Спосіб оцінки якості сперми коропа
Номер патенту: 83803
Опубліковано: 25.09.2013
Автори: Пуговкін Антон Юрійович, Буцький Кирило Ігорович, Копєйка Євген Федорович




Формула / Реферат
Спосіб оцінки якості сперми коропа, який включає інкубацію клітин у гіпотонічному розчині, визначення динаміки пошкодження сперматозоїдів і їх осмотичної резистентності, який відрізняється тим, що при інкубації сперматозоїдів у гіпотонічному розчині за допомогою фотоелектроколориметра реєструють залежність світлопропускання суспензії клітин від часу, звідки отримують динаміку пошкодження сперматозоїдів, при цьому час пошкодження 50 % клітин визначає осмотичну резистентність сперматозоїдів.
Текст
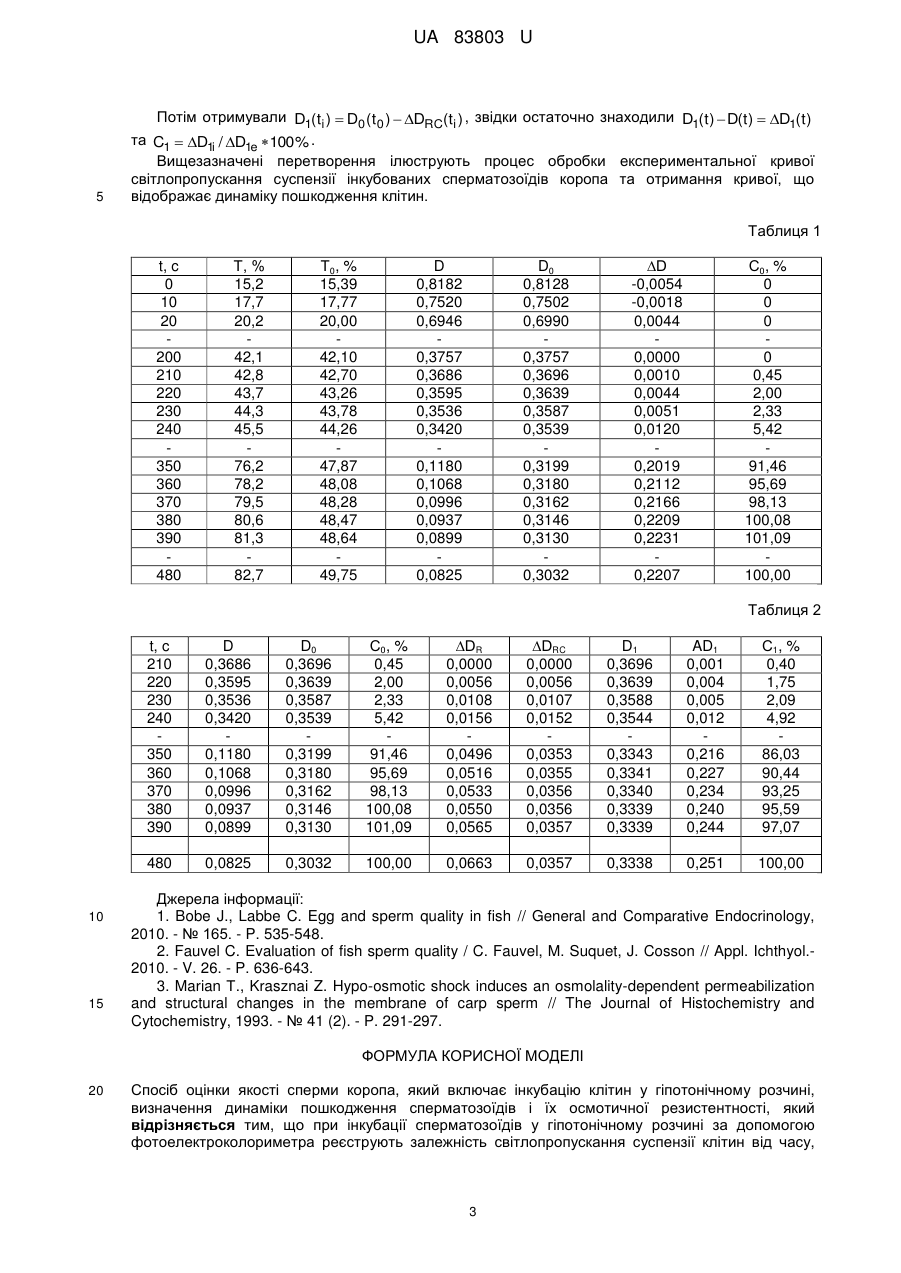
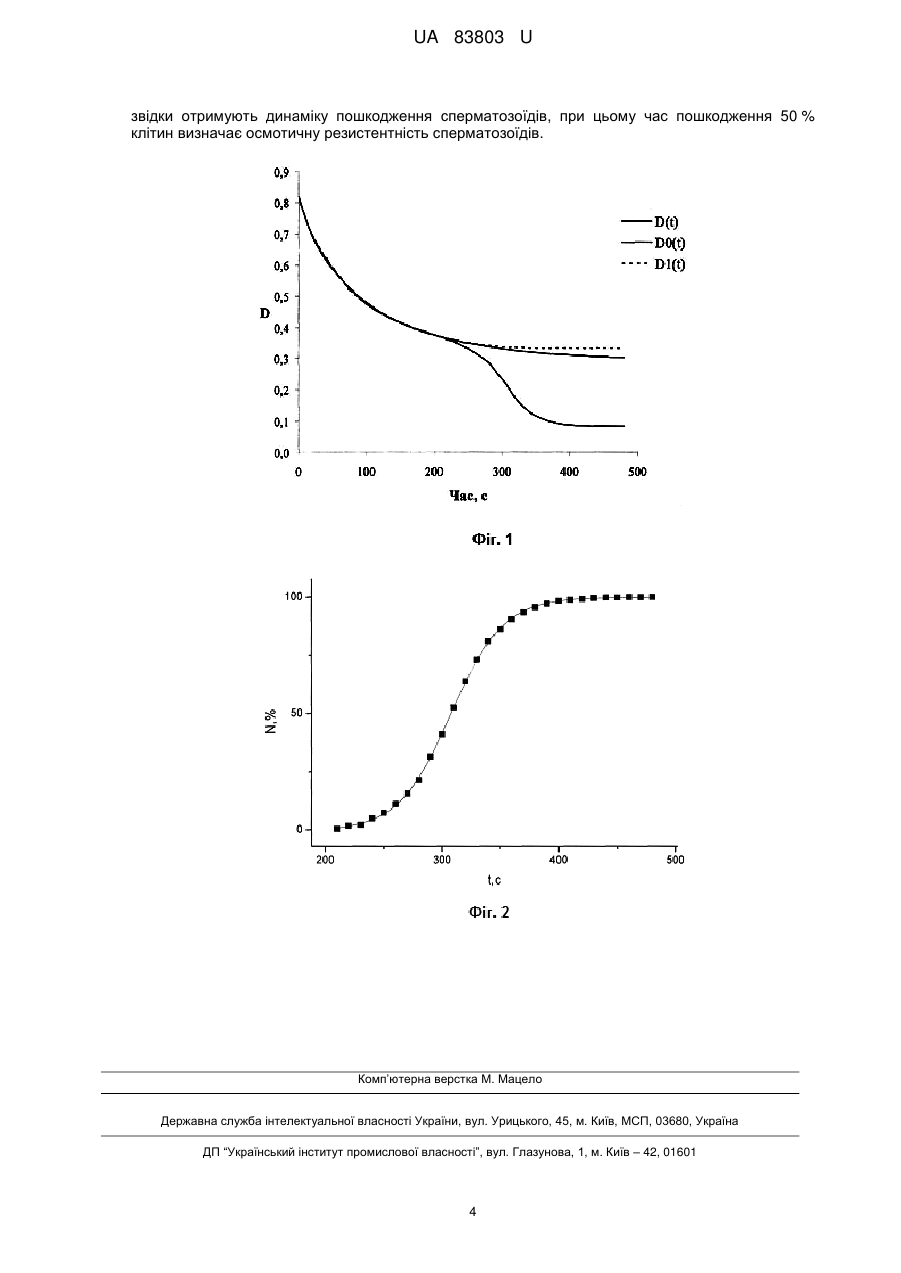
Реферат: Спосіб оцінки якості сперми коропа включає інкубацію клітин у гіпотонічному розчині, визначення динаміки пошкодження сперматозоїдів і їх осмотичної резистентності. При цьому при інкубації сперматозоїдів у гіпотонічному розчині за допомогою фотоелектроколориметра реєструють залежність світлопропускання суспензії клітин від часу, звідки отримують динаміку пошкодження сперматозоїдів, час пошкодження 50 % клітин визначає осмотичну резистентність сперматозоїдів. UA 83803 U (54) СПОСІБ ОЦІНКИ ЯКОСТІ СПЕРМИ КОРОПА UA 83803 U UA 83803 U 5 10 15 20 25 30 35 40 45 50 55 60 Корисна модель належить до галузі кріобіології і може бути використана при розробці середовищ кріоконсервування сперматозоїдів коропа для збереження генофонду промислових риб, а також у рибному господарстві. Якість сперми можна визначається як здатність сперматозоїдів запліднити яйцеклітини та привести до розвитку нормальних ембріонів. Визначення якості сперми включає оцінку параметрів руху сперматозоїдів, морфологічний та біохімічний аналіз клітин. Відомий спосіб оцінки якості сперми риб за відносною кількістю сперматозоїдів, що рухаються. Для цього певну кількість сперми розмішують в активуючому розчині і за допомогою мікроскопу візуально визначають відношення кількості сперматозоїдів, що рухаються, до загальної кількості сперматозоїдів у полі зору [1]. Відомий спосіб оцінки якості сперми риб, в якому визначають час рухливості сперматозоїдів в активуючому середовищі [2]. Значним недоліком зазначених способів є їх суб'єктивність, що обумовлює значну похибку визначення кількості рухливих сперматозоїдів. Також кількість рухливих сперматозоїдів залежить від того, в якому шарі в активуючому розчині вони знаходяться, тобто на різній глибині можуть бути різні показники рухливості, що так само призводить до похибки в оцінці якості сперми. Відомий спосіб оцінки якості сперми риб за показником швидкості руху сперматозоїдів в активуючому середовищі. Він визначається за допомогою комп'ютерної обробки відеозаписів руху клітин [1, 2]. Недоліком цього способу є також неточність оцінки через залежність рухливості клітин від глибини в активаторі, на якій вони знаходяться. Як найближчий аналог вибрано спосіб оцінки якості сперми коропа, який базується на визначенні резистентності мембран до осмотичного шоку [3]. Осмотичну резистентність клітини визначає час, за який пошкоджується певна частина клітин, або навпаки, деякий відсоток пошкоджених клітин за визначений час. 8 Згідно зі способом, сперму розбавляють фізіологічним розчином до концентрації 10 сперм./мл. Далі у відношенні 1:100 інкубують у гіпотонічному розчині хлориду натрію, що містить 50 мкг/мл барвника propidium iodide (PI), і за допомогою спектрофлуориметра визначають залежність інтенсивності флуоресценції РІ, що зв'язується з ДНК, від часу, тобто динаміку пошкодження сперматозоїдів, яка свідчить про їх осмотичну резистентність. На її основі роблять висновок про якість сперми. Недоліком цього способу є необхідність використання дорогих лабораторного приладдя і реактивів, що обмежує його використання у сільському господарстві. В основу корисної моделі поставлено задачу створити більш доступний для використання на практиці спосіб оцінки якості сперми коропа. Ця задача вирішується тим, що в відомому способі оцінки якості сперми, який включає інкубацію клітин у гіпотонічному розчині, визначення динаміки пошкодження сперматозоїдів і їх осмотичної резистентності, згідно з корисною моделлю, при інкубації сперматозоїдів у гіпотонічному розчині за допомогою фотоелектроколориметра (ФЕК) реєструють залежність світлопропускання суспензії клітин від часу, звідки отримують динаміку пошкодження сперматозоїдів, при цьому час пошкодження 50 % клітин визначає осмотичну резистентність сперматозоїдів коропа. Використання більш доступного приладу (ФЕК) без застосування реактиву розширює можливість застосування заявленого способу в сільському господарстві. Спосіб здійснюють таким чином. Сперму поміщують у кювету з гіпотонічним розчином і постійно перемішують суспензію. За допомогою ФЕК реєструють залежність світлопропускання з часом та фіксують на папері з використанням самописця. Потім проводять її апроксимацію при значеннях часу, менших значення часу характерного перегину кривої, та екстраполюють визначену функцію на значення часу, більші за значення характерного перегину. Далі вказану залежність коригують у відповідності до реєстрованої залежності світлопропускання, обидві залежності виражають в термінах оптичної щільності та знаходять різницю оптичних щільностей та нормують її на 1. Таким чином, отримують криву, що характеризує зменшення концентрації непошкоджених сперматозоїдів у суспензії, тобто динаміку пошкодження сперматозоїдів, обумовлену осмотичним шоком. Приклад Сперму коропа (Cyprinus carpio) отримували у чистий сухий посуд. Під час проведення досліджень зберігали при температурі 15-18 °C у закритому посуді. Вимірювання проводили при температурі 18-20 °C. 1 UA 83803 U 5 10 15 20 25 30 35 40 В процесі вимірювання кювету (внутрішній розмір 10×10×44 мм) за допомогою мікродозатора наповнювали дистилятом (3,6 мл) та занурювали мішалку, виставляли значення світлопропускання на відмітці 100 %, потім вмикали самописець. Сперму за допомогою мікродозатора (54 мкл) додавали у кювету, швидко розмішували протягом 1-2 с та переміщували у кюветне відділення ФЕК. Вимірювання проводили при довжині хвилі світла 610 нм. Як тільки зміна світлопропускання припинялась, вимірювання закінчували та ретельно промивали кювету. Отримана крива є суперпозицією двох процесів - процесу набухання клітини, що відображується в зменшенні оптичної щільності, і процесу зменшення кількості непошкоджених клітин, що також призводить до зменшення оптичної щільності суспензії. У першому наближенні криву зміни кількості клітин з часом можна отримати, знаючи різницю функцій оптичної щільності D0 (t ) и D( t ) від часу. При цьому, D( t ) - динаміка оптичної щільності, що отримана експериментально та враховує вклад як набухання, так і зміни концентрації розсіювачів (фіг. 1), a D0 (t ) - динаміка оптичної щільності суспензії, що отримана екстраполяцією D( t ) при t t 0 , де t 0 - час, при якому починають пошкоджуватися клітини, тобто динаміка, що враховує тільки набухання клітин. Різниця D0 (t ) D(t ) D0 (t ) C0 (t )l , тобто пропорційна концентрації частинок у середовищі, коефіцієнту екстинкції і ширині кювети. Вважаючи, що ширина кювети та коефіцієнт екстинкції незмінні, D(t) відображає зміну кількості непошкоджених клітин. Обробку результатів проводили наступним чином. Вводили значення світлопропускання суспензії T у таблицю на ПК через кожні 10 с інкубації (таблиця 1). За допомогою програми OriginPro проводили апроксимацію отриманих значень при t 180 с функцією виду T0 A Вехр(kt ) , у якій в даному випадку коефіцієнти А і В мають зміст граничного значення світлопропускання та діапазону його зміни, а коефіцієнт k - швидкості зміни світлопропускання. Згідно з формулою D lg(1/ T) розраховували відповідно D і D0 , звідки знаходили D D0 D і C0 Di / De 100 % ( Di - значення різниці оптичної щільності в кожний момент часу ti , De , - значення різниці оптичної щільності в кінцевий момент часу t e , - в даному прикладі при t 480 інкубації). Отримані в першому наближенні значення C0 (t) містять відносно незначну помилку (декілька відсотків), пов'язану з тим, що отримана екстраполяцією функція T0 (D0 ) змінюється при постійних значеннях T( t ) (D(t)) , тобто згідно з моделлю T0 (D0 ) визначає набухання 100 % клітин навіть тоді, коли відбувається їхнє пошкодження. Для усунення вищезазначеного, у другому наближенні, враховується те, що величина зміни T0 (D0 ) сперматозоїдів залежить від концентрації клітин у кюветі. Так, при t t 0 , зі зменшенням кількості непошкоджених клітин, величина зміни світлопропускання або оптичної щільності суспензії, отримана екстраполяцією D( t ) , має зменшуватись відповідно. В результаті перетворювань маємо різницю D1(t ) D(t ) D1(t ) ~ C1(t ) . Нормована крива, яка відображає динаміку пошкодження клітин, наведена на фіг. 2. Експериментальні значення апроксимували функцією виду A1 A 2 y A2 . 1 e( x x 0 ) / dx ( A 1,2 - початкове і кінцеве значення D1(t ) , x 0 - абсциса центра, dx - константа часу). У таблиці 2 наведені дані, що демонструють перетворення функції D0 (t ) до функції D1(t ) . Спочатку знаходили 45 50 DR (t ) D0 (t 0 ) D0 (t ) при t t 0 . Ця залежність характеризує набухання 100 % непошкоджених клітин. Далі знаходили DRC(t ) як таблично задану функцію за наступним співвідношенням: D( ti ) D(t e ) DRC( ti ) DRC( ti1) [DR ( ti ) DR ( ti1)] D( t 0 ) D(t e ) за умови DRC(t 0 ) 0 . Залежність DRC(t ) вже характеризує набухання тільки непошкоджених клітин у кожний окремий момент часу. 2 UA 83803 U 5 Потім отримували D1(ti ) D0 (t 0 ) DRC(ti ) , звідки остаточно знаходили D1(t) D(t) D1(t ) та C1 D1i / D1e 100 % . Вищезазначені перетворення ілюструють процес обробки експериментальної кривої світлопропускання суспензії інкубованих сперматозоїдів коропа та отримання кривої, що відображає динаміку пошкодження клітин. Таблиця 1 t, c 0 10 20 200 210 220 230 240 350 360 370 380 390 480 Т, % 15,2 17,7 20,2 42,1 42,8 43,7 44,3 45,5 76,2 78,2 79,5 80,6 81,3 82,7 Т0, % 15,39 17,77 20,00 42,10 42,70 43,26 43,78 44,26 47,87 48,08 48,28 48,47 48,64 49,75 D 0,8182 0,7520 0,6946 0,3757 0,3686 0,3595 0,3536 0,3420 0,1180 0,1068 0,0996 0,0937 0,0899 0,0825 D0 0,8128 0,7502 0,6990 0,3757 0,3696 0,3639 0,3587 0,3539 0,3199 0,3180 0,3162 0,3146 0,3130 0,3032 ∆D -0,0054 -0,0018 0,0044 0,0000 0,0010 0,0044 0,0051 0,0120 0,2019 0,2112 0,2166 0,2209 0,2231 0,2207 С0, % 0 0 0 0 0,45 2,00 2,33 5,42 91,46 95,69 98,13 100,08 101,09 100,00 Таблиця 2 t, c 210 220 230 240 350 360 370 380 390 15 D0 0,3696 0,3639 0,3587 0,3539 0,3199 0,3180 0,3162 0,3146 0,3130 С0, % 0,45 2,00 2,33 5,42 91,46 95,69 98,13 100,08 101,09 ∆DR 0,0000 0,0056 0,0108 0,0156 0,0496 0,0516 0,0533 0,0550 0,0565 ∆DRC 0,0000 0,0056 0,0107 0,0152 0,0353 0,0355 0,0356 0,0356 0,0357 D1 0,3696 0,3639 0,3588 0,3544 0,3343 0,3341 0,3340 0,3339 0,3339 AD1 0,001 0,004 0,005 0,012 0,216 0,227 0,234 0,240 0,244 С1, % 0,40 1,75 2,09 4,92 86,03 90,44 93,25 95,59 97,07 480 10 D 0,3686 0,3595 0,3536 0,3420 0,1180 0,1068 0,0996 0,0937 0,0899 0,0825 0,3032 100,00 0,0663 0,0357 0,3338 0,251 100,00 Джерела інформації: 1. Bobe J., Labbe C. Egg and sperm quality in fish // General and Comparative Endocrinology, 2010. - № 165. - P. 535-548. 2. Fauvel C. Evaluation of fish sperm quality / C. Fauvel, M. Suquet, J. Cosson // Appl. Ichthyol.2010. - V. 26. - P. 636-643. 3. Marian Т., Krasznai Z. Hypo-osmotic shock induces an osmolality-dependent permeabilization and structural changes in the membrane of carp sperm // The Journal of Histochemistry and Cytochemistry, 1993. - № 41 (2). - P. 291-297. ФОРМУЛА КОРИСНОЇ МОДЕЛІ 20 Спосіб оцінки якості сперми коропа, який включає інкубацію клітин у гіпотонічному розчині, визначення динаміки пошкодження сперматозоїдів і їх осмотичної резистентності, який відрізняється тим, що при інкубації сперматозоїдів у гіпотонічному розчині за допомогою фотоелектроколориметра реєструють залежність світлопропускання суспензії клітин від часу, 3 UA 83803 U звідки отримують динаміку пошкодження сперматозоїдів, при цьому час пошкодження 50 % клітин визначає осмотичну резистентність сперматозоїдів. Комп’ютерна верстка М. Мацело Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 4
ДивитисяДодаткова інформація
МПК / Мітки
МПК: G01N 15/14, G01N 33/48
Мітки: оцінки, коропа, спосіб, якості, сперми
Код посилання
<a href="https://ua.patents.su/6-83803-sposib-ocinki-yakosti-spermi-koropa.html" target="_blank" rel="follow" title="База патентів України">Спосіб оцінки якості сперми коропа</a>
Спосіб оцінки якості сперми

Номер патенту: 43524
Опубліковано: 25.08.2009
Автори: Самойлюк В'ячеслав Володимирович, ЛЯШЕНКО ЄВГЕН ВОЛОДИМИРОВИЧ, Буров Валерій Олексійович
МПК: A61D 99/00
Мітки: спосіб, сперми, якості, оцінки
Формула / Реферат:
1. Спосіб оцінки якості сперми, який включає стадії підготовки зразка, нанесення його на термостатоване предметне скельце і мікроскопічний підрахунок сперміїв, який відрізняється тим, що на стадії підготовки сперма розбавляється дистильованою водою, а нанесений на предметне скельце зразок підсушується.2. Спосіб за п. 1, який відрізняється тим, що при підрахунку під мікроскопом визначається кількість добрих, задовільних і фізіологічно...
Спосіб прискореної оцінки якості заморожено-відтаяної сперми бугаїв

Номер патенту: 60740
Опубліковано: 25.06.2011
Автори: Савельєва Марина Сергіївна, Сушко Олексій Борисович
МПК: A61D 19/02, A61D 19/00
Мітки: оцінки, прискореної, сперми, якості, заморожено-відтаяної, спосіб, бугаїв
Формула / Реферат:
Спосіб прискореної оцінки якості заморожено-відтаяної сперми бугаїв, що включає інкубування зразка у біологічному ультратермостаті та періодичне визначення рухливості сперми, який відрізняється тим, що при інкубації використовується температура 45 °С, а інтервали часу між визначенням рухливості складають 10 хвилин.
Спосіб оцінки якості сперми

Номер патенту: 13369
Опубліковано: 28.02.1997
Автори: Зубец Михайло Васильович, Бегма Людмила Олексіївна, Власенко Віталій Валерійович, Пеньков Федір Михайлович, БЕГМА Анатолій Андрійович
МПК: A61D 19/02
Мітки: оцінки, сперми, якості, спосіб
Формула / Реферат:
(57) 1. Способ оценки качества спермы, включающий ее отбор, разбавление, облучение лучом лазера при длине волны 633 нм, измерение рассеянного света и последующую оценку качества спермы, отличающийся тем, что после облучения спермы лучом лазера регистрируют и измеряют флуктуации рассеянного света, определяют корреляционную функцию рассеянного света и по ней определяют скорость движения и число подвижных спермиев, по которым судят о качестве...
Середовище для кріоконсервування сперми коропа

Номер патенту: 49236
Опубліковано: 26.04.2010
Автори: Дрокін Сергій Іванович, Копєйка Євгеній Федорович
МПК: A01K 61/00, A01K 1/02
Мітки: кріоконсервування, середовище, коропа, сперми
Формула / Реферат:
Середовище для кріоконсервування сперми коропа, яке містить хлористий натрій, гідрокарбонат натрію, хлористий калій, глюкозу, трис, 10 % жовтка курячого яйця і 16 % кріопротектора етиленгліколю, який відрізняється тим, що містить хлористий натрій в кількості 0,026 М, гідрокарбонат натрію - 0,030 М, хлористий калій - 0,013 М, глюкозу - 0,005 М, трис - 0,058 М, при цьому сумарна осмотичність розчину без жовтка і кріопротектора становить 201...
Спосіб прогнозування якості сперми бугаїв

Номер патенту: 21530
Опубліковано: 16.12.1997
Автори: Звєрєва Галина Володимирівна, Ісаченко Євгенія Федорівна, Осташко Федір Іванович, Рівіс Йосип Федорович, Свідерко Богдан Дмитрович, Тимчишин Ірина Миколаївна, Палфій Федір Юрієвич, Максим'юк Василь Михайлович, Кокодиняк Ізяслав Петрович
МПК: G01N 33/84
Мітки: бугаїв, спосіб, сперми, якості, прогнозування
Формула / Реферат:
Спосіб прогнозування якості сперми бугаїв за біохімічними показниками, який відрізняється тим, що для оцінки якості свіжоотриманої і розмороженої сперми використовують показник рівня калію в спермі, при цьому сперму з числовим значенням вмісту калію в межах 10-45 мМоль/л приймають за високоякісну, а при значеннях 46 мМоль/л і більше - вибраковують.
Попередній патент: Затискач нирковий
Наступний патент: Термошайба універсальна компенсаційна
Випадковий патент: Пристрій для гранулювання рідкого матеріалу