Спосіб визначення сортової чистоти та типовості партій насіння сільськогосподарських культур з використанням днк-маркерів
Номер патенту: 56555
Опубліковано: 25.01.2011
Автори: Парій Мирослав Федорович, Вдовиченко Жанна Вікторівна, Спиридонов Владислав Геннадійович



Формула / Реферат
Спосіб визначення сортової чистоти та типовості партій насіння сільськогосподарських культур з використанням ДНК-маркерів, що включає створення тест-системи шляхом підбору поліморфних ДНК-маркерів, виділення ДНК із рослинного зразка, що аналізується, проведення ампліфікації ДНК-маркерів у полімеразній ланцюговій реакції, аналіз продуктів ампліфікації, який відрізняється тим, що ДНК виділяють одночасно із репрезентативної сукупності насінин оцінюваного сорту; аналіз продуктів ампліфікації проводять шляхом визначення якісного та кількісного складу алелів маркерних послідовностей, а отримані співвідношення алелів використовують для розрахунку сортової чистоти (типовості) партії насіння.
Текст
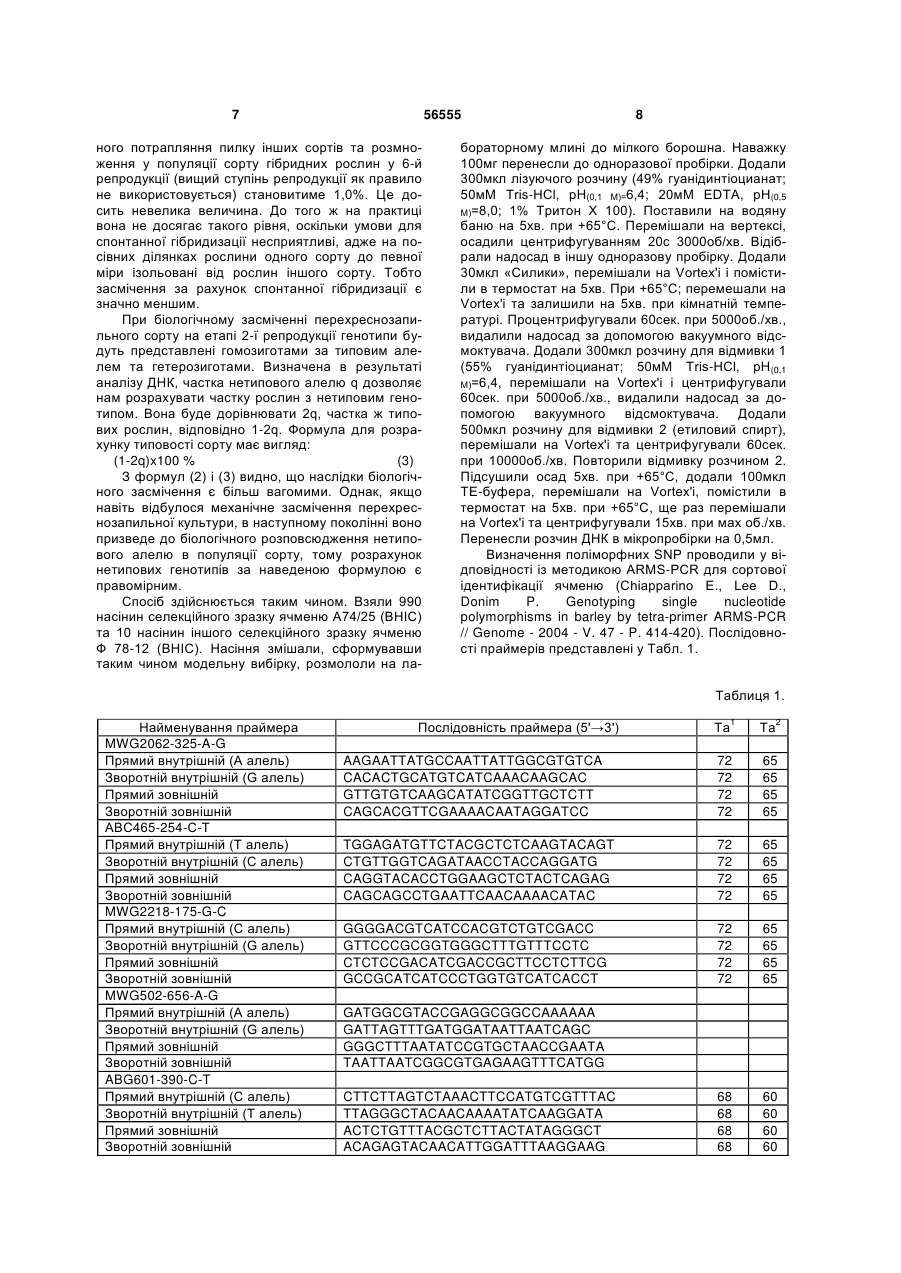
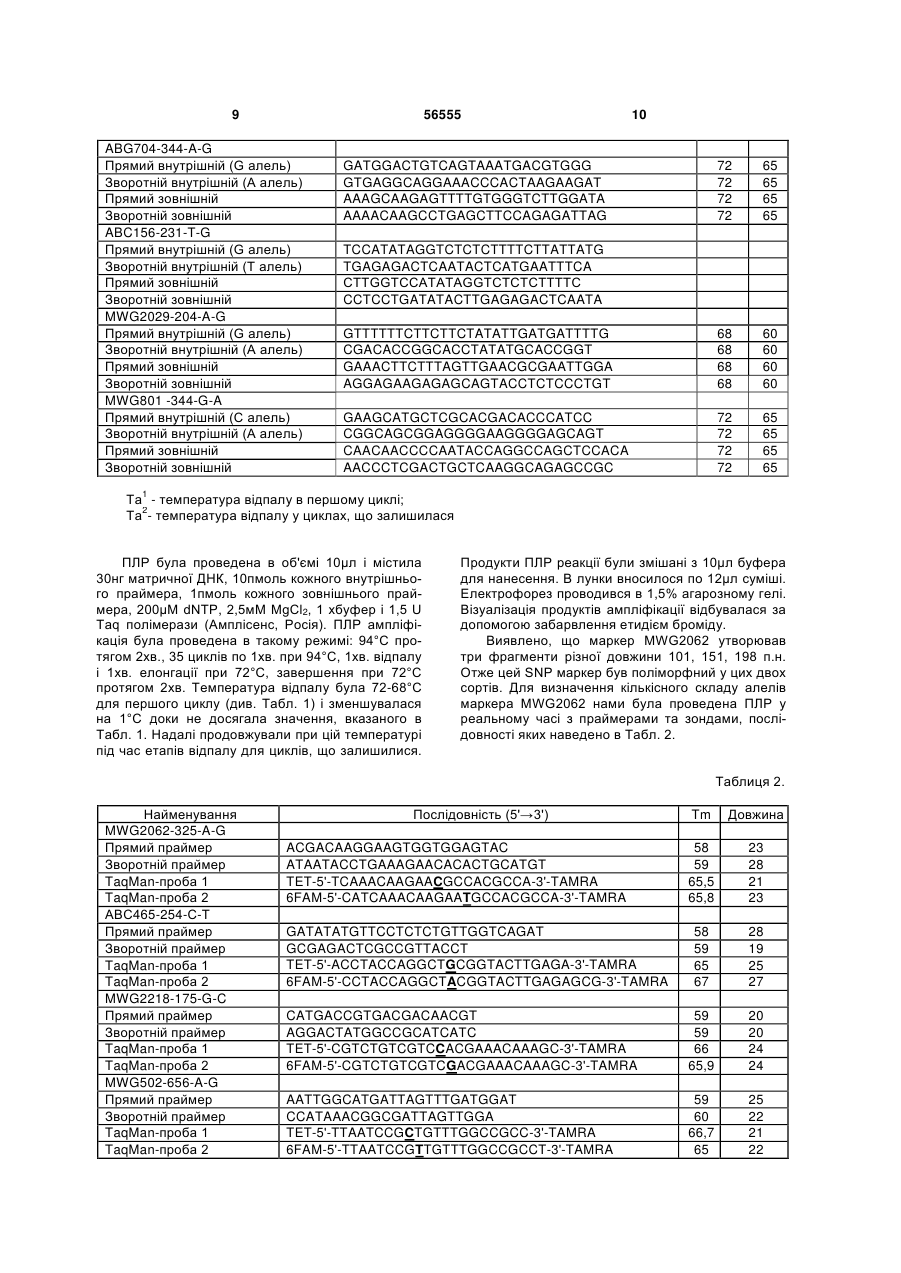
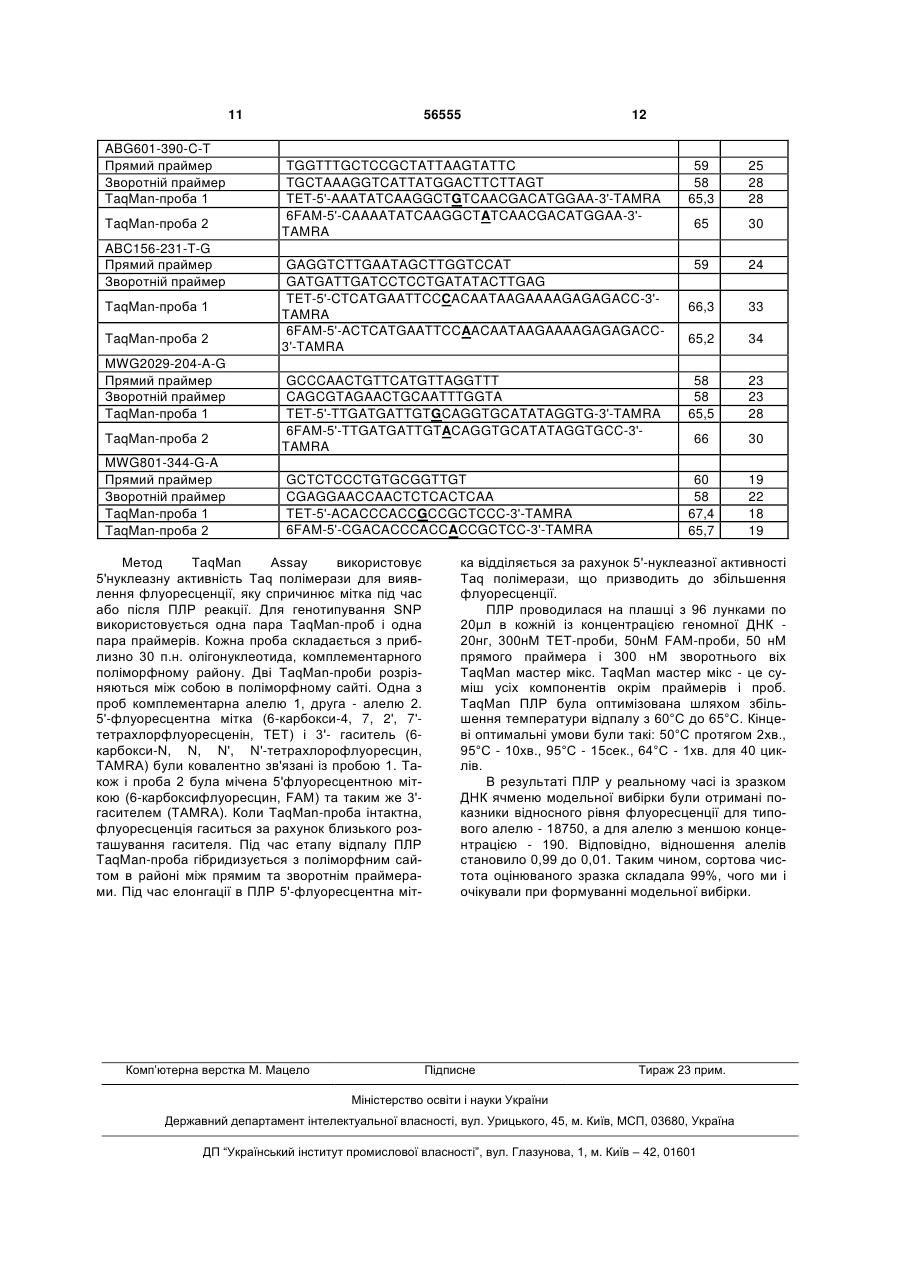
Спосіб визначення сортової чистоти та типовості партій насіння сільськогосподарських культур з використанням ДНК-маркерів, що включає ство 3 водити 400 ПЛР реакцій та розподіляти продукти ампліфікації на 400 треках для визначення типовості лінії. В основу винаходу поставлено задачу в способі визначення сортової чистоти та типовості партій насіння сільськогосподарських культур зменшити витрати праці для аналізу одного зразка (партії) та зменшити термін проведення, шляхом одночасного аналізу частотного складу алелів маркерних послідовностей в сукупності насінин оцінюваного зразка. Поставлена задача вирішується тим, що у способі визначення сортової чистоти та типовості партій насіння сільськогосподарських культур, що включає створення тест-системи шляхом підбору поліморфних ДНК-маркерів, виділення ДНК із рослинного зразка, що аналізується, проведення ампліфікації ДНК-маркерів у полімеразній ланцюговій реакції, аналіз продуктів ампліфікації, згідно винаходу виділення ДНК відбувається одночасно із репрезентативної сукупності насінин оцінюваного сорту, проводиться аналіз частот алелів досліджуваних маркерів шляхом виявлення алельного стану маркерів і визначення співвідношень алелів у зразку, а отримані співвідношення використовуються для розрахунку сортової чистоти (типовості) партії насіння. Запропонований спосіб характеризується такими суттєвими ознаками: створення тест-системи шляхом підбору поліморфних ДНК маркерів; виділення ДНК із репрезентативної сукупності насінин сорту; проведення ампліфікації ДНК-маркерів у полімеразній ланцюговій реакції, аналіз продуктів ампліфікації, який полягає у визначенні якісного та кількісного складу алелів маркерних генів у зразку; використання отриманих співвідношень алелів для розрахунку сортової чистоти (типовості) партії насіння. Такі етапи, як виділення ДНК із репрезентативної сукупності насінин сорту; аналіз продуктів ампліфікації, який полягає у визначенні якісного та кількісного складу алелів маркерних генів у зразку; використання отриманих співвідношень алелів для розрахунку сортової чистоти (типовості) партії насіння є відмінними від прототипу ознаками. Нові, відмінні від прототипу, ознаки забезпечують зменшення витрат праці для виконання аналізу одного зразка та скорочення терміну його проведення. Чим досягається інтенсифікація процесу визначення сортової чистоти (типовості) партії насіння. До переваг запропонованого способу можна віднести такі: - кількість локусів, за якими може бути проведено аналіз, необмежена, локуси рівномірно розподілені по геному рослин; - об'єм вибірки може бути значно збільшений у порівнянні з прототипом і обмежується тільки розподільчою здатністю метода визначення кількісного вмісту алелів в суміші ДНК, що збільшує достовірність аналізу. На відміну від прототипу у винаході можливе визначення засмічення, яке складає менше ніж один відсоток; - виділення сумарної ДНК відбувається одразу із усіх оцінюваних рослин в одній пробі і подальший аналіз цієї сумарної ДНК. В результаті суттєво 56555 4 скорочується кількість реакцій, необхідних для проведення оцінки. - спосіб має невисоку вартість. Визначення сортової чистоти та типовості партій насіння - це необхідна процедура для оцінки якості насіннєвої продукції, в якій зацікавлені як її виробники, так і споживачі цієї сировини. Тому вона має виконуватися швидко і бути максимально спрощеною, при цьому не втративши своєї надійності. Однак існуючі методи не відповідають цим вимогам. Визначення чистоти (типовості) сорту за допомогою електрофорезу білків не забезпечує достатньої інформативності та надійності, оскільки білкові маркери характеризуються незначним поліморфізмом і не охоплюють геном рослини у достатній мірі (Сиволап Ю.М., Солоденко А.Є. Спосіб встановлення типовості та рівня гібридності генотипів соняшника // Деклараційний патент на винахід № UА63265 від 15.01.2004). Спосіб, обраний як прототип - визначення чистоти (типовості) сорту за допомогою SSR-маркерів, хоч і позбавлений недоліків способа-аналога, але є високовартісним. В обох випадках ці методи вимагають значних витрат праці і часу, оскільки потребують оцінки кожної насінини (генотипу) окремо. Це обмежує величину вибірки насінин, що можуть бути залучені до аналізу, а значить і точність аналізу. В способі-аналозі та способі-прототипі визначення сортової чистоти (типовості) проводиться шляхом підрахунку кількості типових та нетипових рослин за профілем розподілу в гельелектрофорезі білків (аналог) або продуктів ампліфікації (прототип). В способі запропоновано проводити оцінку сортової чистоти (типовості) через визначення співвідношень алелів маркерних генів в суміші ДНК всіх рослин оцінюваного зразка. Методи визначення співвідношень алелів під час аналізу ДНК базуються на тому, що кількість продуктів ампліфікації в одних і тих же умовах залежить від вихідної кількості матричної ДНК і, відповідно, кількість ампліфікованих фрагментів кожного алеля буде залежати від вихідної кількості матричної ДНК відповідного типу. Кількість ДНК кожного з алельних варіантів може бути виміряна, наприклад, методом ПЛР у реальному часі. Здійснення винаходу можливе двома шляхами: 1. Одночасний аналіз якісного і кількісного складу апелів ДНК маркерів, тобто ампліфікація в такій кількості реакцій, яка дорівнює кількості маркерів в тест-системі. Якщо в якості методу аналізу ДНК застосовувати ПЛР у реальному часі, то спрощення аналізу відбувається також за рахунок того, що на відміну від існуючих методів, виключається стадія електрофорезу продуктів ампліфікації, оцінка результатів аналізу ДНК відбувається автоматично. 2. Послідовне визначення, спочатку якісного складу алелів маркерів в одній реакції, з наступним визначенням співвідношення тільки тих алелів маркерів, які виявили поліморфізм під час першої реакції. В результаті застосування другого варіанту кількість реакцій може бути зменшеною до двох. Наприклад, при використанні SNP маркерів (маркерів послідовностей ДНК, які відрізняються замі 5 56555 ною одного нуклеотида), проводиться перша мультиплексна полімеразна ланцюгова реакція, в який визначається якісний склад алелів (SNPlex). На другому етапі обирається один з маркерів, що виявився поліморфним, і проводиться ПЛР в реальному часі із зондом до поліморфного маркера, Спосіб Спосіб-аналог Спосіб-прототип Винахід 6 таким чином визначається кількісне співвідношення алелів. Для збільшення точності можливий варіант кількісного аналізу двох поліморфних маркерів. Порівняймо необхідну кількість проб для застосування існуючих методів та винаходу: Кількість проб 100 100 х кількість маркерів Варіанти здійснення: 1) 1 х кількість маркерів; 2) 1+1 x (кількість поліморфних маркерів, виявлених у першій реакції) Сортова чистота (типовість) - це відношення кількості насінин даного сорту до загальної їх кількості, виражена у відсотках. Ці показники мають суттєве значення в процесі насінництва. Встановлено норми, при перевищенні яких партії насіння не допускаються до посіву або до використання. Цей показник визначає цінність партії насіння та її придатність для закладки насінницьких посівів. Рівень сортової чистоти (типовості) залежить від системи розмноження культури і знижується через механічне або біологічне засмічення. Поняття "сортова чистота" використовується для самозапильних культур (ячменю, пшениці, сої та ін.). Система насінництва самозапильних культур складається із кількох ланок: оригінальне насінництво, елітне насінництво, насінництво 1-ої, 2-ої, 3ої і т.д. репродукцій. Відповідно до ланок виділяють і категорії насіння: оригінальне, елітне, 1-ої, 2ої, 3-ої категорій). До певної категорії відносять насіння тієї чи іншої репродукції в залежності від показника сортової чистоти. Сортова чистота елітного насіння пшениці, ячменю, вівса, гороху та ін. має бути не нижче 99,7%, насіння 1-ої категорії 99,5%, 2-ої - 98%, 3-ої - 95% (Семена сельскохозяйственных культур. Сортовые и посевные качества. - М. 1991). У виробництві використовується насіння 1-ої, 2-ої, 3-ої категорій. Насінництво самозапильних культур відбувається простим пересівом сорту. Внаслідок самозапилення отримують насіння, яке представляє собою прямих нащадків однієї або кількох вихідних рослин. Тому зниження сортової чистоти, у самозапильних культур відбувається переважно внаслідок механічного засмічення, тобто шляхом потрапляння насіння інших сортів під час збирання, зберігання та висіву, а також внаслідок посіву сорту на площах, де попередником була та ж культура іншого сорту. Найчастіше засмічення відбувається в ланках репродукційного насінництва. Типовість - показник, що використовується для оцінки партій насіння гомозиготних ліній перехреснозапильних культур, насінництво яких ведеться шляхом перезапилення на просторово ізольованих ділянках. При відтворенні насіння перехреснозапильних культур до механічного засмічення додається біологічне, під яким розуміють потрапляння пилку іншого сорту на насінницькі посіви. Наприклад, вимоги до сортової чистоти перехреснозапильних культур такі: жито, кукурудза: 1 кат. - 99%, 2 кат. - 98%, 3 кат. - 97%, цукрова куку рудза - 1 кат. - 99,5%, 2 кат. - 98%, 3 кат. - 95%, для цукрового буряка - не нижче 94 % (Семена сельскохозяйственных культур. Сортовые и посевные качества." М. 1991). Висновок про сортову чистоту (типовість) у винаході, що заявляється, робиться в залежності від способу розмноження культури та системи її насінництва. Наведемо основні формули, які можуть бути застосовані для розрахунку сортової чистоти (типовості). В результаті аналізу ДНК, виділеної із сукупності усіх оцінюваних рослин ми отримуємо величину (p/q), яка відображає співвідношення алелів, наявних у популяції сорту. Ця величина дозволяє розрахувати частоту алелів по кожному з локусів, що аналізуються. Позначимо через р і q частоти алелів за одним з локусів, при цьому приймемо, що р - це типовий для оцінюваного сорту алель. Частоту q можна виразити як 1-р, тоді p p q 1 p , Звідси p q (1) p p 1 q Підставляючи в рівняння значення p/q, отримане в результаті кількісного аналізу ДНК, отримуємо значення частоти p. При механічному засміченні сорту самозапильної культури в популяції сорту будуть представлені гомозиготи за типовим алелем та гомозиготи за нетиповим алелем у певному співвідношенні. Співвідношення генотипів буде в даному випадку таким самим, як і співвідношення апелів, що оцінюються. Таким чином, визначена частка типових рослин, переведена у відсотки, і є показник сортової чистоти: (2) р 100 % На етапі 1-ої репродукції самозапильної культури засмічення відбувається механічним шляхом. Тому для розрахунку чистоти достатньо брати до уваги лише його. Встановлено, що у пшениці спонтанна гібридизація досягає 0,2%. Приблизно така ж величина спостерігається і для інших самозапильних культур (Гуляев Г.В., Гужов Ю.Л. Селекция и семеноводство полевых культур. - М. Агропромиздат, 1987. - 447 с. С. 348). За нашими розрахунками кількість нетипових рослин за умови щоріч 7 56555 ного потрапляння пилку інших сортів та розмноження у популяції сорту гібридних рослин у 6-й репродукції (вищий ступінь репродукції як правило не використовується) становитиме 1,0%. Це досить невелика величина. До того ж на практиці вона не досягає такого рівня, оскільки умови для спонтанної гібридизації несприятливі, адже на посівних ділянках рослини одного сорту до певної міри ізольовані від рослин іншого сорту. Тобто засмічення за рахунок спонтанної гібридизації є значно меншим. При біологічному засміченні перехреснозапильного сорту на етапі 2-ї репродукції генотипи будуть представлені гомозиготами за типовим алелем та гетерозиготами. Визначена в результаті аналізу ДНК, частка нетипового алелю q дозволяє нам розрахувати частку рослин з нетиповим генотипом. Вона буде дорівнювати 2q, частка ж типових рослин, відповідно 1-2q. Формула для розрахунку типовості сорту має вигляд: (1-2q)x100 % (3) З формул (2) і (3) видно, що наслідки біологічного засмічення є більш вагомими. Однак, якщо навіть відбулося механічне засмічення перехреснозапильної культури, в наступному поколінні воно призведе до біологічного розповсюдження нетипового алелю в популяції сорту, тому розрахунок нетипових генотипів за наведеною формулою є правомірним. Спосіб здійснюється таким чином. Взяли 990 насінин селекційного зразку ячменю А74/25 (ВНІС) та 10 насінин іншого селекційного зразку ячменю Ф 78-12 (ВНІС). Насіння змішали, сформувавши таким чином модельну вибірку, розмололи на ла 8 бораторному млині до мілкого борошна. Наважку 100мг перенесли до одноразової пробірки. Додали 300мкл лізуючого розчину (49% гуанідинтіоцианат; 50мМ Tris-HCl, рН(0,1 М)=6,4; 20мМ EDTA, рН(0,5 М)=8,0; 1% Тритон Х 100). Поставили на водяну баню на 5хв. при +65°С. Перемішали на вертексі, осадили центрифугуванням 20с 3000об/хв. Відібрали надосад в іншу одноразову пробірку. Додали 30мкл «Силики», перемішали на Vortex'i і помістили в термостат на 5хв. При +65°С; перемешали на Vortex'i та залишили на 5хв. при кімнатній температурі. Процентрифугували 60сек. при 5000об./хв., видалили надосад за допомогою вакуумного відсмоктувача. Додали 300мкл розчину для відмивки 1 (55% гуанідинтіоцианат; 50мМ Tris-HCl, рН(0,1 М)=6,4, перемішали на Vortex'i і центрифугували 60сек. при 5000об./хв., видалили надосад за допомогою вакуумного відсмоктувача. Додали 500мкл розчину для відмивки 2 (етиловий спирт), перемішали на Vortex'i та центрифугували 60сек. при 10000об./хв. Повторили відмивку розчином 2. Підсушили осад 5хв. при +65°С, додали 100мкл ТЕ-буфера, перемішали на Vortex'i, помістили в термостат на 5хв. при +65°С, ще раз перемішали на Vortex'i та центрифугували 15хв. при мах об./хв. Перенесли розчин ДНК в мікропробірки на 0,5мл. Визначення поліморфних SNP проводили у відповідності із методикою ARMS-PCR для сортової ідентифікації ячменю (Chiapparino E., Lee D., Donim P. Genotyping single nucleotide polymorphisms in barley by tetra-primer ARMS-PCR // Genome - 2004 - V. 47 - P. 414-420). Послідовності праймерів представлені у Табл. 1. Таблиця 1. Найменування праймера MWG2062-325-A-G Прямий внутрішній (А алель) Зворотній внутрішній (G алель) Прямий зовнішній Зворотній зовнішній АВС465-254-С-Т Прямий внутрішній (Т алель) Зворотній внутрішній (С алель) Прямий зовнішній Зворотній зовнішній MWG2218-175-G-C Прямий внутрішній (С алель) Зворотній внутрішній (G алель) Прямий зовнішній Зворотній зовнішній MWG502-656-A-G Прямий внутрішній (А алель) Зворотній внутрішній (G алель) Прямий зовнішній Зворотній зовнішній ABG601-390-C-T Прямий внутрішній (С алель) Зворотній внутрішній (Т алель) Прямий зовнішній Зворотній зовнішній Tа1 Та2 AAGAATTATGCCAATTATTGGCGTGTCA CACACTGCATGTCATCAAACAAGCAC GTTGTGTCAAGCATATCGGTTGCTCTT CAGCACGTTCGAAAACAATAGGATCC 72 72 72 72 65 65 65 65 TGGAGATGTTCTACGCTCTCAAGTACAGT CTGTTGGTCAGATAACCTACCAGGATG CAGGTACACCTGGAAGCTCTACTCAGAG CAGCAGCCTGAATTCAACAAAACATAC 72 72 72 72 65 65 65 65 GGGGACGTCATCCACGTCTGTCGACC GTTCCCGCGGTGGGCTTTGTTTCCTC CTCTCCGACATCGACCGCTTCCTCTTCG GCCGCATCATCCCTGGTGTCATCACCT 72 72 72 72 65 65 65 65 68 68 68 68 60 60 60 60 Послідовність праймера (5'→3') GATGGCGTACCGAGGCGGCCAAAAAA GATTAGTTTGATGGATAATTAATCAGC GGGCTTTAATATCCGTGCTAACCGAATA TAATTAATCGGCGTGAGAAGTTTCATGG CTTCTTAGTCTAAACTTCCATGTCGTTTAC TTAGGGCTACAACAAAATATCAAGGATA ACTCTGTTTACGCTCTTACTATAGGGCT ACAGAGTACAACATTGGATTTAAGGAAG 9 56555 ABG704-344-A-G Прямий внутрішній (G алель) Зворотній внутрішній (А алель) Прямий зовнішній Зворотній зовнішній ABC156-231-T-G Прямий внутрішній (G алель) Зворотній внутрішній (Т алель) Прямий зовнішній Зворотній зовнішній MWG2029-204-A-G Прямий внутрішній (G алель) Зворотній внутрішній (А алель) Прямий зовнішній Зворотній зовнішній MWG801 -344-G-A Прямий внутрішній (С алель) Зворотній внутрішній (А алель) Прямий зовнішній Зворотній зовнішній 10 GATGGACTGTCAGTAAATGACGTGGG GTGAGGCAGGAAACCCACTAAGAAGAT AAAGCAAGAGTTTTGTGGGTCTTGGATA AAAACAAGCCTGAGCTTCCAGAGATTAG 72 72 72 72 65 65 65 65 GTTTTTTCTTCTTCTATATTGATGATTTTG CGACACCGGCACCTATATGCACCGGT GAAACTTCTTTAGTTGAACGCGAATTGGA AGGAGAAGAGAGCAGTACCTCTCCCTGT 68 68 68 68 60 60 60 60 GAAGCATGCTCGCACGACACCCATCC CGGCAGCGGAGGGGAAGGGGAGCAGT CAACAACCCCAATACCAGGCCAGCTCCACA AACCCTCGACTGCTCAAGGCAGAGCCGC 72 72 72 72 65 65 65 65 TCCATATAGGTCTCTCTTTTCTTATTATG TGAGAGACTCAATACTCATGAATTTCA CTТGGTCCATATAGGTCTCTCTTTTC CCTCCTGATATACTTGAGAGACTCAATA Та1 - температура відпалу в першому циклі; Та2- температура відпалу у циклах, що залишилася ПЛР була проведена в об'ємі 10μл і містила 30нг матричної ДНК, 10пмоль кожного внутрішнього праймера, 1пмоль кожного зовнішнього праймера, 200μМ dNTP, 2,5мМ MgCl2, 1 хбуфер і 1,5 U Taq полімерази (Амплісенс, Росія). ПЛР ампліфікація була проведена в такому режимі: 94°С протягом 2хв., 35 циклів по 1хв. при 94°С, 1хв. відпалу і 1хв. елонгації при 72°С, завершення при 72°С протягом 2хв. Температура відпалу була 72-68°С для першого циклу (див. Табл. 1) і зменшувалася на 1°С доки не досягала значення, вказаного в Табл. 1. Надалі продовжували при цій температурі під час етапів відпалу для циклів, що залишилися. Продукти ПЛР реакції були змішані з 10μл буфера для нанесення. В лунки вносилося по 12μл суміші. Електрофорез проводився в 1,5% агарозному гелі. Візуалізація продуктів ампліфікації відбувалася за допомогою забарвлення етидієм броміду. Виявлено, що маркер MWG2062 утворював три фрагменти різної довжини 101, 151, 198 п.н. Отже цей SNP маркер був поліморфний у цих двох сортів. Для визначення кількісного складу алелів маркера MWG2062 нами була проведена ПЛР у реальному часі з праймерами та зондами, послідовності яких наведено в Табл. 2. Таблиця 2. Найменування MWG2062-325-A-G Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 АВС465-254-С-Т Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 MWG2218-175-G-C Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 MWG502-656-A-G Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 Послідовність (5'→3') Tm Довжина 58 59 65,5 65,8 23 28 21 23 58 59 65 67 28 19 25 27 CATGACCGTGACGACAACGT AGGACTATGGCCGCATCATC TET-5'-CGTCTGTCGTCCACGAAACAAAGC-3'-TAMRA 6FAM-5'-CGTCTGTCGTCGACGAAACAAAGC-3'-TAMRA 59 59 66 65,9 20 20 24 24 AATTGGCATGATTAGTTTGATGGAT CCATAAACGGCGATTAGTTGGA TET-5'-TTAATCCGCTGTTTGGCCGCC-3'-TAMRA 6FAM-5'-TTAATCCGTTGTTTGGCCGCCT-3'-TAMRA 59 60 66,7 65 25 22 21 22 ACGACAAGGAAGTGGTGGAGTAC ATAATACCTGAAAGAACACACTGCATGT TET-5'-TCAAACAAGAACGCCACGCCA-3'-TAMRA 6FAM-5'-CATCAAACAAGAATGCCACGCCA-3'-TAMRA GATATATGTTCCTCTCTGTTGGTCAGAT GCGAGACTCGCCGTTACCT TET-5'-ACCTACCAGGCTGCGGTACTTGAGA-3'-TAMRA 6FAM-5'-CCTACCAGGCTACGGTACTTGAGAGCG-3'-TAMRA 11 ABG601-390-C-T Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 ABC156-231-T-G Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 MWG2029-204-A-G Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 MWG801-344-G-A Прямий праймер Зворотній праймер TaqMan-проба 1 TaqMan-проба 2 56555 12 TGGTTTGCTCCGCTATTAAGTATTC TGCTAAAGGTCATTATGGACTTCTTAGT TET-5'-AAATATCAAGGCTGTCAACGACATGGAA-3'-TAMRA 6FAM-5'-CAAAATATCAAGGCTATCAACGACATGGAA-3'TAMRA 59 58 65,3 25 28 28 65 30 GAGGTCTTGAATAGCTTGGTCCAT GATGATTGATCCTCCTGATATACTTGAG TET-5'-CTCATGAATTCCCACAATAAGAAAAGAGAGACC-3'TAMRA 6FAM-5'-ACTCATGAATTCCAACAATAAGAAAAGAGAGACC3'-TAMRA 59 24 66,3 33 65,2 34 58 58 65,5 23 23 28 66 30 60 58 67,4 65,7 19 22 18 19 GCCCAACTGTTCATGTTAGGTTT CAGCGTAGAACTGCAATTTGGTA TET-5'-TTGATGATTGTGCAGGTGCATATAGGTG-3'-TAMRA 6FAM-5'-TTGATGATTGTACAGGTGCATATAGGTGCC-3'TAMRA GCTCTCCCTGTGCGGTTGT CGAGGAACCAACTCTCACTCAA TET-5'-ACACCCACCGCCGCTCCC-3'-TAMRA 6FAM-5'-CGACACCCACCACCGCTCC-3'-TAMRA Метод TaqMan Assay використовує 5'нуклеазну активність Taq полімерази для виявлення флуоресценції, яку спричинює мітка під час або після ПЛР реакції. Для генотипування SNP використовується одна пара TaqMan-проб і одна пара праймерів. Кожна проба складається з приблизно 30 п.н. олігонуклеотида, комплементарного поліморфному району. Дві TaqMan-проби розрізняються між собою в поліморфному сайті. Одна з проб комплементарна алелю 1, друга - алелю 2. 5'-флуоресцентна мітка (6-карбокси-4, 7, 2', 7'тетрахлорфлуоресценін, ТЕТ) і 3'- гаситель (6карбокси-N, N, N', N'-тетрахлорофлуоресцин, TAMRA) були ковалентно зв'язані із пробою 1. Також і проба 2 була мічена 5'флуоресцентною міткою (6-карбоксифлуоресцин, FAM) та таким же 3'гасителем (TAMRA). Коли TaqMan-проба інтактна, флуоресценція гаситься за рахунок близького розташування гасителя. Під час етапу відпалу ПЛР TaqMan-проба гібридизується з поліморфним сайтом в районі між прямим та зворотнім праймерами. Під час елонгації в ПЛР 5'-флуоресцентна міт Комп’ютерна верстка М. Мацело ка відділяється за рахунок 5'-нуклеазної активності Taq полімерази, що призводить до збільшення флуоресценції. ПЛР проводилася на плашці з 96 лунками по 20μл в кожній із концентрацією геномної ДНК 20нг, 300нМ ТЕТ-проби, 50нМ FAM-проби, 50 нМ прямого праймера і 300 нМ зворотнього віх TaqMan мастер мікс. TaqMan мастер мікс - це суміш усіх компонентів окрім праймерів і проб. TaqMan ПЛР була оптимізована шляхом збільшення температури відпалу з 60°С до 65°С. Кінцеві оптимальні умови були такі: 50°С протягом 2хв., 95°С - 10хв., 95°С - 15сек., 64°С - 1хв. для 40 циклів. В результаті ПЛР у реальному часі із зразком ДНК ячменю модельної вибірки були отримані показники відносного рівня флуоресценції для типового алелю - 18750, а для алелю з меншою концентрацією - 190. Відповідно, відношення алелів становило 0,99 до 0,01. Таким чином, сортова чистота оцінюваного зразка складала 99%, чого ми і очікували при формуванні модельної вибірки. Підписне Тираж 23 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for determining varietal purity and typicalness of lots of seed of farm crops using dna-markers
Автори англійськоюParii Myroslav Fedorovych, Vdovychenko Zhanna Viktorivna, Spyrydonov Vladyslav Hennadiiovych
Назва патенту російськоюСпособ определения сортовой чистоты и типичности партий семян сельскохозяйственных культур c использованием днк-mapkepob
Автори російськоюПарий Мирослав Федорович, Вдовиченко Жанна Викторовна, Спиридонов Владислав Геннадьевич
МПК / Мітки
МПК: A01H 1/04
Мітки: використанням, чистоти, спосіб, культур, визначення, днк-маркерів, типовості, партій, сортової, насіння, сільськогосподарських
Код посилання
<a href="https://ua.patents.su/6-56555-sposib-viznachennya-sortovo-chistoti-ta-tipovosti-partijj-nasinnya-silskogospodarskikh-kultur-z-vikoristannyam-dnk-markeriv.html" target="_blank" rel="follow" title="База патентів України">Спосіб визначення сортової чистоти та типовості партій насіння сільськогосподарських культур з використанням днк-маркерів</a>
Спосіб визначення рівномірності розподілу насіння при сівбі сільськогосподарських культур

Номер патенту: 29795
Опубліковано: 25.01.2008
Автори: Насонов Василь Андрійович, Васківнюк Юрій Миколайович, Білоткач Михайло Петрович, Цибуля Микола Григорович, Романенко Михайло Пилипович
МПК: G01B 5/26
Мітки: сільськогосподарських, насіння, спосіб, культур, визначення, сівби, розподілу, рівномірності
Формула / Реферат:
Спосіб визначення рівномірності розподілу насіння по площі при сівбі сільськогосподарських культур, який включає використання облікової рамки певних розмірів з загальною обліковою площею S, підрахунок кількості рослин n в межах площі облікової рамки і визначення оцінного показника k, що характеризує рівномірність розміщення рослин по площі, який відрізняється тим, що на обліковій рамці закріплюють прозору плівку та на плівці позначають...
Спосіб підвищення /збереження/ сортової типовості однодомних конопель

Номер патенту: 6653
Опубліковано: 16.05.2005
Автори: Лайко Ірина Михайлівна, Ситник Василь Петрович, Вировець В'ячеслав Гаврилович, Щербань Іван Іванович
МПК: A01H 1/04
Мітки: конопель, сортової, збереження, підвищення, спосіб, типовості, однодомних
Формула / Реферат:
Спосіб підвищення (збереження) сортової типовості однодомних конопель, який включає посів для розмноження селекційного і насіннєвого матеріалу без вмісту чоловічих рослин (звичайної плосконі), який відрізняється тим, що процес підвищення (збереження) сортової типовості здійснюється за рахунок посіву насіння потомків окремих рослин (родин) та насіннєвого матеріалу однодомних конопель у яких попередньо була проведена оцінка генотипу за статевим...
Спосіб протруювання насіння сільськогосподарських культур

Номер патенту: 21428
Опубліковано: 15.03.2007
Автори: Крупський Артур Євгенович, Діндорого Володимир Григорович, Клименко Ірина Іванівна
МПК: A01C 1/00
Мітки: спосіб, протруювання, насіння, культур, сільськогосподарських
Формула / Реферат:
Спосіб протруювання насіння сільскогосподарських культур, що включає обробку їх протруйниками системної дії (фунгіцидами), який відрізняється тим, що при висіві в день протруєння використовують повну норму фунгіциду на одиницю маси насіння або зменшують її до 1/4 частини при сумісному внесенні в робочу рідину для протруєння біопрепаратів (бактеріальних добрив) та регуляторів росту (як окремо, так і у суміші), або насіння попередньо озонують...
Спосіб класифікації генетичної чистоти сільськогосподарських культур

Номер патенту: 57636
Опубліковано: 16.06.2003
Автор: Клявзо Сергій Павлович
МПК: A01H 1/04
Мітки: культур, генетично, сільськогосподарських, спосіб, класифікації, чистоти
Формула / Реферат:
Спосіб класифікації генетичної чистоти сільськогосподарських культур, при якому виділяють білкові фракції, підготовляють їх до електрофорезу, проводять електрофорез, проявляють компоненти білка в спектрі, визначають частоти наявності білкових компонентів за відносними електрофоретичними рухомостями, складають білкові формули, який відрізняється тим, що класифікацію генетичної чистоти сільськогосподарських культур проводять за коефіцієнтом...
Спосіб передпосівної обробки насіння сільськогосподарських культур

Номер патенту: 18229
Опубліковано: 15.11.2006
Автори: Герасько Тетяна Володимирівна, Калитка Валентина Василівна, Заславський Олексій Маркович, Малахова Тетяна Олексіївна
МПК: A01N 25/02, A01C 1/00, A01C 1/06
Мітки: насіння, спосіб, культур, обробки, передпосівної, сільськогосподарських
Формула / Реферат:
Спосіб передпосівної обробки насіння сільськогосподарських культур, що передбачає обробку насіння композицією на основі фунгіциду та регулятора росту, який відрізняється тим, що обробку здійснюють композицією, що містить фунгіцид бенлат (фундазол, 50% змочуваний порошок (з.п.)), як регулятор росту - антиоксидант дистинол, суміш поліетиленоксиду 400 і поліетиленоксиду 1500, і воду, при такому співвідношенні компонентів, мас. %: ...