Людська гібридна клітина для експресії гену ссавців, клітинна лінія людини нкв11, спосіб отримання гібридних людських клітин, придатних для експресїї гетерологічного протеїну





Формула / Реферат
1. Клітина, отримана злиттям клітини людської ембріональної нирки , що має походження від 293 та клітини, що має походження від лімфоми Беркітта і є клітиною 2В8 (депозитний номер АТСС: CRL-12569).
2. Клітина за п. 1, в якій клітина людської ембріональної нирки є клітиною, що має походження від 293S.
3. Клітина за п. 1, яка експресує гетерологічний протеїн.
4. Клітина за п. 3, в якій гетерологічний протеїн вибрано з групи, яка складається з FVIII, BDD-FVІІI, моноклонального антитіла, антитіла анти-TNF, rІL4, tPA та ЕРО.
5. Клітинна лінія людини, позначена НКВ11 (депозитний номер АТСС: CRL-12568), придатна для рекомбінантного отримання терапевтичних протеїнів.
6. Клітинна лінія людини, позначена НКВ11 (депозитний номер АТСС: CRL-12568), модифікована шляхом введення гетерологічного гена і здатна експресувати гетерологічний протеїн.
7. Клітинна лінія за п. 6, в якій протеїном є ІСАМ-1.
8. Клітинна лінія за п. 6, в якій протеїном є BDD-FVIII.
9. Клітинна лінія за п. 6, в якій протеїном є моноклональне антитіло.
10. Клітинна лінія за п. 9, в якій моноклональним антитілом є анти-TNF.
11. Клітинна лінія за п. 6, в якій протеїном є rІL4.
12. Клітинна лінія за п. 6, в якій протеїном є FVIII.
13. Клітинна лінія за п. 6, в якій протеїном є tPA.
14. Клітинна лінія за п. 6, в якій протеїном є ЕРО.
15. Клітинна лінія за п. 6, в якій протеїном є IL-4SA (T13D/R121E), клітинна лінія позначена 1G2 (депозитний номер АТСС: РТА-87).
16. Клітинна лінія за п. 6, в якій протеїном є людський протеїн, який має людський профіль глікозилювання.
17. Спосіб отримання гібридних людських клітин, придатних для експресії гетерологічного протеїну, який містить наступні стадії:
a) отримання клітин людської ембріональної нирки, що має походження від 293, які мають першу бажану характеристику,
b) отримання клітин, що мають походження від лімфоми Беркітта і є клітиною 2В8 ((депозитний номер АТСС: CRL-12569), які мають другу бажану характеристику,
c) введення в контакт клітин, отриманих на стадії а) з клітинами, отриманими на стадії b) при умовах, за яких відбувається злиття клітин,
d) відбір клітин, які отримують на стадії с) для одержання клітин, які виявляють щонайменше одну бажану характеристику кожної з клітин, отриманих на стадіях а) та b).
18. Спосіб за п. 17, в якому клітинами людської ембріональної нирки є клітини 293S.
Текст
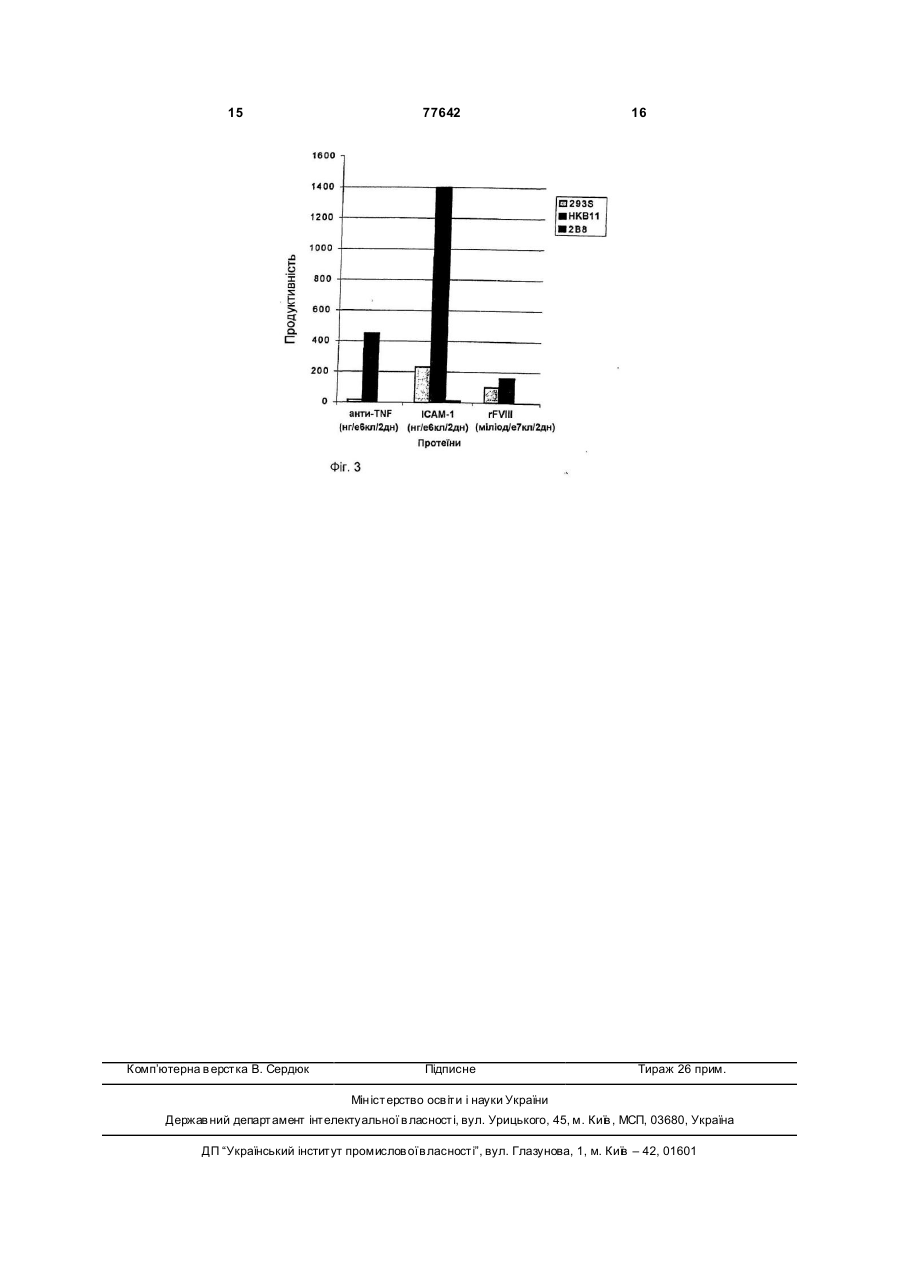
1. Клітина, отримана злиттям клітини людської ембріональної нирки , що має походження від 293 та клітини, що має походження від лімфоми Беркітта і є клітиною 2В8 (депозитний номер АТСС: CRL-12569). 2. Клітина за п.1, в якій клітина людської ембріональної нирки є клітиною, що має походження від 293S. 3. Клітина за п.1, яка експресує гетерологічний протеїн. 4. Клітина за п.3, в якій гетерологічний протеїн вибрано з групи, яка складається з FVIII, BDDFVІІI, моноклонального антитіла, антитіла антиTNF, rІL4, tPA та ЕРО. 5. Клітинна лінія людини, позначена НКВ11 (депозитний номер АТСС: CRL-12568), придатна для рекомбінантного отримання терапевтичних протеїнів. 2 (19) 1 3 77642 Споріднені заявки: Заявка Чо і Чена "Vectors Having Terminal Repeat Sequence of Epstein-Barr Virus", позначена MSB-7254 [US, 09/209,915], і заявка Чо та ін. "Expression system for factor VIII", позначена MSB-7255 [US, 09/209,916], містять споріднений об'єкт винаходу. Обидві заявки були подані 10.12.1998. Цей винахід стосується, в основному, одержаних шля хом генетичної інженерії клітинних ліній ссавців для отримання біологічно активного протеїну. Зокрема, винахід стосується людських гібридних клітинних клонів, отриманих за допомогою процесу злиття людських ембріональних клітин нирки (293S) та клітин лімфоми Беркітта. Ці людські гібридні клітини можуть використовуватися для отримання гетерологічних протеїнів. До даного часу більшість терапевтичних рекомбінантних протеїнів отримувались з клітин ссавців, окрім людини. Деякими прикладами є: (dhfr-) - клітини яєчника китайського хом'яка (СНО) [Urlaub et al. // Proc. Natl. Acad. Sci. USA. – 1980. – 77. – P. 4216-4220] з селективним маркером дигідрофолат редуктази, здатним до ампліфікації [Kaufman et al. // Мої. Biol. – 1982. – 159. – P. 601-621; Gasser et al. // Proc. Natl. Acad. Sci. USA. – 1982. – 79. – P. 6522-6526], використовувались для отримання терапевтичного рекомбінантного протеїну. Різноманітність рекомбінантних терапевтичних протеїнів, відомі як такі, що одержують у клітинах ссавців, наприклад, рекомбінантний фактор VIII (rFVIII) [Kaufman et al. // J. Biol. Chem. – 1988. – 263. – P. 6352-6362], тканинний активатор плазміногену (t-PA) [US, 4766075, 1988], еритропоетин (ЕРО) [US, 4703008, 1987] і моноклональні антитіла (монАт) [US, 4816397, 1989]. Клітини (ВНК) нирки ембріону хом'яка (ВНК21) використовувалися для отримання rFVIII після селекції G418 та метотрексатної (МТХ) ампліфікації резистентних клітин G418 [US, 4965199, 1990]. Клітини мишиної мієломи (NSO) використовувались для отримання одержаного шляхом генетичної інженерії людського антитіла анти-TNF (ЕНАТ) [US, 4816397, 1989]. Проте, ця клітинна лінія генерує протеїни, які мають особливі мишині вуглеводні структури, які не є бажаними для застосування на людях. Людська клітинна лінія Namalwa (джерело лімфоми Беркітта) використовувалась для отримання альфа-інтерферону у Wellcom Research Laboratory і для отримання проурокінази [Saton et al. // Cytotechnology. – 1996. – 18. – P. 167-185], тканинного активатора плазміногену (t-PA) [Khan et al. // Biochem. Soc. Trans. – 1995. – 23. – P. 99], колонієстимулювального фактора гранулоцитів та макрофагів [Okamoto et al. // Arch. Biochem. Biophys. – 1991. – 286. P. 562-568], інтерферонів та лімфотоксинів [Hosoi et al. // C ytotechnology. – 1991. – 5. – P. 17-34] колонієстимулювального фактора гранулоцитів [Hosoi et al. // Cytotechnology. – 4 1991. – 7. – P. 25-32] у Tokyo Research Laboratories. Проте, ці клітини дуже важко піддавалися трансфекції ДНК. Уельс та ін. [1989, Gene 81: 139-149] повідомили про використання стратегії сумісної dhfr/MTX ампліфікації для експресії функціонального протеїну С в ембріональних клітинах людської нирки (клітини 293S) повідомили [Уеллс та ін. // Gene. - 1989. - 81. - P.139-149]. Клітини 293 [Stillman et al. // Мої. Cell. Biol. - 1985. - 5. - P.20512060] відомі як ті, що утворюють великі агрегати в суспензії, зокрема, при високій концентрації кальцію (>100мкМ), яка сприяє збільшенню агрегації та зниженню життєздатності клітин [Peshwa et al. // Biotech. and Bioeng. - 1993. - 41. - P. 179-187]. Усі ці відомості наведені у даній заявці як посилання. Клони гібридизованих людських клітин на даний момент визначені як такі, що легко піддаються трансфекції електропорацією або катіонною ліпосомою і які легко адаптувати до росту в суспензійній культурі. Гетерологічні протеїни можуть експресувати низькі рівні (50-100нМ) МТХ ампліфікації в оточуючому середовищі людської клітини. Окрім того, клітини легко адаптуються до росту у вільному від сироватки середовищі. Ці клітини є продуктом злиття між ембріональними клітинами людської нирки (293S) та клітинами лімфоми Беркітта. Дивіться Фіг.1 для коментарю. Ці гібридні клони, названі НКВ, містять дефектні EBV-геноми, що походять від НН514-16, які є клітинною лінією, що походить з клітин лімфоми Беркітта, P3HR1 [Hinuma et al. // J. Virol. – 1967. – 1. – P. 1045-1051]. Клітини P3HR1 несуть неіморталізований EBV. HH514-16 є клоном P3HR1 [Hinuma et al. // J. Virol. – 1967. – 1. – P. 1045-1051], який несе неіморталізований EBV. HH514-16 втратив гетерологічну ДНК, ДНК, яка перериває латентний стан протягом процесу клонування [Rabson et al. // Proc. Natl. Acad. Sci. USA. – 1983. – 87. – P. 3660-3664]. Тому EBV НКВ клони є неморталізованим вірусом і залишається в латентному стані. НКВ є людськими гібридними клітинами хазяїна, придатними для рекомбінантного отримання терапевтичних протеїнів. Ці клітини хазяїна отримуються шляхом гібридизації різних батьківських клітинних ліній, кожна з яких має різні корисні характеристики. Клітини хазяїна, які мають корисні характеристики кожної з батьківських клітинних ліній, отримуються з клітин, одержаних в результаті гібридизації. Клітини хазяїна можна одержувати шляхом генетичної інженерії для експресії високих рівнів широко поширених протеїнів. Протеїни, які можуть бути одержані за допомогою клітин хазяїна, отриманих в результаті генетичної інженерії, включають, але не обмежуються розчинним ІСАМ-1, рекомбінантним інтерлейкіном-4 (IL-4), t-PA, ЕРО, rFVIII, і BDD-FVIII (варіанти Фактору VIII з делетованим доменом В) і похідні цих протеїнів. Фактор 5 77642 6 VIII має структур у домену А1-А2-В-АЗ-С1-С2 і синНа Фіг.1 зображено спрощений варіант деритезується як одноланцюговий поліпептид з 2351 ватизації клітин НКВ. амінокислот, із яких сигнальний пептид, що має 19 На Фіг.2 зображено фізичні карти векторів ексамінокислот, розщеплюється під час транслокації пресії, зазначених у тексті. Усі плазміди утворюу порожнину ендоплазматичного ретикулюму. Беються на основі pBR322 та містять одиницю ексручи до уваги той факт, що фактор VIII важко гліпресії dhfr. Всі гени, які кодують протеїни, що козилюється, було важко досягнути експресії висопредставляють інтерес, знаходяться під регулякого рівня (>0,2пг/кл/дн.) фактора VIII [Lind et al. // ційним контролем енхансера CMV/промотора Eur. J. Biochem. – 1995. – 232. – P. 19-27; Kaufman (CMV е/р); 5-й інтрон (MIS або CIS) розміщувався et al. // Моl. Cell. Biol. – 1989. – 9. – P. 1233-1242]. біля 5-го кінця генів, за виключенням BZLF1. ПоліЕкспресія фактора VIII у клітинах ссавців, типово Α сигнальна область, позначалася рА. Обидві плана 2-3 порядки нижча по величині ніж та, що спозміди pSH157 та pCIS25DTR містять послідовність стерігалась у решті генів з використанням подібEBV-TR (402 п.о.). них векторів та підходів. Продуктивність отриманНа Фіг.3 зображено порівняння клітинних ліній ня клітинних ліній для фактора VIII знаходилася в хазяїна для генної експресії у перехідних аналізах інтервалі 0,5-1,0од./кл/дн. (0,1-0,2пг/кл/дн.). трансфекції. Трансфекції проводилися при тих же Було продемонстровано, що В-домен фактора умовах: та ж кількість клітин 293S, 2В8 і НКВ11 VIII є несуттєвим для прокоагулянтної активності. трансфікувалася тією ж кількістю плазмідної ДНК, Використовуючи вкорочені варіанти фактора VIII, використовуючи той же трансфекційний агент. різними групами відзначалася поліпшена експреРідкі культури тканин збиралися через 2 дні після сія фактора VIII у клітинах ссавців [Lind et al. // Eur. трансфекції. Рівні отримання протеїну визначалиJ. Biochem. – 1995. – 232. – P. 19-27; Tajima et al. // ся ELISA (IgG та ІСАМ-1; виміряні як нг протеїProc. 6th Int. Symp. H.T. – 1990. – P. 51-63; US, ну/106 клітин/2 дні) і комплектом проби Coatest® 5661008, 1997]. Проте, рівень експресії варіантів (rFVIII; виміряні як міліодиниці/107 клітин/2 дні). фактора VIII зі стабільного клітинного клону залиОсобливі варіанти реалізації шався нижчим 1пг/кл/дн. Також було виявлено, що Матеріали та способи незважаючи на те, що ендогенний імуноглобулін НН514-16 були любязно надані доктором (lg) не експресувався, експресія рекомбінантного Джорджем Міллером (Єльський Університет). Кліlg з клітин хазяїна, отриманих в результаті генетитини 293S були отримані від доктора Бреда Зелечної інженерії, була високою. Протеїни, отримані з ра (Molecular Therapeutic Institute, West Heaven, клітин НКВ11, мають специфічний людський проCT). Клітини 293S є клітинами 293 (ATCC CRLфіль глікозилювання. Тому, клони є оптимальними 1573), які були адаптовані для росту в суспензійній клітинами хазяїна для отримання lg, одержаного культурі [Stillman et al. // Моl. Cell. Biol. – 1985. – 5. за допомогою генетичної інженерії, та решти про– P. 2051-2060]. теїнів. Плазміди Використовувана тут клітина, що має похоВсі вектори експресії, які використовувались в дження від лімфоми Беркітта, є клітиною лімфоми цій доповіді, головним чином, були плазмідою на Беркітта, що походить від клітини лімфоми Беркітоснові pBR322 з функціональним сегментом генної та, що має походження від іншої клітини лімфоми експресії dhfr. Фізичні карти векторів експресії опиБеркітта, або клітиною, яку отримують в результаті суються на Фіг.2. Плазміди pSH157 та pCIS25DTR мітотичного поділу будь-якої з вищезгаданих. "Помають також термінальну повторювану послідовходить від" в цьому контексті має намір включати, ність вірусу Епштейна-Барра (EBV-TR). Дивіться але не обмежуватися нормальним мітотичним заявку на патент Чо та Чена "Vectors Having поділом клітин та способами, такими як трансфекTerminal Repeat Sequence of Epstein-Barr Virus", ція, злиття клітин або іншими способами генетичпозначену MSB-7254 [US, 09/209,915], для посліної інженерії або біології клітин, які використовудовності EBV-TR. Вектор pSH131, який депоноваються для зміни клітин або отримання клітин з ний в Американській Колекції Типових Культур новими властивостями. Подібним чином, клітина АТСС 98879, можна використовува ти з метою гелюдської ембріональної нирки є клітиною людської нерації векторів експресії для вибраного протеїну, ембріональної нирки, отриманої від клітини людсьяк описано у Чо та Чена (MSB-7254). кої ембріональної нирки, отриманої з іншої клітини ELISA джерела людської ембріональної нирки, або клітиДля вимірювання секреції tlCAM-1, моноклоною, яка отримується в результаті мітотичного нального антитіла для ІСАМ-1, С92.5 [McClelland поділу будь-якої з вищезгаданих. Також клітина et al. // Proc. Natl. Acad. Sci. USA. – 1991. – 88. – P. джерела 293S є клітиною 293S, отриманою з клі7993-7997] абсорбувався на мікротитрувальних тини 293S, отриманої з іншої клітини джерела планшетах з круглим дном. Планшети закупорю293S, або клітиною, яка отримується в результаті валися обробкою фосфатно-буферним розчином мітотичного поділу будь-якої з вищезгаданих. Та(PBS), який містив 1% альбуміну з коров’ячої сикож клітина джерела 2В8 є клітиною 2В8, отримароватки (BSA), а потім витримувалися з пробами, ною з клітини 2В8, отриманої з іншої клітини джеякі містили tlCAM-1. Потім планшети промивалися рела 2В8, або клітиною, яка отримується в буферним розчином для промивання (PBS плюс результаті мітотичного поділу будь-якої з вище0.005% Tween 20) та інкубувалися з біотинільовазгаданих. Гетерологічний протеїн є протеїном, ним С78.5, другим моноклональним антитілом для який отримувався в результаті генетичної інженеіншого епітопу на tlCAM-1 [McClelland et al. // Proc. рії клітини. Natl. Acad. Sci. USA. – 1991. – 88. – P. 7993-7997]. Після промивання планшети інкубувалися з HRP 7 77642 8 стрептавідином. Потім планшети промивалися збільшувалась від 5мкг/мл до 30мкг/мл протягом буферним розчином для промивання, вводились у шестимісячного періоду відбору. Потім клітини взаємодію з тетраметилбензидином (ТМБ) і реактестувалися на чутливість до середовища, яке цію зупиняли 1N HCL. Концентрація tlCAM-1 вимістить HAT. Окремі клітинні клони (SCC) отримузначалася зчитуванням OD при 450/570нм і поріввалися з обмежувального клонування розведень нювалася зі стандартною кривою очищеного (одна клітина на лунку у 96-лункових планшетах) tlCAM-1. популяції А5, чутливої до HAT. Один з SCC, Для вимірювання секреції імуноглобуліну (lg) A5/1D7 трансфікувався pSV2neo, який має ген neo використовувалось антитіло анти-lg для покриття під контролем промотора SV40 у векторі pBR для планшету і біотинільоване антитіло анти-lg викоотримання клітин стійких до G418. Один з SCC, ристовувалось як антитіло детекції. Відома концестійких до G418 (1,5мкг/мл), названий 2В8 (депонтрація молекули lg використовувалась як станнований в Американській Колекції Типових Кульдарт. В усьому іншому спосіб залишався тим же. тур, №. CRL-12569) використовувався для злиття. Для вимірювання секреції інтерлейкіну-4 (IL-4) Приклад 1. Злиття клітин та модифікації одиантитіла анти-ІL-4 використовувались для покритничних клітинних клонів тя планшету, а біотинільоване антитіло анти-ІL-4 Злиття клітин спочатку проводилося згідно з використовувалось як антитіло детектування. Мополіетиленгліколевим (ПЕГ) способом злиття, опилекула очищеного IL-4 використовувалась як стасаним [Кеннет // Meth Enzymol. – 1979. – 58. – P. 5ндарт. 359]. П'ять мільйонів кожної з клітин 293S та 2В8 Проба для вимірювання секреції rFVIII промивалися PBS за відсутності Са ++ та Мg++ у Кількісний склад молекули rFVIII визначався за логарифмічній фазі росту (Life Technologies, Rockдопомогою реагентів з Coatest® VIII:C/4 Kit (Chroville, MD) та висівалися в одну лунку 6-лункового mogenix, Molndal, Sweden). Стандартний для США планшету, який попередньо піддавався обробці антигемофілічний фактор (фактор VIII), відомий як арахісовим аглютиніном (Sigma) (5мкг/мл). 6MEGA 1 (Office of Biologies Research and Review, лункові планшети, заповнені клітинами, центрифуBethesda, MD) використовувався як стандарт вигувалися при 400g протягом 6хв. на центрифузі мірювання для планшетів EIA/RIA А/2 (Corning, Бекмана J-6M/E (Beckman, Palo Alto, CA). Після Corning, NY), попередньо нагрітих до 37°С на Севидалення PBS з лунки, клітини вводилися у взаєлективних Нагрівальних Блоках (WVR Scientific, модію з 2мл 40% (вага/об'єм) ПЕГ (Sigma) протяSan Francisco, CA). Фактор ІХа, фактор X, фосфогом 1хв. Для контролю одна лунка не вводилась у ліпід та СаСІ2 додавалися до кожної проби і вивзаємодію з ПЕГ. Клітини промивалися три рази тримувалися протягом 10хв. для активування фак5мл PBS, який містив 5% DMSO після трьох протора X. Потім додавався хромогенний субстрат мивань PBS. Клітини інкубувалися у свіжому сере(S2222) і інкубувався протягом 10хв. для виділендовищі з добавкою 15% FBS протягом 25хв. Клітиня хромогенної групи, pNA. Ця реакція зупинялася ни висівалися на 96-лункові планшети (1,2x106 додаванням 50% оцтової кислоти. Потім вимірюклітин на планшет) з використанням свіжого серевалась колорометрична абсорбція при 405/450нм довища, яке містило G418 (1мг/мл) і HAT (Life на спектрофотометричному зчитувачі для мікроTechnology) з добавкою 15% PBS. Клітини підживпланшетів SPECTRAmax® 250 (Molecular Divices, лювалися двічі на тиждень з використанням селеSunnyvale, CA) і дані обраховувалися за допомоктивного середовища. У цьому прикладі для почагою програмного забезпечення SOFTmax® PRO, ткового відбору клітини 293S мали бажану забезпечуваного Molecular Device. характеристику відсутності чутливості до середоМодифікація НАТ-чутливої та О418вища, яке містить HAT, і подібним чином, клітини резистентної клітинної лінії лімфоми Беркітта 2В8 мали бажану характеристику стійкості до Для отримання клітин чутливи х до HAT, гіпоG418. ксантин-гуанін фосфорибозил трансферази В той час як злиті клітини вирощувались при (НGPRT)-дефіцитні клітинні лінії [Szybalska et al. // селективних умовах, змішані клітини не росли при Proc. Natl. Acad. Sci. USA. – 1962. – 48. - P. 2026тих же умовах. Через три тижні після відбору, по2034; Littlefield // Proc. Natl. Acad. Sci. USA. – 1963. чаткові популяції переносилися на більші форма– 50. – P. 568-576] відновлювалися за стандартти. Для отримання SCC двадцять початкових поним прописом, описаним Сядаком та ін. [US, пуляцій зі стабільним ростом перемішувалися та 4834975, 1989]. піддавалися обмежувальному клонуванню розвеКлітини НН514-16 (отримані від доктора Дж. день (одна клітина на лунку) з використанням сеМіллера, Єльський Університет), в яких була відлективного середовища. Дев'ятнадцять SCC висутня EBV гетерологічна ДНК [Rabson et al. // Proc. биралися з 15х96-лункових планшетів після Natl. Acad. Sci. USA. – 1983. – 87. – P. 3660-3664] обережної реєстрації індивідуальних клонів з вивводилися у взаємодію протягом 24год. з користанням мікроскопу. SCC, отримані з експе300мкг/мл етилового складного ефіру метансульрименту по злиттю, називалися клітинами НКВ фокислоти (СЕМ) (Sigma, St. Louis, MO) у середо(Гібридні клітини людської нирки і В-клітин). Сім вищі суспензії RPMI-1640 (Life Science, GaithersSCC вибиралися з трансфекційного дослідження. burg, MD) з добавкою 15% ембріональної Ці сім SCC надалі тестувалися для стабільного коров'ячої сироватки (FBS) (Hyclone, Logan, UT). отримання різних протеїнів. Один з семи SCC, Після промивання клітин середовищем, вони висіHKB11 вибирався як клітина хазяїна, якій надаєтьвалися на планшети у середовищі, яке містило 6ся перевага у ссавців для отримання гетерологічтіогуанін (6ТГ) (Sigma) (5мкг/мл) для селекції на них протеїнів (Фіг.1). HGPRT-негативних клітинах. Концентрація 6ТГ 9 77642 10 Приклад 2. Характеристика клонів НКВ Хоча всі гібридні клітини вибиралися при селективних умовах, гібридний статус підтверджувався підрахунком кількості хромосом у клітинах. Насправді, всі НКВ показали кількість модальних хромосом 90-110. Ці кількості збігалися з сумою кількостей модальних хромосом 293S та 2В8 (64 та 47, відповідно, згідно з даними АТСС). 293S [Stillman et al. // Моl. Cell. Biol. – 5. – P. 2051-2060] є суспензією, адаптованою 293 (АТСС CRL-1573). Всі гібридні клітинні клони тестувалися для ендогенної (мю та каппа) експресії lg за допомогою прямого імунофлюорисцентного аналізу з використанням фіксованих метанолом клітин для визначення того, які клітинні клони секретують ендогенний lg. У повторних експериментах протягом довшого періоду ці клітини спостерігались як такі, що є негативними для експресії мю- та каппаланцюгів на основі імунофлюорисцентного тесту (табл. 1) та ELISA (дані не зображені). дної ДНК, як передбачалося для функціональної експресії важких (гамма) та легких (каппа) ланцюгів. Для тестування рівнів секреції розчинного ІСАМ-1, клітини НКВ (5x106 клітин) трансфікувалися електропорацією з 10мкг плазмідної ДНК, як передбачалося для функціональної експресії розчинного ІСАМ-1. Рівні секреції визначалися за допомогою ELISA для IgG або ІСАМ-1. Усі 19 SCC секретували відносно високі рівні ІСАМ-1 (100500нг/мл/2дн.) та IgG (30-200нг/мл/2дн.) у тимчасовій трансфекційній пробі (табл. 2). Ці дані вказують на те, що гібридні клітини підтримують експресію трансфікованих генів lg, хоча ендогеннаекспресія гену lg припинялася. Вірус Епштейна-Барра (EBV) існує у вигляді епісом у клітинах 2В8. Усі SCC були позитивними для експресії EBNA-1 (дані не зображені), що відображає той факт, що вони позитивні для EBV. Тим не менше, було невідомо, чи все ще існ ує весь геном EBV у гібридних клітинах. Таблиця 1 Таблиця 2 Детекція генної експресії lg Клітини 293S 2В8 НКВ Генна експресія Важкий ланцюг Легкий ланцюг (мю) (каппа) Негативний Негативний Позитивний Позитивний Негативний Негативний Таблиця 1 зображує результати, які спостерігалися в імунофлюорисцентному тестуванні трьох типів клітин. Клітини повторно суспендувалися у PBS та наносилися як мазок на скляне предметне скло. Після просушування клітин предметні скельця фіксували у холодному (-20°С) метанолі протягом 5хв. Клітини забарвлювалися протягом 45хв. за допомогою анти-FITC каппа та мю ланцюгами людини (розведення 1:20) (Zymed Laboratories, Inc., So. San Francisco, CA) у зволоженій камері при 37°С. Після промивання предметних скелець PBS протягом 10хв., вони покривалися покрівним склом, використовуючи PBS/Гліцерил (1:1). Клітини досліджувалися під флуоресцентним мікроскопом (Carl Zeiss, Inc., Thornwood, NY). Особлива людська структура зв'язку сиалової кислоти, альфа(2-6)сиалілтрансфераза підтверджувалася аналізом FACS клітин НКВ з використанням FITC-зв'язаного лектину Sambucus nigra (SNA) (Sigma, St. Louis, MO). Протеїн, виділений з клітин НКВ (клон 1G2), відображав альфа(2-3) та альфа(2-6) зв'язок сиалової кислоти профілю глікозилювання, проаналізований олігосахаридним способом "відбитків пальців" (дані не зображені). Це дослідження вказує на те, що клітини НКВ, отримані злиттям клітин 293S та 2В8, зберігали особливі людські ферменти глікозилювання. Для перевірки, чи мають ці гібридні клітини функціональну активність для експресії сторонніх генів, клітини трансфікувалися векторами експресії для ІСАМ-1 та IgG ( антитіло анти-TNF). Для тестування секреції IgG, клітини НКВ (5x106 клітин) трансфікувалися електропорацією з 10мкг плазмі Тимчасові трансфекційні проби для секреції IgG та розчинного ІСАМ з клонів НКВ, які отримуються зразу після процесу клонування Клон НКВ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 Рівні секреції протеїну, нг/мл/2дн. IgG Розчинний ІСАМ-1 ~150 ~500 ~30 ~30 ~20 ~50 ~80 ~200 ~100 ~500 ~50 ~300 ~40 ~200 ~30 нз *) ~80 нз ~80 ~100 ~200 нз ~100 нз ~80 ~150 ~80 нз ~30 ~200 ~80 ~200 ~160 ~500 ~80 ~200 ~80 ~100 Примітка: - *) нз - не проводили. Тому, статус геному EBV у гібридних клітинах тестувався за допомогою трансфекції клітин фрагментом BZLF1 геному EBV, trans-активуючим геномом, що перериває латентний період. Клітини НКВ (5x106 клітин) трансфікувалися 10мкг плазмідної ДНК (pSH121) для одержання функціональної експресії BZLF1. Визначення капсидного антигену EBV (EBV-VCA) проводилося непрямою імунофлюорисценцією з використанням людської сироватки, яка містила титр анти EBV-VCA та зв'язаний FITC - кон'югований антилюдський IgG. Технічні 11 77642 12 деталі імунофлюорисцентного тесту описуються ампліфікувалися зі зростаючими концентраціями вище. Як зображено у табл. 3, фіктивно трансфіМТХ (100, 200, та 400нМ) у середовищі. Початкові ковані клітини були негативними для експресії каппопуляції 6-лункових планшетів надалі відбиралисидного антигену EBV (EBV-VC A), що вказує на те, ся для одержання найкращої популяції. Накінець, що вони були негативними для реплікації EBV. ці популяції використовувалися для клонування Тим не менше, незначний процент трансфікованих окремих виділених клітинних клонів. Як зображено клітин був позитивним для антигенної експресії. Ці в табл. 4, клітини НКВ11 були оптимальними для дані вказують на те, що гібридні клітини містять отримання високих рівнів гетерологічних людських геном EBV у латентній формі. протеїнів. Для отримання ІСАМ-1, lg та похідної IL4, клітини НКВ11 були такими ж придатними, як і Таблиця 3 клітини СНО (дані не показані). Проте, клони НКВ росли швидше, ніж клони СНО і могли легше приІндукція реплікації EBV за допомогою стосовуватися до суспензійної культури та до умов трансфекції BZLF1 відсутності сироватки. У випадку отримання BDDFVIII, клони НКВ11 показали наближено в десять раз більшу продуктивність, ніж клони СНО. ВищеЕкспресія вірусного капсидного наведені результати вказують на те, що клітини антигену Клони НКВ НКВ11 є оптимальною людською клітиною хазяїна, Фіктивна BZLF1, % придатною для експресії людських терапевтичних трансфекція протеїнів. НКВ1 Негативна 0,20 НКВ5 Негативна 1-2 Таблиця 4 НКВ7 Негативна 0,20 НКВ11 Негативна 0,20 Отримання гетеролопчного протеїна НКВ13 Негативна 0,50 з клонів НКВ НКВ17 Негативна 0,20 НКВ19 Негативна 1-2 Особлива Клітина МТХ підсипродуктивГібридні клітини адаптувалися до середовища, Протеїн хазяїна лення, нМ ність, щоне містить сироватки, двократним зменшенням пг/кл./дн. FBS у колбах для стр ушування. Після двох тижнів ІСАМ-1 НКВ11 100 101) клітини вирощувались у середовищі без FBS. Кліlg НКВ13 50 12 тини вирощувалися як невеликі агрегати у колбах BDD-FVIII НКВ11 400 102) для струшування. На противагу до клітин 293S, IL-4SA НКВ11 100 53) гібридні клітини легше пристосовувалися до росту суспензійних культур без сироватки. Гібридні кліПримітки: тини у середовищі без сироватки та альбуміну з 1) - Рівень отримання ІСАМ-1 з клонів НКВ був добавкою трансферину та інсуліну могли зберігаподібним до того, що мав місце для клітин тися протягом більше, ніж року в суспензійній 293S. Клон НКВ, який секретував ІС АМ-1, лекультурі з використанням колб для струшування. гше адаптувався до суспензійної культури, в Для порівняння рівнів секреції трансфікованих той час як клони 293S дуже важко пристосовугенних продуктів, один з клонів, НКВ11 та батьківвалися до суспензійної культури. ські клітини 293S та 2В8 трансфікували pSM98 2) - Особлива продуктивність в од./к/дн. Рівень (розчинний ІСАМ-1), pSM125 (IgG анти-TNF) і секреції BDD-FVIII був в десять раз вищим, ніж pCISF8 (rFVIII). Як зображено на Фіг.3, рівні секретой, що мав місце для клонів, виділених з кліції ІСАМ-1 та IgG були набагато вищими (приблизтин СНО, трансфікованих тим же вектором но у 10 разів) у клітин НКВ11, ніж у клітин 293S. експресії. Рівні секреції обох протеїнів з 2В8 були такими, що 3) Грам-кількість отримуваного ILне піддавались детекції. Рівні секреції rFVIII у клі4SA(T13D/R121E) була можливою через 6 мітинах НКВ11 були подібними до тих, що спостерісяців після трансфекції з використанням галися у клітин 293S. Ці дані вказують на те, що НКВ11, і в той же час забирало на декілька міефективність трансфекції клітин НКВ11 набагато сяців більше з використанням клітин СНО в краща, ніж у батьківських клітинах. одночасних експериментах з використанням Приклад 3. Модифікація стабільних клонів того ж вектору експресії та подібного способу. НКВ, які секретують ге терологічні протеїни Гібридні клони тестувалися на стабільність Клітинні лінії, отримані вищезгаданими спосоекспресії протеїнів у генній системі, здатній до бами, містили (див. Фіг.2 для фізичної карти векампліфікації (dhfr/MTX). Клітини спочатку трансфіторів експресії): кували прийнятним вектором експресії, а потім (і) клони, що секретують розчинний ІСАМ-1 трансфіковані клітини (зазвичай 106 клітин на 96(10пг/кл./дн.), отриманих з клітин НКВ11, трансфілунковий планшет) висівалися та відбираликованих pSM98 після ампліфікації у 100нΜ ΜΤΧ; ся/ампліфікувалися селективним середовищем, в (іі) окремі клітинні клони, що секретують моноякому були відсутні гіпоксантин та тимідин, але з клональне антитіло (анти-TNF) (12пг/кл./дн.), отдобавкою FBS та МТХ (50нМ). Після першого відримані з НКВ13, трансфікованих pSS125 після амбору планшетів, клітини з лунок з високою секреціпліфікації у 50нМ МТХ та обмежувального єю відбиралися та переносилися до 6-лункових клонування розведень без МТХ; планшетів. Клітини у 6-лункових планшетах надалі 13 77642 14 (ііі) окремих клітинних клонів, що секретують Хоча явище трансформації або гібридизації, вкорочений rFVIII (BDD-FVIII) (5-10мкод./кл./дн., за необхідне у більшості випадків для отримання умов відсутності сироватки), отримані з клітин стабільної клітинної лінії, може приводити до зміНКВ11, трансфікованих pCIS25DTR після ампліфінених профілів глікозилювання [Yamashita // J. кації (400нМ МТХ) та обмежувального клонування Biol. Chem. – 1989. – 264. – P. 2415-2423], було розведень без МТХ; виявлено, що НКВ11, який є соматичною гібрид(iv) клонів, які секретують ко-похідну IL-4 (сеною клітинною лінією, має типово людські фермелективний агоніст IL-4, IL-4SA; мутовані по двох нти глікозилювання, наприклад, альфа(2-3) та положеннях амінокислот T13D та R121E) альфа(2-6) сиаліл трансферази. Більше того, про(5пг/кл./дн.), отримані з НКВ11, трансфікованих теїни, отримані з трансфікованих клітин НКВ маpSH157 після МТХ ампліфікації. Клон НКВ, що ють нормальні людські моделі глікозилювання секретує IL-4SA, 1G2 використовувався для відноальфа(2-3) та альфа(2-6) зв'язків сиалових кислот. сно швидкого отримання малих кількостей протеїНа довершення клітини НКВ та гібридні людну (грамкількості). ські клітини, які мають бажані характеристики з Обговорення кожної з батьківських клітинних ліній; а саме, які Гібридні людські клітинні лінії, описані тут, вілегко вирощуються у суспензійній культурі без дображують бажані характеристики, якими волоагрегації (що спостерігалося у клітин 2В8) і полегдіють їх батьківські клітинні лінії. Клітинні лінії НКВ, шують трансфекцію та бажані характеристики секякі отримані злиттям людських ембріональних кліреції (що спостерігалося у клітин 293S). Гібридна тин нирки (або клітин, отриманих з людських ембклітинна лінія, якій надавалась перевага, НКВ11 є ріональних клітин нирки) з клітинами лімфоми Бенегативною для іммуноглобулінової генної експреркітта (або клітинами, отриманими з клітин сії, як і клітини 293S. Ця риса є корисною у випадлімфоми Беркітта), придатні для одержані клітинках, коли бажаним є отримання рекомбінантним них ліній з метою експресії гетерологічних протеїспособом моноклональних антитіл з людських клінів. тин. Відкриття того, що злиття людських клітин Початковою метою одержання гібридних клідавало в результаті клітини, які мали корисні хатинних ліній 293S та 2В8 було повторне вирішення рактеристики, слугує для заохочення подальших проблеми, пов’язаної з клітинами 293S, які мають вивчень злиття з використанням 293S та інших тенденцію до агрегації при вирощуванні у суспенклітинних ліній В-клітинного джерела, наприклад, зійній культурі (небажана характеристика). Клітини Namalwa та 6F11 для отримання нових комбінацій НКВ, що отримувалися в процесі гібридизації, вирис у гібридних клітин зі злиттям різних батьківсьрощуються у вигляді моношару при культивуванні ких клітинних ліній. у Т-колбах. Проте, ці клітини вирощуються у виВищенаведені приклади мають намір проілюсгляді клітин суспензії (не утворюють великих скуптрувати винахід, як гадають, для фахівців у цій чень) при культивуванні у вигляді суспензії. Клітигалузі будуть очевидними варіантами цього винани НКВ дуже легко піддаються трансфекції і ходу. Відповідно, вважається, що мета винаходу продуктивність набагато вища ніж у клітин 293S. повинна обмежуватися тільки приведеною нижче формулою. 15 Комп’ютерна в ерстка В. Сердюк 77642 Підписне 16 Тираж 26 прим. Міністерство осв іт и і науки України Держав ний департамент інтелектуальної в ласності, вул. Урицького, 45, м. Київ , МСП, 03680, Україна ДП “Український інститут промислов ої в ласності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюHuman hybrid cell for expressionof gene of mammals, human cell line нкв11, method for producing hybrid human cells suitable for expression of heterologic protein
Назва патенту російськоюЧеловеческая гибридная клетка для экспрессии гена млекопитающих, клеточная линия человека нкв11, способ получения гибридных человеческих клеток, пригодных для экспрессии гетерологического протеина
МПК / Мітки
МПК: C12N 5/10, C12N 5/16, C12N 5/02, C12N 5/06
Мітки: спосіб, експресії, лінія, гетерологічного, отримання, клітин, придатних, людських, протеїну, експрес, людська, гібридних, клітинна, нкв11, гену, клітина, гібридна, ссавців, людини
Код посилання
<a href="https://ua.patents.su/8-77642-lyudska-gibridna-klitina-dlya-ekspresi-genu-ssavciv-klitinna-liniya-lyudini-nkv11-sposib-otrimannya-gibridnikh-lyudskikh-klitin-pridatnikh-dlya-ekspres-geterologichnogo-protenu.html" target="_blank" rel="follow" title="База патентів України">Людська гібридна клітина для експресії гену ссавців, клітинна лінія людини нкв11, спосіб отримання гібридних людських клітин, придатних для експресїї гетерологічного протеїну</a>
Спосіб експресії мітогенного білка ендотеліальних клітин людини

Номер патенту: 13321
Опубліковано: 28.02.1997
Автори: Томас Мейсіаг, Майкл Джей, Вільсон Берджесс, Вільям Дрохан
МПК: C12N 15/24, C12N 15/74
Мітки: білка, мітогенного, експресії, людини, спосіб, ендотеліальних, клітин
Формула / Реферат:
(57) Способ экспресии митогенного белка эндотелиальных клеток человека в Escherichla coll, заключающийся в том, что выделяют мРНК из вытяжки ствола мозга человека, получают из мРНК поли(А+)мРНК, синтезируют кДНК с помощью указанной мРНК, колонируют кДНК в вектор лямбда gt10 или лямбда gt11, отбирают клоны кДН К, кодирующие митогенный белок эндотелиальных клеток путем гибридизации указанных клонов с олигонуклеотидной пробой, имеющей следующую...
Спосіб одержання чужорідного протеїну в клітинах е.соli, плазмідний вектор і трансформований штам e. coli для експресії чужорідного протеїну

Номер патенту: 61066
Опубліковано: 17.11.2003
Автори: Хорн Уве, Вендерот Рольф, Різенберг Дітер, Кнюпфер Уве, Куджау Маріан, Стріттматтер Вольфган, Мацку Зігфред
МПК: C12P 21/00, C12N 15/09, C12P 21/02, C07K 16/28, C07K 16/00, C12N 15/72, C12N 1/21
Мітки: одержання, експресії, плазмідний, клітинах, спосіб, е.соli, трансформований, штам, протеїну, вектор, чужорідного
Формула / Реферат:
1. Способ получения чужеродного протеина в клетках Е. coli, которые трансформированы с помощью плазмиды, несущей чужеродный ген с индуцируемым промотором путем ферментации высокой плотности клеток через порционную стадию и стадию порционной подпитки без какого-либо ограничения роста за счет субстратов или метаболических побочных продуктов, и выделения и очистки экспрессируемого протеина из культуральной среды, причем концентрацию...
Генно-інженерна конструкція днк, що кодує 3-фосфогліцерат дегідрогеназу, вектор експресії, клітина та спосіб одержання цільового продукту, який є серином або похідним серину.

Номер патенту: 39861
Опубліковано: 16.07.2001
Автор: Бурлінгейм Річард
МПК: C12N 15/09, C12N 15/53, C12N 9/04, C12P 13/06, C12N 1/21, C12N 1/19
Мітки: серину, експресії, цільового, дегідрогеназу, клітина, серином, похідним, продукту, днк, генно-інженерна, спосіб, вектор, конструкція, кодує, одержання, 3-фосфогліцерат
Формула / Реферат:
1. Генно-инженерная конструкция ДНК, кодирующая 3- фосфоглицерат дегидрогеназу (РGD), обладающую пониженной чувствительностью к ингибированию серином по сравнению с нативным типом PGD, характеризующаяся тем, что она кодирует PGD, которая имеет изменения на С-концевом участке от VAL 363 до TYR 410 по сравнению с PGD, выделенной из E.coli.2. Генно-инженерная конструкция по п.1, отличающаяся тем, что указанная PGD содержит С-концевую...
Ізольований фрагмент днк промотору гена синтази ацетооксикислот (ahas) для експресії генів в рослинах, вектор для трансформації рослин, спосіб високорівневого експресування гетерологічного гена в рослині та в

Номер патенту: 48951
Опубліковано: 16.09.2002
Автори: Дітріх Габріель, Сміт Джейн, Пенг Джіань Їнг.
МПК: C12N 15/82, C12N 9/88, C12N 5/10, A01H 5/00, C12N 15/09, C12N 15/60, C12N 15/29
Мітки: фрагмент, рослин, рослинах, спосіб, днк, промотору, синтази, високорівневого, експресування, рослини, ahas, трансформації, вектор, ацетооксикислот, гена, ізольований, генів, експресії, гетерологічного
Формула / Реферат:
1. Изолированный фрагмент ДНК промотора гена синтазы ацетооксикислот (AHAS) для экспрессии генов в растениях, выбранный из группы, состоящей из SEQ ID №1,SEQ ID № 2,SEQ ID № 3.2. Изолированный фрагмент ДНК по п. 1, отличающийся тем, что растение является однодольным.3. Изолированный фрагмент ДНК по п. 2, отличающийся тем, что растение представляет собой кукурузу.4. Вектор для...
Спосіб розділення дезамідованої та недезамідованої людських днказ, очищена дезамідована людська днказа, очищена недезамідована людська днказа, фармацевтична композиція (варіанти), спосіб зберігання людської днк

Номер патенту: 46693
Опубліковано: 17.06.2002
Автори: Слівковскі Мері Б., Френз Джон, Шайє Стівен Джей
МПК: C07K 1/18, C07K 14/47, C12N 9/16, C07K 1/22, A61P 11/00, A61K 38/45, C12N 15/09, C12N 9/22
Мітки: дезамідована, днказа, людської, дезамідованої, днказ, фармацевтична, недезамідована, недезамідованої, розділення, людських, очищена, варіанти, зберігання, композиція, людська, спосіб, днк
Формула / Реферат:
1. Способ разделения дезамидированной и недезамидированной человеческой ДНКазы из их смеси, заключающийся в том, что смесь разделяют с помощью триптической или тентаклической хроматографии с использованием синтетических смол.2. Способ по п. 1, отличающийся тем, что используют тентаклическую катионообменную смолу.3. Способ по п. 1, отличающийся тем, что используют иммобилизованную гепариновую смолу.4. Способ по п. 1,...
Попередній патент: Система протокового цитометра для ізоляції бажаних клітин
Наступний патент: Спосіб уловлювання благородних металів у виробництві азотної кислоти
Випадковий патент: Пристрій для дугового зварювання