Спосіб боротьби з грибами та ооміцетами шляхом інгібування гена сахаропіндегідрогенази
Номер патенту: 115132
Опубліковано: 25.09.2017
Автори: Паже Ерік, Шмітт Фредерік, Ессігманн Бернд, Делебарре Томас, Вілалба Француа, Дорме Сесіль







Формула / Реферат
1. Молекула длРНК, яка містить і) перший ланцюг, що містить послідовність, щонайменше на 95 % ідентичну до щонайменше 18 суміжних нуклеотидів гена сахаропіндегідрогенази гриба або ооміцета, та іі) другий ланцюг, що містить послідовність, комплементарну до понад щонайменше 80 % нуклеотидів першого ланцюга, де ген гриба або ооміцета вибирають із групи, що складається з:
a) полінуклеотиду, що містить послідовність, вказану у SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43;
b) полінуклеотиду, що кодує поліпептид, вказаний у SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44;
c) полінуклеотиду, що має щонайменше 95 % ідентичності послідовності до полінуклеотиду, що має послідовність, вказану у SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43;
d) полінуклеотиду, що кодує поліпептид, який має щонайменше 95 % ідентичності до поліпептиду, що має послідовність, вказану у SEQ ID NО: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44;
е) полінуклеотиду, що схрещується за суворих умов з полінуклеотидом, що має послідовність, вказану у SEQ ID NO: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25,27, 29, 31, 33, 35, 37, 39, 41, 43; та
f) полінуклеотиду, що схрещується за суворих умов з полінуклеотидом, який кодує поліпептид, що має послідовність, вказану у SEQ ID NO: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44.
2. Композиція для боротьби з грибами та ооміцетами, яка містить щонайменше молекулу длРНК за п. 1.
3. Композиція за п. 1 або 2, яка відрізняється тим, що додатково містить прийнятну для сільського господарства підложку, носій, наповнювач та/або поверхнево-активну речовину.
4. Композиція за п. 2 або 3, яка додатково містить сполуку, що сприяє росту рослин.
5. Застосування мікроорганізму для виробництва молекули длРНК за п. 1.
6. Генетична конструкція, яка містить щонайменше одну послідовність ДНК, а також щонайменше один гетерологічний регулювальний елемент у положенні 5', яка відрізняється тим, що щонайменше одна послідовність ДНК кодує длРНК за п. 1.
7. Генетична конструкція за п. 6, яка відрізняється тим, що містить додатково щонайменше один гетерологічний регулювальний елемент у положенні 3'.
8. Вектор клонування та/або експресії, який відрізняється тим, що він містить щонайменше одну генетичну конструкцію за п. 6 або п. 7.
9. Клітина трансгенної рослини, що містить молекулу длРНК за п. 1 або конструкцію за п. 6 або п. 7.
10. Трансгенна рослина, насіння або їх частина, що містить клітину трансгенної рослини за п. 9.
11. Клітина трансгенної рослини за п. 9 або трансгенна рослина, насіння або їх частина за п. 10, де вказаною рослиною є рослина соєвих бобів, насіння олійних культур, рису або картоплі.
12. Спосіб одержання клітини трансгенної рослини, здатної експресувати длРНК, що інгібує ген сахаропіндегідрогенази гриба або ооміцета, який відрізняється тим, що включає стадії трансформування клітини рослини генетичною конструкцією за п. 6 або п. 7.
13. Спосіб боротьби з рослинним патогеном грибом або ооміцетом, який включає введення вказаному патогену молекули длРНК за п. 1 або композиції за будь-яким із пп. 2-4.
14. Спосіб за п. 13, який відрізняється тим, що ефективну кількість молекули длРНК за п. 1 або композиції за будь-яким із пп. 2-4 вносять у ґрунт, де рослини ростуть або здатні рости, на листя та/або плоди рослин або на насіння рослин.
15. Спосіб за п. 14, який відрізняється тим, що рослині-хазяїну вказаного рослинного патогену гриба або ооміцета вводять трансформовану клітину рослини за п. 9.
16. Спосіб інгібування експресії гена рослинного патогену гриба або ооміцета, який включає такі стадії:
і) трансформування клітини рослини за допомогою генетичної конструкції за п. 6 або п. 7;
іі) розміщення трансформованих клітин, отриманих на стадії і) в умовах, що дозволяють транскрибування вказаної конструкції;
ііі) контактування трансформованих клітин, отриманих на стадії і) з патогеном рослин.
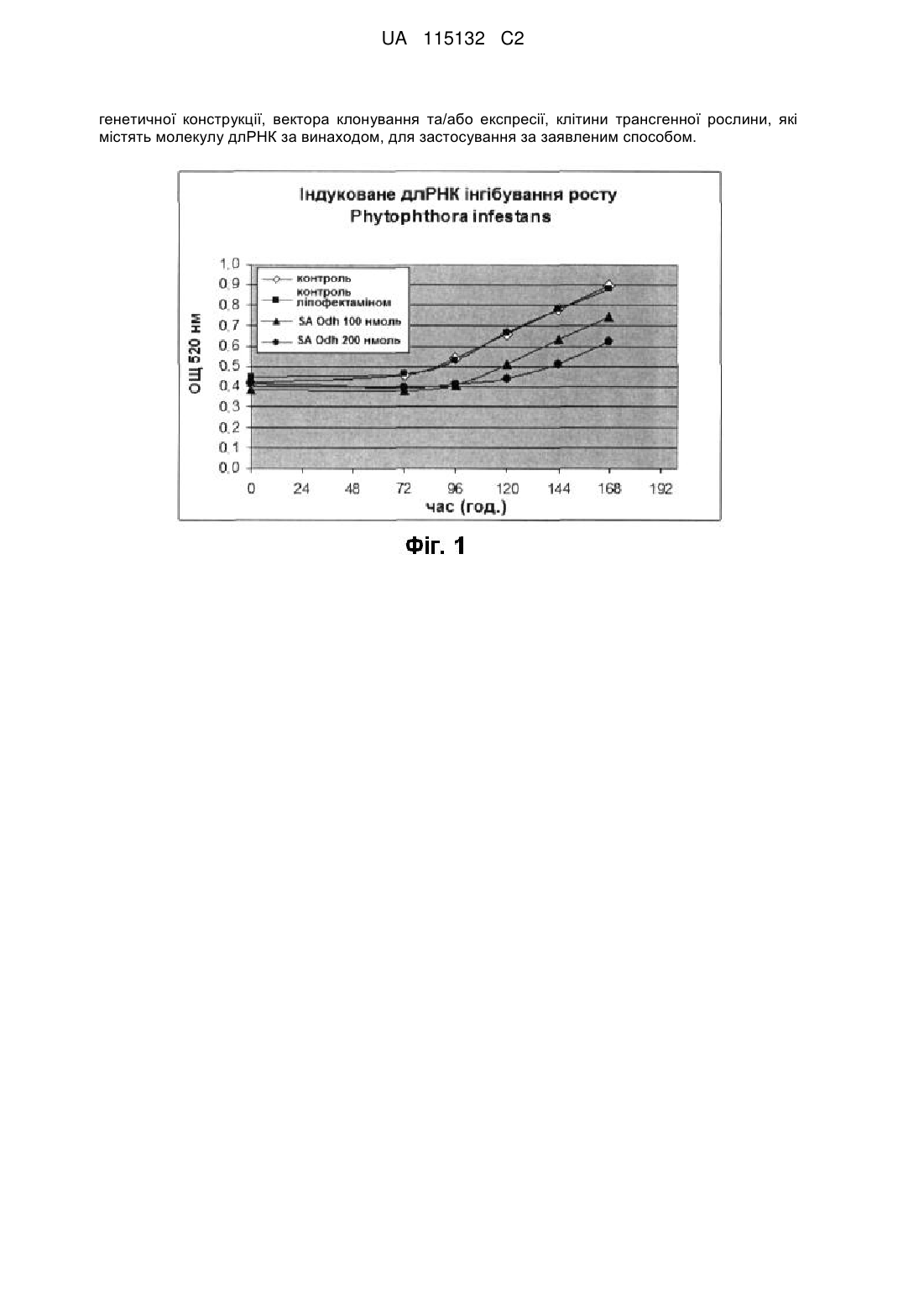
17. Спосіб за будь-яким із пп. 11-16, в якому вказаним патогеном рослин є Magnaporthe grisea, Phytophthora infestans, Sclerotinia sclerotinium або Phakopsora pachyrhizi.
18. Спосіб за будь-яким із пп. 11-16, де вказаною рослиною є соя, насіння олійних культур, рис або картопля.
Текст
Реферат: Винахід стосується способу боротьби з грибами та ооміцетами, шляхом інгібування гена сахаропіндегідрогенази із застосуванням РНК-інтерференції; молекули длРНК та композиції, UA 115132 C2 (12) UA 115132 C2 генетичної конструкції, вектора клонування та/або експресії, клітини трансгенної рослини, які містять молекулу длРНК за винаходом, для застосування за заявленим способом. UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 Цей винахід стосується боротьби з патогенами та шкідниками рослин, особливо грибами або ооміцетами, шляхом інгібування однієї або більше біологічних функцій, особливо шляхом інгібування гену сахаропін дегідрогенази грибів, залученого до шляху α-аміноадипату біосинтезу лізину, та їхніх ооміцетних гомологів із застосуванням РНК-інтерференції. Винахід забезпечує способи та композиції із застосуванням РНК-інтерференції генівмішеней грибів або ооміцетів для такої боротьби. Винахід також стосується способів одержання трансгенних рослин, толерантних до таких грибів або ооміцетів, та одержаних з них трансгенних рослин і насіння. Технологією, що її використовують у контексті цього винаходу, є РНК-інтерференція, або РНКі. Експресія в організмі послідовності, гомологічної до гену-мішені, яка здатна індукувати утворення малих дволанцюгових РНК (длРНК), дозволяє, дуже особливим чином, знищити цей ген і спостерігати за одержаним з нього фенотипом (Xiao et al., 2003). ДлРНК ініціює специфічне розщеплення гомологічної РНК лише у ділянці ідентичності з длРНК (Zamore et al., 2000; Tang et al., 2003). ДлРНК є молекулою РНК, яка містить дволанцюгову послідовність, звичайно зі щонайменше 19 базових пар (бп), у тому числі кодувальний нетранскрибований ланцюг та антисмисловий ланцюг. Молекули длРНК також відрізняються дуже високим ступенем компліментарності між двома комплементарними ланцюгами РНК. ДлРНК розщеплюється на фрагменти РНК звичайно з 18-25 нуклеотидів (міРНК), і сайти розщеплення на РНК-мішені рівномірно рознесені 18-25 нуклеотидами. Одержані малі міРНК проявляють дуже високий рівень ідентичності відносно РНК-мішені; однак, порушення компліментарності 3-4 нуклеотидів між міРНК та відповідною частиною цільової РНК все ж таки дозволяє системі працювати (Tang et al., 2003). З огляду на це, було висловлено припущення, що ці фрагменти з 18-25 нуклеотидів є РНК-гідами для розпізнавання мішені (Zamore et al., 2000). Ці малі РНК також були виявлені в екстрактах, одержаних з клітин Шнайдера 2 Drosophila melanogaster, які були трансфіковані длРНК перед клітинним лізисом (Hammond et al., 2000). Провідну роль фрагментів з 18-25 нуклеотидів у розщепленні мРНК підтверджує спостереження, що ці фрагменти з 19-25 нуклеотидів, виділених з длРНК, можуть бути залучені до розщеплення мРНК (Zamore et al., 2000). Гомологічні молекули РНК значних розмірів також накопичуються в тканинах рослин, які зазнають явища PTGS ("Посттранскрипційний сайленсинг генів", Hamilton and Baulcome, 1999). Ці малі РНК можуть регулювати експресію генів на трьох різних рівнях: - транскрибування (TGS означає "Транскрипційний сайленсинг генів"), - розщеплення матричної інформаційної РНК (PTGS означає "Пост- транскрипційний сайленсинг генів"), - шлях міРНК, - трансляція. Виявляється, що регулювання із залученням розщеплення матричної інформаційної РНК існує у всіх еукаріотів, тоді як регулювання на транскрипційному рівні описували лише у ссавців, рослин, дрозофіл та C. elegans. Стосовно регулювання трансляції, його описували у C. elegans та дрозофіл і виявляється, що воно також існує у ссавців (Hannon, 2002) та рослин (Ruiz Ferrer and Voinnet, 2009). У літературі посилаються на РНК-інтерференцію, PTGS, співсупресію або пригнічення (призначені для грибів) при згадуванні цього явища залежно від організмів, в яких воно досліджується. РНКі, зокрема, довела, що вона є ефективною при введенні дволанцюгової РНК (длРНК) в нематод Caenorhabditis elegans (Fire et al. 1998; Montgomery et al., 1998; WO99/32619). Також спостерігали за інгібуванням експресії гену-мішені у комахи, коли цю комаху годували бактеріями, що експресують малі дволанцюгові РНК, що відповідають згаданому гену-мішені у комах (WO 01/37654). Зовсім нещодавно були розкриті фармацевтичні композиції, що містять длРНК, по суті комплементарну щонайменше до частини гену, щодо якої є підозри в залученні до інфекції вірусу папіломи людини (HPV), разом з фармацевтично прийнятним носієм для лікування згаданої інфекції HPV (WO2009/0247607). Введення длРНК проводили в рослинах для індукування сайленсингу ендогенного генумішені (Hamilton et al., 1998, WO99/15682), для індукування стійкості до РНК-вірусів шляхом застосування трансгену, що експресує длРНК, який має істотну ідентичність відносно вірусних генів (Waterhouse et al., 1998; Pandolfini et al., 2003, WO98/36083, WO99/15682, US 5,175,102), але також для індукування стійкості до нематодів (Chuang and Meyerowitz, 2000, WO01/96584) або, як альтернатива, до бактерій Agrobacterium (WO00/26346, Escobar et al., 2001). Зовсім нещодавно було показано, що рослини, які експресують длРНК, що має істотну ідентичність 1 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 проти грибкового гену, що є важливим для росту гриба або його патогенності, також можуть індукувати стійкість до цього гриба (WO05/071091). Між тим, з того часу існує лише декілька попередніх результатів і жодних комерційних прикладів відносно опосередкованої РНКі стійкості чи толерантності до фітопатогенних грибів, тоді як дволанцюгові (длРНК) або малі інтерферувальні (міРНК) молекули експресуються в рослині або наносяться як частина зовнішньої композиції на насіння, рослину чи плід рослини або вносяться у ґрунт чи на інертний субстрат, де рослина росте або де її вирощування є бажаним. Поряд з іншими, однією зі складностей є виявлення належного гену-мішені, інгібування якого за допомогою длРНК або міРНК індукує добрий рівень толерантності до грибів, аж до рівня, що підходить для комерційного застосування, без шкідливого впливу на рослину, що експресує згадану длРНК чи міРНК або на яку наносять композицію, що містить згадану длРНК чи міРНК. Старкель С. (Stärkel C.) у своїй кандидатській дисертації на тему "Індукований хазяїном сайленсинг генів: стратегії покращення стійкості проти Cercospora beticola у цукрового буряка (B. vulgaris L.) та проти Fusarium graminearum у пшениці (T. aestivum L.) та маїсу (Z. mays L.)" ["Host induced gene silencing - strategies for the improvement of resistance against Cercospora beticola in sugar beet (B. vulgaris L.) and against Fusarium graminearum in пшениця (T. aestivum L.) and maize (Z. mays L.)", defended in June 2011], захищеній в червні 2011 року, намагався інгібувати ріст Fusarium graminearum шляхом трансформування пшениці за допомогою конструкцій сайленсингу, націлених на ген гомоаконітази - важливий ген у шляху біосинтезу лізину. Проте, трансгенних рослин пшениці вивести не вдалося. Крім того, спроби вилучити або приглушити ген гомоаконітази у Fusarium graminearum шляхом трансформування грибів за допомогою конструкції, створеної для індукованого сайленсингу гену гомоаконітази, також були марними. Автори цього винаходу несподівано показали, що інгібування гену дегідрогенази сахаропіну грибів або ооміцетів, залученого до шляху α-аміноадипату (AAA), за допомогою методології РНКі спричиняє зупинення інфекції, росту, розвитку, відтворення та/або патогенності і, зрештою, закінчується загибеллю організму. Ця нова мішень для РНКі технології є особливо доцільною з урахуванням того, що шлях AAA особливо виявляють у деяких патогенах рослин, у тому числі у вищих грибів, а не у рослин, людей або тварин. З-поміж 20 звичайних протеїногенних амінокислот L-лізин є єдиною амінокислотою, відомою як така, що має біосинтетичний шлях, який відрізняється у рослин та вищих грибів. У рослин та бактерій L-лізин надходить через діамінопімелатний (DAP) шлях. У вищих грибів та евгленідів Lлізин надходить через α-аміноадипатний (AAA) шлях. Сахаропін дегідрогеназа, гомоцитрат синтаза, гомоаконітаза, гомоізоцитрат дегідрогеназа, α-аміноадипат амінотрансфераза, αаміноадипат редуктаза та сахаропін редуктаза є ферментами, залученими до αаміноадипатного шляху біосинтезу L-лізину. Жоден із ферментів, залучених до цих відмінних шляхів (DAP та AAA шляхів), не є звичайним (Xu et al., 2006, Bhattacharjee, J. K., 1985; Bhattacharjee, J. K., 1992; Vogel, H. J., 1965). Як приклад, сахаропін дегідрогеназа грибів залучена до біосинтезу лізину, коли лізинкетоглутарат із рослини, що його іноді також називають сахаропін дегідрогеназою, відповідає за катаболізм лізину (Houmard et al., 2007). Для людей і тварин L-лізин є важливою амінокислотою, яку можна одержати лише з білка, що міститься в їжі. Крім того, ферменти, залучені до AAA шляху грибів, є унікальними для синтезу лізину (Umbargar, H. E., 1978; Bhattacharjee, J. K., 1992). Присутність специфічного α-аміноадипатного (AAA) шляху у вищих грибів, якого немає ані в рослинах, ані у людей чи тварин, наводить на думку вважати ферменти, залучені до згаданого ААА шляху, особливо привабливими мішенями для боротьби з патогенами рослин, особливо грибковими патогенами. Цікаво, що хоча повідомляли, що лізин синтезується у ооміцетів через діамінопімеліновий шлях (Born and Blanchard, 1999), у ооміцетів виявили гени, гомологічні до генів грибів AAA шляху, що може свідчити, що обидва шляхи можуть бути присутніми у ооміцетів. Автори показали, що длРНК проти гену сахаропін дегідрогенази ооміцетів істотно знижувала ріст згаданого ооміцету, і що рослини, які експресують длРНК проти генів сахаропін дегідрогенази ооміцетів, могли би бути менш чутливими до згаданої інфекції ооміцетів. Присутність AAA шляху біосинтезу лізину була продемонстрована та досліджена, наприклад, на Saccharomyces cerevisiae (Broquist, H. P., 1971, Bhattacharjee, J. K., 1992), на патогенних грибах людини Candida albicans (Garrad, R. C. and Bhattacharjee, J. K., 1992) та на патогенні рослин Magnaporthe grisea (Umbargar, H. E., 1978). Ферменти дегідрогенази 2 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 сахаропіну, залучені до ААА шляху, ретельно досліджували та порівнювали у різних організмів, а їх технічні ознаки, у тому числі нуклеотидні та амінокислотні послідовності, кінетика, субстратна специфічність, функції, тривимірна структура, а також спосіб їх очищення та характеристики, добре відомі спеціалістові (див. Xu et al., 2001, зміст якого включений до цього опису як посилання). Численні гени різних грибів або ооміцетів і дані їхніх послідовностей розкриті та доступні у відкритих базах, придатних для пошуку, таких як genBank. Randall, T.A. et al. (2005) надає перелік генів AAA шляху ооміцетів Phytophthora infestans, що ідентифіковані за інвентарними номерами, дані їхніх послідовностей, включені до цього опису як посилання, доступні у відкритих базах, придатних для пошуку (Randall, T.A., 2005, MPMI, 18, 229-243; див., зокрема, таблицю 8 на стор. 239). В одному втіленні цей винахід забезпечує молекулу длРНК, яка містить: і) перший ланцюг, що містить послідовність, по суті ідентичну до щонайменше 18 суміжних нуклеотидів гену грибів або ооміцетів, та ii) другий ланцюг, що містить послідовність, по суті комплементарну до першого ланцюга, причому перший ген грибів або ооміцетів є геном дегідрогенази сахаропіну. Використаний тут термін "РНКі", або "РНК-інтерференція", стосується процесу послідовність-специфічного сайленсингу генів, опосередкованого дволанцюговою РНК (длРНК). Використаний тут термін "длРНК", або "длРНК молекула", стосується РНК, яка є частково або повністю дволанцюговою. Дволанцюгова РНК також стосується малої або короткої інтерферувальної РНК (міРНК), короткої інтерферувальної нуклеїнової кислоти (кіНК), короткої інтерферувальної РНК, мікро-РНК (міРНК), кільцевої інтерферувальної РНК (кіРНК), короткої шпількової РНК (кшРНК) тощо. Як використано у цьому описі, з урахуванням заміщення урацилу тиміном при порівнянні послідовностей РНК і ДНК, термін "по суті ідентичний" або "по суті гомологічний" відносно длРНК означає, що нуклеотидна послідовність одного ланцюга длРНК є щонайменше близько на 80 %, щонайменше на 85 % ідентичною до 18 або більше суміжних нуклеотидів гену-мішені, переважно щонайменше на близько 90 % ідентичною до 18 або більше суміжних нуклеотидів гену-мішені і найкраще щонайменше на близько 95 %, 96 %, 97 %, 98 % або 99 % ідентичною або абсолютно ідентичною до 18 або більше суміжних нуклеотидів гену-мішені. 18 або більше нуклеотидів означає ділянку, що становить щонайменше близько 18, 20, 21, 22, 23, 24, 25, 50, 100, 200, 300, 400, 500, 1000, 1500 або 2000 послідовних основ або аж до повної довжини генумішені. Як використано у цьому описі, "комплементарними" полінуклеотидами є ті, що здатні до спаровування основ відповідно до стандартних правил комплементарності Ватсона-Кріка (Watson-Crick). Особливо пурини утворюють пари з піримідинами для утворення комбінації гуаніну, парованого з цитозином (G:C), та аденіну, парованого з тиміном (A:T) у разі ДНК, або аденіну, парованого з урацилом (A:U) у разі РНК. Розуміють, що два полінуклеотиди можуть схрещуватись один з одним навіть якщо вони не є повністю комплементарними один до одного, за умови, що кожний має щонайменше одну ділянку, що є по суті комплементарною до іншої. Використаний тут термін "по суті комплементарний" означає, що дві послідовності нуклеїнової кислоти є комплементарними до понад щонайменше 80 % їхніх нуклеотидів. Краще, якщо дві послідовності нуклеїнової кислоти є комплементарними до понад щонайменше 85 %, 90 %, 95 %, 96 %, 97 %, 98 %, 99 % або більше чи всіх їхніх нуклеотидів. Альтернативно, "по суті комплементарний" означає, що дві послідовності нуклеїнової кислоти можуть схрещуватись за дуже суворих умов. Використаний тут термін "по суті ідентичний" або "що відповідає" означає, що дві послідовності нуклеїнової кислоти мають щонайменше 80 % ідентичності послідовності. Краще, якщо дві послідовності нуклеїнових кислот мають щонайменше 85 %, 90 %, 95 %, 96 %, 97 %, 98 %, 99 % або 100 % ідентичності послідовності. Також використані тут терміни "нуклеїнова кислота" та "полінуклеотид" стосуються РНК або ДНК, яка є лінійною або розгалуженою, з одним або подвійним ланцюгом, або їх гібриду. Цей термін також охоплює гібриди РНК/ДНК. При синтетичному одержанні длРНК менш поширені основи, такі як інозин, 5-метилцитозин, 6-метиладенін, гіпоксантин та інші, також можуть бути використані для атисмислового, длРНК та рибозимного спаровування. Наприклад, було виявлено, що полінуклеотиди, які містять C-5 пропінові аналоги уридину та цитидину, зв'язуються з РНК з високою афінністю і є потужними антисмисловими інгібіторами генної експресії. Також можуть бути одержані інші модифікації, такі як модифікація фосфодіефірного каркаса, зачинена нуклеїнова кислота або 2'-гідрокси в рибозній цукровій групі РНК. Відповідно до винаходу перший ланцюг і другий ланцюг можуть мати ідентичні розміри. Альтернативно, розмір першого ланцюга може бути більшим, ніж розмір другого ланцюга. Як приклад, розмір першого ланцюга може бути близько на 200 нуклеотидів більшим, ніж розмір другого ланцюга. В іншому аспекті цього винаходу розмір другого ланцюга є більшим, ніж розмір 3 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 першого ланцюга. Відповідно до винаходу молекула длРНК містить перший ланцюг, що містить послідовність, по суті ідентичну щонайменше до 18 суміжних нуклеотидів гену сахаропін дегідрогенази грибів або ооміцетів. В окремому втіленні винахід забезпечує молекулу длРНК, що містить і) перший ланцюг, що містить послідовність, по суті ідентичну щонайменше до 18 суміжних нуклеотидів гену грибів або ооміцетів, та ii) другий ланцюг, що містить послідовність, по суті компліментарну до першого ланцюга, причому згаданий ген грибів або ооміцетів вибирають з переліку, що складається з: a) полінуклеотиду, що містить послідовність, вказану у SEQ ID N0: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43; b) полінуклеотиду, що кодує поліпептид, який має послідовність, вказану у SEQ ID N0: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44; c) полінуклеотиду, що має щонайменше 70 % ідентичності послідовності, краще щонайменше 80 %, ще краще щонайменше 90 %, навіть ще краще щонайменше 95 % до полінуклеотиду, який має послідовність, вказану у SEQ ID N0: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43; d) полінуклеотиду, що кодує поліпептид, який має щонайменше 70 % ідентичності послідовності, краще щонайменше 80 %, ще краще щонайменше 90 %, навіть ще краще щонайменше 95 % до поліпептиду, який має послідовність, вказану у SEQ ID N0: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44; e) полінуклеотиду, що схрещується за суворих умов з полінуклеотидом, який має послідовність, вказану у SEQ ID N0: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 33, 35, 37, 39, 41, 43; та f) полінуклеотиду, що схрещується за суворих умов з полінуклеотидом, який кодує поліпептид, що має щонайменше 70 % ідентичності послідовності до поліпептиду, що має послідовність, вказану у SEQ ID N0: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44. Відповідно до цього винаходу термін "ідентичність" треба розуміти як такий, що означає кількість амінокислот / нуклеотидів, що відповідає амінокислотам / нуклеотидам іншого білка / нуклеїнової кислоти, виражену у процентному співвідношенні. Ідентичність переважно визначають шляхом порівняння ідентичності послідовності, вказаної у цьому описі, з іншим білком / нуклеїновою кислотою за допомогою комп'ютерних програм. Якщо послідовності, що порівнюються одна з одною, мають різну довжину, ідентичність визначають таким чином, щоб кількість амінокислот, що мають меншу послідовність спільну з довшою послідовністю, визначала коефіцієнт процентного співвідношення ідентичності. Переважно ідентичність визначають за допомогою комп'ютерної програми ClustalW, яка є добре відомою та загальнодоступною (Thompson et al., 1994). ClustalW є загальнодоступною за адресою: http://www.ebi.ac.uk/tools/clustalW2/index.html. Переважно застосовують версію 2.1 комп'ютерної програми ClustalW для визначення ідентичності між білками відповідно до винаходу та іншими білками. При здійсненні цього необхідно встановлювати такі параметри: KTUPLE=1, TOPDIAG=5, WINDOW=5, PAIRGAP=3, GAPOPEN=10, GAPEXTEND=0.05, GAPDIST=8, MAXDIV=40, MATRIX=GONNET, ENDGAPS(OFF), NOPGAP, NOHGAP. Переважно застосовують версію 2.1 комп'ютерної програми ClustalW для визначення, наприклад, ідентичності між нуклеотидною послідовністю молекул нуклеїнової кислоти відповідно до винаходу та нуклеотидною послідовністю інших молекул нуклеїнової кислоти. При здійсненні цього необхідно встановлювати такі параметри: KTUPLE=2, TOPDIAGS=4, PAIRGAP=5, DNAMATRIX:IUB, GAPOPEN=10, GAPEXT=5, MAXDIV=40, ТРАНЗИЦІЇ: незважені. Відповідно до цього винаходу термін "схрещування за суворих умов" стосується полінуклеотидів або послідовностей нуклеїнової кислоти, які схрещуються з еталонною послідовністю нуклеїнової кислоти на рівні, що є значно вищим, ніж фоновий шум. Фоновий шум може бути пов'язаний зі схрещуванням інших послідовностей присутніх ДНК, зокрема інших кДНК, присутніх в бібліотеці кДНК. Рівень сигналу, утвореного взаємодією між послідовністю, здатною вибірково схрещуватись, та послідовностями, визначеними вказаними вище ідентифікаторами послідовностей відповідно до винаходу, звичайно є у 10 разів, переважно у 100 разів вищим, ніж рівень сигналу від взаємодії інших послідовностей ДНК, що утворюють фоновий шум. Рівень взаємодії може бути виміряний, наприклад, шляхом мічення зонду 32 радіоактивними елементами, такими як P. Селективного схрещування звичайно досягають шляхом застосування дуже суворих умов до середовища (наприклад, 0,03 M NaCl і 0,03 M цитрату натрію при температурі приблизно 50-60 °C). Звичайно, схрещування може бути проведено відповідно до звичних методів, відомих із рівня техніки (зокрема, Sambrook et al., 2001, Molecular Cloning: A Laboratory Manual ("Молекулярне клонування: лабораторний 4 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 посібник"), третє видання). В окремому втіленні винаходу молекулу длРНК наносять на патоген рослин, особливо на гриби або ооміцети, та/або на рослину чи сільськогосподарську культуру, що підлягає захисту. Відповідно, цей винахід також стосується композиції, що містить ефективну та нефітотоксичну кількість визначеної тут молекули длРНК. Молекули длРНК відповідно до винаходу можуть бути одержані класичним хімічним синтезом, способом транскрибування в умовах in vitro або в організмах на зразок клітин тварин, бактерій, дріжджів або рослин гетерологічною експресією (Aalto A.P. et al, 2007 RNA. 13(3):4229.) Відповідно, цей винахід стосується мікроорганізму, який виробляє визначену тут молекулу длРНК. Цей винахід також стосується генетичної конструкції, що містить щонайменше одну послідовність ДНК, а також гетерологічний регуляторний елемент або елементи у 5" та необов'язково у 3" положенні, яка відрізняється тим, що послідовність ДНК здатна утворювати визначену тут молекулу длРНК. Цей винахід також стосується клонувального та/або експресійного вектору, який відрізняється тим, що він містить щонайменше одну визначену тут генетичну конструкцію. Вираз "ефективна і нефітотоксична кількість" означає кількість композиції відповідно до винаходу, яка є достатньою для боротьби з патогеном або його знищення, що присутній на сільськогосподарських культурах або поява якого на них є ймовірною, і яка не викликає будьяких істотних симптомів фітотоксичності для згаданих культур. Така кількість може варіюватись у межах широкого діапазону залежно від патогену, з яким необхідно боротися, типу сільськогосподарської культури, кліматичних умов і сполук, що входять до складу композиції відповідно до винаходу. Ця кількість може бути визначена систематичними польовими випробуваннями, можливість проведення яких є доступною для спеціаліста у цій галузі. Отже, відповідно до винаходу забезпечена композиція, яка містить як активний інгредієнт ефективну кількість визначеної тут молекули длРНК сполуки та прийнятні для сільського господарства підложку, носій, наповнювач та/або поверхнево-активну речовину. Відповідно до винаходу термін "підложка" позначає природну або синтетичну, органічну або неорганічну сполуку, з якою поєднують або асоціюють активну сполуку формули (I) для того, аби полегшити її нанесення, особливо на частини рослини. З огляду на це ця підложка звичайно є інертною і повинна бути прийнятною для сільського господарства. Підложка може бути твердою або рідкою. Прикладами підходящих підложок є глина, природні або синтетичні силікати, кремнезем, смоли, віск, тверді добрива, вода, спирти, зокрема бутанол, органічні розчинники, мінеральні й рослинні олії та їх похідні. Також можуть бути використані суміші таких підложок. Композиція відповідно до винаходу також може містити додаткові компоненти, такі як, наприклад, але це не є обмеженням, поверхнево-активна речовина, захисні колоїди, адгезиви, згущувачі, тиксотропні агенти, агенти просочування, стабілізатори, комплексоутворювальні сполуки. У більшості випадків активні сполуки можуть бути поєднані з будь-якою твердою або рідкою добавкою, що відповідає звичайним технологіям приготування. Загалом, композиція за винаходом може містити від 0,05 до 99 ваг. % активної сполуки, переважно від 10 до 70 ваг. %. Композиції відповідно до винаходу можуть бути використані у різних формах, таких як аерозольний розподільний пристрій, капсульна суспензія, концентрат холодного туману, пилоподібний порошок, емульгований концентрат, емульсія олія-у-воді, емульсія вода-в-олії, інкапсульована гранула, дрібна гранула, текучий концентрат для обробки насіння, газ (під тиском), газоутворювальний продукт, гранули, концентрат гарячого туману, макрогранули, мікрогранули, порошок, здатний до диспергування в олії, змішуваний з олією текучий концентрат, змішувана з олією рідина, паста, рослинні палички, порошок для обробки сухого насіння, насіння, покрите пестицидом, розчинний концентрат, розчинний порошок, розчин для обробки насіння, концентрат суспензії (текучий концентрат), рідина з ультранизьким об'ємом (ULV), суспензія з ультранизьким об'ємом (ULV), гранули і таблетки, здатні до диспергування у воді, порошок, здатний до диспергування у воді, для обробки рідким розчином (суспензією), водорозчинні гранули або таблетки, водорозчинний порошок для обробки насіння і змочуваний порошок. Ці композиції включають не лише композиції, що є готовими для нанесення на рослину або насіння для обробки за допомогою підходящого пристрою, такого як пристрій для розбризкування або обприскування, але також концентровані комерційні композиції, які мають бути розведені до нанесення на культури рослин. Сполуки відповідно до винаходу також можуть бути змішані з однією або більше активною 5 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 фітофармацевтичною сполукою або сполукою, що сприяє росту рослин, такою як фунгіцид, гербіцид, інсектицид, нематицид, акарицид, молюскоцид, стимулятор стійкості, антидоти, сигнальні сполуки, біопрепарати, активна речовина феромону або іншими сполуками з біологічною активністю. Одержані таким чином суміші мають розширений спектр активності. Сумішам з іншими фунгіцидними сполуками надають особливу перевагу. В окремому втіленні винаходу длРНК вводять або вносять у рослину, що підлягає захисту. Після введення або внесення в рослину длРНК додатково може бути перетворена на відносно малі фрагменти (міРНК) і згодом може розподілітися по всій рослині. Альтернативно, длРНК вводять або вносять у рослину із застосуванням регуляторного елемента або промотора, що спричиняє експресію длРНК тканинним, тимчасовим, просторовим або індукованим способом, і може додатково бути перетворена на відносно малі фрагменти за допомогою рослинної клітини, що містить механізм переробки РНКі. Таким чином, винахід стосується генетичної конструкції або химерного гену, здатного утворювати длРНК за винаходом всередині рослинної клітини. Така генетична конструкція або химерний ген містить щонайменше одну послідовність ДНК, а також гетерологічний регуляторний елемент або елементи у 5" та необов'язково у 3" положенні, що здатні функціонувати в рослині, які відрізняються тим, що послідовність (послідовності) ДНК може (можуть) утворювати описану тут молекулу длРНК після експресії в рослині. В окремому втіленні генетична конструкція або химерний ген містить: - паромоторну регуляторну послідовність, що є функціональною в рослинних клітинах, сполучену з можливістю функціонування до - ДНК послідовності, яка, при її транскрибуванні, утворює молекулу РНК, що містить щонайменше смислову послідовність та анти смислову послідовність, які є щонайменше частково комплементарними, причому згадана смислова послідовність містить послідовність, по суті ідентичну до щонайменше 18 суміжних нуклеотидів гену-мішені (тобто в контексті винаходу гену сахаропін дегідрогенази), і причому згадана антисмислова послідовність містить послідовність, що є по суті комплементарною до смислової послідовності, та - необов'язково термінатор ну регуляторну послідовність. У згаданому втіленні послідовність ДНК відповідно до винаходу може мати більш конкретно два аспекти. У першому аспекті вона містить дві нуклеотидні послідовності, які є смисловими та антисмисловими, відокремлені спейсерною нуклеотидною послідовністю або інтроном, що не проявляють будь-якої гомології з геном-мішенню. Послідовністю, клонованою у смисловому та антисмисловому напрямку, є послідовність, експресію якої в патогенні необхідно інгібувати. Транскрибування цієї ДНК-послідовності таким чином дає велику одноланцюгову РНК, що відповідає смисловій/спейсер-інтрон/антисмисловій конструкції. Цей довгий РНК-транскрипт може бути виявлений за допомогою ПЛР зі зворотною транскрипцією. Оскільки смислова та антисмислова послідовності є гомологічними, вони будуть паруватися, і спейсер або інтрон, що їх відокремлює, відіграватиме роль петлі для укладання. Потім одержують длРНК в усіх гомологічних ділянках. ДлРНК згодом спеціально руйнується за допомогою ферментного комплексу під назвою "DICER" (дайсер). Потім від руйнування длРНК утворюються міРНК ("міРНК" на кресленні), малі дволанцюгові РНК, що мають розмір від 19 до 25 основ. Потім вони стають міРНК, які, при паруванні з транскрибованими РНК, виведеними з гену-мішені, спричинятимуть їх руйнування за допомогою ферментного механізму рослини механізму сайленсингу РНК. У другому аспекті послідовність ДНК містить дві нуклеотидні послідовності, які є смисловою та антисмисловою, різного розміру, причому структура їхньої петлі відповідає частині нуклеотидної послідовності, що не проявляє будь-якої гомології з іншою нуклеотидною послідовністю. Нуклеотидна послідовність, клонована у смисловому напрямку, є по суті гомологічною до послідовності гену-мішені, експресію якої необхідно інгібувати. Антисмислова нуклеотидна послідовність є по суті гомологічною до комплементарного ланцюга послідовності згаданого гену-мішені. Таким чином, транскрибування цієї ДНК-послідовності дає велику одноланцюгову ДНК, що відповідає смисловій/антисмисловій конструкції. Цю довгу РНКтранскрипцію можна виявити за допомогою ПЛР зі зворотною транскрипцією. Гомологічні смислова/антисмислова послідовності паруються. Потім одержують длРНК у всіх гомологічних ділянках. ДлРНК згодом спеціально руйнується за допомогою ферментного комплексу під назвою "DICER". Руйнування длРНК потім утворює міРНК ("міРНК" на кресленні), малі дволанцюгові РНК, що мають розмір від 18 до 25 основ. Потім вони стають міРНК, які, при паруванні з РНК-мішенями спричинятимуть їх руйнування за допомогою ферментного механізму рослини механізму сайленсингу РНК. В іншому втіленні генетична конструкція містить: 6 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 - дві промоторні регуляторні послідовності, що є функціональними в рослинних клітинах, де перша промоторна регуляторна послідовність оперативно сполучена з ДНК-послідовністю, яка, при її транскрибуванні, утворює молекулу РНК, що містить щонайменше смислову послідовність, а друга промоторна регуляторна послідовність оперативно сполучена з ДНКпослідовністю, яка, при її транскрибуванні, утворює молекулу РНК, що містить щонайменше антисмислову послідовність, частково компліментарну до смислової послідовності, і де згадана смислова послідовність містить послідовність, по суті ідентичну до щонайменше 18 суміжних нуклеотидів гену-мішені, та - необов'язково термінаторну регуляторну послідовність (послідовності). У цьому конкретному втіленні генетична конструкція може складатися з двох химерних генів, причому один містить першу промоторну регуляторну послідовність, оперативно сполучену з першою послідовністю ДНК, яка, при її транскрибуванні, утворює молекулу РНК, що містить щонайменше смислову послідовність, по суті ідентичну до щонайменше 18 суміжних нуклеотидів гену-мішені, і необов'язково термінаторну регуляторну послідовність, і причому другий химерний ген містить другу промоторну регуляторну послідовність, оперативно сполучену з другою послідовністю ДНК, яка, при її транскрибуванні, утворює молекулу РНК, що містить щонайменше антисмислову послідовність, частково комплементарну до смислової послідовності, та необов'язково термінаторну регуляторну послідовність. Ці два химерні гени переважно вводять у рослинну клітину спільно, але це не є необхідністю, для сприяння схрещуванню двох одиночних ланцюгів РНК для утворення длРНК. Альтернативно, генетична конструкція може являти собою конструкцію, що містить: - перший промотор, оперативно сполучений з - дволанцюговою послідовністю ДНК, де один ланцюг, при його транскрибуванні під контролем першого промотора, утворює молекулу РНК, що містить щонайменше смислову послідовність, по суті ідентичну до щонайменше 18 суміжних нуклеотидів гену-мішені, і необов'язково термінаторну регуляторну послідовність, і де другий ланцюг, при його транскрибуванні під контролем другого промотора, утворює молекулу РНК, що містить щонайменше антисмислову послідовність, частково комплементарну до смислової послідовності, і необов'язково термінаторну регуляторну послідовність, та - другий промотор в протилежному до першого промотора напрямку. Перша і друга промоторні регуляторні послідовності можуть бути різними або ідентичними, краще різними. Винахід також стосується клонувального та/або експресійного вектору для трансформування рослини, який відрізняється тим, що він містить щонайменше один химерний ген або генетичну конструкцію, як визначено тут. Цей винахід також стосується клітини трансгенної рослини, що містить молекулу длРНК за винаходом і як вона визначена тут. Цей винахід також стосується клітини трансгенної рослини, що містить визначені тут генетичну конструкцію або химерний ген за винаходом. В окремому втіленні винаходу клітиною трансгенної рослини є клітина рослини соєвих бобів, олійного насіння, рису або картоплі. Цей винахід також стосується трансгенної рослини, насіння або його частини, що містить клітину трансгенної рослини відповідно до винаходу. В окремому втіленні цього винаходу трансгенною рослиною, насінням або його частиною є рослина, насіння або його частина соєвих бобів, олійного насіння, рису або картоплі. Відповідно до винаходу вирази "химерний ген" або "експресійна касета" означають нуклеотидну послідовність, що містить функціонально сполучені одна з одною у напрямі транскрибування регуляторну промоторну послідовність, що є функціональною у рослин, послідовність, що кодує білок або РНК ланцюг, та, необов'язково, термінатор, що є функціональним в рослинних клітинах. Термін "химерний ген" або "експресійна касета" звичайно означають ген, для якого певні елементи не є присутніми в нативному гені, але були заміщені елементами, присутніми в нативному гені або були додані. Відповідно до винаходу терміни "химерний ген" або "експресійна касета" також можуть відповідати випадку, де всі елементи гену є присутніми в нативному гені, та, альтернативно, термін "ген" може відповідати химерному гену. Вираз "химерний ген" або "експресійна касета" також можуть відповідати випадку, коли послідовність, що кодує білок або РНК ланцюг, безпосередньо не сполучена з промотором, але є частиною, наприклад, поліцистронної конструкції, що містить декілька кодувальних послідовностей під контролем того самого промотора. У цьому випадку кожна кодувальна 7 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 послідовність під контролем промотора сконструйована як "химерний ген" або "експресійна касета". Відповідно до винаходу вираз "функціонально сполучений один до одного" означає, що згадані елементи елементарного химерного гену сполучені один з одним у такий спосіб, що їхня функція координована і дозволяє експресію кодувальної послідовності. Як приклад, промотор функціонально сполучений з кодувальною послідовністю, коли він здатний забезпечити експресію згаданої кодувальної послідовності. Конструювання химерного гену відповідно до винаходу та збирання його різних елементів може бути здійснене із застосуванням технологій, відомих спеціалістам у цій галузі, зокрема тих, що описані Sambrook et al. (2001, Molecular Cloning: A Laboratory Manual (third edition) ("Молекулярне клонування: Лабораторний посібник" (третє видання)), Nolan C. ed., New York: Cold Spring Harbor Laboratory Press). Вибір регуляторних елементів, що складають химерний ген, залежить по суті від рослини та виду клітини, в якій вони мають функціонувати, і спеціалісти у цій галузі здатні вибрати регуляторні елементи, що є функціональними у конкретній рослині. Відповідно до винаходу термін "промоторна регуляторна послідовність" має означати будьяку промоторну регуляторну послідовність гену, що природно експресується в рослинах, зокрема промотор, що експресується особливо в листях рослин, наприклад, промотори, які мають бактерійне, вірусне або рослинне походження, або інші промотори, які називають світлозалежними, такими як промотори рослинного рибулоза-біскарбоксилаза/оксигеназа (RuBisCO) гену малої субодиниці, або будь-який відомий промотор, який може бути використаний. Серед промоторів рослинного походження можна зазначити промотори гістону, як описано в заявці EP 0 507 698, або промотор рисового актину (US 5,641,876). Серез промоторів гену вірусу рослин можна зазначити промотор мозаїчного вірусу цвітної капусти (CaMV 19S або 35S), мозаїчний вірус cassava vein (CsVMV: WO97/48819) або промотор цирковірусу (AU 689 311). Також можна застосовувати промоторну регуляторну послідовність, специфічну для конкретних ділянок або тканин рослин, і, більш конкретно, специфічні до насіння промотори (Datla, R. et al., 1997, Biotechnology Ann. Rev., 3, 269-296), особливо промотори напіну (EP 255 378), фазеоліну, глютеніну, геліантиніну (WO 92/17580), альбуміну (WO 98/45460) та олеозину (WO 98/45461). Також може бути використаний індукований промотор, його можна переважно вибрати з промоторів фенілаланін аміакліази (PAL), HMG-CoA редуктази (HMG), хітиназ, глюканаз, інгібіторів протеїнази (PI), генів сімейства PR1, нопалін синтази (nos) або гену vspB (US 5 670 349), промотора HMG2 (US 5 670 349), промотора бетагалактозидази яблуні (ABG1) або промотора аміноциклопропан карбоксилатсинтази яблуні (ACC синтази) (WO 98/45445). Термін "термінаторна регуляторна послідовність" означає будь-яку послідовність, що є функціональною в рослинних клітинах або рослинах, яка також містить послідовності поліаденілування, які можуть мати як бактерійне походження, наприклад, термінатор nos або ocs Agrobacterium tumefaciens, вірусне походження, наприклад, термінатор CaMV 35S, або рослинне походження, наприклад, гістоновий термінатор, описаний в заявці EP 0 633 317. Стадія вибору для ідентифікації трансформованих клітин та/або рослин, в яких інтегрована конструкція за винаходом, може бути проведена завдяки присутності селектованого гену, присутнього в конструкції за винаходом або в плазміді, що її застосовують для трансформації клітин або рослин, та який складає згадану конструкцію. Селектований ген може бути у вигляді химерного гену, що містить такі елементи, функціонально сполучені у напрямку транскрибування: промоторна регуляторна послідовність, що є функціональною в рослинних клітинах, послідовність, що кодує селектований маркер, і термінаторна регуляторна послідовність, що є функціональною в рослинних клітинах. Серед селектованих маркерів, які можуть бути застосовані, можна зазначити маркери, що містять гени для стійкості до антибіотиків, такі як, наприклад, маркери гену гігроміцин фосфотрансферази (Gritz et al., 1983, Gene 25: 179-188), гену неоміцин фосфотрансферази II, що індукують стійкість до канаміцину (Wirtz et al., 1987, DNA, 6(3): 245-253), або гену аміноглікозид 3"-аденілтрансферази, але також маркери, що містять гени для толерантності до гербіцидів, такі як ген bar (White et al., NAR 18: 1062, 1990) для толерантності до біалафосу, ген EPSPS (US 5,188,642) для толерантності до гліфосату або ген HPPD (WO 96/38567) для толерантності до ізоксазолів. Можна зазначити також гени, що кодують ферменти, які можна легко ідентифікувати, такі як фермент GUS, білок GFP або гени, що кодують пігменти, або ферменти, що регулюють вироблення пігментів в трансформованих клітинах. Такі селектовані маркерні гени, зокрема, описані в міжнародних патентних заявках WO 91/02071, WO 95/06128, WO 96/38567 та WO 97/04103. Цей винахід також стосується способу одержання трансгенної рослинної клітини або 8 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 рослини, здатної експресувати длРНК, що інгібує ген сахаропін дегідрогенази грибів або ооміцетів, де згаданий спосіб містить стадії трансформування рослинної клітини за допомогою химерного гену або генетичної конструкції за винаходом. Спосіб може додатково містити стадію вибору рослинної клітини, яку було трансформовано. В окремому втіленні винахід стосується способу одержання трансгенної рослинної клітини або рослини, здатної експресувати длРНК, що інгібує ген сахаропін дегідрогенази грибів або ооміцетів, де згаданий спосіб містить стадії трансформування рослинної клітини за допомогою химерного гену або генетичної конструкції за винаходом, і де згаданою рослинною клітинною є рослинна клітина соєвих бобів, олійного насіння, рису чи картоплі або згаданою рослиною є рослина соєвих бобів, олійного насіння, рису або картоплі. Цей винахід також стосується трансформованих рослин або їх частин і рослин або їх частин, які виведені шляхом вирощування та/або схрещування згаданих регенерованих рослин та насіння трансформованих рослин. Цей винахід також стосується кінцевих продуктів, таких як борошно, олія, волокно, що їх одержують з рослин, їх частин або насіння за винаходом. Для одержання клітин або рослин відповідно до винаходу спеціаліст у цій галузі зможе застосувати один з численних відомих способів трансформування. Один з цих способів полягає у приведенні клітин або тканин організмів-хазяїнів, що підлягають трансформуванню, у контакт з поліетиленгліколем (PEG) та векторами за винаходом (Chang and Cohen, 1979, Mol. Gen. Genet. 168(1), 111-115; Mercenier and Chassy, 1988, Biochimie 70(4), 503-517). Іншим способом є електропорація, що полягає у зазнаванні клітинами або тканинами, що підлягають трансформуванню, та векторами за винаходом дії електричного поля (Andreason and Evans, 1988, Biotechniques 6(7), 650-660; Shigekawa and Dower, 1989, Aust. J. Biotechnol. 3(1), 56-62). Інший спосіб полягає у безпосередньому введенні векторів у клітини або тканини шляхом мікроін'єкцій (Gordon and Ruddle, 1985, Gene 33(2), 121136). Переважно може бути застосований "біолістичний" метод. Він полягає в бомбардуванні клітин або тканин частками, в які адсорбуються вектори за винаходом (Bruce et al., 1989, Proc. Natl. Acad. Sci. USA 86(24), 9692-9696; Klein et al., 1992, Biotechnology 10(3), 286-291; патент США №4,945,050). Переважно трансформування рослинних клітин або тканин може бути здійснене із застосуванням бактерій роду Agrobacterium, переважно інфікуванням клітин або тканин згаданих рослин A. tumefaciens (Knopf, 1979, Subcell. Biochem. 6, 143-173; Shaw et al., 1983, Gene 23(3): 315-330) або A. rhizogenes (Bevan and Chilton, 1982, Annu. Rev. Genet. 16: 357384; Tepfer and Casse-Delbart, 1987, Microbiol. Sci. 4(1), 24-28). Переважно трансформування рослинних клітин або тканин Agrobacterium tumefaciens здійснюють відповідно до протоколу, описаного Hiei et al., (1994, Plant J. 6(2): 271-282). Спеціалісти у цій галузі виберуть підходящий метод відповідно до характеру організмів-хазяїнів, що підлягають трансформуванню. Рослини відповідно до винаходу містять трансформовані рослинні клітини, як визначено вище. Зокрема, трансформовані рослини можуть бути одержані шляхом регенерації описаних вище трансформованих рослинних клітин. Регенерації досягають будь-яким підходящим способом, що залежить від властивостей виду. Винахід також містить частини цих рослин та потомство цих рослин. Термін "частина цих рослин" означає будь-який орган цих рослин, наземний або підземний. Наземними органами є стебла, листя та квіти, що містять чоловічі та жіночі репродуктивні органи. Підземними органами є здебільшого коріння, але ними також можуть бути бульби. Термін "потомство" означає здебільшого насіння, що містить зародки, виведені розмноженням цих рослин один від одного. Крім того, термін "потомство" стосується всього насіння, утвореного з кожного нового покоління, виведеного шляхом схрещування між трансформованими рослинами відповідно до винаходу. Потомство та насіння також можуть бути одержані вегетативним розмноженням згаданих трансформованих рослин. Насіння відповідно до винаходу може бути покрите агрохімічною композицією, що містить щонайменше один активний продукт, який має активність, вибрану з фунгіцидної, гербіцидної, інсектицидної, нематицидної, бактерицидної або віроцидної активностей. Винахід також стосується способу боротьби з патогеном рослин, зокрема з грибами або ооміцетами, що полягає в обробці згаданого патогену молекулою длРНК за винаходом і як визначено тут. В окремому втіленні винаходу спосіб стосується способу боротьби з патогеном рослин, зокрема з грибами або ооміцетами, що полягає в обробці згаданого патогену длРНК за винаходом і як визначено тут або композицією, що містить згадану длРНК, де згаданим патогеном рослин є Magnaporthe grisea, Phytophthora infestans, Sclerotinia sclerotiorum або Phakopsora pachyrhizi. 9 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 В окремому втіленні винаходу спосіб стосується способу боротьби з патогеном рослин, зокрема з грибами або ооміцетами, що полягає в обробці згаданого патогену молекулою длРНК за винаходом і як визначено тут або композицією, що містить згадану длРНК, де згаданою рослиною є рослина соєвих бобів, олійного насіння, рису або картоплі. Винахід також стосується способу боротьби з патогеном рослин, сільськогосподарських культур або насіння, особливо з грибами або ооміцетами, який відрізняється тим, що агрономічно ефективну та по суті нефітотоксичну кількість молекули длРНК за винаходом або композиції за винаходом наносять для обробки насіння, листя, стебел, зрошення або кроплення (внесення з поливною водою) насіння, рослини або плоду рослини або ґрунту або вносять до інертного субстрату (наприклад, неорганічних субстратів, таких як пісок, мінеральна вата, скловата; спучені мінерали типу перліту, вермікуліту, цеоліту або спучена глина), пемзи, пірокластичного матеріалу або наповнювача, до синтетичних органічних субстратів (наприклад, поліуретану), органічних субстратів (наприклад, торфу, компосту, деревинних відходів типу кокосового волокна, деревного волокна або тирси, деревної кори) або до рідкого субстрата (наприклад, в плавальних гідропонних системах, при застосуванні техніки поживного шару, аеропоніки), де рослина росте або де її вирощування є бажаним. Відповідно, винахід стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, який відрізняється тим, що ефективну кількість молекули длРНК за винаходом або композиції за винаходом вносять у ґрунт, де ростуть рослини або де вони здатні рости, наносять на листя та/або плоди рослин або на насіння рослин. В окремому втіленні винаходу винахід стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, який відрізняється тим, що ефективну кількість молекули длРНК за винаходом або композиції за винаходом вносять у ґрунт, де ростуть рослини або де вони здатні рости, наносять на листя та/або плоди рослин або на насіння рослин, де згаданим патогеном рослин є Magnaporthe grisea, Phytophthora infestans, Sclerotinia sclerotinium або Phakopsora pachyrhizi. В окремому втіленні винаходу винахід стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, який відрізняється тим, що ефективну кількість молекули длРНК за винаходом або композиції за винаходом вносять у ґрунт, де ростуть рослини або де вони здатні рости, наносять на листя та/або плоди рослин або на насіння рослин, де згаданою рослиною є рослина соєвих бобів, олійного насіння, рису або картоплі. Вираз "наносять на рослини, що підлягають обробці" розуміють як такий, що означає, для цілей цього винаходу, що пестицидна композиція, яка є об'єктом цього винаходу, може бути нанесена за допомогою різноманітних способів обробки, таких як: - обприскування надземних частин згаданих рослин рідиною, що містить одну зі згаданих композицій, - опилювання, внесення в ґрунт гранул або порошків, обприскування навколо згаданих рослин та, у разі дерев, упорскування або обмазування, - покриття або плівкове покриття насіння згаданих рослин за допомогою рослино-захисної суміші, що містить один зі згаданих компонентів. Спосіб відповідно до винаходу може бути або лікувальним, або превентивним, або ерадикативним (нищівним). У цьому способі композиція, що її використовують, може бути приготована заздалегідь шляхом змішування двох або більше активних сполук за винаходом. Відповідно до альтернативи цьому способу також можна застосовувати одночасно, послідовно або окремо сполуки (A) та (B) для досягнення кон'югованого ефекту (A)/(B) відмінних композицій, кожна з яких має один із двох або трьох активних інгредієнтів (A) або (B). Доза активної сполуки длРНК, яку звичайно застосовують у способі обробки відповідно до винаходу, звичайно та переважно становить: - для обробки листя: від 0,0001 до 10,000 г/га, краще від 0,0001 до 1000 г/га, ще краще від 0,001 до 300 г/га; у разі зрошування або кроплення доза навіть може бути знижена, особливо при застосуванні інертних субстратів типу мінеральної вати або перліту; - для обробки насіння: від 0,0001 до 200 г на 100 кг насіння, краще від 0,001 до 150 г на 100 кг насіння; - для обробки ґрунту: від 0,0001 до 10,000 г/га, краще від 0,001 до 5,000 г/га. При змішуванні длРНК за винаходом з іншої активною фітофармацевтичною сполукою або сполукою, що сприяє росту рослин, згадану фітофармацевтичну сполуку або сполуку, що сприяє росту рослин, використовують у дозі, яку використовують звичайно. Згаданими фітофармацевтичними сполуками або сполуками, що сприяють росту рослин, можуть бути фунгіцидна, гербіцидна, інсектицидна, нематицидна, акарицидна, молюскоцидна 10 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 сполука, стимулятор стійкості, антидоти або сигнальні сполуки. Доза фітофармацевтичної активної сполуки, яку звичайно застосовують у способі обробки відповідно до винаходу, звичайно і переважно становить від 10 до 800 г/га, краще від 50 до 300 г/га для обробки листя. Доза активної речовини, що її наносять, звичайно і переважно становить від 2 до 200 г на 100 кг насіння, переважно від 3 до 150 г на 100 кг насіння у разі обробки насіння. Вказані тут дози наведені як ілюстративні приклади способу відповідно до винаходу. Спеціаліст у цій галузі знатиме, як адаптувати дози нанесення, особливо з урахуванням характерних властивостей рослини або сільськогосподарської культури, що підлягає обробці. За особливих умов, наприклад, відповідно до характерних властивостей патогену, фітопатогенних грибів або ооміцетів, що підлягають обробці або боротьбі, менша доза може забезпечити адекватний захист. Певні кліматичні умови, стійкість або інші природні фактори (характерні особливості) патогену або ступінь зараження, наприклад, рослин цими патогенами, можуть вимагати більших доз комбінованих активних інгредієнтів. Оптимальна доза звичайно залежить від декількох факторів, наприклад, від виду патогену, що підлягає обробці, від виду або рівня розвитку зараженої рослини або рослинного матеріалу, від щільності вегетації або, альтернативно, від способу застосування. Не становлячи обмеження, сільськогосподарською рослиною, що її обробляють пестицидною композицією або комбінацією відповідно до винаходу, є, наприклад, виноградна лоза, хлібні злаки, овочі, люцерна, соєві боби, городні культури, торф, деревина, дерева або садові культури. Спосіб обробки відповідно до винаходу також може бути придатним для обробки матеріалу для розмноження, такого як бульби або кореневища, але також насіння, розсада або висаджувана розсада та рослини або висаджувані рослини. Цей спосіб обробки також може бути придатним для обробки коріння. Спосіб обробки відповідно до винаходу також може бути придатним для обробки надземних частин рослини, таких як стовбури, стебла або черешки, листя, квіти та плоди відповідної рослини, і загалом кожний матеріал, чутливий до грибкової інфекції (наприклад, внаслідок зберігання типу сіна). Винахід також стосується способу боротьби з патогеном рослин, особливо грибами або ооміцетами, що полягає в забезпеченні рослини-хазяїна згаданого патогену рослин трансформованою рослинною клітиною відповідно до винаходу. Винахід також стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, що полягає в забезпеченні рослини-хазяїна згаданого патогену рослин трансформованою рослинною клітиною, що містить длРНК, визначену тут. Винахід також стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, що полягає в трансформуванні рослини генетичною конструкцією відповідно до винаходу. Винахід також стосується способу боротьби з патогеном рослин, особливо з грибами або ооміцетами, що включає такі стадії: i) трансформування рослинної клітини химерним геном відповідно до винаходу; ii) розміщення трансформованих таким чином клітин в умовах, що дозволяють транскрибування згаданої конструкції; iii) приведення клітин у контакт з патогеном. В окремому втіленні винаходу способи за винаходом полягають у боротьбі з патогеном рослин, вибраним з Magnaporthe grisea, Phytophthora infestans, Sclerotinia sclerotinium або Phakopsora pachyrhizi. В окремому втіленні винаходу способи за винаходом полягають у боротьбі з патогеном рослин, де рослиною є рослина соєвих бобів, олійного насіння, рису або картоплі. Винахід також стосується способу інгібування експресії патогену рослин, особливо грибів або ооміцетів, гену сахаропін дегідрогенази, що включає такі стадії: i) трансформування рослинної клітини химерним геном відповідно до винаходу; ii) розміщення трансформованих таким чином клітин в умовах, що дозволяють транскрибування згаданої конструкції; iii) приведення клітин у контакт з патогеном. В окремому втіленні винаходу спосіб за винаходом полягає в інгібуванні гену сахаропін дегідрогенази грибів або ооміцетів, де грибами або ооміцетами є Magnaporthe grisea, Phytophthora infestans, Sclerotinia sclerotinium або Phakopsora pachyrhizi. В окремому втіленні винаходу спосіб за винаходом інгібує ген сахаропін дегідрогенази грибів або ооміцетів, де метод включає такі стадії: i) трансформування рослинної клітини химерним геном відповідно до винаходу; 11 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 ii) розміщення трансформованих таким чином клітин в умовах, що дозволяють транскрибування згаданої конструкції; iii) приведення клітин у контакт з патогеном; де рослиною є рослина соєвих бобів, олійного насіння, рису або картоплі. Відповідно до винаходу можуть бути оброблені всі рослини та частини рослин. Під рослинами розуміють всі рослини та популяції рослин, такі як бажані та небажані дикорослі рослини, культивари та сорти рослин (що охороняються або не охороняються правами на сорти рослин або правами агрономів-селекціонерів). Культиварами та сортами рослин можуть бути рослини, одержані звичайними методами розмноження та вирощування, яким можуть допомагати або які можуть бути доповнені одним або більше біотехнологічними методами, такими як застосування подвійних гаплоїдів, злиття протопластів, випадковий та спрямований мутагенез, молекулярні або генетичні маркери, або методами біоінженерії та генної інженерії. Під частинами рослин розуміють всі частини та органи рослин, що розміщені над ґрунтом та під ґрунтом, такі як паростки, листя, цвіт та коріння, до яких відносять, наприклад, листя, голки, стебла, гілки, цвіт, плодоносні тіла, плоди та насіння, а також корені, бульбоцибулини та кореневища. Сільськогосподарські культури та вегетативний і генеративний розмножувальний матеріал, наприклад, черенки, бульбоцибулини, кореневища, пагони та насіння, також відносяться до частин рослин. Серед рослин, які можуть бути захищені за допомогою способу відповідно до винаходу, можна зазначити основні польові культури, такі як кукурудза, соя, бавовна, олійне насіння сімейства Brassica, таке як Brassica napus (наприклад, канола), Brassica rapa, B. juncea (наприклад, гірчиця) та Brassica carinata, рис, пшениця, цукровий буряк, цукрова тростина, овес, жито, ячмінь, просо, тритикале, льон, виноградна лоза та різні фрукти та овочі різних ботанічних таксонів, такі як види Rosaceae (наприклад, односім'яні плоди, такі як яблука та груші, але також кісточкові фрукти, такі як абрикоси, вишні, мигдаль і персики, ягідні культури, такі як полуниця), Ribesioidae, Juglandaceae, Betulaceae, Anacardiaceae, Fagaceae, Moraceae, Oleaceae, Actinidaceae, Lauraceae, Musaceae (наприклад, бананові дерева та плантації), Rubiaceae (наприклад, кава), Theaceae, Sterculiceae, Rutaceae (наприклад, лимони, апельсини та грейпфрут); Solanaceae (наприклад, томати, картопля, перець, баклажани), Liliaceae, Compositiae (наприклад, салат-латук, артишок і цикорій, у тому числі корінь цикорію, цикорійендивій або звичайний цикорій), Umbelliferae (наприклад, морква, петрушка та селера), Cucurbitaceae (наприклад, огірки - у тому числі корнішони, гарбузи, кавуни, бахчеві культури та дині), Alliaceae (наприклад, цибуля та лук-порей), Cruciferae (наприклад, білокачанна капуста, червона капуста, броколі, цвітна капуста, брюссельська капуста, пекінська капуста, кольрабі, редис, хрін, крес-салат, китайська капуста), Leguminosae (наприклад, арахіс, горох та бобові такі як квасоля, що в'ється, та кормові боби), Chenopodiaceae (наприклад, буряк кормовий (мангольд), буряк листовий, шпінат, столовий буряк), Malvaceae (наприклад, окра), Asparagaceae (наприклад, аспарагус); садові та лісові культури; декоративні рослини; а також генно-модифіковані гомологи цих культур. Спосіб обробки за винаходом може бути використаний при обробці генетично модифікованих організмів (ГМО), наприклад, рослин або насіння. Генетично модифікованими рослинами (або трансгенними рослинами) є рослини, до геному яких був стійко інтегрований гетерологічний ген. Вираз "гетерологічний ген" по суті означає ген, який передбачений або складений поза рослиною і при введенні до ядерного, хлоропластного або мітохондрійного геному надає трансформованій рослині нових або покращених агрономічних чи інших властивостей шляхом експресії білка або поліпептиду, що становить інтерес, або шляхом даунрегуляції чи сайленсингу іншого гену або генів, присутніх у рослині (із застосуванням, наприклад, антисмислової технології, технології співсупресії або РНК інтерференції - RNAiтехнології). Гетерологічний ген, розміщений у геномі, також називають трансгеном. Трансген, визначений своїм особливим розташуванням у геномі рослини, називають трансформаційною або трансгенною подією. Залежно від видів або культиварів рослин, їхнього розташування та умов вирощування (ґрунти, клімат, період вегетації, раціон), обробка за винаходом може також спричинити суперадитивні ("синергійні") ефекти. Так, наприклад, можливе зниження норм нанесення та/або розширення спектру активності та/або підвищення активності активних сполук та композицій, які можуть бути використані за винаходом, кращий ріст рослин, підвищена стійкість до високих або низьких температур, підвищена стійкість до посухи або вмісту солі у воді чи ґрунті, підвищена продуктивність цвітіння, полегшення збирання врожаю, прискорене дозрівання, підвищена врожайність, більші плоди, більша висота рослин, більш зелений колір листя, більш раннє цвітіння, вища якість та/або вища поживна цінність зібраних продуктів, більш висока 12 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 концентрація цукру у плодах, краща стійкість при зберіганні та/або можливість обробки зібраних продуктів, які перевершують звичайно очікувані ефекти. При певних нормах нанесення комбінації активної сполуки за винаходом можуть також мати зміцнювальний ефект у рослин. Відповідно, вони також є підходящими для мобілізації захисної системи рослини проти нападу небажаних мікроорганізмів. Це може бути, якщо це має місце, однією з причин підвищеної активності комбінацій за винаходом, наприклад, проти грибів. Речовини, що зміцнюють рослини (які індукують стійкість), треба розуміти як такі, що означають, у даному контексті, ті речовини або комбінації речовин, які здатні стимулювати захисну систему рослин таким чином, що при подальшому інокулюванні небажаними мікроорганізмами оброблені рослини проявляють істотний ступінь стійкості до цих мікроорганізмів. У даному випадку небажані мікроорганізми треба розуміти як такі, що означають фітопатогенні гриби, бактерії та віруси. Отже, речовини за винаходом можуть бути застосовані для захисту рослин проти нападу згаданих вище патогенів впродовж певного періоду часу після обробки. Період часу, впродовж якого відбувається захист, звичайно триває від 1 до 10 днів, краще від 1 до 7 днів після обробки рослин активними сполуками. Як вже зазначалося вище, можливо обробляти всі рослини та їх частини відповідно до винаходу. У кращому втіленні обробляють дикорослі види рослин та культивари рослин або рослини, одержані традиційними методами біологічного вирощування, такими як схрещування або злиття протопластів, а також їх частини. В іншому кращому втіленні обробляють трансгенні рослини та культивари рослин, одержані методами генної інженерії, за потреби у поєднанні з традиційними методами (генетично-модифіковані організми), та їх частини. Терміни "частини" або "частини рослин" або "рослинні частини" було пояснено вище. Ще краще, якщо рослини культиварів рослин, які є доступними у продажу або які застосовують, обробляють відповідно до винаходу. Розуміють, що культивари рослин означають рослини, що мають нові властивості ("риси") і які були одержані традиційним вирощуванням, мутагенезом або за допомогою технологій рекомбінантної ДНК. Вони можуть бути культиварами, сортами, біо- або генотипами. Спосіб обробки за винаходом може бути використаний при обробці генетично модифікованих організмів (ГМО), наприклад, рослин або насіння. Генетично модифікованими рослинами (або трансгенними рослинами) є рослини, до геному яких був стійко інтегрований гетерологічний ген. Вираз "гетерологічний ген" по суті означає ген, який передбачений або складений поза рослиною і при введенні до ядерного, хлоропластного або мітохондрійного геному надає трансформованій рослині нових або покращених агрономічних чи інших властивостей шляхом експресії білка або поліпептиду, що становить інтерес, або шляхом даунрегуляції чи сайленсингу іншого гену або генів, присутніх у рослині (із застосуванням, наприклад, антисмислової технології, технології співсупресії або РНК інтерференції - РНКiтехнології або мікро-РНК - міРНК - технології). Гетерологічний ген, розміщений у геномі, також називають трансгеном. Трансген, визначений своїм особливим розташуванням у геномі рослини, називають трансформаційною або трансгенною подією. Залежно від видів або культиварів рослин, їхнього розташування та умов вирощування (ґрунти, клімат, період вегетації, раціон), обробка за винаходом може також спричинити суперадитивні ("синергійні") ефекти. Так, наприклад, можливе зниження норм нанесення та/або розширення спектру активності та/або підвищення активності активних сполук та композицій, які можуть бути використані за винаходом, кращий ріст рослин, підвищена стійкість до високих або низьких температур, підвищена стійкість до посухи або вмісту солі у воді чи ґрунті, підвищена продуктивність цвітіння, полегшення збирання врожаю, прискорене дозрівання, підвищена врожайність, більші плоди, більша висота рослин, більш зелений колір листя, більш раннє цвітіння, вища якість та/або вища поживна цінність зібраних продуктів, більш висока концентрація цукру у плодах, краща стійкість при зберіганні та/або можливість обробки зібраних продуктів, які перевершують звичайно очікувані ефекти. При певних нормах нанесення комбінації активної сполуки за винаходом можуть також мати зміцнювальний ефект у рослин. Відповідно, вони також є підходящими для мобілізації захисної системи рослини проти нападу небажаних мікроорганізмів. Це може бути, якщо це має місце, однією з причин підвищеної активності комбінацій за винаходом, наприклад, проти грибів. Речовини, що зміцнюють рослини (які індукують стійкість), треба розуміти як такі, що означають, у даному контексті, ті речовини або комбінації речовин, які здатні стимулювати захисну систему рослин таким чином, що при подальшому інокулюванні небажаними мікроорганізмами оброблені рослини проявляють істотний ступінь стійкості до цих мікроорганізмів. У даному випадку небажані мікроорганізми треба розуміти як такі, що означають фітопатогенні гриби, бактерії та віруси. Отже, речовини за винаходом можуть бути застосовані для захисту рослин проти нападу згаданих вище патогенів впродовж певного періоду часу після обробки. Період 13 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 часу, впродовж якого відбувається захист, звичайно триває від 1 до 10 днів, краще від 1 до 7 днів після обробки рослин активними сполуками. Рослини та культивари рослин, які краще обробляти відповідно до винаходу, включають усі рослини, які мають генетичний матеріал, що надає особливо сприятливих, корисних рис цим рослинам (одержаних вирощуванням та/або біотехнологічними засобами). Рослини та культивари рослин, які також краще обробляти відповідно до винаходу, є стійкими проти одного або більше біотичного стресу, тобто згадані рослини проявляють кращий захист проти тваринних та мікробних паразитів, таких як нематоди, комахи, кліщі, фітопатогенні гриби, бактерії, віруси та/або віроїди. Приклади стійких до нематодів або комах рослин описані, наприклад, у патентних заявках США №№ 11/765,491, 11/765,494, 10/926,819, 10/782,020, 12/032,479, 10/783,417, 10/782,096, 11/657,964, 12/192,904, 11/396,808, 12/166,253, 12/166,239, 12/166,124, 12/166,209, 11/762,886, 12/364,335, 11/763,947, 12/252,453, 12/209,354, 12/491,396, 12/497,221, 12/644,632, 12/646,004, 12/701,058, 12/718,059, 12/721,595, 12/638,591. Рослинами та культиварами рослин, які також можна обробляти відповідно до винаходу, є рослини, які є стійкими до одного або більше абіотичного стресу. Умови абіотичного стресу можуть включати, наприклад, посуху, зазнання холодної температури, зазнання спеки, осмотичний стрес, затоплення, підвищену солоність ґрунту, підвищений вплив мінералів, вплив озону, вплив яскравого світла, обмежену наявність азотних поживних речовин, обмежену наявність фосфорних поживних речовин, недостатність тіні. Рослинами та культиварами рослин, які також можна обробляти відповідно до винаходу, є рослини, які відрізняються характеристиками підвищеної врожайності. Підвищена врожайність у таких рослин може бути результатом, наприклад, покращеної фізіології, росту та розвитку рослини, таких як ефективність використання води, ефективність утримання води, покращене використання азоту, покращене поглинання вуглецю, покращений фітосинтез, покращена ефективність пророщування та прискорене дозрівання. На врожайність, крім того, може впливати покращена архітектура рослини (за умов стресу та без стресу), у тому числі, але не обмежуючись цим, раннє цвітіння, контролювання цвітіння для одержання гібридного насіння, міцність сіянців, розмір рослини, кількість та відстань міжвузел, ріст коріння, розмір насіння, розмір плодів, розмір стручків, кількість стручків або качанів, кількість насіння на стручок або качан, маса насіння, підвищене наповнення насіння, знижене розосередження насіння, зниження розтріскування стручків та стійкість до полягання. Інші риси врожаю включають склад насіння, такий як вміст вуглеводню, білка, вміст та склад олії, поживна цінність, зниження антипоживних сполук, покращена здатність до обробляння та краща стійкість при зберіганні. Рослинами, які можна обробляти відповідно до винаходу, є гібридні рослини, які вже проявляють характеристики гетерозу або гібридної сили, результатом яких звичайно є більш висока врожайність, міцність, здоров'я та стійкість до факторів біотичного та абіотичного стресу. Такі рослини звичайно одержують шляхом схрещування інбредної батьківської лінії з чоловічим безпліддям (батьки жіночої статі) з іншою інбредною лінією з чоловічою плідністю (батьки чоловічої статі). Гібридне насіння звичайно збирають з чоловічих стерильних рослин і продають садоводам-городникам. Іноді чоловічі стерильні рослини можна одержати (наприклад, у кукурудзи) шляхом видалення мітелок або китиць, тобто механічним видаленням чоловічих репродуктивних органів (або чоловічих квіток), але, що є більш типовим, чоловіча стерильність є результатом генетичних детермінантів у геномі рослини. У такому разі, і особливо тоді, коли насіння є бажаним продуктом, врожай якого треба зібрати з гібридних рослин, звичайно є корисним забезпечити, щоб чоловіча плідність гібридних рослин була повністю відновлена. Це може бути здійснене шляхом забезпечення, щоб батьки чоловічої статі мали відповідні гени відновлення плідності, які здатні відновлювати чоловічу плідність в гібридних рослинах, які містять генетичні детермінанти, що відповідають за чоловіче безпліддя. Генетичні детермінанти чоловічої стерильності можуть бути розташовані у цитоплазмі. Приклади цитоплазмової чоловічої стерильності (CMS), наприклад, були описані у сортів Brassica (WO 92/05251, WO 95/09910, WO 98/27806, WO 05/002324, WO 06/021972 і US 6,229,072). Проте, генетичні детермінанти для чоловічої стерильності також можуть бути розташовані в ядерному геномі. Рослини з чоловічою стерильністю також можуть бути одержані методами рослинної біотехнології, такими як генна інженерія. Особливо корисний засіб одержання рослин з чоловічою стерильністю описаний у WO 89/10396, в якому, наприклад, рибонуклеазу, таку як барназа, селективно експресують у клітинах тапетуму в тичинках. Після цього плідність може бути відновлена експресією у клітинах тапетуму інгібітора рибонуклеази, такого як барстар (наприклад, у WO 1991/02069). Рослини або культивари рослин (одержані методами рослинної біотехнології, такими як 14 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 генна інженерія), які можна обробляти відповідно до винаходу, є толерантними до гербіцидів рослинами, тобто рослинами, що є виносливими до одного або більше конкретних гербіцидів. Такі рослини можуть бути одержані або генною трансформацією, або селекцією рослин, що містять мутацію, що надає таку виносливість відносно гербіцидів. Стійкими до гербіцидів рослинами є, наприклад, гліфосат-толерантні рослини, тобто рослини, яким надана толерантність до гербіцидного гліфосату або його солей. Рослини можна зробити толерантними до гліфосату різними методами. Наприклад,гліфосат-толерантні рослини можна одержати трансформуванням рослини за допомогою гену, що кодує ензим 5енолпірувілшикімат-3-фосфатсинтази (EPSPS). Прикладами таких генів EPSPS є ген AroA (мутант CT7) бактерії Salmonella typhimurium (Science 1983, 221, 370-371), ген CP4 бактерії виду Agrobacterium (Curr. Topics Plant Physiol. 1992, 7, 139-145), гени, що кодують EPSPS петунії (Science 1986, 233, 478-481), EPSPS томатів (J. Biol. Chem. 1988, 263, 4280-4289) або EPSPS коракану (WO 01/66704). Це також може бути мутований EPSPS, як це описано, наприклад, у EP 0837944, WO 00/66746, WO 00/66747 або WO 02/26995. Гліфосат-толерантні рослини також можна одержати експресією гену, що кодує гліфосатний оксидо-редуктазний фермент, як це описано у патентах США №№ US 5,776,760 і US 5,463,175. Гліфосат-толерантні рослини також можна одержати експресією гену, що кодує гліфосатний ацетилтрансферазний фермент, як це описано, наприклад, у WO 02/036782, WO 03/092360, WO 2005/012515 і WO 2007/024782. Гліфосат-толерантні рослини також можна одержати селекцією рослин, що містять природні мутації вищезгаданих генів, як описано, наприклад, у WO 01/024615 або WO 03/013226. Рослини, що експресують гени EPSPS, що надають толерантність до гліфосату, описані, наприклад, у патентних заявках США №№ 11/517,991, 10/739,610, 12/139,408, 12/352,532, 11/312,866, 11/315,678, 12/421,292, 11/400,598, 11/651,752, 11/681,285, 11/605,824, 12/468,205, 11/760,570, 11/762,526, 11/769,327, 11/769,255, 11/943801 або 12/362,774. Рослини, що містять інші гени, що надають толерантність до гліфосату, такі як гени декарбоксилази, описані, наприклад, у патентних заявках США №№ 11/588,811, 11/185,342, 12/364,724, 11/185,560 або 12/423,926. Іншими стійкими до гербіцидів рослинами є, наприклад, рослини, яким надана толерантність до гербіцидів, що інгібують ензимглютамінсинтазу, таких як біалафос, фосфінотрицин або глюфозинат. Такі рослини можна одержати експресією ензиму, що детоксифікує гербіцид, або мутантного глютамінсинтазного ензиму, стійкого до інгібування, наприклад, описаного у патентній заявці США №11/760,602. Одним із таких ефективних детоксифікувальних ензимів є ензим, що кодує фосфінотрицин ацетилтрансферазу (такий як білок бар або пет виду Streptomyces). Рослини, що експресують екзогенну фосфінотрицин ацетилтрансферазу, описані, наприклад, в патентах США №№ 5,561,236; 5,648,477; 5,646,024; 5,273,894; 5,637,489; 5,276,268; 5,739,082; 5,908,810 і 7,112,665. Іншими гербіцид-толерантними рослинами є також рослини, яким надана толерантність до гербіцидів, що інгібують ензимну гідроксифенілпіруват-діоксигеназу (HPPD). HPPD є ензимом, що каталізують реакцію, в якій пара-гідроксифенілпіруват (HPP) трансформується у гомогентисат. Рослини, толерантні до HPPD-інгібіторів, можуть бути трансформовані за допомогою гену, що кодує природний стійкий HPPD ензим, або гену, що кодує мутований або химерний HPPD ензим, як це описано у міжнародних заявках WO 96/38567, WO 99/24585, WO 99/24586, WO 09/144079, WO 02/046387, або патенті США US 6,768,044. Толерантність до HPPD-інгібіторів також може бути досягнута шляхом трансформування рослин за допомогою генів, які кодують певні ензими, що дозволяє утворення гомогентисату, незважаючи на інгібування нативного HPPD ензиму HPPD-інгібітором. Такі рослини та гени описані в міжнародних заявках WO 99/34008 і WO 02/36787. Толерантність рослин до HPPD інгібіторів може бути також покращена трансформуванням рослин за допомогою гену, що кодує ензим, який має активність префенатдегідрогенази (PDH), на додаток до гену, що кодує HPPDтолерантний ензим, як це описано у міжнародній заявці WO 04/024928. Крім того, рослинам може бути надана більша толерантність до HPPD-інгібіторних гербіцидів шляхом додавання до їхнього геному гену, що кодує ензим, здатний до засвоєння або руйнування HPPD-інгібіторів, таких як ензими CYP450, показані в міжнародних заявках WO 2007/103567 і WO 2008/150473. Ще іншими стійкими до гербіцидів рослинами є рослини, яким надана толерантність до інгібіторів ацетолактатсинтази (ALS). Відомі інгібітори ALS включають, наприклад, сульфонілсечовину, імідазолінон, тріазолпіримідини, піримідинілокси(тіо)бензоати та/або сульфоніламінокарбонілтріазолiнонові гербіциди. Різні мутації в ензимі ALS (також відомої як ацетогідроксикислотна синтаза, AHAS) відомі як такі, що надають толерантність до різних гербіцидів і груп гербіцидів, як це описано, наприклад, у Tranel і Wright (Weed Science ("Наука про бур'яни") 2002, 50, 700-712), але також у патентах США №№ 5,605,011, 5,378,824, 5,141,870 15 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 і 5,013,659. Одержання толерантних до сульфонілсечовини рослин і толерантних до імідазолінону рослин описано у патентах США №№ 5,605,011; 5,013,659; 5,141,870; 5,767,361; 5,731,180; 5,304,732; 4,761,373; 5,331,107; 5,928,937; і 5,378,824; та в міжнародній заявці WO 96/33270. Інші толерантні до імідазолінону рослини також описані, наприклад, у міжнародних заявках WO 2004/040012, WO 2004/106529, WO 2005/020673, WO 2005/093093, WO 2006/007373, WO 2006/015376, WO 2006/024351 та WO 2006/060634. Інші толерантні до сульфонілсечовини та імідазолінону рослини також описані, наприклад, у міжнародній заявці WO 2007/024782 та патентній заявці США № 61/288958. Інші рослини, толерантні до імідазолінону та/або сульфонілсечовини, можуть бути одержані індукованим мутагенезом, селекцією в культурах клітин у присутності гербіциду або мутаційним вирощуванням, як описано, наприклад, для бобів сої у патенті США 5,084,082, для рису в міжнародній заявці WO 97/41218, для цукрового буряку у патенті США 5,773,702 і міжнародній заявці WO 99/057965, для салата-латука у патенті США 5,198,599 або для соняшника в міжнародній заявці WO 01/065922. Рослини або культивари рослин (одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є стійкими до комах трансгенними рослинами, тобто рослинами, яким надана стійкість до нападу певних цільових комах. Такі рослини можуть бути одержані генетичною трансформацією або селекцією рослин, що містять мутацію, яка забезпечує таку стійкість до комах. Використаний у цьому описі вираз "стійка до комах трансгенна рослина" включає будь-яку рослину, що містить принаймні один трансген з кодувальною послідовністю, яка кодує: 1) інсектицидний кристалічний білок Bacillus thuringiensis або його інсектицидну частку, такий як інсектицидні кристалічні білки, перелік яких наведено авторами Crickmore та ін. (1998, Microbiology and Molecular Biology Reviews ("Огляди мікробіології та молекулярної біології"), 62: 807-813), видання було оновлене авторами Crickmore та ін. (2005) у номенклатурі токсинів Bacillus thuringiensis, адреса он-лайн: http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/), або його інсектицидні частки, наприклад, білки класів білків Cry Cry1Ab, Cry1Ac, Cry1B, Cry1C, Cry1D, Cry1F, Cry2Ab, Cry3Aa або Cry3Bb або їхні інсектицидні частки (наприклад, EP-A 1 999 141 і WO 2007/107302), або такі білки, кодовані синтетичними генами, як, наприклад, описані у патентній заявці США № 12/249,016; або 2) кристалічний білок виду Bacillus thuringiensis або його частку, що є інсектицидною у присутності другого іншого кристалічного білка виду Bacillus thuringiensis або його частки, такого як бінарний токсин, складений з кристалічних білків Cry34 і Cry35 (Nat. Biotechnol. 2001, 19, 66872; Applied Environm. Microbiol. 2006, 71, 1765-1774) або бінарний токсин, складений з білків Cry1A або Cry1F і білків Cry2Aa або Cry2Ab або Cry2Ae (патентні заявки США № 12/214,022 і EP-A 2 300 618); або 3) гібридний інсектицидний білок, що містить частки різних інсектицидних кристалічних білків виду Bacillus thuringiensis, такий як гібрид білків 1) вище або гібрид білків 2) вище, наприклад, білок Cry1A.105, продукований подією у кукурудзі MON98034 (WO 2007/027777); або 4) білок будь-якого з 1) по 3) вище, в якому деякі, зокрема з 1 по 10, амінокислоти були заміщені іншою амінокислотою для одержання більш високої інсектицидної активності до цільового виду комах, та/або для розширення діапазону уражених цільових видів комах, та/або внаслідок змін, введених до кодувальної ДНК при клонуванні або трансформуванні, такий як білок Cry3Bb1 у подіях у кукурудзі MON863 або MON88017, або білок Cry3A у події у кукурудзі MIR604; або 5) інсектицидний секретований білок з виду Bacillus thuringiensis або Bacillus cereus або його інсектицидна частка, такий як вегетативні інсектицидні білки (VIP), перелік яких наведено на сайті: http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/vip.html, наприклад, білки з класу білків VIP3Aa; або 6) секретований білок з Bacillus thuringiensis або Bacillus cereus, який є інсектицидним у присутності другого секретованого білка з Bacillus thuringiensis або B. cereus, такий як бінарний токсин, складений з білків VIP1A і VIP2A (WO 94/21795); або 7) гібридний інсектицидний білок, що містить частки різних секретованих білків з Bacillus thuringiensis або Bacillus cereus, такий як гібрид білків за 1) вище або гібрид білків за 2) вище; або 8) білок будь-якого з 5) по 7) вище, в якому деякі, зокрема з 1 по 10, амінокислоти були замінені іншою амінокислотою для одержання більш високої інсектицидної активності відносно цільового виду комах, та/або для розширення діапазону уражених цільових видів комах, та/або внаслідок змін, внесених до кодувальної ДНК при клонуванні або трансформуванні (при 16 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 продовженні кодування інсектицидного білка), такий як білок VIP3Aa у події в бавовні COT102; або 9) секретований білок з Bacillus thuringiensis або Bacillus cereus, який є інсектицидним у присутності кристалічного білка з Bacillus thuringiensis, такий як бінарний токсин, складений з VIP3 і Cry1A або Cry1F (патентні заявки США №№ 61/126083 і 61/195019), або бінарний токсин, складений з білка VIP3 та білків Cry2Aa або Cry2Ab або Cry2Ae (патентні заявки США № 12/214,022 і EP-A 2 300 618); 10) білок за 9) вище, в якому деякі, особливо з 1 по 10, амінокислоти були заміщені іншою амінокислотою для одержання більш високої інсектицидної активності відносно цільового виду комах, та/або для розширення діапазону уражених цільових видів комах, та/або внаслідок змін, внесених до кодувальної ДНК при клонуванні або трансформуванні (при продовженні кодування інсектицидного білка). Звичайно, до стійких до комах трансгенних рослин, як це використано у цьому описі, також відносяться будь-які рослини, що містять комбінацію генів, що кодують білки будь-якого із вказаних вище з 1 по 10 класів. В одному втіленні, стійка до комах рослина містить понад один трансген, що кодує білок будь-якого із вказаних вище з 1 по 10 класів, для розширення діапазону уражених цільових видів комах при використанні різних білків, спрямованих проти різних цільових видів комах або для затримання розвитку стійкості до комах у рослинах із використанням різних білків, що є інсектицидними для тих самих цільових видів комах, але мають інший механізм дії, такий як зв'язування з різними сайтами звязування рецепторів у комасі. Термін "стійка до комах трансгенна рослина", використаний у цьому описі, також охоплює будь-яку рослину, що містить щонайменше один трансген, який містить послідовність, що продукує після експресії дволанцюгову РНК, яка після усмоктування комахою-шкідником рослин інгібує ріст цієї комахи-шкідника, як описано, наприклад, у міжнародних патентних заявках WO 2007/080126, WO 2006/129204, WO 2007/074405, WO 2007/080127 і WO 2007/035650. Рослини або культивари рослин (одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є толерантними до абіотичних стресів. Такі рослини можна одержати генетичною трансформацією або селекцією рослин, які містять мутацію, що забезпечує таку стійкість до стресу. До особливо придатних рослин, толерантних до стресу, відносяться: 1) рослини, які містять трансген, здатний знижувати експресію та/або активність полі(ADPрибоза)полімеразного (PARP) гену в клітинах рослини або рослинах, як це описано у заявках WO 00/04173, WO 2006/045633, EP-A 1 807 519 або EP-A 2 018 431; 2) рослини, які містять трансген, що підсилює толерантність до стресу, здатний знижувати експресію та/або активність PARG кодувальних генів рослин або клітин рослин, як це описано, наприклад, у WO 2004/090140; 3) рослини, які містять трансген, що підсилює толерантність до стресу, який кодує рослиннофункціональний ензим шляхом нікотинамідаденін динуклеотидного реутилізаційного (відновлювального) синтезу, у тому числі нікотинамідазу, нікотинат фосфорибозилтрансферазу, мононуклеотид аденілтрансферазу нікотинової кислоти, нікотинамідаденін динуклеотидсинтетазу або нікотинамід фосфорибозилтрансферазу, як це описано, наприклад, у EP-A 1 794 306, WO 2006/133827, WO 2007/107326, EP-A 1 999 263 або WO 2007/107326. Рослини або культивари рослин (одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, проявляють змінену кількість, якість та/або стабільність зберігання зібраного продукту та/або змінені властивості конкретних інгредієнтів зібраного продукту, наприклад: 1) трансгенні рослини, які синтезують модифікований крохмаль, що за своїми фізикохімічними характеристиками, зокрема за вмістом амілози або співвідношенням амілоза/амілопектин, ступенем розгалуження, середньою довжиною ланцюга, розподілом бокового ланцюга, поведінкою в'язкості, стійкістю гелеутворення, розміром зернин крохмалю та/або морфологією зернини крохмалю, змінюється порівняно із синтезованим крохмалем у клітинах рослин або рослинах дикого типу, так що він кращим чином підходить для спеціального застосування. Такі трансгенні рослини, що синтезують модифікований крохмаль, розкриті, наприклад, у EP-A 0 571 427, WO 95/04826, EP-A 0 719 338, WO 96/15248, WO 96/19581, WO 96/27674, WO 97/11188, WO 97/26362, WO 97/32985, WO 97/42328, WO 97/44472, WO 97/45545, WO 98/27212, WO 98/40503, WO 99/58688, WO 99/58690, WO 99/58654, WO 00/08184, WO 00/08185, WO 00/08175, WO 00/28052, WO 00/77229, WO 01/12782, WO 01/12826, WO 02/101059, WO 03/071860, WO 04/056999, WO 05/030942, WO 2005/030941, WO 2005/095632, WO 2005/095617, WO 2005/095619, WO 2005/095618, WO 2005/123927, WO 2006/018319, WO 17 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 2006/103107, WO 2006/108702, WO 2007/009823, WO 00/22140, WO 2006/063862, WO 2006/072603, WO 02/034923, WO 2008/017518, WO 2008/080630, WO 2008/080631, EP 07090007.1, WO 2008/090008, WO 01/14569, WO 02/79410, WO 03/33540, WO 2004/078983, WO 01/19975, WO 95/26407, WO 96/34968, WO 98/20145, WO 99/12950, WO 99/66050, WO 99/53072, US 6,734,341, WO 00/11192, WO 98/22604, WO 98/32326, WO 01/98509, WO 01/98509, WO 2005/002359, US 5,824,790, US 6,013,861, WO 94/04693, WO 94/09144, WO 94/11520, WO 95/35026, WO 97/20936, WO 2010/012796, WO 2010/003701, 2) трансгенні рослини, які синтезують некрохмальні вуглеводневі полімери або некрохмальні вуглеводневі полімери зі зміненими властивостями порівняно з рослинами дикого типу без генної модифікації. Прикладами є рослини, що виробляють поліфруктозу, особливо інулінового та леванового типів, розкриті у EP-A 0 663 956, WO96/01904, WO 96/21023, WO 98/39460 та WO 99/24593, рослини, що виробляють альфа-1,4-глюкани, розкриті у WO 95/31553, US 2002031826, US 6,284,479, US 5,712,107, WO 97/47806, WO 97/47807, WO 97/47808 та WO 00/14249, рослини, що виробляють альфа-1,6 розгалужені альфа-1,4-глюкани, розкриті у WO 00/73422, рослини, що виробляють алтернан, розкриті, наприклад, у WO 00/47727, WO 00/73422, EP 06077301.7, US 5,908,975 та EP-A 0 728 213, 3) трансгенні рослини, які виробляють гіалуронан, як, наприклад, ті, що розкриті у WO 2006/032538, WO 2007/039314, WO 2007/039315, WO 2007/039316, JP-A 2006-304779 та WO 2005/012529, 4) трансгенні рослини або гібридні рослини, такі як цибуля, з характеристиками на зразок "високий вміст розчинних твердих речовин", "низька гострота" (LP) та/або "тривале зберігання" (LS), як описано у патентних заявках США №№ 12/020,360 та 61/054,026. Рослинами або культиварами рослин (що можуть бути одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є рослини, такі як рослини бавовни, зі зміненими характеристиками волокна. Такі рослини можуть бути одержані генною трансформацією або селекцією рослин, які містять мутацію, що забезпечує такі змінені характеристики волокна, і до них відносяться: a) рослини, такі як рослини бавовни, які містять змінену форму целюлозасинтазних генів, як ті, що описані у WO 98/00549; b) рослини, такі як рослини бавовни, які містять змінену форму rsw2 або rsw3 гомологічних нуклеїнових кислот, як ті, що описані у WO 2004/053219; c) рослини, такі як рослини бавовни, з підвищеною експресією цукрозафосфатсинтази, як ті, що описані у WO 01/17333; d) рослини, такі як рослини бавовни, з підвищеною експресією цукрозасинтази, як ті, що описані у WO 02/45485; e) рослини, такі як рослини бавовни, в яких змінена синхронізація плазмодесматального стробування в основі клітини волокна, наприклад шляхом даун-регуляції волокноселективної β1,3-глюканази, як ті, що описані у WO 2005/017157, або як ті, що описані у WO 2009/143995; f) рослини, такі як рослини бавовни, які мають волокна зі зміненою реактивністю, наприклад шляхом експресії N-ацетилглюкозамін-трансферазного гену, у тому числі nodC та хітинсинтазних генів, як ті, що описані у WO 2006/136351. Рослинами або культиварами рослин (що можуть бути одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є рослини, такі як олійний рапс або споріднені рослини сорту Brassica, зі зміненими характеристиками олійного профілю. Такі рослини можна одержати генною трансформацією або селекцією рослин, які містять мутацію, що надає таких змінених характеристик олійного профілю, до яких відносяться: a) рослини, такі як рослини олійного рапсу, які виробляють олію, що має високий вміст олеїнової кислоти, як ті, що описані, наприклад, у патентах США US 5,969,169, US 5,840,946 або US 6,323,392 або US 6,063,947; b) рослини, такі як рослини олійного рапсу, які виробляють олію, що має низький вміст лінолевої кислоти, як ті, що описані у патентах США US 6,270,828, US 6,169,190 або US 5,965,755; c) рослини, такі як рослини олійного рапсу, які виробляють олію, що має низький рівень насичених жирних кислот, як ті, що описані, наприклад, у патенті США No. 5,434,283 або патентній заявці США №12/668303. Рослинами або культиварами рослин (що можуть бути одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є рослини, такі як олійний рапс або споріднені рослини сорту Brassica, зі зміненими характеристиками обсипання зерна. Такі рослини можуть бути одержані генною 18 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 трансформацією або селекцією рослин, які містять мутацію, що надає таких змінених характеристик обсипання зерна, і до них відносяться рослини, такі як рослини олійного рапсу, із уповільненим або зниженим обсипанням зерна, як це описано у патентній заявці США № 61/135,230, міжнародних заявках WO 2009/068313 і WO 2010/006732. Рослинами або культиварами рослин (що можуть бути одержані методами рослинної біотехнології, такими як генна інженерія), які також можна обробляти відповідно до винаходу, є рослини, такі як рослини тютюну, зі зміненими моделями пост-трансляційної модифікації білка, наприклад, як ті, що описані у міжнародних заявках WO 2010/121818 та WO 2010/145846. Особливо придатними трансгенними рослинами, які можна обробляти відповідно до винаходу, є рослини, що містять трансформаційні події або комбінацію трансформаційних подій, що є предметом клопотань для одержання нерегульованого статусу в США до Служби інспекції здоров'я тварин і рослин (APHIS) Міністерства сільського господарства США (USDA), незалежно від того, чи такі клопотання були задоволені або все ще перебувають на розгляді. Ця інформаціяє завжди доступною у APHIS (4700 Рівер Роуд Рівердейл, штат Меріленд 20737, США), наприклад, на її сайті в Інтернеті (URL http://www.aphis.usda.gov/brs/not_reg.html). На дату подання цієї заявки клопотаннями про надання нерегульованого статусу, що перебували на розгляді в APHIS або були задоволені APHIS, були ті, що містять таку інформацію: - Клопотання: ідентифікаційний номер клопотання. Технічні описи трансформаційних подій можна знайти в документах окремих клопотань, які можна одержати в APHIS, наприклад, на вебсайті APHIS, із посиланням на номер цього клопотання. Ці описи включені до цієї заявки шляхом посилання. - Поширення клопотання: посилання на попереднє клопотання, про поширення якого подана заява. - Організація: найменування юридичної особи, що подає клопотання. - Об'єкт регулювання: відповідний сорт рослин. - Трансгенний фенотип: характерна ознака, надана рослинам за допомогою трансформаційної події. - Трансформаційна подія або лінія: назва події або подій (іноді також позначені як лінія або лінії), щодо якої подано клопотання про надання нерегульованого статусу. - Документи APHIS: різні документи, опубліковані APHIS відносно клопотання і які можуть бути запитані в APHIS. Додаткові особливо придатні рослини, що містять одиночні трансформаційні події або комбінації трансформаційних подій, перелічені, наприклад, у базах даних різних національних і регіональних органів управління (див., наприклад, http://gmoinfo.jrc.it/gmp_browse.aspx і http://www.agbios.com/dbase.php). Особливо придатні трансгенні рослини, які можна обробляти відповідно до винаходу, є рослини, які містять трансформаційні події або комбінацію трансформаційних подій, які перелічені, наприклад, у базах даних різних національних і регіональних органів управління, у тому числі такі: подія 1143-14A (бавовна, боротьба з комахами, не депонована, описана у WO 2006/128569); подія 1143-51B (бавовна, боротьба з комахами, не депонована, описана у WO 2006/128570); подія 1445 (бавовна, толерантність до гербіцидів, не депонована, описана у US-A 2002-120964 або WO 02/034946); подія 17053 (рис, толерантність до гербіцидів, депонований як PTA-9843, описаний у WO 2010/117737); подія 17314 (рис, толерантність до гербіцидів, депонований як PTA-9844, описаний у WO 2010/117735); подія 281-24-236 (бавовна, боротьба з комахами - толерантність до гербіцидів, депонована як PTA-6233, описана у WO 2005/103266 або US-A 2005-216969); подія 3006-210-23 (бавовна, боротьба з комахами - толерантність до гербіцидів, депонована як PTA-6233, описана у US-A 2007-143876 або WO 2005/103266); подія 3272 (кукурудза, ознака якості, депонована як PTA-9972, описана у WO 2006/098952 або US-A 2006-230473); подія 40416 (кукурудза, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-11508, описана у WO 2011/075593); подія 43A47 (кукурудза, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-11509, описана у WO 2011/075595); подія 5307 (кукурудза, боротьба з комахами, депонована як ATCC PTA-9561, описана у WO 2010/077816); подія ASR-368 (польовиця, толерантність до гербіцидів, депонована як ATCC PTA-4816, описана у US-A 2006-162007 або WO 2004/053062); подія B16 (кукурудза, толерантність до гербіцидів, не депонована, описана у US-A 2003-126634); подія BPS-CV127-9 (соя, толерантність до гербіцидів, депонована як NCIMB No. 41603, описана у WO 2010/080829); подія CE43-67B (бавовна, боротьба з комахами, депонована як DSM ACC2724, описана у US-A 2009-217423 або WO2006/128573); подія CE44-69D (бавовна, боротьба з комахами, не депонована, описана у US-A 2010-0024077); подія CE44-69D (бавовна, боротьба з комахами, не депонована, описана у WO 2006/128571); подія CE46-02A (бавовна, боротьба з 19 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 комахами, не депонована, описана у WO 2006/128572); подія COT102 (бавовна, боротьба з комахами, не депонована, описана у US-A 2006-130175 або WO 2004/039986); подія COT202 (бавовна, боротьба з комахами, не депонована, описана у US-A 2007-067868 або WO 2005/054479); подія COT203 (бавовна, боротьба з комахами, не депонована, описана у WO 2005/054480); подія DAS40278 (кукурудза, толерантність до гербіцидів, депонована як ATCC PTA-10244, описана у WO 2011/022469); подія DAS-59122-7 (кукурудза, боротьба з комахами толерантність до гербіцидів, депонована як ATCC PTA 11384, описана у US-A 2006-070139); подія DAS-59132 (кукурудза, боротьба з комахами - толерантність до гербіцидів, не депонована, описана у WO 2009/100188); подія DAS68416 (соя, толерантність до гербіцидів, депонована як ATCC PTA-10442, описана у WO 2011/066384 або WO 2011/066360); подія DP-098140-6 (кукурудза, толерантність до гербіцидів, депонована як ATCC PTA-8296, описана у US-A 2009137395 або WO 2008/112019); подія DP-305423-1 (соя, ознака якості, не депонована, описана у US-A 2008-312082 або WO 2008/054747); подія DP-32138-1 (кукурудза, система схрещування, депонована як ATCC PTA-9158, described in US-A 2009-0210970 або WO 2009/103049); подія DP-356043-5 (соя, толерантність до гербіцидів, депонована як ATCC PTA-8287, описана у US-A 2010-0184079 або WO 2008/002872); подія EE-1 (баклажан, боротьба з комахами, не депонована, описана у WO 2007/091277); подія FI117 (кукурудза, толерантність до гербіцидів, депонована як ATCC 209031, описана у US-A 2006-059581 або WO 98/044140); подія GA21 (кукурудза, толерантність до гербіцидів, депонована як ATCC 209033, описана у US-A 2005086719 або WO 98/044140); подія GG25 (кукурудза, толерантність до гербіцидів, депонована як ATCC 209032, описана у US-A 2005-188434 або WO 98/044140); подія GHB119 (бавовна, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-8398, описана у WO 2008/151780); подія GHB614 (бавовна, толерантність до гербіцидів, депонована як ATCC PTA-6878, описана у US-A 2010-050282 або WO 2007/017186); подія GJ11 (кукурудза, толерантність до гербіцидів, депонована як ATCC 209030, описана у US-A 2005-188434 або WO 98/044140); подія GM RZ13 (цукровий буряк, стійкість до вірусів, депонований як NCIMB-41601, описаний у WO 2010/076212); подія H7-1 (цукровий буряк, толерантність до гербіцидів, депонований як NCIMB 41158 або NCIMB 41159, описаний як US-A 2004-172669 або WO 2004/074492); подія JOPLIN1 (пшениця, толерантність до хвороб, не депонована, описана у USA 2008-064032); подія LL27 (соя, толерантність до гербіцидів, депонована як NCIMB41658, описана у WO 2006/108674 або US-A 2008-320616); подія LL55 (соя, толерантність до гербіцидів, депонована як NCIMB 41660, описана у WO 2006/108675 або US-A 2008-196127); подія LLcotton25 (бавовна, толерантність до гербіцидів, депонована як ATCC PTA-3343, описана у WO 03/013224 або US-A 2003-097687); подія LLRICE06 (рис, толерантність до гербіцидів, депонований як ATCC-23352, описаний у US 6,468,747 або WO 00/026345); подія LLRICE601 (рис, толерантність до гербіцидів, депонований як ATCC PTA-2600, депонований як US-A 20082289060 або WO 00/026356); подія LY038 (кукурудза, ознака якості, депонована як ATCC PTA5623, описана у US-A 2007-028322 або WO 2005/061720); подія MIR162 (кукурудза, боротьба з комахами, депонована як PTA-8166, описана у US-A 2009-300784 або WO 2007/142840); подія MIR604 (кукурудза, боротьба з комахами, не депонована, описана у US-A 2008-167456 або WO 2005/103301); подія MON15985 (бавовна, боротьба з комахами, депонована як ATCC PTA-2516, описана у US-A 2004-250317 або WO 02/100163); подія MON810 (кукурудза, боротьба з комахами, не депонована, описана у US-A 2002-102582); подія MON863 (кукурудза, боротьба з комахами, депонована як ATCC PTA-2605, описана у WO 2004/011601 або US-A 2006-095986); подія MON87427 (кукурудза, контроль запилення, депонована як ATCC PTA-7899, описана у WO 2011/062904); подія MON87460 (кукурудза, толерантність до стресу, депонована як ATCC PTA-8910, описана у WO 2009/111263 або US-A 2011-0138504); подія MON87701 (соя, боротьба з комахами, депонована як ATCC PTA-8194, описана у US-A 2009-130071 або WO 2009/064652); подія MON87705 (соя, ознака якості - толерантність до гербіцидів, депонована як ATCC PTA9241, описана у US-A 2010-0080887 або WO 2010/037016); подія MON87708 (соя, толерантність до гербіцидів, депонована як ATCC PTA9670, описана у WO 2011/034704); подія MON87754 (соя, ознака якості, депонована як ATCC PTA-9385, описана у WO 2010/024976); подія MON87769 (соя, ознака якості, депонована як ATCC PTA-8911, описана у US-A 2011-0067141 або WO 2009/102873); подія MON88017 (кукурудза, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-5582, описана у US-A 2008-028482 або WO 2005/059103); подія MON88913 (бавовна, толерантність до гербіцидів, депонована як ATCC PTA-4854, описана у WO 2004/072235 або US-A 2006-059590); подія MON89034 (кукурудза, боротьба з комахами, депонована як ATCC PTA-7455, описана у WO 2007/140256 або US-A 2008-260932); подія MON89788 (соя, толерантність до гербіцидів, депонована як ATCC PTA-6708, описана у US-A 2006-282915 або WO 2006/130436); подія MS11 (олійний рапс, контроль запилення 20 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 толерантність до гербіцидів, депонований як ATCC PTA-850 або PTA-2485, описаний у WO 01/031042); подія MS8 (олійний рапс, контроль запилення - толерантність до гербіцидів, депонований як ATCC PTA-730, описаний у WO 01/041558 або US-A 2003-188347); подія NK603 (кукурудза, толерантність до гербіцидів, депонована як ATCC PTA-2478, описана у US-A 2007292854); подія PE-7 (рис, боротьба з комахами, не депонована, описана у WO 2008/114282); подія RF3 (олійний рапс, контроль запилення - толерантність до гербіцидів, депонований як ATCC PTA-730, описаний у WO 01/041558 або US-A 2003-188347); подія RT73 (олійний рапс, толерантність до гербіцидів, не депонована, описана у WO 02/036831 або US-A 2008-070260); подія T227-1 (цукровий буряк, толерантність до гербіцидів, не депонована, описана у WO 02/44407 або US-A 2009-265817); подія T25 (кукурудза, толерантність до гербіцидів, не депонована, описана у US-A 2001-029014 або WO 01/051654); подія T304-40 (бавовна, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-8171, описана у US-A 2010077501 або WO 2008/122406); подія T342-142 (бавовна, боротьба з комахами, не депонована, описана у WO 2006/128568); подія TC1507 (кукурудза, боротьба з комахами - толерантність до гербіцидів, не депонована, описана у US-A 2005-039226 або WO 2004/099447); подія VIP1034 (кукурудза, боротьба з комахами - толерантність до гербіцидів, депонована як ATCC PTA-3925, описана у WO 03/052073), подія 32316 (кукурудза, боротьба з комахами-толерантність до гербіцидів, депонована як PTA-11507, описана у WO 2011/084632), подія 4114 (кукурудза, боротьба з комахами - толерантність до гербіцидів, депонована як PTA-11506, описана у WO 2011/084621). Композиція за винаходом може також бути використана проти грибкових хвороб, схильних до розростання на або всередині деревини. Термін "деревина" означає усі типи сортів дерева та усі типи обробки цього дерева, призначеної для будівництва, наприклад, тверде дерево, дерево з високою щільністю, ламіноване дерево та клеєна фанера. Спосіб обробки деревини відповідно до винаходу здебільшого полягає у контактуванні з однією або більше сполуками за винаходом або композицією за винаходом; до цього відноситься, наприклад, безпосереднє нанесення, обприскування, занурювання, упорскування або будь-які інші підходящі способи. Серед хвороб рослин або сільськогосподарських культур, з якими можна боротися за допомогою способів за винаходом, можна зазначити такі: Хвороби борошнистої роси, такі як: Хвороби блюмерії (Blumeria), спричинені, наприклад, Blumeria graminis; Хвороби подосфери (Podosphaera), спричинені, наприклад, Podosphaera leucotricha; Хвороби сфаротеки (Sphaerotheca), спричинені, наприклад, Sphaerotheca fuliginea; Хвороби унцинули (Uncinula), спричинені, наприклад, Uncinula necator; Хвороби іржавіння, такі як: Хвороби гімноспорангію (Gymnosporangium), спричинені, наприклад, Gymnosporangium sabinae; Хвороби гемілеї (Hemileia), спричинені, наприклад, Hemileia vastaтриx; Хвороби факопсори (Phakopsora), спричинені, наприклад, Phakopsora pachyrhizi або Phakopsora meibomiae; Хвороби пуцинії (Puccinia), спричинені, наприклад, Puccinia recondite, Puccinia graminis або Puccinia striiformis; Хвороби уроміцесу (Uromyces), спричинені, наприклад, Uromyces appendiculatus; Хвороби ооміцету (Oomycete), такі як: Хвороби альбуго (Albugo), спричинені, наприклад, Albugo candida; Хвороби бремії (Bremia), спричинені, наприклад, Bremia lactucae; Хвороби пероноспори (Peronospora), спричинені, наприклад, Peronospora pisi або P. brassicae; Хвороби фітофтори (Phytophthora), спричинені, наприклад, Phytophthora infestans; Хвороби плазмопари (Plasmopara), спричинені, наприклад, Plasmopara viticola; Хвороби псевдопероноспори (Pseudoperonospora), спричинені, наприклад, Pseudoperonospora humuli або Pseudoperonospora cubensis; Хвороби пітію (Pythium), спричинені, наприклад, Pythium ultimum; Хвороби плямистості листя: плями, прищі, тля, такі як: Хвороби альтернарії (Alternaria), спричинені, наприклад, Alternaria solani; Хвороби церкоспори (Cercospora), спричинені, наприклад, Cercospora beticola; Хвороби кладіоспоруму (Cladiosporum), спричинені, наприклад, Cladiosporium cucumerinum; Хвороби кохліоболусу (Cochliobolus), спричинені, наприклад, Cochliobolus sativus (форма конідій: Дрехслера (Drechslera), синонім: гельмінтоспоріум (Helminthosporium) або Cochliobolus miyabeanus; 21 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 Хвороби колетотрихуму (Colletotrichum), спричинені, наприклад, Colletotrichum lindemuthanium; Хвороби циклоконію (Cycloconium), спричинені, наприклад, Cycloconium oleaginum; Хвороби діапорте (Diaporthe), спричинені, наприклад, Diaporthe citri; Хвороби ельсине (Elsinoe), спричинені, наприклад, Elsinoe fawcettii; Хвороби глеоспорію (Gloeosporium), спричинені, наприклад, Gloeosporium laeticolor; Хвороби гломерели (Glomerella), спричинені, наприклад, Glomerella cingulata; Хвороби гігнардії (Guignardia), спричинені, наприклад, Guignardia bidwelli; Хвороби лептосферії (Leptosphaeria), спричинені, наприклад, Leptosphaeria maculans; Leptosphaeria nodorum; Хвороби магнапорте (Magnaporthe), спричинені, наприклад, Magnaporthe grisea; Хвороби мікосфарели (Mycosphaerella), спричинені, наприклад, Mycosphaerella graminicola; Mycosphaerella arachidicola; Mycosphaerella fijiensis; Хвороби фасфарії (Phaeosphaeria), спричинені, наприклад, Phaeosphaeria nodorum; Хвороби піренофори (Pyrenophora), спричинені, наприклад, Pyrenophora teres або Pyrenophora tritici repentis; Хвороби рамуларії (Ramularia), спричинені, наприклад, Ramularia collo-cygni або Ramularia areola; Хвороби ринхоспорію (Rhynchosporium), спричинені, наприклад, Rhynchosporium secalis; Хвороби септорії (Septoria), спричинені, наприклад, Septoria apii або Septoria lycopercisi; Хвороби тифули (Typhula), спричинені, наприклад, Typhula incarnata; Хвороби вентурії (Venturia), спричинені, наприклад, Venturia inaequalis; Хвороби коріння та стебла, такі як: Хвороби кортицію (Corticium), спричинені, наприклад, Corticium graminarum; Хвороби фузарію (Fusarium), спричинені, наприклад, Fusarium oxysporum; Хвороби геуманоміцію (Gaeumannomyces), спричинені, наприклад, Gaeumannomyces graminis; Хвороби ризоктонії (Rhizoctonia), спричинені, наприклад, Rhizoctonia solani; Хвороби сарокладію (Sarocladium), спричинені, наприклад, Sarocladium oryzae; Хвороби склеротію (Sclerotium), спричинені, наприклад, Sclerotium oryzae; Хвороби тапесії (Tapesia), спричинені, наприклад, Tapesia acuformis; Хвороби тилавіопсису (Thielaviopsis), спричинені, наприклад, Thielaviopsis basicola; Хвороби колосся та суцвіття у вигляді мітли, такі як: Хвороби альтернарії (Alternaria), спричинені, наприклад, видом Alternaria; Хвороби аспергілусу (Aspergillus), спричинені, наприклад, Aspergillus flavus; Хвороби кладоспорію (Cladosporium), спричинені, наприклад, видом Cladosporium; Хвороби клавісепсу (Claviceps), спричинені, наприклад, Claviceps purpurea; Хвороби фузарію (Fusarium), спричинені, наприклад, Fusarium culmorum; Хвороби гіберели (Gibberella), спричинені, наприклад, Gibberella zeae; Хвороби монографели (Monographella), спричинені, наприклад, Monographella nivalis; Хвороби головні та мокрої головні, такі як: Хвороби сфацелотеки (Sphacelotheca), спричинені, наприклад, Sphacelotheca reiliana; Хвороби тилетії (Tilletia), спричинені, наприклад, Tilletia caries; Хвороби уроцистису (Urocystis), спричинені, наприклад, Urocystis occulta; Хвороби устилаго (Ustilago), спричинені, наприклад, Ustilago nuda; Хвороби гниття та плісняви фруктів, такі як: Хвороби аспергілусу (Aspergillus), спричинені, наприклад, Aspergillus flavus; Хвороби ботритису (Botrytis), спричинені, наприклад, Botrytis cinerea; Хвороби пеніцилію (Penicillium), спричинені, наприклад, Penicillium expansum; Хвороби ризофусу (Rhizopus), спричинені, наприклад, Rhizopus stolonifer; Хвороби склеротинії (Sclerotinia), спричинені, наприклад, Sclerotinia sclerotiorum; Хвороби вертицилію (Verticilium), спричинені, наприклад, Verticilium alboatrum; Хвороби гниття, плісняви, зів'янення, гнилизни і випрівання (чорної ніжки) насіння та ґрунту, такі як: Хвороби альтернарії (Alternaria), спричинені, наприклад, Alternaria brassicicola; Хвороби афаноміцесу (Aphanomyces), спричинені, наприклад, Aphanomyces euteiches; Хвороби аскохіта (Ascochyta), спричинені, наприклад, Ascochyta lentis; Хвороби аспергілусу (Aspergillus), спричинені, наприклад, Aspergillus flavus; Хвороби кладоспорію (Cladosporium), спричинені, наприклад, Cladosporium herbarum; Хвороби кохліоболусу (Cochliobolus), спричинені, наприклад, Cochliobolus sativus (форма 22 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 конідій: Drechslera, Bipolaris, синонім: Helminthosporium); Хвороби колетотрихуму (Colletotrichum), спричинені, наприклад, Colletoтриchum coccodes; Хвороби фузарію (Fusarium), спричинені, наприклад, Fusarium culmorum; Хвороби гіберели (Gibberella), спричинені, наприклад, Gibberella zeae; Хвороби макрофоміни (Macrophomina), спричинені, наприклад Macrophomina phaseolina; Хвороби монографели (Monographella), спричинені, наприклад Monographella nivalis; Хвороби пеніцилію (Penicillium), спричинені, наприклад, Penicillium expansum; Хвороби фома (Phoma), спричинені, наприклад, Phoma lingam; Хвороби фомопсису (Phomopsis), спричинені, наприклад, Phomopsis sojae; Хвороби фітофтори (Phytophthora), спричинені, наприклад, Phytophthora cactorum; Хвороби піренофори (Pyrenophora), спричинені, наприклад, Pyrenophora graminea; Хвороби пірикуларію (Pyricularia), спричинені, наприклад, Pyricularia oryzae; Хвороби пітію (Pythium), спричинені, наприклад, Pythium ultimum; Хвороби ризоктонії (Rhizoctonia), спричинені, наприклад, Rhizoctonia solani; Хвороби ризопусу (Rhizopus), спричинені, наприклад Rhizopus oryzae; Хвороби склероцію (Sclerotium), спричинені, наприклад, Sclerotium rolfsii; Хвороби септорії (Septoria), спричинені, наприклад, Septoria nodorum; Хвороби тифули (Typhula), спричинені, наприклад, Typhula incarnata; Хвороби вертицилію (Verticillium), спричинені, наприклад, Verticillium dahliae; Хвороби у вигляді червоточини, засихання вершин та мітел, такі як: Хвороби нектрії (Nectria), спричинені, наприклад, Nectria galligena; Хвороби, які характеризуються зів'яненням, гниттям або припиненням росту, такі як: Хвороби моніліозу (Monilinia), спричинені, наприклад, Monilinia laxa; Хвороби пухирчастості або закручування листя, такі як: Хвороби екзобазидію (Exobasidium), спричинені, наприклад, Exobasidium vexans; Хвороби тафрини (Taphrina), спричинені, наприклад, Taphrina deformans; Хвороби погіршення стану дерев'янистих рослин, такі як: Хвороби еска (Esca), спричинені, наприклад, Phaemoniella clamydospora; Хвороби еутипа дайбек (Eutypa dyeback), спричинені, наприклад, Eutypa lata; Хвороби ганодерма (Ganoderma), спричинені, наприклад, Ganoderma boninense; Хвороби ригідопорусу (Rigidoporus), спричинені, наприклад, Rigidoporus lignosus; Хвороби квітів і насіння, такі як: Хвороби ботритису (Botrytis), спричинені, наприклад, Botrytis cinerea; Хвороби бульби, такі як: Хвороби ризоктонію (Rhizoctonia), спричинені, наприклад, Rhizoctonia solani; Хвороби гельмінтоспорію (Helminthosporium), спричинені, наприклад, Helminthosporium solani; Хвороби кіли хрестоцвітих, такі як: Хвороби плазмодіофори (Plasmodiophora), спричинені, наприклад, Plamodiophora brassicae; Хвороби, спричинені бактерійними організмами, такими як: Вид ксантоманас (Xanthomanas), наприклад, Xanthomonas campestris pv. oryzae; Вид псевдомонас (Pseudomonas), наприклад, Pseudomonas syringae pv. lachrymans; Вид ервінія (Erwinia), наприклад, Erwinia amylovora. Умовні позначення на кресленнях: Фіг. 1: Вимірювання інгібування росту Phytophthora infestans у присутності длРНК, мішенню якої є матрична інформаційна ДНК сахаропін дегідрогенази. Фіг. 2: Аналіз рівня мРНК сахаропін дегідрогенази за допомогою кількісної ПЛР зі зворотною транскрипцією. Перелік послідовностей: SEQ ID N°1: Сахаропін дегідрогеназа (LYS1) з Aspergillus clavatus. SEQ ID N°2: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°3: Сахаропін дегідрогеназа (LYS1) з Aspergillus fumigatus. SEQ ID N°4: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°5: Сахаропін дегідрогеназа (LYS1) з Botrytis cinerea. SEQ ID N°6: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°7: Сахаропін дегідрогеназа (LYS1) з Fusarium graminearum. SEQ ID N°8: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°9: Сахаропін дегідрогеназа (LYS1) з Fusarium oxysporum. SEQ ID N°10: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°11: Сахаропін дегідрогеназа (LYS1) з Fusarium verticilloides. 23 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 SEQ ID N°12: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°13: Сахаропін дегідрогеназа (LYS1) з Fusarium verticilloides. SEQ ID N°14: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°15: Сахаропін дегідрогеназа (LYS1) з Mycosphaerella fijiensis. SEQ ID N°16: Поліпептид, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°17: Сахаропін дегідрогеназа (LYS1) з Magnaporthe grisea. SEQ ID N°18: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°19: Сахаропін дегідрогеназа (LYS1) з Monoliophthora perniciosa. SEQ ID N°20: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°21: Сахаропін дегідрогеназа (LYS1) з Puccinia graminis. SEQ ID N°22: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°23: Сахаропін дегідрогеназа (LYS1) з Phytophthora infestans. SEQ ID N°24: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°25: Сахаропін дегідрогеназа (LYS1) з Phytophthora ramorum. SEQ ID N°26: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°27: Сахаропін дегідрогеназа (LYS1) з Phytophthora sojae. SEQ ID N°28: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°29: Сахаропін дегідрогеназа (LYS1) з Pyrenophora tritici-repentis. SEQ ID N°30: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°31: Сахаропін дегідрогеназа (LYS1) з Sclerotinia sclerotiorum. SEQ ID N°32: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°33: Сахаропін дегідрогеназа (LYS1) з Trichoderma reesei. SEQ ID N°34: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°35: Сахаропін дегідрогеназа (LYS1) з Ustilago maydis. SEQ ID N°36: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°37: Сахаропін дегідрогеназа (LYS1) з Verticillium albo-atrum. SEQ ID N°38: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°39: Сахаропін дегідрогеназа (LYS1) з Mycosphaerella graminicola. SEQ ID N°40: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°41: Сахаропін дегідрогеназа (LYS1) з Fusarium moniliform. SEQ ID N°42: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°43: Сахаропін дегідрогеназа (LYS1) з Claviceps purpurea. SEQ ID N°44: Білок, кодований вищевказаною послідовністю нуклеїнової кислоти. SEQ ID N°45: Праймер SACdh_Pi_T7_F. SEQ ID N°46: Праймер SACdh_Pi_T7_R. SEQ ID N°47: Праймер актину прямий. SEQ ID N°48: Праймер актину зворотний. SEQ ID N°49: Праймер β-тубулін прямий. SEQ ID N°50: Праймер β-тубулін зворотний. SEQ ID N°51: Праймер SACdh прямий. SEQ ID N°52: Праймер SACdh зворотний. SEQ ID N°53: Праймер pBINB33-1. SEQ ID N°54: Праймер pBINB33-2. SEQ ID N°55: Праймер SacdhPI R. SEQ ID N°56: Праймер SacdhPI F. SEQ ID N°57: Праймер LYS1 Pot 117-F. SEQ ID N°58: Праймер LYS1 Pot 117-R. Різні аспекти винаходу будуть зрозумілими більш повно за допомогою наведених нижче експериментальних прикладів. Всі способи або операції, описані нижче, наведені як приклад та відповідають вибору, зробленому з-поміж різних методів, доступних для досягнення того самого результату. Вибір не має жодного впливу на якість результату, і, як наслідок, будь-який підходящий метод може бути використаний спеціалістами у цій галузі для досягнення того самого результату. Зокрема, і якщо не вказано інакше в прикладах, всі застосовані технології рекомбінантної ДНК проводять відповідно до стандартних протоколів, описаних у Sambrook and Russel (2001, Molecular cloning: A laboratory manual ("Молекулярне клонування: лабораторний посібник), третє видання, Cold Spring Harbor Laboratory Press, Нью-Йорк), у Ausubel at al. (1994, Current Protocols in Molecular Biology, Current protocols ("Сучасні протоколи в молекулярній біології"), США, томи 1 і 2), та у Brown (1998, Molecular Biology LabFax ("Молекулярна біологія LabFax") LabFax, друге видання, Academic Press, Велика Британія). Стандартні матеріали та методи молекулярної біології 24 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 рослин описані у Croy R.D.D. (1993, Plant Molecular Biology LabFax ("Молекулярна біологія рослин LabFax"), BIOS Scientific Publications Ltd (Велика Британія) та Blackwell Scientific Publications (UK)). Стандартні матеріали та способи ПЛР (полімеразної ланцюгової реакції) також описані у Dieffenbach and Dveksler (1995, PCR Primer: A laboratory manual ("Праймер ПЛР: Лабораторний посібник"), Cold Spring Harbor Laboratory Press, Нью-Йорк) та у McPherson et al. (2000, PCR-Basics: From background to bench, перше видання, Springer Verlag, Німеччина). Довідкова бібліографія: - Bevan, 1984, Nucl Acids Res 12: 8711-8721; - Bhattacharjee, J. K., 1985, Crit. Rev.Microbiol. 12, 131-151; - Bhattacharjee, J. K., 1992, у Handbook of Evolution of Metabolic Function (Підручник з еволюції метаболічної функції), Mortlock, R. P., ed., CRC Press, Boca Raton, FL, стор. 47-80; - Born T.L. і Blanchard J.S., 1999, Curr. Opin. Chem. Biol., 3:607-613; - Broquist, H. P., 1971, Methods Enzymol. 17, 112-129; - Chuang and Meyerowitz, 2000, PNAS, 97: 4985-4990; - Ehmann at al., 1999, biochemistry, 38,6171-6177; - Escobar et al., 2001, Proc. Natl. Acad. Sci. USA., 98(23): 13437-13442; - Fire at al., 1998, Nature 391: 806-811; - Garrad, R. C. and Bhattacharjee, J. K., 1992, J. Bacteriol. 174, 7379-7384; - Gielen at al., 1984, EMBO Journal 3, 835-846; - Hamilton and Baulcome, 1999, Science 286: 950-952; - Hamilton et al., 1998, Plant J, 15: 737746; - Hammond at al., 2000, Nature 404: 293-296; - Hannon, 2002, Nature, 418 (6894): 244-51; Herrera-Estrella at al., 1983 Nature, 303, 209-213; - Hofgen and Willmitzer, 1990, Plant Science 66, 221-230; - Houmard at al., 2007, Plant Biotechnology Journal 5, 605-614; - Montgomery at al., 1998, PNAS 95: 15502-15507; - Pandolfini at al., 2003, Biotechnol., 25; 3(1): 7; - Pietrzak at al., 1986 Nucleic Acids Res. 14, 5858; - Thompson at al., Nucleic Acids Research 22 (1994), 4673-4680; - Umbargar, H. E., 1978, Annu. Rev. Biochem. 47, 533-606; - Randall, T.A., 2005, MPMI, 18, 229-243; - Rocha-Sosa at al., 1989, EMBO J. 8, 23-29; - Ruiz Ferrer and Voinnet, Annu. Rev. Plant Biol. 2009. 60:485-510; - Sambrook at al., 2001, Molecular Cloning: A Laboratory Manual ("Молекулятрне клонування: лабораторний посібник"), третє видання, ISBN978-087969577-4; - Silué at al., Physiol. Mol. Plant Pathol., 53 239-251, 1998; - Stärkel C., кандидатська дисертація "Host induced gene silencing - strategies for the improvement of resistance against Cercospora beticola in sugar beet (B. vulgaris L.) and against Fusarium graminearum in corn (T. aestivum L.) and maize (Z. mays L.)", ("Сайленсинг генів, індукований хазяїном - стратегії покращення стійкості проти Cercospora beticola в цукровому буряку (B. vulgaris L.) та проти Fusarium graminearum в пшениці (T. aestivum L.) та маїсі (Z. mays L.)"), захищена в червні 2011 р., XP002668931, одержана в Інтернеті (2012-02-07) за адресою http://ediss.sub.uni-hamburg.de/vollteste/2011/5286/pdf/Dissertation.pdf; - Tang at al., 2003 Gene Dev., 17(1): 49-63; - Villalba at al., Fungal Genetics and Biology ("Грибкова генетика та біологія"), 45, 68-75, 2008; - Vogel, H. J., 1965, у Handbook of Evolving Genes and Proteins ("Підручник з генів та білків, що еволюціонують", Bryson, V видання, Academic Press, Нью-Йорк, стор. 25-40; - Waterhouse at al., 1998, PNAS 95: 13959-13964; - Wesley at al., The Plant Journal ("Журнал з рослин") (2001); 27(6), 581-590; - Xiao at al., 2003, Plant Mol Biol., 52(5): 957-66; - Xu at al., 2006, Cell Bio chemistry and Biophysics ("Клітинна біохімія та біофізика"), том. 46, 43-64; - Zamore at al., 2000, Cell ("Клітина"), 101: 25-33. Приклади Приклад 1: Розведення в умовах іn vitro Magnaporthe Grisea Були проведені аналізи із застосуванням штамів Magnaporthe grisea дикого типу P1.2, одержаного з колекції фітопатологічної лабораторії CIRAD (Centre de coopération internationale en recherche agronomique pour le développement - Сільськогосподарського дослідницького центру з міжнародного розвитку) в Монпельє. Умови вирощування, склад середовища рис-агар, утримання та спороутворення, а також препарати протопластів описані Silué та ін. (1998). Приклад 2: Трансфікування Magnaporthe grisea длРНК, мішенню якої є сахаропін дегідрогеназа, та вимірювання інгібування росту Послідовність генів сахаропін дегідрогенази (SACdh) Magnaporthe grisea Lys-1 (MGG_01359.6: 1426 bp) одержали в інституті Броад (Broad Institute) (http://www.broadinstitute.org/). Вибрали ділянку з близько 325 бп для длРНК, що містила 25 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 нуклеотиди від 301 до 626, компанія Geneart її синтезувала та клонувала у плазміду. Для трансфікування одержали длРНК сахаропін дегідрогенази з Magnaporthe grisea із використанням набору РНКі MEGAscript (Ambion) згідно з інструкцією виробника. Різні кількості (200 мкг - 2 мг дл РНК) обробили трансфекційним агентом Ліпофектамін РНКі макс (Invitrogen) 6 згідно з інструкцією виробника. Додали комплекси Ліпофектамін-дл РНК до 2,5 × 10 протопластів Magnaporthe grisea у мікротитраційному планшеті із середовищем TB3 (Villalba at al., 2008), і спостерігали за ростом впродовж 5-7 днів при ОЩ 600 нм із використанням зчитувача мікропланшетів Infinite M1000 (Tecan). За ростом протопластів Magnaporthe grisea, оброблених длРНК сахаропін дегідрогенази, порівняно з необробленими контрольними зразками спостерігали у декількох моментах часу, і це спостереження показало значну відмінність у рості. Приклад 3: Вирощування в умовах іn vitro Phytophthora infestans та одержання зооспор Штам PT78 Phytophthora infestans вирощували в умовах in vitro у 9 см чашках Петрі в середовищі горох-агар (125 г/л вареного та м'ятого гороху, 20 г/л агар-агару, карбеніцилін 100 мг/л) при температурі 21 °C в темряві. Кожні 15 днів інокулювали нове середовище чотирма 5 мм кубічними заглушками міцелію. Для вивільнення зооспор 12 мл крижаної води вилили на культуру віком 10 днів, і культуру помістили при температурі 4 °C на 2 години. Потім супернатант зібрали не порушуючи цільності міцелію та відфільтрували через 100 мкм сито для видалення гіфових фрагментів. Зооспори виклали на лід та підрахували за допомогою рахувальної камери. Приклад 4: Трансфікування длРНК Phytophthora infestans, мішенню якої є сахаропін дегідрогеназа, та вимірювання інгібування росту Послідовність гену Lys-1 сахаропін дегідрогенази Phytophthora infestans (PITG_03530: 3020 бп) одержали з бази даних P. infestans інституту Броад. Ділянку близько 500 бп з кращими міРНК відповідно до програмного статку РНКі-конструктора BLOCKit (Invitrogen) та із вмістом нуклеотидів від 2251 до 2750 синтезувала компанія Geneart та клонувала в плазміду 0920357_SacDH_Pi_pMA. Синтез длРНК проводили із використанням набору РНКі Megascript (Ambion) відповідно до протоколу виробника та із використанням як шаблону продукту ПЛР, збільшеного з плазміди 0920357_SacDH_Pi_pMA. Прямим праймером, що його використовували, був SACdh_Pi_T7_F: 5" TAATACGACTCACTATAGGGTTGCAGGAGAGCGCAGAAAGC, а зворотним праймером був SACdh_Pi_T7_R: TAATACGACTCACTATAGGGTCAGTTGGAGTCCGCGTGGTGT. Потім длРНК осадили за допомогою 100 %-ного етанолу та ацетату натрію 3M, pH 5.2, двічі промили 70 %-ним етанолом, і гранули повторно суспендували у вільній від рибонуклеаз воді. Трансфекційні суміші приготували у 48-лунковому планшеті додаванням послідовно середовища V8 (5 % соку V8 (Campbell Foods Belgium), pH5), відповідна кількість длРНК та 10 мкл ліпофектаміну РНКі макс (Invitrogen) у кінцевому об'ємі 200 мкл. Трансфекційну суміш інкубували впродовж 15 хвилин при кімнатній температурі. 4 Зооспори розбавили у середовищі V8 до концентрації 5 × 10 зооспор/мл. Потім у кожну лунку планшета додали 800 мкл розчину зооспор. Кінцева концентрація зооспор становила 4 × 5 10 зооспор/мл. У кожний планшет додали три контрольні зразки: середовище V8, середовище V8 + зооспори, середовище V8 + зооспори + ліпофектамін. Планшети інкубували при температурі 21 °C у темряві. За ростом грибів спостерігали шляхом вимірювання поглинання при 620 нм у планшетному зчитувачі (Infinite 1000, Tecan) через 8 днів. Процентний вміст інгібування росту обчислювали із використанням такої формули: 100-(ОЩдлРНК x 100 / ОЩконтрольний ліпофектамін). Ріст грибів знизився у присутності длРНК, спрямованого проти сахаропін дегідрогенази, залежно від концентрації (100 нмоль та 200 нмоль відповідно), як показано на Фігурі 1. Приклад 5: Аналіз кількісної ПЛР матричної інформаційної РНК сахаропін дегідрогенази P. Infestans Для одержання достатньої кількості РНК для синтезу кДНК та ПЛР зі зворотною транскрипцією у режимі реального часу декілька лунок 48-лункового планшета звели для однієї концентрації випробуваної длРНК: 10 лунок для 72 год. моменту часу, 6 лунок для 96 год. моменту часу, 3 лунки для 120 год. моменту часу. Через 72 год., 96 год. і 120 год. обробки длРНК зібрали міцелій. Зразки центрифугували для видалення середовища. Зразки заморозили в рідкому азоті, а потім ліофілізували впродовж ночі. Перед екстрагуванням РНК міцелій розмололи. Всю РНК екстрагували із використанням 26 UA 115132 C2 5 10 15 міні-набору RNeasy Plant (Qiagen) згідно з протоколом виробника. Забруднення ДНК зразків РНК видалили перетравлюванням дезоксирибонуклеази (без ДНК, Ambion). Цільність РНК аналізували на Біоаналізаторі 2100 (нано-набір РНК 6000, Agilent) згідно з наданим виробником протоколом. кДНК синтезували з 2 мкг загальної РНК оліго дТ праймінгом із використанням набору Термоскрипт системи ПЛР зі зворотною транскрипцією (Invitrogen) згідно з протоколом виробника. кДНК осадили 100 %-ним EtOH та ацетатом натрію 3M, pH 5.2, двічі промили 70 %ним EtOH, і кульки повторно суспендували у 10 мкл вільної від рибонуклеази води. кДНК розбавили 100 разів для аналізу кількісної ПЛР. Пари праймерів призначили для кожної генної послідовності із застосуванням програмного забезпечення "Primer Express 3" (Applied Biosystems). ПЛР зі зворотною транскрипцією у режимі реального часу проводили в системі ПЛР в режимі реального часу 7900 (Applied Biosystems) з зеленим мастер-розчином ПЛР Power SYBR (Applied Biosystems) згідно з протоколом виробника. К-ПЛР проводили таким чином: при температурі 95 °C впродовж 10 хв., 45 циклів при температурі 95 °C впродовж 15 сек. і при 60 °C 1 хв. з наступною стадією дисоціації при температурі 95 °C впродовж 15 сек., при 60 °C впродовж 1 хв. і при 95 °C впродовж 15 сек. Гени актину і β-тубуліну використовували як ендогенний контроль. Відносну експресію генів обчислювали методом 2ΔΔCt. На фігурі 2 показане значне зниження рівня матричної інформаційної РНК сахаропін дегідрогенази, і це зниження корелює з інгібуванням росту. Таблиця 1 Послідовність праймерів кПЛР: Назва праймера актин β-Tub SACdh Пряма праймерна послідовність Зворотна праймерна послідовність CGACTCTGGTGACGGTGTGT GCGTGAGGAAGAGCGTAACC CCGCCCAGACAATTTCGT CCTTGGCCCAGTTGTTACCA TGGGTGGTTTCCAAGGTCTTC AAAGGCACCAAGCCACTGAA 20 25 30 35 40 45 50 Приклад 6: Конструювання трансформаційних векторів із вмістом гену сахаропін дегідрогенази Phytophthora infestans a) Одержання експресійного вектору рослини IR 47-71 Плазміда pBinAR є похідною бінарної векторної плазміди pBin19 (Bevan, 1984), яка сконструйована таким чином: фрагмент довжиною 529 бп, складений з нуклеотидів 6909-7437 промотора 35S мозаїчного вірусу цвітної капусти, ізолювали як фрагмент EcoR \IKpn I з плазміди pDH51 (Pietrzak et al, 1986) та лігували між сайтами рестрикції EcoR I та Kpn I полілінкера pUC18. Таким чином була сформована плазміда pUC18-35S. Із використанням ендонуклеаз рестрикції Hind III та Pvu II, фрагмент довжиною 192 бп, що містив сигнал ' поліаденілування (3 -кінець) гену октопінсинтази (ген 3) Т-ДНК Ti плазміди pTiACHδ (Gielen et al, 1984) (нуклеотиди 11 749-11 939), ізолювали з плазміди pAGV40 (Herrera-Estrella et al., 1983). Після додавання лінкерів Sph I до сайту рестрикції Pvu Il цей фрагмент лігували між сайтами рестрикції Sph I та Hind III pUC18-35S. Це дало плазміду pA7. Тут був вилучений весь полілінкер, що містив промотор 35S та Ocs-кінець, із використанням EcoR I та Hind III та лігований у належним чином розщеплений вектор pBin19. Це дало експресійний вектор рослини pBinAR (Hofgen and Willmitzer, 1990). Промотор гену пататину B33 Solanum tuberosum (Rocha-Sosa et al., 1989), як фрагмент Dra I (нуклеотиди -1512 - +14), лігували у Ssf l-розщеплений вектор pUC19, кінці якого затупили із використанням T4-ДНК полімерази. Це дало плазміду pUC19-B33. З цієї плазміди промотор B33 видалили із використанням EcoR I та Sma I та лігували у належним чином обмежений вектор pBinAR. Це дало експресійний вектор рослини pBinB33. Для полегшення подальших стадій клонування розширили СМК (сайт множинного клонування). Для цього синтезували два комплементарні олігонуклеотиди, нагріли до 95 °C впродовж 5 хвилин, повільно охолодили до кімнатної температури для досягнення доброї фіксації (гартування) та клонували у сайти рестрикції Sal I та Kpn I pBinB33. Олігонуклеотиди, що їх використовували з цією метою, мали таку послідовність: pBINB33-1: 5'-TCG ACA GGC CTG GAT CCT TAA TTA AAC TAG TCT CGA GGA GCT CGG TAC-3" ’ pBINB33-2: 5'-CGA GCT CCT CGA GAC TAG TTT AAT TAA GGA TCC AGG CCT G-3 Одержану плазміду назвали IR 47-71. b) Одержання експресійних векторів рослини pEPA248 та pEPA262, що містять послідовності нуклеїнової кислоти для гену сахаропін дегідрогенази Phytophthora infestans. 27 UA 115132 C2 5 10 15 20 25 30 35 40 45 50 55 60 Послідовність сахаропін дегідрогенази (PITG_03530: 3020 бп) отримали з бази даних P. infestans ORF Prot V1. Ділянку близько 500 бп, що мала кращу міРНК відповідно до конструкторського програмного статку РНКі BLOCKit (Invitrogen), синтезувала компанія Geneart. Фрагмент 300 бп посилили ПЛР з цієї послідовності ДНК праймерами SacdhPI R (5’agaggtaccaagcttgcgtagctgg-3") та SacdhPI F (5’-tatctcgagtctagacaacgccattggttac-3"). Посилений фрагмент клонували у pCRII-Topo (Invitrogen) для одержання плазміди pEPA250. Відповідно до Wesley et al. (2001) послідовність, що становила інтерес, клонували у вектор pHannibal для одержання плазміди pEPA241. Потім експресійну касету длРНК суб-клонували в різні бінарні (експресійні рослинні) вектори pART27 (Gleave AP, PMB 20, (1992), 1203-1207) та IR 47 для одержання експресійних векторів рослин pEPA248 та pEPA262 відповідно. Вектори pEPA248 та pEPA262 ввели відповідно у GV3101 та C58C1RIF (pGV2260) клітини агробактерій електропорацією (Rocha-Sosa et al.(1989)) для подальшої трансформації рослин картоплі. Приклад 7: Конструювання трансформаційних векторів, мішенню яких є ген сахаропін дегідрогенази lys1 Sclerotinia sclerotiorum 351 бп ділянки кодувальної послідовності S. sclerotiorum Lys1 (сахаропін дегідрогеназа SS1G_06166.1) синтезувала компанія Geneart (pEPA293) з розташуванням по боках внутрішніх (XbaI, HindIII) та зовнішніх (XhoI, KpnI) сайтів рестрикції, які були сконструйовані для здійснення двостадійного клонування у векторі pHannibal (Wesley et al., 2001). Проміжна плазміда приховувала дві інвертовані копії фрагменту гену Lys1, відокремлені нітроном pHannibal PdK та регульовані промотором (CaMV) 35S мозаїчного вірусу цвітної капусти та OCS-кінцем. Всю касету ДНК потім вирізали за допомогою NotI та вставили у бінарний вектор pART27 (Gleave, 1992), даючи кінцевій плазміді pEPA307 касету селекції рослин на основі стійкості до канаміцину (ген nptII, регульований промотором і кінцем Nos). Таку саму касету NotI також вставили у бінарний вектор (pFCO31) з маркером селекції рослин на основі стійкості до інгібіторів ГФПД для використання у трансформації сої. Кінцева плазміда потім може трансформувати рослини за допомогою T-ДНК, що містить між правим та лівим кордоном нашу касету, що становить інтерес, та ген ГФПД, що його регулює промотор CsVMV, хлоропластну транзитну пептидну послідовність та 3’Nos кінець. Приклад 8: Конструювання трансформаційних векторів, мішенню яких є ген сахаропін дегідрогенази lys1 Phakopsora pachirizi 364 бп ділянки Phakopsora pachirizi Lys1 E.S.T. (сахаропін дегідрогеназа PHAPC_EH247326.1) синтезувала компанія Geneart (pCED42)) з розташуванням по боках внутрішніх (XbaI, HindIII) та зовнішніх (XhoI, KpnI) сайтів рестрикції, які були сконструйовані для здійснення двостадійного клонування у векторі pHannibal (Wesley et al., 2001). Проміжна плазміда приховувала дві інвертовані копії фрагменту гену Lys1, відокремлені нітроном pHannibal PdK та регульовані промотором (CaMV) 35S мозаїчного вірусу цвітної капусти та OCS-кінцем. Всю касету ДНК потім вирізали за допомогою NotI та вставили у бінарний вектор pART27 (Gleave, 1992), даючи кінцевій плазміді pCED45 касету селекції рослин на основі стійкості до канаміцину (ген nptII, регульований промотором і кінцем Nos). Таку саму касету NotI також вставили у бінарний вектор (pFCO31) з маркером селекції рослин на основі стійкості до інгібіторів ГФПД для використання у трансформації сої. Кінцева плазміда (pCED87) потім може трансформувати рослини за допомогою T-ДНК, що містить між правим та лівим кордоном нашу касету, що становить інтерес, та ген ГФПД, що його регулює промотор CsVMV, хлоропластну транзитну пептидну послідовність та 3’Nos кінець. Приклад 9: Трансформування рослин картоплі векторами експресії рослин, що містять молекули нуклеїнової кислоти, що кодують для шпилькової конструкції сахаропін дегідрогенази pEPA262 Рослини картоплі трансформували за допомогою агробактерії із використанням експресійного вектору рослини pEPA262, що містить кодувальну послідовність нуклеїнової кислоти сахаропін дегідрогенази під контролем промотора гену пататину B33 від Solanυm tuberosum, як описано Rocha-Sosa et al. (1989). Трансгенні рослини картоплі, трансформовані плазмідою pEPA262, назвали "537 ES". Молекулярний аналіз подій "537 ES" проводили із використанням стандартних методів ПЛР (Sambrook et al.) для виявлення присутності послідовності нуклеїнової кислоти сахаропін дегідрогенази із використанням таких праймерів SacDH PI F: 5'TATCTCGAGTCTAGACAACGCCATTGGTTAC-3' та SacDH PI R: 5'AGAGGTACCAAGCTTGCGTAGCTGG-3'. Подальший вибір проводили або Нозерн-блоттингом, або аналізом експресії послідовності нуклеїнової кислоти сахаропін дегідрогенази за допомогою 28
ДивитисяДодаткова інформація
Назва патенту англійськоюRnai for the control of fungi and oomycetes by inhibiting saccharopine dehydrogenase gene
Автори англійськоюDelebarre, Thomas, Dorme, Cecile, Essigmann, Bernd, Schmitt, Frederic, Villalba, Francois, Paget, Eric
Автори російськоюДэлэбаррэ Томас, Дормэ Сэсиль, Эссигманн Бэрнд, Шмитт Фрэдэрик, Вилалба Француа, Пажэ Эрик
МПК / Мітки
МПК: C12N 15/113, C12N 15/82, A01H 5/00
Мітки: гена, ооміцетами, інгібування, грибами, сахаропіндегідрогенази, спосіб, боротьби, шляхом
Код посилання
<a href="https://ua.patents.su/118-115132-sposib-borotbi-z-gribami-ta-oomicetami-shlyakhom-ingibuvannya-gena-sakharopindegidrogenazi.html" target="_blank" rel="follow" title="База патентів України">Спосіб боротьби з грибами та ооміцетами шляхом інгібування гена сахаропіндегідрогенази</a>
Похідні тетразолу, фунгіцидна композиція на їх основі, застосування їх для радикальної або запобіжної боротьби із фітопатогенними грибами рослин або злакових культур та спосіб боротьби із фітопатогенними грибами

Номер патенту: 94468
Опубліковано: 10.05.2011
Автори: Рінолфі Філіпп, Відал Джекі, Гросжан-Курноєр Марі-Клєр, Утін П'єр, Туш Арунаріт, Крістіан Ізабель
МПК: C07D 405/14, A01P 3/00, C07D 409/14, A01N 43/713
Мітки: рослин, спосіб, радикальної, застосування, злакових, композиція, основі, запобіжної, фунгіцидна, боротьби, похідні, грибами, культур, фітопатогенними, тетразолу
Формула / Реферат:
1. Похідні тетразолу формули (І), (I)деТ представляє заміщену або незаміщену тетразолільну групу формули Та або Тс: Та або , ТсдеЕ1 вибирають з водню, [С1-С8]-алкілу,...
Похідні тетразолу, фунгіцидна композиція на їх основі, застосування їх для радикальної або запобіжної боротьби із фітопатогенними грибами рослин або злакових культур та спосіб боротьби із фітопатогенними грибами
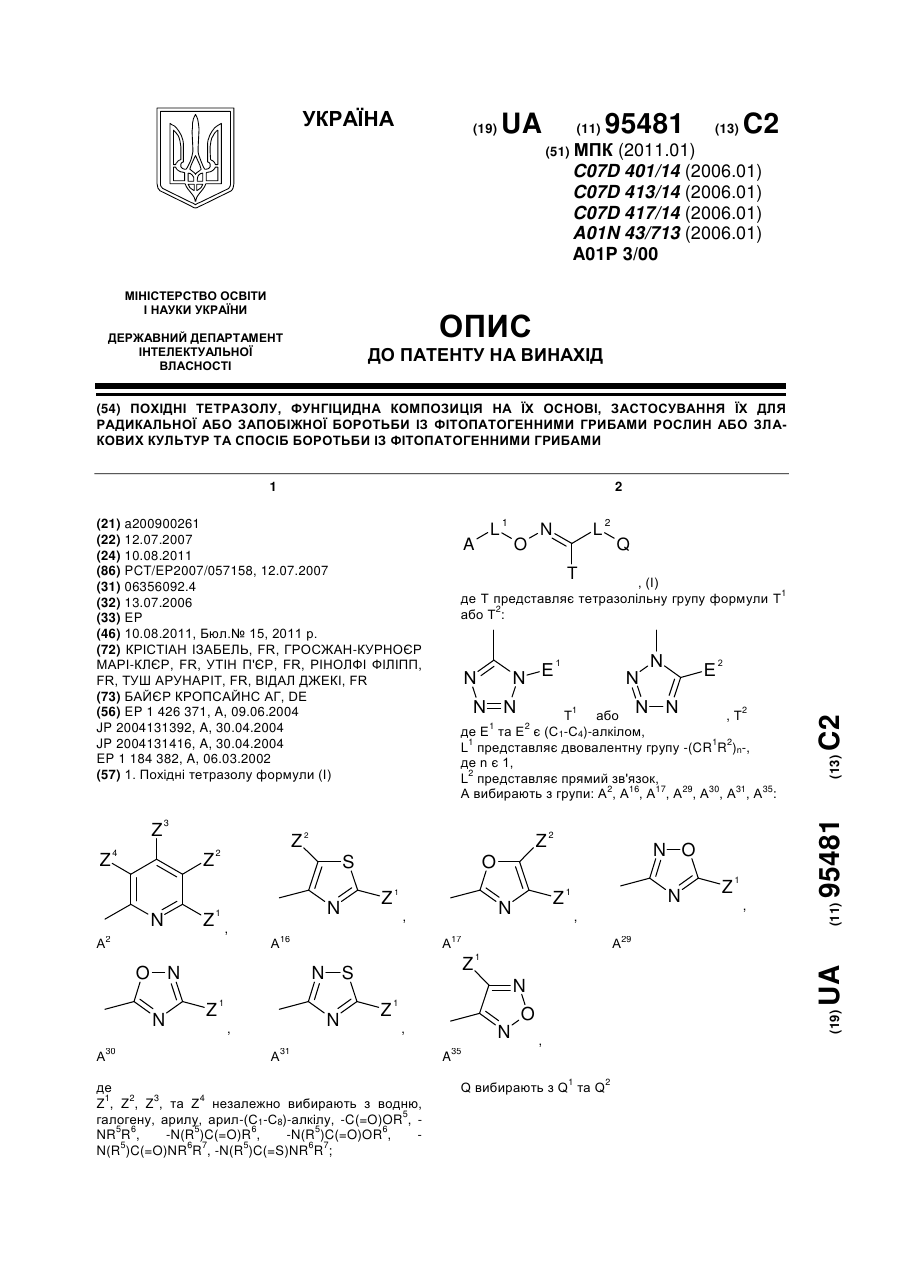
Номер патенту: 95481
Опубліковано: 10.08.2011
Автори: Рінолфі Філіпп, Крістіан Ізабель, Відал Джекі, Гросжан-Курноєр Марі-Клєр, Утін П'єр, Туш Арунаріт
МПК: A01P 3/00, C07D 413/14, C07D 417/14, A01N 43/713, C07D 401/14
Мітки: культур, спосіб, основі, похідні, злакових, фунгіцидна, композиція, боротьби, запобіжної, застосування, рослин, тетразолу, фітопатогенними, грибами, радикальної
Формула / Реферат:
1. Похідні тетразолу формули (І) , (І)де Т представляє тетразолільну групу формули Т1 або Т2:Т1 або , Т2де Е1 та Е2 є (С1-С4)-алкілом,L1 представляє двовалентну групу -(CR1R2)n-,де n є...
Похідні тетразолу, фунгіцидна композиція на їх основі, застосування їх для радикальної або запобіжної боротьби із фітопатогенними грибами рослин або злакових культур та спосіб боротьби із фітопатогенними грибами

Номер патенту: 94467
Опубліковано: 10.05.2011
Автори: Крістіан Ізабель, Відал Джекі, Рінолфі Філіпп, Гросжан-Курноєр Марі-Клєр, Утін П'єр, Туш Арунаріт
МПК: C07D 403/12, C07D 417/12, C07D 401/12, A01N 43/713, C07D 257/04, A01P 3/00
Мітки: фунгіцидна, тетразолу, злакових, рослин, боротьби, запобіжної, радикальної, похідні, культур, основі, спосіб, грибами, застосування, композиція, фітопатогенними
Формула / Реферат:
1. Похідні тетразолу формули (І) ., (І)в якійТ представляє тетразолільну групу формули Т1 або Т2 Т1 або Т2,деЕ1 і Е2 незалежно вибирають з водню, [С1-С8]-алкілу, [С1-С8]-галоалкілу,...
Засіб боротьби з шкідливими грибами, що містить інгібуючу дихання за цитохромним комплексом ііі активну речовину та феназахін і спосіб боротьби з шкідливими грибами

Номер патенту: 40670
Опубліковано: 15.08.2001
Автори: Рецлаф Гюнтер, Лоренц Гізела, Байєр Херберт, Аммерманн Еберхард, Келе Харальд, Штратманн Зігфрид, Заутер Хуберт
МПК: A01N 37/52, A01N 43/54
Мітки: боротьби, ііі, цитохромним, спосіб, комплексом, речовину, дихання, засіб, шкідливими, активну, феназахін, інгібуючу, містить, грибами
Формула / Реферат:
1. Засіб боротьби з шкідливими грибами, що містить в твердому або рідкому наповнювачі принаймні одну активну речовину І, інгібуючу дихання за цитохромним комплексом III, який відрізняється тим, що він містить феназахін формули2. Засіб за п. 1, який відрізняється тим, що містить як активну речовину І сполуку формули ІА або ІВв яких означає подвійний або простий зв’язок;R' являє собою -С [СО2СН3]=СНОСН3,...
Азолілметилоксирани, їх застосування для боротьби з фітопатогенними грибами, засіб для захисту рослин, спосіб боротьби з фітопатогенними грибами та посівний матеріал

Номер патенту: 96450
Опубліковано: 10.11.2011
Автори: Глеттлі Аліс, Реннер Йєнс, Гроте Томас, Мюллер Бернд, Дітц Йохен, Ульмшнайдер Сара, Ломанн Ян Клаас
МПК: A01N 43/653, A01P 3/00, C07D 405/06, A01C 1/08
Мітки: рослин, захисту, матеріал, грибами, фітопатогенними, посівний, спосіб, застосування, азолілметилоксирани, боротьби, засіб
Формула / Реферат:
1. Азолілметилоксирани загальної формули І, (І)в якійА означає феніл, який заміщений двома F,В означає феніл, який заміщений від одного до трьох наступних замісників галоген, NO2, аміно, С1-С4-алкіл, С1-С4-алкокси, С1-С4-галогеналкокси, С1-С4-алкіламіно, С1-С4-діалкіламіно, тіо або С1-С4-алкілтіо,за умови, що В не означає о-метилфеніл,...
Попередній патент: Сполуки боронатного ефіру і його фармацевтичні рецептури
Наступний патент: Сполуки 1-арилкарбоніл-4-оксипіперидину, придатні для лікування нейродегенеративних захворювань
Випадковий патент: Піщаний фільтрувальний пристрій