Активний протеазостійкий мутант антитіла fс
Номер патенту: 113725
Опубліковано: 10.03.2017
Автори: Строл Уілльям, Брезскі Рендалл, Джордан Роберт







Формула / Реферат
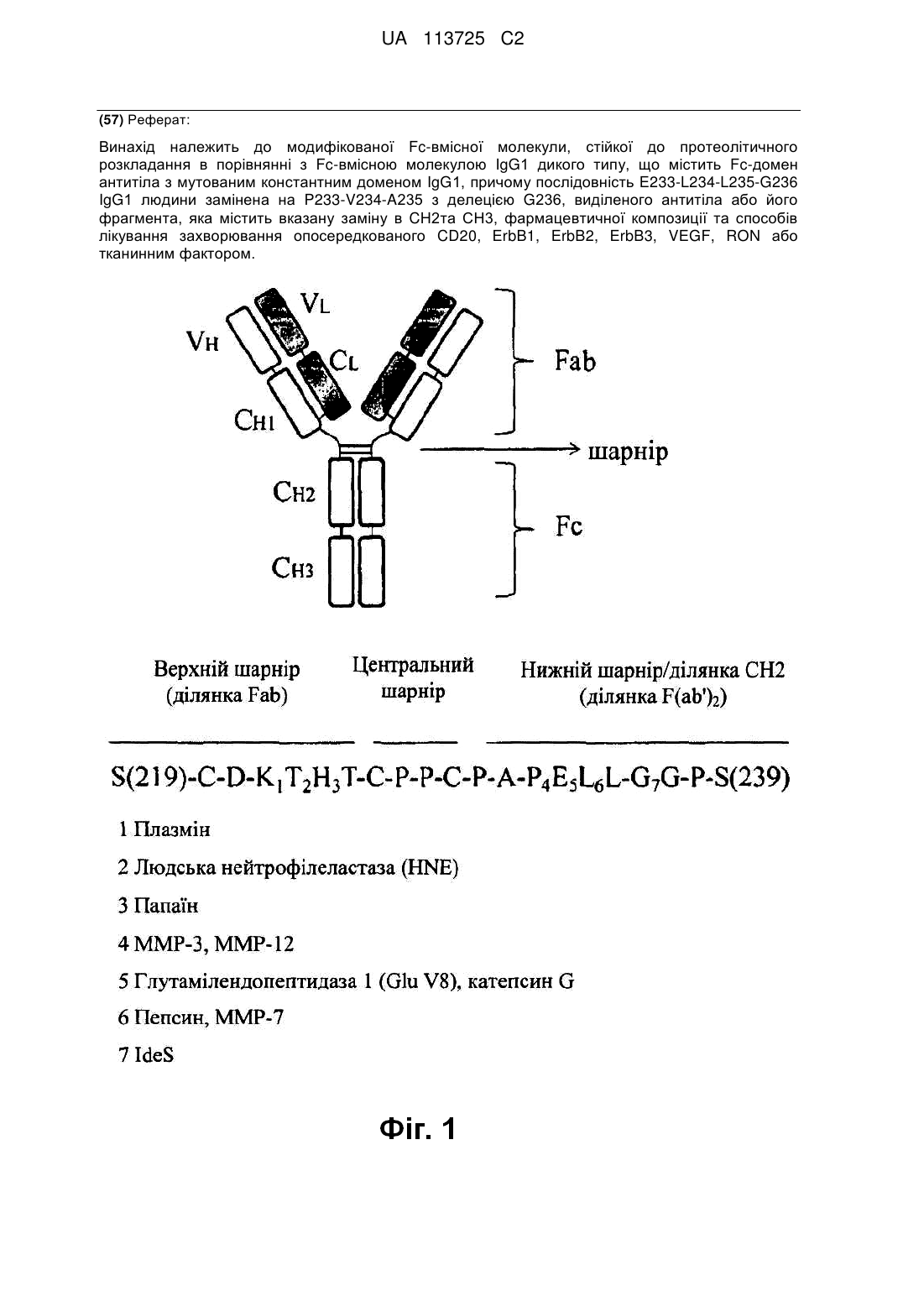
1. Модифікована Fc-вмісна молекула, стійка до протеолітичного розкладання, в порівнянні з Fc-вмісною молекулою IgG1 дикого типу, яка містить Fc-домен антитіла з мутованим константним доменом IgG1, причому послідовність E233-L234-L235-G236 IgG1 людини замінена на P233-V234-A235 з делецією G236, як визначено відповідно до нумерації по EU, і яка додатково, містить одну або більше заміни в послідовності IgG1 людини дикого типу, вибраних із S239D/I332E; K326А/Е333А; Е333А/K334А; H268F/S324T/I332E; F243L/R292P/Y300L; S239D/H268F/S324T/I332E; S267E/H268F/S324T/I332E; K326А/І332Е/Е333А; S239D/K326A/E333A; S267E/I332E і G237X/S239D/I332E, де X являє собою A, D, P, Q або S.
2. Fc-вмісна молекула за п. 1, яка стійка до розкладання протеазою, здатною розкладати молекулу IgG1 між залишками 222-237 (нумерація по EU).
3. Fc-вмісна молекула за п. 2, яка стійка до розкладання ММР-3, ММР-7, ММР-12, катепсином G, пепсином, який розкладає імуноглобулін ферментом Strep. Pyrongenes (IdeS) або глутамілендопептидазою І Staph. aureus (GluV8) в порівнянні з IgG1 дикого типу.
4. Fc-вмісна молекула за п. 3, яка стійка до розкладання ММР-3, ММР-7, IdeS або GluV8 в порівнянні з IgG1 дикого типу.
5. Fc-вмісна молекула за п. 1, яка здатна стимулювати ADCP, виміряний в присутності мононуклеарних клітин крові CD14 pos і/або CD11b pos, що містить послідовність, вибрану з групи SEQ ID NO: 8, 10-15 і 18-20.
6. Fc-вмісна молекула за п. 1, яка здатна стимулювати ADCC, виміряний в присутності мононуклеарних клітин крові.
7. Fc-вмісна молекула за п. 6, що містить послідовність, вибрану з групи SEQ ID NO: 8, 10-12, 15, 18-20.
8. Fc-вмісна молекула за п. 1, яка здатна стимулювати комплементозалежну цитотоксичність (CDC), виміряну по лізису клітин в присутності комплементу.
9. Fc-вмісна молекула за п. 8, що містить послідовність, вибрану з групи SEQ ID NO: 13, 14 і 18-20.
10. Fc-вмісна молекула за п. 1, яка здатна зв'язувати Fcg-рецептор з порівнянною або більшою афінністю, ніж Fc-домен IgG2 дикого типу.
11. Fc-вмісна молекула за п. 1, яка здатна зв'язувати Fcg-рецептор з порівнянною або більшою афінністю, ніж Fc-домен IgG1 дикого типу.
12. Fc-вмісна молекула за п. 6, що містить SEQ ID NO: 8.
13. Fc-вмісна молекула за будь-яким з пп. 1-12, яка являє собою антитіло або Fc-злитий білок.
14. Fc-вмісна молекула за п. 13, де антитіло зв'язується з антигеном на пухлинній клітині, пухлинній матриці або судинній сітці пухлини.
15. Fc-вмісна молекула за п. 14, де антитіло зв'язується з одним із CD20, ErbBl, ЕrbВ2, ЕrbВ3, VEGF, RON і тканинним фактором.
16. Fc-вмісна молекула за п. 1, в якій послідовність Fc-домену щонайменше на 90 % ідентична IgG1 людини дикого типу із залишку 214 до приблизно залишку 340 за системою нумерації EU.
17. Виділене антитіло або його фрагмент, які включають модифіковану Fc-вмісну молекулу, яка містить ділянку Fc людського IgG1 дикого типу з SEQ ID NO: 1, яка містить шарнір, ділянку СН2 та ділянку СН3, де послідовність Е233-L234-L235-G236 в шарнірі замінюють на P233-V234-A235 з делецією G236; і
ділянка СН2 включає щонайменше одну заміну, вибрану з S239D/I332E; K326А/Е333А; H268F/S324T/I332E; F243L/R292P/Y300L; S239D/H268F/S324T/I332E; S267E/H268F/S324T/I332E; K326А/І332Е/Е333А; S239D/K326A/E333A; S267E/I332E і G237X/S239D/I332E, де X являє собою A, D, P, Q або S; де нумерація здійснена відповідно до нумерації EU.
18. Зв'язувальна молекула за п. 17, в якій Fc-домен додатково містить одну або більше заміни із послідовності IgG1 людини дикого типу, вибраних із S239D/I332E; K326А/Е333А; Е333А/K334А; H268F/S324T/I332E; F243L/R292P/Y300L; S239D/H268F/S324T/I332E; S267E/H268F/S324T/I332E; K326А/І332Е/Е333А; S239D/K326A/E333A; S267E/I332E і G237X/S239D/I332E, де X являє собою A, D, P, Q aбo S.
19. Зв'язувальна молекула за п. 17, в якій Fc-домен стійкий до розкладання протеазою, здатною розщеплювати молекулу IgG1 між залишками 222-237 (нумерація по EU).
20. Fc-вмісна молекула за п. 19, яка стійка до розкладання ММР-3, ММР-7, ММР-12, катепсином G, пепсином, IdeS або GluV8 в порівнянні з IgG1 дикого типу.
21. Зв'язувальна молекула за п. 17, в якій зв'язувальний домен вибраний із домену, який містить паратоп антитіла; ферменту; гормону; рецептора; цитокіну; поверхневого антигену імунної клітини і адгезійної молекули.
22. Зв'язувальна молекула за п. 21, яка містить два або більше мішеневих зв'язувальних доменів і має авідність.
23. Зв'язувальна молекула за п. 22, в якій зв'язувальний домен містить паратоп антитіла, який зв'язується з антигеном на пухлинній клітині або судинній сітці пухлини.
24. Зв'язувальна молекула за п. 23, яка зв'язується з одним із CD20, ErbB1, ErbВ2, ErbВ3, VEGF, RON і тканинним фактором.
25. Фармацевтична композиція, що містить Fc-вмісну молекулу за п. 1 або 24.
26. Спосіб лікування захворювання, що характеризується небажаною проліферацією або міграцією клітин, який включає введення пацієнту фармацевтичної композиції за п. 25.
27. Спосіб за п. 26, в якому захворювання являє собою злоякісне захворювання, фіброзне захворювання або захворювання, що характеризується небажаним ангіогенезом.
28. Спосіб за п. 27, в якому Fc включає заміни, вибрані тільки із І332Е або в комбінації з іншими заміщеннями, такими як S239D/I332E, S239D/H268F/I332E, S239D/H268F/S324T/I332E, S267E/H268F/S324T/I332E, G237X/S239D/I332E, де X являє собою А або S; K326A/E333A і F243L/R292P/Y300L.
29. Спосіб за п. 27, в якому Fc включає заміни, вибрані тільки із S239D або в комбінації з іншими заміщеннями, такими як S239D/I332E, S239D/H268F/I332E, S239D/H268F/S324T/I332E, S267E/H268F/S324T/I332E, G237X/S239D/I332E, де X являє собою А або S; K326A/E333A; і F243L/R292P/Y300L.
30. Спосіб лікування захворювання, яке характеризується як інфекція прокаріотичним організмом, що включає введення зв'язувальної молекули за п. 17.
31. Спосіб за п. 29, в якому Fc стійкий до прокаріотичної протеази і виявляє CDC.
32. Спосіб за п. 29, в якому Fc-домен має послідовність людського IgG1 в шарнірній ділянці і ділянці СН2 від приблизно EU залишків 214 до приблизно залишку 330, причому щонайменше залишки 233-237 заміщені PVA/(делеція G236), і додатково містить одну або більше замін в домені СН2, вибраних із K326А/Е333А, S267E/H268F/S324T/I332E, K326А/І332Е/Е333А, S239D/K326A/E333A і S267/I332E.
33. Спосіб за п. 31, в якому зв'язувальна молекула містить послідовність, вибрану із групи SEQ ID NO: 13, 14 і 18-20.
Текст
Реферат: Винахід належить до модифікованої Fc-вмісної молекули, стійкої до протеолітичного розкладання в порівнянні з Fc-вмісною молекулою IgG1 дикого типу, що містить Fc-домен антитіла з мутованим константним доменом IgG1, причому послідовність E233-L234-L235-G236 IgG1 людини замінена на P233-V234-A235 з делецією G236, виділеного антитіла або його фрагмента, яка містить вказану заміну в CH2та СН3, фармацевтичної композиції та способів лікування захворювання опосередкованого CD20, ErbB1, ErbB2, ErbB3, VEGF, RON або тканинним фактором. UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 ПЕРЕДУМОВИ СТВОРЕННЯ ВИНАХОДУ Попередня заявка Дана заявка вимагає пріоритет по заявці США № 61/426,619, поданій 23 грудня 2010 р., і заявці США № 61/540,882, поданій 29 вересня 2011 р., зміст яких повністю включений в цей документ шляхом посилання. Галузь застосування винаходу Даний винахід стосується константних ділянок людських антитіл IgG, зокрема, ділянок Fc, мутованих таким чином, щоб змінити сайти протеолітичного розщеплення, додаючи стійкість відносно ендогенних протеаз і протеаз, які мають патогенне походження, а також додатково змінених таким чином, щоб специфічно зв'язувати рецептори Fcγ і активувати мітогенні відповіді імунних клітин за рахунок опосередкованого рецепторами Fc перехресного зшивання або активації комплементарного каскаду. Нові послідовності можуть бути включені в терапевтичні композиції антитіл, в яких необхідна протеолітична стійкість і функціональні властивості цитотоксичного ефектора. ОПИС ГАЛУЗІ ЗАСТОСУВАННЯ Ізотип людських антитіл IgG складається з підтипів IgG1, IgG2, IgG3 і IgG4, кожен з яких містить два антигензв'язувальних плеча (Fab), з'єднаних із загальним доменом Fc шарнірною ділянкою. IgG1, переважаючий підклас, представлений в терапевтичних моноклональних антитілах, прийнято відносити до стабільних молекул з тривалим періодом напівжиття від 17,6 до 56,2 діб (Salfeld, Nat Biotechnol 25(12):1369-72, 2007 р.). Однак IgG1 схильний протеолізу в шарнірній ділянці під впливом ряду фізіологічно значущих протеаз, пов'язаних з інвазивним раком (наприклад, матриксні металопротеїнази) і патологічними мікроорганізмами. Розщеплення вище дисульфідних зв'язків (центральний шарнір) між тяжкими ланцюгами вивільняє одновалентний Fab, а двостороннє розщеплення нижче дисульфідних зв'язків вивільняє двовалентну структуру - фрагмент F(ab')2. Деякі металопротеїнази і два бактерійних ферменти - глутамілендопептидаза V8 Staphylococcus aureus (GluV8) і розкладаючий імуноглобулін фермент Streptococcus pyogenes (IdeS) - впливають на IgG1 в нижньому шарнірі (нижче дисульфідних зв'язків (фігура 1)) і, зрештою, приводять до утворення фрагментів F(ab') 2 і Fc (Ryan et al., Mol Immunol 45(7):1837-46, 2008 р.). Якщо ефективність терапевтичних моноклональних антитіл (mAb), спрямованих проти поверхневих антигенів клітини, пов'язана із знищенням клітини-мішені, "ефекторні функції" антитіла, які йому додає домен Fc, безсумнівно, грають роль і мають важливе значення для терапевтичного ефекту антитіла загалом. (Bibeau et al., J Clin Oncol 27:1122-1129, 2009 р.; Cartron et al., Blood 99:754-758, 2002 р. і Musolino et al., J Clin Oncol 26:1789-1796, 2008 р.). Передбачається, що домен Fc антитіла, який взаємодіє з гамма-рецепторами Fc (FcγR), експресованими на імунних клітинах, а також взаємодію домену Fc з комплементом визначають вплив декількох моноклональних антитіл (mAb), спрямованих проти поверхневих антигенів клітини. Ці взаємодії можуть привести до усунення клітин-мішеней mAb за рахунок антитілозалежної клітинноопосередкованої цитотоксичності (ADCC), антитілозалежного клітинноопосередкованого фагоцитозу (ADCP) і комплементозалежної цитотоксичності (CDC). У цей час показано, що одиничне протеолітичне розщеплення в одному з поліпептидів тяжкого ланцюга IgG1 не порушує зв'язку ланцюгів і, отже, зберігає тривалість антигензв'язувальної здатності в організмі, але приводить до втрати здатності IgG1 зв'язувати FcγR і визначати опосередковані Fc ефекторні функції (Brezski et al., Proc Natl Acad Sci USA 106:17864-17869, 2009 р.). Як одиничні, так і множинні розщеплення терапевтичних моноклональних антитіл можуть приводити до утворення компонентів, які зв'язуються з мішенню, але частково або повністю втратили ефективність. Таким чином, інженерія протеазостійких платформ Fc з підвищеною ефекторною функцією крім інших застосувань може забезпечити значну перевагу, яка полягає в підвищенні ефективності протиракових і протиінфекційних терапевтичних засобів, коли необхідно усунення клітин-мішеней або тканинмішеней. ВИКЛАД СУТІ ВИНАХОДУ Даний винахід представляє композиції модифікованих константних доменів імуноглобуліну, які використовуються при інженерії терапевтичних засобів на основі антитіл або їм подібних. Імуноглобуліни класу IgG, які використовуються як терапевтичні засоби, які містять ділянку Fc і мають ліганди клітинної поверхні як мішені, є конкретними кандидатами для модифікації при використанні даних композицій. Модифіковані імуноглобуліни, які складають предмет даного винаходу, стійкі відносно протеолітичних ферментів середовища, мішенню яких є антигензв'язувальний домен IgG, наприклад, внутрішньопухлинного середовища, яке містить 1 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 антиген, який супутній раковим клітинам або судинній сітці пухлини, однак при цьому вони зберігають важливі ефекторні функції, пов'язані з ділянкою Fc антитіла. Відповідно з даним винаходом запропоновані способи отримання протеазостійких моноклональних антитіл IgG1 за допомогою введення мутацій в ділянку Fc IgG1, які роблять їх протеазостійкими. У додатковому аспекті функціональна активність IgG, що містять такі мутації, може бути відновлена за рахунок введення додаткових змін в послідовності амінокислот в дистальному положенні в ділянці Fc. Композиція, що складає предмет даного винаходу, являє собою протеазостійке антитіло IgG1, яке містить змінену послідовність амінокислотних залишків в нижньому шарнірі IgG1, які беруть участь у взаємодії Fc і FcγR. Крім того, було встановлено, що вони є сайтами фізіологічно значущого протеолітичного розщеплення. У одному аспекті залишки підкласу людського IgG1 замінюють відповідними залишками нижнього шарніра людського IgG2. У одному варіанті здійснення послідовність E233-L234-L235-G236 людського IgG1 замінюють на IgG2 P233-V234-A235 з делецією G236 (нумерація по EU (Edelman et al., Proc Natl Acad Sci USA 63:78-85, 1969 р.)). У іншому варіанті здійснення залишки IgG1 E233-L234-L235-G236-G237 замінюють на P233-V234-A235 з делецією G236, а G237 замінюють амінокислотою, вибраною із А, D, Р, Q або S. В іншому варіанті здійснення всі залишки константного домену IgG1, які відповідають залишкам 214-236, KVEPKSCDKT HTCPPCPAPEL LG (SEQ ID NO: 3), замінюють на послідовність відповідних положень в людському IgG2 або TVERKCCVEC PPCPAPPVA (SEQ ID NO: 4). У другому аспекті здатність протеазостійкого мутанта IgG1, що складає предмет даного винаходу, зв'язувати FcγR і/або комплемент може відновлюватися за рахунок введення додаткових мутацій. Таким чином, без таких додаткових замін протеазостійкий мутант IgG1, який містить протеазостійку шарнірну ділянку, яка містить P233-V234-A235 з делецією G236, або мутант, в якому всі послідовності залишків 214-236 константної ділянки IgG1 замінюють на відповідні послідовності IgG2 (SEQ ID NO: 4), відрізняється зниженою здатністю зв'язувати FcγR. Щоб вирішити проблему втрати ефекторної функції, що стосується зв'язування FcR і C1q, в константну ділянку, що не стосується шарніра, вводять додаткові мутації таким чином, щоб модифікована молекула IgG1 зберігала вимірну або навіть підвищену здатність зв'язувати FcγR в порівнянні з людським IgG1 дикого типу. Таким чином, в одному варіанті здійснення даного винаходу протеазостійкий IgG1 містить такі молекули, які містять домен Fc з послідовністю людського IgG1 в шарнірній ділянці і ділянці CH2 від приблизно залишків 214 до приблизно залишку 330 за системою EU, де щонайменше залишки 233-237 заміщені на PVA/ (делеція G236), а також введені спеціальні заміни, вибрані із S239D/I332E; K326A/E333A; H268F/S324T/I332E; F243L/R292P/Y300L; S239D/H268F/S324T/I332E; S267E/H268F/S324T/I332E; E333A/K334A; G237A/S239D/I332E; а також G237S/S239D/I332E; S298A/E333A/K334A; S239D/K326A/E333A і S267E/I332E. У конкретному варіанті здійснення молекула IgG, отримана на основі послідовностей константної ділянки IgG1 дикого типу, містить SEQ ID NO: 6. У іншому варіанті здійснення протеазостійкий IgG отримують із константних доменів IgG2 дикого типу із замінами при S239D і I332E. Таким чином, введення цих додаткових мутацій забезпечує функції Fc, зберігаючи при цьому протеазостійкість, а в деяких випадках поліпшує опосередковану Fc ефекторну функцію в порівнянні з диким типом IgG1 як в аналізах зв'язування FcγR in vitro, так і в клітинних аналізах in vitro, таких як антитілозалежна клітинноопосередкована цитотоксичність (ADCC). Протеазостійке моноклональне антитіло з ефекторною функцією, яке включає домен Fc, що складає предмет даного винаходу, використовується як протираковий і протимікробний терапевтичний засіб, причому можливе розщеплення IgG внаслідок підвищеного локального рівня протеази в сайтах росту пухлини або інфекції. У іншому аспекті композиції, описані в цьому документі, знаходять застосування як компоненти молекул терапевтичних засобів, які вводяться пацієнтам, що їх потребують, наприклад, ссавцям. У одному способі використання композиції антитіло, яке містить послідовності людського IgG1 дикого типу і лігандзв'язувальний домен, яке використовується для лікування пацієнтів, які потребують цього, модифіковано для включення протеазостійких шарнірних послідовностей P233-V234-A235 з делецією G236 і за необхідності включає модифікації, які поновлюють ефекторну функцію, вибрані із S239D/I332E; K326A/E333A; H268F/S324T/I332E; F243L/R292P/Y300L; K326A/I332E/E333A, S239D/K326A/E333A і S267E/I332E S239D/H268F/S324T/I332E; S267E/H268F/S324T/I332E; E333A/K334A; G237A/S239D/I332E; а також G237S/S239D/I332E. У одному варіанті здійснення композицію мутанта IgG1 Fc використовують при показаннях, коли для ефективності композиції важливе значення має активація FcγR, пов'язана з імунною і ефекторною функціями, такими як i) 2 UA 113725 C2 5 10 15 20 25 30 35 40 45 антитілозалежна клітинноопосередкована цитотоксичність (ADCC), ii) комплементозалежна цитотоксичність (CDC), iii) антитілозалежний клітинноопосередкований фагоцитоз (ADCP) і iv) опосередкована FcR клітинна активація (наприклад, вивільнення цитокінів за допомогою перехресного зшивання FcR) і v) опосередкована FcR активація/елімінація тромбоцитів. У одному аспекті мутації IgG1 Fc вводять в терапевтичні антитіла або Fc-злиті полівалентні зв'язувальні, що мають як мішені ліганди на клітинах проліферативних розладів, таких як клітини пухлини; клітини судинної сітки пухлини; фібробласти; активовані В-клітини або Тклітини, або клітини патогенного хазяїна, або клітини, отримані не від хазяїна, особливо бактерійні клітини. У іншому варіанті здійснення мутант IgG1 Fc містить фармацевтичну композицію. У іншому варіанті здійснення мутант IgG1 Fc містить частину фармацевтично активної молекули. Фармацевтичні композиції, що містять мутант IgG1 Fc або активні молекули, які містить мутант IgG1 Fc, використовують для лікування захворювань, що характеризуються небажаною або неконтрольованою проліферацією або міграцією клітин. У одному аспекті композиції даного винаходу з протеазостійким константним доменом в комбінації з лігандзв'язувальним доменом вводять пацієнту, який їх потребує, з використанням способу введення в ту частину організму, де були виявлені одна або більше протеаз, здатних розкладати молекули підкласу IgG1, з використанням способів системної доставки або місцевої доставки. КОРОТКИЙ ОПИС ФІГУР На фігурі 1 представлене зображення антитіла людського IgG1, доповнене амінокислотною послідовністю, присутньою в шарнірній ділянці, яка має важливе значення для взаємодії FcγR і комплементу, а також із вказуванням точок розщеплення протеазою. На фігурі 2 представлене зіставлення амінокислотних послідовностей частин константних ділянок людського IgG1 дикого типу (SEQ ID NO: 1) і IgG2 (SEQ ID NO: 2), зіставлених з новими конструкціями 2h S239D/I332E (SEQ ID NO: 8) і 2h E333A/K334A (SEQ ID NO: 9), із вказуванням відповідної нумерації за системою EU для кожного залишку; де шарнірна ділянка як IgG1, так і IgG2 в обох випадках містить цистеїнові залишки при залишках EU 226 і 229, як і в нових конструкціях. На фігурах 3A-B представлені аналізи розщеплення антитіл, які містять різні ізотипи IgG (А) і їх нові конструкції, як описано на фігурі 2 (В) в присутності протеази IdeS в різні моменти часу. На фігурах 4A-D представлене розщеплення конструкцій IgG в присутності IdeS (А), GluV8 (В), MMP-3 (С) і MMP-12 (D) через 24 години інкубації (n=2). На фігурах 5A-G представлені результати аналізу зв'язування FcγR за допомогою AlphaScreen® для груп протеазостійких конструкцій mAb: FcγRI (А), FcγRIIa (В і С), FcγRIIb (D і Е) і FcγRIIIa (F і G), де відновлення в % максимального сигналу відображає здатність неміченої конструкції конкурувати з біотинілованим IgG1 при зв'язуванні (n=2). На фігурах 6A-C представлені графіки окремих аналізів ADCP, проведених з протеазостійкими конструкціями mAb і IgG1, і IgG2 дикого типу, де % фагоцитозу по осі Y відображає загальне число відібраних клітин. На фігурах 7A-C представлені графіки окремих аналізів ADCC, проведених з протеазостійкими конструкціями mAb і IgG1, і IgG2 дикого типу, де % лізису по осі Y визначається відносно 100 % лізису тієї ж кількості клітин під впливом детергенту (n=2). На фігурі 8 представлений графік аналізу CDC, проведеного з протеазостійкими конструкціями mAb і IgG1, і IgG2 дикого типу, де % лізису по осі Y визначається відносно 100 % лізису тієї ж кількості клітин під впливом детергенту (n=2). КОРОТКИЙ ОПИС СПИСКУ ПОСЛІДОВНОСТЕЙ SEQ ID NO: 1 2 3 4 5 6 Опис IgG1-Fc; людський Ig класу гамма, підклас, шарнір, домени CH2 і CH3 IgG2-Fc; людський Ig класу гамма, підклас 2, шарнір, домени CH2 і CH3 Шарнірна ділянка IgG1, EU 214-236 Шарнірна ділянка IgG2 (2 hc), EU 214235 Гібридна шарнірна ділянка IgG (2h), IgG2 EU 233-235 2hc (EU 214-447) 3 UA 113725 C2 7 8 9 10 11 12 13 14 15 16 17 18 19 20 5 10 15 20 25 30 35 40 45 2h (EU 214-447) 2h S239D/I332E 2h E333A/K334A 2h F243L/R292P/Y300L 2h H268F/S324T/I332E 2h S239D/H268F/S324T/I332E 2h S267E/H268F/S324T/I332E 2h K326A/E333A 2h G237X/S239D/I332E, де X являє собою А, D, Р, Q або S 2hc S239D/I332E IgG2 S239D/I332E 2h K326A/I332E/E333A 2h S239D/K326A/E333A 2h S267E/I332E ДОКЛАДНИЙ ОПИС ВИНАХОДУ Скорочення ADCC = антитілозалежна клітинна цитотоксичність; ADCP = антитілозалежний клітинний фагоцитоз; CDC = комплементозалежна цитотоксичність; FDCR=Fc-залежне вивільнення цитокінів; FcγR або Fc-гамма-R=Fc-гамма рецептор; GluV8 = глутамілендопептидаза V8 Staphylococcus aureus; IdeS = розкладаючий імуноглобулін фермент Streptococcus pyogenes; IgG = імуноглобулін G; ITAM = мотив активації імунорецептора на основі тирозину; ITIM = мотив інгібування імунорецептора на основі тирозину; mAb = моноклональне антитіло; MMP = матриксна металопротеїназа; термін "протеаза" еквівалентний терміну "протеїназа" і вони використовуються рівнозначно; PR = протеазостійкий. Визначення і пояснення термінології Терміни "антитілозалежна клітинна цитотоксичність", "антитілозалежна клітинноопосередкована цитотоксичність" або ADCC стосуються такої форми цитотоксичності, при якій секретований Ig, зв'язаний з рецепторами Fc (FcR), які присутні на певних цитотоксичних клітинах (наприклад, на клітинах натуральних кілерах (NK), нейтрофілах і макрофагах), сприяє специфічному зв'язуванню цих цитотоксичних ефекторних клітин з клітиною-мішенню, яка є носієм антигену, з подальшим знищенням клітини-мішені цитотоксинами. Лігандспецифічні високоафінні антитіла IgG, націлені на поверхню клітинмішеней, стимулюють цитотоксичні клітини і абсолютно необхідні для такого роду знищення. Лізис клітини-мішені має позаклітинну природу, вимагає прямого контакту клітина-клітина, і в даному процесі комплемент не задіяний. Можна проводити аналіз здатності будь-якого конкретного антитіла опосередковувати лізис клітини-мішені за рахунок ADCC. Для оцінки активності ADCC антитіло, яке аналізується, додають до клітин-мішеней, що мають цільовий ліганд, в комбінації з імунними ефекторними клітинами, які можуть бути активовані комплексами антиген-антитіло з подальшим цитолізом клітини-мішені. Цитоліз по суті виявляють за вивільненням мітки (наприклад, радіоактивних субстратів, флуоресцентних барвників або природних внутрішньоклітинних білків) із лізованих клітин. Придатні для таких аналізів ефекторні клітини включають мононуклеарні клітини периферичної крові (PBMC) і клітини натуральні кілери (NK). Конкретні приклади аналізів in vitro ADCC описані в роботах Wisecarver et al., 1985 р., 19:211; Bruggemann et al., 1987 р., J Exp Med 166:1351; Wilkinson et al., 2001 р., J Immunol Methods 258:183; Patel et al., 1995 р., J Immunol Methods 184:29 (кожна з яких включена в даний документ шляхом посилання). У альтернативному варіанті здійснення або додатково активність ADCC антитіла, що розглядається, може оцінюватися in vivo, наприклад, в моделі на тваринах, такій як описана в роботі Clynes et al., 1998 р., PNAS USA 95:652, зміст якої повністю включений в цей документ шляхом посилання. Якщо ефекторна клітина діє в основному за допомогою фагоцитозу, процес може називатися антитілозалежним клітинним фагоцитозом (ADCP). "Комплементорієнтована цитотоксичність" або CDC означає таку форму цитотоксичності, при якій каскад комплементу активується зв'язуванням компонента комплементу C1q з антитілом Fc. Термін "ефекторні функції" включає такі взаємодії Fc з Fc-гамма-рецепторами (FcγR), експресованими на імунних клітинах, а також взаємодії домену Fc з комплементом, які приводять до усунення експресувальної антиген-клітини за допомогою літичних процесів або в ході фагоцитозу ефекторними клітинами і компонентами комплементу. 4 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 У даному документі терміни "Fc", "Fc-вмісний білок" або "Fc-вмісна молекула" означають мономерні, димерні або гетеродимерні білки, які мають щонайменше домен CH2 і CH3 імуноглобуліну. Домени CH2 і CH3 можуть утворювати щонайменше частину димерної ділянки білка/молекули (наприклад, антитіла). У цьому документі термін "антитіло" означає конкретну форму Fc-вмісного білка, який містить щонайменше один лігандзв'язувальний домен, який містить або зберігає значну гомологію відносно щонайменше одного тяжкого або легкого ланцюга варіабельного домену антитіла щонайменше одного виду антитіла тварин. Константні послідовності людського підкласу IgG дикого типу каталогізовані в базі даних UniProt, яка доступна в режимі он-лайн, під кодами P01857 (IgG1), P01859 (IgG2), P01860 (IgG3) і P01861 (IgG4). У даному документі термін "ділянка людського IgG1 Fc дикого типу" означає ділянку людського IgG Fc, яка містить амінокислотну послідовність SEQ ID NO: 1 або її фрагмент, який містить від залишку K214 до залишку K447 тяжкого ланцюга людського IgG згідно з нумерацією EU по Kabat. Амінокислоти в константній ділянці нумеруються згідно із зіставленням з антитілом людського IgG1 за EU (див. Cunningham et al., 1970 р., Biochemistry, 9: 3161-70). Таким чином, тяжкий і легкий ланцюги антитіла зіставляються з тяжким і легким ланцюгами EU для досягнення максимальної ідентичності амінокислотної послідовності, і кожній амінокислоті в антитілі привласнюється той же номер, що і відповідній амінокислоті в EU. Систему нумерації EU по загальній угоді прийнято використовувати фахівцями в даній галузі (див., загалом роботу, Kabat et al., Sequences of Protein of Immunological Interest, NIH Publication No. 91-3242, US Department of Health and Human Services (1991 р.)). Згідно з угодою при зіставленні константної ділянки IgG2 дикого типу і EU виявляється відсутність амінокислот в положеннях 221-223 і 236 (фігура 2, SEQ ID NO: 2). Послідовності константного домену тяжкого ланцюга IgG ссавців позначаються в послідовності як CH1-шарнір-CH2-CH3. "Шарнір", "шарнірна ділянка" або "шарнірний домен" IgG по суті визначається як ділянка, що включає Glu216 і закінчується на Pro230 людського IgG1 за системою Kabat, але з функціональної точки зору можна вважати, що гнучка частина ланцюга містить додаткові залишки, якими закінчуються верхня і нижня ділянка шарніра, як, наприклад, з Glu216 по Gly237 (Roux et al., J. Immunol. 1998 р., 161:4083), а під нижнім шарніром розуміють ділянку Fc залишків 233-239, де по суті відбувалося зв'язування Fc-гамма-R. Шарнірні ділянки інших ізотипів IgG можуть порівнюватися з послідовністю IgG1 з вирівнюванням першого і другого цистеїнових залишків, які утворюють зшивання S-S між тяжкими ланцюгами. Незважаючи на можливе незначне розходження по межах згідно з нумерацією за системою Kabat, домен CH1 прилягає до домену VH, а аміно-кінець - до шарнірної ділянки тяжкого ланцюга молекули імуноглобуліну, і включає перший (в основному аміно-кінець) домен в константній ділянці тяжкого ланцюга імуноглобуліну, наприклад, приблизно з положень EU 118215. Домен Fc проходить від 231 до 447 амінокислотних залишків; домен CH2 - від приблизно Ala231 до Lys340 або Gly341, а CH3 - від приблизно Gly341 або Gln342 до Lys447. Залишки константної ділянки тяжкого ланцюга IgG ділянки CH1 закінчуються на Lys. Термін "протеазостійкий" означає здатність молекули, яка містить пептидні зв'язки, протистояти гідролітичному розщепленню одного або більше із своїх пептидних зв'язків в присутності протеолітичного ферменту. Стійкість до протеолітичних ферментів є відносною властивістю і порівнюється з молекулою, яка в меншому ступені здатна протистояти гідролітичному розщепленню одного або більше своїх пептидних зв'язків за вказаний період часу і у вказаних умовах, включаючи рН і/або температуру, при яких перевіряється стійкість до розщеплення. Один результат протеолітичного розщеплення, який вказує на те, що такий розпад стався, являє собою створення більш дрібних фрагментів (з більш низькою молекулярною масою) в порівнянні з молекулярною масою інтактної нерозщепленої материнської молекули. Термін "терапевтично ефективна кількість" означає кількість терапевтичного агента, описаного в цьому документі, який містить домен Fc, що може являти собою антитіло, фрагмент антитіла або його похідне для лікування того або іншого захворювання або розладу у пацієнта. Загальний опис винаходу Створення даного винаходу визначено спробами визначити домен Fc для використання при виробництві терапевтичних антитіл, злитих Fc і подібних біофармацевтичних засобів з підвищеною стійкістю до протеолізу in situ і із здатністю, яка зберігає здатність викликати вивільнення цитокінів, пошкоджувати або знищувати клітини-мішені, які мають антиген, і тканини, які оточують клітини-мішені. Протеази поділяються на п'ять основних груп у відповідності із структурою каталітичного сайта і амінокислоти (як однієї із складових), що має велике значення для їх активності: 5 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 серинові протеїнази, треонінові протеїнази, цистеїнові (тіолові) протеїнази, аспарагінові протеїнази і металопротеїнази. Різні позаклітинні протеази функціонують всередині організму і у відділах тіла, здійснюючи важливі регулюючі і метаболічні процеси. Кислотостійкі протеази, секретовані в шлунок (такі як пепсин), і серинові протеази, які присутні в дванадцятипалій кишці (трипсин і хімотрипсин), забезпечують розкладання білків із їжі в шлунково-кишковому тракті; протеази, присутні в крові або сироватці (тромбін, плазмін, фактор Хагемана і т. п.), грають важливу роль в процесах згортання крові, а також лізису тромбів і регуляції клітин імунної системи. Протеази присутні або вивільняються в лейкоцитах (еластаза, катепсин G). Протеази визначають час життя інших білків і, таким чином, грають важливу роль в метаболізмі. На відміну від гормонів, інтерлейкінів або хемокінів, механізм експресії білка не вимагає ніякої внутрішньоклітинної передачі або змін, що робить протеолітичний контроль одним із самих швидких механізмів регуляторного переключення. Спільна дія протеаз, наприклад, в каскадних реакціях, додатково приводить до швидкої і ефективної ампліфікації відгуку організму на фізіологічний сигнал. Ізотипи людського IgG (підкласи зрілих антитіл гамма-глобуліну класу G; IgG1, IgG2, IgG3 і IgG4) показують диференційовану здатність посилювати імунні функції. Наприклад, антитілозалежна клітинна цитотоксичність (ADCC) стимулюється IgG1 і IgG3, антитілозалежний клітинний фагоцитоз (ADCP) стимулюється IgG1, IgG2, IgG3 і IgG4, а комплементозалежна цитотоксичність (CDC) стимулюється IgG1 і IgG3. Ізотипспецифічне включення таких імунних функцій засноване на селективності рецепторів Fc відносно окремих імунних клітин і здатності зв'язувати C1q, за допомогою цього активуючи утворення мембраноатакуючого комплексу (MAC). Серед різних ізотипів відносна афінність до рецепторів Fcγ, які включають FcγRI, FcγRIIa/b/с і FcγRIIIa/b, є високою для IgG1 і IgG3. Однак афінність Fcγ до IgG2 значно нижче, за винятком поліморфізму FcγRIIa H131, і IgG4 виявляє лише афінність до FcγRI, яка вимірюється. На основі порівняльного аналізу послідовності і структур співкристалізації ключові контактні залишки для зв'язування рецептора співставлялися з амінокислотними залишками, які входять в ділянку нижнього шарніра і ділянку CH2 (Hezereh et al., J Virol 75:12161-12168 2001 р.; Shields et al., J Biol Chem 276:6591-6604, 2001 р.). У багатьох дослідженнях, проведених раніше, був зроблений висновок про те, що заміщення в залишках нижнього шарніра IgG2 (положення за EU 233-235) порушують опосередковану FcγR функцію і активацію комплементу. У одній роботі заміщення E233PL234V-L235A з делецією G236 в нижньому шарнірі IgG1 було одним варіантом панелі конструкцій Fc людського IgG1, для яких була продемонстрована втрата опосередкованої Fc ефекторної функції (Armour et al., 1999 р., Eur J Immunol 29:2613-2624). Ця інформація показує, що заміна залишків в шарнірі IgG1 або поруч з ним на відповідні залишки IgG2 приводила до помітної втрати афінності зв'язування з FcγRs і втрати здатності впливати на фіксацію комплементу і знищення клітин. По-друге, деякі із вказаних положень залишків стосуються саме тих, в яких протеази розщеплюють послідовність IgG1 (фігура 1) і які зайняті різними амінокислотами в більш протеолітично стійкій послідовності IgG2, як продемонстровано при співставленні людських IgG1 і IgG2 (фігура 2). Автори даного винаходу використовували цю інформацію як відправну точку для створення протеазостійкого IgG-Fc, але з компетентною ефекторною функцією. У даному винаході вперше продемонстровані заміщення у множині положень константних ділянок IgG1 (Fc), які несподіваним чином дозволяють створити протеазостійкий і функціональний (FcgR-зв'язувальний) домен Fc. Фахівцям в даній галузі буде зрозуміло, що, якщо відомі властивості домену Fc, який має специфічну амінокислотну послідовність, інформацію можна використати для конструювання або модифікації інженерії існуючих антитіл або поліпептидних злитих Fc. Протеазостійкість, яку мають композиції, що складають предмет даного винаходу, розповсюджується, наприклад, на протеази, які розщеплюють в залишках IgG1 при шарнірній ділянці, заміщених на альтернативні амінокислоти, отримані із відповідного залишку IgG2, наприклад, MMP-3, MMP-7, MMP-12, HNE, плазмін, катепсин G, пепсин, IdeS або глутамілендопептидаза I Staph aureus (фігура 1) (Ryan et al., 2008 р. supra). Заміщені по множині положень мутанти IgG1 вибирали на основі їх відносної афінності до людських FcR (FcγRI, FcγRIIa, FcγRIIb, FcγRIIIa, оцінку проводили по конкурентних аналізах AlphaScreen®). Ці мутанти додатково тестували у відповідних клітинних системах по їх здатності індукувати ADCC в присутності PBMC і ADCP в присутності макрофагів, диференційованих in vitro. У експериментальних даних, представлених в цьому документі, IgG1 з вказаними введеними мутаціями (таблиця 2) являли собою рекомбінантні протеазостійкі mAb, похідні від IgG1 з мутацією, що посилює Fc. 6 UA 113725 C2 5 Альтернативні композиції залишків 214-237 в шарнірній ділянці IgG1 за системою нумерації EU (SEQ ID NO: 3) отримали з використанням людського IgG1 дикого типу (wt) (SEQ ID NO: 1), а для забезпечення протеазостійкості проводили заміну всієї послідовності IgG2 (SEQ ID NO: 4) шарніра або химерного шарніра (SEQ ID NO: 5), причому, як показано в таблиці 1, мінялися тільки залишки 233-235. Таблиця 1 Каркас IgG (EU 214-447) Назва Fc IgG1 wt (UniProt P01857, 96-329) hIgG1 IgG2 wt (UniProt P01859, 97-326) hIgG2 Шарнір/проксимальна CH2 (EU 214-236) KVEPKSCDKTHTCPPCPAPELLG (SEQ ID NO:3) TVERKCCVECPPCPAPPVA (SEQ ID NO:4) 2 hc hIgG1 із усією шарнірною ділянкою IgG2 SEQ ID NO:4 2h hIgG1 KVEPKSCDKTHTCPPCPAPPVA (SEQ ID NO:5) Компенсувальні мутації в ділянках, які посилюють ефекторну функцію, як показано нижче в таблиці 2, можуть бути вибрані із раніше описаних заміщень. 10 Таблиця 2 Мутації (нумеровані за EU положеннями) S239D/I332E E333A/K334A F243L/R292P/Y300L H268F/S324T/I332E S239D/H268F/S324T/I332E S267E/H268F/S324T/I332E K326A/E333A K326A/I332E/E333A S239D/K326A/E333A S267E/I332E G237X/S239D/I332E, де X являє собою А, D, Р, Q або S S298A/E333A/K334A 15 20 Контроль 1 5 3 4 4 4 2 1, 2 1, 2 1, 4 1 5 Конструкції з підвищеною функцією Fc раніше цитувалися в наступних роботах: 1. Lazar, Proc Natl Acad Sci USA 103:4005-4010 (2006 р.), 2. Idusogie, J Immunol 166:2571-2575 (2001 р.), 3. Stavenhagen, Cancer Res 67(18):8882-90 (2007 р.), 4. Moore et al. MAbs 2(2):181-189 (2010 р.), 5. Shields et al., J Biol Chem 276:6591-6604 (2001 р.). За різними показниками функції Fc, отриманими на основі аналізів in vitro, було визначено декілька протеазостійких послідовностей Fc, які при введенні в повну структуру IgG (H2L2) забезпечували стійкість відносно однієї або більше протеаз, які діють за залишками нижнього шарніра (EU232-237), зберігаючи при цьому здатність зв'язувати FcγR або викликати цитоліз. Зміни активності, пов'язаної з Fc, для вибраних конструкцій, які містять зміну в шарнірі, а також в ділянці CH2, і поділених за афінністю рецептора і біологічною активністю in vitro, представлені в таблиці 3. 25 7 UA 113725 C2 Таблиця 3 Ізотип/конструкція IgG1 IgG2 IgG1 2h (PVA) IgG1 2h E333A/K334A IgG1 2h S239D/I332E IgG1 2h S239D/H268F/S324T/I332E IgG1 2h S267E/H268F/S324T/I332E IgG1 2h K326A/E333A 5 10 15 20 25 30 35 40 45 FcγRI +++++ +++ FcγRIIa ++++ ++ + +++++ FcγRIIb ++++ ++++ FcγIIIa ADCC ADCP CDC +++ +++++ +++++ +++++ +/++ +/+ ++ +/++ не визн. ++++ +++++ ++++++ не визн. ++++ +++++ ++++ ++++++ ++++++ не визн. +++++ ++++++ +++ +/ + +++++ не визн. +/ +/ +++ +++++ Спосіб отримання молекул із зміненою Fc-вмісною ділянкою Сайти для заміщення вибирають в залежності від необхідності отримати композицію, яка має структурні елементи нативного антитіла Fc, зберегти стабільність, утримувати зв'язування FcR і здатність стимулювати каскад комплементу, лізис клітин, фагоцитоз клітин або вивільнення цитокінів. Білки, зокрема багатомірні молекули з довгими поліпептидними ланцюгами, із зміненими або мутованими амінокислотами, зручно отримувати за допомогою модифікації нуклеїнової кислоти в межах вектора експресії, який кодує материнську послідовність для зміни відповідного генетичного коду для необхідної амінокислоти. Генетичний код і такі способи добре відомі фахівцям в даній галузі. При створенні химерної послідовності, такої як Fc, яка складає предмет даного винаходу, що містить частини IgG1 і частини IgG2, можна провести сплайсинг більш великих сегментів відповідних кодуючих нуклеїнових кислот або заміну сегментів за допомогою стандартних методик клонування. Протеолітичне тестування Щоб визначити, чи є та або інша Fc-вмісна композиція або антитіло більш протеазостійкою в порівнянні з іншою або композицією дикого типу, оцінюються швидкість або ступінь розщеплення різних виділених Fc-вмісних композицій або антитіл в присутності протеолітичного ферменту. Після певного періоду часу вимірюють розкладання різних композицій за допомогою способу, який дозволяє виявити безпосереднє розщеплення ланцюга, таке як утворення унікальної структури в сайті розщеплення або вимірювання фрагментів, які знову утворилися. У альтернативному варіанті здійснення, якщо розщеплення приводить до втрати активність, може проводитися функціональний аналіз, включаючи аналіз зв'язування. Протеолітичне розщеплення IgG1 може пройти по будь-якому із чотирьох поліпептидних ланцюгів димерної гетеродимерної структури. Розщеплення IgG приводить до створення добре охарактеризованих фрагментів, таких як Fab, (Fab')2, і фрагментів Fc із приблизними, але унікальними молекулярними масами. Розділення таких фрагментів, отриманих в ході експерименту протеолізу, може досягатися за допомогою ексклюзійної хроматографії (SEC), гель-електрофорезу, аналізів MALDI-TOF-MS (час-пролітної мас-спектрометрії з матриксною лазерною десорбцією/іонізацією) після деглікозилування з використанням PNGаse F (пептид-Nглікозидаза F), як це було описано раніше (WO2007024743A2, WO2009045894A1). Отже, під стійкою до протеолітичного розщеплення Fc-вмісною композицією, розуміють те, що та або інша композиція з меншою імовірністю буде розщеплюватися, втрачати активність, втрачати афінність до зв'язуючого Fc партнеру, такому як FcR, після впливу протеолітичного ферменту в порівнянні з порівнюваною молекулою, такою як дикий тип людського IgG1. Біологічні характеристики мутантів Функціональність Fc-вмісних білків можна порівнювати за допомогою декількох добре відомих аналізів in vitro. Зокрема, являє інтерес афінність до членів сімейства FcγRI, FcγRII і FcγRIII рецепторів Fcγ. Дані вимірювання можна проводити з використанням рекомбінантних розчинних форм рецепторів або клітинноасоційованих форм рецепторів. Крім того, можна вимірювати афінність до FcRn, рецептору, відповідальному за тривалий період напівжиття IgG, за допомогою набору у вигляді лігандзв'язаних гранул, такого як ALPHASCREEN з рекомбінантним розчинним FcRn. AlphaScreen®, що використовується в скринінзі з високою пропускною спроможністю, являє собою технологію гомогенного аналізу, який дозволяє реєструвати події на молекулярному рівні, такі як зв'язування. В основі технології аналізу AlphaScreen® лежать гранули "донора" і "акцептора" з покриттям. Аналіз AlphaScreen® з використанням гранул забезпечується за рахунок взаємодії гранул при зближенні одна з одною, 8 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 що приводить до каскаду хімічних реакцій, які приводять до значно більш ампліфікованого сигналу. Наприклад, при конкурентному зв'язуванні для оцінки відносної афінності або авідності білків можуть застосовуватися прямі або непрямі оцінки. Внаслідок природної еволюції ізотипів людського IgG (наприклад, IgG1, IgG2, IgG3 і IgG4) кожен із них показує різний спектр здатностей мобілізувати імунні функції, такі як антитілозалежна клітинна цитотоксичність (ADCC, наприклад, IgG1 і IgG3), антитілозалежний клітинний фагоцитоз (ADCP, наприклад, IgG1, IgG2, IgG3 і IgG4) і комплементозалежна цитотоксичність (CDC, наприклад, IgG1, IgG3). Ізотипспецифічне включення даних функцій засноване на диференційованій селективності до рецепторів Fc, які знаходяться в різних імунних клітинах, і здатності зв'язувати C1q і активувати збирання мембраноатакуючого комплексу (MAC) з подальшими CDC і CDP (комплементозалежний фагоцитоз) за допомогою специфічного рецепторного зв'язування компонентів комплементу на ефекторних макрофагах. Ієрархія здатності зв'язувати вихідний компонент, C1q, каскаду комплементу для людських ізотипів виглядає таким чином: IgG1>IgG3>IgG2>IgG4, причому здатність IgG2 і IgG4 активувати комплемент при мікробних інфекціях підтверджена документально. Функціональні клітинні аналізи, такі як аналізи ADCC і CDC, дають уявлення про вірогідні функціональні наслідки конкретних конструкційних структур. Антитілозалежна клітинноопосередкована цитотоксичність (ADCC) являє собою клітинноопосередковану реакцію, при якій неспецифічні цитотоксичні клітини, які експресують рецептори Fc (FcR) (наприклад, клітини натуральні кілери (NK), нейтрофіли і макрофаги), виявляють зв'язане антитіло на клітині-мішені і потім викликають лізис клітини-мішені. У одному варіанті здійснення аналіз ADCC виконаний з можливістю присутності NK-клітин як початкової клітини-ефектора, яка відображає функціональні ефекти впливу на FcγRIIIa, який, як відомо, є єдиним активуючим рецептором Fcγ-типу, експресованим даними клітинами. Для порівняння імунних ефекторних функцій різних мутантів також можна використовувати аналіз фагоцитозу, а також аналізи, які вимірюють клітинні реакції, такі як вивільнення супероксиду або медіатору запалення. Також можна використати моделі in vivo, наприклад, при використанні мутантів анти-CD3-антитіл для вимірювання активації Т-клітин у мишей активності, яка залежить від зв'язування доменів Fc із специфічними лігандами, такими як рецептори Fcγ. Антитілонаправлена активація макрофагів опосередковує антитілозалежний клітинний фагоцитоз (ADCP), внаслідок чого опсонізовані клітини-мішені поглинаються і перетравлюються макрофагами. In vitro диференційовані макрофаги, експресуючі високі рівні FcR, можуть диференціюватися по фенотипу М1 під впливом INFγ або GM-CSF до експресованих підвищених рівнів всіх FcR (FcγRI, FcγRIIa, FcγRIIIa) в порівнянні з моноцитами. Отримання мутантів антитіл Композиції, що складають предмет даного винаходу, являють собою складні білки, які найбільш зручно отримувати в рекомбінантній клітині-хазяїні. Як описано в цьому документі, клітина-хазяїн, вибрана для експресування рекомбінантного Fc-вмісного білка або моноклонального антитіла, впливає на кінцеву композицію, включаючи без обмежень варіацію в композиції оточуючих білок фрагментів олігосахаридів в домені CH2 імуноглобуліну. Таким чином, один аспект даного винаходу включає вибір відповідної клітини-хазяїна, яка несе полінуклеотидні послідовності, які кодують конструкції, що складають предмет даного винаходу, що містять Fc, для застосування як продуктивної клітини, яка експресує необхідний терапевтичний білок, або для створення такої клітини. Клітина-хазяїн може додатково походити від ссавця або може бути вибрана із клітин COS-1, COS-7, HEK293, BHK21, СНО, BSC-1, Hep G2, 653, SP2/0, 293, HeLa, клітин мієломи, лімфоми, дріжджів, клітин комах або рослин, або будь-яких їх похідних, іморталізованих або трансформованих клітин. В альтернативному варіанті здійснення клітина-хазяїн може бути вибрана із клітин видів або організмів, нездатних глікозилувати поліпептиди, наприклад, прокаріотичних клітин або організмів, таких як природні або рекомбінантні бактерії виду Е. coli, Klebsiella або Pseudomonas, клітини рекомбінантних рослин або комах. Глікозилування при природному сайті глікозилування у тяжкому ланцюгу IgG (N297, нумерація за EU) також робить внесок в афінність зв'язування Fc до FcγR. Оскільки в залежності від ізотипу міняються константні ділянки, кожен ізотип має відмінний набор Nзв'язаних вуглеводних структур, які по-різному впливають на збирання, секрецію і функціональну активність білка (Wright А. і Morrison S. L., Trends Biotech. 15:26-32 (1997 р.)). Структура приєднаного N-зв'язаного вуглеводу істотно змінюється в залежності від ступеня обробки і може включати залишки розгалужених і 2-антенарних складних олігосахаридів з високим вмістом манози і сіалової кислоти (N-ацетилнейрамінова кислота або NANA), фукози, 9 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 галактози і GlcNAc (N-ацетилглюкозамін) як кінцевих цукрів. Відмічали вплив на ефекторні функції клітини-хазяїна і вміст олігосахаридів в антитілах (Lifely M. R., et al., 1995 р., Glycobiology 5: 813-822; Jefferis R., et al., 1998 р., Immunol Rev. 163: 59-76; Wright А. і Morrison S. L., supra; Presta L., 2003 р., Curr Opin Struct Biol. 13(4): 519-25). Більш того, як відомо, відносно цукрового ланцюга в антитілі додавання або модифікація фукози в проксимальному N-ацетилглюкозаміні на поновлюючому кінці N-глікозид-зв'язаного цукрового ланцюга антитіла значно змінює ADCCактивність антитіла (WO00/61739). Відносний внесок галактозилування 2-антенарних олігосахаридів, присутність GlcNac, яка розщеплюється, і фукозилування додатково вказують на те, що нефукозиловані mAb виявляють більшу здатність активізувати ADCC, як показують оцінки in vitro і in vivo, ніж інші модифікації Nзв'язаних 2-антенарних олігосахаридних структур (Shields, et al., 2002 р., J Biol Chem. 277: 26733-40; Niwa, et al., 2004 р., Cancer Res. 64: 2127-2133). Експресія або виробництво протеазостійких mAb, що складають предмет даного винаходу, за допомогою клітини-хазяїна, здатної, рекомбінантної (наприклад, нокаутом або пригніченням конкретних ферментів Shinkawa, et al., 2003 р., J. Biol. Chem., 278: 3466-3473; EP1176195A1) або індукованої, наприклад, за допомогою контролю зовнішніх умов або живлення таким чином, щоб виробляти mAb з низьким змістом фукози, входить в об'єм даного винаходу. Антитіла, Fc і Fc-злиті білки Антитілозв'язувальний домен або його фрагменти можуть бути отримані за допомогою способів, відомих фахівцям в даній галузі, і в комбінації з інформацією, наведеною в цьому документі, можуть включати послідовності або бути отримані від будь-якого ссавця, такого як без обмежень чоловік, миша, кролик, щур, примат, коза або будь-яка їх комбінація. Такі антитіла можуть забезпечувати основні структури і компоненти зв'язувальних доменів для використання при виробництві конструкцій антитіл, що складають предмет даного винаходу. У одному аспекті даного винаходу антитілозв'язувальні домени отримують із гібридом, приготованих за допомогою імунізації миші або іншої тварини цільовими пептидами, клітинами або екстрактами тканин. Антитіла можуть бути отримані з використанням будь-яких гібридомних методик, добре відомих фахівцям в даній галузі, див., наприклад, роботу Harlow і Lane, antibodies, а Laboratory Manual, Cold Spring Harbor, NY (1989 р.), зміст якої повністю включений в цей документ шляхом посилання, або за допомогою вибору продукуючого антитіла лімфоциту і клонування послідовностей нуклеїнових кислот, які кодують зв'язувальні домени, з використанням методик, відомих фахівцям в даній галузі. Даний винахід стосується константної ділянки людського IgG. Отже, в об'єм даного винаходу входять будь-яке антитіло або злитий білок, що містить людський Fc-домен, для яких необхідно, щоб кінцева композиція виявляла як протеолітичну стійкість (як описано в даному винаході), так і здатність зв'язувати FcgR і стимулювати ADCC, ADCP і/або CDC в аналізах in vitro. Цільовий фрагмент, який включає без обмежень Fv антитіла або одиничний варіабельний домен, може за необхідності зливатися з Fc-структурою. Fv містить димер домену варіабельної ділянки одного тяжкого і одного легкого ланцюга, жорстко зв'язаних нековалентним зв'язком. Внаслідок фолдинга даних двох доменів виникає шість гіперваріабельних петель (3 петлі, кожна із Н- і Lланцюга), які надають амінокислотні залишки для зв'язування антигену і забезпечують специфічність зв'язування антитіла з антигеном. Однак навіть одиничний варіабельний домен (або половина Fv, що містить тільки три CDR, специфічних відносно антигену) мають здатність розпізнавати і зв'язувати антиген, хоч часто з більш низькою афінністю, ніж весь сайт зв'язування. "Одноланцюгові Fv", які також скорочуються як sFv або scFv, являють собою фрагменти антитіла, які містять домени антитіла VH і VL, зв'язані в єдиний поліпептидний ланцюг. Поліпептид scFv по суті містить поліпептидний лінкер між VH і VL. Цільовий фрагмент також може бути вибраний із паратопу антитіла (зв'язувальні залишки без обмежень з CDR або структур варіабельного домену); ферменту; гормону; рецептора; цитокіну; поверхневого антигену імунної клітини і адгезивної молекули, в якій конструкцію отримують повністю рекомбінантними способами. Цільовий фрагмент також може мати небілкову природу, як наприклад, вуглевод, ліпід, ліпополісахарид, органічна молекула або метал, або комплекс металу. По суті, якщо вводиться цільова молекула, вона буде зв'язана з Fc за допомогою лінкера, який може являти собою поліпептид або неполіпептид. Fc-вмісні білки або Fc-фрагменти, описані в цьому документі, можуть бути отримані декількома способами, добре відомими в даній галузі. Антитілозв'язувальні домени або злиті Fc-білки або їх компоненти і домени також можуть бути отримані із бібліотек таких доменів або компонентів, як, наприклад, бібліотеки фагів. Бібліотеку фагів можна створити за допомогою вставки бібліотеки випадкових олігонуклеотидів або бібліотеки полінуклеотидів, які містять послідовності, такі В-клітини імунізованої тварини або людини (Smith G. P., 1985 р., Science 228: 10 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 1315-1317). Бібліотеки фагів антитіл містять пари варіабельних ділянок тяжкого (Н) і легкого (L) ланцюгів в одному фазі, який дозволяє експресувати фрагменти Fv із одним ланцюгом або фрагменти Fab (Hoogenboom, et al., 2000 р., Immunol. Today 21(8) 371-8). Різноманітність фагомідної бібліотеки можна адаптувати для підвищення і/або зміни імунологічних специфічностей моноклональних антитіл бібліотеки для отримання і подальшої ідентифікації додаткових необхідних моноклональних антитіл людини. Наприклад, кодувальні гени тяжкого (Н) і легкого (L) ланцюгів молекули імуноглобуліну можуть бути випадково перемішані (перетасовані), щоб при збиранні молекули імуноглобуліну утворити нові пари HL. Крім того, кодувальні гени одного або обох із Н- і L-ланцюгів можуть бути мутовані в ділянку, яка визначає комплементарність (CDR), варіабельної ділянки імуноглобулінового поліпептиду з подальшим скринінгом необхідної афінності і здатності до нейтралізації. Бібліотеки антитіл або Fc можуть також створюватися синтетично за допомогою вибору однієї або більше людських каркасних послідовностей і введення колекцій кластерів CDR, отриманих із репертуарів людських антитіл або за допомогою цілеспрямованої варіації (Kretzschmar і von Ruden, 2000 р., Current Opinion in Biotechnology, 13: 598-602). Положення, які визначають різноманітність, не обмежуються CDR, але можуть також включати каркасні сегменти варіабельних ділянок або можуть включати варіабельні ділянки, які не стосуються антитіла, такі як пептиди. В одному аспекті фагова бібліотека, здатна забезпечувати дисплей димерних структур Fc, зв'язаних з білком оболонки фага pIX, як описано в заявці на патент США (№ 61/261767), що направлена заявниками, яка одночасно розглядається, може використовуватися для вибору нових Fc-вмісних структур відповідно з даним винаходом. Бібліотеки зв'язувальних компонентів-мішеней, які можуть включати зв'язувальні компоненти-мішені, відмінні від варіабельних ділянок, що стосуються антитіла, можуть включати бібліотеки рибосомного дисплея, бібліотеки дріжджового дисплея і бібліотек бактерійного дисплея. Рибосомний дисплей являє собою спосіб трансляції мРНК у відповідні білки із збереженням білка прикріпленим до РНК. Кодувальну послідовність нуклеїнової кислоти визначають методом ОТ-ПЦР (Mattheakis L.C. et al., 1994 р., Proc. Natl. Acad. Sci. USA 91, 9022). Дріжджовий дисплей заснований на конструюванні злитих білків зв'язаного з мембраною альфааглютинінового рецептора зв'язування дріжджів, aga1 і aga2, частини системи типу спаровування (Broder et al., 1997 р., Nature Biotechnology, 15: 553-7). Бактерійний дисплей заснований на злитті мішені з експортованими бактерійними білками, які зв'язуються з клітинною мембраною або стінкою клітини (Chen і Georgiou, 2002 р., Biotechnol Bioeng, 79: 496503). Даний винахід також представляє нуклеїнові кислоти, які кодують композиції, що складають предмет даного винаходу, у вигляді виділених полінуклеотидів або частин векторів експресії, включаючи вектори, сумісні з експресією в прокаріотичних, еукаріотичних абонитчатих фагах, виділенням і/або дисплеєм композицій або їх спрямованих мутагенів. Використання Fc-вмісних молекул Композиції (антитіло, Fc-злиття, Fc-фрагменти), отримані з використанням будь-яких описаних вище способів, можна застосовувати для діагностики, лікування, виявлення або модуляції хвороб людини або конкретних патологій в клітинах, тканинах, органах, рідинах або по суті хазяїні. Як описано в цьому документі, модифікація частини Fc антитіла, Fc-злитого білка або Fc-фрагмента для зниження або виключення протеолітичного розкладання при збереженні вимірного зв'язування з Fc-гамма-рецептором або вказаних ефекторних функцій можуть комбінуватися із зв'язувальним доменом, таким як паратоп антитіла або лігандзв'язувальний домен, який зберігає вихідну специфічність до мішені і біологічну активність. Приклади зв'язувальних доменів являють собою паратоп антитіла, один або більше CDR антитіла або один або більше варіабельних доменів антитіла; ферменту; гормону; рецептора; позаклітинний домен мембранного рецептора; цитокіну; поверхневого антигену імунної клітини і адгезійної молекули. Отримані конструкції додають антитілам і Fc-конструкціям прекрасний спектр активності, біофізичних властивостей, стабільність і здатність зберігатися в організмі хазяїна. Відкриті заявниками послідовності Fc з унікальними комбінаціями стійкості до фізіологічно важливих протеаз і здатності або нездатності зв'язуватися з одним або більше Fc-гаммарецепторами і/або здатності здійснювати лізис клітин за допомогою активації ефекторних клітин або комплементу, забезпечують можливість цілеспрямовано орієнтувати зв'язувальну молекулу для максимальної ефективності при конкретних показаннях. Наприклад, здатність вибирати як мішені аберантну клітину-хазяїна, таку як задіяна в неоплазії або інших небажаних процесах проліферації, таких як неадекватний ангіогенез, неадекватний фіброз, в найбільшому ступені буде підходити для молекули, яка містить еукаріотичний протеазостійкий Fc, що складає предмет даного винаходу, здатний до ADCC і ADCP. На відміну від цього бактерійні клітини 11 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 легко знищуються комплементопосередкованими механізмами. Отже, бактерійні інфекції можна лікувати придатними Fc-конструкціями, що складають предмет даного винаходу, які стійкі до бактерійних протеаз і мають здатність викликати CDC. Заявники визначили способи вибору Fc, що мають відповідну комбінацію властивостей, і показали діючі приклади цілеспрямовано модифікованих специфічних Fc-вмісних молекул. Молекули, стійкі до еукаріотичних протеаз і здатні виявляти одну або більше функцій, зв'язаних з ADCC, ADCP і CDC, включають домен Fc, що містить послідовність людського IgG1 в шарнірній ділянці і ділянці CH2, від приблизно EU залишку 214 до приблизно залишку 330, де щонайменше залишки 233-237 замінені PVA/(делеція G236), і додатково, який містить одне або більше заміщення в домені CH2, причому молекула здатна виявляти одну або більше функцій, зв'язаних з ADCC, ADCP і CDC, і включає конструкції 5, 7, 8, 9, 10, 11, 12, 14, 15, 16 і 17. У конкретному варіанті здійснення такі молекули включають заміщення, вибрані із I332E в комбінації з іншими заміщеннями, такими як S239D/I332E (5, 14), S239D/H268F/I332E (не проводили), H268F/S324T/I332E (8), S239D/H268F/S324T/I332E (9), S267E/H268F/S324T/I332E (10), G237X/S239D/I332E, де X являє собою А або S (12); K326A/I332E/E333A (15) і S267E/I332E (17). Молекули, стійкі до прокаріотичних протеаз і здатні виявляти CDC, включають дані молекули, які містять домен Fc, що має послідовність людського IgG1 в шарнірній ділянці і ділянці CH2, від приблизно EU залишку 214 до приблизно залишку 330, де щонайменше залишки 233-237 замінені PVA/ (делеція G236), і додатково, який містить одне або більше заміщень в домені CH2, вибраних із K326A/E333A (11), S267E/H268F/S324T/I332E (10), K326A/I332E/E333A (15), S239D/K326A/E333A (16) і S267E/I332E (17). Молекули, стійкі до еукаріотичних протеаз, але не сприяючі лізису клітин-мішеней, можуть також ефективно використовуватися для лікування захворювання або станів, при яких метою є не усунення клітин-мішеней, а їх модуляція, як наприклад, за рахунок використання антитіла або іншої Fc-конструкції, як описано в цьому документі, яка відрізняється протеазостійкістю, але демонструє низький рівень ADCC, ADCP або CDC в порівнянні з IgG1 дикого типу. Молекули, які містять неприродний домен Fc не дикого типу, які є протеазостійкими, включають дані молекули, що містять домен Fc з послідовністю людського IgG1 в шарнірній ділянці і ділянці CH2 від приблизно EU залишків 214 до приблизно залишку 330, де щонайменше залишки 233237 замінені PVA/ (делеція G236). Таким чином, на основі представлених в цьому документі концепцій і прикладів, Fc-вмісні конструкції, які отримують в даному винаході, що демонструють підвищену стійкість до протеаз, які присутні в організмі ссавців, і необов'язково, які мають здатність вибирати як мішень клітину і антиген на клітині, забезпечують більш ефективні терапевтичні молекули в порівнянні з терапевтичною молекулою, яка не є протеазостійкою. Вступ Оскільки протеолітичні ферменти локалізуються у відповідності із швидкістю утворення або накопичення, наприклад, пепсин в травному тракті, або матриксні металопротеїнази (MMP) в ділянках, де відбувається перебудова тканини або ріст злоякісних утворень, композиції, що складають предмет даного винаходу, зокрема, підходять для застосувань, коли відомо, що та або інша область організму містить протеази або відрізняється аномально високим вмістом протеаз. Даний винахід представляє стабільні склади композиції протеазостійкого IgG, такі як антитіла, які переважно являють собою водний фосфатно-буферний розчин або змішаний сольовий розчин, а також розчини і склади з консервантами, склади з консервантами для багаторазового використання, придатні в фармацевтичних або ветеринарних цілях, що містять щонайменше одне протеазостійке антитіло в фармацевтично прийнятному складі. Придатні середовища, що несуть, і їх склад з можливістю введення інших білків людини, наприклад, сироватковий альбумін людини, описані, наприклад, в роботі Remington: The Science and Practice of Pharmacy, 21-е вид., під ред. Troy D. B., Lipincott Williams and Wilkins, Philadelphia, PA 2006 р., частина 5, Pharmaceutical Manufacturing, стор. 691-1092, див. особливо стор. 958-989. Протеазостійка композиція IgG з ефекторними функціями в стабільному складі або в складі, який містить консерванти, як описано в цьому документі або відомо фахівцям в даній галузі, може вводитися пацієнту відповідно з даним винаходом за допомогою ряду способів доставки, включаючи введення внутрішньовенно (в/в); внутрішньом'язово (в/м); підшкірно (п/ш); трансдермально; через легені; трансмукозально; з використанням складу в імплантаті, осмотичному насосі, картриджі, мікронасосі або з використанням інших засобів, добре відомих фахівцеві в даній галузі. 12 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 Можливе направлене введення в ту або іншу частину тіла або порожнину, таке як внутрішньосуглобове введення, всередину бронха, внутрішньоочеревинне, всередину капсули, хряща, порожнини, інтрацеліальне, всередину мозочка, шлуночка мозку, всередину товстої кишки, шийки, шлунка, печінки, міокарда, всередину кістки, таза, перикарда, порожнини живота, плеври, простати, легень, всередину прямої кишки, всередину нирки, сітчатки, хребта, інтрасуглобно, грудної клітки, всередину матки, сечового міхура, внутрішньоосередково, вагінально, ректально, букально, під язик, інтраназально або трансдермально. Захворювання або патології, які можуть піддаватися лікуванню за допомогою композиції, наведеної в даному винаході, включають без обмежень захворювання, при яких несприятливою є небажана проліферація, активація або міграція клітин, така як злоякісні утворення, гіперактивні або незбалансовані імунні відповіді, утворення фіброзної тканини або інфекція, оскільки композиції забезпечують активацію цитотоксичного або цитолітичного механізмів імунної системи хазяїна за допомогою механізмів, регульованих FcγR. Такі захворювання включають злоякісні утворення: лейкемію, гостру лейкемію, гострий лімфобластний лейкоз (ALL), ALL В-клітин, Т-клітин або FAB, гостру мієлоїдну лейкемію (AML), хронічну мієлоцитарну лейкемію (CML), хронічну лімфоцитарну лейкемію (CLL), лейкоз ворсистих клітин, мієлодиспластичний синдром (MDS), лімфопроліферативне захворювання, шкірну Т-клітинну лімфому, хворобу Ходжкіна, хворобу Кастлемана, гліому, гліобластому, астрацитому, злоякісну лімфому, неходжкінську лімфому, лімфому Беркітта, множинну мієлому, саркому Капоши, колоректальну карциному, панкреатичну карциному, гіпернефроїдний рак, рак молочної залози, протокову карциному, ліпому, рак носоглотки, рак простати, рак яєчок, рак яєчників, ретинобластому, злоякісний гістіоцитоз, гіперкальціємію злоякісного утворення, плазмацитоми, хондросаркоми, саркоми, трабекулярний рак, гепатоклітинну карциному, гепатому, рак базальних клітин, аденокарциноми, плоскоклітинний рак, саркоми (такі як саркома Юінга, Капоши, дитяча м'яких тканин, доросла), меланоми, метастатичні меланоми, гемангіому, метастатичне захворювання, остеосаркому, рабдоміосаркому, тімому і тимічну карциному, пов'язану з раком резорбцію кістки, рак тіла матки, вагінальний рак, рак матки, пухлини Вільмса, пов'язану з раком біль в кістках і т. п. Цільові молекули, здатні зв'язувати антигени на злоякісних лімфоцитах, включають антигени В-клітин, такі як CD19, CD20 і CD22. Солідні пухлини, які формуються на епідермальній тканині, часто демонструють і стимулюються зв'язуванням лігандів з рецепторами епідермального фактора росту, відомими як ErbB1, ErbB2, ErbB3 і іншими рецепторами, здатними передавати сигнал або викликати проліферативну відповідь або антиапоптозну відповідь, які приводять до неконтрольованого росту пухлини. Інші поширені антигени на солідних пухлинах являють собою тканинний фактор або RON. При комбінації з відповідним зв'язувальним доменом з мішенню композиції також застосовуються для лікування інфекційних захворювань, що викликаються бактерійними (такими як Streptococcus, Staphylococcus і Е. coli.), вірусними (такими як грип, СНІД, РСВ, ТОРС і вірус лихоманки Західного Нілу), грибковими (такими як аспергільоз, кокцидіоїдомікоз, криптококоз або кандидоз) або протозойними інфекціями (такими як трипаносомоз, токсоплазмоз, лямблія або малярія). Композиції застосовуються для лікування загальних імунологічних і аутоімунних порушень, включаючи без обмежень ревматичні захворювання, псоріаз і склеродерму. Композиції застосовуються для лікування захворювань, пов'язаних з неадекватним ангіогенезом. Ангіогенез являє собою процес формування нових капілярних кровоносних судин, і він є наслідком активованої проліферації клітин ендотелію. Неоваскуляризація жорстко регулюється і відбувається тільки під час ембріонального розвитку, перебудови тканини, загоєння ран і періодичного циклу розвитку жовтого тіла (Folkman і Cotran, Relation of vascular proliferation to tumor growth, Int. Rev. Exp. Pathol.'16, 207-248 (1976 р.)). Проліферація клітин ендотелію звичайно відбувається набагато повільніше, ніж у інших видів клітин організму. Однак, якщо швидкість проліферації даних клітин виходить з-під контролю, результатом може стати патологічний ангіогенез. Патологічний ангіогенез виявляється в багатьох захворюваннях. Наприклад, серцевосудинних захворюваннях, таких як ангіома, ангіофіброма, судинна деформація, атеросклероз, синехія і едемічний склероз; а також офтальмологічних захворюваннях, таких як неоваскуляризація після імплантації рогівки, неоваскулярна глаукома, діабетична ретинопатія, ангіогенне захворювання рогівки, дегенерація жовтої плями, птеригіум, дегенерація сітчатки, ретролентальна фіброплазія і гранулярний кон'юнктивіт, пов'язані з ангіогенезом. Хронічні запальні захворювання, такі як артрит; дерматологічні захворювання, такі як псоріаз, телеангіектазія, піогенна гранулема, себорейний дерматит, варикозні виразки, 13 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 вугровий висип, рожеві вугри (розацеа або еритематоз), папіломи (бородавки), екзема, гемангіоми, лімфангіогенез, також залежать від ангіогенезу. Діабетична ретинопатія може мати дві форми, непроліферативну або проліферативну. Проліферативна ретинопатія характеризується аномальним утворенням нових судин (неоваскуляризація), які ростуть на поверхні скловидного тіла або проходять всередину порожнини скловидного тіла. Дегенерація жовтої плями також може мати дві форми, суху і вологу. При ексудативній дегенерації жовтої плями (волога форма), яка зустрічається набагато рідше, відмічається утворення субретинальної сітки хороїдальної неоваскуляризації, часто пов'язаної з інтраретинальною кровотечею, субретинальної рідини, епітеліальне відшаровування пігменту і гіперпігментація. Неоваскулярна глаукома зустрічається у пацієнтів, страждаючих діабетом або оклюзією центральної ретинальної вени, або при пов'язаних з увеїтом запальних виділеннях, які видавлюють райдужну оболонку у верхній кут (Гл. 99. The Merck Manual, 17-е вид., 1999 р.). Ревматоїдний артрит, запальне захворювання, також приводить до неадекватного ангіогенезу. Ріст васкулярних клітин ендотелію в синовіальній порожнині активується запальними цитокінами і приводить до пошкодження хряща і заміни на панус у суглобі (Koch A. K., Polverini P. J. і Leibovich S. J., Arth; 15 Rhenium, 29, 471-479 (1986 р.); Stupack D. G., Storgard C. M. і Cheresh D. A., Braz. J. Med. Biol. Res., 32, 578-581 (1999 р.); Koch A. K., Arthritis Rheum, 41, 951-962 (1998 р.)). Композиції можуть використовуватися для лікування псоріазу, який викликається неконтрольованою проліферацією клітин шкіри. Клітини, які швидко ростуть, потребують достатнього притоку крові, і при псоріазі індукується аномальний ангіогенез (Folkman J., J. Invest. Dermatol. 59, 40-48 (1972 р.)). У процесах і явищах, які ведуть до ангіогенезу, задіяний ряд факторів: молекули клітинної адгезії, інтегріни, фактор росту ендотелію судин (VEGF), TNFalpha, bFGF і цитокіни, включаючи IL-6 і IL-12. Наприклад, було показано, що близькоспоріднені, але різні інтегріни alphaVbeta3 і aVb5 опосередковують різні шляхи ангіогенного процесу. Антитіло, що виробляється проти alphaVbeta3, блокувало ангіогенез, який індукується основним фактором росту фібробластів (bFGF), тоді як антитіло, специфічне до aVb5, інгібувало ангіогенез, який індукований фактором росту ендотелію судин (VEGF) (Eliceiri et al., J. Clin. Invest.103: 1227-1230 (1999 р.); Friedlander et al., Science 270: 1500-1502 (1995 р.)). Отже, даний винахід включає застосування цільових зв'язувальних доменів, орієнтованих на дані мішені в композиціях, що складають предмет даного винаходу, для лікування захворювань, при яких показано інгібування ангіогенезу. Заявники опублікованої заявки на міжнародний патент WO2009023457A1, що одночасно розглядається, яка включена в дану заявку шляхом посилання, розкривають стратегію відновлення ефекторної функції розщеплених IgG з використанням моноклональних антитіл, специфічних до епітопу, який перешкоджає розщепленню в сайті шарніра. Включення протеазостійкого, який має компетентну ефекторну функцію Fc, відповідно з даним винаходом з антишарнірним доменом для утворення двовалентного антитіла дозволить отримати терапевтичне mAb, здатне до відновлення ефекторної функції Fc у розщеплених IgG, забезпечуючи при цьому терапевтичну стійкість до пригнічення за рахунок протеолітичного розщеплення. Відповідно, даний винахід передбачає включення протеазостійкої константної ділянки Fc, яка складає предмет даного винаходу, з протишарнірною варіабельною ділянкою mAb, описаною вище. Хоч даний винахід описаний вище в загальних рисах, варіанти здійснення даного винаходу будуть додатково описані в наступних прикладах, які нічим не обмежують об'єм формули винаходу. ПРИКЛАД 1. КОНСТРУЮВАННЯ І ТЕСТУВАННЯ МУТАНТІВ Fc З використанням стандартної рекомбінантної технології був отриманий набір конструкцій (показаних на фігурі 2, які об'єднують шарнірну ділянку, представлену в таблиці 1, з поновлюючими активність мутаціями в ділянці CH2, вибраними із таблиці 2). Позначення 2hc означає, що константний домен IgG1, відповідний по нумерації Kabat EU антитіла (нумерація EU) від 214 до 236 (SEQ ID NO: 3), був замінений відповідною послідовністю IgG2 (SEQ ID NO: 4). Позначення 2h означає, що залишки IgG1 E233-L234-L235-G236 замінені відповідними залишками IgG2 P233-V234-A235 з делецією G235. 14 UA 113725 C2 Таблиця 4 Конструкція 1 2 3 4 5 6 7 8 9 10 11 12 13 14 5 10 15 20 25 30 35 Позначення/ конструкція IgG1 wt IgG2 wt 2 hc (всі шарнірні залишки від IgG2) 2h 2h S239D/I332E 2h E333A/K334A 2h F243L/R292P/Y300L 2h H268F/S324T/I332E 2h S239D/H268F/S324T/I332E 2h S267E/H268F/S324T/I332E 2h K326A/E333A 2h G237X/S239D/I332E, де X являє собою А, D, Р, Q або S 2hc S239D/I332E IgG2 S239D/I332E Каркас Послідовність EU ізотипу IgG 214-447 hIgG1 SEQ ID NO: 1 hIgG2 SEQ ID NO: 2 CD142 CD20 X X X X hIgG1 SEQ ID NO: 6 X hIgG1 hIgG1 hIgG1 hIgG1 hIgG1 SEQ ID NO: 7 SEQ ID NO: 8 SEQ ID NO: 9 SEQ ID NO: 10 SEQ ID NO: 11 X X X X X X X hIgG1 SEQ ID NO: 12 X X hIgG1 SEQ ID NO: 13 X X hIgG1 SEQ ID NO: 14 X X hIgG1 SEQ ID NO: 15 X hIgG1 hIgG2 SEQ ID NO: 16 SEQ ID NO: 17 X X X X Варіабельні ділянки одного набору конструкцій антитіл зв'язуються з CD142 (тканинний фактор), що дозволяє тестувати Fc-залежне знищення клітин антитілам в клітинних аналізах з використанням MDA-MB-231 (ATCC, HTB-26™), експресуючого тканинний фактор (Brezski et al., Proc Natl Acad Sci USA 106:17864-17869, 2009 р.). Була підготовлена додаткова панель з варіабельними ділянками, які зв'язуються з CD20, що дозволяло тестувати активність CDC за допомогою клітин WIL2-S, які демонструють CD20 (ATCC, CRL-8885) (Brezski et al., Proc Natl Acad Sci USA 106:17864-17869, 2009 р.). Всі антитіла проміжно експресували в клітинах 293T за допомогою стандартних способів і процедур клонування. Перед початком експериментального аналізу mAb очищали на колонках білка А до чистоти більше 95 %. Протеазне розщеплення конструкцій mAb дикого типу Протеазне розщеплення обчищених IgG проводили при pH 7,5 в фізіологічному розчині з фосфатно-буферним розчином (ФСБ) або у випадку MMP (MMP-3, MMP-7, MMP-12 і MMP-13 отримані від Enzo Life Sciences) в фізіологічному розчині з трис-буфером при 37 °C. IdeS отримували від Genovis, а GluV8 отримували від Pierce. CaCl 2 включали в реакції MMP в концентрації 5 мM для всіх тестованих MMP. Концентрації антитіла становили 0,5 мг/мл, і реакції ініціювали додаванням ферменту до IgG у співвідношенні приблизно 1-2 % (в./в.), якщо не вказане інше. Розщеплення IgG оцінювали за допомогою аналізу електрофорограм після мікрокапілярного електрофорезу Agilent Biosizing (Agilent Technologies). Всі реакції розщеплення проводили в двох повторюваннях. Аналізи конкурентного зв'язування AlphaScreen® Дослідження конкурентного зв'язування проводили з половиною об'єму ямки в 96-ямкових непрозорих планшетах (Corning) в аналітичному буферному розчині (ФСБ, 0,05 % BSA, 0,01 % Tween-20) при pH 7,4. Всі конкурентні дослідження проводили для біотинілованого IgG1 (1 IgG: 2 біотин з використанням EZ Link™ NHS-LC-біотину, Pierce) при фіксованій концентрації і конкуруючих конструкціях дикого типу і протеазостійких конструкціях при послідовних 3-кратних розбавленнях. Концентрації FcγR становили 0,2 мкг/мл в кінцевій концентрації в аналізах. Біотинілований IgG1 (кінцева концентрація 0,2 мкг/мл) і антитіла дикого типу і протеазостійкі (10 мкл) послідовно додавали до кожного ряду 96-ямкового планшета в двох повторюваннях. Потім додавали задані FcγR з подальшим додаванням по 10 мкл розведених 1/50 гранул акцептора хелату нікелю (Ni) і гранул донора стрептавідину (SA). Непрозорий планшет покривали алюмінієвою фольгою, щоб виключити потрапляння світла під час струшування на шейкері Orbital протягом 30 хвилин. Потім фольгу знімали і реєстрували флуоресценцію на зчитувачі планшетів ENVISION™ (PerkinElmer), забезпеченому відповідним набором фільтрів AlphaScreen® для спектрів збудження/поглинання. Отримані дані обробляли програмою 15 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 GraphPad PRISM™ і нормалізували по максимальному сигналу, а також будували криві конкуренції за допомогою програми нелінійної регресії для підгонки кривих. Результати Початковий інтерес був пов'язаний із спробами визначити чутливість конструкцій mAb до ряду фізіологічно важливих протеаз, для яких раніше було показано розщеплення IgG1 в нижній шарнірній ділянці; MMP-3, MMP-7, MMP-12, MMP-13, GluV8 і IdeS. Конструкції 1-5, 7, 9-11, 12 (G237A), 13-14 тестували як CD142-зв'язувальні антитіла. Протягом 24 годин протеази демонстрували різний ступінь розщеплення IgG1. Тоді як MMP-3, MMP-12, IdeS руйнували весь інтактний IgG1 (конструкція 1) протягом 24 годин, MMP-7 розщеплювала приблизно 30 %, MMP13 - приблизно 40 %, а GluV8 - приблизно 60 %. Конструкція 4 (2h) і дані конструкції з 2h модифікацією нижнього шарніра в більш-менш однаковому ступені виявляли стійкість відносно всіх MMP. Конструкція 4 була стійкою до GluV8, але не до IdeS. Конструкція 2 була також більш стійкою до розщеплення GluV8. Було показано, що IdeS розщеплює всі ізотипи людського IgG (von Pawel-Rammingen et al., EMBO 21:1607-1615, 2002 р.). Дані дослідження залежності від часу (фігура 3А) показують, що IdeS швидко трансформувала IgG1 (1) і IgG2 (2) у фрагмент F(ab')2 (протягом 5 хвилин інкубування), тоді як проба IgG1 2h (4) протягом 120 хвилин утворювала інтермедіат з одиничним розривом. Із всіх тестованих конструкцій 2h S239D/I332E (5) була найбільш протеазостійкою до IdeS (фігура 3B), причому інтактний IgG виявлявся навіть після 24 годин інкубації (в інших конструкціях антитіл інтактний IgG не виявлявся вже через 5 хвилин). Мутація S239D в (5), яка знаходиться поруч з місцем розщеплення IdeS між G236 і G237, може сприяти додатковій протеазостійкості в порівнянні з IgG1 2h (4). Панель із 12 анти-CD142 конструкцій антитіл (1-5, 7, 9-13 і 14) разом з її аналогами IgG1 і IgG2 дикого типу тестували на предмет чутливості до протеолізу IdeS, GluV8, MMP-3 і MMP-12. Дані для інтактного IgG, що залишається через 24 години після інкубації, розраховували на основі електрофорограм і наведені на фігурах 4A-D. Дані показали, що IgG1 wt (1), IgG2 wt (2), 2hc (3), IgG1 2h (4), 2hc S239D/I332E (13), IgG2 S239D/I332E (14) і 2h K326A/E333A (11) були піддані протеолізу IdeS після 24 годин інкубації. На відміну від цього конструкції 2h S239D/I332E (5), 2h G237A/S239D/I332E (12), 2h F243L/R292P/Y300L (7), 2h S239D/H268F/S324T/I332E (9) і 2h S267E/H268F/S324T/I332E (10) були стійкі до протеолізу за допомогою IdeS. Несподівано було виявлено, що конструкції з великим числом заміщень шарнірної ділянки IgG2 (які містять SEQ ID NO: 4) не були стійкі до протеолізу IdeS. Конструкція IgG2 S239D/I332E містила менше 40 % інтактного IgG після 24 годин розщеплення в присутності IdeS. Розщеплення протеазою GluV8 із Staph aureus показало, що конструкції IgG2 S239D/I332E (14), 2h S239D/I332E (5), 2h G237A/S239D/I332E (12) і 2h F243L/R292P/Y300L (7) забезпечували збереження інтактного IgG, який залишається через 24 години після розщеплення, в діапазоні 40-60 %, аналогічно рівню розщеплення, який спостерігався для IgG1 wt (1). Конструкції IgG2 wt (2), 2hc (3), 2hc S239D/I332E (14), IgG1 2h (4), IgG1 2h K326A/E333A (11), 2h S239D/H268F/S324T/I332E (9) і 2h S267E/H268F/S324T/I332E (10) продемонстрували підвищену стійкість до протеолізу в присутності GluV8 (у всіх випадках залишалося більше 75 % інтактного IgG) в порівнянні з IgG1 wt. MMP-3 і MMP-12 являють собою два типи зв'язаних з раком протеаз. Менше 5 % інтактного IgG1 wt було зареєстровано після розщеплення MMP-3 і MMP-12. На відміну від цього людський IgG2 wt і всі тестовані конструкції виявляли підвищену протеазостійкість у відношенні як MMP-3, так і MMP-12, що підтверджувалося більш ніж 60 % інтактного IgG, який залишається після 24 годин розщеплення. Результати зв'язування Fcγ-рецептора Оцінювали здатність деяких Fc-конструкцій конкурувати із зв'язування з рецепторами Fcγсімейства з IgG1 wt. Здатність конструкцій знижувати максимальний сигнал біотинілованого IgG1 показана на фігурах 5A-G. Початковий скринінг включав конструкції 1-2, 4-6. У вихідній групі тестованих конструкцій IgG2 (2), IgG1 2h (4) і 2h E333A/K334A (6) не демонстрували виявлене зв'язування до високоафінних FcγRI, тоді як дикий тип IgG1 демонстрував стійке зв'язування. 2h S239D/I332E (5) демонстрував виявлене, але знижене зв'язування з FcγRI в порівнянні з IgG1. Для IgG1 2h (4) і 2h E333A/K334A (6) не демонструвалося виявлене зв'язування з FcγRIIa, тоді як конструкція 2h S239D/I332E (5) демонструвала порівнянне зв'язування з IgG1 wt (фігура 5B). Три конструкції: IgG2 (2), IgG1 2h (4) і 2h E333A/K334A (6) не демонстрували виявлене зв'язування з FcγRIIb, тоді як IgG1 (1) і 2h S239D/I332E (5) демонстрували порівнянне зв'язування (фігура 5D). IgG2 (2), IgG1 2h (4) і 2h E333A/K334A (6) демонстрували порівнянне, але знижене зв'язування з FcγRIIIa в порівнянні з IgG1. Конструкція 16 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 2h S239D/I332E (5) виявляла найбільший рівень зв'язування з FcγRIIIa, навіть більше, ніж IgG1 wt (фігура 5F). Крім того, конструкція 2h S239D/H268F/S324T/I332E (9) виявляла порівнянне зв'язування з IgG1, тоді як конструкція 2h S267E/H268F/S324T/I332E (10) відрізнялася підвищеним зв'язуванням з FcγRIIa в порівнянні з IgG1 wt (фігура 5C). Обидві конструкції 2h S239D/H268F/S324T/I332E (9) і 2h S267E/H268F/S324T/I332E (10) виявляли підвищене зв'язування з FcγRIIb в порівнянні з IgG1 wt (фігура 5E). Конструкція 2h K326A/E333A (11) демонструвала мінімальне виявлене зв'язування як з FcγRIIa (фігура 5C), так і з FcγRIIb (фігура 5E). Конструкція 2h S267E/H268F/S324T/I332E (10) демонструвала декілька знижене зв'язування з FcγRIIIa в порівнянні з IgG1 wt, тоді як конструкція 2h S239D/H268F/S324T/I332E (9) демонструвала підвищене зв'язування з FcγRIIIa (фігура 5G). 2h K326A/E333A (11) виявляла слабе зв'язування з FcγRIIIa в порівнянні з IgG1 wt (фігура 5G). Зведена інформація Дані результати показують, що протеазостійкі конструкції, які містять 2h S239D/I332E (5), 2h S239D/H268F/S324T/I332E (9) і 2h S267E/H268F/S324T/I332E (10), були здатні в різному ступені зв'язувати FcγRIIa, IIb і IIIa, і кожна з трьох даних конструкцій виявляє підвищене зв'язування відносно однієї мутації 2h (4). Мутація залишків в нижньому шарнірі IgG1 в комбінації з іншими мутаціями CH2, які посилюють афінність зв'язування FcγRIIa в порівнянні з IgG1 wt, як і для конструкції 2h S267E/H268F/S324T/I332E (10), підвищена афінність 2h S239D/H268F/S324T/I332E (9) і 2h S267E/H268F/S324T/I332E (10) до FcγRIIb, підвищена афінність зв'язування FcγRIIIa в порівнянні з IgG1 wt, як і для конструкцій 2h S239D/I332E (5) і 2h S239D/H268F/S324T/I332E (9) - були несподіваними результатами. ПРИКЛАД 2. АНТИТІЛОЗАЛЕЖНИЙ КЛІТИННИЙ ФАГОЦИТОЗ (ADCP) Для тестування здатності протеазостійких mAb опосередковувати Fc-залежне знищення клітин in vitro проводили аналізи ADCP. У даному аналізі фагоцитарні клітини впливають на клітину-мішень, яка несе антиген, за допомогою зв'язування антитіла, і вимірюється знищення клітин-мішеней. Процедура PBMC були виділені із зразків від здорових людських донорів за допомогою центрифугування в градієнті Ficoll. Моноцити CD14pos виділяли із PBMC методом негативної елімінації за допомогою набору CD14 Isolation, при якому не відбувається елімінація моноцитів 6 2 CD16pos (Stem Cell Technologies). Моноцити висівали по 0,1×10 клітин/см в середовищі XVIVO-10 (Lonza), що містить 10 % FBS і 20 нг/мл GM-CSF (R&D Systems), і витримували протягом 7 діб. У останні 24 години диференціювання додавали 100 нг/мл IFNγ (R&D Systems). Клітини-мішені для аналізу ADCP являли собою GFP-експресуючі клітини MDA-MB-231. Виділені макрофаги інкубували з GFP-експресуючими MDA-MB-231 у співвідношенні 4:1 протягом 4 годин в присутності або за відсутності конструкцій mAb дикого типу і протеазостійких конструкцій в 96-ямковому планшеті з U-подібним дном ямок. Після інкубування клітини видаляли з 96-ямкових планшетів з використанням Accutase (Sigma). Макрофаги ідентифікували анти-CD11b і анти-CD14 антитілами (обидва засоби від BD Biosciences), пов'язаними з AlexaFluor 647 (Invitrogen), після чого клітини відділяли на FACS Calibur (BD Biosciences). Дані аналізували за допомогою програмного забезпечення FloJo Software (Tree Star). Процент фагоцитозу визначали за наступним рівнянням: (клітини GFPpos, CD11bpos, CD14pos)/(клітини GFPpos, CD11bpos, CD14pos плюс тільки клітини GFPpos)×100 %. Виділені моноцити диференціювали in vitro з використанням GM-CSF і IFNγ, як описано в цьому документі. Як було показано в інших роботах, диференційовані макрофаги експресували весь набір FcγR(FcγRI, FcγRIIa, FcγRIIb і FcγRIIIa) (дані не показані). Результати Дані, представлені на фігурі 6A, показують, що IgG1 wt (1) і 2h S239D/I332E (5) забезпечували досягнення самих високих рівнів ADCP. Здатність до ADCP для IgG2 wt (2), IgG1 2h (4) і 2h E333A/K334A (6) була низькою, але виявленою. Ці результати показують, що протеазостійка конструкція 2h S239D/I332E (5) була здатна до фагоцитозу клітин пухлини на рівні, порівнянному з IgG1 wt. У окремому експерименті додаткову панель конструкцій CH2, що містять шарнірну ділянку IgG1 2h, тестували на здатність до ADCP (фігура 6B). У даній групі конструкції 2h S239D/I332E (5), 2h F243L/R292P/Y300L (7) і 2h S239D/H268F/S324T/I332E (9) виявляли рівень ADCP, аналогічний IgG1 wt (1), тоді як конструкції 2h S267E/H268F/S324T/I332E (10), 2h H268F/S324T/I332E (8) і 2h G237A/S239D/I332E (12) демонстрували декілька знижений максимальний фагоцитоз в порівнянні з IgG1 wt. Конструкції 2h K326A/E333A (11) виявляли низький, але виявлений рівень ADCP, аналогічний IgG2 wt (2) і IgG1 2h (4). Нарешті, конструкції, 17 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 60 що містять повний шарнір IgG2, також тестували на ADCP. Конструкція IgG2 S239D/I332E (14) виявляла рівень ADCP, аналогічний IgG1 wt, тоді як конструкції 2hc (3) і 2hc S239D/I332E (13) виявляли низький, але виявлений рівень ADCP. ПРИКЛАД 3. АНТИТІЛОЗАЛЕЖНА КЛІТИННА ЦИТОТОКСИЧНІСТЬ (ADCC) У цьому аналізі мононуклеарні клітини впливають на клітину-мішень, яка несе антиген, і вимірюється видалення клітин-мішеней. Процедура Аналізи ADCC проводили, як описано в попередній роботі (Scallon et al., Mol Immunol 44: 1524-1534, 2007 р.). Іншими словами, PBMC виділяли з людської крові з використанням градієнта Ficoll і використовували як ефекторні клітини для аналізу ADCC. Клітини карциноми молочної залози людини MDA-MB-231 (ATCC HTB-26) використовували як клітини-мішені у співвідношенні 1 клітина-мішень до 50 ефекторних клітин. Клітини-мішені заздалегідь мітили BATDA (PerkinElmer) протягом 20 хвилин при 37 °C, двічі промивали і повторно розчиняли в модифікованому за способом Дульбекко середовищі Ігла, 10 % термоінактивованої FBS, 2 мM 4 L-глутаміну (всі виробництва Invitrogen). Клітини-мішені (1×10 клітин) і ефекторні клітини 6 (0,5×10 клітин) об'єднували і додавали 100 мкл клітин до ямок 96-ямкового планшета з Uподібним дном. Додавали додаткові 100 мкл в присутності або за відсутності конструкцій mAb дикого типу і протеазостійких конструкцій. Всі проби були представлені в двох повторюваннях. Планшети центрифугували при 200g протягом 3 хвилин, інкубували при 37 °C протягом 2 годин, а потім знову центрифугували при 200g протягом 3 хвилин. Відбирали по 20 мкл супернатанту з кожної ямки і вимірювали лізис клітин за допомогою додавання 200 мкл реагенту DELPHIA на основі європію (PerkinElmer). Інтенсивність флуоресценції вимірювали за допомогою Envision 2101 Multilabel Reader (PerkinElmer). Дані нормалізували за максимальною цитотоксичністю з 0,67 % Triton X-100 (Sigma Aldrich) і мінімальним контролем, які визначали по вивільненню BATDA із клітин-мішеней за відсутність будь-якого антитіла. Дані апроксимували сигмоїдною моделлю ефекту дози за допомогою GraphPad Prism, версія 5. Результати Дані відображалися на графіку таким чином, щоб рівень лізису клітин був поданий на осі Y як функція концентрації антитіла. Дані, представлені на фігурі 7А, показують, що конструкція 2h S239D/I332E (5) демонструвала самий високий рівень ADCC, що відповідало приблизно восьмиразовому (як свідчить зсув в EC50, яка спостерігається) поліпшенню в порівнянні з IgG1 wt у даному аналізі. У іншому експерименті зіставляли здатність до ADCC розширеної панелі конструкцій. На фігурі 7В представлені криві, побудовані на основі даних. Три конструкції (2h S239D/I332E (5), 2h F243L/R292P/Y300L (7) і 2h S239D/H268F/S324T/I332E (9)) виявляли підвищену здатність до ADCC в порівнянні з IgG1 wt. Конструкції 2h G237A/S239D/I332E (12) і 2h H268F/S324T/I332E (8) демонстрували дещо підвищений рівень ADCC в порівнянні з IgG1 wt, тоді як конструкції 2h K326A/E333A (11) і 2h S267E/H268F/S324T/I332E (10) демонстрували виявлений, але знижений рівень ADCC в порівнянні з IgG1 wt. На фігурі 7C представлені результати ADCC для панелі конструкцій, які містили повну шарнірну ділянку IgG2. Конструкція IgG2 S239D/I332E (14) демонструвала більш низький рівень EC50 в порівнянні з IgG1 wt, але також і більш низький рівень максимального лізису. Конструкції 2hc (3) і 2hc S239D/I332E (13) демонстрували виявлений рівень ADCC вище IgG2 wt, але нижче IgG1 wt. У сукупності ці результати показують, що мутація найважливіших залишків в нижньому шарнірі для відновлення ADCC і FcγRзв'язування може компенсуватися рядом мутацій CH2. Однак не всі проаналізовані мутації CH2 в головному ланцюгу IgG1 2h (3) були здатні відновити/підвищити рівень ADCC в порівнянні з IgG1 wt. Ці результати погоджуються з аналізом зв'язування FcγRIIIa (фігури 5F-G), який демонструє підвищену афінність 2h S239D/I332E (5) і 2h S239D/H268F/S324T/I332E (9), оскільки експресуючі FcγRIIIa NK-клітини вважаються компетентними ефекторними клітинами в ADCC. ПРИКЛАД 4. КОМПЛЕМЕНТОЗАЛЕЖНА ЦИТОТОКСИЧНІСТЬ (CDC) У даному аналізі компоненти комплементу впливають на клітини-мішені, які несуть антиген, і оцінюється руйнування клітин-мішеней. Процедура Аналізи CDC проводили, як описано в попередній роботі (Brezski et al. J Immunol. 181(5): 3183-3192, 2008 р.). Клітини WIL2-S використовували як клітини-мішені при аналізах CDC. 50 4 мкл клітин додавали в 96-ямкові планшети до кінцевої концентрації 810 клітин на ямку в RPMI, 5 % термоінактивованої FBS, 0,1 мМ замінних амінокислот, 1 мМ пірувату натрію (всі виробництва Invitrogen). Додаткові 50 мкл додавали в ямки в присутності або за відсутності антитіл, і планшети інкубували при кімнатній температурі протягом 2 годин. У ямки додавали 50 18 UA 113725 C2 5 10 15 20 25 30 35 40 45 50 55 мкл 10 % комплементу кролика (Invitrogen), і планшети інкубували протягом 20 хвилин при 37 °C. Всі проби були представлені в двох повторюваннях. Планшети центрифугували при 200g протягом 3 хвилин і переносили 50 мкл супернатанту в окремі планшети, після чого вимірювали CDC за допомогою набору вимірювання цитотоксичності LDH (Roche). Поглинання вимірювали на Spectra max Plus 384 (PerkinElmer). Дані нормалізували за максимальною цитотоксичністю з Triton X-100 (Sigma Aldrich) і мінімальним контролем, який містить тільки клітини і один лише комплемент. Дані апроксимували сигмоїдною моделлю ефекту дози з використанням GraphPad Prism, версія 5. Результати Дані, представлені на фігурі 8, показують, що конструкції 2h S267E/H268F/S324T/I332E (10) і 2h K326A/E333A (11) забезпечували рівні лізису клітин, аналогічні IgG1 wt. Конструкції IgG1 2h (4), 2h S239D/I332E (5), 2h F243L/R292P/Y300L (7) і 2h S239D/H268F/S324T/I332E (9) мали мінімальну здатність до CDC, що було аналогічно вимірюванням для IgG2 wt (2). ПРИКЛАД 5. ДОДАТКОВІ ПРОТЕАЗОСТІЙКІ КОНСТРУКЦІЇ Тільки дві мутації CH2 в комбінації з E233P/L234V/L235A з делецією G236 демонстрували CDC активність, порівнянну з IgG1 wt, а саме - 2h K326A/K334A (11) і 2h S267E/H268F/S324T/I332E (10). Однак 2h K326A/K334A (11) виявляла мінімальну активність за ADCC і ADCP, а 2h S267E/H268F/S324T/I332E (10) демонструвала знижену активність за ADCC в порівнянні з IgG1 wt. Важливе значення мала б можливість створювати протеазостійкі конструкції, які б виявляли всі три види активності (ADCC, ADCP і CDC). Для одних тільки мутацій H268F/S324T раніше була продемонстрована відсутність збільшення афінності до FcγRs (Moore et al.), тоді як конструкція 2h H268F/S324T/I332E виявляла підвищену активність за ADCC в порівнянні з 2h. Отже, мутація I332E окремо може відновити активність за ADCC у шарнірній протеазостійкій конструкції 2h. Отже, будуть створені конструкції, які поєднують відновлення активності як за ADCC/ADCP, так і за CDC для материнської шарнірної конструкції 2h, включаючи 2h K326A/I332E/E333A (15) (SEQ ID NO: 18), S239D/K326A/E333A (SEQ ID NO: 19) (16) і S267E/I332E (17) (SEQ ID NO: 20). Три конструкції тестували з використанням матеріалів і способів, описаних в прикладі 1. Три конструкції виявляли стійкість у відношенні MMP-3 і MMP-12 в порівнянні з IgG1 wt. Як було показано раніше, IgG2 wt (2) був стійкий до GluV8, тоді як у випадку IgG1 wt (1) через 24 години після розщеплення залишалося менше 60 % інтактного IgG. Три конструкції виявляли підвищену стійкість до GluV8 в порівнянні з IgG1 wt. Однак конструкції 2h K326A/I332E/E333A (15) і 2h S267E/I332E (17) відрізнялися зниженою стійкістю до GluV8 в порівнянні з IgG2 wt, тоді як 2h S239D/K326A/E333A (16) виявляла стійкість, порівнювану з IgG2 wt. Отримані дані свідчать про те, що мутації, які вводять додатковий Glu в CH2 в комбінації з мутацією нижньої шарнірної ділянки, формують новий сайт розщеплення для GluV8 (наприклад, 2h S239D/I332E (5), 2h K326A/I332E/E333A (15) і 2h S267E/I332E (17)), тоді як мутації, які не вводять Glu в CH2, демонструють стійкість до GluV8, аналогічну IgG2 wt (наприклад, 2h K326A/E333A (11) і 2h S239D/K326A/E333A (16)). Як IgG1 wt, так і IgG2 wt були піддані протеолізу під впливом IdeS. У разі двох конструкцій 2h K326A/I332E/E333A (15) і 2h S267E/I332E (17) залишалося більше 90 % інтактного IgG після 24 годин інкубації в IdeS, тоді як у разі конструкції 2h S239D/K326A/E333A (16) залишалося менше 20 % інтактного IgG. Дані результати показують, що додавання Glu в CH2 в комбінації з мутацією в нижньому шарнірі збільшує протеазостійкість до IdeS - властивість, яку не здатна забезпечити одна мутація нижнього шарніра, 2h (4). Три конструкції тестували на здатність виявляти активність по ADCP, ADCC і CDC. Три конструкції відрізнялися підвищеною активністю по ADCP в порівнянні з IgG2 wt і 2h (4), але зниженим максимумом по ADCP в порівнянні з IgG1 wt. Дві конструкції 2h K326A/I332E/E333A (15) і 2h S239D/K326A/E333A (16) відрізнялися декілька підвищеною активністю по ADCC в порівнянні з IgG1 wt. Конструкція 2h S267E/I332E (17) відрізнялася зниженою активністю ADCC в порівнянні з IgG1 wt, але підвищеним рівнем ADCC в порівнянні з IgG2 wt і 2h (4). Всі три конструкції демонстрували підвищений рівень CDC в порівнянні з IgG2 wt і 2h (4); однак рівень CDC для всіх трьох конструкцій був трохи нижче в порівнянні з IgG1 wt. ПРИКЛАД 6. ЗВЕДЕНА ІНФОРМАЦІЯ ПО СПРИЯТЛИВИХ МУТАЦІЯХ Було показано, що наступні одинадцять варіантів Fc забезпечують композицію антитіла, стійку до однієї або більше протеаз, здатних розщеплювати IgG1 в нижньому шарнірі, зберігаючи при цьому одну або більше ефекторних функцій, показаних диким типом людського IgG1. Символом 2h позначається IgG1 з E233P/L234V/L235A - делецією G236. 19 UA 113725 C2 Таблиця 5 Конструкція IgG1 2h S239D/I332E IgG1 2h F243L/R292P/Y300L IgG1 2h H268F/S324T/I332E IgG1 2h S239D/H268F/S324T/I332E IgG1 2h S267E/H268F/S324T/I332E IgG1 2h K326A/E333A IgG1 2h G237A/S239D/I332E IgG2 S239D/I332E IgG1 2h K326A/I332E/E333A IgG1 2h S239D/K326A/E333A IgG1 2h S267E/I332E 5 Скорочення (№) 2h DE (5) 2h LPL(7) 2h FTE (8) 2h DFTE (9) 2h EFTE (10) 2h AA (11) 2h XDE (12) IgG2 DE (14) 2h AEA (15) 2h DAA (16) 2h EE (17) При достатньому об'ємі даних нижче представлені значення EC50, які використовуються як наближення для оцінки різних ефекторних функцій і розраховуються в тих випадках, коли при аналізі in vitro знищення клітин було повним або практично повним. Дані, наведені в таблицях 6A і 6B, отримані в ідентичних умовах, однак джерело донорських PBMC було різним. Отже, для стандартизації відносної біологічної активності в кожному експерименті використовували кратність зміни в порівнянні з IgG1 wt (1). Таблиця 6A Конструкція (№) IgG1 wt (1) 2h DE (5) 2h G237A/DE (13) 2h LPL (7) 2h AA (11) 2h DFTE (9) 2h EFTE (10) ADCC EC50 (нг/мл) 4,8 0,53 Кратність Кратність 1 9 ADCP EC50 (нг/мл) 27 54 Кратність 1 0,5 CDC EC50 (нг/мл) 96 н/д 0,47 10 24 1,1 не визн. не визн. 0,31 21 0,70 н/д 15 0,2 7 н/д 30 н/д 34 44 0,9 н/д 0,8 0,6 н/д 157 н/д 77 н/д 0,6 н/д 1,2 1 н/д 10 Таблиця 6B Конструкція (№) IgG1 wt 2h AEA (15) 2h DAA (16) 2h EE (17) ADCC EC50 (нг/мл) 0,42 0,17 0,14 5,6 Кратність 1 2,5 3,4 0,08 ADCP EC50 (нг/мл) 186 44* 42* 52* Кратність 1 4,2 4,4 3,6 CDC EC50 (нг/мл) 114 202 308 592 Кратність 1 0,56 0,37 0,19 н/д = немає даних (недостатньо даних на кривій зв'язування для визначення EC50), * досягнутий субмаксимальний лізис; не визн. = не визначено; кратність = EC50 IgG1 wt/EC50 конструкції. 15 Вплив на ADCC і ACDP додаткової модифікації в G237 в нижньому шарнірі в доповнення до S239D/I332E і 233PVA/236 аналізували з використанням конструкцій, перерахованих нижче в таблиці 7. Тестували конструкцію G237A, і було встановлено, що вона має стійкість до MMP, IdeS і GluV8. В аналізах розщеплення інші конструкції не оцінювали. Наведені дані показують, що в положенні 237 допустимо використовувати Ala(А) і Ser (S), але вони не збільшують цитолітичну активність Fc зверх тієї, яку виявляла материнська молекула, 2h DE (5). 20 UA 113725 C2 Таблиця 7 Конструкція IgG1 2h DE IgG1 2h ADE IgG1 2h DDE IgG1 2h PDE IgG1 2h QDE IgG1 2h SDE 5 ADCC ++++++ ++++++ + ++ + +++++ ADCP +++++ ++++ + + + ++ У таблиці 8 представлена зведена інформація відносно комбінованої протеазостійкості (PR) до конкретних фізіологічно важливих протеаз, а також результати in vitro для приблизних аналізів, які вказують на потенційну ефекторну функцію (ADCC, ADCP і CDC), де ці конструкції з комбінованою протеазостійкістю і однієї або більше ефекторними функціями, що демонструються, представлені білим. Таблиця 8 Ізотип/конструкція IgG1 wt (1) IgG2 wt (2) IgG1 2hc (3) IgG1 2hc DE (13) IgG1 2h (4) IgG1 2h DE (5) IgG1 2h LPL (7) IgG1 2h FTE (8) IgG1 2h DFTE (9) IgG1 2h EFTE (10) 2h AA (11) 2h ADE (12) IgG2 DE (14) 2h AEA (15) 2h DAA (16) 2h EE (17) 10 15 20 25 30 PR MMP +++++ +++++ +++++ +++++ +++++ +++++ +++++ +++++ +++++ +++++ +++++ +++++ ++++ ++++ ++++ PR IdeS ++++ ++++ ++++ +++++ ++++ ++++ +++ +++++ + +++++ PR GluV8 + +++++ +++++ +++++ +++++ ++ ++ +++++ ++++ ++++ +++++ ++ ++ +++ +++++ +++ ADCC +++++ + + + ++++++ ++++++ ++++++ ++++++ + +++ ++++++ ++++++ ++++++ +++++ + ADCP +++++ ++ ++ ++ ++ +++++ +++++ +++ +++++ ++++ ++ ++++ ++++ +++ +++ +++ CDC +++++ не визн. не визн. +++++ +++++ не визн. не визн. ++++ ++++ ++++ Зведена інформація про результати Дослідження конструкцій Fc, які представлені в цьому документі, продемонструвало, що заміщення залишків по EU 233-236 на PVA/ сайтів, де, як відомо, протеази розщеплюють молекулу IgG1, дозволяє отримати Fc, які стійкі до MMP-3, MMP-12 і GluV8; протеаз, які розщеплюють молекулу між залишками 232 і 234 (фігура 1). В комбінації з даними заміщеннями додаткові модифікації забезпечували стійкість до протеази Staphylococcus IdeS незалежно від того, чи проводилася модифікація положень заміщених залишків в передбачуваному сайті розщеплення (EU 236-237) або в більш дистальному положенні. Одне тільки заміщення залишків EU 233-236 на PVA/ (2h, конструкція 4) приводило до втрати цитолітичних функцій, що оцінювали по аналізу in vitro, описаному для ADCC, ADCP і CDC. Що стосується комбінації модифікацій IgG1 Fc, які, як стверджували раніше, посилювали одну або більше ефекторних функцій (таблиця 2), заміщення нижнього шарніра PVA/ несподівано відновлювало один або більше аспектів цитолітичної активності in vitro. Таким чином, жодна з конструкцій не виявляла як протеазостійкість, так і помітну або підвищену активність відносно всіх трьох ефекторних функцій, які оцінювали по знищенню клітин in vitro або аналізах на лізис клітин для ADCC, ADCP і CDC. 1. Вісім конструкцій мали протеазостійкість і підвищену або порівнювану активність по ADCC в порівнянні з IgG1 wt: 5, 7, 8, 9, 12, 14, 15 і 16. Шість із даних конструкцій містили заміщення I332E, включаючи IgG1 2h DE (5), IgG1 2h FTE (8), IgG1 2h DFTE (9), IgG1 2h ADE (12), IgG2 DE (14) і IgG1 2h AEA (15). 2. Три конструкції PR демонстрували аналогічну активність по ADCP в порівнянні з IgG1 wt, включаючи IgG1 2h DE (5), IgG1 2h LPL (7) і IgG1 2h DFTE (9). Три конструкції виявляли декілька 21 UA 113725 C2 5 10 знижену активність по ADCP в порівнянні з IgG1 wt, включаючи IgG1 2h FTE (8), IgG1 2h EFTE (10), IgG1 2h ADE (12) і IgG2 DE (14). 3. П'ять мутацій PR забезпечували відновлення активності по CDC, IgG1 2h AA (11), IgG 2h EFTE (10), 2h AEA (15), 2h DAA (16) і 2h EE (17). Крім того, всі п'ять мутацій відрізнялися виявленою, але зниженою активністю по ADCP в порівнянні з IgG1 wt. Два варіанти, 2h AEA (15), 2h DAA (16), також демонстрували підвищену активність по ADCC в порівнянні з IgG1 wt (1). Дві конструкції (8 і 9), що містять мутації H268F/S324T, не виявляли відновлену активність по CDC при наявності протеазостійкого шарніра. Мутація S267E (EFTE (10)) відновлювала CDC, але знижувала зв'язування з FcγRIIIa (також вказано в роботі Moore et al. mAbs, 2010 р., 2(2): 181.) Мутація S267E підвищувала афінність до FcγRIIb. 22 UA 113725 C2 23 UA 113725 C2 24 UA 113725 C2 25 UA 113725 C2 26 UA 113725 C2 27 UA 113725 C2 28
ДивитисяДодаткова інформація
Назва патенту англійськоюActive protease-resistant antibody fc mutants
Автори англійськоюStrohl, William, Jordan, Robert, Brezski, Randall
Автори російськоюСтрол Уилльям, Джордан Роберт, Брезски Рэндалл
МПК / Мітки
МПК: C12N 15/13, A61K 39/395
Мітки: протеазостійкий, мутант, антитіла, активний
Код посилання
<a href="https://ua.patents.su/61-113725-aktivnijj-proteazostijjkijj-mutant-antitila-fs.html" target="_blank" rel="follow" title="База патентів України">Активний протеазостійкий мутант антитіла fс</a>
Спосіб радіоактивного мічення кон’югованого з комплексоном антитіла або фрагмента антитіла радіоактивним ізотопом ітрій-90 та комплект для здійснення способу

Номер патенту: 75036
Опубліковано: 15.03.2006
Автор: Чінн Пол
МПК: A61K 51/10, A61K 51/08, G01N 33/534
Мітки: антитіла, способу, комплект, комплексоном, ітрій-90, фрагмента, спосіб, мічення, ізотопом, кон'югованого, радіоактивного, радіоактивним, здійснення
Формула / Реферат:
1. Спосіб радіоактивного мічення кон'югованого з комплексоном антитіла або фрагмента антитіла з 90Y для введення пацієнту, у якому:і) змішують антитіло або фрагмент антитіла, що кон'юговане з біфункціональним комплексоном, вибраним із групи, яка включає MX-DTPA, феніл-DTPA, бензил-DTPA, CHX-DTPA, DOTA та їх похідні, з розчином, що містить 90Y або його сіль у прийнятному буфері, таіі) інкубують цю суміш протягом достатнього,...
Мутант глікозилування fsh

Номер патенту: 88879
Опубліковано: 10.12.2009
Автори: Кампбелл Роберт К., Аркінстелл Стефен Дж., Дзянь Сюлянь, Теппер Марк, Герон Луіз М., Маккена Шон Д., Брондик Уільям Х.
МПК: A61K 38/24
Мітки: глікозилування, мутант
Формула / Реферат:
1. Рекомбінантний FSH, в якому альфа-субодиниця FSH містить ПОСЛІДОВНІСТЬ №3, а бета-субодиниця FSH містить ПОСЛІДОВНІСТЬ №4.2. Рекомбінантний FSH за п. 1, який відрізняється тим, що N55 бета-субодиниці є глікозилованим.3. Рекомбінантний FSH за п. 1, який відрізняється тим, що N83 альфа-субодиниці є глікозилованим і Т57 бета-субодиниці є глікозилованим.4. Композиція, яка містить рекомбінантний FSH за п. 2 або 3 і...
Мутант людського інтерлейкіну-2, що активує переважно т-клітини у порівнянні з природними кілерними клітинами, фармацевтична композиція, полінуклеотид, вектор, прокаріотична клітина, спосіб стимуляції імунної с

Номер патенту: 73719
Опубліковано: 15.09.2005
Автори: Грів Джеффрі М., Лембах Кеннет Дж., Уетцел Гейл Д., Джесмок Гарі, Шанафелт Армен Б.
МПК: C12N 15/09, A61K 38/00, G01N 33/15, C07K 14/55, C12N 15/26, G01N 33/50, C12Q 1/02
Мітки: людського, клітина, т-клітини, природними, прокаріотична, спосіб, фармацевтична, композиція, мутант, порівнянні, кілерними, переважно, активує, інтерлейкіну-2, імунної, вектор, клітинами, полінуклеотид, стимуляції
Формула / Реферат:
1. Мутант людського інтерлейкіну-2, пронумерований у відповідності з диким типом інтерлейкіну-2, що має амінокислотну заміну в одному або двох положеннях 20, 88 та 126, який відрізняється тим, що амінокислотна заміна у положенні 20 містить амінокислоту з групи, що включає аланін, гістидин, ізолейцин, метіонін, глутамін, серин, валін та тирозин;амінокислотна заміна у положенні 88 містить амінокислоту з групи, що включає аланін, аргінін,...
Злитий поліпептид проти пухлини, індукованої вірусом eb, і мутант коліцину ia

Номер патенту: 104933
Опубліковано: 25.03.2014
Автор: Цю Сяоцин
МПК: C07K 19/00, A61K 35/00, A61K 38/16
Мітки: поліпептид, індукованої, пухлини, злитий, мутант, вірусом, коліцину
Формула / Реферат:
1. Поліпептид проти пухлини, викликаної вірусом ЕВ, який утворюють шляхом функціонального зв'язування мутантного поліпептиду коліцину, здатного утворювати іонні канали, з поліпептидом антитіла до вірусу ЕВ або поліпептидом міметиків антитіл до вірусу ЕВ, мутантний поліпептид коліцину, здатний утворювати іонні канали, одержують шляхом мутації амінокислотних залишків G11A, H22G, A26G, V31L і H40D у пептидному ланцюзі коліцину дикого типу Іа,...
Толерантний до інгібуючого als гербіциду мутант буряку звичайного

Номер патенту: 113721
Опубліковано: 10.03.2017
Автори: Шпрінгманн Клеменс, Донн Гюнтер, Хольтшульте Бернд, Бентінг Юрген, Янсен Рудольф, Кніттель-Оттлебен Наталі, Хаін Рюдігер, Лоокк Андреас
МПК: C12N 15/82, C12N 15/29, A01H 5/00, C07K 14/415
Мітки: інгібуючого, мутант, звичайного, гербіциду, буряку, толерантний
Формула / Реферат:
1. Толерантна до інгібуючих ALS гербіцидів рослина буряку звичайного та її органи, які включають мутацію в положенні, відповідному положенню 1705-1707 ендогенного гена ацетолактатсинтази (ALS), показаного еталонною нуклеотидною послідовністю SEQ ID NO: 1, причому ALS ген кодує поліпептид ALS, який містить лейцин замість триптофану, у позиції 569 поліпептиду ALS, і який є гомозиготним щодо зазначеної мутації ендогенного...
Попередній патент: Спосіб одержання кулінарного смакоароматичного харчового продукту
Наступний патент: Замок (запор) з рухомим елементом
Випадковий патент: Спосіб автоматизованої обробки документів