Моноклональне антитіло, яке специфічно зв’язується з позаклітинним доменом ilt7 людини
Номер патенту: 97946
Опубліковано: 10.04.2012
Автори: Камоґава Юміко, Чо Мінквон, Ішида Кої, Арай Наоко







Формула / Реферат
1. Моноклональне антитіло, що специфічно зв'язується із позаклітинним доменом ILT7 людини, або фрагмент антитіла, який містить його антиген-зв'язувальну ділянку, де моноклональне антитіло містить амінокислотні послідовності згідно з будь-якою з наступних від і) до ііі), такі як CDR1, CDR2 та CDR3 у варіабельній ділянці важкого ланцюга та варіабельній ділянці легкого ланцюга:
і) CDR1 варіабельної ділянки важкого ланцюга: SDYAWN (SEQ ID NO: 58);
CDR2 варіабельної ділянки важкого ланцюга: YISYSGSTSYNPSLKSR (SEQ ID NO: 59); та
CDR3 варіабельної ділянки важкого ланцюга: SPPYYAMDY (SEQ ID NO: 60);
CDR1 варіабельної ділянки легкого ланцюга: KASQDVGTAVA (SEQ ID NO: 61);
CDR2 варіабельної ділянки легкого ланцюга: WASTRHT (SEQ ID NO: 62); та
CDR3 варіабельної ділянки легкого ланцюга: QQYSSYPLT (SEQ ID NO: 63);
іі) CDR1 варіабельної ділянки важкого ланцюга: SYWIH (SEQ ID NO: 64);
CDR2 варіабельної ділянки важкого ланцюга: RIYPGTGSTYYNEKFKG (SEQ ID NO: 65); та
CDR3 варіабельної ділянки важкого ланцюга: YPTYDWYFDV (SEQ ID NO: 66);
CDR1 варіабельної ділянки легкого ланцюга: RASQSISNYLH (SEQ ID NO: 67);
CDR2 варіабельної ділянки легкого ланцюга: YASQSIS (SEQ ID NO: 68);
CDR3 варіабельної ділянки легкого ланцюга: QQSNSWPLT (SEQ ID NO: 69);
ііі) CDR1 варіабельної ділянки важкого ланцюга: SDYAWN (SEQ ID NO: 70);
CDR2 варіабельної ділянки важкого ланцюга: YISYSGSTSYNPSLKSR (SEQ ID NO:
71);
CDR3 варіабельної ділянки важкого ланцюга: ALPLPWFAY (SEQ ID NO: 72);
CDR1 варіабельної ділянки легкого ланцюга: KASQDVGTAVA (SEQ ID NO: 73);
CDR2 варіабельної ділянки легкого ланцюга: WASTRHT (SEQ ID NO: 74); та
CDR3 варіабельної ділянки легкого ланцюга: QQYSSYPYT (SEQ ID NO: 75).
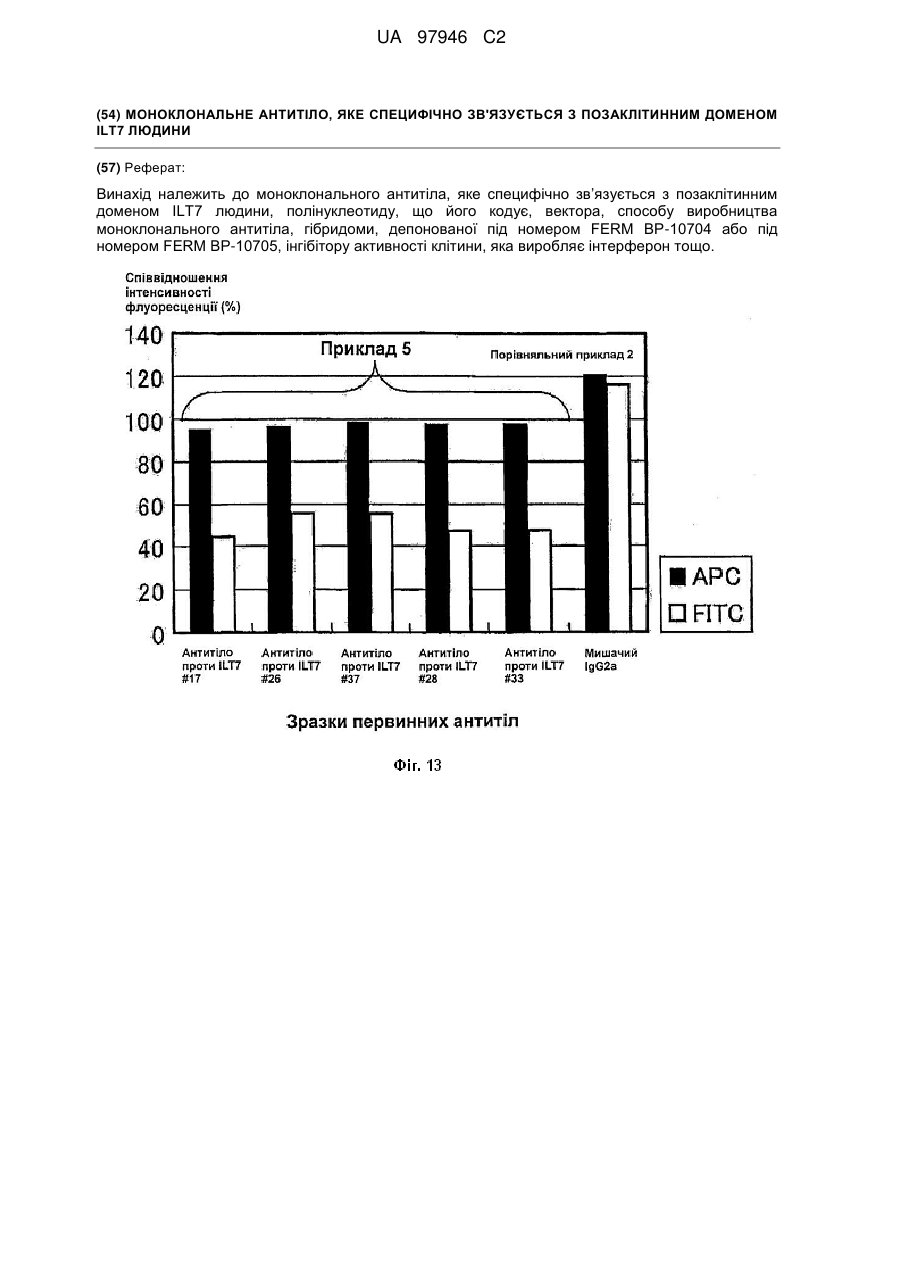
2. Моноклональне антитіло або фрагмент антитіла за п. 1, де моноклональне антитіло специфічно зв'язується з клітиною людини, що виробляє інтерферон.
3. Моноклональне антитіло або фрагмент антитіла за п. 1, де моноклональне антитіло містить зрілу послідовність амінокислотної послідовності, що вибрана з будь-якої з наступних комбінацій від (а) до (с) як варіабельна ділянка важкого ланцюга та варіабельна ділянка легкого ланцюга:
а) варіабельна ділянка важкого ланцюга SEQ ID NO: 39 та варіабельна ділянка легкого ланцюга SEQ ID NO: 41;
b) варіабельна ділянка важкого ланцюга SEQ ID NO: 43 та варіабельна ділянка легкого ланцюга SEQ ID NO: 45; та
c) варіабельна ділянка важкого ланцюга SEQ ID NO: 47 та варіабельна ділянка легкого ланцюга SEQ ID NO: 49.
4. Моноклональне антитіло, що специфічно зв'язується із позаклітинним доменом ILT7 людини, або фрагмент антитіла, який містить його антиген-зв'язувальну ділянку, де моноклональне антитіло продукується гібридомою ILT7#11, депонованою за номером доступу FERM ВР-10704, або гібридомою ILT7#17, депонованою за номером доступу FERMBP-10705.
5. Полінуклеотид, який кодує моноклональне антитіло або фрагмент антитіла за п. 1 або п. 3.
6. Вектор, який містить полінуклеотид, що кодує моноклональне антитіло або фрагмент антитіла за п. 1 або п. 3.
7. Трансформована клітина, яка утримує вектор за п. 6 у стані здатності до експресії.
8. Спосіб виробництва моноклонального антитіла або фрагмента антитіла за п. 1 або п. 3, який передбачає етапи культивування трансформованої клітини за п. 7 та одержання моноклонального антитіла або фрагмента антитіла, який містить його антиген-зв'язувальну ділянку, з цієї культури.
9. Гібридома, яка виробляє будь-яке моноклональне антитіло за п. 1 або п. 2.
10. Гібридома, яка виробляє моноклональне антитіло, яке специфічно зв'язується із позаклітинним доменом ILT7, при цьому гібридома депонована за номером доступу FERM BP-10704 або за номером доступу FERM BP-10705.
11. Спосіб виробництва моноклонального антитіла, який передбачає етапи: культивування гібридоми за п. 10 та збирання моноклонального антитіла з цієї культури.
12. Спосіб виробництва клітини, що виробляє моноклональне антитіло за п. 1, який передбачає наступні етапи:
(1) введення імунній тварині клітини, що експресує екзогенний білок, який містить позаклітинний домен ILT7 людини, та екзогенну молекулу, яка зв'язана з ILT7 людини; та
(2) вибір виробляючої антитіла клітини, яка виробляє антитіло, яке специфічно зв'язується з ILT7 людини, з виробляючих антитіла клітин імунних тварин.
13. Спосіб за п. 12, де молекула, яка зв'язана з ILT7 людини, - це білок клітинної мембрани.
14. Спосіб за п. 13, де білок клітинної мембрани - це γ-ланцюг рецептора Fc.
15. Спосіб за п. 14, де клітина, що експресує ILT7 людини та молекулу, що зв'язана з ILT7 людини, - це клітина, що утримує у стані здатності до експресії наступні (а) та (b):
(a) екзогенний полінуклеотид, що кодує амінокислотну послідовність, яка містить позаклітинний домен ILT7 людини; та
(b) екзогенний полінуклеотид, що кодує γ-ланцюг рецептора Fc.
16. Спосіб за п. 15, де клітина - це тваринна клітина.
17. Спосіб за п. 16, де клітина - це клітина, що походить від людини.
18. Спосіб за п. 17, де клітина, що походить від людини, - це клітина 293Т.
19. Спосіб за п. 12, який додатково включає етап клонування клітини, що виробляє антитіло, отриманої за способом за п. 12.
20. Спосіб виробництва моноклонального антитіла за п. 1, який передбачає етапи культивування клітини, що виробляє антитіло, за способом за п. 12 та одержання моноклонального антитіла з цієї культури.
21. Моноклональне антитіло за п. 1, яке можна отримати за наступними етапами:
(1) введення імунній тварині клітини, яка екзогенно експресує білок, який містить позаклітинний домен ILT7 людини, та молекулу, яка зв'язана з ILT7 людини;
(2) вибір виробляючої антитіла клітини, яка виробляє антитіло, яке специфічно зв'язується з ILT7 людини, з виробляючих антитіла клітин імунних тварин; та
(3) культивування вибраної на етапі (2) клітини, яка виробляє антитіла, та одержання антитіла, здатного розпізнавати ILT7 людини, з цієї культури.
22. Імуноген для виробництва антитіла за п. 1, який включає тваринну клітину, у якій утримуються у стані здатності до екзогенної експресії (а) полінуклеотид, що кодує амінокислотну послідовність, яка містить позаклітинний домен ILT7 людини, та (b) полінуклеотид, що кодує γ-ланцюг рецептора Fc; або її фракцію клітинної мембрани.
23. Імуноген за п. 22, де тваринна клітина - це клітина, що походить від людини.
24. Спосіб виявлення клітини, що виробляє інтерферон, який передбачає етапи: контактування моноклонального антитіла за п. 1 або фрагмента антитіла, який містить його антиген-зв'язувальну ділянку, із випробовуваною клітиною та
виявлення моноклонального антитіла або фрагмента антитіла, який містить його антиген-зв'язувальну ділянку, зв'язаного з клітиною.
25. Реагент для виявлення клітини, що виробляє інтерферон, який містить моноклональне антитіло за п. 1 або фрагмент антитіла, який містить його антиген-зв'язувальну ділянку.
26. Спосіб інгібування in vitro активності клітини, що виробляє інтерферон, який передбачає етап контактування будь-якого з наступних компонентів з клітиною, яка виробляє інтерферон:
(а) моноклонального антитіла за п. 1, що інгібує активність клітини, яка виробляє інтерферон, або фрагмента антитіла, який містить його антиген-зв'язувальну ділянку;
(b) імуноглобуліну, у який уведена гіперваріабельна ділянка моноклонального антитіла, описаного у (а), або фрагмента імуноглобуліну, який містить його антиген-зв'язувальну ділянку.
27. Спосіб інгібування у живому організмі активності клітини, яка виробляє інтерферон, який передбачає етап введення живому організму будь-якого з наступних компонентів:
(a) моноклонального антитіла за п. 1, що інгібує активність клітини, яка виробляє інтерферон, або фрагмента антитіла, який містить його антиген-зв'язувальну ділянку;
(b) імуноглобуліну, у який уведена гіперваріабельна ділянка моноклонального антитіла, описаного у (а), або фрагмента імуноглобуліну, який містить його антиген-зв'язувальну ділянку; та
(c) полінуклеотиду, що кодує будь-який компонент, описаний у (а) або (b).
28. Спосіб за п. 26 або 27, де активність клітини, яка виробляє інтерферон, є наслідком або активності клітини, що виробляє інтерферон, або виживання клітини, що виробляє інтерферон, або їх обох.
29. Інгібітор активності клітини, яка виробляє інтерферон, який містить будь-який з наступних компонентів як активний інгредієнт:
(a) моноклональне антитіло за п. 1, що інгібує активність клітини, яка виробляє інтерферон, або фрагмент антитіла, який містить його антиген-зв'язувальну ділянку;
(b) імуноглобулін, у який уведена гіперваріабельна ділянка моноклонального антитіла, описаного у (а), або фрагмент імуноглобуліну, який містить його антиген-зв'язувальну ділянку;
(c) полінуклеотид, що кодує будь-який компонент, описаний у (а) або (b).
30. Інгібітор активності клітини, яка виробляє інтерферон, за п. 29, де активність клітини, яка виробляє інтерферон, є наслідком або активності клітини, що виробляє інтерферон, або виживання клітини, що виробляє інтерферон, або їх обох.
Текст
Реферат: Винахід належить до моноклонального антитіла, яке специфічно зв’язується з позаклітинним доменом ILT7 людини, полінуклеотиду, що його кодує, вектора, способу виробництва моноклонального антитіла, гібридоми, депонованої під номером FERM BP-10704 або під номером FERM BP-10705, інгібітору активності клітини, яка виробляє інтерферон тощо. UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 Галузь техніки Цей винахід стосується антитіла, що зв'язується з ILT7 людини. Попередній рівень техніки Відомо, що інтерферон (IFN далі у цьому описі "інтерферон" скорочується як IFN) та інтерферон (IFN) є інтерферонами типу 1, яким притаманна противірусна активність або протипухлинна активність. З іншого боку, було також виявлено, що IFN пов'язаний з автоімунними хворобами. Наприклад, повідомлялося про аномальне виробництво IFN у пацієнтів з переліченими далі автоімунними хворобами. Також припустили, що симптоми автоімунних хвороб можна зменшити шляхом нейтралізації IFN. Системний червоний вовчак (Shiozawa et al., Arthr. & Rheum. 35, 412, 1992). Хронічний ревматизм (Hopkins et al., Clin. Exp. Immunol. 73, 88, 1988). Повідомлялося про випадки, при яких симптоми автоімунних хвороб були явними або загострювалися внаслідок введення рекомбінантного IFN2 або IFN (Wada et al., Am. J. Gastroenterol. 90, 136,1995; Perez et al., Am. J. Hematol. 49, 365, 1995; Wilson LE et al., Semin Arthritis. Rheum. 32, 163-173, 2002). Крім того, було також виявлено, що IFN індукує диференціювання дендритних клітин. Дендритна клітина - це також клітина з присутнім антигеном. Отже, вважають, що індукція диференціювання дендритних клітин складає важливий механізм в автоімунних хворобах. Припустили, що існує глибокий зв'язок між індукцією диференціювання дендритних клітин IFN та початком системного червоного вовчака (Blanco et al., Science, 16:294, 1540-1543, 2001). Отже, було вказано нате, що IFN є тісно пов'язаним з протипухлинною активністю, так само, як і з автоімунними хворобами. Крім того, IFN глибоко залучається у початок псоріазу (Nestle FO et аl., J. Exp. Med. 202, 135-143, 2005). Клітини, що виробляють інтерферон (ІРС), ідентифікували як клітини, що виробляють IFN типу 1 у великих кількостях у зв'язку з вірусною інфекцією. Невелика кількість ІРС є присутньою у крові. Вважають, що лімфоцити периферійної крові становлять 1% або менше ІРС. Проте, ІРС мають дуже велику здатність виробляти IFN. Здатність ІРС виробляти IFN сягає, наприклад, 4 3000 пг/мл/10 клітин. Тобто, можна говорити про те, що більша частина IFN або IFN у крові, що виробляються під час вірусної інфекції, є результатом ІРС, незважаючи на невелику кількість таких клітин. З іншого боку, ІРС - це недиференційовані лімфоїдні дендритні клітини, яких вважають клітинами-попередниками дендритних клітин. ІРС можна назвати плазмоцитоїдними дендритними клітинами. ІРС диференціюються у дендритні клітини внаслідок стимуляції вірусом та індукують виробництво IFN або IL-10 Т-клітинами. ІРС також диференціюються у дендритні клітини внаслідок стимуляції IL-3. Диференційовані внаслідок стимуляції IL-3 дендритні клітини індукують виробництво цитокіну Th2 (IL-4, IL-5 та IL-10) Т-клітинами. Отже, ІРС мають властивості, що дозволяють їм диференціюватися у певні дендритні клітини шляхом різних стимуляцій. Отже, ІРС мають два профілі: клітини, що виробляють IFN, та клітини-попередники дендритних клітин. Обидва типи клітин відіграють важливу роль в імунній системі. Іншими словами, ІРС - це одна з важливих клітин, яка підтримує імунну систему у різних аспектах. Непатентний документ 1: Shiozawa et al., Arthr. & Rheum. 35, 412, 1992. Непатентний документ 2: Hopkins et al., Clin. Exp. Immunol. 73, 88, 1988. Непатентний документ 3: Wada et al., Am. J. Gastroenterol. 90, 136, 1995. Непатентний документ 4: Parez et al., Am. J. Hematol. 49, 365, 1995. Непатентний документ 5: Bianco et al., Science, 16:294, 1540-1543, 2001. Непатентний документ 6: Ju et al., Gene. 2004 Apr 28; 331: 159-64. Непатентний документ 7: Colonna Μ et al., Seminars in Immunology 12: 121-127, 2000. Непатентний документ 8: Nakajima Η. et al., J. Immunology 162: 5-8, 1999. Непатентний документ 9: Wilson LE et al., Semin Arthritis. Rheum. 32, 163-173, 2002. Непатентний документ 10: Nestle FO et al., J. Exp. Med. 202, 135-143, 2005. Патентний документ 1: WO03/12061 (опублікована заявка на патент США №. 2003-148316). Суть винаходу Задачі, що вирішуються винаходом Завданням цього винаходу є забезпечення антитілом, що зв'язується з імуноглобулінподібним транскриптом-7 (ILT-7), та виявлення, ідентифікування або виділення ІРС. Іншим завданням цього винаходу є регулювання активності ІРС. Засоби для вирішення задач Для регулювання активності гуморального фактору, такого як IFN, ефективним є введення антитіл, що розпізнають цей фактор. Наприклад, було здійснено спробу лікувати автоімунні 1 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 хвороби антитілами проти інтерлейкіну IL-1 або IL-4 (Guler et al., Arthritis Rheum., 44. S307, 2001). Крім того, припускають, що нейтралізуючі антитіла можуть служити як терапевтичні агенти автоімунних хвороб, як у випадку з інтерфероном (Stewart, ТА. Cytokine Growth Factor Rev. 14; 139-154, 2003). Можна передбачити, що такий саме підхід, як описано вище, є ефективним у випадку з інтерфероном, що виробляється ІРС. Проте, такий підхід засновується на інгібуванні ефекту гуморального фактору після виробництва цього фактору. Якщо можна безпосередньо контролювати виробництво бажаного гуморального фактору, тоді можна досягти більш суттєвих терапевтичних ефектів. Повідомлялося про антитіла, що розпізнають ІРС людини. Наприклад, анти-ВDСА-2 моноклональне антитіло - це ІРС-специфічне моноклональне антитіло людини (Dzionek А. et al. J. Immunol. 165: 6037-6046, 2000). Визначають, що анти-ВSDСА-2 моноклональне антитіло є ефективним при інгібуванні виробництва інтерферону ІРС людини (J. Ехр. Med. 194: 1823-1834, 2001). Крім того, також повідомлялося, що моноклональні антитіла, які розпізнають мишачі клітини, що виробляють інтерферон, інгібують виробництво інтерферону (Blood 2004 Jun 1; 103/11: 4201-4206.Epub 2003 Dec). Повідомлялося, що зменшення кількості дендритних клітин було наслідком дії моноклональних антитіл проти плазмоцитоїдних дендритних клітин у мишей (J. Immunol. 2003, 171: 6466-6477). Подібно до цього, якщо є антитіла, які розпізнають ІРС людини та можуть регулювати їх активність, це може бути корисним. Наприклад, автори цього винаходу вже продемонстрували, що антитіло, яке розпізнає Ly49Q, специфічно зв'язується з ІРС мишей. Проте, антитіло проти Ly49Q не впливає на активність ІРС мишей (Blood, I April 2005, Vol. 105, No. 7, та pp. 2787-2792.; WO2004/13325). З іншого боку, відомо, що ILT7 є молекулою, специфічна експресія якої спостерігається у плазмоцитоїдних дендритних клітинах (Ju XS et al. and Gene. 2004 Apr 28; 331: 159-64.; WO03/12061). Проте, не було отримано жодних антитіл проти ILT7. Отже, вплив антитіл на ІРС є також невідомими. ILT7 - це мембранний білок, що містить імуноглобулін-подібний мотив. Про нього повідомляли, як про одну з молекул, що експресуються у клітинах мієлоїдної системи або лімфатичної системи (Colonna Μ et al., Seminars in Immunology 12:121-127, 2000). Численні молекули, структура яких є аналогічною до ILT7, називаються родиною ILT. Родина ILT є також структурно подібною до інгібіторних рецепторів клітин-кілерів (KIR). ILT7 має чотири імуноглобулін-подібні домени С-типу, як і інші молекули родини ILT. Вважають, що ILT7 посилає сигнали активації у клітини, як і у випадку з ILT1, ILT1-подібним білком, ILT8 та LIR6a. Було підтверджено, що молекула, яка належить до родини ILT, експресується у клітинах кровотворної системи (Young et al., Immunogenetics 53: 270-278, 2001; "The KIR Gene Cluster", Carrington, Mary and Norman, Paul. Bethesda (MD): National Library of Medicine (US), NCBI; 2003). Далі, високу експресію ILT7 визначили у плазмацитоїдних дендритних клітинах (PDC), а низьку експресію ILT7 визначили у моноцит-похідних дендритних клітинах (MDDC) за допомогою віднімальної гібридизації. ILT2 та ILT3 експресувалися не тільки у PDC, а також у DC (дендритних клітинах), отриманих з MDDC- або CD34- позитивних клітин. Проте, оскільки мРНК у ILT7 специфічно експресувалася у PDC, визначили, що мРНК може служити як маркер PDC. Крім того, визначили, що у той самий час експресія ILT7 зменшувалася внаслідок зменшення стимуляції CpG (Ju XS et al. Gene. 2004 Apr 28; 331: 159-64.; WO03/12061). Цей винахід доводить, що специфічній експресії ILT7 у ІРС сприяло дослідження на ІРС людини. Отже, автори цього винаходу зробили спробу отримати антитіла до ILT7 та пояснити ці ефекти. Наприклад, молекули, що складають родину ILT, такі як ILT2 та ILT3, мають високу консервативність, особливо в амінокислотних послідовностях позаклітинних доменів (Фіг. 9). Ці родини ILT демонструють характерні профілі експресії у різних клітинах крові, відповідно. Отже, дуже важливо отримати антитіло, яке може імунологічно розрізняти між іншими молекулами родини ILT та ILT7. Проте, фактично, було важко отримати антитіло, що зв'язується специфічно з ІРС людини, застосовуючи при цьому ILT7 як імуноген, що є наслідком перешкод, описаних нижче. Взагалі, білок, що виробляється шляхом технології генної рекомбінації, застосовується як імуноген для того, щоб отримати антитіло, що розпізнає невелику кількість білків, що походять з живих організмів. У цьому винаході зроблена спроба експресувати ILT7 людини на основі інформації послідовності основ кДНК ILT7 людини, яку вже знайшли, та амінокислотної послідовності, що кодується послідовністю основ (GenBank Accession No. NM_012276). Проте, авторам цього винаходу не вдалося отримати ILT7 людини як рекомбінант за нормальних умов. Часткову амінокислотну послідовність природного білка часто намагаються застосовувати як імуноген з метою отримання антитіла білка. Проте, існує мало амінокислотних послідовностей, що є специфічними до ILT7 людини у білках, оскільки гомологія амінокислотних 2 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 послідовностей є надзвичайно високою у родині ILT. Крім того, необхідно вибирати ділянку, що складається з частини, яка розпізнається як епітоп антитілами на поверхні клітин для того, щоб дозволити антитілам розпізнавати молекули на поверхні клітин. Отже, вважали, що утворення антитіла, що є специфічним до ILT7, застосовуючи фрагмент амінокислотної послідовності як імуноген, є неможливим. Автори цього винаходи показали, що антитіло, яке зв'язується з ІРС, можна отримати, застосовуючи спеціальний імуноген за таких умов. Крім того, автори цього винаходу визначили, що отримане таким способом антитіло специфічно розпізнає ІРС людини та, крім того, має ефект регулювання активності, таким чином їм вдалося виконати цей винахід. Отже, цей винахід стосується наступного антитіла проти ILT7, способу його виробництва та його застосування. Ефекти винаходу Цим винаходом пропонується імуноген, що є корисним при виробництві антитіла, яке розпізнає ILT7 людини, та спосіб виробництва антитіла проти ILT7 людини, при якому застосовується цей імуноген. ILT7 - це мембранний білок, що належить до родини ILT. Зокрема, амінокислотна послідовність позаклітинної ділянки є надзвичайно консервативною серед родини ILT. Отже, надзвичайно важко отримати антитіло, що розрізняє представників родини ILT, застосовуючи загальні способи імунізації. Автори цього винаходу показали, що антитіло, яке розпізнає ILT7 людини, можна легко отримати, застосовуючи тваринні клітини, у яких ILT7 експресується разом з білком клітинної мембрани. Антитіло проти ILT7, яке можна отримати, застосовуючи цей винахід, має високу специфічність, яка дозволяє відрізняти клітини, що експресують інші родини ILT, від тих, що експресують ІРС людини. У переважному варіанті здійснення винаходу антитіло проти ILT7 людини, запропоноване цим винаходом, зв'язується з ІРС людини. Крім того, антитіло цього винаходу специфічно розпізнає ІРС людини. Отже, воно є корисним при виявленні та виділенні ІРС. ІРС - це клітина, що виробляє більшу частину інтерферону типу 1. Отже, таке виявлення та виділення є важливими для постановки діагнозу та вивчення хвороб, які залучають ІРС, таких як автоімунні хвороби. Зокрема, згідно з даними авторів цього винаходу експресія ILT7 у ІРС не знижується у присутності IFN. Експресія IFN часто легко відбувається у пацієнтів з автоімунними хворобами. Це означає, що антитіло проти ILT7 цього винаходу можна застосовувати для виявлення та виділення ІРС у пацієнтів з автоімунними хворобами, у яких відбувається експресія IFN. Антитіло проти ILT7, запропоноване цим винаходом, має ефект, який регулює активність ІРС людини у переважному варіанті здійснення цього винаходу. Отже, антитіло проти ILT7 цього винаходу можна застосовувати для інгібування активності ІРС. Як описано раніше, експресія ILT7 у ІРС не зменшується у присутності IFN. Отже, якщо застосовується інгібування активності ІРС антитілом цього винаходу, можна очікувати терапевтичний ефект у пацієнтів з автоімунними хворобами, у яких спостерігається експресія IFN. Невелика кількість ІРС виробляє велику кількість інтерферону. Для нейтралізації інтерферону необхідно стільки антитіл, скільки є молекул інтерферону. На відміну від цього, згідно з цим винаходом безпосередньо інгібується активація клітин, які виробляють інтерферон. Внаслідок цього можна очікувати сильний інгібіторний ефект щодо інтерферону, навіть якщо застосовується менша кількість антитіл порівняно з нейтралізацією антитілом проти IFN. Крім того, у випадку, коли інтерферон виробляється безперервно, вважають, що нейтралізація антитілами проти інтерферону є тимчасовим інгібуванням. У цьому винаході, оскільки інгібується активність ІРС, ефект інгібування виробництва інтерферону можна очікувати протягом тривалого періоду часу. Стислий опис ілюстративних матеріалів Фіг. 1а - це фотографія, на якій експресія мРНК гена ILT7 досліджується способом полімеразної ланцюгової реакції зі зворотною транскриптазою (RT-PCR). Це результат аналізу експресії мРНК гена ILT7 в імуноцитах людини. Фіг. 1b - це діаграма, на якій експресія мРНК гена ILT7 у різних тканинах та клітинах людини порівнюється та досліджується із застосуванням способу кількісної полімеразної ланцюгової реакції (PCR). Горизонтальна вісь представляє клітини та тканини, що досліджуються, а вертикальна вісь демонструє рівень експресії ILT7, який стандартизується згідно з рівнем експресії гена GAPDH. Фіг. 2 - це діаграма, що демонструє структури білка ILT7, де Фіг. 2(а) демонструє амінокислотну послідовність білка ILT7 і також демонструє визначену сигнальну послідовність секреції та трансмембранний домен на зображенні, та 3 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 Фіг. 2(b) демонструє схематичну діаграму білків ILT7, які кодуються побудованими векторами експресії. Фіг. 3 - це діаграма, що демонструє результати дослідження за допомогою FCM того, що вектор експресії ILT7 та вектор експресії FcR ввели у клітини, та поверхнево-клітинної експресії молекул ILT7. Горизонтальна вісь представляє інтенсивність флюоресценції, визначену в антитілі проти FLAG, а саме, інтенсивність поверхнево-клітинної експресії молекул ILT7, до яких мітка FLAG була приєднана, а вертикальна вісь представляє кількість клітин. Фіг. 4 демонструє фотографію, де вектор експресії ILT7 та вектор експресії FcR ввели у клітини та пов'язані молекули проаналізували шляхом імунопреципітації та вестерн-блотингу. Діаграми з лівого боку демонструють результати того, що молекула ILT7 зазнала блотингу антитілом проти FLAG після імунопреципітації молекули FcR антитілом проти myc (зображення зверху), а молекула FcR зазнала блотингу антитілом проти myc (зображення знизу). Аналогічно, діаграми з правого боку демонструють результати того, що молекула ILT7 зазнала блотингу антитілом проти FLAG після імунопреципітації молекули FcR антитілом проти FLAG (зображення зверху), а молекула FcR зазнала блотингу антитілом проти myc (зображення знизу). Фіг. 5 - це фотографія, де глікозилування молекули ILT7 досліджували шляхом введення вектора експресії ILT7 та вектора експресії FcR у клітину та обробки N-глікозидазою. Лівий бік діаграми демонструє розмір ILT7 у випадку, коли ILT7 не обробляли N-глікозидазою, а правий бік фотографії демонструє розмір ILT7 у випадку, коли виконували обробку N-глікозидазою. Фіг. 6а - це діаграма, яка демонструє дослідження реактивності отриманого моноклонального антитіла проти ILT7 шляхом аналізу FCM. (а) демонструє результат того, що зв'язування антитіла проти ILT7 з фракцією BDCA-2-позитивних ІРС проаналізували, застосовуючи лімфоцити периферійної крові людини та подвійне забарвлення антитілом проти ILT7 та антитілом проти BDCA-2. Вертикальна вісь демонструє реактивність до антитіла проти BDCA-2, а горизонтальна вісь демонструє реактивність до кожного з отриманих антитіл проти ILT7. Фіг. 6b - це діаграма, яка демонструє результати дослідження реактивності отриманих моноклональних антитіл проти ILT7 за допомогою FCM-аналізу. (b) демонструє результат дослідження зв'язування антитіла проти ILT7 з молекулою ILT7 із застосуванням клітин 293Т, у які ввели вектори експресії ILT7 та FcR. Вертикальна вісь демонструє реактивність антитіла проти FLAG, а саме інтенсивність експресії молекул ILT7, до яких приєднали мітку FLAG, а горизонтальна вісь демонструє реактивність відповідних антитіл проти ILT7. Фіг. 7 - це діаграма, що демонструє результати дослідження отриманих моноклональних антитіл проти ILT7, реактивності двох клонів до лімфоцитів периферійної крові людини за допомогою FCM-аналізу. Три графіки зліва демонструють результати #11, та три графіки справа демонструють результати #17. На діаграмах з лівого боку кожна вісь з відміткою ILT7 демонструє реактивність ILT7 #11. Подібно до цього, на діаграмах з правого боку кожна вісь з відміткою ILT7 демонструє реактивність ILT7 #17. Фіг. 8 - це результат порівняння та дослідження зв'язувальної активності отриманих моноклональних антитіл ILT#11 та ILT# 17 проти ILT7 з лімфоцитами людини зі зв'язувальною активністю антитіла проти BDCA-2. Вертикальна вісь демонструє реактивність антитіла проти CD123, а горизонтальна вісь демонструє реактивність кожного антитіла. Тобто, кожне антитіло зв'язується з частиною CD123 позитивної клітини. Це діаграма, що демонструє результати аналізу реактивності, де клітини-лімфоцити стимулювали двома типами CpG та IFN. Фіг. 9а - це діаграма, що демонструє амінокислотні послідовності молекул родини з високою гомологією молекул ILT7. Кожну амінокислотну послідовність зовнішньо-клітинної ділянки здебільшого зображено як результат порівняльного аналізу, Фіг. 9b - це продовження Фіг. 9а, а Фіг. 9с - це продовження Фіг. 9b. Фіг. 10 - це результат дослідження реактивності отриманих моноклональних антитіл ІLT#11 та ILT7#17 проти ILT7 до молекул ILT1, ILT2 та ILT3, при якому застосовувалися клітини, у які ввели їх вектори експресії. Верхня діаграма демонструє результати, де знов підтвердилася реактивність до клітин, у яких молекули ILT7 з міткою FLAG експресувалися разом з FcR. Нижня діаграма демонструє результати дослідження реактивності до клітин, у які ввели ILT1, ILT2, ILT3 та FcR (ліва діаграма: ILT7#11, права діаграма: ILT7#17). Горизонтальна вісь демонструє реактивність кожного антитіла проти ILT7. Фіг. 11 - це діаграма, що демонструє вплив отриманих моноклональних антитіл ILT7#11 та ILT7#17 проти ILT7 на інтерферогенну спроможність лімфоцитів людини. На діаграмі горизонтальна вісь демонструє концентрацію інтерферону- у культуральній надосадовій рідині, коли лімфоцити людини стимулювали вірусом грипу, а вертикальна вісь показує 4 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 оброблені антитіла. Термін "без інфекції" вказує на результати з клітинами, які не стимулювали вірусом грипу. Фіг. 12 - це діаграма, що демонструє активність CDC отриманих моноклональних антитіл ILT7#37, ILT7#28 та ILT7#33 проти ILT7. Навіть коли застосовували моноклональні антитіла проти ILT7, отримані з гібридоми, 80% або більше активності CDC спостерігали при концентрації антитіл 0,1 мкг/мл або більше. У випадку з антитілами, відмінними від моноклонального антитіла проти ILT7, активність CDC до клітин-мішеней не спостерігали. Фіг. 13 - це діаграма, що демонструє інтерналізацію до клітин-мішеней отриманих моноклональних антитіл ILT7#17, ILT7#26, ILT7#37, ІLT#28 та ІLT7#33 проти ILT7. Інтенсивність флюоресценції антигенпрезентувальної клітини (АРС) - це індикатор кількості імунного комплексу ILT7 - антитіло проти ILT7, який був присутнім на поверхні клітин перед інкубацією, та вона визначається незважаючи на те, чи є присутнім на поверхні клітини-мішені імунний комплекс ILT7 - антитіло проти ILT7, чи його було включено у клітину після інкубації. З іншого боку, інтенсивність флюоресценції FITC - це індикатор кількості імунного комплексу ILT7 - антитіло проти ILT7, яка залишилася на поверхні клітин після інкубації. Тобто, інтенсивність флюоресценції FITC зменшується шляхом інтерналізації. Переважні варіанти здійснення винаходу Повідомлялося про те, що ILT7 людини (імуноглобулін-подібний транскрипт-7) - це молекула, яка специфічно експресується у плазмацитоїдних дендритних клітинах (Gene. 2004 Apr 28; 331:1 59-64.; WO03/12061). Альтернативно, також відомо, що ILT7 людини можна застосовувати як індикатор-попередник для прогнозування лімфоми (WO2005/24043). Проте, способу отримання антитіла, здатного розпізнавати ILT7 людини, ще було не розроблено. ILT7 людини складається з 499 амінокислотних залишків, як зображено на SEQ ID NO: 2, та він є трансмембранним білком, що включає чотири імуноглобулін-подібні домени у структурі та одну трансмембранну ділянку (445-466; від 429 до 450 у SEQ ID NO: 2). Серед 444 амінокислотних залишків, що включають N-закінчення, 16 амінокислотних залишків (від -15 до 1, у SEQ ID NO: 2) є сигнальними послідовностями, а від 17 до 444 амінокислотного залишку (від 1 до 428 у SEQ ID NO: 2) складають позаклітинний домен. З іншого боку, ділянка Сзакінчення - це внутрішньоклітинний домен. Більшість частин ILT7 людини - це позаклітинні домени, а 33 амінокислотні залишки складають внутрішньоклітинний домен (від 467 до 499, від 451 до 483 у SEQ ID NO: 2). Не передбачається, що мотив, який залучається у сигналізацію, є присутнім у внутрішньоклітинному домені. Амінокислотну послідовність повної довжини ILT7 людини зображено на SEQ ID NO: 2, а послідовність основ кДНК, що кодує амінокислотну послідовність, зображено на SEQ ID NO: 1. Тут: кодувальні ділянки зрілого пептиду (72)..(1520), зображені на SEQ ID NO: 1, не включають кодони термінації та ініціації. Тобто, послідовності, що кодують білок та включають кодони термінації та ініціації у SEQ ID NO: 1 - це від 24 до 1523. Вважають, що сигнал ліганду передається клітинам у зв'язку з ILT7 людини молекулою трансдукції сигналу. Наприклад, більшість -ланцюгів Fc рецептора є присутніми у клітинах. Крім того, внутрішньоклітинний домен містить мотив активації на основі тирозину імунорецептора (ІТАМ), який залучається у сигналізацію. ІТАМ - це частина амінокислотної послідовності, яку звичайно можна спостерігати у молекулах-адапторах, які пов'язуються з імунорецепторами, такими як рецептори Fc. Мотив, такий як YxxL (SEQ ID NO: 76), який є мішенню фосфорилування тирозину, включається у ІТАМ, і сигнал передається шляхом фосфорилування. Відомі приклади молекули трансдукції сигналу, яка включає ІТАМ у внутрішньоклітинному домені, окрім -ланцюга рецептора Fc, включають CD3 та DAP 12. Серед цих молекул трансдукції сигналу передбачається, що молекула, пов'язана з ILT7 людини, буде -ланцюгом рецептора Fc. На цей час не знайдено ліганду, який зв'язується з ILT7 людини. Автори цього винаходу підтвердили шляхом аналізу генної експресії, що ILT7 специфічно експресується у ІРС людини. Автори цього винаходу вважали, що може бути корисним досліджувати ІРС, якщо можна отримати антитіло, здатне імунологічно відрізняти ILT7 людини від інших молекул. Проте, у родині ILT, що включає ILT7, існує багато молекул з подібними структурами. Молекули, такі як ILT1, ILT2, ILT3, ILT4, ILT5, ILT6 або LIR-8 містять високо гомологічні амінокислотні послідовності, особливо у своїх позаклітинних доменах. Отже, автори цього винаходу вважали, що дуже важко отримати антитіло, здатне розрізняти ці молекули, застосовуючи пептид домену, який включає часткову амінокислотну послідовність, що утворює позаклітинний домен, як імуноген. Отже, автори цього винаходу намагалися отримати антитіло проти ILT7 людини, застосовуючи клітини, що експресують ILT7 людини як імуногени. Проте, застосування загальних векторів експресії не викликало експресію кДНК ILT7 людини у тваринних клітинах. Повідомлялося, що молекула ILT1, що має структуру, дуже подібну до ILT7, пов'язується з -ланцюгом рецептора Fc. Тобто, коли клітини, у яких експресувався 5 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 ланцюг рецептора Fc, такі як клітини RBL (базофільного лейкозу щурів) та клітини Р815 (мастоцитоми мишей), застосовувалися як клітини-хазяїни, спостерігали експресію ILT1 на поверхні цих клітин. Проте, якщо ILT1 примушували експресуватися у клітинах 293, у яких ланцюг рецептора Fc первинно не експресується, експресію на поверхні клітин не спостерігали. З іншого боку, було продемонстровано, що експресію ILT1 на поверхні клітин можна підтвердити, коли ILT1 експресується разом з -ланцюгом рецептора Fc (Nakajima H. et al., J. Immunology 162:5-8.1999). Проте, не існує інформації про імуноген для виробництва антитіл до ILT7. Наприклад, у цьому повідомленні клітини RBL, у які вводять ген ILT1, застосовуються як імуноген для отримання антитіл до ILT1. Автори цього винаходу намагалися отримати антитіла до ILT7, застосовуючи комбінацію клітин RBL з геном ILT7 таким самим способом, як описано. Проте, навіть якщо ILT7 примушували експресуватися у клітинах RBL (P815), експресію ILT7 на поверхні клітин не спостерігали, та внаслідок цього його не можна застосовувати як імуноген. Автори цього винаходу здійснили цільове дослідження з метою отримати антитіло, здатне розпізнавати ILT7 людини. Внаслідок цього автори цього винаходу визначили, що бажане антитіло можна отримати, застосовуючи специфічну трансформовану клітину як імуноген, та тим самим виконали цей винахід. Тобто, цей винахід стосується моноклонального антитіла, яке зв'язується з позаклітинним доменом ILT7 людини, та стосується фрагменту, що включає його антиген-зв'язувальну ділянку. У цьому винаході ILT7 людини можна визначити як природну молекулу, що експресується у ІРС людини, або як молекулу, яка є імунологічно еквівалентною до ILT7, що експресується у ІРС людини. У цьому винаході зв'язування антитіл з ILT7 людини можна підтвердити, наприклад, наступним способом. Підтвердження на основі реактивності до клітин людини Згідно з даними авторів цього винаходу специфічну експресію ILT7 людини спостерігали у ІРС людини. Спочатку ILT людини виділили як ген, експресію якого спостерігають у плазмацитоїдних дендритних клітинах (Blood. 2002 100; 3295-3303, Gene. 2004 Apr 28; 331:15964). Крім того, також відомо, що його можна застосовувати як маркер плазмацитоїдних дендритних клітин (WO03/12061). Припускають, що плазмацитоїдні дендритні клітини та ІРС є здебільшого ідентичними популяціями клітин, або їх великі частини є однаковими. Отже, не існує жодних протиріч між цими повідомленнями та даними авторів цього винаходу. З точки зору такого профілю експресії ILT7 людини, по-перше, зв'язувальна активність ІРС або плазмацитоїдних дендритних клітин з принаймні певною субпопуляцією - це одна з важливих характеристик антитіла, що зв'язується з ILT людини у цьому винаході. Маркери поверхні клітин, специфічні до відповідних клітинних популяцій, можна застосовувати для визначення, чи є певна клітина ІРС або плазмацитоїдною дендритною клітиною. Наприклад, зв'язування з бажаними клітинами можна підтвердити шляхом подвійного забарвлення антитілом, що зв'язується з маркерами поверхні клітин, та антитілом, зв'язувальну активність якого слід перевірити. Тобто ІРС у цьому винаході включають, наприклад, клітини, які експресують BDCA-2. Підтвердження на основі реактивності до трансформованих клітин, що експресують ген ILT7 людини Автори цього винаходу визначили, що імунологічна характеристика ILT7, що експресувався у ІРС людини, була перебудована, коли експресію гена ILT7 людини здійснювали за специфічною умовою. Отже, реактивність до ILT7 людини можна підтвердити на основі реактивності антитіл до клітин, до яких ген, що кодує ILT7, штучно вводиться. А саме, цей винахід стосується моноклонального антитіла, яке включає амінокислотну послідовність, яка утворює позаклітинний домен як певний позаклітинний домен та зв'язується з молекулою, що експресується разом з молекулою трансдукції сигналу, або стосується фрагмента, який включає його антиген-зв'язувальну ділянку. У цьому описі позаклітинний домен складається з амінокислотної послідовності, яка відповідає від 17-ої до 444-ої позиції N-кінцевої амінокислотної послідовності, зображеної на SEQ ID NO: 2 (від 1 до 428 у SEQ ID NO: 2). Наприклад, імунологічну характеристику ILT7, що експресується у ІРС людини, підтримували у клітинах, які трансфектували вектором експресії, який включає ДНК, що кодує ILT7 людини, разом з вектором експресії, який включає ДНК, що кодує молекулу трансдукції сигналу. Отже, трансформована клітина, яка експресує разом ILT7 людини та молекулу трансдукції сигналу, є переважною для підтвердження афінності зв'язування антитіл із позаклітинним доменом ILT7 людини у цьому винаході. У цьому винаході бажано застосовувати клітину, яка не трансформується, як контрольні клітини, коли реактивність антитіл підтверджується застосуванням при цьому трансформованої клітини. Крім того, також важливо 6 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 підтвердити те, що зв'язування антитіл не визначається при застосуванні такої ж самої клітинихазяїна, яка експресує тільки молекулу трансдукції сигналу, як контрольної. У цьому винаході молекулу, яка індукує експресію ILT7 людини на клітинній поверхні, можна застосовувати як молекулу трансдукції сигналу для спільної експресії. Молекулу трансдукції сигналу у цьому винаході можна також визначити як молекулу, яка може надавати імунологічної характеристики природного ILT7 людини принаймні позаклітинному домену молекули ILT7 у клітині, яка експресує ILT7. Як застосовується у цьому описі, термін "імунологічна характеристика" природного ILT7 людини означає розпізнавання антитілом, яке зв'язується з ІРС людини. Специфічно, переважно застосовувати -ланцюг рецептора Fc або DAP 12 як молекулу трансдукції сигналу. У цьому винаході -ланцюг рецептора Fc є особливо переважним як молекула трансдукції сигналу. -Ланцюг рецептора Fc - це молекула, що складається з амінокислотних послідовностей, зображених на SEQ ID NO: 16. Молекула трансдукції сигналу може бути фрагментом, доки ILT7 людини, який мусить спільно експресуватися, розташовується на клітинній поверхні. Доки ILT7 людини, який мусить спільно експресуватися, розташовується на клітинній поверхні, доти мутація або додавання амінокислотної послідовності дозволяється в амінокислотних послідовностях, зображених на SEQ ID NO: 16. Тобто, цим винаходом пропонуються способи отримання клітин, що виробляють моноклональне антитіло, яке зв'язується з позаклітинним доменом ILT7 людини, при цьому спосіб включає наступні етапи: (1) введення імунним тваринам клітини, яка екзогенно експресує білок, що включає позаклітинний домен ILT7 людини, та молекулу, що включає амінокислотні послідовності, вказані у SEQ ID NO: 16; та (2) вибір виробляючої антитіла клітини, яка виробляє антитіло, яке зв'язується з ILT7 людини, з виробляючих антитіла клітин імунних тварин. Внаслідок цього, стосовно антитіла, що зв'язується з ILT7 людини у цьому винаході, є переважним застосовувати антитіло, в якому схрещування з клітинними популяціями, про які відомо, що вони експресують родини ILT, відмінні від ILT7, не спостерігається. Специфічно, стосовно антитіла, що зв'язується з ILT7 людини у цьому винаході, є переважним застосовувати антитіло, у якому зв'язування з клітинними популяціями, про які відомо, що вони експресують родини ILT, відмінні від ILT7, не може спостерігатися за такою ж самою умовою, як і умова, за якої підтвердилося зв'язування з ІРС. Як вже було описано, наприклад, ILT2 та ILT3 експресуються не тільки у PDC, а також у DC, отриманих з MDDC- або CD34-позитивних клітин (Gene. 2004 Ар 28; 331: 159-64). З іншого боку, експресію ILT7 не можна визначити у дендритних клітинах через диференціювання ІРС. Отже, антитіло, яке не може виявляти зв'язування з DC, отриманих від MDDC- або CD34-позитивних клітин за умовою, при якій зв'язування з ІРС можна підтвердити, є антитілом, яке зв'язується з ILT7 людини у цьому винаході. Повідомлялося про наступні конфігурації експресії стосовно інших молекул родини ILT ("The KIR Gene Cluster", Carrington, Mary and Norman, Paul. Bethesda (MD): National Library of Medicine (US), NCBI; 2003, Gene. 2004 Apr 28; 331: 159-64). Отже, антитіло, яке зв'язується з ІРС або PDC людини, та зв'язування якого з наступними клітинами не можна підтвердити, включається в антитіло, що має специфічність до ILT7: ILT1; послідовність генерацій мієлоїдних клітин (моноцити, DC, що походять від моноцитів, макрофаги); ILT2; PDC, В-клітини, CD34-позитивні клітини, DC, що походять від CD34-позитивних клітин, та DC, що походять від моноцитів; ILT3; PDC та DC; ILT5; моноцити, DC, що походять від СD34-позитивних клітин, та DC, що походять від моноцитів; та ILT8; послідовність генерацій клітин-моноцитів. Тобто, моноклональне антитіло, яке зв'язується із позаклітинним доменом ILT7 людини, у цьому винаході переважно включає моноклональне антитіло, що має наступні імунологічні характеристики: a) моноклональне антитіло зв'язується з ІРС людини та b) зв'язування моноклонального антитіла з однією або більше клітинами, що вибрані з групи, яка складається з моноцитів, макрофагів, В-клітин, CD34-позитивних клітин та дендритних клітин, що походять з цих клітин, не можна підтвердити в умовах для зв'язування з ІРС людини. Стосовно моноклонального антитіла цього винаходу переважно застосовувати антитіло, у якому зв'язування з моноцитами, макрофагами, В-клітинами, CD34-позитивними клітинами та 7 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 дендритними клітинами, що походять з цих клітин, не можна підтвердити в умовах для зв'язування, особливо, з ІРС людини. Альтернативно, моноклональне антитіло, яке зв'язується із позаклітинним доменом ILT7 людини у цьому винаході, переважно є моноклональним антитілом, яке має наступні імунологічні характеристики: c) моноклональне антитіло зв'язується з трансформованою клітиною, яка трансфектується вектором експресії, який є виразним носієм ДНК, що кодує ILT7 людини, разом з вектором експресії, який є виразним носієм ДНК, що кодує молекулу трансдукції сигналу; d) зв'язування з клітиною-хазяїном до трансформації не можна підтвердити в умовах для зв'язування з клітинами, що зазнали спільної трансфекції, як описано у (с); або моноклональне антитіло цього винаходу є моноклональним антитілом, яке має наступні імунологічні характеристики: e) зв'язування з клітиною-хазяїном, яка експресує тільки молекулу трансдукції сигналу, не можна підтвердити в умовах для зв'язування з клітинами, що зазнали спільної трансфекції, як описано у (с). У цьому винаході той факт, що моноклональне антитіло проти ILT7 не перехрещується з родиною ILT інших молекул, можна підтвердити, застосовуючи клітини, у яких кожну родину ILT примушували експресуватися. Тобто для примусової експресії, кДНК, що кодує кожну родину ILT амінокислотних послідовностей, вводиться у відповідну клітину-хазяїна. Моноклональне антитіло проти ILT7, схрещування якого слід підтвердити, виробляють так, щоб воно контактувало з отриманою трансформованою клітиною. Потім можна підтвердити, що, коли зв'язування антитіла з клітиною, яка експресує молекули родини ILT, відмінні від ILT7, не спостерігається, антитіло є здатним імунологічно відрізняти ILT7 від інших молекул родини ILT. Наприклад, у прикладах, описаних далі, спостерігається той факт, що моноклональне антитіло проти ILT7, отримане за наступним винаходом, не перехрещується з ILT1, ILT2 та ILT3. Отже, переважним прикладом моноклонального антитіла у цьому винаході є моноклональне антитіло, що зв'язується з ILT7, у якому зв'язування з ILT1, ILT2 та ILT3 неможливо виявити в тих же самих умовах. Зокрема, ILT2 та ILT3 - це гени, експресію яких у ІРС підтвердили (Ju et al. Gene 331,159164, 2004). Проте, ці молекули можуть демонструвати профілі експресії, які є унікальними для кожного клітинного типу, залежно від відповідних рівнів диференціювання у ІРС або умов, таких як стимуляція вірусами або іншими цитокінами. Застосування антитіла, яке є здатним імунологічно відрізняти ці молекули родини ILT від ILT7, дозволяє специфічно виявляти зміни в експресії ILT7. Зв'язування моноклонального антитіла, зв'язувальну активність якого слід підтвердити з різними типами клітин, можна підтвердити на основі, наприклад, принципу проточної цитометрії. Для того, щоб підтвердити реактивність антитіл на основі принципу проточної цитометрії, переважно заздалегідь помітити антитіла молекулою або атомною групою, яка виробляє помітний сигнал. Взагалі застосовуються флуоресцентні або люмінесцентні мітки. Клітинний сортер зі збудженням флюоресценції (FACS) можна застосовувати для того, щоб проаналізувати зв'язування мічених флюоресценцією антитіл з клітинами на основі принципу проточної цитометрії. Застосування клітинного сортера зі збудженням флюоресценції дозволяє ефективно підтвердити зв'язування численних антитіл з численними клітинами. Специфічно, наприклад, антитіло А, стосовно якого попередньо визначили, що воно здатне ідентифікувати ІРС, та антитіло В, характеристики зв'язування з ІРС якого слід проаналізувати, реагували з клітинними популяціями, які у той самий час включали ІРС. Антитіло А та антитіло В мітяться флуоресцентним сигналом, який заздалегідь взаємно розрізняється цими антитілами. У випадку, коли обидва сигнали виявляються з однакових клітинних популяцій, зв'язування цих антитіл з такими ж самими клітинними популяціями можна підтвердити. Іншими словами, визначили, що як антитіло А, так і антитіло В мають однакові характеристики зв'язування. У випадку, коли вони зв'язуються з різними клітинними популяціями, є очевидним, що обидва антитіла мають відмінні характеристики зв'язування. Переважний приклад моноклонального антитіла у цьому винаході може бути моноклональним антитілом, що виробляється гібридомою ILT7#11 або ILT7#17. Гібридома ILT7#11 та гібридома ILT7#17 зберігаються у Національному інституті провідної промислової науки та техніки, Міжнародному сховищі патентних організмів за номерами FERM ВР-10704 та FERM BP-10705 з 21 жовтня 2005 року. Специфікація сховища включає наступне. (a) Назва та адреса закладу зберігання 8 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 Назва: Національний інститут провідної промислової науки та техніки, Міжнародне сховище патентних організмів Адреса: AIST Tsukuba Central 6, 1-1-1, Higashi, Tsukuba-shi, Ibaraki, Japan (zip code 3058566). (b) Дата початку зберігання: 21 жовтня 2005 року. (c) Номер доступу: FERM ВР-10704 (гібридома ILT7#11). (d) Номер доступу: FERM BP-10705 (гібридома ILT7#17). Моноклональне антитіло цього винаходу може бути фрагментом, що включає його антигензв'язувальну ділянку. Наприклад, фрагмент антитіла, що включає антиген-зв'язувальну ділянку, яку отримують шляхом ферментативного перетравлення IgG, можна застосовувати як антитіло у цьому винаході. Специфічно, фрагменти антитіла, такі як Fab та F(ab') 2, можна отримати шляхом перетравлення папаїном або пепсином. Добре відомо, що ці фрагменти антитіла можна застосовувати як молекули антитіла, що мають афінітет до антитіл. Альтернативно, антитіла, побудовані шляхом генної рекомбінації, можна також застосовувати доти, доки зберігається задовільна антиген-зв'язувальна активність. Приклади антитіл, побудованих шляхом генної рекомбінації, включають химерні антитіла, CDR-трансплантовані антитіла, Fvs з єдиним ланцюгом, діатіла, лінійні антитіла та поліспецифічні антитіла, утворені з фрагментів антитіл. Загально відомо, що ці антитіла можна отримати із застосуванням моноклональних антитіл або клітин, що виробляють антитіла. Моноклональне антитіло цього винаходу можна отримати, застосовуючи специфічну трансформовану клітину як імуноген. Тобто, цей винахід стосується способу отримання клітин, що виробляють моноклональне антитіло, яке зв'язується із позаклітинним доменом ILT7 людини, при цьому цей спосіб включає наступні етапи: (1) введення імунним тваринам клітини, що експресує екзогенний білок, який включає позаклітинний домен ILT7 людини, та екзогенну молекулу, яка зв'язана з ILT7 людини; та (2) вибір виробляючої антитіла клітини, яка виробляє антитіло, яке зв'язується з ILT7 людини, з виробляючих антитіла клітин імунних тварин. Отримані таким способом клітини, що виробляють антитіло, або іморталізовані клітини, що виробляють антитіло, культивують, та з культур можна одержувати бажані моноклональні антитіла. Відомо багато способів стосовно іморталізації клітин, що виробляють антитіло. У способі отримання моноклонального антитіла цього винаходу корисні приклади молекули, яка зв'язана з ILT7 людини, для виробництва трансформованої клітини, яку слід застосовувати як імуноген, включають білки клітинної мембрани. Серед них молекулу трансдукції сигналу, яка розташовується у клітинних мембранах, переважно застосовувати як білок клітинної мембрани у цьому винаході. Термін "молекула трансдукції сигналу" означає молекулу, яка пов'язується з білками та клітинами, які мають рецепторні структури у позаклітинному домені, та яка передає стимуляцію зв'язування лігандів до рецепторів у клітинах. Приклади молекули трансдукції сигналу включають -ланцюг рецептора Fc, DAP 12 або подібні. Наприклад, -ланцюг рецептора Fc є переважним для застосування як білка клітинної мембрани у цьому винаході. Амінокислотні послідовності DAP 12 людини та -ланцюг рецептора Fc, а також послідовність основ кДНК, що кодує ці послідовності, є широко відомими. Послідовність основ -ланцюга рецептора Fc та амінокислотна послідовність, що кодується цією послідовністю основ, наведені в SEQ ID NOs: 15 та 16, відповідно. У цьому винаході трансформовану клітину, яку слід застосовувати як імуноген, можна отримати шляхом приготування, наприклад, клітини, яка є виразним носієм наступних (а) та (b): (a) екзогенного полінуклеотиду, що кодує амінокислотну послідовність, яка включає позаклітинний домен ILT7 людини; та (b) екзогенного поілнуклеотиду, що кодує -ланцюг рецептора Fc. У цьому винаході екзогенний полінуклеотид означає полінуклеотид, який штучно вводиться у клітину-хазяїна. Коли клітини людини застосовуються як такі клітини, гени людини вводяться у клітини людини. У такій комбінації штучно введений полінуклеотид означає екзогенний полінуклеотид. Отже, ектопічна експресія ILT7 людини або -ланцюга рецептора Fc людини включається в експресію екзогенного полінуклеотиду. Як застосовується у цьому описі, термін "позаклітинний домен ILT7 людини" означає амінокислотну послідовність від 17-ої до 444-ої позиції амінокислотної послідовності, наведеної в SEQ ID NO: 2, яка відповідає позаклітинному домену амінокислотної послідовності (від 1 до 428 у SEQ ID NO: 2). Стосовно амінокислотної послідовності, що включає позаклітинний домен ILT7 людини, у цьому винаході переважно застосовувати амінокислотну послідовність, яка включає кожну ділянку, наприклад, починаючи з N-закінчення, у наступному порядку: 9 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 сигнальна послідовність + позаклітинний домен + трансмембранний домен + внутріклітинна ділянка. Альтернативною є амінокислотна послідовність, у якій частково не вистачає внутріклітинної ділянки, як описано далі, включається в амінокислотну послідовність, яка включає позаклітинний домен ILT7 людини, у цьому винаході: сигнальна послідовність + позаклітинний домен + трансмембранний домен + частина внутріклітинної ділянки. Крім того, структура, у якій не вистачає внутріклітинної ділянки, як згадується далі, включається в амінокислотну послідовність, яка включає позаклітинний домен ILT7 людини, у цьому винаході, тобто: сигнальна послідовність + позаклітинний домен + трансмембранний домен. У структурі, ділянки, відмінні від позаклітинного домену, можуть бути амінокислотними послідовностями, що вибираються з амінокислотних послідовностей, наведених у SEQ ID NO: 2, або можуть об'єднуватися з іншими амінокислотними послідовностями, що мають гомологію з цими ділянками. Наприклад, амінокислотна послідовність, що складає сигнальну послідовність, трансмембранний домен та внутріклітинну ділянку, може бути амінокислотною послідовністю молекул родини ILT, відмінних від ILT7. Або її можна об'єднати з амінокислотною послідовністю родини ILT у зразках, відмінних від зразків людини. Крім того, амінокислотна послідовність, яка складає ділянки, відмінні від позаклітинного домену, може включати мутацію у діапазоні, придатному для зберігання функції кожної ділянки. Альтернативно, інші ділянки можуть втручатися між кожною ділянкою. Наприклад, мітку епітопу, таку як FLAG, можна також вставити між сигнальною послідовністю та позаклітинним доменом. Зокрема, сигнальна послідовність видаляється шляхом процесингу під час її перенесення до поверхні клітинної мембрани після того, як її транслювали у білок. Отже, довільну амінокислотну послідовність, яка індукує перехід трансльованого білка до клітинної мембрани, можна застосовувати як сигнальну послідовність. Більш специфічно, переважно застосовувати амінокислотну послідовність (SEQ ID NO: 2) ILT7 людини як амінокислотну послідовність, що включає позаклітинний домен ILT7 людини. Отже, у цьому винаході довільну послідовність основ, що кодує амінокислотну послідовність, яка складає вищезгадану структуру [сигнальна послідовність + позаклітинний домен + трансмембранний домен + внутріклітинна ділянка], можна застосовувати як полінуклеотид, який складає екзогенний полінуклеотид, описаний у (а). Наприклад, амінокислотна послідовність SEQ ID NO: 2 кодується послідовністю основ, визначеній у SEQ ID NO: 1. У цьому винаході вектор експресії, який є виразним носієм вищезгаданих полінуклеотидів (а) та (b), можна ввести у відповідну клітину-хазяїна для отримання трансформованої клітини, яку слід застосовувати як імуноген. Один вектор або різні вектори можуть бути носіями полінуклеотидів (а) та (b). Коли різні вектори є носіями кожного полінуклеотиду, клітини-хазяїни трансфектуються спільно двома типами векторів. Переважні приклади клітини-хазяїна у цьому винаході включають клітини ссавців. Специфічні приклади клітини-хазяїна включають клітини, що походять від людей, мавп, мишей або щурів. Зокрема, клітини, що походять від людей, є переважними клітинами-хазяїнами. Наприклад, у цьому винаході переважно застосовувати як клітину-хазяїна клітини 293Т, що походять від людей. Клітини 293Т можна отримати як АТСС CRL-11268. Крім того, клітини, що походять від імунних тварин, можна також застосовувати як клітини-хазяїни. Коли клітини, що походять від імунних тварин, застосовуються як і імуноген, отримують незначну імунологічну реакцію на клітини-хазяїнів. Внаслідок цього можна отримати ефективне антитіло проти позаклітинного домену ILT7, який експресується екзогенно. Отже, наприклад, коли мишей застосовують як імунних тварин, клітини, що походять від мишей, можна також застосовувати як клітини-хазяїнів. Вищезгадані полінуклеотиди можна трансфектувати у клітини, якщо їх носієм буде вектор, здатний індукувати експресію у клітинах-хазяїнах. Можна застосовувати комерційно доступні вектори, які можуть індукувати експресію у клітинах ссавців. Для цього винаходу можна застосовувати вектори експресії, такі як pCMV-Script(R) Vector, PSG5 Vector (виробляється Stratagene), pcDNA3.1 (виробляється Invitrogen). Трансформовані таким способом клітини вводяться імунним тваринам за необхідністю разом з додатковими компонентами, такими як ад'юванти. Корисні приклади ад'юванту включають повний ад'ювант Фрейнда та йому подібні. У випадку застосування мишей як імунних 4 9 тварин трансформовані клітини можна вводити у діапазоні від 10 до 10 клітин, більш 4 6 специфічно від 10 до 10 клітин. Взагалі, багаторазові дози імуногена вводяться з регулярними інтервалами доти, поки титр антитіла не підвищиться. Наприклад, у випадку нетривалої 10 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 імунізації трансформовані клітини вводять з інтервалами від 2 до 4 днів, більш специфічно з інтервалами у 3 дні. Після введення двічі або тричі можна одержати клітини, що виробляють антитіло. Альтернативно, їх вводять один раз на тиждень, а клітини, що виробляють антитіло, можна також одержати після введення п'ять або шість разів. У цьому винаході одержані клітини, що виробляють антитіла, клонуються, внаслідок чого отримують моноклональні антитіла. Переважно, щоб клітини, що виробляють антитіла, були іморталізовані для клонування. Наприклад, спосіб злиття клітин, типовим зразком якого є спосіб гібридом або спосіб трансформації вірусом Епштейна-Бара (EBV), можна застосовувати як спосіб іморталізації клітин, що виробляють антитіла. Стосовно клітин, що виробляють антитіла, одна клітина виробляє один тип антитіла. Отже, створення клітинних популяцій, що походять від однієї клітини (тобто клонування), дозволяє отримати моноклональні антитіла. Спосіб гібридом включає процес, при якому клітини, що виробляють антитіла, зливаються з відповідною клітинною лінією, яка є іморталізованою, а потім піддають клонуванню. Іморталізовані клітини, що виробляють антитіла, можна клонувати за допомогою способу, такого як спосіб серійних розведень. Відомо, що існує багато клітинних ліній, корисних для способу гібридом. Ці клітинні лінії є відмінними стосовно ефективності іморталізації лімфоцитних клітин та мають різні генетичні маркери, необхідні для вибору успішно злитих клітин. Крім того, коли намагаються отримати клітини, що виробляють антитіла, можна також застосовувати клітинну лінію, у якої нема здатності виробляти антитіла. Наприклад, мієлому мишей P3x63Ag8.653 (ATCC CRL-1580) та P3x63Ag8U.l (ATCC CRL1597) широко застосовують як корисні клітинні лінії для способу злиття клітин для мишей або щурів. Взагалі, гібридома виробляється шляхом злиття гомогенних клітин, проте моноклональне антитіло можна також отримати з гетерогенної гібридоми з різних зразків серед близько споріднених зразків. Специфічні протоколи злиття клітин є широко відомими. А саме, виробляючі антитіла клітини імунних тварин змішуються з відповідними партнерами по злиттю, щоб виконати злиття клітин. Прийнятні приклади клітини, що виробляє антитіло, включають клітини селезінки, лімфоцитні клітини, взяті з лімфатичного вузла, та В-клітини периферійної крові. Як партерів по злиттю можна застосовувати різні клітинні лінії, які описано раніше. Для злиття клітин можна застосовувати спосіб поліетиленгліколю та спосіб електричного злиття. Далі успішно злиті клітини вибирають на основі маркерів селекції злитих клітин. Наприклад, коли НАТ-чутливу клітинну лінію застосовують для злиття клітин, успішно злиті клітини вибираються шляхом селекції клітин, що зростають у середовищі HAT. Далі підтверджують, що антитіла, вироблені вибраними клітинами, мають бажану реактивність. Кожну гібридому піддають скринінгу на основі реактивності антитіл. А саме, гібридома, що виробляє антитіла, які зв'язуються з ILT7 людини, вибирається за способом, описаним раніше. Переважно, коли вибрану гібридому субклонують, а потім зрештою підтверджується виробництво бажаного антитіла, і підтверджене антитіло стає підставою для вибору гібридоми, що виробляє моноклональне антитіло цього винаходу. Специфічно, бажану гібридому можна вибирати на основі реактивності до клітин людини або реактивності до трансформованої клітини, яка експресує ген ILT7 людини. Антитіла, що зв'язуються з клітинами, можна виявити на основі принципу імунологічного аналізу. Наприклад, аналіз ЕЛАЙЗА, в якому клітини застосовуються як антигени, можна використовувати для виявлення бажаного антитіла. Специфічно, виготовляється культуральна надосадова рідина гібридоми, яка мусить контактувати з основою, на якій ІРС людини або трансформована клітина застосовується як імуноген. У випадку, коли культуральна надосадова рідина включає бажане антитіло, це антитіло захоплюється у клітину, іммобілізовану на основі. Потім тверду фазу відокремлюють від культуральної надосадової рідини та за необхідністю промивають. Після цього можна виявити антитіло, що потрапило у тверду фазу. Антитіло, що розпізнає антитіло, можна застосовувати для виявлення антитіл. Наприклад, антитіло миші можна виявити антитілом анти-мишачого імуноглобуліну. Виявлення здійснити легко, якщо антитіло, яке розпізнає антитіло, є міченим. Придатні приклади мітки включають ферменти, флуоресцентні барвники, люмінесцентні барвники та їм подібні. З іншого боку, частинки та внутрішню стінку титраційного мікропланшета можна застосовувати як основу, на якій клітини іммобілізуються. Клітини можна іммобілізувати на частинках, виготовлених з пластмаси, або на поверхні контейнера шляхом фізичної адсорбції. Придатні приклади основи для іммобілізації клітин включають гранули, виготовлені з полістиролу, та реакційні судини. 11 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 У селекції гібридом можна передбачати виробництво антитіла не проти ILT7, а проти клітини-хазяїна трансформованої клітини, що застосовується як імуноген. Наприклад, як проілюстровано у Прикладах, у випадку, коли клітину людини застосовують як імуноген, а мишу застосовують як імунну тварину, клітина людини розпізнається як стороння речовина. Отже, передбачається виробництво антитіла, що зв'язується зі сторонньою речовиною. У цьому винаході наполегливо намагаються отримати антитіло, здатне розпізнавати ILT7 людини. Отже, не має необхідності отримати антитіло, що розпізнає антигени клітини людини, відмінні від ILT7 людини. Для того, щоб видалити гібридоми, які виробляють таке антитіло, при скринуванні, небажані антитіла можна абсорбувати до підтвердження реактивності антитіла. Небажані антитіла можна абсорбувати антигеном, з яким зв'язується антитіло, існування якого передбачається. Специфічно, наприклад, антитіло проти антигенів-клітин людини, відмінних від ILT7 людини, можна абсорбувати клітиною, яка не може детектувати експресію ILT7 людини. У цьому винаході переважно застосовувати клітину-хазяїна, що застосовується для імуногена, як антиген для абсорбції небажаних антитіл. Альтернативно, клітину-хазяїна, яка не експресує позаклітинний домен ILT7 людини, проте експресує молекулу, яка пов'язується з ILT7, можна застосовувати як антиген для абсорбції цих антитіл. Стосовно моноклонального антитіла, зв'язувальна активність якого з антигеном підтверджується, його дійсний вплив на активність ІРС підтверджується за необхідністю. Вплив на ІРС можна підтвердити за способами, які описані далі. Стосовно моноклонального антитіла цього винаходу, гібридома, що виробляє моноклональне антитіло, культивується, а моноклональне антитіло цього винаходу одержується з отриманої культури. Гібридому можна культивувати in vitro та in vivo. У випадку культивування in vitro гібридому можна культивувати, застосовуючи відоме культуральне середовище, таке як RPMI1640. Імуноглобулін, що виділяється гібридомою, накопичується у культуральній надосадовій рідині. Отже, моноклональне антитіло цього винаходу можна отримати шляхом збирання культуральної надосадової рідини та очищення її за необхідністю. Легше очистити імуноглобулін, коли сироватку не додають до культурального середовища. Проте, маючи на меті більш швидку проліферацію гібридоми та сприяння виробництву антитіла, 10% фетальну бичачу сироватку можна також додати до культурального середовища.Гібридому можна також культивувати in vivo. Специфічно, внутрішньочеревну культивацію гібридоми можна здійснювати шляхом інокуляції гібридоми у черевну порожнину голих мишей. Моноклональні антитіла накопичуються в асцитах. Отже, якщо отримують асцит та за необхідністю його очищують, бажане моноклональне антитіло можна отримати. Отримані моноклональні антитіла можна відповідним способом модифікувати або обробляти згідно з призначенням їх застосування. Моноклональне антитіло цього винаходу може експресуватися шляхом отримання кДНК, що кодує антиген-зв'язувальну ділянку антитіла, з гібридоми, та вставлення її у відповідний вектор експресії. Відомим є спосіб, при якому отримують кДНК, яка кодує варіабельну ділянку антитіла, а потім вона експресується у відповідній клітині-хазяїні. Крім того, також відомим є спосіб, при якому химерне антитіло отримують шляхом вшивання варіабельної ділянки, яка включає антиген-зв'язувальну ділянку, у константну ділянку. Переважні приклади моноклонального антитіла у цьому винаході включають моноклональні антитіла, що виробляються гібридомою #11 (Номер доступу: FERM ВР-10704), гібридомою #17 (Номер доступу: FERM BP-10705) або гібридомою #37. Амінокислотні послідовності, що складають варіабельні ділянки цих моноклональних антитіл, а також послідовності основ кДНК, що їх кодує, описано далі. Отже, наприклад, химерні антитіла, які слід отримати шляхом кон'югації цих варіабельних ділянок з константними ділянками інших імуноглобулінів, є переважними у цьому винаході. В амінокислотних послідовностях, описаних у Переліку послідовностей, амінокислотна послідовність від 1 до С-закінчення складає зрілий білок. А саме, отримана внаслідок цього, амінокислотна послідовність від 1 до С-закінчення для кожної амінокислотної послідовності - це зріла послідовність кожної амінокислотної послідовності. З іншого боку, амінокислотна послідовність, представлена числовим значенням від N-закінчення до -1, - це сигнальна послідовність. 12 UA 97946 C2 5 10 Наприклад, химерне антитіло миші (варіабельна ділянка) - людини (константна ділянка) можна отримати шляхом вшивання генів цих варіабельних ділянок у константну ділянку важкого ланцюга lgG1 людини та ген, що кодує константну ділянку легкого ланцюга Ig каппа людини, відповідно. Амінокислотні послідовності такого химерного антитіла та послідовності основ, що його кодують, відповідно описано далі. Химерні антитіла, що визначаються цими послідовностями, демонструють побудову переважного варіанту моноклонального антитіла проти ILT7 у цьому винаході. У наступних амінокислотних послідовностях химерних антитіл амінокислотна послідовність від N-закінчення до -1 відповідає сигнальній послідовності, а амінокислотна послідовність від 1 до С-закінчення відповідає зрілому білку. Тобто, химерне антитіло, що включає важкий та легкий ланцюги, які складаються з амінокислотної послідовності від 1 до С-закінчення для кожної амінокислотної послідовності, є переважним у цьому винаході. 15 20 25 30 Крім того, антиген-зв'язувальну активну ділянку моноклонального антитіла можна також трансплантувати у інші імуноглобуліни. Варіабельна ділянка імуноглобуліну включає гіперваріабельну ділянку (CDR) та каркасну ділянку. Антиген-зв'язувальна властивість кожного імуноглобуліну визначається CDR, а каркасна ділянка підтримує структуру антигензв'язувальної ділянки. Амінокислотна послідовність CDR є надзвичайно багатою на різноманітність, тоді як амінокислотна послідовність частини каркасної ділянки є надзвичайно консервативною. Відомо, що амінокислотна послідовність, що складає CDR, включається у каркасну ділянку інших молекул імуноглобуліну, що дозволяє переносити антиген-зв'язувальну активність. Розроблено спосіб, при якому антиген-зв'язувальна властивість різних імуноглобулінів переноситься на імуноглобулін людини із застосуванням цього процесу. Як застосовується у цьому описі, термін "антиген-зв'язувальна ділянка" може включати CDR, що трансплантується у каркасну ділянку. Отже, термін "фрагмент, що включає антигензв'язувальну ділянку" певного моноклонального антитіла включає фрагмент імуноглобуліну людини, який включає варіабельну ділянку, до якої трансплантується CDR моноклонального антитіла. Наприклад, кожна амінокислотна послідовність вищезгаданих варіабельних ділянок включає наступні амінокислотні послідовності (SEQ ID NO) як CDR. 13 UA 97946 C2 5 10 15 20 25 30 35 40 45 На основі інформації стосовно послідовності основ, яка кодує вищезгадані амінокислотні послідовності, та інформації стосовно послідовності основ, яка кодує каркасну ділянку (FR) імуноглобуліну людини, можна побудувати праймер та ампліфікувати кДНК, що має послідовність основ, отриману шляхом кон'югації обох послідовностей основ. Операцію повторюють для кожної каркасної ділянки, та можна побудувати варіабельну ділянку, у якій CDR1, CDR2 та CDR3 мишей з'єднуються каркасною ділянкою людини. Далі, коли послідовність основ, що кодує константу ділянку імуноглобуліну людини, зазнає кон'югації, за необхідністю можна отримати гуманізоване антитіло з константною ділянкою. Стосовно химерного антитіла, що включає вищезгадані варіабельні ділянки, або гуманізованого антитіла, до якого трансплантується CDR, яка складає варіабельну ділянку, антитіло з константною ділянкою, що походить від IgG або IgM, включається у переважне антитіло у цьому винаході. Автори цього винаходу підтвердили, що моноклональне антитіло проти ILT7 демонструє CDC-дію на клітинах, що експресують ILT7. Отже, антитіло, що має константу ділянку, яка походить від IgG або IgM, демонструє цитотоксичність проти клітин, що експресують ILT7, внаслідок CDC-ефекту. Такі антитіла є корисними для інгібування ряду клітин, що експресують ILT7, таких як ІРС. Химерне антитіло, здатне розпізнавати ILT7, або гуманізоване антитіло, що пропонується цим винаходом, можна отримати шляхом генної інженерії, застосовуючи полінуклеотиди, що кодують ці антитіла. Наприклад, полінуклеотид, який є послідовністю основ, представленою наступними SEO ID NO, та кодує амінокислотну послідовність, що складає зрілий білок для амінокислотної послідовності, можна застосовувати як полінуклеотид, що кодує варіабельну ділянку #11 або #17. Отримана внаслідок цього амінокислотна послідовність від 1 до Сзакінчення для кожної амінокислотної послідовності відповідає зрілому білку. У випадку, коли кожний зрілий білок експресується як окремий білок, переважно розташовувати сигнал секреції на N-закінченні кожної амінокислотної послідовності. Наприклад, в амінокислотних послідовностях, представлених в цих SEQ ID NO, амінокислотну послідовність від N-закінчення до -1 можна застосовувати як сигнальну послідовність, коли такі білки експресуються у тваринних клітинах. Альтернативно, ці варіабельні ділянки можна виділяти як зрілі білки, застосовуючи довільну сигнальну послідовність, що робить можливою секрецію імуноглобуліну. Таким самим способом, як описано вище, стосовно полінуклеотиду, що кодує гуманізоване антитіло, полінуклеотид, який експресує гуманізоване антитіло, можна отримати, застосовуючи послідовність основ, яка кодує білок, що має сигнальну послідовність, яку слід додати до Nзакінчення. Коли важкий та легкий ланцюги переносяться на окремих векторах, обидва вектори спільно трансфектуються в одну клітину-хазяїна. Важкий та легкий ланцюги, що експресуються з кожного вектора, застосовуються для побудови молекули імуноглобуліну з обома ланцюгами. Або полінуклеотид, що кодує важкий ланцюг, та полінуклеотид, що кодує легкий ланцюг, можна також переносити на одному векторі. Клітина-хазяїн, у яку вектор, який є носієм обох полінуклеотидів, трансфектується, експресує важкий та легкий ланцюги та виробляє імуноглобулін, що має обидва ланцюги. Ці полінуклеотиди можна експресувати як антитіла, застосовуючи систему вектор-хазяїн, здатну експресувати ген антитіла. Крім того, у випадку, коли вони експресуються як молекула сигнального білка шляхом з'єднання варіабельної ділянки важкого ланцюга та варіабельної ділянки легкого ланцюга, сигнальну послідовність можна розташувати на N-закінченні молекули 14 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 білка. Відомий приклад такої молекули антитіла включає молекулу scFv, у якій варіабельна ділянка важкого ланцюга та варіабельна ділянка легкого ланцюга з'єднуються лінкером. Кожне моноклональне антитіло, яке отримали за таким способом, є моноклональним антитілом цього винаходу. Іншими словами, моноклональне антитіло, що складається з імуноглобуліну, який включає антиген-зв'язувальну ділянку, що кодується полінуклеотидом, який походить від кДНК, яка кодує антиген-зв'язувальну ділянку вищезгаданих моноклональних антитіл, є моноклональним антитілом цього винаходу. Як описано раніше, клітини RBL, у яких ген ILT1 примусово експресувався, можна застосовувати як імуноген для отримання антитіл ILT1. Проте, експресію ILT7 на поверхні клітин RBL (P815) не можна підтвердити, та внаслідок цього її не можна застосовувати як імуноген. Автори цього винаходу визначили, що експресію ILT7 людини на поверхні клітин можнаіндукувати шляхом спільної експресії ILT7 людини та інших білків клітинної мембрани, які пов'язані з ILT7 людини. Потім автори цього винаходу визначили, що антитіло, яке зв'язується з ІРС людини, можна отримати, застосовуючи трансформовану клітину, експресія якої таким способом індукується, як імуноген, та виконали цей винахід. Тобто, цим винаходом пропонується імуноген для виробництва антитіла, яке зв'язується з позаклітинним доменом ILT7 людини, та він є або тваринними клітинами, у яких (а) полінуклеотид, який кодує амінокислотну послідовність, що включає позаклітинний домен ILT7 людини; та (б) полінуклеотид, що кодує -ланцюг рецептора Fc, зберігаються так, щоб експресуватися екзогенно, або мембранними фракціями цих клітин. Вже минуло шість або більше років з того часу, як у 1998 році відкрили структуру ILT7 людини. Проте, антитіло, здатне специфічно розпізнавати ILT7, досі не отримали. Антитіло, здатне розпізнавати ILT7 людини, вперше отримали, застосовуючи імуноген цього винаходу. Тобто, цим винаходом запропоновано антитіло, здатне розпізнавати ILT7 людини, яке можна отримати, виконуючи наступні етапи: (1) введення імунним тваринам клітини, яка екзогенно експресує білок, який включає позаклітинний домен ILT7 людини, та молекулу, яка зв'язана з ILT7 людини; (2) вибір виробляючої антитіла клітини, яка виробляє антитіло, яке зв'язується з ILT7 людини, з виробляючих антитіла клітин імунних тварин; та (3) культивування вибраних на етапі (2) клітин, які виробляють антитіла, та одержання антитіла, здатного розпізнавати ILT7 людини, з цих культур. Визначили, що ILT7 людини специфічно експресується у ІРС людини. Під час аналізу генної експресії шляхом SAGE, який здійснювали автори цього винаходу, специфічну експресію ILT7 людини у ІРС людини також підтвердили. Проте, в останніх повідомленнях рівні експресії ILT7 в обох випадках проаналізували на основі мРНК. Оскільки антитіло, здатне виявляти ILT7 людини, не отримали, стан експресії білка не проаналізували традиційним способом. Аналіз білка ILT7 людини виконували з урахуванням антитіла, яке зв'язується з позаклітинним доменом ILT7 людини, цього винаходу. Автори цього винаходу дійсно підтвердили, що моноклональне антитіло, яке зв'язується з позаклітинним доменом ILT7 людини, на основі цього винаходу специфічно виявляло ІРС людини. Отже, цей винахід стосується способу виявлення клітин, що виробляють інтерферон, який включає етапи: контактування моноклонального антитіла, яке зв'язується з позаклітинним доменом ILT7 людини, або фрагмента, що включає його антиген-зв'язувальну ділянку, з тестовою клітиною та виявлення моноклонального антитіла, зв'язаного з клітинами, або фрагмента, який включає його антиген-зв'язувальну ділянку. Виявлення ILT7 людини на основі цього винаходу дозволяє визначити, чи є певна клітина ІРС. Тобто, цим винаходом пропонується спосіб ідентифікації ІРС, застосовуючи ILT7 людини як індикатор. Або, ІРС людини можна відокремити шляхом відокремлення клітин, у яких виявили ILT7 людини, на основі цього винаходу. Тобто, цим винаходом пропонується спосіб відокремлення ІРС, застосовуючи ILT7 людини як індикатор. На основі аналізу антитілом ILT7 людини підтвердили, що рівень експресії ILT7 у ІРС, диференціювання яких індукувалася CpG та йому подібним, знизився. Тобто, ІРС до того, як індукується їх диференціювання, можна специфічно виявити, застосовуючи ILT7 як індикатор. Іншими словами, моноклональне антитіло цього винаходу є корисним, зокрема для виявлення ІРС до початку їх диференціювання у дендритні клітини. Як застосовується у цьому описі, терміном "ІРС до їх диференціювання" можна визначити клітинні популяції, що зберігають здатність виробляти інтерферон. У цьому винаході моноклональне антитіло, яке зв'язується з позаклітинним доменом ILT7 людини, або фрагмент, що включає його антиген-зв'язувальну ділянку, можна заздалегідь помітити. Наприклад, антитіла можна легко виявити завдяки міченню люмінесцентними 15 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 барвниками або флуоресцентними барвниками. Більш специфічно, антитіло, мічене флуоресцентним барвником, отримують, щоб воно контактувало з клітинною популяцією, яка може включати ІРС, а потім клітини, з якими антитіло цього винаходу зв'язано, можна виявити, застосовуючи флуоресцентний барвник як індикатор. Далі, ІРС можна відокремити шляхом відокремлення клітин, у яких виявляється флуоресцентний барвник. Серію етапів можна легко здійснити на основі принципу FACS (клітинного сортера з активацією флюоресценції). Альтернативно, антитіло цього винаходу можна заздалегідь зв'язати з твердофазною основою, такою як магнітні частинки. Антитіло, зв'язане з твердофазною основою, розпізнає ILT7 людини, а потім ІРС захоплюються у твердофазну основу. Внаслідок цього ІРС можна виявити або відокремити. Як реагент для виявлення ІРС можна отримати антитіло, необхідне для способу виявлення ІРС на основі цього винаходу. Тобто, цим винаходом пропонується реагент для виявлення клітин, що виробляють інтерферон, який включає моноклональне антитіло, яке зв'язується з позаклітинним доменом ILT7 людини, або фрагмент, який включає його антиген-зв'язувальну ділянку. Реагент для виявлення ІРС цього винаходу можна застосовувати у додаток до антитіл у комбінації з позитивними контрольними клітинами або негативними контрольними клітинами. Наприклад, трансформовані клітини, що експресують позаклітинний домен ILT7 людини та застосовуються як імуноген, а також ІРС, отримані від людини, можна застосовувати як позитивні контрольні клітини. Звичайно, тільки невелику кількість ІРС людини можна отримати з периферійної крові. Тому переважно застосовувати особливо трансформовану клітину як позитивну контрольну клітину у реагенті цього винаходу. З іншого боку, довільну клітину, яка не експресує ILT7 людини, можна застосовувати як негативну контрольну клітину. Тобто, цим винаходом пропонується набір для виявлення ІРС людини, який містить: (a) моноклональне антитіло, що зв'язується з позаклітинним доменом ILT7 людини, або фрагмент, що включає його антиген-зв'язувальну ділянку; та (b) клітину, що експресує екзогенний білок, який включає позаклітинний домен ILT7 людини, та екзогенну молекулу, що зв'язана з ILT7 людини. Автори цього винаходу проаналізували вплив антитіла, що зв'язується з позаклітинним доменом ILT7 людини, на ІРС. Внаслідок цього, підтвердилося, що антитіло, яке зв'язується із позаклітинним доменом ILT7 людини, інгібує активність ІРС. Тобто, цей винахід стосується способу інгібування активності клітин, що виробляють інтерферон, який включає етап контактування будь-якого з наступних компонентів з клітинами, що виробляють інтерферон: (а) моноклонального антитіла, що зв'язується з ILT7 людини та інгібує активність клітин, що виробляють інтерферон, або фрагмента, що включає його антиген-зв'язувальну ділянку; та (b) імуноглобуліну, до якого гіперваріабельна ділянка моноклонального антитіла, описаного у (а), трансплантована, або фрагмента, що включає його антиген-зв'язувальну ділянку. Або цей винахід стосується способу інгібування активності клітин, що виробляють інтерферон, у живих організмах, який включає етап введення живим організмам будь-якого з наступних компонентів: (a) моноклонального антитіла, що зв'язується з ILT7 людини та інгібує активність клітин, що виробляють інтерферон, або фрагмента, що включає його антиген-зв'язувальну ділянку; (b) фрагмента, що включає імуноглобулін, до якого гіперваріабельна ділянка моноклонального антитіла, описаного у (а), трансплантована, або фрагмента, що включає його антиген-зв'язувальну ділянку; та (c) полінуклеотиду, що кодує компоненти, описані у (а) або (b). Як застосовується у цьому описі, термін "клітини, що виробляють інтерферон, (ІРС)" означає клітини, що мають здатність виробляти інтерферон та експресувати ILT7 на клітинній поверхні. Далі у цьому описі, якщо інше не вказується, термін "ІРС" охоплює не тільки клітини, що є клітинами-попередниками дендритних клітин, а також клітини, що мають здатність виробляти інтерферон та експресувати ILT7 на клітинній поверхні. Способи ідентифікації таких ІРС є загально відомими. ІРС можна відрізнити від інших клітин крові, застосовуючи деякі маркери клітинної поверхні як індикатори. Специфічно, профіль маркерів клітинної поверхні ІРС людини описано далі (Shortman, K. and Liu, YJ. Nature Reviews 2: 151-161, 2002). За останні роки в деяких повідомленнях також було запропоновано, що BDCA-2-позитивні клітини визначаються як ІРС (Dzionek, A. et al. J. Immunol. 165: 6037-6046, 2000). Профіль антигенів клітинної поверхні ІРС людини СD4-позитивні, СD123-позитивні. Послідовність клітинних генерацій (CD3, CD14, CD16, CD19, CD20, CD56)-негативні та CD11с-негативні 16 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 Отже, можна також сказати, що ІРС - це клітини, що мають профіль експресії цих відомих маркерів та мають здатність виробляти інтерферон. Далі, клітини у живих організмах зі здатністю виробляти інтерферон включають у ІРС, навіть якщо клітини є клітинною популяцією з профілями, відмінними від структури експресії профілю експресії цих маркерів. Далі, приклади характеристик, які звичайно спостерігаються у ІРС людини, є наступними. Морфологічна характеристика клітин - Подібні до клітин плазми, - круглі клітини з гладкою клітинною поверхнею, - ядро є відносно великим. Функціональна характеристика клітин - Під час вірусної інфекції велика кількість інтерферонів типу 1 виробляється протягом короткого періоду часу; - диференціюються у дендритні клітини після вірусної інфекції. Як застосовується у цьому описі, термін "інгібування активності ІРС" означає інгібування принаймні однієї з функцій ІРС. Приклади функції ІРС включають виробництво інтерферону та виживання клітин. Виживання клітин можна також інтерпретувати як кількість клітин. Отже, у випадку інгібування обох або однієї з цих функцій говорять, що активність ІРС інгібується. Визначили, що інтерферон типу 1, що виробляється ІРС, викликає різні хвороби. Отже, інгібування ряду ІРС та виробництва інтерферону є корисним для стратегії медичного лікування цих хвороб. Наприклад, вказується на взаємозв'язок між патологічним станом автоімунних хвороб та інтерфероном-. Більша частина IFN- виробляється ІРС. Отже, патологічні стани, що спричиняються IFN-, можна полегшити шляхом інгібування виробництва інтерферону-. Як застосовується у цьому описі, термін "інгібування виробництва інтерферону ІРС" означає інгібування виробництва принаймні одного з інтерферонів, що виробляються ІРС. Переважний інтерферон у цьому винаходу - це інтерферон типу 1. Серед них важливим є IFN-. Тобто, цей винахід стосується інгібітора виробництва інтерферону, який включає антитіло, яке зв'язується з позаклтинним доменом ILT7, як активний інгредієнт. Або цим винаходом пропонується спосіб інгібування виробництва інтерферону, який включає етап введення антитіла, що зв'язується із позаклітинним доменом ILT7. Далі, цей винахід стосується застосування антитіла, яке зв'язується з позаклітинним доменом ILT7, у виробництві медичної композиції для інгібування виробництва інтерферону. Клітини, у яких велика кількість інтерферону виробляється невеликою кількістю клітин, включаються до ІРС. Наприклад, клітини-попередники дендритних клітин, стимульовані вірусами та їм подібними, виробляють більшу частину інтерферону, що виробляється живим організмом. Наслідком інгібування ряду ІРС, які виробляють багато інтерферону, є супресія виробництва інтерферону. Завдяки цьому патологічні стани, спричинені IFN-, можна зменшити шляхом інгібування ряду ІРС. Підтвердили, що зв'язок моноклонального антитіла проти ILT7 з клітинами, що експресують ILT7, і наступний ефект цитотоксичності були наслідком комплемент-залежної цитотоксичності (CDC) у переважному варіанті здійснення цього винаходу. Ефект CDC - це один з важливих механізмів ліків з антитілом. Моноклональне антитіло проти ILT7 цього винаходу також має сильну цитотоксичність проти клітин, що експресують ILT7, таких як ІРС, внаслідок його ефекту CDC. Тобто, стосовно моноклонального антитіла проти ILT7 у переважному варіанті здійснення винаходу, окрім механізму інгібування виробництва інтерферону, можна очікувати ефект інгібування виробництва інтерферону внаслідок цитотоксичності проти ІРС. Антитіло, що розпізнає позаклітинний домен ILT7 людини, що слід застосовувати для цього винаходу, можна отримати на основі способу, описаного раніше. Антитіло у цьому винаході може бути будь-якого класу. Види організмів, з яких походить антитіло, також не обмежуються. Далі, фрагмент, що включає антиген-зв'язувальну ділянку антитіла, можна застосовувати як антитіло. Наприклад, фрагмент антитіла, який включає антиген-зв'язувальну ділянку, яку отримують шляхом ферментного перетравлення IgG, можна застосовувати як антитіло цього винаходу. Специфічно, фрагменти антитіла, такі як Fab та F(ab') 2, можна отримати шляхом перетравлення папаїном або пепсином. Добре відомо, що ці фрагменти антитіл можна застосовувати як молекули антитіл, які мають афінітет до антитіл. Альтернативно, антитіла, побудовані шляхом генної рекомбінації, можна також застосовувати доти, поки зберігається задовільна антиген-зв'язувальна активність. Приклади антитіл, побудованих шляхом генної рекомбінації, включають химерні антитіла, CDR-трансплантовані антитіла, Fvs з єдиним ланцюгом, діатіла, лінійні антитіла та поліспецифічні антитіла, утворені з фрагментів антитіл. Загально відомо, що ці антитіла можна отримати, застосовуючи моноклональні антитіла. 17 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 У цьому винаході антитіла можна за необхідністю модифікувати. Згідно з цим винаходом антитіло, яке розпізнає позаклітинний домен ILT7 людини, має інгібувальний вплив на активність ІРС. Тобто, передбачається, що само антитіло має цитотоксичність проти ІРС. Підкласи антитіл, які демонструють сильну ефекторну активність, є відомими. Альтернативно, інгібувальну дію на активність ІРС можна далі підсилити шляхом модифікування антитіл цитотоксичним агентом. Приклади цитотоксичних агентів описані далі. Токсини: ендотоксин синьогнійного сепсису, токсин дифтерії, рицин. 99m 89 131 90 Радіоізотопи: Tc , Sr , I , Y . Протиракові агенти: каліхеаміцин, мітоміцин, паклітаксел. Токсини, що складаються з білків, можна кон'югувати з антитілами або їх фрагментами за допомогою біфункціонального реагенту. Альтернативно, токсин, що кодує ген, з'єднується з антитілом, що кодує ген, і злиті білки обох генів можна також отримати. Спосіб кон'югації антитіл з радіоізотопами є також відомим. Наприклад, відомим є спосіб мічення антитіл радіоізотопами, застосовуючи хелатоутворювальний агент. Крім того, протиракові агенти можна кон'югувати з антитілами, застосовуючи цукрові ланцюги або біфункціональний реагент. Автори цього винаходу підтвердили феномен, при якому моноклональне антитіло, що зв'язується з ILT7, що експресується на клітинній мембрані, включається у клітини після зв'язування (інтерналізації). Отже, цитотоксичні агенти можна доставити у клітини, здійснивши контакт антитіл, кон'югованих з цитотоксичними агентами цього винаходу, з клітинами, що експресують ILT7. Тобто, цим винаходом пропонується активний інгібітор клітин, що експресують ILT7, який включає моноклональне антитіло проти ILT7, з яким цитотоксичний агент кон'югується як активний інгредієнт. Або цей винахід стосується застосування моноклонального антитіла проти ILT7, з яким цитотоксичний агент кон'югується, у виробництві активного інгібітора клітин, що експресують ILT7. Далі, цим винаходом пропонується спосіб інгібування активності клітин, що експресують ILT7, який включає етап введення моноклонального антитіла проти ILT7, з яким кон'югується цитотоксичний агент. У цьому винаході антитіло, структура якого штучно модифікується, можна також застосовувати як активний інгредієнт. Наприклад, відомо багато способів модифікації, призначених для покращення цитотоксичності та стабільності антитіл. Специфічно, відомим є імуноглобулін, у якому модифікуються цукрові ланцюги важких ланцюгів (Shinkawa, Т. et al. J. Biol. Chem. 278:3466-3473. 2003). Активність імуноглобуліну щодо опосередкованої клітинами залежної від антитіла цитотоксичності (ADCC) підвищили шляхом модифікації цукрових ланцюгів. Або імуноглобулін, у якому амінокислотна послідовність ділянки Fc модифікується, також відомий. Тобто, активність ADCC була поліпшена шляхом штучного підвищення зв'язувальної активності імуноглобуліну з рецептором Fc (Shield, RL. et al. J. Biol. Chem. 276; 6591-6604, 2001). IgG, що зв'язується з рецептором Fc, включається у клітини один раз. Потім IgG зв'язується з рецептором Fc, що експресується в ендосомі, та він виділяється у кров знов. Цей феномен було визначено. IgG з високою зв'язувальною активністю з рецептором Fc має кращий шанс бути виділеним у кров знов після включення його у клітини. Внаслідок цього час утримання IgG у крові подовжується (Hinton, PR. et al. J Biol Chem. 279: 6213-6216, 2004). Окрім цього, говорять, що модифікація амінокислотної послідовності ділянки Fc спричиняє зміну активності комплемент-залежної цитотоксичності (CDC). Ці модифіковані антитіла можна застосовувати як антитіло цього винаходу. Коли антитіло, що зв'язується із позаклітинним доменом ILT7 людини, контактує з ІРС, тоді активність ІРС інгібується. Завдяки цьому антитіла можна застосовувати для інгібітора або для способу інгібування активності ІРС. Тобто, цим винаходом пропонується активний інгібітор ІРС, який включає принаймні один компонент, вибраний з групи, яка складається з перелічених далі від (а) до (с), як активний інгредієнт. Або цей винахід стосується способу інгібування активності ІРС, який включає етап введення принаймні одного компонента, що вибирають з групи, яка складається з перелічених далі від (а) до (с). Далі, цей винахід стосується застосування принаймні одного компонента, вибраного з групи, що складається з перелічених далі від (а) до (с), у виробництві активного інгібітора ІРС: (a) моноклонального антитіла, що зв'язується з ILT7 людини, або фрагмента, що включає його антиген-зв'язувальну ділянку; (b) імуноглобуліну, до якого гіперваріабельна ділянка моноклонального антитіла, описаного у (а), трансплантована, або фрагмента, що включає його антиген-зв'язувальну ділянку; та (c) полінуклеотиду, що кодує компоненти, описані у (а) або (b). У цьому винаході моноклональне антитіло, що розпізнає позаклітинний домен ILT7 людини, можна застосовувати як моноклональне антитіло, що інгібує активність ІРС. У цьому винаході 18 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 можна застосовувати одне або більше моноклональних антитіл. Наприклад, одне або більше моноклональних антитіл, що розпізнають позаклітинний домен ILT7 людини, змішують для застосування у цьому винаході. Можна підтвердити, що антитіла мають інгібуючу дію на виробництво інтерферону ІРС, таку як описано далі. ІРС виробляють велику кількість інтерферону внаслідок вірусної стимуляції. Антитіла додають до ІРС до, після або під час стимуляції ІРС вірусами. Здатність кожної такої клітини ІРС, що виробляє інтерферон, порівнюється зі здатністю виробляти інтерферон кожної контрольної клітини ІРС, до якої не додавали антитіла. Здатність виробляти інтерферон можна оцінити шляхом вимірювання IFN та IFN, що містяться у культуральній надосадовій рідині ІРС. Внаслідок порівняння можна підтвердити, що антитіла, які тестуються, є ефективними при інгібуванні здатності виробляти інтерферон, коли кількість інтерферону у надосадовій рідині суттєво знижується внаслідок додавання антитіл. Ці способи вимірювання інтерферону є відомими. ІРС виробляють більшу частину інтерферону у живому організмі. Отже, стан виробництва інтерферону у живому організмі можна регулювати шляхом інгібування здатності ІРС виробляти інтерферон. У цьому винаході активність ІРС охоплює підтримання кількості ІРС. Внаслідок цього, інгібування активності ІРС у цьому винаході включає інгібування кількості ІРС. Коли підтверджується, що кількість ІРС зменшується у присутності антитіл, можна визначити, що антитіла інгібують активність ІРС. Стосовно виробництва інтерферону, інертний імуноглобулін, що походить з такого ж самого тваринного зразка, що і антитіло, активність якого слід підтвердити, можна застосовувати як порівняльний контрольний імуноглобулін. Кількість ІРС можна кількісно порівняти шляхом рахування клітин. Кількість клітин можна рахувати за допомогою клітинного сортера з активацією флюоресценції або за допомогою мікроскопа. Далі, говорять, що внаслідок інфекції вірусом або йому подібним ІРС диференціюються у клітини, що індукують Th2, які називаються дендритними клітинами 2 (DC2). Якщо виробництво інтерферону ІРС внаслідок вірусної стимуляції можна інгібувати, їх диференціювання у Th2 можна також інгібувати. Таким чином, можна очікувати, що моноклональне антитіло цього винаходу, яке інгібує виробництво інтерферону, може також мати терапевтичний вплив на різні алергічні хвороби. Коли антитіло, що розпізнає позаклітинний домен ILT7 людини, вводиться хазяїну, відмінному від виду організму, з якого походить антитіло, бажано надати антитілу форму, яку хазяїну важко розпізнавати як сторонню речовину. Наприклад, імуноглобулін не можна легко розпізнати як сторонню речовину, перетворивши антитіло у перелічені далі молекули (спосіб процесингу молекул імуноглобуліну, який описано далі, є відомим): - фрагмент, що включає антиген-зв'язувальну ділянку, у якій не вистачає константної ділянки (Monoclonal Antibodies: Principles and Practice, third edition, Academic Press Limited. 1995; Antibody Engineering, A Practical Approach, IRL PRESS, 1996); - химерне антитіло, що складається з антиген-зв'язувальної ділянки моноклонального антитіла та константної ділянки імуноглобуліну хазяїна ("Gene Expression Experiment Manual", Isao Ishida, Tamie Ando, eds., Kodansha, 1994); - CDR-заміщене антитіло, у якому гіперваріабельна ділянка (CDR) імуноглобуліну хазяїна замінюється на CDR моноклонального антитіла ("Gene Expression Experiment Manual", Isao Ishida, Tamie Ando, eds., Kodansha, 1994). Альтернативно, антитіло людини можна отримати шляхом використання тварин, які не є людиною, у які поміщений ген антитіла людини, як імунних тварин, незважаючи на те, що застосовуються тварини, які не є людиною. Наприклад, з метою отримання антитіл людини, трансгенних мишей з генами антитіла людини застосовували на практиці як імунних тварин (Ishida et al., Cloning and Stem Cells, 4:85-95, 2002). Застосування таких тварин дозволяє отримати антитіло людини, яке розпізнає ILT7, застосовуючи імуногени, як описано раніше. Переважно вводити антитіло людини людям. Альтернативно, ген варіабельної ділянки імуноглобуліну людини можна також отримати за способом фагової індикації (McCafferty J. et al., Nature 348:552-554, 1990; Kretzschmar Τ et. al., Curr Opin Biotechnol. 2002 Dec; 13(6): 598-602). У способі фагової індикації ген, що кодує варіабельну ділянку імуноглобуліну людини, включається у ген фага. Бібліотеку фагів можна також отримати шляхом застосування різних генів імуноглобуліну як додатків. Фаг експресує варіабельну ділянку як злитий білок складеного білка фага. Варіабельна ділянка, що експресується фагом на поверхні фага, зберігає зв'язувальну активність з антигеном. Внаслідок цього вибираються фаги, що зв'язуються з антигенами або клітинами, у яких антигени експресуються, тим самим дозволяючи вибрати з бібліотеки фагів фаг, у якому експресується варіабельна ділянка, що має бажану зв'язувальну активність. Далі, ген, що кодує варіабельну 19 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 ділянку, яка має бажану зв'язувальну активність, утримується у вибраних таким чином фагових частинках. Тобто, у способі фагової індикації можна отримати ген, що кодує варіабельну ділянку з бажаною зв'язувальною активністю, використовуючи зв'язувальну активність варіабельної ділянки як індикатор. В активному інгібіторі ІРС або у способі інгібування активності ІРС цього винаходу антитіло, що розпізнає позаклітинний домен ILT7 людини, або фрагмент антитіла, який включає принаймні антиген-зв'язувальну ділянку антитіла, можна вводити як білок або як полінуклеотид, що кодує білок. Стосовно введення полінуклеотидів бажано застосовувати вектор, у якому полінуклеотид, що кодує бажаний білок, розташовується під контролем відповідного промотору, так щоб експресувати бажаний білок. У векторі можна також розташувати енхансер та термінатор. Відомим є вектор, що є носієм гена важкого та легкого ланцюгів, що створює імуноглобулін та є здатним експресувати молекулу імуноглобуліну. Вектор, здатний експресувати імуноглобулін, можна вводити шляхом введення його у клітини. При введенні до живих організмів вектор, яким можна інфікувати шляхом введення у живі організми разом з клітинами, можна вводити й безпосередньо. Якщо з живих організмів виділяються лімфоцити, тоді вектор вводиться у лімфоцити, які можна знов повернути у живі організми (ex vivo). В активному інгібіторі ІРС або у способі інгібування активності ІРС на основі цього винаходу, з точки зору кількості моноклонального антитіла, яке слід вводити живим організмам, імуноглобулін вводиться звичайно у діапазоні від 0,5 мг до 100 мг, наприклад, від 1 мг до 50 мг, переважно від 2 мг до 10 мг на кг маси тіла. Інтервали введення антитіла живим організмам можна належно відрегулювати, так щоб у живих організмах підтримувалася ефективна концентрація імуноглобуліну протягом періоду лікування. Специфічно, наприклад, антитіло можна вводити з інтервалами від 1 до 2 тижнів. Спосіб введення є довільним. Фахівці у галузі можуть вибрати належний ефективний спосіб введення під час лікування. Специфічні приклади способів введення включають пероральне введення або парентеральне введення. Антитіла вводять системно або локально, наприклад, шляхом внутрівенної ін'єкції, внутрішньом'язової ін'єкції, внутрішньочеревної ін'єкції, підшкірної ін'єкції тощо. Приклади відповідних лікарських форм для парентерального введення у цьому винаході включають розчини для ін'єкцій, супозиторії та спреї. Коли антитіло вводиться до клітин, імуноглобулін додають до культурального середовища звичайно у діапазоні 1 мкг/мл, переважно 10 мкг/мл, більш переважно 50 мкг/мл, найбільш переважно 0,5 мг/мл. В активному інгібіторі ІРС або у способі інгібування активності ІРС цього винаходу моноклональне антитіло можна вводити живим організмам довільними способами. Звичайно моноклональне антитіло змішується з фармацевтично прийнятною основою. За необхідністю моноклональне антитіло можна змішувати з додатковими агентами, такими як загусники, стабілізатори, консерванти та солюбілізатори. Приклади такої основи або додаткового агента включають лактозу, лимонну кислоту, стеаринову кислоту, магнію стеарат, сахарозу, крохмаль, тальк, желатин, агар, рослинну олію та етиленгліколь. Термін "фармацевтично прийнятний" означає ухвалені компетентним органом кожного уряду або перелічені у фармакопеї у кожній країні або інші загально визнані фармацевтичні агенти для застосування щодо тварин, ссавців та, зокрема, до людей. Активний інгібітор ІРС цього винаходу може також бути у формі єдиної дози або багаторазових доз ліофілізованих порошків або таблеток. Далі, ліофілізовані порошки або таблетки можна застосовувати у комбінації зі стерильною водою для ін'єкцій, з фізіологічним сольовим розчином або буферним розчином для розчинення композицій, щоб досягти бажаної концентрації перед введенням. Далі, коли моноклональне антитіло вводиться як вектор, який експресує імуноглобулін, тоді важкий та легкий ланцюги спільно трансфектуються до іншої плазміди, та кожну плазміду можна вводити у діапазоні від 0,1 до 10 мг, наприклад, від 1 до 5 мг на кілограм маси тіла. Для того, щоб ввести плазміди у клітини in vitro, вміст векторів для застосування становить від 1 до 5 6 мкг/10 клітин. Далі у цьому описі цей винахід буде специфічно описано з посиланням на Приклади. Усі документи з попереднього рівня техніки, які процитовано у цьому описі, включено у їх повному обсязі шляхом посилання. Приклади Приклад 1 А. Аналіз експресії ILT7 А-1) Аналіз із застосуванням бібліотеки SAGE Експресію генів у моноцитах, ІРС та оброблених вірусом простого герпесу (HSV) ІРС людини порівнювали та проаналізували, застосовуючи спосіб серійного аналізу експресії генів 20 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 (торгівельна назва; SAGE). Спосіб аналізу полягає у наступному. Моноцити виділили як BDCA4-позитивні клітини, а ІРС виділили як CD14-позитивні клітини з периферійної крові людини, застосовуючи клітинний сортер. Далі, ІРС культивували протягом 12 годин у присутності вірусу простого герпесу (HSV), а потім приготували диференційовані ІРС. З відповідних клітин отримали РНК, потім приготували бібліотеку SAGE, застосовуючи набір I-SAGE (торгівельна назва) (вироблено Invitrogen). Дані на отриманих послідовностях основ з приблизно 100000 міток проаналізували, застосовуючи аналітичне програмне забезпечення SAGE (розроблено Invitrogen). Внаслідок цього, виявили ген, у якому значення кількості моноцитів/IPC/IPC+HSV становить 0/16/0, а саме ILT7 (Gen Bank Acc#NM_012276), відомий як ген, що демонструє ІРСспецифічну експресію. ILT7 - це мембранний білок з імуноглобулін-подібними доменами, що кодується послідовністю основ, наведеною на SEQ ID NO: 1 (Фіг. 2(а)). Повідомлялося, що мРНК ILT7 експресується ІРС (Blood 100, 3295-3303 (2002)). А-2) Полімеразна ланцюгова реакція (ПЛР) зі зворотною транскриптазою (RT PCR) Експресію ILT7 у клітинах-гемоцитах досліджували більш детально. Кожну клітину заздалегідь виділили з периферійної крові людини за допомогою клітинного сортера. РНК екстрагували з кожної з виділених клітинних популяцій, з яких синтезували кДНК. Кількісну ПЛР здійснювали за звичайним способом, застосовуючи отриману кДНК як матрицю, та проаналізували рівень експресії мРНК ILT7. Умови, що застосовувалися для послідовностей основ праймерів та ПЛР, є наступними: Передній праймер: 5' СТС САА ССС СТА ССТ GCT GTC 3' (SEQ ID NO: 3), Зворотній праймер: 5' ТТС CCA AGG СТС САС САС ТСТ 3' (SEQ ID NO: 4), 1 цикл ПЛР (при 94°С протягом 3 хвилин), 25 циклів ПЛР [94°С протягом 30 секунд, при 58°С протягом 30 секунд та при 72°С протягом 1 хвилини], 1 цикл ПЛР (при 72°С протягом 6 хвилин). Коли досліджували ІРС, стимульовані моноцитами, ІРС, HSV, та CD19-позитивні клітини (тобто В-клітини), CD3-позитивні клітини (тобто Т-клітини), Т-клітини, стимульовані РМА, та CD56-позитивні клітини (тобто NK-клітини), визначили, що ILT7 експрєсувався специфічно у ІРС (Фіг. 1(а)). А-3) Кількісна RT PCR Далі експресію в інших органах та тканинах досліджували шляхом кількісної ПЛР, застосовуючи АВІ PRISM 7000 (виробляється Applied Biosystem). Як панель кДНК застосовували панель кДНК численної тканини МТС BD (торгівельна назва) (Human I; Cat.No.636742, Human immune; Cat.No.636748, фракції крові людини; Cat.No.636750; усі вони вироблені Becton Dickinson), та таку ж саму кДНК, що походить з клітин-гемоцитів, як описано у 2). Використані послідовності основ прайм єрів є наступними: Передній праймер для ILT7: 5' ССТ САА ТСС AGC АСА ААА GAA GT 3' (SEQ ID NO: 5), Зворотній праймер для ILT7: 5' CGG ATG AGA ТТС ТСС ACT GTG ТАА 3' (SEQ ID NO: 6), Передній праймер для GAPDH: 5' CCA CCC ATG GCA ААТ ТСС 3' (SEQ ID NO: 7), Зворотній праймер для GAPDH: 5' TGG GAT ТТС CAT TGA TGA САА G 3' (SEQ ID NO: 8). ПЛР здійснювали, застосовуючи АВІ PRISM 7000 (виробляється Applied Biosystem) та набір SYBR green PCR master mix (виробляється тією ж самою компанією). Для аналізу застосовували програмне забезпечення Sequence Detection System Software (розроблено тією ж самою компанією). Умови реакції є наступними: Етап 1: 1 цикл ПЛР (при 50°С протягом 2 хвилин), Етап 2: 1 цикл ПЛР (при 95°С протягом 10 хвилин), Етап 3: 40 циклів ПЛР (при 95°С протягом 15 секунд, при 60°С протягом 1 хвилини). Експресію гена ILT7 порівнювали між кожною тканиною шляхом стандартизації рівня експресії гена гліцеральдегід-3-фосфат дегідрогенази (GAPDH), про який відомо, що він експресується конститутивно. Внаслідок цього спостерігали, що ILT7 не експресується у будьяких органах, відмінних від лімфатичних тканин, та експресується специфічно у ІРС. В. Виробництво векторів експресії ILT7 та FcR Далі здійснювали клонування генів та виробництво векторів експресії з метою експресувати білки ILT7. В-1) Клонування генів ILT7 + Роlу(А) РНК, виділену з периферійної крові людини, екстрагували з ІРС, з чого синтезували кДНК, застосовуючи праймер oligo dT та Super Script Choice System для набору синтезу кДНК. Адаптер EcoRI вшили у синтезовану кДНК, яку вшили у вектор pME18S, розщеплений EcoRI, 21 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 внаслідок чого отримали бібліотеку кДНК ІРС людини. Ген ILT7 ампліфікували за способом ПЛР, застосовуючи отриману бібліотеку кДНК як матрицю, а також застосовуючи праймери з наступними послідовностями основ. 1 одиницю ДНК-полімерази KOD Plus (виробляється TOYOBO CO., LTD.) застосовували для ПЛР. Умови реакції були наступними: 25 циклів ПЛР [при 94°С протягом 15 секунд, при 55°С протягом 30 секунд та при 68°С протягом 2 хвилин] після 1 циклу ПЛР при 94°С протягом 2 хвилин. Передній праймер: 5' CAG GGC CAG GAG GAG GAG ATG 3' (SEQ ID NO: 9). Зворотній праймер: 5' TCA GCA GAC ACT TCC CCA ACT 3' (SEQ ID NO: 10). Ампліфікований фрагмент з 2 т.п.н. кДНК ILT7 виділили та одержали шляхом електрофорезу, застосовуючи 1% агарозний гель, а потім клонували у вектор плазміди pCR4 Blunt-TOPO (виробляється Invitrogen), застосовуючи набір Zero Blunt TOPO PCR Cloning (виробляється Invitrogen). Послідовності основ отриманих генів проаналізували та визначили, що отримали бажаний ген ILT7, наведений у SEQ ID NO: 1. В-2) Виробництво векторів експресії FLAG-міченого ILT7 Відповідно побудували плазміду, що експресує білок, де мітки FLAG було злито з N-та Сзакінченнями ILT7. ILT7 злили з міткою, що дозволило підтвердити експресію білка ILT7 завдяки виявленню мітки. Бажану послідовність ампліфікували за способом ПЛР, застосовуючи ген ILT7, отриманий так, як описано у (1), як матрицю, а також застосовуючи праймери з наступними послідовностями основ. 1 одиницю ДНК-полімерази KOD Plus (виробляється TOYOBO CO., LTD.) застосовували для ПЛР. Умови реакції були наступними: 25 циклів ПЛР [при 94°С протягом 15 секунд, при 55°С протягом 30 секунд та при 68°С протягом 2 хвилин] після 1 циклу ПЛР при 94°С протягом 2 хвилин. Для N-FLAG ILT7 Передній праймер (SEQ ID NO: 11): 5' CCG etc gag ATG ACC CTC ATT CTC АСА AGC CTG CTC TTC TTT GGG CTG AGC CTG GGC [GAT TAC AAG GAT GAC GAC GAT AAG] CCC AGG ACC CGG GTG CAG GCA GAA 3'. Зворотній праймер (SEQ ID NO: 12): 5' С TAG act agt TCA GAT CTG TTC CCA AGG CTC 3'. Для C-FLAGI LT7 Передній праймер (SEQ ID NO: 13): 5' CCG etc gag ATG ACC CTC ATT CTC АСА AGC 3'. Зворотній праймер (SEQ ID NO: 14): 5' С TAG act agt ТСА [СТТ АТС GTC GTC АТС CTT GTA ATC] GAT CTG TTC CCA AGG CTC 3'. У вищезгаданих послідовностях основ кожна підкреслена частина у дужках показує послідовність основ, що кодує приєднану мітку FLAG, а кожна маленька літера показує сайт розщеплення для рестрикційного ферменту Xhol або Spel. Фрагменти ДНК, ампліфіковані ПЛР, розщепили Xhol та Spel, які потім відокремили шляхом гель-електрофорезу. Одержали фрагменти ДНК 2 т.п.н., а потім їх вшили у вектор рМЕ18Х, розщеплений Xhol та Spel за таким самим способом, як описано вище. Потім побудували два типи плазмід, здатних експресувати бажаний злитий білок, тобто pME18X-N-FLAG ILT7 та рМЕ18 X-C-FLAG ILT7, відповідно. В-3) Клонування генів FcR Білок FcR вважали білком, здатним зв'язуватися з білком ILT7. Ця молекула є геном з послідовностями основ та амінокислотними послідовностями SEQ ID NO: 15 та 16 (Genbank Acc#NM_004106, J. Biol. Chem. 265, 6448-6452 (1990)). Ця молекула є молекулою ( ланцюг), що утворює FcRI, тобто рецептор IgE з високим афінітетом. Незважаючи на те, що його також позначають як FcRI , далі у цьому описі його позначають як FcR. Стосовно цього, ця молекула також відома як компонент FcR або FcR. З метою отримання векторів експресії, цей ген клонували за способом ПЛР, як показано далі. Ген FcR ампліфікували за способом ПЛР, застосовуючи бібліотеку кДНК ІРС людини, отриману так, як описано у (1), як матрицю, а також застосовуючи праймери з наступними послідовностями основ. 1 одиницю ДНК-полімерази KOD Plus (виробляється TOYOBO CO., LTD.) застосовували для ПЛР. Умови реакції були наступними: 25 циклів ПЛР [при 94°С протягом 15 секунд, при 55°С протягом 30 секунд та при 68°С протягом 1 хвилини] після 1 циклу ПЛР при 94°С протягом 2 хвилин. Передній праймер: 5' ССС AAG ATG ATT CCA GCA GTG 3' (SEQ ID NO: 17). j Зворотній праймер: 5' GGA AGA АСС AGA AGC САА AGA 3' (SEQ ID NO: 18). | Ампліфікований фрагмент FcRкДHK 0,3 т.п.н. виділили та одержали електрофорезом, застосовуючи 2% агарозний гель, потім його клонували до плазмідного вектора pCR4 BluntTOPO (виробляється Invitrogen), застосовуючи набір Zero Blunt TOPO PCR Cloning (виробляється Invitrogen). Послідовності основ отриманих генів проаналізували та підтвердили, що бажаний ген FcR, наведений на SEQ ID NO: 15, клонувався. В-4) Виробництво векторів експресії Мус-міченого FcR 22 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 Плазміду, що експресує білок, де мітку Мус приєднали до С-закінчення, побудували так, щоб можна було б підтвердити експресію білка FcR. Бажану послідовність ампліфікували за способом ПЛР, застосовуючи ген FcR, отриманий так, як описано у (3), як матрицю, а також застосовуючи праймери з наступними послідовностями основ. 1 одиницю ДНК-полімерази KOD Plus (виробляється TOYOBO CO., LTD.) застосовували для ПЛР. Умови реакції були наступними: 25 циклів ПЛР [при 94°С протягом 15 секунд, при 55°С протягом 30 секунд та при 68°С протягом 1 хвилини] після 1 циклу ПЛР при 94°С протягом 2 хвилин. Передній праймер: (SEQ ID NO: 19): 5' CCG etc gag ATG ATT CCA GCA GTG GTC TTG 3'. Зворотній праймер: (SEQ ID NO: 20): 5' СТА Gac tag tCT А[СА GAT CCT CTT CAG AGA TGA GTT TCT GCT C]CT GTG GTG GTT TCT CAT G 3'. У вищезгаданих послідовностях праймерів підкреслена частина у дужках показує послідовність основ, що кодує приєднану мітку Мус, а кожна маленька літера показує сайт розщеплення для рестрикційного ферменту Xhol або Spel. Фрагменти ДНК, ампліфіковані ПЛР, розщепили Xhol та Spel, які потім відокремили шляхом гель-електрофорезу. Фрагменти ДНК приблизно 0,3 т.п.н. одержали, а потім вшили у вектор рМЕ18Х, розщеплений Xhol та Spel за таким самим способом, як описано вище. Потім побудували плазміду, здатну експресувати бажаний злитий білок, тобто рМЕ18Х-Мус-FcR. С. Експресія ILT7 у тваринних клітин Експресію ILT7 у тваринних клітин досліджували, застосовуючи вектори експресії, отримані так, як описано вище. С-1) Експресія у клітин 293Т 5 ДНК, що складаються з наступних п'яти комбінацій, ввели у клітини 293Τ (7xΙ10 клітин), застосовуючи набір трансфекції effectene (виробляється Qiagen). Через два дні після введення здійснили аналіз проточної цитометрії (аналіз FCM). (1) pME18X-N-FLAG ILT7 2 мкг (2) pME18X-C-FLAG ILT7 2 мкг (3) pME18X-N-FLAG ILT7 1 мкг + pME18X-Myc-FcR 1 мкг (4) pME18X-C-FLAGILT7 1 мкг + pME18X-Myc-FcR 1 мкг (5) pME18X-Myc-FcR 2 мкг Спосіб аналізу проточної цитометрії виконували таким самим чином, як описано у А-4 з наступного Прикладу 2. Су3-кон'юговане анти-Flag антитіло (виробляється Sigma) застосовували для реакції та FACScan (виробляється Becton Dickinson) застосовували для аналізу. Внаслідок цього визначили, що тільки невелика кількість ILT7 експресувалася на клітинній поверхні, коли у реакції брало участь тільки антитіло, проте ILT7 експресувався за межами клітини та суттєво у присутності FсR (Фіг. 3). Відомо, що FсR миші має високу гомологію з FcR людини. Проте, коли клітини р815 (мастоцитома миші), які експресують FсR миші, застосовувалися як хазяїни, експресію ILT7 не могли спостерігати. С-2) Аналіз способами імунопреципітації та вестерн-блотингу ILT7 експресувався у супроводі FcRyira клітинній поверхні, що підтвердили наступним способом. Після імунопреципітації проаналізували різні антитіла для кожної клітини 293Т, яка експресувалася спільно з обома генами у відповідних комбінаціях, описаних у від (1) до (5). ДНК 5 ввели у клітини 293Τ (7x10 клітин), де клітини 293Τ одержали через два дні після введення за таким самим способом, як описано у (1). Клітинні фракції розчинили у лізисному буфері (0,5% Triton, 150 мМ NaCl), який залишили на льоду на 20 хвилин. Після цього декілька разів повторювали аспірацію, застосовуючи голку (27G), потім центрифугували зі швидкістю 15000 обертів за хвилину протягом 20 хвилин. Анти-мус антитіло (2 мкг, виробляється Santa Cruz biotechnology) або анти-Flag антитіло (виробляється Sigma) додали до 200 мкг лізату отриманих продуктів, який потім перемішували шляхом обертання при 4°С протягом 4 годин. Потім суміш Protein A/G Sepharose 4 Fast Flow (виробляється Amersham bioscience) додали, а потім перемішували шляхом обертання при 4°С протягом 1 години. Потім отримані преципітовані фракції тричі промили лізисним буфером з наступним складом. Лізисний буфер: 0,5% TritonX-100, 50 мМ HEPES (pH 7,6), 150 мМ NaCl, 1 мМ EDTA, 10% гліцерин, 1 мМ DTT (дитіотреїтол), 2 мМ PMSF (фенілметилсульфонілфторид), 1 мкг/мл Апротиніну, 23 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 1 мкг/мл Леупептину, 1 мкг/мл Пепстатину А, 0,1 мкг/мл Хімостатину, 1 мМ Na3VO4, 0,1 мМ -гліцерофосфат. Зразок буферу для електрофорезу в поліакриламідному гелі у присутності додецилсульфату натрію (SDS-PAGE) додали до промитих осадів, а потім кип'ятили протягом 5 хвилин та центрифугували, потім здійснювали електрофорез на гелі з 10% додецилсульфату натрію. Зразки перенесли з гелю після електрофорезу до мембрани PVDF (мембрана Immobilon-p-transfer: виробляється Millipore) згідно зі звичайним способом. Блотинг виконували з анти-Flag антитілом та анти-мус антитілом. Підтвердили, що ILT7, пов'язаний з FcR, був присутнім у клітинах 293Т, тому що їх присутність спостерігали у кожному імунному преципітаті (Фіг. 4). С-3) Аналіз цукрового ланцюга Оскільки під час вестерн-аналізу спостерігали декілька смуг ILT7, то досліджували можливість того, що ILT7 глікозилувався. 200 мкг лізату клітин 293Т, що експресують NFLAGILT7 та Myc-FcR, піддали імунопреципітації анти-Flag антитілом за способом, описаним у (1) та (2). Після цього осаджені фракції суспендували у 60 мкл N-глікозидазному буферу з наведеним далі складом, та 30 мкл кожного отриманого розчину розподілили без залишків у дві трубки. N-глікозидазний буфер: 10 мМ EDTA, 0,2% додецилсульфат натрію (SDS), 0,5% TritonX100, 1% 2-меркаптоетанол у PBS (фосфатний буфер). Потім 3 одиниці з 3 мкл N-глікозидази (#1365177, виробляється Roche) додали до однієї пробірки, у якій відбувалася реакція при 37°С протягом 15 годин. Потім туди додали 7 мкл буферу зразка та нагрівали при 100°С протягом 5 хвилин, після чого здійснили електрофорез на гелі з 10% додецилсульфату натрію. Після електрофорезу гель перенесли до мембрани PVDF, до якої додали 1 мкг поліклонального антитіла проти ILT7, як описано у (4), та здійснювали реакцію при 4°С протягом ночі. Отриманий продукт промили буфером TBS-T та здійснили реакцію з розведеним у 100000 разів HRP-міченим анти-кролячим антитілом (виробляється Jackson) при кімнатній температурі. Потім його забарвили системою ECL Western Blotting Detection System (виробляється Amersham bioscience). Внаслідок цього очевидну молекулярну масу зменшили обробкою N-глікозидазою. Отже, очікували, що цукрові ланцюги додалися до ILT7 (Фіг. 5). С-4) Виробництво поліклонального антитіла проти ILT7 Поліклональне антитіло проти ILT7, що застосовувалося, як описано у (3), отримали за наступним способом. Пептид з 23 амінокислот, що відповідають С-закінченню ILT7 (CSQEANSRKDNAPFRWEPWEQI; SEQ ID NO: 21), синтезували хімічним способом та зв'язали з білком KLH, який є носієм, а отриманий продукт застосовували як імуноген. Кролів крізьшкірно імунізували імуногеном, змішаним з повним ад'ювантом Фрейнда. Після всього шести імунізацій (одна на тиждень) підтвердився підвищений титр антитіла у сироватці, та потім зібрали суцільну кров. Потім деяку кількість сироватки афінно очистили, застосовуючи пептидну колонку такої ж самої послідовності. Отриманий продукт визначили як поліклональне антитіло проти ILT7. Приклад 2 А. Виробництво моноклонального антитіла проти ILT7 А-1) Виробництво імуногена Клітини, які слід застосовувати як імуногени, приготували шляхом введення генів у клітини 293Т за способом, який описано далі. 46,4 мкг трансгена (рМЕ18 X-C-FLAG ILT7 23,2 мкг та ME18X-Myc-FcR 23,2 мкг) додали до дна 100 мм покритої колагеном чашки (IWAKI), покритої 3 мл opti-MEM (GIBCO), та змішали. Після цього азид з трангенного розчину, 58 мкл Lipofectamine (торгівельна назва) 2000 (Invitrogen) розвели 3 мл opti-MEM та залишили при кімнатній температурі на 5 хвилин, та приготували розчин Lipofectamine. Після цього розчин Lipofectamine повільно додавали до чашки, що містила трансгенний розчин, та перемішували. Після витримування протягом 20 хвилин при кімнатній температурі 10 мл клітин 293Т, розведених до 6 1x10 клітин/мл із застосуванням культурального середовища DMEM (SIGMA), що містило 10% FBS (фетальної бичачої сироватки), повільно додавали до чашки. Отримане середовище піддали статичному культивуванню в інкубаторі при 37°С у присутності СО 2 протягом 48 годин. 24 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 З цього середовища клітини отримали за допомогою піпетки. Отримані клітини застосовували як трансфектанти для імуногенів. А-2) Виробництво гібридом За день до імунізації клітин 50 мкл емульсії, отриманої шляхом змішування 200 мкл фізіологічного розчину з фосфатним буфером (PBS) з 200 мкл повного ад'юванту Фрейнда (FREUND) (RM606-1, виробляється Mitsubishi Kagaku Iatron, Inc.), ввели шляхом ін'єкції у нижні частини обох ніг чотирьох самок мишей Balb/c (віком чотири тижні) для імунізації. Наступного 7 дня імунізували 50 мкл 2x10 клітин, суспендованих у 400 мкл фізіологічного розчину з фосфатним буфером. Другу та третю імунізації виконували кожні чотири дні. Через три дні після третьої імунізації здійснювали злиття клітин наступним способом. Клітини зібрали з лімфатичних вузлів мишей, імунізованих у ногу. Клітини мієломи мишей P3-X63-Ag8-U1, культивовані у культуральному середовищі RPMI1640 (SIGMA), що містить 10% FBS, змішали з клітинами, що походять з лімфатичних вузлів та мієломи, так щоб відношення клітин мієломи мишей до клітин, що походять з лімфатичних вузлів, становило від 2:1 до 10:1. З цього середовища клітини одержали шляхом центрифугування. PEG4000 (MERCK), еквівалентно розведений культуральним середовищем RPMI1640, додали до отриманих клітинних фракцій, які потім піддали клітинному злиттю. Після промивання клітин отриманий продукт суспендували у 160 мл 15% середовища FBS-HAT, що містило конглютинін, а потім його інокулювали у 16 96лункових планшетів по 200 мкл/лунку. Культуральне середовище поміняли через три дні. Через один-два тижні після спостереження утворення колонії виконали первинний скринінг. А-3) Скринінг гібридоми способом ЕЛАЙЗА для клітин Гібридому, що виробляє антитіло-мішень, скринували за наступним способом ЕЛАЙЗА для 7 клітин. Отримані клітини, як описано у (1), застосовували по 1x10 клітин на 96-лунковий планшет, їх суспендували у 0,5% BSA/2 мМ EDTA/PBS, а потім розподілили без залишку у чашці для ЕЛАЙЗА клітин (NUNC 249570 96V NW PS) по 100 мкл/лунку. Центрифугування здійснювали зі швидкістю 2000 обертів за хвилину при 4°С, а потім надосадову рідину відкинули. Культуральну надосадову рідину зразка додали по 50 мкл/лунку, реакція здійснювалася при кімнатній температурі протягом 30 хвилин. Двічі здійснювали операцію промивання, яка включає додавання 0,5% BSA/2 мМ та EDTA/PBS до кожної лунки, центрифугування зі швидкістю 2000 обертів за хвилину при 4°С протягом 2 хвилин та відкидання надосадової рідини. 50 мкл/лунку розведеного у 10000 разів міченого пероксидазою анти-мишачого IgG антитіла козла (ІМ0819; Beckman coulter) додали до кожної лунки після промивання, та реакція здійснювалася протягом 30 хвилин. Операцію промивання із застосуванням 0,5% BSA/2 мМ-EDTA/PBS виконували двічі, а потім додали забарвлювальний розчин. Отриманий розчин антитіла замістили фізіологічним розчином з фосфатним буфером () мембраною діалізу (10000 розрізів, вироблених PIERCE), внаслідок чого отримали очищені химерні антитіла проти ILT7. А-4) Дослідження реактивності антитіла аналізом проточної цитометрії (FCM) Культуральну надосадову рідину гібридоми проаналізували шляхом проточної цитометрії (FCM). Отримані клітини, які описано у (1), суспендували у 0,5% BSA/2 мМ EDTA/PBS, що потім 5 перенесли у пробірку для центрифугування при 1x10 на один зразок, а потім додали 40 мкл кожної культури. Реакцію здійснювали при кімнатній температурі протягом 30 хвилин. Двічі здійснювали операцію промивання, яка включає додавання до кожної пробірки 1 мл 0,5% BSA/2 мМ та EDTA/PBS, центрифугування при 1200 об./хв. при 4°С протягом 3 хвилин та відкидання надосадової рідини. 40 мкл розведеного у 100 разів міченого FITC анти-мишачого IgG антитіла козла (ІМ0819; Beckman coulter) додали до кожної лунки після промивання, та реакція здійснювалася протягом 30 хвилин при кімнатній температурі. Операцію промивання із застосуванням 0,5% BSA/2 мМ-EDTA/PBS виконували двічі, а потім аналізували із застосуванням проточної цитометрії FC500 (Beckman coulter). Вибрали гібридому, що виробляла антитіло, яке не реагувало на саму тільки клітину-хазяїна, а реагувало специфічно на клітину, у яку ввели ген. Вибрану гібридому клонували способом обмежувального розведення та отримали гібридоми #11 та #17, що виробляють моноклональні антитіла. В. Дослідження реактивності антитіла проти ILT7 ILT7, у якому мітку FLAG приєднали до N-закінчення, експресували спільно з молекулою FcR y клітинах 293Т за таким способом, як описано у (С-1) Прикладу 1. Потім реактивність антитіла, отриманого у Прикладі 2, підтвердили аналізом FCM, застосовуючи FACScan (Becton Dickinson). Внаслідок цього підтвердили, що усі антитіла, що виробляються гібридомами #11 та #17, які отримали так, як описано у (А), реагували на клітини, до яких ввели ген ILT7, та які експресували ILT7 (Фіг. 6b). Далі, лімфоцити виділили з периферійної крові людини, застосовуючи Ficoll, та потім виконували подвійне забарвлення отриманим антитілом проти 25 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 ILT7 та РЕ-міченим антитілом проти BDCA-2 (Miltenyi). Потім досліджували реактивність до лімфоцитів. Внаслідок цього виявили зв'язування моноклонального антитіла, виробленого гібридомами #11 та #17, з BDCА-2-позитивною клітиною. Тобто, підтвердили, що обидва моноклональні антитіла розпізнавали молекули ILT7, що експресуються на ІРС людини (Фіг. 6а). Ці моноклональні антитіла позначили як антитіло #11 проти ILT7 та антитіло #17 проти ILT7, відповідно. Виконали більш детальний аналіз. Аналіз за допомогою багаторазового забарвлення для лімфоцитів периферійної крові людини здійснювали із застосуванням отриманого антитіла проти ILT7, антитіла проти послідовності клітинних генерацій-1 (антитіла проти CD3, CD14, CD16, CD19, CD56; Becton Dickinson), антитіла проти CD123 (Becton Dickinson) та антитіла проти BDCA-2 (Miltenyi). Стосовно позитивних фракцій антитіла ILT7 маркер послідовності клітинних генерацій був негативним, стосовно CD123 був позитивним та стосовно BDCA-2 теж був позитивним. На підставі результатів підтвердили, що ІРС були забарвлені тільки ILT7#11 та ІLT7#17 (Фіг. 7). Далі, експресію різних молекул досліджували за допомогою аналізу проточної цитометрії, коли лімфоцити периферійної крові людини стимулювали за допомогою CpG або IFN протягом 24 годин. CpGODN2216 застосовували як CpGA, який індукує виробництво інтерферону з ІРС, та CpGODN2006 застосовували як CpGB, який сприяє визріванню дендритних клітин (Moseman et al. J. Immunology. 173, 4433-4442, 2004). Вікно було встановлено для негативної фракції маркера послідовності клітинних генерацій. Коли аналізували реактивність антитіла проти BDCA-2 та антитіла проти ILT7 на популяцію позитивних клітин CD123, більшість позитивних фракцій ILT7 зникла навіть після 24-годинної стимуляції за допомогою CpG. З іншого боку, деякі клітини BDCA-2 демонстрували позитивні фракції після 24-годинної стимуляції за допомогою CpG (Фіг. 8). Вважали, що ІРС диференціюються на різні клітини одразу після стимуляції за допомогою CpG. Вказали, що антитіло проти ILT7 цього винаходу є корисним як стадійно специфічне антитіло до ІРС. Далі підтвердили, що ІРС у лімфоцитах периферійної крові людини не диференціювалися у присутності INF, у тому випадку, коли коефіцієнт виживання був високим, експресія ILT7 зберігалася на ІРС, далі ILT7 був стабільно присутнім на ІРС при автоімунних хворобах з можливістю, що інтерферон у сироватці був на високому рівні. С. Дослідження специфічності антитіла проти ILT7 ILT7 належить до родини ILT/LIR, та існує багато молекул з високою гомологією, особливо з високою гомологією у позаклітинній ділянці (Фіг. 9). Повідомлялося, що мРНК молекул, особливо таких як ILT2 та ILT3, експресуються у ІРС (Ju et al. Gene 331, 159-164, 2004). Внаслідок цього реактивність цих молекул підтверджували із застосуванням трансгенних клітин. С-1) Клонування молекули ILT1 та виробництво векторів експресії кДНК синтезували з РНК, що походить з мигдалини людини, застосовуючи праймер oligo dT та Superscript Choice System для набору синтезу кДНК. Далі, адаптер NotI вшили у вектор pME18S, розщеплений NotI, внаслідок чого отримали бібліотеку кДНК мигдалини людини. Ген ILT1 з міткою FLAG на С-закінченні ампліфікували шляхом ПЛР, застосовуючи отриману бібліотеку кДНК як матрицю, а також застосовуючи праймери з наступними послідовностями основ. 1 одиницю полімерази ДНК KOD Plus (виробляється TOYOBO CO., LTD.) застосовували для ПЛР. Умови реакції були наступними: 25 циклів ПЛР [при 94°С протягом 15 секунд, при 55°С протягом 30 секунд та при 68°С протягом 2 хвилин] після 1 циклу ПЛР при 94°С протягом 2 хвилин. Передній праймер (SEQ ID NO: 22): 5' CCG etc gag ATG ACC CCC АТС CTC ACG GTC С 3'. Зворотній праймер (SEQ ID NO: 23): 5' СТА Gac tag tTC А[СТ TAT CGT CGT CAT CCT TGT AAT C]CC TCC CGG CTG CAT CTT G 3'. У вищезгаданих послідовностях праймерів кожна підкреслена частина у дужках показує послідовність основ, що кодує приєднану мітку FLAG, а кожна маленька літера показує сайт розщеплення для рестрикційного ферменту Xhol або Spel. Фрагменти ДНК, ампліфіковані ПЛР, розщепили Xhol та Spel, які потім відокремили шляхом гель-електрофорезу. Одержали фрагменти ДНК приблизно у 2 т.п.н., а потім їх вшили у вектор рМЕ18Х, розщеплений Xhol та Spel за таким самим способом, як описано вище. Потім побудували плазміду, здатну експресувати бажаний злитий білок, тобто рМЕ18Х-С-FLAGILT1. Послідовність основ та амінокислотну послідовність наведено у SEQ ID NOs: 24 та 25. С-2) Виробництво експресуючих клітин та дослідження реактивності антитіла Стосовно ILT2 (SEQ ID NO: 26) та ILT3 (SEQ ID NO: 28), застосовували вектори експресії, у яких відповідні гени клонувалися до сайтів ХbаІ або Xhol pcDNA4.1 (виробляється Invitrogen). 5 ДНК наступних комбінацій ввели у клітини 293Т (7x10 клітин) за таким способом, як описано у (С-1). Через два дні після введення здійснили аналіз проточної цитометрії, а потім проаналізували антитіло проти ILT7. 26 UA 97946 C2 5 10 15 20 25 30 35 40 45 50 55 60 (1) 1 мкг pMEl 8X-N-FLAG ILT7 + 1 мкг pME18X-Myc-FcR (2) 0,5 мкг pMEl 8X-C-FLAG ILT1 + 0,5 мкг pMEl 8X-Myc-FcR + 0,5 мкг pcDNA4.1-ILT2 + 0,5 мкг pcDNA4.1-ILT3 Внаслідок цього будь-які антитіла не реагували на клітини, у яких експресувався ILT1. Тому припустили, що ці антитіла проти ILT7 специфічно розпізнавали молекули ILT7 на ІРС (Фіг. 10). ' Приклад 3 Вплив антитіла проти ILT7 на здатність виробляти IFN людини 5 Лімфоцити периферійної крові людини інокулювали у 96-лунковому планшеті по 2x10 клітин/лунку, які вступили у реакцію з 5 мкг/мл різних антитіл при 37°С. Через одну годину культивування до реакції додали вірус грипу PR8. Через 24 години культивування вимірили IFN у культуральній надосадовій рідині за допомогою набору ЕЛАЙЗА (Bender Med System). Внаслідок цього виробництво інтерферону інгібувалося після додавання антитіла проти ILT7 (Фіг. 11). А саме, визначили, що на виробництво інтерферону ІРС впливало антитіло проти ILT7 цього винаходу. Приклад 4 CDC-активність антитіла проти ILT7 A. Виробництво моноклонального антитіла проти ILT7 Клон, що виробляє моноклональне антитіло, отримали за таким самим способом, як описано у від (А-1) до (А-4) Прикладу 2. Реактивність досліджували за таким самим способом, як описано у (В) Прикладу 2, а специфічність досліджували за таким самим способом, як описано у (С) Прикладу 2. Внаслідок цього отримали гібридоми #37, #28 та #33, які виробляли моноклональні антитіла проти ILT7 з гарною реактивністю та специфічністю. CDC-активність вимірювали так, як описано далі, застосовуючи моноклональне антитіло проти ILT7, де вироблялося три види цих гібридом. B. Визначення CDC-активності В-1) Попереднього дня перед цільовим виробництвом лінії клітин-мішеней (клітинна лінія 5 ILT7-CHO) наступну ДНК ввели у клітини CHO-k1, які інокулювали по 6х10 клітин на чашку (діаметром 6 см), застосовуючи Effectene Transfection Reagent (виробляється QLAGEN), а потім резистентні штами зібрали, застосовуючи 800 мкг/мл Zeocin (виробляється Invitrogen). Введена ДНК: 1 мкг pcDNA3.1-C-FLAG ILT7 + 2 мкг рМЕ18Х-Мус FcR. Після цього клітинну лінію, яка суттєво експресувала ILT7, отримали, застосовуючи клітинний сортер (BD FACSAria, виробляється Becton Dickinson). Аналізом проточної цитометрії підтвердили, що вибрана клітинна лінія суттєво експресувала ILT7. Операцію аналізу із застосуванням проточної цитометрії здійснювали згідно зі способом, описаним у (А-4) Прикладу 2, за виключенням того, що для проточної цитометрії застосовували BD FACSCaliber (виробляється BD). Наступні антитіла застосовували для первинного антитіла та вторинного антитіла, відповідно. Первинне антитіло: 5 мкг/мл антитіла проти ILT7 миші (#37). Вторинне антитіло: R-рикоеритрин (R-PE)-кон’юговане анти-мишаче імуноглобулінспецифічне поліклональне антитіло козла (BD). В-2) Реакція клітин-мішеней на антитіла проти ILT7 Отримані клітини-мішені, як описано у (В-1) (клітини ILT7-CHO), одержали, застосовуючи 5 мМ розчин EDTA/PBS, а потім суспендували у середовищі CDC з наведеним далі складом, так 5 щоб концентрація становила 4x10 клітин/мл. Суспензію розподілили без залишків на 96лунковому планшеті по 50 мкл/лунку. Середовище CDC: RPMI1640, 0,1% BSA (альбумін бичачої сироватки), 100 одиниць/мл пеніциліну, 100 мкг/мл стрептоміцину, 10 мМ Hepes (pH 7,6), 2 мМ L-глутаміну. 50 мкл розчину антитіла проти ILT7, приготовленого у середовищі CDC, додали до кожної лунки та змішали так, щоб кінцева концентрація антитіл становила 0,1 мкг/мл, 0,5 мкг/мл, 1 мкг/мл та 5 мкг/мл. Потім туди додали 50 мкл середовища CDC, що містило комплемент з наведеним далі складом, та змішали так, щоб кінцева концентрація комплементу становила 6%, а потім культивували при 37°С протягом 2 годин. Середовище CDC, що містило комплімент: 1 мл комплементу молодого кроля (Catalog No.: CL3441, виробляється CEDARLANE), Середовище CDC (див. вище). 27 UA 97946 C2 5 10 15 20 25 Потім суспензію центрифугували (умови центрифугування: при 250 g протягом 4 хвилин) та надосадову рідину одержали, звертаючи увагу на те, щоб вона не була забруднена клітинами. LDH у надосадовій рідині вимірювали за звичайним способом, що було визначено як "Кількість LDH, що витікали з клітини-мішені завдяки активності комплементу" (Експериментальний зразок). Наступні параметри також приготували, щоб визначити CDC-активність. Спонтанне виділення LDH з клітин-мішеней: тільки клітини-мішені культивували у такому ж самому об'ємі, як і зразок, та приготували. Максимальне виділення LDH з клітин-мішеней: тільки клітини-мішені культивували у такому ж самому об'ємі, як і зразок, а потім до них додали розчин TritonX-100, включений у набір, за 60 хвилин до одержання надосадової рідини, так щоб кінцева концентрація становила 0,8%, та приготували. Контроль корекції об'єму: таку ж саму кількість TritonX-100, яку додавали, коли готували максимальне виділення LDH з клітин-мішеней, додали до культурального середовища такого ж об'єму, що й у зразка, та приготували. Основа культурального середовища: приготували культуральне середовище такого ж самого об'єму, що й у зразка, та розчин, до якого додавали CDC-середовище, що містило комплемент, додавали до культурального середовища, так щоб був приготований такий же самий об'єм, що й у зразка. Такий самий об'єм культурального середовища, що й у зразка, відняли від абсорбції Максимальної мішені та Спонтанної мішені. Розчин, до якого додавали CDC-середовище, що містило комплемент, додавали до культурального середовища так, щоб мати такий самий об'єм, що й у зразка, відняли від абсорбції Експериментального зразка та коректували. CDCактивність підраховували, застосовуючи наступне рівняння. Результати наведено у Таблиці 1 та Фіг. 12. Навіть у тому випадку, коли застосовувалися моноклональні антитіла проти ILT7, отримані з гібридоми, 80% або більше CDC-активності спостерігали, коли концентрація антитіла становила 0,5 мкг/мл або більше. 30 35 40 Порівняльний приклад 1 Точно таку ж операцію виконували за таким самим способом, як описано у (В) та (С) Прикладу 4, за тим винятком, що IgG2a миші застосовували замість антитіла проти ILT7. Результати наведено у Прикладі 4, а також у Таблиці 5 та Фіг. CDC-активність до клітинмішеней не спостерігали в антитілах, відмінних від моноклонального антитіла проти ILT7. Приклад 5 Інтерналізація антитіла проти ILT7 до клітин-мішеней A. Моноклональне антитіло проти ILT7 28
ДивитисяДодаткова інформація
Назва патенту англійськоюMonoclonal antibody which specifically binds to an extracellular domain of human ilt7
Автори англійськоюKamogawa, Yumiko, Cho Minkwon, Arai Naoko, Ishida Koji
Назва патенту російськоюМоноклональное антитело, специфически связывающееся с внеклеточным доменом ilt7 человека
Автори російськоюКамогава Юмико, Чо Минквон, Арай Наоко, Ишида Кои
МПК / Мітки
МПК: C07K 16/28, C12N 5/20, G01N 33/577, A61K 39/395, C12N 15/13, A61P 37/00, C12P 21/08
Мітки: доменом, людини, зв'язується, позаклітинним, яке, моноклональне, специфічно, антитіло
Код посилання
<a href="https://ua.patents.su/102-97946-monoklonalne-antitilo-yake-specifichno-zvyazuehtsya-z-pozaklitinnim-domenom-ilt7-lyudini.html" target="_blank" rel="follow" title="База патентів України">Моноклональне антитіло, яке специфічно зв’язується з позаклітинним доменом ilt7 людини</a>
Моноклональне антитіло, яке спричиняє апоптоз кров’яних клітин, що містять ядро та інтегрин-пов’язаний протеїн людини (іпп), фрагмент моноклонального антитіла, гібридома, яка продукує моноклональне антитіло (ва

Номер патенту: 72192
Опубліковано: 15.02.2005
Автори: Уно Шінсуке, Фукушіма Наоші
МПК: C07K 16/28, C12N 5/20, C12P 21/08
Мітки: кров'яних, людини, яке, іпп, моноклонального, інтегрин-пов'язаний, антитіла, моноклональне, спричиняє, гібридома, містять, ядро, антитіло, ва, фрагмент, клітин, яка, протеїн, апоптоз, продукує
Формула / Реферат:
1. Моноклональне антитіло, яке зв’язується із інтегрин-пов’язаним протеїном людини (ІПП) і спричиняє апоптоз кров'яних клітин, що містять ядро та ІПП.2. Моноклональне антитіло за п.1, яке відрізняється тим, що антитіло отримують способом, що включає:а) імунізацію тварин з використанням ІПП людини як сенсибілізуючого антигена,б) одержання гібридом;в) відбір антитіл, що розпізнають ІПП людини та спричиняють апоптоз...
Моноклональне антитіло людини, яке специфічно зв’язується з рецептором інсуліноподібного фактора росту і (igf-ir)

Номер патенту: 87804
Опубліковано: 25.08.2009
Автори: Кохен Брюс Д., Мойєр Джеймс Д., Галло Майкл, Міллер Пенелопа Е., Корвалан Хосе Р., Біб Жан
МПК: A61K 49/00, A61K 39/395, G01N 33/574, A61P 35/00, C07K 16/18, C12N 15/13, C12N 5/20, C12P 21/08, C12N 15/64
Мітки: інсуліноподібного, людини, антитіло, рецептором, яке, моноклональне, росту, igf-ir, специфічно, зв'язується, фактора
Формула / Реферат:
1. Людське моноклональне антитіло або його антигензв'язувальна частина, яка специфічно зв'язується з рецептором інсуліноподібного фактора росту I (IGF-IR), де вказане антитіло містить важкий ланцюг і легкий ланцюг, де вказаний легкий ланцюг кодується геном Vκ А30 зародкової лінії людини, і де амінокислотні послідовності CDR1, CDR2 і CDR3 важкого ланцюга та амінокислотні послідовності CDR1, CDR2 і CDR3 легкого ланцюга вибрані з групи, що...
Виділене моноклональне антитіло людини, яке специфічно зв’язується з cd25 людини та інгібує зв’язування il-2 з сd25

Номер патенту: 90082
Опубліковано: 12.04.2010
Автори: Ван де Вінкель Ян Г.Й., Уільямс Деніс Лі, Паррен Пауль, Баадсгаард Оле Д.М.С., Шуурман Янін, Хавеніт Катаріна Емануель Герарда, Петерсен Йорген
МПК: A61K 51/10, A61P 7/00, G01N 33/577, C07K 16/28, A61K 39/395, A61P 35/00
Мітки: інгібує, моноклональне, сd25, яке, зв'язується, людини, антитіло, специфічно, зв'язування, виділене
Формула / Реферат:
1. Виділене моноклональне антитіло людини, яке специфічно зв'язується з СD25 людини та інгібує зв’язування IL-2 з CD25, вибране з групи, що містить:а) варіабельну ділянку важкого ланцюга, представлену в SEQ ID NO: 2, та варіабельну ділянку легкого ланцюга, представлену в SEQ ID NO: 4;b) варіабельну ділянку важкого ланцюга, представлену в SEQ ID NO: 6, та варіабельну ділянку легкого ланцюга, представлену в SEQ ID NO: 8;с)...
Людське моноклональне антитіло, яке специфічно зв’язується з m-csf людини та блокує зв’язування m-csf з c-fms

Номер патенту: 90457
Опубліковано: 11.05.2010
Автори: Моблі Джеймс Леслі, Келлєрманн Зірід-Аймеє, Лоу Джозеф Едвін, Девалараджа Мадхав Нарасімха, Хаак-Френдшо Мері, Фолтц Ян, Бедіан Вахе
МПК: A61P 25/00, A61K 39/395, A61P 37/00, C07K 16/22
Мітки: людини, людське, антитіло, m-csf, c-fms, зв'язування, моноклональне, зв'язується, яке, специфічно, блокує
Формула / Реферат:
1. Людське моноклональне антитіло, яке специфічно зв'язується з M-CSF людини і блокує зв'язування M-CSF з c-fms, або його антигензв'язувальна частина, де антитіло містить амінокислотну послідовність варіабельного домену важкого ланцюга (VН) і амінокислотну послідовність варіабельного домену легкого ланцюга (VL), у вказаному порядку:a) SEQ ID NO:2 і 4;b) SEQ ID NO:6 і 8;c) SEQ ID NO:10 і 12;d) SEQ ID NO:14 і...
Модифіковане моноклональне антитіло людини, що специфічно зв’язується з рецептором подібного до інсуліну фактора росту і людини (igf-ir)

Номер патенту: 85058
Опубліковано: 25.12.2008
Автори: Коен Брюс Девід, Бедіан Вахе
МПК: C07K 16/28, C12N 15/13, A61K 39/395, A61P 35/00
Мітки: росту, модифіковане, людини, рецептором, специфічно, igf-ir, моноклональне, антитіло, фактора, зв'язується, подібного, інсуліну
Формула / Реферат:
1. Модифіковане моноклональне антитіло людини, що специфічно зв’язується з рецептором подібного до інсуліну фактора росту І людини (IGF-IR), яке містить важкий ланцюг, що містить амінокислотну послідовність варіабельної ділянки, яка відповідає амінокислотним залишкам в позиціях з 20 до 144 SEQ ID NO: 3, і легкий ланцюг, що містить амінокислотну послідовність варіабельної ділянки, яка відповідає амінокислотним залишкам в позиціях з 23 до 130...
Попередній патент: Спосіб одержання кульок пінополістиролу
Наступний патент: Сліпа заклепка і пристрій для неї
Випадковий патент: Спосіб одержання солей лужних металів (варіанти)