Спосіб дослідження дихання мітохондрій in situ
Номер патенту: 60528
Опубліковано: 25.06.2011
Автори: Манько Володимир Васильович, Манько Богдан Олексійович, Мерлавський Володимир Михайлович, Великопольська Ольга Юріївна



Формула / Реферат
Спосіб дослідження дихання мітохондрій in situ, за яким пермеабілізують плазматичну мембрану клітин неіонним детергентом-дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, який відрізняється тим, що пермеабілізацію здійснюють оптимальною кількістю детергенту, яка знаходиться у сталому співвідношенні з кількістю досліджуваних клітин, яке для кожної тканини і для кожного типу клітин визначають емпірично.
Текст
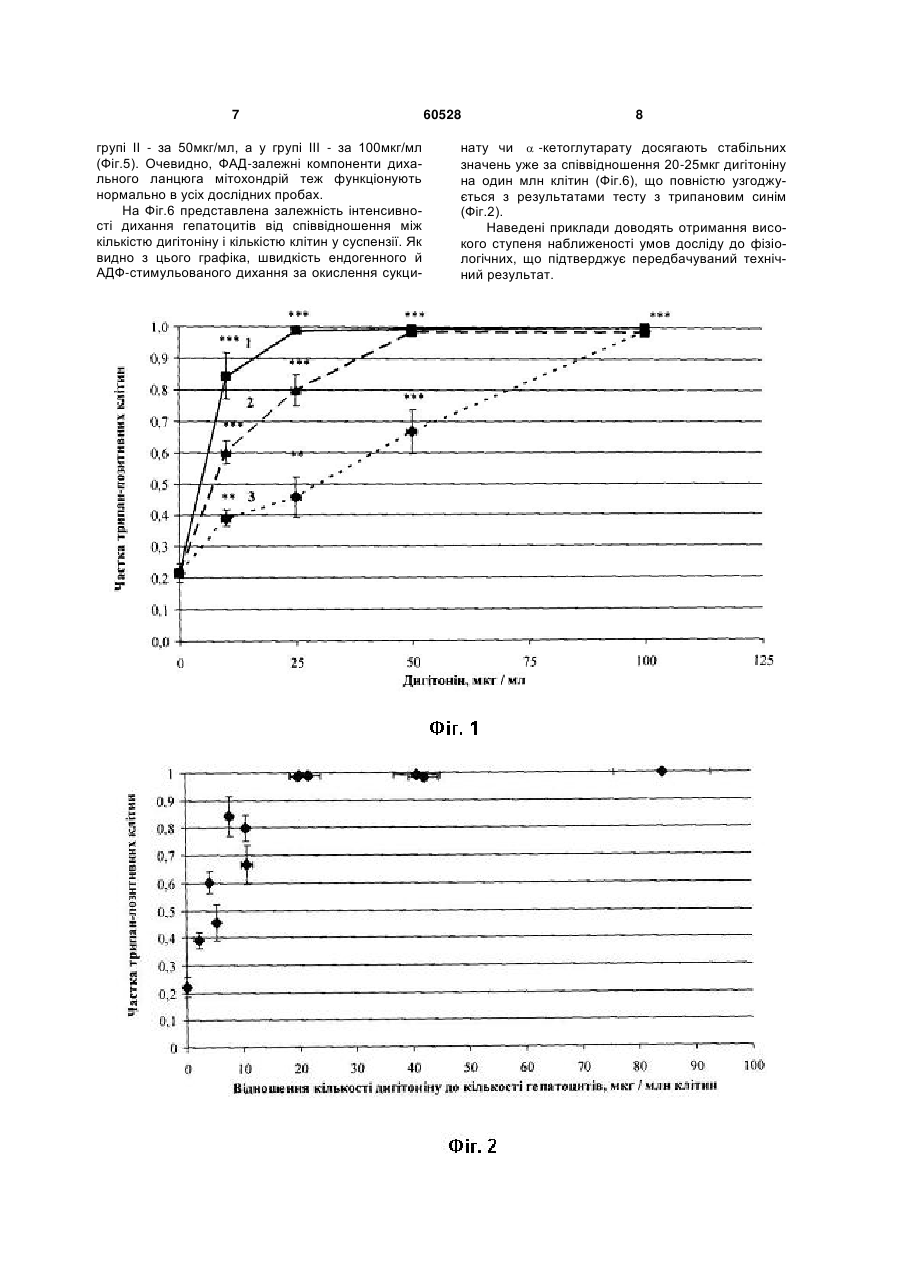
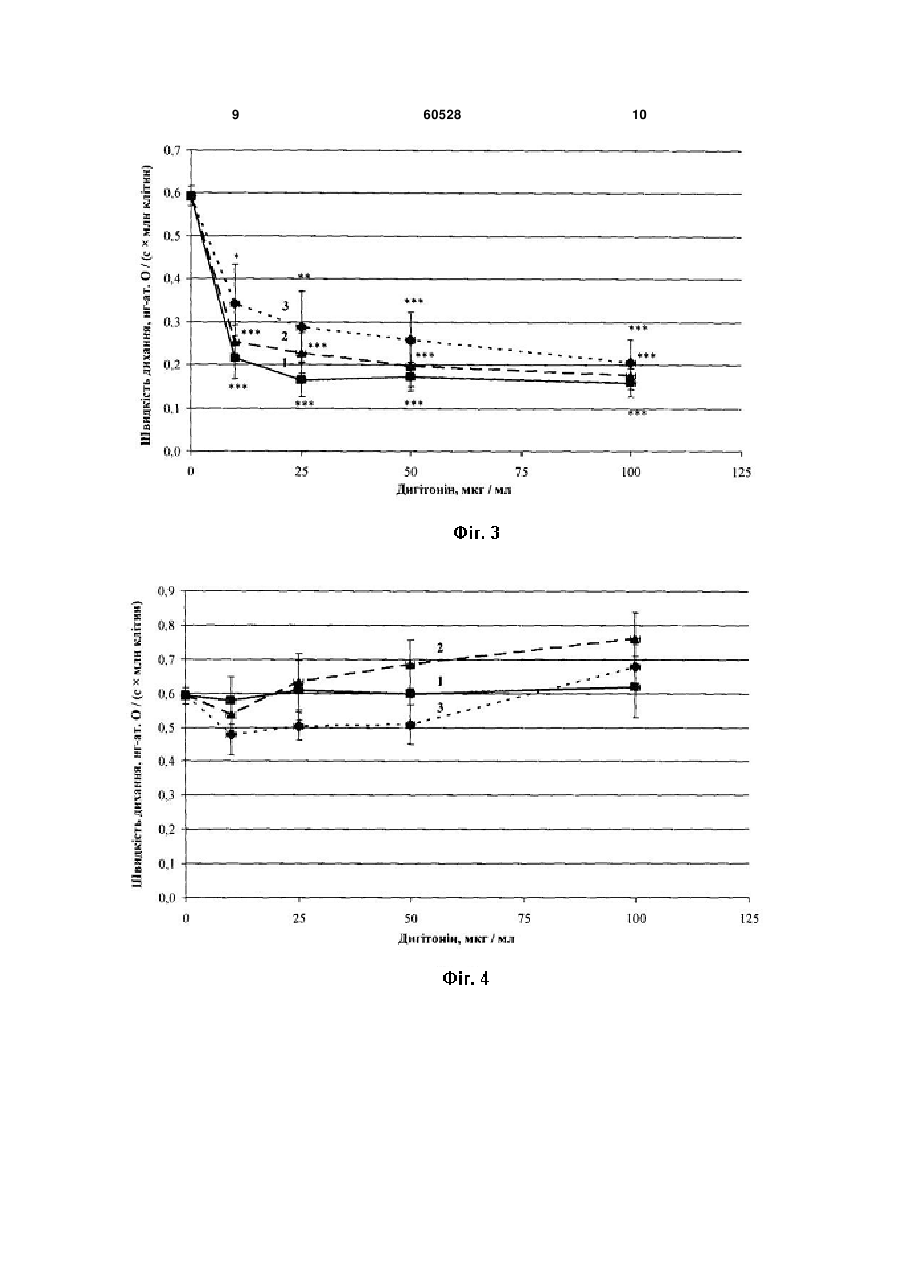
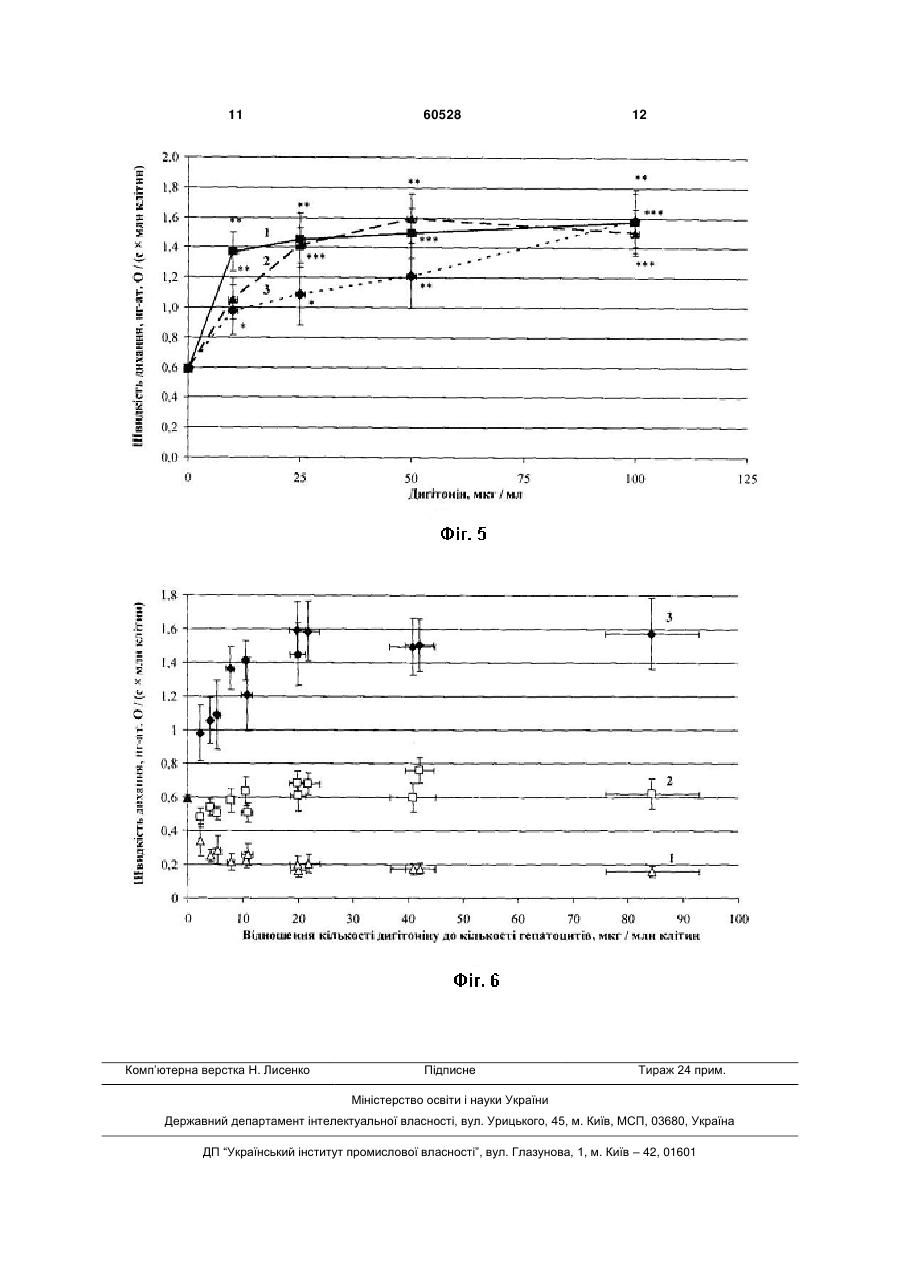
Спосіб дослідження дихання мітохондрій in situ, за яким пермеабілізують плазматичну мем брану клітин неіонним детергентом-дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, який відрізняється тим, що пермеабілізацію здійснюють оптимальною кількістю детергенту, яка знаходиться у сталому співвідношенні з кількістю досліджуваних клітин, яке для кожної тканини і для кожного типу клітин визначають емпірично. Корисна модель стосується галузі біології, а саме клітинної фізіології та може бути використана для дослідження процесів мітохондріального окислення в умовах, максимально наближених до фізіологічних. Відомий спосіб дослідження мітохондріального дихання на ізольованих мітохондріях (Chance В., Williams G. R. Respiratory enzymes in oxidative phosphorylation. Kinetics of oxygen utilization // J. Biol. Chem. 1955. Vol. 217. P. 383-393). Ізольовані мітохондрії отримують шляхом диференційного центрифугування. Для вивчення окисних процесів швидкість дихання реєструють полярографічним методом за п'яти метаболічних станів мітохондрій, в кожному з яких потік електронів дихальним ланцюгом лімітується різними чинниками: або доступністю субстратів окислення (стан 1), або концентрацією АДФ і фосфату, спряженою з окисним фосфорилюванням (стан 2), або концентрацією кисню у випадку його дефіциту (стан 5). За наявності всіх необхідних інгредієнтів кінетичний контроль дихання здійснюється надлишком позамітохондріального АДФ (стан 3, або «активний»). Після фосфорилювання АДФ встановлюється новий, більш енергізований стан 4. Недоліком цього способу є втрата зв'язків ізольованих мітохондрій з іншими клітинними органелами, що не дає змоги дослідити усі аспекти регуляції клітинного дихання, бо мітохондрії можуть знаходитися в тісному структурному і функціональному зв'язку з ендоплазматичним ретикулумом і комплексом Гольджі (Dolman N. J. et al. Stable Golgi-mitochondria complexes and formation of Golgi Ca gradients in pancreatic acinar cells // J. Biol. Chem. 2005. Vol. 280, № 16. P. 15794-15799; Rizzuto R., Pozzan T. Microdomains of intracellular 2+ Ca : molecular determinants and functional consequences // J. Physiol. Rev. 2006. Vol. 86. P. 2+ 369-408; Манько В.В. Концепція Са функціональних одиниць у застосуванні до секреторних клітин слинних залоз личинки Chironomus plumosus II Біологічні Студії / Studia Biologica. 2008. Т.2, №1. С.33-50), а інтенсивність окисного фосфорилювання регулюється сусідніми органелами, наприклад, шляхом зміни споживання АТФ 2+ чи концентрації Са в цитозолі (Rizzuto R., Pozzan 2+ T. Microdomains of intracellular Ca : molecular determinants and functional consequences // J. Physiol. Rev. 2006. Vol. 86. P. 369-408). Найближчим за технічною сутністю - прототипом - є спосіб дослідження дихання пермеабілізованих волокон скелетних м'язів зі застосуванням дигітоніну (Saks V. A. et al. Permeabilised cell and skinned fiber techniques in studies of mitochondrial function in vivo II Мої. Cell. Biochem. 1998. Vol. 184. P. 81-100). Пермеабілізація здійснюється для забезпечення проникнення екзогенних агентів, що в нормі не проникають крізь плазматичну мембрану і впливають на мітохондріальне дихання. Такими агентами можуть бути субстрати циклу Кребса, регулятори його ферментів, аденілові нуклеотиди, інгібітори комплексів дихального ланцюга тощо. Цей спосіб дає змогу зберегти взаємозв'язки між мітохондріальним диханням та іншими клітинними процесами. (19) UA (11) 60528 (13) U 2+ 3 Недоліком способу є відсутність системного підходу до визначення оптимальної концентрації детергенту, оскільки у недостатніх концентраціях дигітонін пермеабілізує плазматичну мембрану не всіх клітин, а у надмірних - може ушкоджувати мембрани мітохондрій і ендоплазматичного ретикулуму (Cook G. A. et al. Structural changes of isolated hepatocytes during treatment with digitonin // Biochim. Biophys. Acta. 1983. Vol. 763. P. 356-367). В основу корисної моделі поставлено завдання удосконалити «Спосіб дослідження дихання мітохондрій in situ» шляхом встановлення оптимальної концентрації неіонного детергенту дигітоніну, що дасть змогу підвищити ступінь наближеності умов досліду до фізіологічних. Поставлене завдання вирішується так, що у способі дослідження дихання мітохондрій in situ, за яким пермеабілізують плазматичну мембрану клітин неіонним детергентом - дигітоніном, контролюють ступінь проникності плазматичної мембрани і полярографічно реєструють швидкість поглинання кисню, причому пермеабілізацію здійснюють оптимальною кількістю детергенту, яка знаходиться у сталому співвідношенні з кількістю досліджуваних клітин, яке для кожної тканини і для кожного типу клітин визначають емпірично. Відомо, що дигітонін зв'язується із холестерином і утворює з ним нерозчинні комплекси, що й призводить до виникнення пор (Nishikawa M. et al. Interaction of digitonin and its analogs with membrane cholesterol // Biochem. J. 984. Vol. 96, № 4. P. 12311239). Вважається, що дигітонін досить селективно діє саме на плазматичну мембрану, не порушуючи цілісності інших мембран клітини, оскільки вміст холестерину в них на кілька порядків нижчий (Colbeau A., Nachbaur J., Vignais P. M. Enzymic characterization and lipid composition of rat liver subcellular membranes // Biochim. Biophys. Acta. 1971. Vol. 249 (2). P. 462-492). Доведено, що у низьких концентраціях (50-150мкг/мл) він не спричиняє порушень ультраструктури внутрішньоклітинних органел і цитоскелету (Fiskum G. et al. The cytoskeleton of digitonin-treated rat hepatocytes // Proc. Nati. Acad. Sci. USA. 1980. Vol. 77, № 6. P. 3430-3434). Проте за концентрації 800мкг/мл дигітонін суттєво пошкоджує внутрішньоклітинні органели, що підтверджено зміною ультраструктури цих органел і виходом їхніх маркерних ферментів у середовище. Для повної селективної пермеабілізації плазматичної мембрани, наприклад, гепатоцитів дослідники використовують дигітонін у досить широкому діапазоні концентрацій. Так, за даними Холмухамедова, дигітонін здатний повністю пермеабілізувати плазматичну мембрану гепатоцитів вже у концентрації 10мкг/мл, а у концентрації 100мкг/мл він призводить до ушкодження мітохондріальних мембран (Холмухамедов Э.Л. Роль митохондрий в обеспечении нормальной жизнедеятельности и выживания клеток млекопитающих // Автореферат дисс. ... д-ра биол. наук. Пущино, 2008. 35с.). Інші дослідники показали, що у концентрації 10мкг/мл дигітонін взагалі не змінює цілісність плазматичної мембрани гепатоцитів і порушує її за концентрації 50-150мкг/мл, не впливаючи 60528 4 на стан мітохондрій (Fiskum G. et al. The cytoskeleton of digitonin-treated rat hepatocytes // Proc. Nati. Acad. Sci. USA. 1980. Vol. 77, № 6. P. 3430-3434). Ці неузгодженості, очевидно, пов'язані із різними концентраціями клітин у суспензії, що використовувались дослідниками. Авторами вперше запропоновано враховувати співвідношення між кількостями дигітоніну і досліджуваних клітин, що дало змогу встановити оптимальну концентрацію детергенту для пермеабілізацІЇ плазматичної мембрани і підвищити ступінь наближеності умов досліду до фізіологічних. Фіг.1. Залежність ступеня пермеабілізацІЇ плазматичної мембрани гепатоцитів від концентрації дигітоніну за різної густини суспензії: 1 - 0,9-1,7млн/мл гепатоцитів; 2 - 2,0-3,0млн/мл гепатоцитів; 3 - 4,0-5,6млн/мл гепатоцитів; тут і надалі: * - статистично вірогідна різниця порівняно з контролем (Р 0,05); ** - статистично вірогідна різниця порівняно з контролем (Р 0,01); *** - статистично вірогідна різниця порівняно з контролем (Р 0,001). Фіг.2. Залежність ступеня пермеабілізації плазматичної мембрани гепатоцитів від відношення кількості дигітоніну до кількості гепатоцитів Фіг.3. Залежність швидкості ендогенного дихання пермеабілізованих гепатоцитів від концентрації дигітоніну за різної густини суспензії: 1 - 0,9-1,7млн/мл гепатоцитів; 2 - 2,0-3,0млн/мл гепатоцитів; 3 - 4,0-5,6млн/мл гепатоцитів Фіг.4. Залежність швидкості дихання пермеабілізованих гепатоцитів, стимульованого кетоглутаратом і АДФ, від концентрації дигітоніну за різної густини суспензії: 1 - 0,9-1,7млн/мл гепатоцитів; 2 - 2,0-3,0млн/мл гепатоцитів; 3 - 4,0-5,6млн/мл гепатоцитів Фіг.5. Залежність швидкості дихання пермеабілізованих гепатоцитів, стимульованого сукцинатом і АДФ, від концентрації дигітоніну за різної густини суспензії: 1 - 0,9-1,7млн/мл гепатоцитів; 2 - 2,0-3,0млн/мл гепатоцитів; 3 - 4,0-5,6млн/мл гепатоцитів Фіг.6. Залежність швидкості дихання пермеабілізованих гепатоцитів від відношення кількості дигітоніну до кількості гепатоцитів: 1 - ендогенне дихання; 2 - -кетоглутарат + АДФ; 3 - сукцинат + АДФ Спосіб можна проілюструвати на прикладі дослідження мітохондріального дихання гепатоцитів in situ. Гепатоцити ізолювали за двостадійним методом Сеглена (Seglen P. О. Preparation of isolated rat liver cells // Methods Cell Biol. 1976. Vol. 13. P. 29-83). Підрахунок гепатоцитів здійснювали за допомогою камери Горяєва. Для перевірки цілісності плазматичних мембран клітини фарбували 0,1% розчином трипанового синього. Відразу після ізоляції кількість клітин з цілісними плазматичними мембранами становила 75-90%. Пермеабілізацію 5 здійснювали за допомогою дигітоніну у внутрішньоклітинному розчині протягом 10хв. Після цього клітини тричі відмивали від дигітоніну і метаболітів, які вийшли з цитозолю внаслідок пермеабілізації, центрифугуючи при 50g. Цілісність плазматичної мембрани гепатоцитів після пермеабілізації оцінювали тестом з трипановим синім, а також за рівнем ендогенного дихання та дихання, стимульованого субстратами окислення і АДФ. Останнє також використовували як критерій для оцінки інтактності мітохондрій. Швидкість поглинання кисню визначали полярографічним методом, який ґрунтується на реєстрації електрохімічного відновлення фізично розчиненого кисню на катоді за різниці потенціалів 0,60,7В: + О2+2Н +2е =Н2О2, + Н2О2+2Н +2е =2Н2О. Величину дифузного струму реєстрували за допомогою полярографічної установки, зібраної на базі закритого електрода Кларка, полярографа YSI 5300, самописця КСП-4, магнітної мішалки для розмішування суспензії та скляної термостатованої закритої комірки об'ємом 1,6мл. Поглинання кисню пермеабілізованими гепатоцитами реєстрували за окислення ендогенних субстратів, кетоглутарату (1ммоль/л) і сукцинату (0,35ммоль/л) при температурі 37°С. Дихання стимулювали додаванням 1,2мкмоль АДФ (концентрація у полярографічній комірці 750мкмоль/л). Базовий позаклітинний розчин мав такий склад, ммоль/л: NaCl -140,0, КСІ - 4,7, СаСl2 - 1,3, MgCl2 - 1,0, HEPES - 10,0, глюкоза - 5,0; рН 7,4. На певних етапах виділення гепатоцитів використовували модифіковані розчини: 1) без СаСl2; 2) з додаванням колагенази (120од./мл) і СаСІ2 (3 ммоль/л). Внутрішньоклітинний розчин мав такий склад, ммоль/л: КСІ -90,0, NaCl - 15,0, КН2РО4 2,0, MgCl2 - 1,0, HEPES-10,0, ЕГТА - 0,5; рН 7,2. Склад розчину відповідає іонному складу внутрішньоклітинного середовища гепатоцитів (Tanaka A., Chance В., Quistorffs В. A possible role of inorganic phosphate as a regulator of oxidative phosphorylation in combined urea synthesis and gluconeogenesis in perfused rat liver. A phosphorus magnetic resonance spectroscopy study // J. Biol. Chem. 1989. Vol. 264, № 17. P. 10034-10040; Gasbarrini A. et al. Effect of + 2+ 2+ anoxia on intracellular ATP, Na ,-, Ca j, Mg j and cytotoxicity in rat hepatocyta // J. Biol. Chem. 1992. Vol. 267, № 10. P. 6654-6663; Wang K... Wondergem R. Hepatocyte water volume and potassium activity during nypotonic stress // 1. Membr. Biol. 1993. Vol. 135 (2). P. 137-144). Необхідні статистичні розрахунки проводили за допомогою комп'ютера з використанням пакету програм Microsoft Office Excel. Для встановлення залежності ступеня пермеабілізації від співвідношення між концентрацією дигітоніну і кількістю гепатоцитів у суспензії змінювали обидва параметри. Тобто, дигітонін використовували у кінцевих концентраціях 10, 25, 50 і 100мкг/мл, додаючи його до суспензій з різною кількістю гепатоцитів. Згодом експериментальні дані були згруповані за кількістю клітин у суспензії у три дослідні групи. В одному мл суспензії дослідної групи І було від 0,9 до 1,7млн клітин (в серед 60528 6 ньому 1,27±0,09млн), дослідної групи II - 2,03,0млн клітин (2,45 ±0,11млн), а дослідної групи III - 4,0-5,6млн клітин (4,67±0,46млн). За відсутності дигітоніну (контроль) 21,67±2,94% гепатоцитів зафарбовувались трипановим синім (Фіг.1). У дослідній групі І повна пермеабілізація плазматичної мембрани спостерігалась вже за концентрації дигітоніну 25мкг/мл, у дослідній групі II - за 50мкг/мл, а в групі III - лише за концентрації 100мкг/мл (Фіг.1). За додавання детергенту у нижчих концентраціях у всіх трьох групах пермеабілізація плазматичної мембрани відбувалась лише у частини гепатоцитів (Фіг.1). Отже, густина суспензії суттєво впливає на ступінь пермеабілізації плазматичної мембрани дигітоніном: чим більша кількість гепатоцитів, тим більшою повинна бути концентрація дигітоніну. Ця залежність є прямопропорційною у досліджуваному діапазоні. Для кращої візуалізації цього факту побудовано графік залежності ступеня пермеабілізації гепатоцитів від співвідношення між кількістю дигітоніну та кількістю клітин у суспензії (Фіг. 2). Як видно із графіка, для повної пермеабілізації гепатоцитів достатньо 20-25мкг дигітоніну на один млн клітин. Крім тесту з трипановим синім, ступінь пермеабілізації оцінювали за швидкістю ендогенного дихання та дихання, стимульованого субстратами окислення й АДФ Відомо, що клітинам із порушеною цілісністю плазматичної мембрани притаманний низький рівень ендогенного дихання (Гулак П.В., Дудченко А.М., Зайцев В.В. и др. Гепатоцит: функционально-метаболические свойства. М.: Наука, 1985. 272с.). Причиною цього є зниження концентрації субстратів окислення у цитоплазмі внаслідок їхньої дифузії у позаклітинне середовище. Обробка дигітоніном призводила до зниження швидкості ендогенного дихання гепатоцитів. У дослідній групі і швидкість ендогенного дихання досягала мінімуму за концентрації дигітоніну 25мкг/мл, у групі II - за концентрації 50мкг/мл, а у групі III - лише за концентрації 100мкг/мл (Фіг.3). Ці дані повністю узгоджуються із результатами тесту з трипановим синім. За внесення -кетоглутарату й АДФ у полярографічну комірку рівень дихання пермеабілізованих гепатоцитів у всіх випадках був такий самий, як і інтактних (Фіг.4). Тобто, внесення цього субстрату відновлювало дихання пермеабілізованих клітин. Причому, це не залежало від ступеня пермеабілізації. Відомо, що в інтактних клітинах дихання в основному відбувається завдяки окисленню НАД-залежних субстратів. Отже, відновлення -кетоглутаратом дихання до рівня інтактних гепатоцитів доводить те, що дигітонін не порушує функціонування НАД-залежних компонентів дихального ланцюга мітохондрій. Сукцинат значно сильніше стимулював дихання пермеабілізованих клітин, причому залежність інтенсивності АДФ-стимульованого дихання за окислення сукцинату від концентрації дигітоніну та кількості клітин добре корелює з результатами тесту з трипановим синім: Інтенсивність дихання у дослідній групі І досягала максимуму за використання дигітоніну у кінцевій концентрації 25мкг/мл, у 7 групі II - за 50мкг/мл, а у групі III - за 100мкг/мл (Фіг.5). Очевидно, ФАД-залежні компоненти дихального ланцюга мітохондрій теж функціонують нормально в усіх дослідних пробах. На Фіг.6 представлена залежність інтенсивності дихання гепатоцитів від співвідношення між кількістю дигітоніну і кількістю клітин у суспензії. Як видно з цього графіка, швидкість ендогенного й АДФ-стимульованого дихання за окислення сукци 60528 8 нату чи -кетоглутарату досягають стабільних значень уже за співвідношення 20-25мкг дигітоніну на один млн клітин (Фіг.6), що повністю узгоджується з результатами тесту з трипановим синім (Фіг.2). Наведені приклади доводять отримання високого ступеня наближеності умов досліду до фізіологічних, що підтверджує передбачуваний технічний результат. 9 60528 10 11 Комп’ютерна верстка Н. Лиcенко 60528 Підписне 12 Тираж 24 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for study of mitochondria breathing in situ
Автори англійськоюManko Bohdan Oleksiiovych, Merlavskyi Volodymyr Mykhailovych, Velykopolska Olha Yuriivna
Назва патенту російськоюСпособ исследования дыхания митохондрий in situ
Автори російськоюМанько Богдан Алексеевич, Мерлавский Владимир Михайлович, Великопольская Ольга Юрьевна
МПК / Мітки
МПК: G01N 33/50, A01N 61/00
Мітки: мітохондрій, дихання, дослідження, спосіб
Код посилання
<a href="https://ua.patents.su/6-60528-sposib-doslidzhennya-dikhannya-mitokhondrijj-in-situ.html" target="_blank" rel="follow" title="База патентів України">Спосіб дослідження дихання мітохондрій in situ</a>
Спосіб встановлення ступеня токсичності комбінованих пестицидів за допомогою визначення швидкості і ефективності дихання мітохондрій печінки

Номер патенту: 41847
Опубліковано: 10.06.2009
Автори: Стегній Борис Тимофійович, Жукова Ірина Олексійовна, Малінін Олег Олексійович
МПК: G01N 27/48
Мітки: пестицидів, мітохондрій, спосіб, встановлення, комбінованих, допомогою, печінки, ступеня, швидкості, визначення, дихання, ефективності, токсичності
Формула / Реферат:
Спосіб встановлення ступеня токсичності комбінованих пестицидів за допомогою визначення швидкості і ефективності дихання мітохондрій печінки, що включає гомогенізацію, інкубацію, вимірювання до додавання АДФ (адинозиндифосфат), після додавання АДФ, перетворення його в АТФ та визначення коефіцієнтів Ларді-Вельмана, Чанса-Вільямса і фосфорилювання, який відрізняється тим, що використовують як окисник буфер-трис-HCL, використовують стандартні...
Спосіб прогнозування рівня онкозахворюваності органів дихання залежно від забруднення атмосфери бенз(а)піреном

Номер патенту: 44924
Опубліковано: 26.10.2009
Автори: Баленко Ніна Василівна, Першегуба Ярослав Володимирович, Черниченко Ігор Олексійович, Литвиченко Ольга Миколаївна
МПК: G01N 31/00, A61B 10/00
Мітки: онкозахворюваності, забруднення, залежно, рівня, спосіб, органів, дихання, прогнозування, бенз(а)піреном, атмосфери
Формула / Реферат:
Спосіб прогнозування рівня онкозахворюваності органів дихання залежно від забруднення атмосфери бенз(а)піреном, що включає визначення середньорічної концентрації забруднюючих атмосферу шкідливих речовин, аналіз динаміки показників забруднення і захворюваності, розрахунок коефіцієнтів кореляції між ними за ідентичними часовими інтервалами і послідовним зміщенням у часі та визначення латентного періоду розвитку онкозахворювань, який...
Спосіб визначення функціонального стану системи зовнішнього дихання

Номер патенту: 12604
Опубліковано: 15.02.2006
Автори: Паненко Анатолій Васильович, Романчук Олександр Петрович
МПК: A61B 10/00, G01N 33/48, A61B 5/00
Мітки: спосіб, визначення, зовнішнього, дихання, системі, стану, функціонального
Формула / Реферат:
Спосіб визначення функціонального стану системи зовнішнього дихання (ФССЗД), що включає дослідження параметрів дихання та їх оцінку, який відрізняється тим, що після реєстрації параметрів паттерну та варіабельності дихання проводять оцінку з попередньою їх частковою кореляцією та визначенням рангу з присвоюванням відповідного бала, після чого за сумою балів визначають рівень вегетативного забезпечення зовнішнього дихання та рівень функції...
Фармацевтична композиція, що містить похідну гідроксамової кислоти, спосіб лікування з її використанням та протектор мітохондріального геному та/або мітохондрій

Номер патенту: 57712
Опубліковано: 15.07.2003
Автори: Шумегі Балаш, Літераті Надь Петер, Мареска Бруно, Віг Ласло
МПК: A61P 25/28, A61K 31/16, A61P 9/00
Мітки: лікування, геному, спосіб, містить, мітохондріального, гідроксамової, протектор, композиція, фармацевтична, кислоти, використанням, мітохондрій, похідну
Формула / Реферат:
1. Фармацевтична композиція для захисту мітохондріального геному та/або мітохондрії від пошкоджень або для лікування захворювань, що пов'язані з такими пошкодженнями, яка включає від 0,1 до 95 мас.% похідної гідроксамової кислоти формули І: І,де R1 є атомом водню або С1-5 алкільною групою,R2 є атомом водню, С1-5 алікільною групою, С3-8 циклоалкільною групою або фенільною групою, яка необов'язково може бути заміщена...
Спосіб забезпечення аналгезії при амбулаторних та малоінвазивних втручаннях за умови збереження самостійного дихання

Номер патенту: 24670
Опубліковано: 10.07.2007
Автори: Сергійчук Олена Віталіївна, Гомон Микола Лонгвинович, Шлапак Ігор Порфирович, Дацюк Олександр Іванович, Скиба Андрій Георгійович
МПК: A61K 31/00
Мітки: забезпечення, амбулаторних, дихання, самостійного, умови, збереження, втручаннях, аналгезії, малоінвазивних, спосіб
Формула / Реферат:
Спосіб забезпечення аналгезії при амбулаторних та малоінвазивних втручаннях за умови збереження самостійного дихання, що передбачає внутрішньовенну загальну анестезію, який відрізняється тим, що додатково одноразово болюсно вводять розчин реміфентанілу в дозі 75-200 мсд дорослому хворому в діагностичній кімнаті за наявності моніторування функції дихання та проведення відповідних заходів при її пригніченні.
Попередній патент: Горілка особлива “франкофф сілвер”
Наступний патент: Процес отримання монокристалів телуру
Випадковий патент: Пристрій для створення поперечного магнітного поля при дуговому зварюванні і наплавленні