Спосіб виробництва біомаси гідробіонтів





Формула / Реферат
1. Спосіб виробництва біомаси гідробіонтів шляхом культивування методом квазібезперервної непропорційної проточної культури, який включає підготовку поживного середовища і його протоку через культиватор, внесення біомаси, забезпечення заданими температурою й освітленістю, збір урожаю, який відрізняється тим, що для культиватора попередньо визначають робочий діапазон щільності культури (Вmах![]() Вmіn), а в процесі культивування вимірюють щільність культури, питому швидкість зростання й розраховують зміну щільності культури по формулі:
Вmіn), а в процесі культивування вимірюють щільність культури, питому швидкість зростання й розраховують зміну щільності культури по формулі:
![]() ,
,
де
В0 - поточна концентрація клітин (біомаса) у культиваторі, г/л;
![]() - питома швидкість зростання мікроводоростей;
- питома швидкість зростання мікроводоростей;
К - коефіцієнт непропорційності;
![]() - питома швидкість протоки;
- питома швидкість протоки;
t-tн - час культивування;
потім порівнюють отриману величину з діапазоном (Вmах![]() Вmіn) і підтримують щільність культивованої культури в заданому діапазоні, змінюючи питому швидкість протоки.
Вmіn) і підтримують щільність культивованої культури в заданому діапазоні, змінюючи питому швидкість протоки.
2. Спосіб за п. 1, який відрізняється тим, що, якщо задають питому швидкість протоки ![]() то щільність культивованої культури підтримують у заданому діапазоні, змінюючи коефіцієнт непропорційності.
то щільність культивованої культури підтримують у заданому діапазоні, змінюючи коефіцієнт непропорційності.
3. Спосіб за п. 1, який відрізняється тим, що, якщо К=1, В![]() =Р, де
=Р, де ![]() - питома швидкість протоки середовища через культиватор, то культивування ведуть у стаціонарному динамічному процесі пропорційно-проточної культури.
- питома швидкість протоки середовища через культиватор, то культивування ведуть у стаціонарному динамічному процесі пропорційно-проточної культури.
4. Спосіб за п. 1, який відрізняється тим, що, якщо ![]() =0,
=0, ![]() й культивування ведуть у накопичувальному режимі.
й культивування ведуть у накопичувальному режимі.
Текст
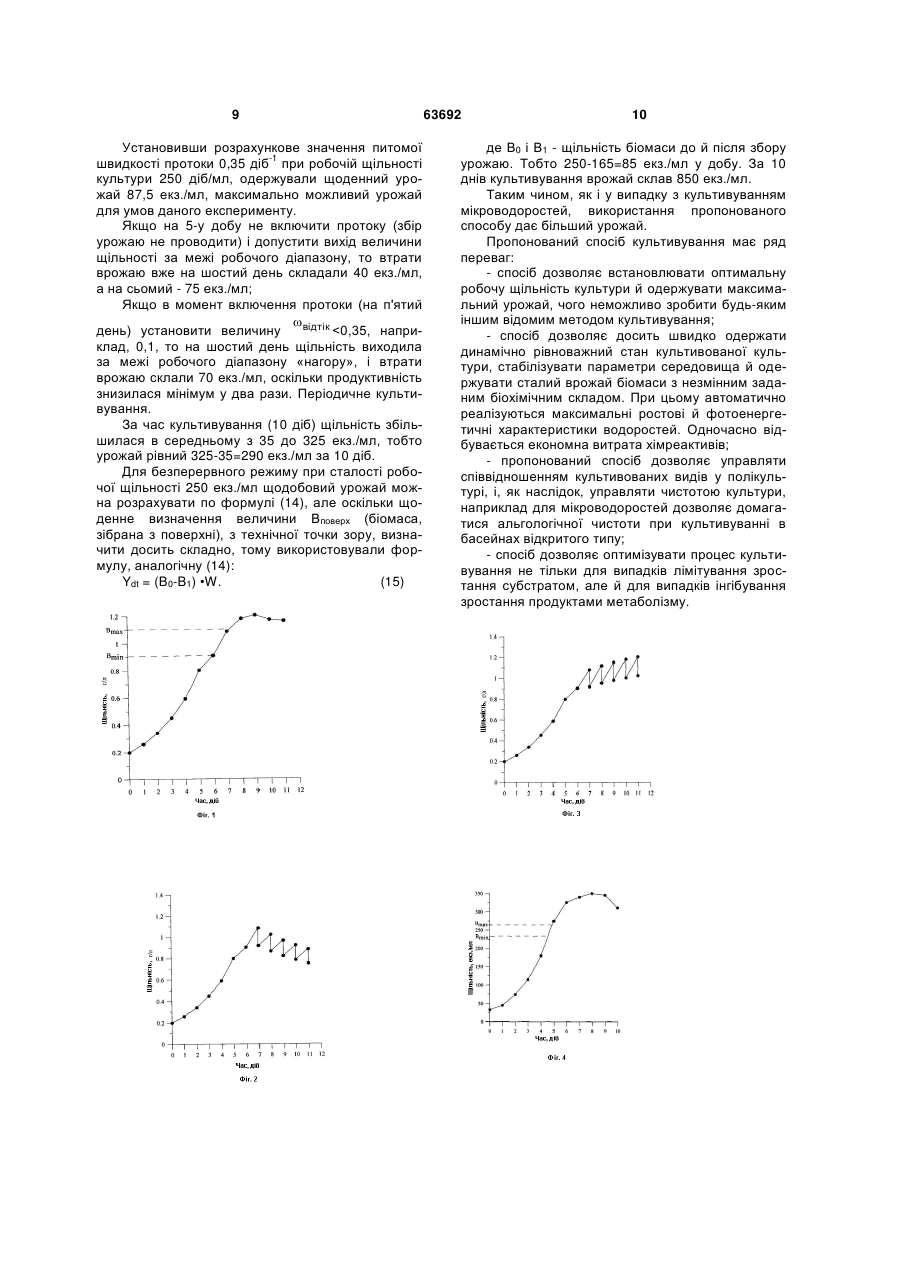
1. Спосіб виробництва біомаси гідробіонтів шляхом культивування методом квазібезперервної непропорційної проточної культури, який включає підготовку поживного середовища і його протоку через культиватор, внесення біомаси, забезпечення заданими температурою й освітленістю, збір урожаю, який відрізняється тим, що для культиватора попередньо визначають робочий діапазон щільності культури (Вmах Вmіn), а в процесі культивування вимірюють щільність культури, питому швидкість зростання й розраховують зміну щільності культури по формулі: 2 3 зазначити, що параметр щоденної заміни середовища був обраний довільно, неформалізованим способом. Основним недоліком відомого способу є низька ефективність внаслідок неоптимальних умов зростання й біосинтезу, що відбивається на кількості зібраної біомаси. Крім того, недоліком також є відсутність можливості керування процесом культивування з метою одержання заданої кількості біомаси з заданим хімічним складом. В основу корисної модулі - спосіб виробництва біомаси гідробіонтів, поставлено задачу підвищення продуктивності широкого спектра культивованих гідробіонтів шляхом забезпечення оптимальних умов зростання. Поставлена задача вирішується тим, що в способі виробництва біомаси гідробіонтів попередньо розраховують величини максимальної щільності Вmах і мінімальної щільності Bmjn культури (робочий діапазон). У процесі культивування вимірюють щільність культури, продуктивність або питому швидкість зростання й розраховують зміну щільності культури по формулі: 1 t 0 exp ommok t t н Потім порівнюють отриману величину з робочим діапазоном (Вmах Вmіn) і підтримують поточну щільність культури в заданому діапазоні, змінюючи питому швидкість протоки. Спосіб пояснюється кресленнями. На Фіг. 1 Накопичувальна крива зростання з робочим діапазоном для культивування спіруліни; Фіг. 2 - Проточне культивування, де питома швидкість зростання меньше питомої швидкості протоки; Фіг. 3 Проточне культивування, де питома швидкість зростання більше питомої швидкості протоки. Фіг. 4 - Накопичувальна крива зростання з вказівкою робочого діапазона для культивування коловерток. При культивуванні гідробіонтів у промислових масштабах важливою є підтримка щільності культури в робочому діапазоні. На прикладі мікроводоростей доведено, що найбільш оптимальною є робоча щільність, що максимально близька до робочої щільності в точці переходу у фазу вповільнення зростання [Gevorgiz R.G., Borovkov А.В., Shiryaev A.V. Optimization model of the regime for cultivation of microalgae in chemostat of Dunaliella salina Teod. // International Journal on algae. - 2007. - Vol. 9, iss. 3. -P. 294-302)]. Це пов'язане з тим, що, з одного боку, продуктивність максимальна, а з іншого - витрата поживних середовищ мінімальна, що позначається на собівартості продукції. Необхідно безупинно контролювати співвідношення біомаси, що відбирається (врожай) і біомаси, яка залишається в культиваторі. Є два крайні випадки: 1) біомаса відбирається більше, чим приростає в культиваторі, тоді щільність у культиваторі буде зменшуватися до повного вимивання разом із протокою (див. Фіг. 2). Продуктивність буде зменшуватися, оскільки щільність вийде за межі робочого діапазону «униз»; 2) біомаса відбирається менше, чим приростає в культиваторі, тоді щільність у культиваторі буде збільшуватися, при цьому продуктивність буде падати, оскільки щільність 63692 4 вийде за межі робочого діапазону «нагору» (див. Фіг. 3). Введемо в розгляд наступні величини, справедливі для безперервного процесу культивування: W - об'єм культиватора, л; В - поточна концентрація клітин (біомаса) у культиваторі, г/л; m0 - маса водоростей у культиваторі, г. Нескінченно мала маса гідробіонтів dm буде дорівнювати добутку величини нескінченно малої щільності помноженої на об'єм, тобто . . W dmо = W dВ, [г] = [л гл]. (1) Далі введемо: Mвідтік - маса гідробіонтів, що виноситься протокою з культиватора, г; V відтік - об'єм середовища, що виноситься протокою з культиватора, л; К - коефіцієнт непропорційності; аналогічно (1) можна записати з урахуванням коефіцієнта непропорційності . . K.dm відтік = B dV відтік, [г] = [гл л]; (2) Масу гідробіонтів, яка приросла за час dt, г, позначимо через dmnpupіcm, . . . . . dmnpupіcm = B Wdt,[г] = [1/діб л г/л діб], або 1 dmприріст WB dt , (3) де - питома швидкість зростання гідробіон-1 тів, 1/діб . Звідси можна записати формулу для продуктивності по біомасі Р: 1 dmприріст W dt . (4) З (4) випливає формула для продуктивності системи культивування П: П dm приріст dt г г W, л л діб діб . (5) З іншого боку, розділивши співвідношення (2) на dt, одержимо вираз для продуктивності: dmвідтік 1 dmвідтік 1 відтік , dt dt т.ч. для непропорційно проточної культури справедлива рівність . відтік . . B =K P W. (6) Слід зазначити, що якщо К=1, то з (6) виходить широко використовувана формула для стаціонарного динамічного процесу пропорційно-проточної культури відтік B =Р W. або В =Р, де ( - питома швидкість протоки середовища через культиватор. Приведемо основні співвідношення для квазібезперервного непропорційно проточного культивування й позначимо: dVприток - об'єм середовища, що додається в культиватор, л; . . 5 63692 dVприріст - об'єм, займаний прирослою за термін dt масою гідробіонтів, л. Для нескінченно малих об'ємів і мас справедлива рівність dVприток = dVвідтік + dVприріст, і dmприріст = dm0 + dmвідтік. Закон збереження об'єму й маси: dVприток dVвідтік dVприріст dmприріст dm0 dmвідтік (7) звідси d приток dVприріст dt dt dVвідтік W dt W dB B dVвідтік , (8) де приток- питома швидкість припливу поживного середовища. Перетворення виразу (8) дає балансове рівняння для динаміки щільності квазібезперервної непропорційної проточної культури в диференціальній формі: dB 1 відтік dt . (9) Легко помітити, що вираз (9) при К=1 перетворюється в балансове рівняння для динаміки щільності безперервної пропорційно проточної культури (Тренкеншу Р.П. Простейшие модели. 2. Квазинепрерывная культура // Экология моря. 2005. - Вып. 67. - С. 98-110) Задаючи початкові умови й вирішуючи рівняння (9), можна одержати закон зміни щільності культури для різних фаз зростання. Наприклад, для експонентної фази = const, отже, рішення (9) можна записати в наступному вигляді: 1 ( t ) B0 exp відтік t t н . (10) Питома швидкість зростання μ, що входить у формулу (10), є функцією багатьох факторів, у тому числі й неконтрольованих на виробництві: опромінення культиваторів (у хмарні дні), температурний режим навколишнього середовища, інтенсивність перемішування суспензії та ін. Якщо К=1, то залежність (10) перетворюється в загальноприйнятий вираз експонентного зростання (Тренкеншу Р.П. Простейшие модели. 2. Квазинепрерывная культура // Экология моря. 2005. - Вып. 67. - С. 98 - ПО). Якщо також питома швидкість протоки дорівнює нулю, тобто відтік =0, то вираз (10) перетворюється в загальноприйнятий вираз експонентного зростання для накопичувальної культури: ( t ) B0 exp( t t н ) . (11) Аналогічні вираз можна записати для будьякої фази зростання. Таким чином, задаючи умови культивування з використанням математичних розрахунків, можна оптимізувати умови культивування й, здійснюючи контроль над процесом вирощування, досягти підвищення врожаю біомаси. 6 Приклади реалізації способу. Приклад 1. Культивування Spirulina platensis в умовах виробництва фірми Агро-Вікторія (м. Сочі, Росія). Культиватором служив прямокутний басейн 2x2,5 м. Об'єм суспензії в культиваторі становив 500 л при висоті шару 10 см. Цей об'єм підтримували протягом усього експерименту. Для культивування спіруліни використовували поживне середовище Заррука. Розраховували необхідний об'єм інокуляту й поживного середовища, які складали робочий об'єм культиватора. Співвідношення об'ємів повинне бути таким, щоб щільність культури після засівання становила 0,2 г (сухої біомаси) на літр суспензії. Для розрахунку заданої початкової щільності достатньо знайти коефіцієнт розведення , розділивши величину щільності інокуляту на робочу щільність, потім отриманий коефіцієнт і робочий об'єм підставити у формулу 1 w W 1 , W-w , (12) де W - робочий об'єм у культиваторі, - коефіцієнт розведення. В експерименті щільність інокуляту становила 0,8 г/л, а робочий об'єм - 500л, тобто w=500(11/(0,8/0,2))=375 л, отже, щоб досягти робочої щільності в 500 л необхідно 500-375=225 л інокуляту й 375 л поживного середовища. Протягом 10 днів щільність культури збільшувалася й досягла щільності 1,2 г/л. На цьому етапі культивування збір урожаю не проводили, тому його можна розглядати як періодичне (накопичувальне) культивування. У даних умовах динаміку щільності культури можна описати виразом (11). Було відзначено, що на 7 день експерименту швидкість зростання знижувалася. Робочий діапазон був обраний у наступних межах: Bmin=0,9 та Вmах =1,1 г/л (див. Фіг. 1) Безперервний (проточний) процес В міру підйому біомаси на поверхню її збирали. Щодня в заданий час робили контроль параметрів культури. Вимірювали температуру, рН, оптичні характеристики й щільність культури.По оптичним характеристикам і результатам мікроскопії визначали альгологічну чистоту культури і якість біомаси. По вимірюванням щільності розраховували об'єм зливу суспензії й доливу поживного середовища. У нашому експерименті питома -1 швидкість протоки становила 0.1 діб. . Для розрахунку об'єму зливу /доливу (w) використовували вираз (12). Якщо величину біомаси, зібраної на двох етапах, розділити на величину об'єму зливу суспензії, то одержана величина буде відмінною від щільності в басейні, тобто питома швидкість протоки по біомасі виявляється більше, ніж швидкість протоки поживного середовища, тобто виконується умова непропорційності двох потоків. Щоб розрахувати величину протоки середовища й коефіцієнт непропорційності, необхідно у формулу (10) підставити значення питомої швидкості зростання , а величину відтік /К підібрати 7 63692 таким чином, щоб величина B(t) була близька до робочої щільності. Розрахуємо величину питомої швидкості зростання з 6 по 7 добу: 1 0 1,0824 0,9059 0,177 діб 1 0 1 0 / 2 0,9059 (1,0824 0,9059 ) / 2 . (12а) Дотримуючи вимоги про рівність одиниці робочої щільності, з формули (10) можна легко знайти величину відтік /К: 1 1 1,024 ехр (0,177 відтік )(8 7) 1 1 ln 0,177 відтік 1,024 . або Звідки відтік /К 0,2, тобто при заданому коефіцієнті К (наприклад, з технологічних міркувань) відтік одержимо величину . Наприклад, при відтік К=0,67 =0,134. Підраховували врожай, отриманий при використанні різних методів культивування. Періодичне (накопичувальне) культивування. Оскільки вихідна щільність культури становила 0,2 г/л, наприкінці періодичного культивування – 1,2 г/л, у кожному літрі приросло 1 г біомаси за 10 днів. Отже, урожай склав 1 • 500=500 г. Безперервне пропорційно проточне культивування. Щодобовий урожай розраховували по формулі: Y = В • w, де w - об'єм зливу суспензії. (13) Оскільки робоча щільність культури була постійною й становила 1 г/л (В=1), при питомій швид-1 кості протоки 0,1 діб урожай склав: 500•0,1•1 = 50 г Множимо цю величину на час культивування 10 днів, одержуємо 500 г. Якщо до врожаю віднести й біомасу, яка залишається в культиваторі, то одержимо 500 + 400 = 900 г. Квазібезперервне непропорційно проточне культивування Тут формула (13) не підходить, оскільки не враховує зібраний урожай з поверхні суспензії. Для розрахунку щодобового врожаю використовується наступний вираз: Ydt = W•Bпврх + Y = Bпвpx•B•w, (14) де Впврх - щільність біомаси, зібраної з поверхні суспензії. У цьому випадку Впврх=0,05 г/л. Отже, щодобовий урожай становив 500•0,05+50=75 г. Множачи цю величину на час культивування, маємо: 75•10= 750 г. У цьому випадку коефіцієнт непропорційності рівний (див. (2)): 75/50/1=1,5, тобто К=1/1,5=0,67 Таким чином, застосування пропонованого способу забезпечувало оптимальні умови зростання для мікроводоростей. Продуктивність системи культивування підвищувалася в 1,25 разу в порівнянні із пропорційно-проточним методом ку 8 льтивування без контролю над умовами вирощування. Приклад 2. Вирощування коловерток. В експерименті використовували солоноватоводну коловертку Brachionus plicatilis. Культиватор об'ємом 300 л являв собою циліндричну ємність з дном конічної форми зі зливальним отвором у нижній частині. Другий зливальний отвір розташовувався у верхній частині культиватора. При додаванні нової порції поживного середовища частина суспензії самопливом стікала через отвір у ємність для врожаю. Таким чином, автоматично підтримували робочий об'єм на постійному рівні. Підтримку оптимального кисневого режиму й рівномірного розподілу корму здійснювали за допомогою подачі повітря в культиватор. Температуру підтримували на рівні 28-30 °С. Поверхню суспензії висвітлювали лампою ДРЛ-750 до збору врожаю. Періодичне культивування. У культиватор поміщали маточну культуру об'ємом не менш 3-5 л щільністю 20-50 екз./мл. У міру зростання коловерток в екстенсивному режимі вирощування об'єм суспензії доводили до 300 л. Потім, протягом 8-10 днів стежили за динамікою щільності аміктичних самок, яка описується виразом (11). При годуванні коловерток корм вносили з розрахунку 0,5 г на 1 млн. коловерток. Після досягнення максимальної щільності 300 -350 екз./мл і вповільненні швидкості зростання переходили до безперервного вирощування (Фіг. 4). Безперервний процес. Щодня в заданий час здійснювали контроль параметрів культури. Вимірювали температуру, рН, щільність популяції. По вимірам щільності аміктичних самок розраховували об'єм зливу суспензії й доливу поживного середовища. В експерименті робоча щільність була визначена в діапазоні від 240 до 260 екз./мл, оскільки в цьому діапазоні продуктивність максимальна. Щоб розрахувати величину протоки середовища й коефіцієнт непропорційності, необхідно у формулу (10) підставити значення питомої швидкості зростання відтік , а величину /К підібрати таким чином, щоб величина B(t) була близькою до робочої щільності. Величину питомої швидкості зростання розраховуємо аналогічно (12а), але за час з 4 по 5 добу (В0=180): 1 0 275 180 0,42 діб 1. 0 1 0 / 2 180 (275 180 ) / 2 Дотримуючи вимоги про рівність робочої щільності 250, з формули (10) можна легко знайти ве личину відтік /К: 1 250 275 ехр (0,42 відтік )( 6 5) 300 1 ln 0,42 відтік 245 . Звідки відтік =0,35. відтік /К 0,5, або (14б) тобто при К=0,7 9 Установивши розрахункове значення питомої -1 швидкості протоки 0,35 діб при робочій щільності культури 250 діб/мл, одержували щоденний урожай 87,5 екз./мл, максимально можливий урожай для умов даного експерименту. Якщо на 5-у добу не включити протоку (збір урожаю не проводити) і допустити вихід величини щільності за межі робочого діапазону, то втрати врожаю вже на шостий день складали 40 екз./мл, а на сьомий - 75 екз./мл; Якщо в момент включення протоки (на п'ятий відтік день) установити величину
ДивитисяДодаткова інформація
Назва патенту англійськоюMethod for the production of biomass of aquatic organisms
Автори англійськоюHevorhis Ruslan Heorhiiovych
Назва патенту російськоюСпособ производства биомассы гидробионтов
Автори російськоюГеворгиз Руслан Георгиевич
МПК / Мітки
МПК: A01K 61/00, C12N 1/12
Мітки: виробництва, біомаси, гідробіонтів, спосіб
Код посилання
<a href="https://ua.patents.su/6-63692-sposib-virobnictva-biomasi-gidrobiontiv.html" target="_blank" rel="follow" title="База патентів України">Спосіб виробництва біомаси гідробіонтів</a>
Спосіб виробництва біомаси бета-каротину

Номер патенту: 83361
Опубліковано: 10.07.2008
Автори: Тюренков Володимир Олександрович, Тюренков Олексій Олександрович, Туриянський Юрій Давидович, Кунщикова Інна Сергієвна, Кунщикова Євгенія Олександрівна
МПК: C12P 23/00, C12N 1/14, C12R 1/645
Мітки: виробництва, бета-каротину, спосіб, біомаси
Формула / Реферат:
1. Спосіб виробництва біомаси бета-каротину шляхом глибинного культивування штамів Вlakeslea trispora на живильному середовищі, основою якого є джерела органічного вуглецю й азоту, з додаванням біостимулятора каротиноутворення й антиоксиданту, який відрізняється тим, що як продуцент використовують штами Вlakeslea trispora ТКСТ (+) і ТКСТ (-), джерела органічного вуглецю й...
Спосіб виробництва бактерійної біомаси в реакційному об’ємі

Номер патенту: 95152
Опубліковано: 11.07.2011
Автори: Ткаченко Віктор Іванович, Токарев Віктор Сергійович, Шульга Олександр Миколайович, Юдбаровський Давид Моносович, Олександров Олександр Миколайович, Піх Зорян Григорьєвич
Мітки: реакційному, виробництва, біомаси, бактерійної, спосіб, об'ємі
Формула / Реферат:
1. Спосіб виробництва бактерійної біомаси в реакційному об'ємі, що включає періодичне культивування і підживлення її збалансованим живильним середовищем, який відрізняється тим, що вирощують біомасу природних штамів сіркобактерій в збагаченому сірководнем шарі морів і водоймищ шляхом занурення в цей шар біореактора з інокулятом живих бактерій, при цьому завантажують біореактор субстратом, в якому використовується як джерело енергії для...
Спосіб виготовлення продукту з біомаси та установка з виробництва палива з біомаси

Номер патенту: 95795
Опубліковано: 12.09.2011
Автори: Уорнік Томас А., Лескайн Сьюзан
МПК: C08L 1/02, C08L 97/00, C12R 1/145, C12N 1/22, C12P 7/06
Мітки: установка, спосіб, палива, продукту, виготовлення, виробництва, біомаси
Формула / Реферат:
1. Спосіб виготовлення продукту з біомаси, де спосіб включає: надання біомаси, що містить геміцелюлозний або лінгоцелюлозний матеріал; об'єднання біомаси, що містить геміцелюлозний або лінгоцелюлозний матеріал, з клітинами Clostridium, що здатні гідролізувати і ферментувати біомасу, що містить геміцелюлозний або лінгоцелюлозний матеріал, в середовищі; і гідроліз і ферментацію біомаси, що містить...
Спосіб виробництва біомаси, що містить лікопін

Номер патенту: 82355
Опубліковано: 10.04.2008
Автори: Тюренков Володимир Олександро, Кунщикова Інна Сергієвна, Туриянський Юрій Давидович, Тюренков Олексій Олександрови, Кунщикова Євгенія Олександрівна
МПК: C12N 1/14, C12R 1/645, C07C 11/00
Мітки: лікопін, виробництва, біомаси, спосіб, містить
Формула / Реферат:
1. Спосіб виробництва біомаси, що містить лікопін, шляхом ферментації штамів міцеліального гриба Blakeslea trispora ЛК1 (+) і ЛК1 (-) на живильному середовищі, яке містить джерела вуглецю і азоту, при постійному перемішуванні і безперервній подачі стерильного повітря у певному співвідношенні до об'єму середовища, який відрізняється тим, що як джерела вуглецю і азоту використовують живильне середовище, вибране з групи: патока зелена та...
Спосіб виробництва біомаси, що містить фітоїн

Номер патенту: 86556
Опубліковано: 27.04.2009
Автори: Тюрєнков Алєксєй Алєксандровіч, Туриянський Юрій Давидович, Сорока Ярослав Михайлович, Тюрєнков Владімір Алєксандровіч, Бондар Ірина Володимирівна, Дмитренко Галина Миколаївна
МПК: C12N 1/14, C12P 23/00
Мітки: біомаси, спосіб, містить, фітоїн, виробництва
Формула / Реферат:
Спосіб виробництва біомаси, що містить фітоїн, шляхом інкубації штамів мікроорганізмів, що продукують каротиноїди, до певного рівня накопичення фітоїну, який відрізняється тим, що як продуцент фітоїна використовують штам гриба Blakeslea trispora Pht1+ і Pht1-, який депоновано в Депозитарії Інституту мікробіології і вірусології НАН України за реєстраційним номером Blakeslea trispora ІМВ F-100053, у співвідношенні 1:1-1:12, а інкубацію ведуть у...
Попередній патент: Амортизатор автоматичний регульований з диференціальною характеристикою
Наступний патент: Спосіб отримання гіпохлориту натрію
Випадковий патент: Радіолокатор беззупинного випромінювання