценкой размера частиц в негативно окрашенных препаратах посредством электронной микроскопии, определением чувствительности изолята к хлороформу и нейтрализаци ей цитопатического эффекта у изолята сыворотками с известной специфичностью (табл. 3). Всякий раз, когда изолят оказывался специфически нейтрализованным сывороткой, направленной против известного вируса, считали, что изолят является представительным для этого известного вируса. Изоляты, которые обнаруживали цитопатический эффект на макрофаговых культурах, также подвергали исследованию окрашиванием с проведением иммунопероксидазномонослойного анализа в условиях постинфекции сыворотками от свиней с 329 или b 822. Изоляты реинокулировапи на макрофаговых культурах и фиксировали на второй день после инокуляции до проявления изолятом цитопатического эффекта. Всякий раз, когда изолят обнаруживал реактивность в условиях проведения иммунопероксидазномонослойого анализа с постинфекцией сыворотками от свиней с 823 или Ь822, считали, что изолят является представительным для фактора Лелистада. Представительные образцы других изолятов, выращенных в макрофагах или в неинфицированых макрофагах, также подвергали окрашиванию этими сыворотками, что делали с целью проверки специфичности сывороток. Дальнейшая идентификация фактора Лелистада ФакторЛелистада подвергали дальнейшему изучению методом гемагглютинации, проводимой при 4°С и при 37°С с использованием красных кровяных телец 0, взятых у цыпленка, морской свинки, свиньи, овцы или человека. При проведении гемагглютинационных исследований в качестве положительного контрольного теста использовали тест с вирусом свиного гриппа, подтип H3N2. Связывание свиных антисывороток, специфически направленных против вируса псевдобешенства, вируса передаваемого гастроэнтерита, вируса свиной эпидемической диареи, вируса гемагглютинирующего энцефалита, вируса африканской лихорадки свиней, вируса свиной холеры и вируса свиного гриппа типа H1N1 и H3N2, бычьих антисывороток, специфически направленных против вирусов бычьего герпеса типа 1 и 4, вируса злокачественной герпетической лихорадки, вируса парагриппа 3, бычьего респираторно-синцитиального вируса и вируса бычьего лейкоза, и птичих антисывороток, специфически направленных против вируса лейкоза птиц и вируса инфекционного бронхита, изучали с использованием специфических импуноглобулиновых конъюгатов пероксидазы хрена обыкновенного в условиях проведения иммунопероксидазномонослойного анализа на свиных легочных макрофагах, инфектированных и неинфектированных фактором Лелистада, что делали так, как это описано выше. Проведением иммунопероксидазномонослойного анализа подвергали тестированию также антисыворотки различных видов, направленные против вируса эпидемического паротита, вируса Сендаи (Sendai), вируса собачей чумы, вируса чумы рогатого скота, вируса кори, вируса пневмонии мышей, бычьего респираторно-синцитиального вируса, вируса бешенства, ( ra-bies virus ), 27788 вируса бешенства (foamy virus), вируса заболевания маеди-висна (maedi-visna virus), вируса бычьего и муринового (mrine) лейкоза, вируса иммунодефицита человека, кошек и обезьян, вируса лимфоцитарного хориоменингита, вируса кошачьего инфекционного перитонита, вируса мышиного гепатита, вируса Бреда (Breda), вируса Гантаана (Hantaan), вируса найробийской болезни овец, вируса восточного, западного и венесуэльского энцефалолиелита у лошадей, вируса коревой краснухи, вируса артериита у лошадей, молочного дегидрогеназного вируса, вируса желтой лихорадки, вируса клещевого энцефалита и вируса гепатита С. Фактор Лелистада вслепую подвергали пассажу в клетках РК2, РК-15 и SK-6 и в куриных яйцах с развивающимся эмбрионом. После двух пассажей материал снова инокулировали в свиных легочных макрофаговых культурах для повторного выделения фактора Лелистада. Фактор Лелистада титровали в свиных легочных макрофагах до и после пропускания через фильтр с размером отверстий 0,2 мкм (Schleicher and Schuell). Фактор Лелистада обнаруживали проведением иммунопероксидазномонослойного анализа и по его цитопатическому эффекту. Титры рассчитывали согласно Риду и Мюнку (Reed and Muench, 1938). Затем готовили свиную антисыворотку, направленную против фактора Лелистада. Две свиньи (21 и 23), свободные от этого специфического патогенного микроорганизма, инфектировали интраназально с дозой 1О5ТСЮ5о материалом, взятым от пятого пассажа клеточной культуры фактора Лелистада. Две другие свиньи, свободные от этого специфического патогенного микроорганизма, инфектировали интраназально свежей суспензией с дозой по фактору Лелистада в Ю^СЮЙО, которая была приготовлена из легочной ткани поросят, свободных от этого специфического патогенного микроорганизма, но инфектированных им. Образцы крови брали на 0, 14, 28 и 42 постинфекционные дни. Проводили, далее, выращивание фактора Лелистада в свиных альвеолярных макрофагах, что делали с целью установления их временной характеристики роста. Свиные альвеолярные макрофаги засевали в склянках F25 на 25 мл (Грейнера), инфектировали фактором Лелистада с множественностью заражения величиной 0,01 ТСЮ5о на клетку. Через 8, 16, 24, 32, 40, 48, 56, и 64 ч после заражения исследовали одну склянку и определяли процентную величину у цитопатического эффекта в сравнении с неинфектированной контрольной культурой. Производили сбор клеток из среды с культурой, и изъятое количество заменяли равным объемом фосфатно-солевого буферного раствора. Среду и склянку хранили при 70°С. После сбора всех культур определяли титры у фактора Лелистада и выражали их в виде lodTCIDsoMn'1. Морфологию фактора Лелистада изучали проведением электронной микроскопии. Фактор Лелистада культивировали так, как это было описано выше. Через 48 часов культуры подвергали замораживанию-оттаиванию и центрифугировали в течение 10 мин. при 6000 G. Примерно 30 мл надосадочной жидкости смешивали затем с 0,3 мл специфической в отношении фактора Лелистада свиной сыворотки и инкубировали в течение 1,5 ч при 37°С. После центрифугирования в течение 30 мин. при 125 000 G результирующий твердый остаток суспендировали в 1%-ной агарозной среде Сикема (Seakem), находящейся в фосфатносолевом буферном растворе при 40°С. После коагуляции агарозный блок погружали в 0,8%-ный глутаровый альдегид и 0,8%-ный тетраоксид осмия ( Hersch et al, 1968), находящиеся в веронально-ацетатном буферном растворе с величиной рН 7,4 (230 мМ осмия на 1 кг воды), и фиксировали микроволновым облучением. Эту методику повторяли единожды с использованием свежего фиксатора. Образец промывали водой, погружали в 1%-ный раствор уранилацетата и окрашивали микроволновым облучением На протяжении всех этапов образец держали при 0°С, и микроволновое излучение (фирма "Самсунг", тип прибора RE211D) подавали при оттаивании на 5 мин. Тонкие срезы готовили по стандартным методикам, окрашивали цитратом свинца (Venable et al., 1965) и обследовали на электронном микроскопе типа СМ 10 производства фирмы "Филипс". Продолжали, далее, выделение фактора Лелистада ілз сывороток свиней, которые были получены в случаях заболевания таинственной болезнью свиней. Образцы сыворотки происходили из Нидерландов (случай заболевания Нидерланды 2), из Германии (случаи заболевания Германия 1 и Германия 2); благодаря любезности д-ров Бернера (Brner), Мюнхен, и Ниенхофа (Nienhoff), и из Соединенных Штатов [экспериментальный случай Соединенные Штаты 1 (эксперимент осуществлен с использованием образца VR-2332, взятого из Американской коллекции типовых культур микроорганизмов; благодаря любезности д-ров Коллинза (Collins), Сент-Пол, и Хладека (Chiadek), Сент-Джозеф) и случаи заболевания Соединенные Штаты 2 и Соединенные Штати 2; благодаря любезности д-ров ван-Алстина (van Alstine), ВестЛафейетт, и Слайфа (Slife), Гейлсберг]. Все образцы направляли в институт "Сентраал .диергенеескундиг институт, Лелистад" для диагноза в отношении фактора Лелистада. Все образцы использовали для выделения вируса, проводимого на свиных альвеолярных макрофагах, как это было описано выше. Цитопатические изоляты трижды подвергали пассажу и идентифицировали как содержащие фактор Лелистада проведением специфического иммуноокрашивания с использованием постинфекционных сывороток с номерами b 822 и с 829, направленных против фактора Лелистада. Изучали также антигенные взаимосвязи у изолятов NL1 (первый изолят фактора Лелистада; код CDI-NL-2.91), NL2, GE1, GE2, US1, US2 и US3 Изоляты выращивали в макрофагах, делая это так, как говорили выше, и подвергали тестированию проведением иммунопероксидазномонослойного анализа с использованием совокупности из сывороток, выделенных при реальном заболевании, и двух наборов экспериментальных сывороток. Сыворотки также тестировали проведением иммунопероксидазномонослойного анализа с использованием неинфектированых макрофагов. 27788 Лелистада и выращивались в течение 24 ч или находились в неинфектированном состоянии. Среду, в которой находилась культура клеток, отделяли, и клетки дважды промывали фосфатносолевым буферным раствором. ДНК экстрагировали, как это описано в литературе (Strauss, 1987). Цитоплазматическую РНК экстрагировали так, как это описано в литературе (Tavarolo et al., 1980), очищали центрифугированием через хлорид цезия с концентрацией 5,7 М (Setger et al., 1980), обрабатывали свободной от рибонуклеазы дезоксирибонуклеазоя (фирма "Фармация") и анализировали в 0,8%-ном нейтральном агарозном геле (Мооктапп and Hulst, 1988). Сыворотки от действительных заболеваний были следующими. Две сыворотки, положительные в отношении вируса Лелистада (ТН-187 и ТО36), были выбраны из совокупности сывороток от действительных заболеваний в Дании, положительных в отношении фактора Лелистада. Двадцать две сыворотки были выбраны из сывороток от действительных заболеваний, присланных изза границы в Лелистад для установления серологического диагноза. Сыворотки были получены из Германии (ВЕ-352, ВЕ-392 и NI-2, благодаря любезности д-ра Бернера (Bemer), Мюнхен, и д-ра Ниенхофа (Nienhoff), Мюнстер), из Великобритании (РА-141615, РА-141617 и РА-142440; благодаря любезности д-ра Пейтона (Paton), Уэйбридж), из Бельгии (РЕ-1960; благодаря любезности проф. Пенсаэрта (Pensaert), Гент), из Франции (ЕА-2975 и ЕА-2985, благодаря любезности д-ра Альбина (Albina), Плуфраган), из Соединенных Штатов (SL441, SL-451, AL-PP9577, AL-P110814/33, AL-4994A, AL-7252, JC-M 41, JC-M 44 и JC-M 45; благодаря любезности д-ра Слайфа (Slife), Гейлсберг, д-ра ван Алстина (van Alstine), Вест-Лафейетт, и д-ра Коллинза (Collins), Сент-Пол) и из Канады (RB-16, RB-I9, RB-22 и RB-23; благодаря любезности д-ра Робинсона (Robinson), Квебек). Экспериментальными сыворотками были следующие сыворотки: описанный выше набор сывороток от свиней 21 23 25 и 29, взятых на 0, 14, 28 и 42 постинфекционные дни; набор экспериментальных сывороток (полученных благодаря любезности д-ров Хладека (Chladek), Сент-Джозеф, и Коллинза (Collins), Сент-Пол), которые происходили от четырех шестимесячных поросят, зараженных интраназально с дозой величиной 105'1 TCID50 от изолята VR-2332 из Американской коллекции типовых культур микроорганизмов. Образцы крови брали у молодой свиньи 2В на 0, 20, 36 и 63 постинфекционные дни; у молодой свиньи 9G на 0, 30, 44 и 68 постинфекционные дни; у молодой свиньи 16W на 0, 25, 40 и 64 постинфекционпые дни и у молодой свиньи 16V на 0, 36 и 64 постинфекционные дни. Для изучения радиоиммуноосадительным анализом (de Mazan-court et al, 1986) белков фактора Лелистада, находящихся в свиных альвеолярных макрофагах, выращивали инфектированные фактором Лелистада и неинфектированные макрофаги, делая это в течение 16 ч в присутствии меченой среды, содержащей 358-цистеин Клетки с меткой осаждали, следуя стандартным методикам, с использованием постинфекционных сывороток 42-го постинфекционного дня от свиней b 822 и 23 и с использованием сыворотки MN8, которая была получена через 26 дней после инфектирования свиноматки изолятом VP-2332, взятым из Американской коллекции типовых культур микроорганизмов (благодаря любезности д-ра Коллинза (Collins), Сент-Пол). Осажденные белки анализировали проведением электрофореза в 12%-ном полиакриламидном геле с додецилсульфатом натрия и визуализировали проведением флюорографии. Для определения характеристик у генома фактора Лелистада экстрагировали ядерную ДНК и цитоплазматическую РНК из макрофаговых культур, которые были ннфектированы фактором Клонирование и секвенирование С целью клонирования РНК вируса Лелистада внутриклеточную РНК инфектированных вирусом Лелистада свиных легочных альвеолярных макрофагов (10 мг) инкубировали вместе с 10 мл гидроксида метилртути в течение 10 мин. при комнатной температуре. Денатурированную РНК инкубировали при 42°С совместно с 50 мМ Tris-HCI при рН 7,8,10 мМ хлорида магния, 70 мМ хлорида калия, 0,5 мМ дАТФ, дЦТФ, дГТФ и дТТФ, 0,6 мкг олигонуклеотидных примеров из тимуса теленка pd(N)6 (фирма "Фармация") и 300 ед. обратной транскриптазы вируса муринового лейкоза Молони (Moloney murine leukaemia virus), находящимися в общем объеме в 100 мкл. По прошествии часа добавляли 20 мМ этилендиаминтетрауксусной кислоты; реакционную смесь экстрагировали затем смесью фенола с хлороформом, пропускали через колонку с сефадексом G50 и осаждали этанолом. Для синтеза второй кДНК-спирали использовали ДНК-полимеразу 1 (фирма "Боерингер") и рибонуклеазу Н (фирма "Фармация") (Jubler and Hoffman, 1983). Для образования тупых концов на окончаниях двухспиральную кДНК инкубировали совместно с ДНК-полимеразой Т4 (фирма "Фармация"), делая это в реакционной смеси, которая содержала 0,05 мМ дезоксинуклеотидтрифосфатов. Затем кДНК фракционировали в 0,8%-ном нейтральном агарозном геле (Moormann and Hulst, 1988). Фрагменты размером от 1000 до 4000 оснований электроэлюировали, вставляли в сайт Sma1 плазмиды pGEM-4Z (фирма "Промега") и использовали для трансформации штамма ОН5а культуры Escherichia coli (Hanahan, 1985). Фильтры с колониями подвергали гибридизации с использованием односпиральнор кДНК-пробы с меткой в виде фосфора-32. Пробу подвергали обратному транскрибированию из РНК вируса Лелистада, которая была подвергнута фракционированию в нейтральном агарозном геле (Moormann and Hulst, 1988). Перед использованием односпиральную ДНК-пробу инкубировали совместно с цитоплазматической РНК, выделенной из ложно инфектированных легочных альвеолярных макрофагов. Взаимосвязь между кДНК-клонами у вируса Лелистада определяли проведением анализа с использованием ограничительных ферментов и проведением гибридации с образованием пятен по Садерну (Southern) на расщепленной ДНК при 10 27788 необходимые для выделения вирусов и проведения серологии. использовании транслированных в определенном месте кДНК-проб (Sambrook et a\, 1989). Для получения S'-конца у вирусного генома конструировали вторую кДНК-библиотеку, для чего использовали олигообразование (dT)i2-is олигонуклеотид, специфический в отношении S'-конца вируса Лелистада, который был комплементарным к вирусному геному со спиралью отрицательного направления; его использовали в качестве примера в реакции с первой спиралью. Реакционные условия, имевшие место при проведении синтеза первой и второй спиралей, были идентичными описанным выше. Эту библиотеку сортировали, используя олигонуклеотидные пробы, специфические в отношении вирусного S'-конца. Большую часть (более 95%) кДНКпоследовательности определяли, используя автоматический лазерный флюоресцентный (A.L.F.™-Toproeafl марка) ДНК-секвенсер, полученный от фирмы Pharmacia-LKB. Определение последовательного порядка расположения аминокислотных остатков, направляемое флюоресцентным олигонуклеотидным примером, проводили на двухспиральной ДНК при использовании набора Авторид м-секвенсинг-кит (торговая марка; фирма "Фармация"), что в существенной мере соответствовало методикам С и D, описанным в инструкции к набору Авторид™-секвенсинг-кит. Флюоресцентные примеры готовили с использованием материала ФлюореПрим™ (торговая марка; фирма "Фармация"). Оставшуюся часть последовательности определяли посредством секвенсирования двухспиральной ДНК с использованием олигонуклеотидных примеров в сочетании с 17полимеразой, основываясь на наборе, предназначенном для последовательного расположения аминокислотных остатков (Секвенсинг-кит; фирма "Фармация"), и на веществе а-328-дАТФ (фирма "Амершам"). Данные для последовательности анализировали с использованием предназначенных для этой цели программ PCGENE (фирма "Интеллигенетикс, инк.", Маунтин-Вью, США) и FASTA (Pearson and Lipman, 1988). РЕЗУЛЬТАТЫ Иммунофлюоресценция Тканевые срезы, взятые у свиней, зараженных таинственной болезнью свиней, окрашивали проведением иммунофлюоресцентного тестирования с использованием FITC-конъюгатов, направленных против вируса африканской лихорадки свиней, вируса свиной холеры, вируса псевдобешенства, свиного парвовируса, вируса свиного гриппа, вируса энцефаломиокардита и культуры Chlamydia psittaci Срезы подвергали окрашиванию, их обследовали методом флюоресцентной микроскопии; и было установлено, что все они дают отрицательный результат. Выделение вируса из поросят, взятых на фермах, зараженных таинственной болезнью свиней Цитопатические изоляты обнаруживали в макрофаговых культурах, инокулированных тканевыми образцами, взятыми у поросят в возрасте от двух до десяти дней, которые были инфектированы таинственной болезнью свиней. Положительными оказались 16 из 19 поросят, взятых на пяти различных фермах (табл. 1А). Все эти изоляты взаимодействовали при проведении иммунопероксидазномонослойного анализа с постинфекционной сывороткой свиньи с 829, тогда как неинокулированные контрольное культуры не взаимодействовали. Эти изоляты, следовательно, оказались представительными для фактора Лелистада. Один раз цитопатический изолят был обнаружен в культуре клеток SK-6, инокулированной суспензией мазга со слизистой оболочки рта поросенка шестой фермы (ферма VE) (табл. 1А). Этот изолят обнаруживал характеристики пикорнавирусов и его нейтрализовали сывороткой, специфической в отношении свиного энтеровируса 2; по этой причине изолят был идентифицирован как свиной энтеровирус PEV 2 (табл. 3). Клетки РК2, РК-15 и куриные яйца, инокулированные образцами этой группы, оставались всегда отрицательными. Экспериментальная репродукция таинственной болезни свиней Обычным образом разведенных беременных свиноматок в количестве 14 особей, беременность которых продолжалась уже в течение 10-11 недель, тестировали на наличие антитела, направленного против фактора Лелистада, для чего проводили иммунопероксидазномонослойный анализ. Все они дали отрицательный результат. Затем были образованы две группы по четыре свиноматки и доставлены в Центральный ветеринарный институт. На 12-ю неделю беременности эти свиноматки были инокулированы интраназально 2 мл фактора Лелистада (число пассажей равнялось трем, титр составляет 104>8ТСЮ50 на 1 мл). Сыворотку и этилендиаминтетрауксуснокислотные образцы крови брали на 10-й день после инокуляции. Ежедневно регистрировали потребление пищи, ректальную температуру и другие клинические симптомы. При опоросе регистрировали дату рождения и число мертвых и живых поросят, приходящихся на одну свиноматку; и брали образцы, Выделение вирусов в случае свиноматок, взятых на фермах, зараженных таинственной болезнью свиней Цитопатические изоляты обнаруживали в макрофаговых культурах, инокулированных образцами, взятыми у свиноматок, инфектированных таинственной болезнью свиней. У 41-й свиноматки из 63, взятых на 11 фермах, результат оказался положительным (таб 1В) Все эти изоляты при проведении иммунопероксидазномонослойного анализа взаимодействовали с постинфекционной сывороткой свиньи b 822, и они, следовательно, были представительными для фактора Лелистада. В одном случае цитопатический изолят был обнаружен в культуре клеток РК2, инокулированной суспензией лейкоцитовой фракции, взятой у свиноматки с фермы HV (табл. 1В). Этот изолят обнаруживал характеристики пикорнавирусов, и 11 27788 Во второй группе - у инфектированных таинственной болезнью свиней свиноматок - фактор Лелистада был выделен у 41 из 63 свиноматок; один раз был выделен вирус энцефаломиокардита. Кроме того, у 123 из 165 свиноматок, инфектированных таинственной болезнью свиней, имела место сероконверсия в фактор Лелистада, как это было установлено тестированием с проведением иммунопероксидазномонослойного анализа. Такая массовая сероконверсия не проявлялась против каких-либо других вирусных патогенных микроорганизмов, подвергнутых тестированию. В третьей группе - у свободных от этого специфического патогенного микроорганизма свиней, находившихся в контакте со свиноматками, зараженными таинственной болезнью свиней, фактор Лелистада был выделен у четырех из 12 поросят; у семи поросят был выделен свиной энтеровирус 7 (PEV7). У всех 12 свиней имела место сероконверсия в фактор Лелистада и в свиной энтеровирус 7 (PEV 7). В четвертой группе - у свободных от этого специфического патогенного микроорганизма свиней, инокулированных кровью свиноматок, зараженных таинственной болезнью свиней, - фактор Лелистада был выделен у двух свиней. Все восемь свиней обнаружили сероконверсию в фактор Лелистада. его нейтрализовали сывороткой, специфической в отношении вируса энцефаломиокардита; по этой причине изолят был идентифицирован как вирус энцефаломиокардита (табл. 3). Клетки SK-6, РК-15 и куриные яйца, инокулированные образцами этой группы, давали отрицательный результат. Выделение вирусов из свиней, свободных от этого специфического патогенного микроорганизма, которые находились в контакте со свиноматками, зараженными таинственной болезнью свиней Цитопатические изоляты обнаруживали в макрофаговых культурах, инокулированных образцами, взятыми у свободных от этого специфического патогенного микроорганизма свиней, находившихся в контакте со свиноматками, зараженными таинственной болезнью свиней. Четыре из 12 свиней дали положительный результат (табл. 2). Все эти изоляты при проведении иммунопероксидазномонослойного анализа взаимодействовали с постинфекционной сывороткой свиньи с 829 и свиньи b 822; и они, следовательно, были представительными для фактора Лелистада. Цитопатические изоляты обнаруживали также в клеточных культурах РК2, РК-15 и SK-6, инокулированных образцами этих свободных от специфического патогенного микроорганизма свиней. Семь из 12 свиней дали положительный результат (табл. 2); все эти изоляты были нейтрализованы сывороткой, направленной против свиного энтеровируса 7 (PEV 7). Один из этих семи изолятов был подвергнут дополнительному исследованию; и другие характеристики также идентифицировали изолят как PEV 7 (табл. 3). Серология свиноматок, взятых на фермах, зараженных таинственной болезнью свиней Парные сыворотки, взятые у свиноматок, зараженных таинственной болезнью свиней, тестировали в отношении разнообразных вирусных патогенных микроорганизмов и в отношении изолятов, полученных при проведении этого исследования (табл.4). Поразительно сильная антительная реакция, направленная против фактора Лелистада, была измерена в условиях проведения иммунопероксидазномонослойного анализа (75% свиноматок оказались сероконвертировэнными; сероконверсию обнаружили на 23 фермах из 26), тогда как ни один из других вирусных патогенных микроорганизмов не давал четкой картины сероконверсии. Нейтрализующие антитела, направленные против фактора Лелистада, обнаружены не были. Выделение вирусов из свиней, свободных от этого специфического патогенного микроорганизма, которые были инокулированы кровью свиноматок, зараженных таинственной болезнью свиней Цитопатические изоляты обнаруживали в макрофаговых культурах, инокулированных образцами, взятыми у свободных от этого специфического патогенного микроорганизма свиней, инокулированных кровью свиноматок, зараженных таинственной болезнью свиней. У двух из восьми свиней результат оказался положительным (табл. 2). Все эти изоляты при проведении иммунопероксидазномонослойного анализа взаимодействовали с постинфекционной сывороткой свиньи с 829 и свиньи b 822; и они, следовательно, были представительными для фактора Лелистада. Клетки РК2, SK-6 и РК-15, инокулированные образцами этой группы, давали отрицательный результат. Суммируя сказанное выше, следует сказать, что четыре группы свиней были подвергнуты тестированию на наличие факторов, которые могут быть связаны с таинственной болезнью свиней. В одной группе - у поросят, инфектированных таинственной болезнью свиней, - фактор Лелистада был выделен у 16 поросят из 20; один раз был выделен свиной энтеровирус 2 (PEV2). Серология свободных от этого специфического патогенного микрообразования свиней, находившихся в контакте со свиноматками, зараженными таинственной болезнью свиней Все восемь свободных от этого специфического патогенного микрообразования свиней обнаруживали антитело, направленное в условиях проведения имиунопероксидазномонослойного анализа против фактора Лелистада (табл.5). Ни одна из этих сывороток не давала положительного результата при проведении иммунопероксидазномонослойного анализа, выполненного на неинфектированных макрофагах. Ни одна из этих сывороток не давала положительного результата в сывороточном нейтрализационном тесте в отношении фактора Лелистада. Все сыворотки, взятые через две недели после возникновения контакта, обла 12 27788 дали высокими нейтрализующими антительными титрами (более 1280) против свиного энтеровируса 7 (PEV7), тогда как прединфекционные сыворотки давали отрицательный результат (менее 10), указывая на то, что все свиньи были также инфектированы свиным энтеровирусом 7 (PEV 7). цательной сывороткой, или в отрицательных контрольных препаратах. Определяли закономерности поведения изолятов из Нидерландов, Германии и Соединенных Штатов. Все семь изолятов были выделены в свиных альвеолярных макрофагах и подвергнуты пассажу три-пять раз. Все изоляты обуславливали появление цитопатического эффекта в макрофагах и могли быть подвергнуты специфическому иммуноокрашиванию направленными против фактора Лелистада сыворотками от свиньи Ь822 и сывороткой 23 42 - постинфекционного дня. Эти изоляты были названы как NL 2, GE1, GE2, US 1, US 2 и US 3. Устанавливали антигенные взаимосвязи у изолятов NL1, NL2, GE1, GE 2, US 1, US 2 и US 3. Ни одна из полученных сывороток не взаимодействовала в условиях проведения иммунопероксидазномонослоного анализа с неинфектированным макрофагами; одако все сыворотки содержали антитела, направленные против одного или нескольких из семи изолятов (табл. 7). Ни одна из экспериментальных сывороток не взаимодействовала в условиях проведения иммунопероксидазномонослойного анализа с неинфектированными макрофагами; и ни одна из экспериментальных сывороток, отвечающих нулевому постинфекционному дню, не взаимодействовала с каким-либо из семи изолятов при проведении иммунопероксидазномонослойного анализа (табл. 8). Все семь изолятов фактора Лелистада взаимодействовали со всеми или большинством сывороток, взятых из совокупности экспериментальных сывороток от свиней 21, 23, 25 и 29, которые были взяты после нулевого постинфекционного дня. Только изоляты US 1, US 2 и US 3 взаимодействовали со всеми или большинством сывороток из совокупности экспериментальных сывороток от поросят 2В, 9G, 16\Л/и 16Y, взятых после нулевого постинфекционного дня. Проводили исследования по радиоммуноосаждению Семь специфических в отношении фактора Лелистада белка были обнаружены в макрофагах, инфектированных фактором Лелистада, но при этом они не были обнаружены в неинфектированных макрофагах, осажденных сыворотками 42-го постинфекционного дня от свиней b 822 и 23. По оценкам белки обладали следующими молекулярными весами: 65, 39, 35, 26, 19, 16 и 15 кДа. Только два из этих специфических в отношении факторах Лелистада белка, а именно, белки с молекулярным весом 16 и 15 кДа, осаждались также сывороткой MN 8 26-го постинфекционного дня. Серология свободных от этого специфического патогенного микрообразования свиней, инокулированных кровью свиномматок зараженных таинственной болезнью свиней Все восемь свободных от итого специфического патогенного микроорганизма свиней обнаруживали антительную реакцию в иммунопероксидазномонослойном анализе, направленную против фактора Лелистада (табл. 5). Ни одна из этих сывороток не давала положительного результата в иммунопероксидазномонослойном анализе, выполненном на неинффектированных макрофагах. Ни одна из этих сывороток не давала положительного результата в сывороточном нейтрализационном тесте в отношении фактора Лелистада. Прединфекционная и двухнедельная постинфекционная сыворотки давали отрицательный результат (менее 10) против свиного энтеровируса 7 (PEV 7). Дальнейшая идентификация фактора Лелистада Фактор Лелистада не гемагглютинирует с 0красными кровяными тельцами цыпленка, морской свинки, свиньи, овцы или человека. Фактор Лелистада не взаимодействует в иммунопероксидазномонослойном анализе с сыворотками, направленными против вируса псевдобешенства, вируса передаваемого гастроэнтерита, вируса свиной эпидемической диареи, вируса африканской лихорадки свиней и т.д. После двух неконтролируемых пассажей фактор Лелистада не растет в клетках РК2, РК-15 или SK-6 или в куриных яйцах с развивающимся эмбрионом, инокулированных аллантоическим путем. Фактор Лелистада по-прежнему сохраняет инфекционную способность после пропускания через фильтр с размером отверстия 0,2 мкм; как обнаружено проведением иммунопероксидазномонослойного анализа титры до и после фильтрования составляли 105'05. и 105'3 TCID5o. Получена кривая роста фактора Лелистада (см. фиг. 3). Максимальные титры у свободного от клетки вируса примерно составляют 10 TCIDso мл"1, что имеет место по прошествии 32-48 ч после инокуляции. По прошествии указанного времени макрофаги становятся убитыми под воздействием цитопатического эффекта фактора Лелистада. Была проведена электронная микроскопия. Были обнаружены кластеры из сферических частиц фактора Лелистада. Частицы характеризовались диаметром 45 - 55 им и содержали нуклеокапсид размером 30 - 35 нм, который был окружен липидной двухслойной мембраной. Частицы фактора Лелистада не обнаруживали в инфектированных культурах, подвергнутых обработке отри Последовательность у генома фактора Лелистада и его организация Природа генома фактора Лелистада была установлена анализом ДНК и РНК, взятых из инфектированных свиных легочных альвеолярных макрофагов. Не обнаружена никакая специфическая для вируса Лелистада кислота ДНК. Однако же, в самом деле, была обнаружена специфическая для вируса Лелистада кислота РНК. В 0,8%-ном нейтральном агарозном геле РНК-вируса Лелистада мигрирует несколько медленнее, чем препарат 13 27788 ности, отвечающей вирусу Лелистада (сравните с фиг. 1). Пример 25 гибридизируется с группой из сети кислот РНК с оцененным молекулярным весом, меняющимся по размеру в пределах от 700 до 3300 оснований (субгеномные кислоты мРНК), а также с геномной РНК. Наиболее вероятным объяснением гибридизационной схемы, отвечающей примеру 25, является то, что 5'-концевые геномные последовательности, длина которых пока еще остается неизвестной, сливаются с основанием кислот мРНК которые транскрибируются от З'-конца генома. В самом деле, из гибридизационной схемы, полученной с использованием примера 25 следует, что 5'-концевые геномные последовательности функционируют как так называемая "лидерная последовательность" в субгеномных кислотах мРНК, Такая транскрипционная схема является отличительным признаком коронавирусов (Spaan et al., 1988) и вируса лошадиного артериита (de Vries et al., 1990). Единственное заметное расхождение между вирусом Лелистада и вирусом лошадиного артериита, которое может быть установлено по приведенным выше данным, состоит в том, что геномный размер у вируса Лелистада примерно на 2500 оснований больше, чем у вируса лошадиного артериита. Согласованная нуклеотидная последовательность из перекрывающихся кДНК-клонов показана на фиг. 1. Длина последовательности оставляет 15038 пар оснований, что находится в хорошем согласии с оцененным размером геномной РНК вируса Лелистада. Поскольку кДНК-библиотека для вируса Лелистада была образована в условиях случайного воздействия примера при обратном транскриптазном взаимодействии с примерами pd (N) 6 из тимуса теленка, не были получены какие-либо кДНКклоны, которые начинаются с полиадениловокислотного участка их З'-конца. С целью клонирования З'-конца вирусного генома была сконструирована вторая кДНК-библиотека, для чего были использованы олигообразование (dT) и пример 39U183R в обратной транскриптазной реакции. Пример 39U83R является комплементарным к спиральной РНК минусового направления вируса Лелистада, которая, по-видимому, присутствует в препарате РНК, выделенном из клеток, инфектированных вирусом Лелистада. Эту библиотеку сортировали с использованием специфических в отношении вируса проб (транслированный в необходимой области кДНК-клон 119 и олигонуклеотид 119R64R), результатом чего явилось выделение пяти дополнительных кДНК-клонов (например, кДНК-клон 151, фиг. 2). При выявлении порядка расположения аминокислотных остатков в этих кДНК-клонах было установлено, что вирус Лелистада содержит З'-полиадениловокислотный хвост. Длина этого полиадениловокислотного хвоста является различной у разных кДНК-клонов; однако максимальная длина составляет двадцать нуклеотидов. Помимо клонов 25 и 155 (фиг. 2) на 5'-конце генома были выделены четыре дополнительных кДНК-клона, которые оказались всего лишь на 2 - 3 нуклеотида короче, чем предельный 5'-нуклеотид, показанный на фиг. 1. С учетом этого факта и способа синтеза кДНК было высказано РНК вируса свиной холеры с размером 12300 оснований (Moormann et al., 1990). Хотя в нейтральных агарозных гелях и не может быть осуществлено определение точного размера, из сделанных оценок следует, что длина у РНК, специфической для вируса Лелистада, составляет примерно от 14500 до 15500 оснований. Для установления сложного строения специфических для вируса Лелистада кислот РНК, находящихся в инфектированных клетках, и выявления нуклеотидной последовательности у генома вируса Лелистада, готовили кДНК из РНК, взятой из свиных легочных альвеолярных макрофагов, ифектированных вирусом Лелистада, и выбирали и картировали специфические для вируса Лелистада кДНК-клоны, делая это так, как было описано в разделе материалы и методы. Специфичность кДНК-клонов переподтверждали проведением гибридизации специфических клонов, локализованных по перекрывающейся «ДНК последовательности, для чего получали пятна по Нордерну (Northern), несущие РНК инфектированных вирусом Лелистада и неинфектированных макрофагов. Поразительным было то, что некоторые из кДНКклонов гибридизировались с РНК размером от 14500 до I5500 оснований, обнаруженных лишь в инфектированных макрофагах, тогда как другие гибри-дизировались с РНК с размером от 14500 до 15500 оснований, а также с группой из четырех или пяти кислот РНК с меньшим молекулярным весом (согласно сделанным оценкам их размер составлял от 1 до 4 тысяч оснований). Все последние клоны являлись кластерными на одном конце кДНК-карты и охватывали приме ло 4000 оснований у ДНК. Из этих данных следует, что геномная организация вируса Лелистада может оказаться схожей с таковой у коронавирусов (Spaan et al., 1988), вируса Берне (Berne virus (BEV); Snijder et a)., 1990b), торовируса и вируса лошадиного артериита (de virus et al.,1990), т.е. помимо геномной РНК там имеются субгеномные кислоты мРНК, образующие сгруппированную совокупность, которая локализуется на З'-конце. Это предположение было подтверждено, когда стали доступными последовательности, отвечающие кДНК-клонам, и могли быть выбраны специфические примеры в качестве пробы, необходимой для получения пятен. Совокупность собранных гибридизационных данных, полученных с использованием кДНК-клонов и специфических примеров, которые были подвергнуты гибридизации с образованием пятен по Нордерну (Northern), несущих РНК инфектированных вирусом Лелистада и неинфектированных макрофагов, показана на фиг. 2. Клоны 12 и 20, которые расположены в 5'части и в центре последовательности, соответственно гибридизируются с геномьой РНК с размером от 14500 до 15500 оснований, обнаруженной только в клетках, инфектированных вирусом Лелистада. Клоны 41 и 39 распознают, однако, геномную РНК с размером от 14500 до 15500 оснований и совокупность из четырех и пяти кислот РНК с более низким молекулярным весом, соответственно. Наиболее поучительная и убедительная гибридизационная схема была получена, однако, при использовании примера 25, который располагается на самом конце 5' последователь 14 27788 нуклеотидном положении 12394 или на стартовом триплете АУГ (AUG) в нуклеотидном положении 12556 и затем кодировать белки, соответственно содержащие 265 и 211 аминокислот. Белок из 265 остатков содержит семь поедполагаемых Nсвязанных сайтов гликозилирования, тогда как белок из 211 остатков содержит четыре сайта (табл. 9). На N-конце белка, состоящего из 265 остатков, идентифицируется гидрофобная последовательность. Судя по результатам проведения анализа на гидрофобность, топология белка, закодированного кадром открытого считывания ORF 4, является аналогичной таковой у белка, закодированного кадром открытого считывания ORF 2, если считать, что проявление кадра открытого считывания ORF 4 инициируется на стартовом триплете АУГ (AUG) в нуклеотидном положении I2936. Однако кадр открытого считывания ORF 4 может также инициироваться в двух иных АУГ-кодонах (сравните фиг. 1 и 2) со стартовыми положениями в последовательности, приходящимися соответственно на нуклеотиды 12981 и 13068. Пока еще остается неясным, какой используется стартовый кодон. В зависимости от используемого стартового кодона кадр открытого считывания ORF 4 может кодировать белки из 183 аминокислот, содержащих четыре предполагаемых N-связанных сайта гликозилирования, из 168 аминокислот, содержащих четыре предполагаемых N-связанных сайта гликозилирозания, или из 139 аминокислот, содержащих три предполагаемых N-связанных сайта гликозилирования (табл. 9). Считают, что кадр открытого считывания ORF 5 кодирует белок из 201 аминокислоты, содержащие два предполагаемых N-связанных сайта гликозилирования (табл. 9). Характерной особенностью проявления кадра открытого считывания ORF 5 является наличие внутренней гидрофобной последовательности, расположенной в области от аминокислоты 108 до аминокислоты 132. Из анализа мембранных соединительных сегментов и гидрофильности продукта проявления кадра открытого считывания ORF 6 следует, что он содержит три трансмембранных соединительных сегмента на N-концевом участке своей последовательности, состоящей из 90 аминокислот. Эта отличительная особенность является также характеристикой небольшого оболочечного гликопротеида М или Е1 у нескольких коронавирусов, например, у вируса инфекционного бронхита (1BV; Boursnell et al., 1984) и у вируса мышиного гепатита (MHV; Rottier et al., 1986). Можно, следовательно, считать, что блок, закодированный кадром открытого считывания ORF 6, обладает мембранной топологией, аналогичной такой у белка М или Е1 коронавирусов (Rottier et al., 1986). Второй характерной особенностью белка М или Е1 является наличие так называемой поверхностной спирали, которая располагается в непосредственной близости от предполагаемой третьей трансмембранной области. Эта последовательность, состоящая примерно из 25 аминокислот, которая является вполне консервативной у коронавирусов, также распознается, хотя и в значительно более вырожденном состоянии, у вируса Лелистада. Все же предполагается, что продукт проявления кадра предположение, что получена картина, которая является очень близкой к имеющей место в случае 5'-конца последовательности геномной РНК вируса Лелистада. Почти 75% геномной последовательности вируса Лелистадта кодирует кадры открытого считывания ORF 1A и ORF 1B. Кадр открытого считывания ORF 1A инициируется, по-видимому, в первом триплете АУГ (AUG, нуклеотидное положение 212, фиг. 1), встречающемся в последовательности вируса Лелистада. У кадра открытого считывания ORF 1A С-конец перекрывает предполагаемый N-конец кадра открытого считывания ORF 1B, что происходит на небольшом участке в 16 нуклеотидов. Представляется, тем самым, что трансляция кадра открытого считывания ORF 1B осуществляется через рибосомальное смещение кадра, что является характерным признаком способа трансляции полимеразного или репликазного гена у коронавирусов (Boursnell et al., 1987) и торовирусов вируса Берне (BEV, Berne virus Snijder et al., 1990a). Характерная РНК-псевдоузельная структура, которая, как считают, должна образовываться в месте рибосомального сдвига кадра, также обнаруживается в этом местоположении у последовательности вируса Лелистада (результаты не показаны). Кадр открытого считывания ORF 1B кодирует аминокислотную последовательность, состоящую почти из 1400 остатков, которая много меньше, чем у кадров открытого считывания ORF 1В коронавирусов, таких как вирус инфекционного гепатита (MHV) и вирус инфекционного бронхита (IBV) (примерно 3700 аминокислотных остатков1 Bredenbeek et al., 1990; Boursnell et al., 1987) и вирус Берне (Berne) (примерно 2300 аминокислотных остатков, Snijder et al., 1990a). Характерные особенности проявления кадра открытого считывания ORF 1В у членов суперсемейства коронавирусов, такие как репликазный лейтмотив и наличие цинкфингерной области, могут быть также найдены у кадра открытого считывания ORF 1B вируса Лелистада (результаты не показаны). Тогда как кадры открытого считывания ORF 1А и ORF 1B кодируют вирусную полимеразу и, следовательно, как это считают, кодируют неструктурированный вирусный белок, кадры открытого считывания от ORF 2 до ORF 7 кодируют, как это полагают, структурированные вирусные белки. Продукты от наличия кадров открытого считывания от ORF 2 до ORF 6 обнаруживают специфические особенности, напоминающие связанные с мембраной (оболочкой) белки. Кадр открытого считывания ORF 2 кодирует белок из 249 аминокислот, содержащий два предсказанных Nсвязанных сайта гликозилирования (табл. 9). На N-конце идентифицируется гидрофобная последовательность, которая может функционировать как так называемая сигнальная последовательность. Конец С также завершается гидрофобной последовательностью, которая в этом случае может функционировать как трансмембранная область, которая захватывает результат проявления кадра открытого считывания 2 со встраиванием в вирусную оболочечную мембрану. Кадр открытого считывания ORF 3 моет инициироваться на стартовом триплете АУГ (AUG) в 15 27788 открытого считывания ORF 6 у вируса Лелистада обладает аналогичной связанной с мембраной функцией, как это наблюдается в случае белка М или Е1 коронавирусов. Крое того, белок, закодированный кадром открытого считывания ORF 6, обнаруживает сильную схожесть (53% идентичных аминокислот) с образованием Vpx (Jodeny et al., 1990) вируса элевации лактатдегидрогеназы. Белок, закодированный кадром открытого считывания ORF 7, обладает длиной в 128 аминокислотных остатков (табл. 9), которая на 13 аминокислот превышает длину последовательности Vpi у вируса элевации лактатдегидрогеназы (Jodeny et al, 1990). Все же наблюдается значительная схожесть (43% идентичных аминокислот) меду белком, закодированным кадром открытого считывания ORF 7, и Vpl. Еще одна общая характерная особенность продукта проявления кадра открытого считывания ORF 7 и Vpl сводится к наличию высокой концентрации основных остатков (аргинин, лизин и гистидин (Arg, Lys, His) на N-концевой половине белка. До аминокислоты 55 последовательность у вируса Лелистада содержит 26% аргинина, лизина и гистидина. Этот результат находится в полном согласии с предполагаемой функцией продукта проявления кадра открытого считывания ORF 7 или Vpl (Jodeny et al., 1990), a именно, с инкапсидацией вирусной геномной РНК На основании изложенных выше данных делается предположение, что продукт проявления кадра открытого считывания ORF 7 у вируса Лелистада представляет собой нуклеокапсидный белок Nвируса. Схематическое представление организации генома у вируса Лелистада показано на фиг. 2 Карта из перекрывающихся клонов, использованная для определения последовательности у вируса Лелистада, показана в верхней группе. Линейное изображение этой карты с указанием концов 51 и 3' у нуклеотидной последовательности вируса Лелистада показано на фиг 1; масштаб в тысячах оснований приводится под картой из кДНК-клонов, чем обеспечивается возможность последовательного расположения этих клоноа в последовательности. Положение кадров открытого считывания, идентифицированных у генома вируса Лелистада, указывается ниже под линейной картой последо вательности вируса Лелистада. Нижняя группа изображает сведенную воедино совокупность субгеномных кислот мРНК и положение этих кислот РНК по отношению к последовательности у вируса Лепистада. В согласии с представлениями о трансляционной стратегии у субгеномных кислот мРНК коронавирусов, торовирусов и артеривирусов делается предположение, что кадры открытого считывания ORF от 1 до 6 транслируются с единого 51конца их геномных кислот или кислот мРНК. Считается, что эта специфическая часть кислот мРНК представляет собой ту часть РНК, которая получается при "вычитании" РНК меньшего молекулярного веса из РНК с большим молекулярным весом, последовательно идущей за ней. Хотя РНК 7 и образует З'-конец у всех других геномных и субгеномных кислот РНК и, тем самым, не характеризуется наличием некой уникальной области, полагают, что кадр открытого считывания ORF 7 транслируется лишь с этой самой маленькой по размеру мРНК. "Лидерная последовательность", расположенная на 5'-конце у субгеномных кислот РНК, указывается зачерненным прямоугольником. Длина этой последовательности составляет примерно 200 оснований; однако точное место слияния с основанием геномных кислот РНК предстоит еще определить. Экспериментальная репродукция таинственной болезни свиней Восемь беременных свиноматок были инокулированы фактором Лелистада; и у этих свиноматок воспроизводили клинические признаки таинственной болезни свиней, такие как отсутствие аппетита и репродуктивные потери. По прошествии от четырех до 10-12 дней после инокуляции все свиноматки стали обнаруживать отвращение к еде. Ни одна из свиноматок не характеризовалась наличием повышенной температуры. У двух свиноматох стали синеватыми уши, что обнаружилось на 9-й и 10-й постинфекционные дни. В табл. 6 приведены данные о дне рождения и числе живых и мертвых поросят, приходящихся на свиноматку. Фактор Лелистада выделяли из 13 рожденных поросят. Таблица 1 Описание и результаты, полученные при выделении вируса из реальных образцов А. Образцы, взятые у поросят, подозреваемых в заражении таинственной болезнью свиней Ферма Число Результаты Использованный материВозраст свиней ал (дней) RB 5 2 легкое, миндалина, го5xLA ловной мозг, DV 4 3 легкое, мозги, пулы из 3xLA почки, селезенка ТН 3 3-5 3xLA легкое, пулы из почек, миндалина DO 3 10 Легкое, миндалина 2xLA ZA 4 1 Легкое, миндалина 3xLA VE 1 ? Оральный мазок 1xLA Всего 20 16 27788 Продолжение таблицы 1 В. Образцы, взятые у свиноматок, подозреваемых в заражении таинственной болезнью свиней Ферма Число свиномаИспользованный маРезультаты ток териал ТН 2 Плазма и лейкоциты 1 xLA ни TS НК LA VL 5 10 5 6 6 Плазма и лейкоциты « U и Сыворотка и лейкоциты ТА Сыворотка 15 LO 4 Плазма и лейкоциты JA 8 и VD 1 VW 1 Сыворотка Всего 63 ) - Результаты приведены в виде числа свиней, у которых производили выделение вируса. Иногда изолят обнаруживали не в одном, а в нескольких образцах, приходившихся на одну свинью. 2 х LA.1 х EMCV 6 xL A 2 xL A 2 xL A 5 xL A 11 xLA 2 xL A 8 xL A 1xLA 1xLA 41 x LA,1 x EMCV LA - фактор Лелистада. PEV 2 - свиной знтеровирус типа 2. EMCV - вирус зндефаломиокардита. Таблица 2 Описание и результаты, полученные при выделении вируса из образцов, взятых у свиней, подвергнутых преднамеренному экспериментальному заражению Свиноматка A(LO) B(JA) C(JA) D(VD) Всего Свинья Использованный материал с 835 Легкое, миндалина с 836 Назальные мазки с 837 Назальные мазки с 825 Легкое, миндалина с 821 Назальные мазки с 823 Назальные мазки с 833 Легкое, миндалина с 832 Назальные мазки с 829 Назальные мазки, плазма, лейкоциты с 816 Легкое, миндалина С813 Назальные мазки с 815 Назальные мазки Изоляты от контактных свиней d ) Свиней, свободных от этого специфического патогенного микроорганизма, либо держали в контакте (с) со свиноматкой, подозреваемой в заражении таинственной болезнью свиней, либо на нулевой день эксперимента внутримышечно вводили 10 мл этилендиаминтетрауксуснокислотной крови (Ь) от такой свиноматки. Группы из одной свиноматки и трех свободных от этого специфического патогенного микроорганизма поросят (с) держали в одном загоне, и все четыре такие группы помещали в одном хлеву. На шестой день одну свинью, свободную от этого специфического патогенного микроорганизма, которую брали из каждой группы, убивали, и миндалину и легкие использовали для выделе Результаты 2 xL A 2 х PEV 7 1 х PEV 7 4 х PEV 7 1 x LA, 1 x PEV 7 2 x PEV 7 3 x LA, 2 x PEV 7 1xLA 1 x PEV 7 7 x LA, 13 x PEV 7 ния вируса, Четыре группы свободных от этого специфического патогенного микроорганизма свиней, инокулированных кровью (Ь), помещали в четыре других загона, находящихся в отдельном хлеву. Назальные мазки у свободных от этого специфического микроорганизма свиней брали на второй, пятый, седьмой и девятый дни, считая от начала эксперимента, и этилендиаминтетрауксуснокислотную кровь, предназначенную для выделения вируса из плазмы и лейкоцитов, брали всегда, когда у свиньи обнаруживалась лихорадка. ) Результаты приводятся в виде числа изолятов, приходящихся на свииью. LA - фактор Лелистада. 17 27788 PEV 7 - свиной энтеровирус типа 7. () 6 с кобках приведены началь ны е буквы названия фермы , откуда проис ходила свиноматка Таблица 3 Идентификация вирусных изолятов Пловучая ' Размер * плотн ос ть в ч ас т и ц п о данным CsCl флю оресцентной микрос копии (нм^ 1,33 28-30 Чувствите3 льность к хлороформ у Отсутствует E MC V( 1 2 8 0) Вирус энцефаломиокардита Оральные мазки Свиноферма VE SK6 Не опреде- 28-30 ляли Отсутствует PEV2(>1280) Свиной энтеровирус Оральные мазки, мин-далина Св инь и без п ат ог ен а CV1 РК15, РК2, SK6 Не опреде- 28-30 ляли Отсутствует Pev7(>1280) Различные образцы, Различны е фермы , Свиные легочны е макрофаги 1,19 Имеется Нет (все
ДивитисяДодаткова інформація
Назва патенту англійськоюComposition comprising the causative lelystad agent, vaccine composition for animal vaccination (variants) and diagnostic kit for antibody detection
Автори англійськоюWensvoort Gert, Terpstra Catharinus, Pol Joannes Maria Anthonis, Moormann Robertus Jacobus Mari, Meulenberg Johanna Jacoba Maria
Назва патенту російськоюКомпозиция, содержащая выделенный фактор лелистада, вакцинная композиция для вакцинации животных (варианты), диагностический набор для выявления антитела
Автори російськоюВенсворт Герт, Терпстра Катаринус, Пол Йоаннес Мария Антонис, Морманн Робертус Йокобус Мария, Меуленберг Йоханна Йакоба Мария
МПК / Мітки
МПК: G01N 33/53, C07K 14/08, A61P 31/12, C12N 15/09, C07K 14/435, C07K 16/00, A61P 31/14, A61K 39/395, C12P 21/00, A61K 39/12, C12P 21/02, C12N 7/00, C07K 16/08
Мітки: містить, варіанти, тварин, виділений, вакцинації, вакцинна, діагностичний, композиція, антитіла, лелістада, фактор, виявлення, набір
Код посилання
<a href="https://ua.patents.su/50-27788-kompoziciya-shho-mistit-vidilenijj-faktor-lelistada-vakcinna-kompoziciya-dlya-vakcinaci-tvarin-varianti-diagnostichnijj-nabir-dlya-viyavlennya-antitila.html" target="_blank" rel="follow" title="База патентів України">Композиція, що містить виділений фактор лелістада, вакцинна композиція для вакцинації тварин (варіанти), діагностичний набір для виявлення антитіла</a>
Макролідні антибіотики або їх солі, спосіб їх одержання (варіанти), композиція для лікування тварин і людей, композиція і спосіб боротьби з шкідниками сільського господарства

Номер патенту: 27261
Опубліковано: 15.09.2000
Автори: НОБЛ Девід, Ремсей Майкл Вінсент Джон, Вард Джон Беррі, ПОРТЕР Ніл, Сазерленд Дерек Рональд, Нобл Хейзел Мері, Флеттон Річард Алан
МПК: A61K 31/365, A01P 5/00, A01P 3/00, C07D 493/22, A01P 7/02, A01N 43/32
Мітки: людей, сільського, шкідниками, спосіб, боротьби, одержання, антибіотики, варіанти, господарства, тварин, солі, макролідні, лікування, композиція
Формула / Реферат:
(57) 1. Макролидные антибиотики общей формулы (lІ)где R1 - метил, этил или изопропил,R2 - атом водорода или группа OR5, где OR5 - гидроксильная или замещенная гидроксильная группа, содержащая до 25 атомов углерода, и R3 - атом водорода, или R2 и R3 вместе с атомом углерода, к которому они присоединяются, представляют > С=О группу, иOR4 представляет группу OR5, как указано выше, причем, когда R2 представляет...
Спосіб одержання препарату, який містить фактор

Номер патенту: 8072
Опубліковано: 26.12.1995
Автор: Мінору Матіда
МПК: A61K 38/22, A61P 39/00
Мітки: препарату, одержання, фактор, містить, спосіб
Формула / Реферат:
(57) 1. Способ получения препарата, содержащего фактор (G-CSF), стимулирующий рост колоний гранулоцитов, отличающийся тем, что смешивают G-CSF при температуре от 1 до 30°С по меньшей мере с одним веществом, выбранным из группы, включающей фармацевтически приемлемое поверхностно-активное вещество, сахарид, белоки высокомолекулярное соединение в соответствующих указанному порядку перечисления количества: 1-10000 мас.ч., 1-10000мас.ч., 1-20000...
Спосіб прямого відновлювання матеріалу, що містить оксиди заліза (варіанти), і установка для його здійснення (варіанти)

Номер патенту: 27017
Опубліковано: 28.02.2000
Автори: ЦІП Герхальд, МІЛІОНІС Константин, Гушех Мортеза Садат, УІПП Рой Хуберт мол.
МПК: C21B 13/02, C21B 13/00
Мітки: оксиди, здійснення, установка, варіанти, спосіб, відновлювання, прямого, містить, заліза, матеріалу
Формула / Реферат:
1. Способ прямого восстановления содержащего оксиды железа материала, в котором газ синтеза смешивают с колошниковым газом, образующимся при прямом восстановлении содержащего оксиды железа материала, и используют для прямого восстановления в качестве восстановительного газа, который не разлагается при температуре восстановления, отличающийся тем, что объемное содержание Н2О в восстановительном газе поддерживают равным от 1 до 2%,...
Композиція, що містить ліпосомний пухирець та введений у нього білковий медикамент, і спосіб її приготування

Номер патенту: 26640
Опубліковано: 11.10.1999
Автори: Ча Янсик, Коллінз Девід
МПК: A61K 9/127, A61K 38/19
Мітки: композиція, містить, пухирець, білковий, приготування, спосіб, введений, нього, ліпосомний, медикамент
Формула / Реферат:
1. Композиция, содержащая липосомный пузырек и введенный в него белковый медикамент, отличающаяся тем, что в качестве липосомного пузырька содержит отрицательно заряженный фосфолипид, а в качестве белкового медикамента - протеин, способный к переходу в распрямленное состояние глобулы, при соотношении липида к протеину не менее 10 : 1.2. Композиция по п.1, отличающаяся тем, что она имеет pH 3,0 - 7,5.3. Композиция по п.1,...
Поліферментна композиція для годівлі сільськогосподарських тварин

Номер патенту: 10246
Опубліковано: 25.12.1996
Автори: Кас'яненко Зоя Іванівна, Баралевич Анатолій Олександрович, Федоренко Ярослав Дмитрович
МПК: A23K 1/165, A23K 1/175
Мітки: композиція, тварин, сільськогосподарських, поліферментна, годівлі
Формула / Реферат:
Полиферментная композиция для откорма сельскохозяйственных животных, содержащая пектофоетидин ГЗх, отличающаяся тем, что она дополнительно содержит целлотеррин ГЗх и глюкоамилазу Г20х при следующем соотношении компонентов в единицах активности на 1 кормовую единицу:пектофоетидин ГЗх 2,0-2,8целлотеррии ГЗх 17,3-23,0глюкоамилаза Г20х 5,8-7,7
Попередній патент: Спосіб обробки газоподібних відходів, які містять шкідливі речовини, для регенеративного теплообмінника та пристрій для його здійснення
Наступний патент: Спосіб каталітичної ізомеризації с8 – ароматичних вуглеводнів
Випадковий патент: Муфта