Застосування s-аденозилметіоніну (sam) та супероксид-дисмутази (sod) для лікування хвороби альцгеймера
Номер патенту: 97440
Опубліковано: 10.02.2012
Автори: Фусо Андреа, Россіні Мауро, Скарпа Сіґфрідо, Даміані Роселліна




Формула / Реферат
1. Застосування S-аденозилметіоніну (SAM) у комбінації з супероксид-дисмутазою (SOD) для отримання ліків для лікування хвороби Альцгеймера.
2. Застосування за п. 1, де ліки інгібують надмірно велику експресію PS1 та ВАСЕ.
3. Застосування за п. 1 або п. 2, де ліки уводять перорально.
4. Комплект ліків для одночасного або послідовного застосування у лікуванні хвороби Альцгеймера, який містить (і) S-аденозилметіонін та (іі) супероксид-дисмутазу.
5. Комплект за п. 4, де SAM є у формі тозилату, бутадієнсульфонату, дисульфату тозилату, дисульфату дитозилату або дисульфату монотозилату, або міститься у клітинах Saccharomyces cerevisiae, збагачених SAM.
Текст
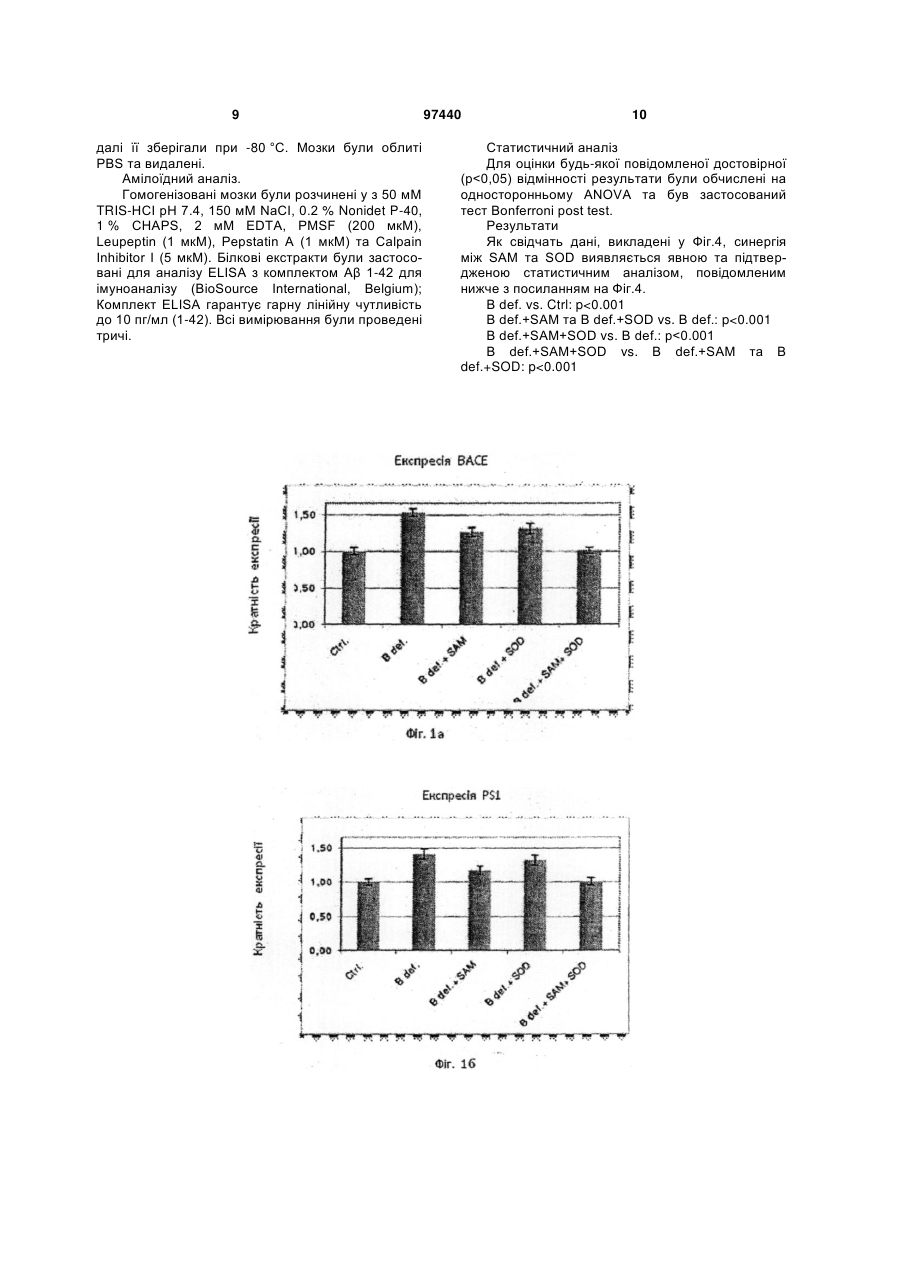
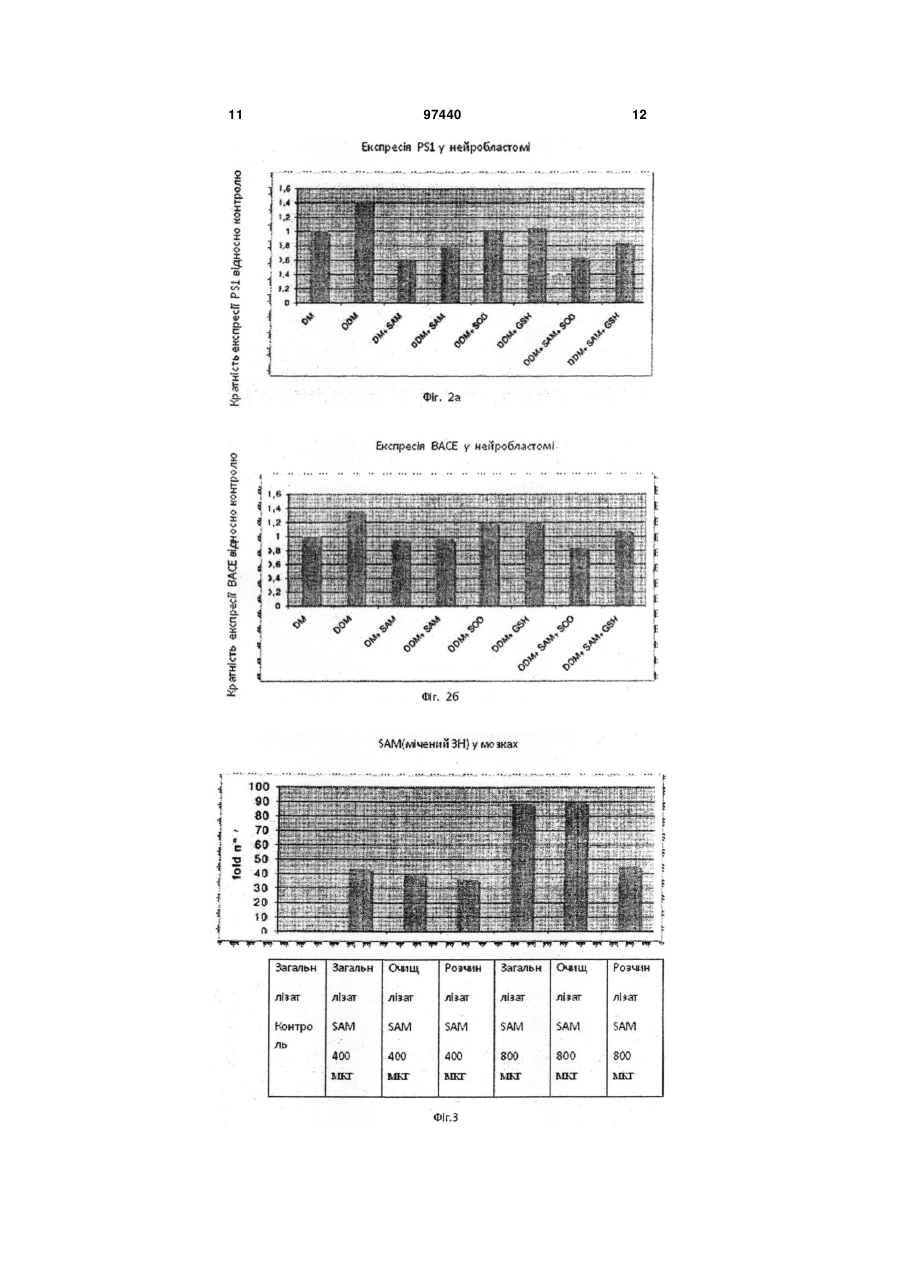
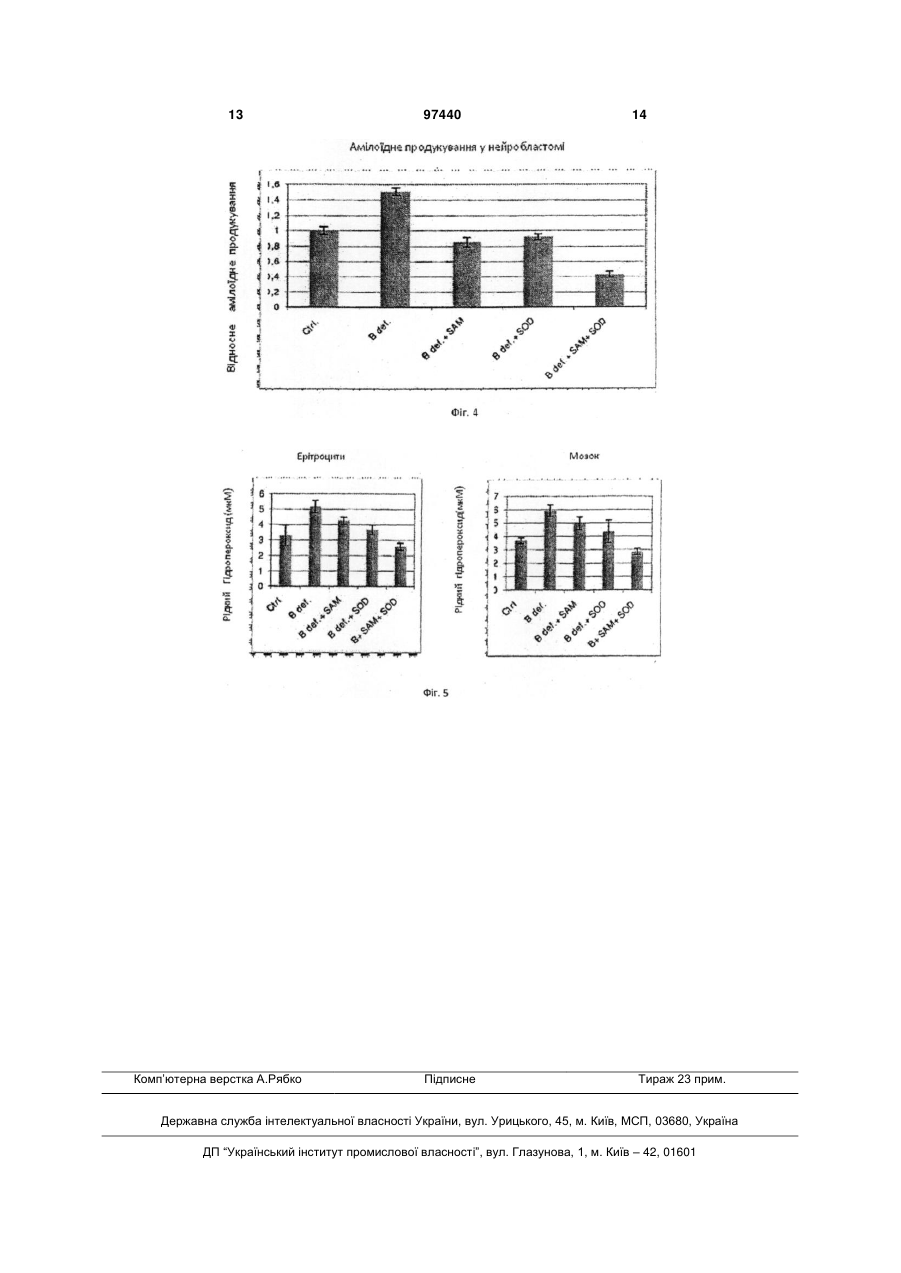
1. Застосування S-аденозилметіоніну (SAM) у комбінації з супероксид-дисмутазою (SOD) для отримання ліків для лікування хвороби Альцгеймера. 2. Застосування за п. 1, де ліки інгібують надмірно велику експресію PS1 та ВАСЕ. 3. Застосування за п. 1 або п. 2, де ліки уводять перорально. 4. Комплект ліків для одночасного або послідовного застосування у лікуванні хвороби Альцгеймера, який містить (і) S-аденозилметіонін та (іі) супероксид-дисмутазу. 5. Комплект за п. 4, де SAM є у формі тозилату, бутадієнсульфонату, дисульфату тозилату, дисульфату дитозилату або дисульфату монотозилату, або міститься у клітинах Saccharomyces cerevisiae, збагачених SAM. UA (21) a201010400 (22) 25.02.2009 (24) 10.02.2012 (86) PCT/EP2009/001323, 25.02.2009 (31) 08425123.0 (32) 29.02.2008 (33) EP (46) 10.02.2012, Бюл.№ 3, 2012 р. (72) СКАРПА СІҐФРІДО, IT, ФУСО АНДРЕА, IT, ДАМІАНІ РОСЕЛЛІНА, IT, РОССІНІ МАУРО, IT (73) ҐНОСІС С.П.А., IT (56) US 2003/129261 A1, 10.07.2003 WO 2005/041996 A, 12.05.2005 FUSO A ET AL: "B-vitamin deprivation induces hyperhomocysteinemia and brain Sadenosylhomocysteine, depletes brain Sadenosylmethionine, and enhances PS1 and BACE expression and amyloid-[beta] deposition in mice" MOLECULAR AND CELLULAR NEUROSCIENCE 200804 US, vol. 37, no. 4, April 2008 (2008-04), pages 731-746, XP002486515 FUSO ANDREA ET AL: "gamma-Secretase is differentially modulated by alterations of homocysteine cycle in neuroblastoma and glioblastoma cells." JOURNAL OF ALZHEIMER'S DISEASE : JAD JUN 2007, vol. 11, no. 3, June 2007 (2007-06), pages 275-290, XP009102313 SCARPA SIGFRIDO ET AL: "Gene silencing through methylation: an epigenetic intervention on Alzheimer C2 2 (19) 1 3 чинює дегенерацію мозку та смерть, але у період у 10 або більше років. Мозок містить велику кількість бляшок у міжнейронних проміжках та типово нейрофібрилярні вузли у нейронах, особливо у церебральному кортексі, гіпокампусі, мигдалевидному тілі, та у інших частинах мозку, що відповідають за розумові функції. Амілоїдні бляшки, також відомі, як старечі бляшки, є полімерами бета-амілоїдного пептиду (Аβ), що походить від більшого білку, що має назву бета-амілоїдного попередника (АРР). АРР належить до родини високо консервативних трансмембранних глікопротеїнов. У останні десять років численні дослідження, що були зроблені вченими, які вивчають AD, забезпечили розуміння його етіології, особливо що стосується молекулярних механізмів. Дослідження молекулярних компонентів та їх регуляції може прояснити їх терапевтичні та діагностичні перспективи. Увага була зосереджена на презенілінах, що грають дуже важливу роль у процесінгу АРР, отже у продукуванні Αβ. Як було показано, ці білки, презенілін 1 (PS1) та презенілін 2 (PS2), відповідають за ензиматичну активність або регулюють активність інших ферментів, що мають назву секретази, які розщеплюють АРР у катаболіти, що здатні нормально розпадатися (альфа-секретаза) або у пептид Αβ (бета - та гамма-секретаза). У спадковій AD, мутації генів, необхідних для кодування PS1 та PS2 ведуть до непомірного продукування Αβ та окремого накопичення ізоформи Αβ-42,що є дуже амілогеничним процесом. У останній час, експерименти на PS1-модифікованих мишах показали велике зниження активності гамма-секретази, що демонструє те, що PS1, а також комплекс, що належить до гамма-секретази, також головним чином відповідають за продукування та накопичення Αβ. Стосовно бета-секретази, то вважається, що продукт ВАСЕ-гена може її самостійно розщеплювати. Розвиток клінічно корисного застосування інгібіторів гамма- та бета-секретази може стати вирішальною зброєю проти хвороби Альцгеймера, та є тепер одним з найбільш зворушливих конкурентних напрямків у нейрологічній науці. Безумовно показано, що активність PS1 не може бути повністю припинена, тому що цей білок є необхідним для процесу трансдукції фактору Notch-1, критичного фактору для дозрівання численних стовбурових клітин, таких як клітини, що причетні до кровотворення. Регуляція експресії гену способами метилування ДНК може бути добре вивчена у системі культури клітин, що здатна до експресії генів причетних до AD. Згідно з результатами наших досліджень було знайдено дуже інтересне проявлення регуляторних механізмів, причетних до старіння, що проявляється у послідовному, глобальному збільшенню гіпометилування ДНК у літніх людей, та накопичуванні гомоцистеїну.що було спостережено у пацієнтів, які страждають від старечого недоумства. Накопичення гомоцистеїну та гіпометилування ДНК є метаболічно співвідносними, тому що відсутність гомоцистеїн, що може бути перетворений у метіонін змінює метаболізм у напрямок до синтезу S-аденозилгомоцистеїну, що, 97440 4 як відомо, є сильним інгібітором ДНКметилтрансферази, і тому спричинює гіпометилування ДНК. У відповідності з добре обґрунтованою теорією, багато генів починають свою експресію, коли цитозіни специфічних послідовностей є деметіловані, ці біохімічні структури можуть привести до експресії неекспресованих генів та надмірно великої експресії генів з зазвичай нормальною експресією. Це може спричинювати AD, тому що надмірно велика експресія PS1 (гамма-секретази), може дискретно перевищувати активність альфасекретази, отже буде продукуватися пептид Αβ, що стане накопичуватися, та може спричинити захворювання через декілька років. Подальше проявлення можливої ролі метилування ДНК у AD - це знахідка того, що у хворих на AD пацієнтів присутні більш низькі посмертні рівні донорів метилу у мозку. Низька придатність Саденозилметіоніну (SAM) може легко привести до змінення або збільшення експресії генів, причетних до метаболізму АРР, та зрештою до продукування та накопичення пептиду Αβ у старечих мозкових бляшках. Попередні експерименти були проведені на лінії нейробластоми (SK-N-SH) що продукує АРР, PS1, PS2, ВАСЕ, альфа-секретазу, інші компоненти гамма-секретази, та NotcM. Культури були оброблені культурним середовищем, що було позбавлене фолату, вітаміну В12 та вітаміну В6 (для того, щоб змінити метаболізм гомоцистеїну), до якої було додано SAM у різних концентраціях (щоби збалансувати позбавлення вітаміну). Ми знайшли підвищення експресії PS1 та ВАСЕ у середі, що була позбавлена вітаміну В, та а відмічено зниження експресії PS1 та ВАСЕ після введення SAM. У експериментах з Hpall/PCR на промоторах АРР та PS1 ми виявили важливу різницю у метилування одного з CpG сайтів PS1 промотору. Ми дійшли до висновку, що дію PS1 гену можна частково усунути шляхом введення екзогенного SAM. Введення SAM може скоротити експресію PS1, поновлюючи метаболічний баланс у бік альфасекретази. Експерименти також проводилися з трансгенними мишами лінії TgCRND8, та відповідними контролями; ці миші характеризуються присутністю людського мутованого гену АРР, та отже можуть формувати амілоїдні бляшки у короткий час. Ці тварини були відгодовані повною дієтою, або дієтою без вітамінів групи В, та знову, як і у клітинах, було відмічено збільшення експресії PS1 та ВАСЕ. У обох експериментальних моделях, змінення експресії гену приводило до ефекту зростання активності гамма - та бета-секретази з наступним надмірним продукуванням Αβ, що накопичувався у формі бляшок старіння більш швидко, чим у тварин, відгодованих контрольною дієтою. Результати досліджень, що коротко викладені вище, були надруковані у наступних публікаціях: Fuso Α., Nicolia V., Cavallaro R.A., Ricceri L, D'Anselmi F., Coluccia P., Calamandrei G. and Scarpa S. 2008. B-Vitamin Deprivation Induces Hyperhomocysteinemia and Brain Sadenosylhomocsyteine, Depletes Brain Sadenosylmethionine, and Enhances PS1 and BACE 5 Expression and Amyloid-β Deposition in Mice. МоІ. Cell. Neurosci. 37: 731-746. Fuso Α., Cavallaro R.A., Zampelli Α., D'Anselmi F., Piscopo P., Confaloni A. and Scarpa S. 2007. γsecretase is differentially modulated by alterations of Homocysteine cycle in neuroblastoma and glioblastoma cells. J. Alz. Dis. 11: 275-290. Cavallaro R. Α., Fuso Α., D'Anselmi F. and Scarpa S. 2006. The effect of S-adenosylmethyonine on CNS gene expression studied by cDNA mycroarrays analysis. J. Alz. Disease. 9: 415-419. Scarpa S., Cavallaro R.A., D'Anselmi F. and Fuso A. 2006. Gene silencing through methylation: an epigenetic intervention on Alzheimer Disease. J. Alz. Disease. 9: 407-414. Fuso Α., Seminara L, Cavallaro R.A., D'Anselmi F. and Scarpa S. 2004. Homocysteine/Sadenosylmethionine Cycle Alterations Unbalance DNA Methylation Status with Consequent Upregulation of Beta-amyloid Production. МоІ. Cell. Neurosci. 28(1): 195-204. Scarpa S., Fuso Α., D'Anselmi F., Cavallaro R.A. 2003. Presenilin 1 gene silencing by Sadenosylmethionine: a treatment for Alzheimer disease? FEBS Letters 541 (1-3):145-148. Fuso Α., Cavallaro R. Α., Orrù L, Buttarelli F.R. and Scarpa S. 2001. Gene silencing by Sadenosylmethionine in muscle differentiation. FEBS Letters 508 (3): 337-340. Застосування SAM для лікування AD було також запропоновано у US 2002/025926 та US 2004/0048827. Нещодавно отримана інформація свідчить про здатність SAM змінювати експресію генів бета-секретази, презеніліну 1 та 2 та гена попередника бета-амілоїдного білку. Дослідження, що проводилися з міченим (тритієм) SAM вже після вказаних вище запатентованих заяв показали здатність цієї молекули впливати, як описано вище, на центральну нервову систему після перорального введення. О наявності корисного ефекту SOD у лікуванні AD було припущено у патентах US 5519058 та CN 1099224. Фармацевтичні композиції, що містять SAM та SOD разом з деякою кількістю активних інгредієнтів для застосування у інших, ніж AD випадках, вказані у US 2003/129261 та у WO 2005/041996. Як тепер відомо, активність SаденозилІметіоніну може бути надзвичайно підвищена, коли його введено у комбінації з супероксид-дисмутазою (SOD). Цей фермент не тільки здатен полегшити проходження Sаденозилметіоніну через гемато-енцефалічний бар'єр, він також синергетично взаємодіє з SAM у зменшенні експресії PS1 та ВАСЕ генів, які мають надмірну експресію у результаті нестачі вітаміну В. Один аспект винаходу стосується виробництва містящих S-аденозилметіонін або його похідних форм та супероксид-дисмутази у формі об'єднаного препарату для одночасного, окремого, або послідовного введення у лікуванні хвороби Альцгеймера. Дозування може змінюватися у широкому діапазоні, зважаючи на дуже малу токсичність SAM та SOD, та залежить від кількості таких факторів, як 97440 6 вага, стать та вік пацієнта. Тим не менше, в загальних рисах, вона буде коливатися між 200 та 2000 мг на добу для SAM та між 50 та 1000 мг на добу для SOD. SAM бажано вводити перорально, або у своєму вигляді або у формі стабільної солі, такої як тозилат, бутандисульфонат, дисульфат тозилату, дисульфат дітозилату або дисульфат монотозилату. Сприятливою формою введення є клітини Saccharomyces cerevisiae, збагачені SAM, що описані у WO 2006/131382. SOD, отриманий шляхом ферментації з штамів Saccharomyces cerevisiae, як описано у WO 2006/131382, що подано Заявником, може також бути введено перорально, з підтримкою гліадинової плівки, або з використанням інших шлункозахисних способів. Як альтернатива пероральному шляху введення, обидва SAM та SOD можуть бути введені перентерально, шляхом внутрішньом'язевої ін'єкції. Приклади цих композицій містять також вкриті плівкою з акрилового або метакрилового полімеру пігулки, капсули гастрорезистентні капсули, мікрокапсули та ін. SAM та SOD можуть бути присутні разом, у одній дозі, або приготовані та введені: у цьому разі комплекти можуть містити, два ліки у окремих формах дозування у супроводі з інструкціями по їх послідовному, одночасному, або окремому застосуванню. Опис фігур. Фіг.1а та 16 показують ефект SAM та SOD у мишей, протягом надмірно великої експресії ΡS1 та ВАСЕ, яку викликано застосуванням дієти з нестачею вітамінів групи В. Фіг.2 показує ефект SOD та глутатіону (GSH) під час надмірно великої експресії PS1 та ВАСЕ, у клітинах нейробластоми лінії SK-N-BE, окремо, та у комбінації з SAM. Фіг.3 показує рівні міченого тритієм SAM у мозках мишей після перорального введення SAM 400 мкг/добу. Фіг.4 показує вимірювання бета-амілоїдного продукування у клітинах людської нейробластоми через один тиждень лікування. Фіг.5 показує ефект SAM та SOD під час оксидативного стану у еритроцитах та мозку мишей після застосування дієти з нестачею вітамінів групи В. Далі винахід буде більш детально описаний за допомогою фармакологічних випробувань, що викладені нижче у якості прикладів. Приклад 1 - Експресія гену. Зокрема дію SAM було перевірено у фармакологічних концентраціях (400 мкг/добу) окремо у мишей TgCRND8 та у відповідних контрольних мишей дикого типу. Канали експресії гену. З клітин культури та гомогенізованих мозків було виділено РНК та синтезовано кДНК. По 0.5 мкг з цієї кДНК було використано для кожної реакції у реальному часі на Opticon2 DNA Engine (MJ Research) з використанням реагентів SYBR-Green. Ефективність ампліфікації для кожної пари праймерів булла заздалегідь визначена ампліфікацією 7 за стандартною лінійною кривою. Експериментальні проби були порівняні зі стандартною кривою специфічного гену для визначення кількості специфічних кДНК, присутніх у стандартній реакції. Стандарти, що отримані з високо очищених PCR продуктів були ампліфіковані з позитивними контролями. Підсумкові рівні кДНК були підпорядковані до β-актинового контролю (ген "домашнього господарства"). Результати показали, що SAM також змінює у протилежний напрямок надмірно велику експресію PS1 та ВАСЕ, викликану позбавленою дієтою in vivo (Фіг.1), та навіть зменшує її до рівнів, що нижче, ніж рівні у мишей з контрольною дієтою. Тоді як метаболізм гомоцистеїну є присутнім у обох реакціях, як метилування, так і окиснення, ми перевірили ефект різних антиоксидантів на експресію двох генів. Перші дані, отримані з клітин нейробластом лінії SK-N-BE продемонстрували, що обидві речовини, як SOD, так і глутатіон (GSH) інгібують надмірно велику експресію PS1 та ВАСЕ, викликану нестачею вітаміну, хоча і в меншій мірі, ніж SAM. Проте, цікаво буде звернути увагу на те, що, коли SOD та SAM введені разом, вони дають синергічний ефект (Фіг.2) з більш поглибленим зменшенням експресії двох генів до більш низьких рівнів, чим τі, що було спостережено з SAM окремо (на 15-20 % нижче). Приклад 2 - Засвоювання SAM. Проведені на мишах та на клітинах експерименти показують, що SAM проходить скрізь гемато-енцефалічний бар'єр. Для культури клітин, 100 мкМ SAM було додано до клітинної культури середи F14 (повної або позбавленої вітаміну В, відповідно до мети досліду) та ці культури було зупинено через 96 годин. Для мишей, 400 мкг/добу SAM було введено перорально зондом-голкою для годування, та через два місяці тварин було вбито; тварин годували повною дієтою, або дієтою, позбавленою вітаміну В, відповідно до мети досліду. Рівні SAM були аналізовані HPLC з Varian HPLC системою. Клітини та гомогенізовані мозки були розчинені у дистильованій воді, та макромолекули осаджувалися з 1.5 Μ РСА. Стандартні криві SAM обчислювалися перед експериментальними пробами та після них. Для аналізу міченого тритієм SAM, клітини були оброблені так, як описано вище, у той час як мишей було витримано чотири доби для того, щоби звести до мінімуму вплив радіоактивного елементу. Клітини та гомогенізовані мозки були розчинені у дистильованій воді. Частина розчинених мозків була піддана дії ультразвуку та центрифугована для відокремлення мембран та клітинних органел; інша частина була оброблена перхлоратною кислотою (РСА) після обробки ультразвуком для відокремлення макромолекул. Засвоювання радіоактивного SAM було поміряно сцинтиляцією у бета-лічильнику. Було знайдено, що внутрішньоклітинні рівні SAM підвищилися від 1 (контроль) до 2.5-3 мкМ. Щоб встановити, чи це збільшення було обумовлено засвоюванням екзогенного SAM, а не зростанням ендогенних резервів у присутності підвищених позаклітинних концентрацій молекули, було 97440 8 проведено випробування з радіоактивним SAM. 100 мкМ міченого тритієм SAM (SAM[3H]) було додано до культури клітин SK-N-BE, та засвоювання було обчислено лізисом клітин та підрахунком радіоактивності. У лізатах клітин було знайдено 1.5 мкМ радіоактивного SAM; цей рівень є порівняльним до збільшення від 1 до 2.5 разів, знайденого для неміченого тритієм SAM, та ясно показує, що внутрішньоклітинне збільшення обумовлено засвоюванням екзогенного SAM. Подібне збільшення рівнів SAM було знайдено у мозкових лізатів мишей, яким було введено перорально 400 мкг/добу SAM; та ще раз знову, коли більш поглиблений тест був проведений SAM[3H] у концентрації 400 та 800 мкг/добу. Підвищення радіоактивності у мозкових лізатів в цілому є порівняним до 0.5 нг SAM на мозок миші, оброблений 400 мкг SAM, та 1 нг на мозок миші, оброблений 800 мкг SAM (Фіг.3). Частина загальних мозкових лізатів була також оброблена ультразвуком та центрифугована для отримання очищеного лізату (цитоплазми), та далі частину цього очищеного лізату було осаджене з перхлоратною кислотою (РСА) для усунення білків (розчинний лізат). Цікаво звернути увагу на те, що розчинний лізат з миші, яка отримала 800 мкг SAM показує радіоактивний рівень порівняно до 0.5 нг SAM (тоді як загальний та очищений лізати показали більш високі рівні), що вказує на те, функція надлишкового метилування, що поєднана з екзогенним SAM була зв'язана з молекулами інших клітин. Приклад 3 - Амілоїдне продукування у клітинних лініях нейробластоми. Способи: Середовище та культури клітин. Клітинна лінія SK-N-BE людської нейробластоми зберігалася у середі F14 з 10 % FCS та була перенесена у повну спеціалізоване середовище (контрольна середа, з 1 % FCS та 10 мкМ ретиноєвої кислоти) або у спеціалізоване середовище, що позбавлене фолату, вітаміну В12 та вітаміну В6 (В- позбавлене). Культури підживлювалися кожну другу доби та були зупинені після 96. Тварини та дієти. Миші у віці приблизно 3 тижня, що були систематично розподілені у одну з двох груп - групу з контрольною дієтою або групу з неповною дієтою, отримували їжу та воду ad libitum. Контрольна (AIN-93M; дієта А: фолат 1.98 мг; вітамін В12 0.025 мг; вітамін В6 7мг) та експериментальна дієта (AIN-93M В; дієта В, позбавлена фолату, вітаміну В12 та вітаміну В6) були закуплені у Mucedola (Italy). Обидві дієти містили 1 % сульфатіазолу для інгібування утворення фолату за допомогою кишкових бактерій та забезпечення того, щоби єдиним джерелом фолієвої кислоти була дієта. Більш того, інші три групи тварини отримували SAM (800 мкг/добу) або SOD (10 од/добу) або комбінацію обох ліків (SAM 400 мкг/добу та SOD 5 од/добу). Через один тиждень обробки, миші були піддані наркозу та вбиті для отримання мозку та крові. Кров була зібрана пункцією з серця у тестову тубу, що містила ΕDTA 2 г/дл та негайно центрифугована для відокремлювання плазми та еритроцитів, 9 далі її зберігали при -80 °C. Мозки були облиті PBS та видалені. Амілоїдний аналіз. Гомогенізовані мозки були розчинені у з 50 мМ TRIS-HCI рН 7.4, 150 мМ NaCI, 0.2 % Nonidet P-40, 1 % CHAPS, 2 мМ EDTA, PMSF (200 мкМ), Leupeptin (1 мкМ), Pepstatin A (1 мкМ) та Calpain Inhibitor I (5 мкМ). Білкові екстракти були застосовані для аналізу ELISA з комплектом Αβ 1-42 для імуноаналізу (BioSource International, Belgium); Комплект ELISA гарантує гарну лінійну чутливість до 10 пг/мл (1-42). Всі вимірювання були проведені тричі. 97440 10 Статистичний аналіз Для оцінки будь-якої повідомленої достовірної (р
ДивитисяДодаткова інформація
Назва патенту англійськоюUse of s-adenosylmethionine (sam) and superoxide dismutase (sod) for the preparation of medicaments for the treatment of alzheimer's disease
Автори англійськоюScarpa, Sigfrido, Fuso, Andrea, Damiani, Rosellina, Rossini, Mauro
Назва патенту російськоюПрименение s-аденозилметионина (sam) и супероксид-дисмутази (sod) для лечения болезни альцгеймера
Автори російськоюСкарпа Сигфридо, Фусо Андреа, Дамиани Роселлина, Россини Мауро
МПК / Мітки
МПК: A61P 25/28, A61K 31/7076, A61K 38/44
Мітки: sod, альцгеймера, лікування, сам, хвороби, супероксид-дисмутази, s-аденозилметіоніну, застосування
Код посилання
<a href="https://ua.patents.su/7-97440-zastosuvannya-s-adenozilmetioninu-sam-ta-superoksid-dismutazi-sod-dlya-likuvannya-khvorobi-alcgejjmera.html" target="_blank" rel="follow" title="База патентів України">Застосування s-аденозилметіоніну (sam) та супероксид-дисмутази (sod) для лікування хвороби альцгеймера</a>
Застосування фрагмента аb16-23 для лікування хвороби альцгеймера

Номер патенту: 87453
Опубліковано: 27.07.2009
Автори: Бард Фредерік, Сьюберт Пітер А., Єднок Тед, Васквез Нікі
МПК: A61P 25/28, A61K 39/00, A61K 38/08
Мітки: застосування, хвороби, лікування, фрагмента, аb16-23, альцгеймера
Формула / Реферат:
1. Застосуванняі) фрагмента Аβ, що складається з Аβ16-23 і має амінокислотну послідовність KLVFFAED, абоіі) фрагмента Аβ, що складається з Аβ16-23 і має амінокислотну послідовність KLVFFAED, де фрагмент зв'язаний з молекулою-носієм з метою утворення кон'югата, де молекула-носій допомагає викликати імунну реакцію на фрагмент, у виробництві лікарського засобу для ефективного лікування або профілактики...
Застосування препарату ненасиченої жирної кислоти для лікування депресії та хвороби альцгеймера

Номер патенту: 79744
Опубліковано: 25.07.2007
Автор: Хорробін Дейвід Фредерік
МПК: A61K 31/19, A61P 25/24, A61K 31/405, A61P 25/28, A61K 31/20, A61K 31/415
Мітки: кислоти, лікування, ненасиченої, жирної, хвороби, застосування, препарату, альцгеймера, депресії
Формула / Реферат:
1. Застосування препарату жирної кислоти, що містить (а) більше за 70 % ейкозапентенової кислоти (ЕРА) або похідної ейкозапентенової кислоти, менше за 10 % докозагексенової кислоти або похідної докозагексенової кислоти та менше за 10 % лінолевої кислоти або похідної лінолевої кислоти; та (b) інгібітор ферментів, вибраний з групи, до якої входять інгібітор СОХ-1 та/або СОХ-2, інгібітор LOX та інгібітор одного або...
Застосування комбінації донепезилу та мемантину для лікування хвороби альцгеймера

Номер патенту: 82480
Опубліковано: 25.04.2008
Автори: Томсен Ларс Люкке, Педерсен Аннерс Ґерсел
МПК: A61P 25/28, A61K 31/13, A61K 31/445
Мітки: мемантину, застосування, альцгеймера, хвороби, лікування, комбінації, донепезилу
Формула / Реферат:
Застосування композиції, яка містить ефективну кількість донепезилу або його фармацевтично ефективної солі та ефективну кількість мемантину або його фармацевтично ефективної солі, для виготовлення лікарського препарату для лікування слабкого порушення пізнавальної здатності або деменції, пов’язаної з хворобою Альцгеймера у стадії від помірної до важкої.
Похідні піримідину та їх застосування у терапії та у виробництві лікарського засобу для запобігання та/або лікування хвороби альцгеймера

Номер патенту: 92181
Опубліковано: 11.10.2010
Автори: Педерсен Торбен, Арзель Ерван, БЕРГ Стефан, Берроуз Джеремі, Стааф Карін, Турек Домініка, Геллберґ Свен, Рейн Тобіас, Андерссон Ларс, Роттічі Дідьє, Уерта Фернандо
МПК: A61P 25/28, C07D 405/14, A61K 31/506, A61P 3/10, C07D 401/14, C07D 403/04, A61P 25/16
Мітки: хвороби, похідні, піримідину, запобігання, засобу, альцгеймера, лікування, лікарського, виробництві, терапії, застосування
Формула / Реферат:
1. Сполука формули І: де R1 є вибраним з водню, ціано, С1-3галоалкілу, ORa, SO2NRbRc, C0-2алкілC(O)NRbRc, С1-4алкілNRbRc, CH2ORh, SO2Ri, C(O)ORa, CH(OH)Rj та C(O)Rj; R2 та R4 є незалежно вибраними з водню, гало, ціано, NO2, С1-4алкілу, С1-3галоалкілу, ORa, C(O)NRbRc, SO2Rі та C(O)ORa;...
Спосіб лікування хвороби альцгеймера

Номер патенту: 59946
Опубліковано: 15.09.2003
Автори: Букреєва Ірина Володимирівна, Волошин Петро Власович, Павлов Анатолій Юлійович, Букреєв Володимир Іванович, Пайкова Людмила Миколаївна, Підкоритов Валерій Семенович
МПК: A61K 31/00
Мітки: лікування, спосіб, хвороби, альцгеймера
Формула / Реферат:
Спосіб лікування хвороби Альцгеймера шляхом транскраніальної анодної мікрополяризації (ТКАМП), який відрізняється тим, що спочатку до хворого застосовують трансорбітальний електрофорез (ТЕФ) з ноотропними препаратами протягом 7 днів, а потім проводять 3 - 5 сеансів ТКАМП.