Поліпептид ліганду mpl (варіанти), ізольована молекула нуклеїнової кислоти, що кодує поліпептид ліганду mpl, вектор експресії, клітина-хазяїн, спосіб продукування поліпептиду ліганду mpl (варіанти), композиція,































































































































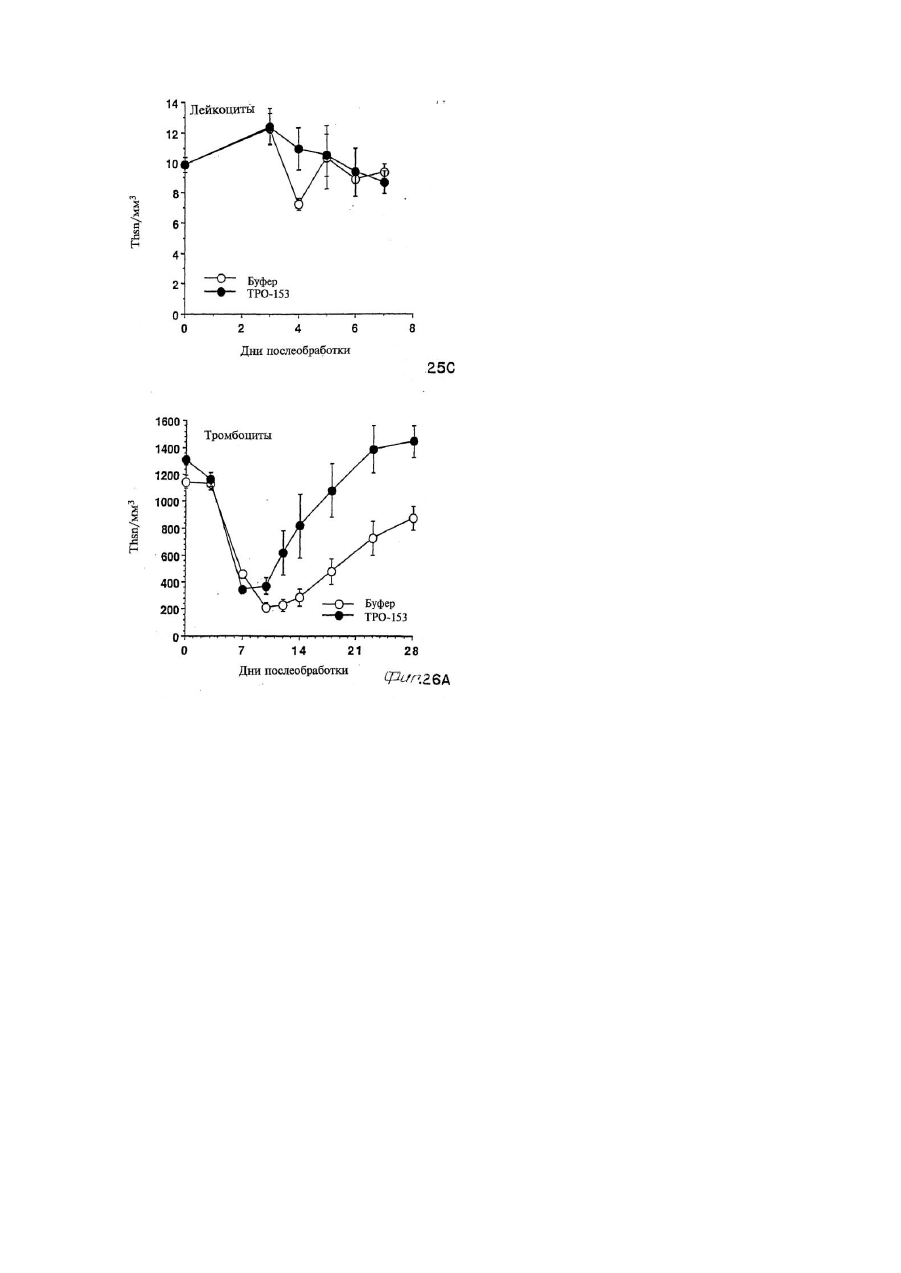
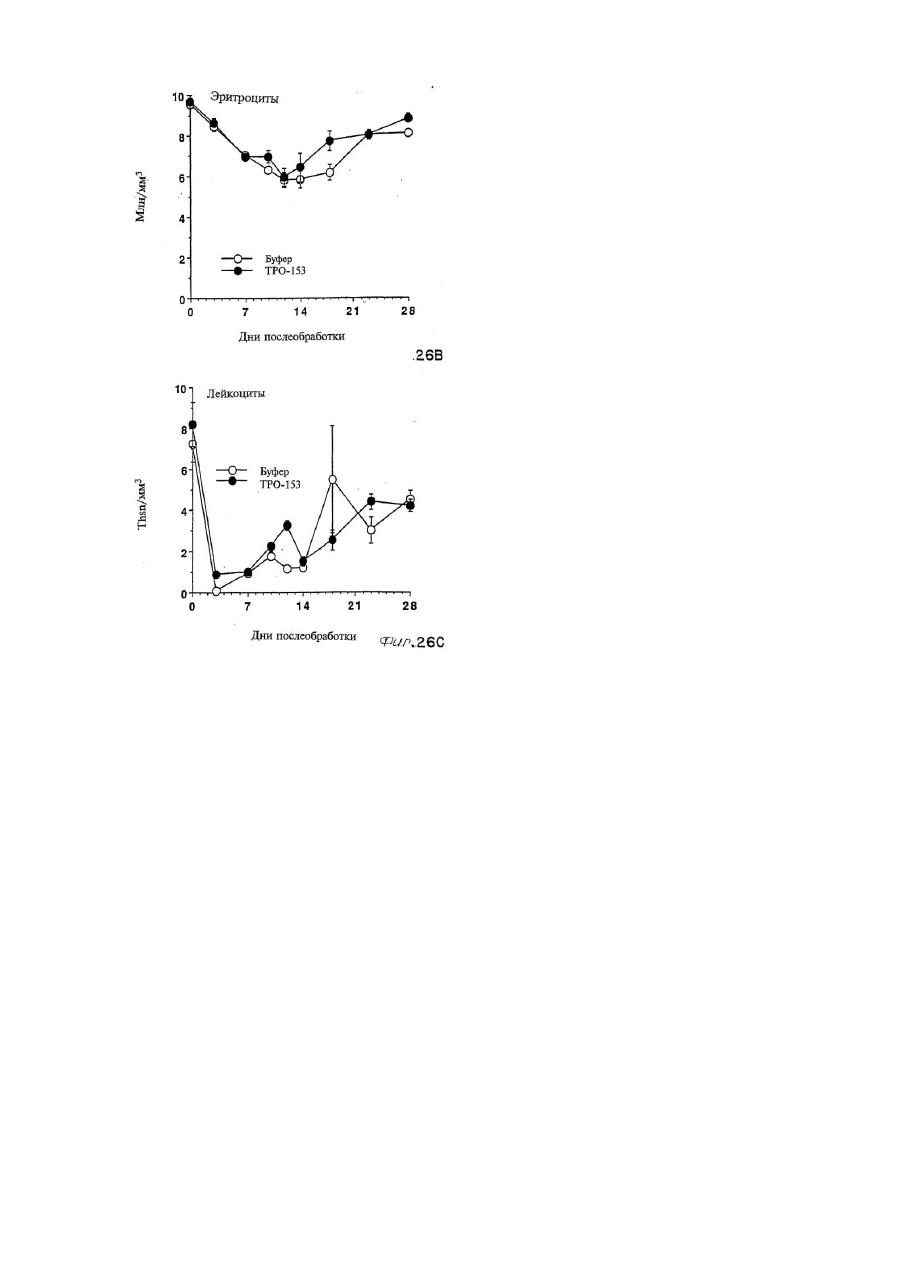
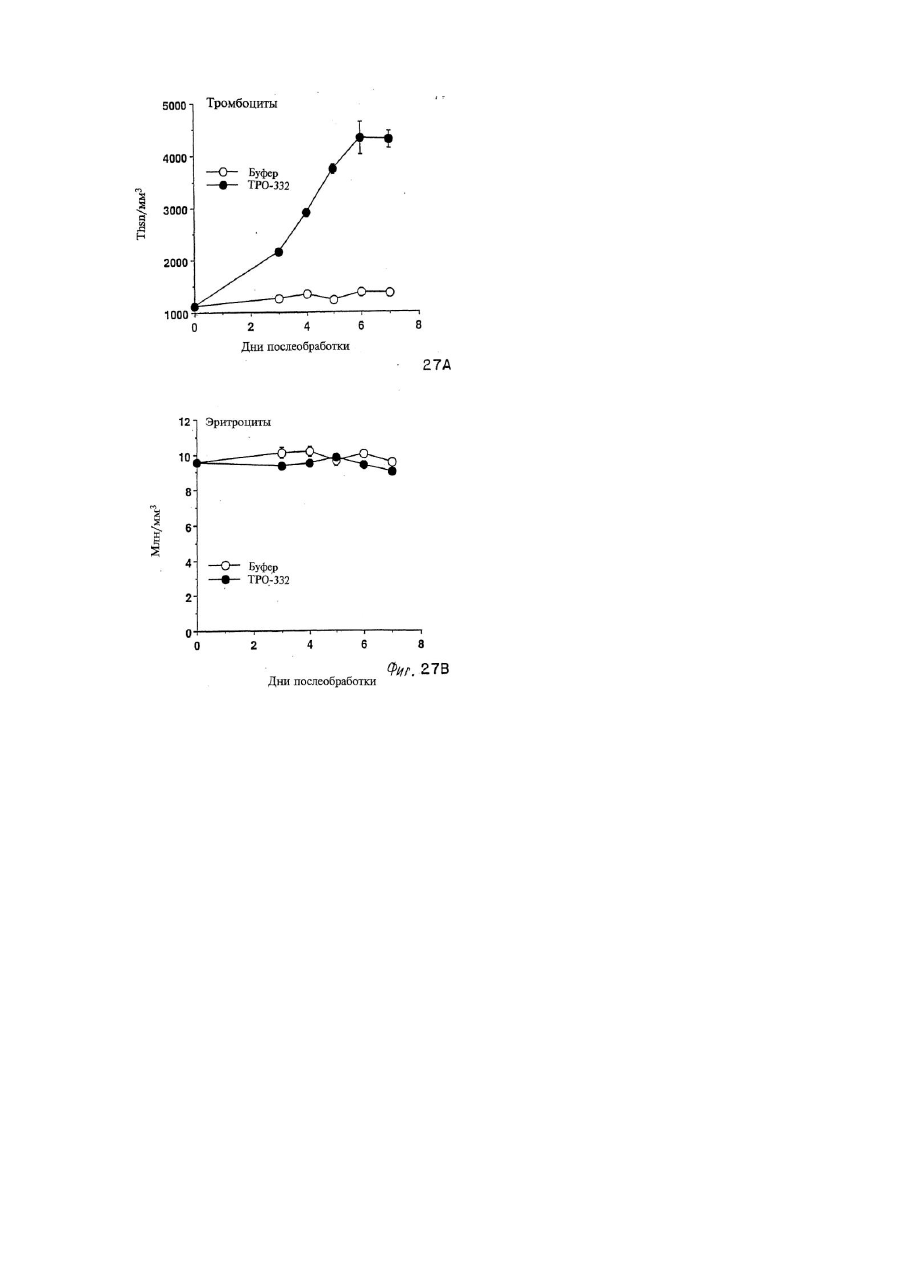
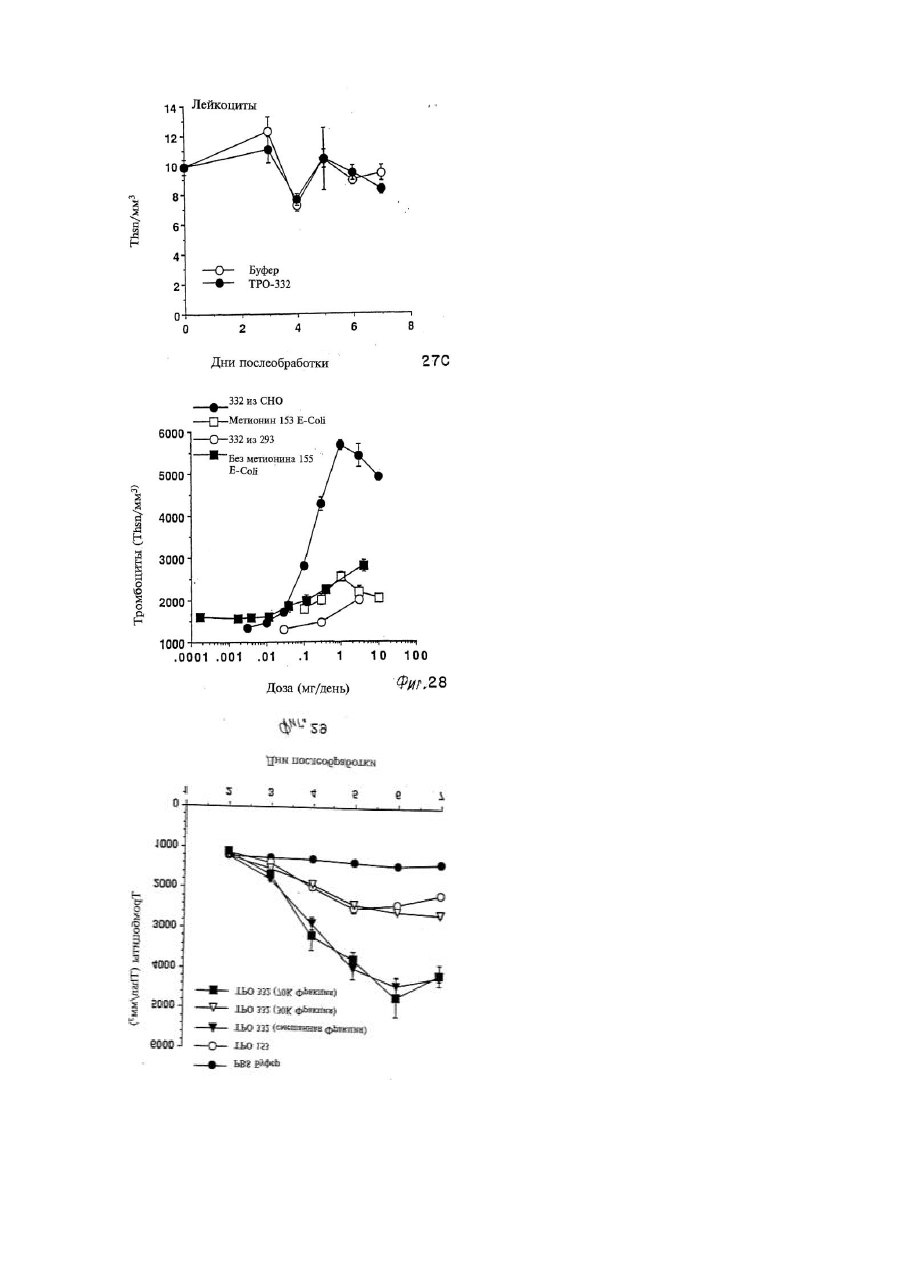















Формула / Реферат
1. Полипептид лиганда mpl, получаемый по способу, включающему:
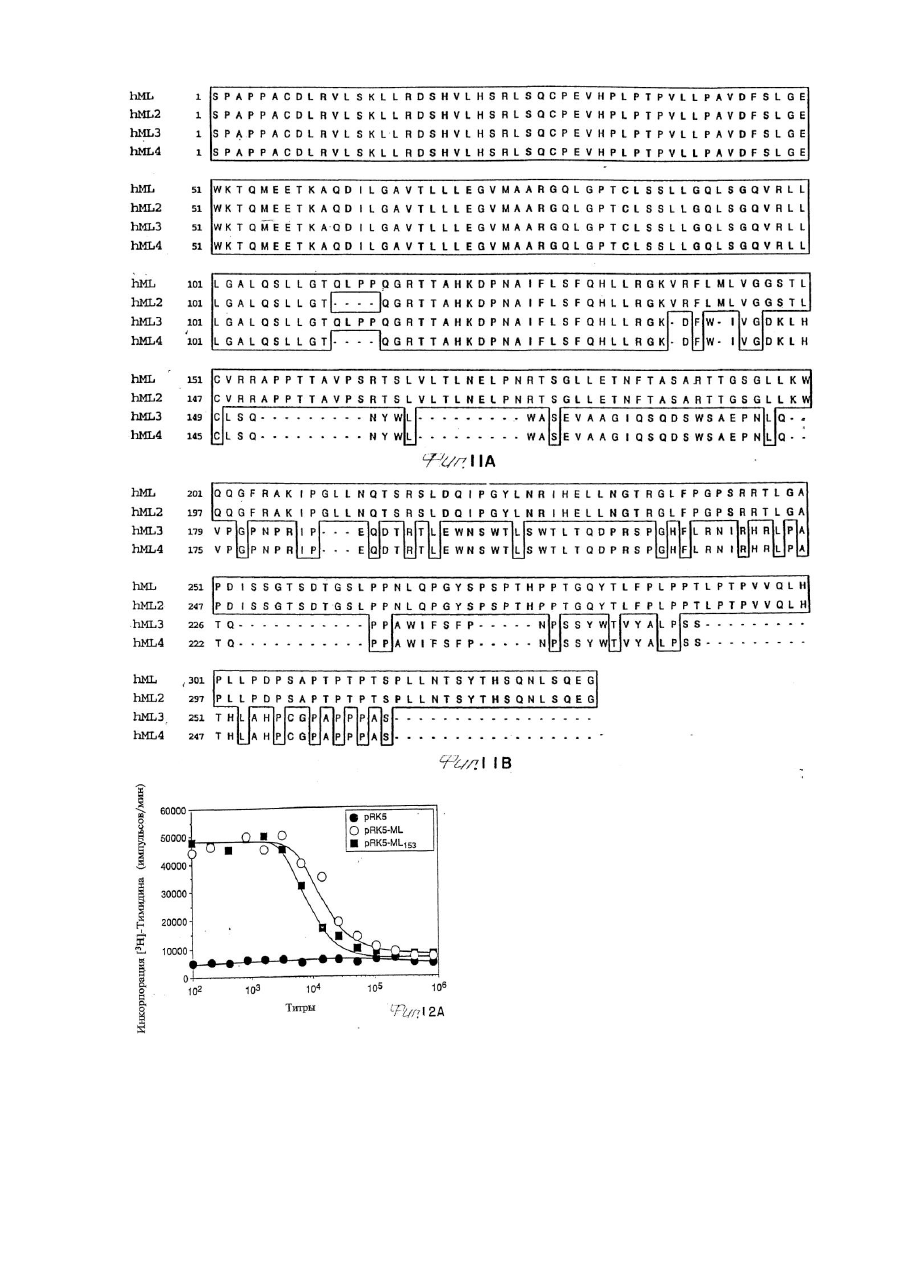
(і) просеивание человеческой геномной библиотеки с олигонуклеотидом на основе геномной последовательности, представленной на фиг. 9, для изоляции геномной ДНК, которая включает кодирующую последовательность экзона лиганда mpl, представленную на фиг. 9, вместе с остальными экзонами гена,
(ii) вставку ДНК в вектор экспрессии,
(iii) трансфекцию клеток млекопитающих вектором и экспрессию гена, и
(iv) извлечение полипептида лиганда mpl из среды клеточной культуры, при этом амино-конечная последовательность полипептида содержит:
SPAPPACDLRVLSKLLRDSHVLHSRL.
2. Полипептид лиганда mpl, получаемый по способу, включающему:
(і) изоляцию из геномной библиотеки геномной ДНК, гибридизируемой в нестрогих условиях с последовательностью ДНК
5' GCCCTGAAGGACGTGGTCGTCACGAAGCAGTTTATTTAGGAGTCG 3'
и содержащей экзоны, кодирующие полипептид лиганда mpl, (ii) вставку ДНК в вектор экспрессии,
(iii) трансфекцию клеток млекопитающих вектором и экспрессию гена, и (iv) извлечение полипептида лиганда mpl из среды клеточной культуры, при этом амино-конечная последовательность полипептида содержит:
SPAPPACDPRLLNKLLRDDHVLHGR, или
SPAPPACDLRVLSKLLRDSHVLHSRL.
3. Полипептид лиганда mpl, получаемый по способу, включающему:
(і) идентификацию подходящего клеточного источника РНК для человеческого лиганда mpl и приготовление одной или нескольких библиотек кДНК из упомянутой РНК,
(ii) гибридизацию-просеивание библиотеки или библиотек с меченым олигонуклеотидом на основе кодирующей последовательности, представленной на фиг. 9, для идентификации и изоляции клона, содержащего кодирующую последовательность лиганда mpl,
(iii) вставку кодирующей ДНК в вектор экспрессии, подходящий для экспрессии в клетках млекопитающего,
(iv) трансфекцию клеток млекопитающих вектором и выражение кДНК и
(v) извлечение полипептида лиганда mpl из среды клеточной культуры,
при этом амино-конечной последовательностью полипептида является SPAPPACDLRVLSKLLRDSHVLHSRL.
4. Полипептид лиганда mpl, получаемый по способу, включающему:
(і) приготовление одной или нескольких библиотек кДНК из мРНК, экстрагированной из клеток человеческой почки,
(ii) гибридизацию-просеивание библиотеки или библиотек с меченым олигонуклеотидом на основе кодирующей последовательности, представленной на фиг. 9, для идентификации и изоляции клона, содержащего кодирующую последовательность лиганда mpl,
(iii) вставку кодирующей ДНК в вектор экспрессии, подходящий для экспрессии в клетках млекопитающего,
(iv) трансфекцию клеток млекопитающих вектором и экспрессию кДНК и (v) извлечение полипептида лиганда mpl из среды клеточной культуры,
при этом амино-конечной последовательностью полипептида является SPAPPACDLRVLSKLLRDSHVLHSRL.
5. Изолированный, в основном гомогенный, лиганд mpl, отличающийся тем, что
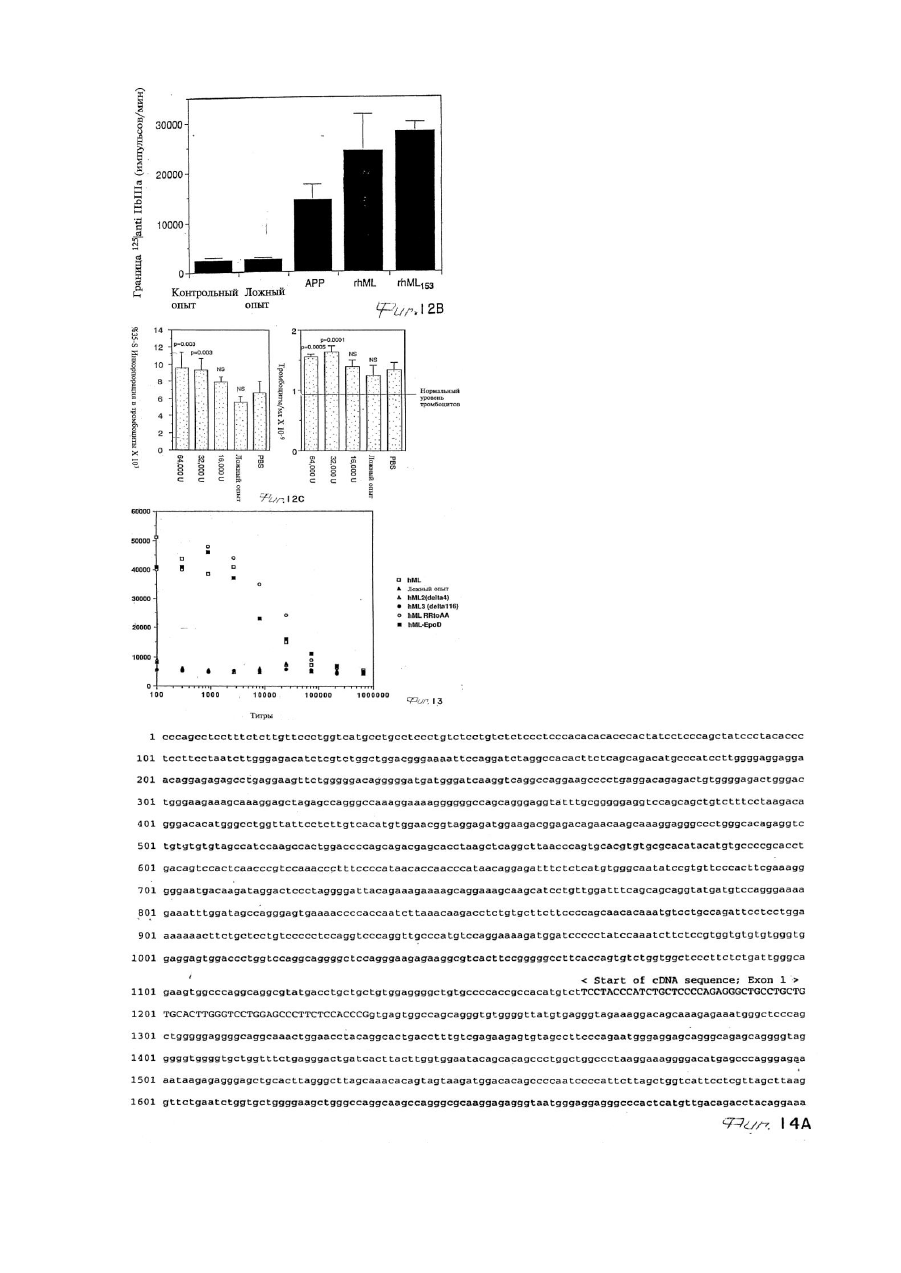
(a) лиганд стимулирует инкорпорацию меченых нуклеотидов (3H-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, зараженных человеческим mpl Р;
(b) лиганд стимулирует инкорпорацию 35S в циркулирующие тромбоциты при анализе ответных тромбоцитов;
(c) лиганд стабилен в среде рН 2,5, SDS 0,1% и 2М мочевины;
(d) лиганд является гликопротеином;
(e) амино-конечная последовательность полипептида следующая:
SPAPPACDPRLLNKLLRDDHVLHGR или
SPAPPACDLRVLSKLLRDSHVLHSRL.
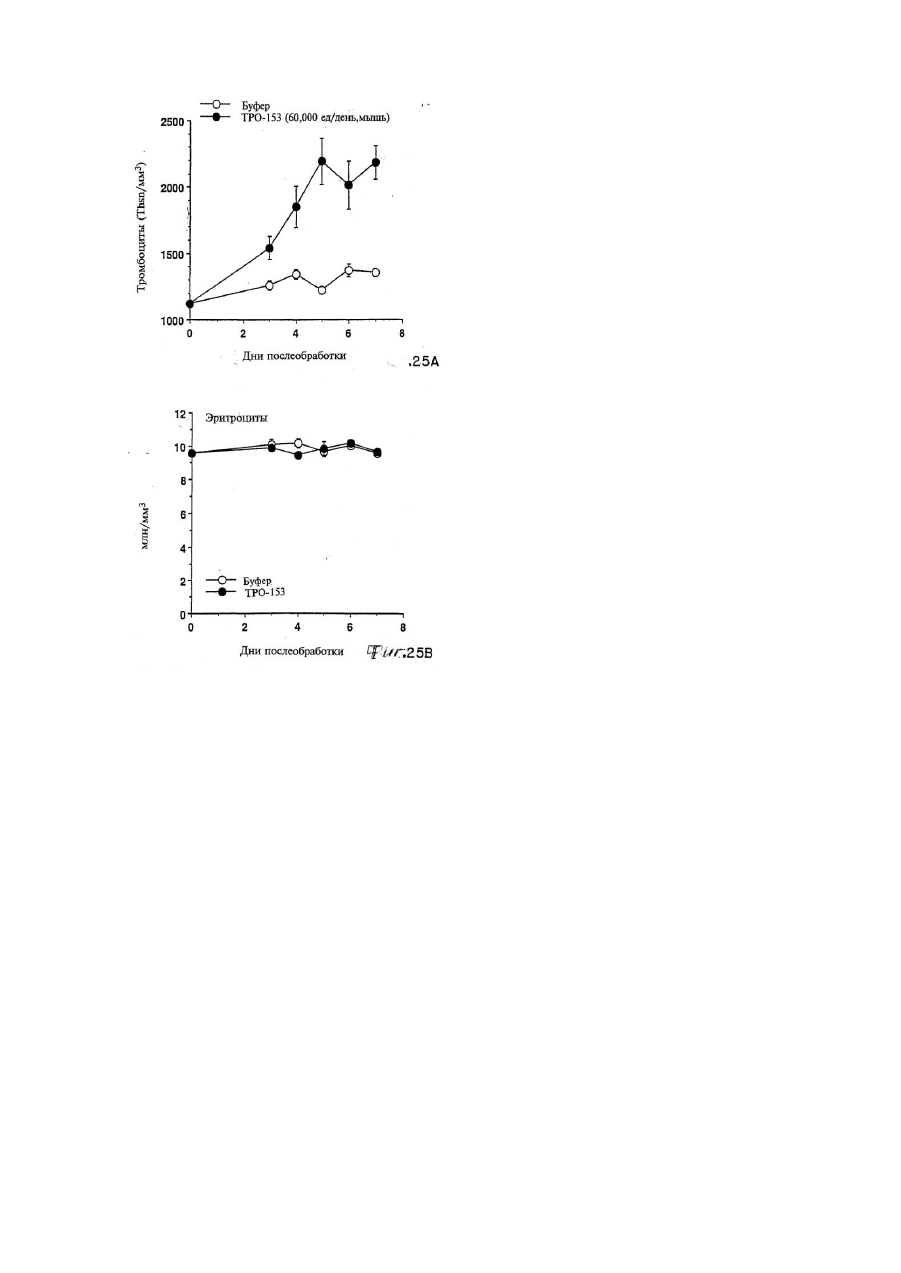
6. Лиганд mpl, получаемый очисткой из апластической плазмы при помощи гидрофобной интерактивной хроматографии, иммобилизованной окрашивающей хроматографии и хроматографии по mpl-сродству, и имеющий очевидный молекулярный вес, обнаруживаемый при помощи SDS-PAGE в условиях восстановления, в диапазоне 18-22 кДа, 28 кДа или 30 кДа.
7. Изолированный полипептид лиганда mpl, кодируемый нуклеиновой кислотой, имеющей последовательность, которая гибридизируется в умеренно строгих условиях с нуклеиновой кислотой, имеющей последовательность нуклеотидов 119-390 последовательностей 4 или 5.
8. Изолированный лиганд mpl по любому из пп. 1-7, отличающийся тем, что является биологически активным.
9. Полипептид лиганда mpl, который имеет как минимум 70% идентичности последовательности с полипептидом по любому из пп. 1-7 и имеет биологическую активность лиганда mpl.
10. Полипептид лиганда mpl, являющийся аллелью или вариантом полипептида по любому из пп. 1-7, который сохраняет биологическую активность лиганда mpl.
11. Полипептид лиганда mpl no любому из пп. 1-10, отличающийся тем, что является производным от человеческих видов.
12. Полипептид лиганда mpl по любому из пп. 1-11, отличающийся тем, что является производным от нечеловеческих видов.
13. Полипептид лиганда mpl по любому из пп. 1-12, отличающийся тем, что является негликозилированным.
14. Полипептид, включающий лиганд mpl по любому из пп. 1-13, отличающийся тем, что является слитым с гетерологичным полипептидом.
15. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид по любому из пп. 1-14.
16. Изолированная молекула нуклеиновой кислоты, выбранная из:
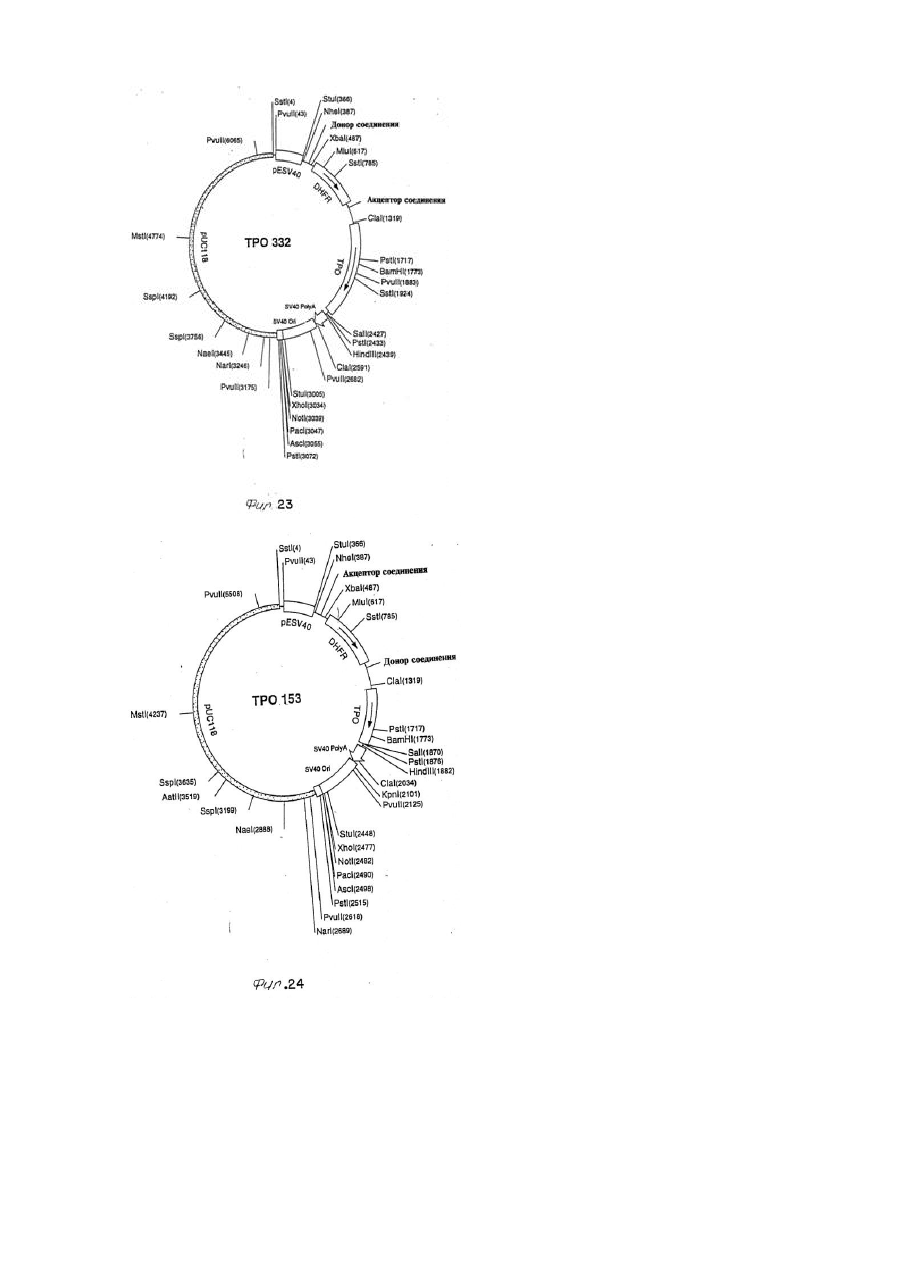
(a) клона кДНК, получаемого на этапе (iii) пункта 3 или этапе (ii) пункта 4;
(b) последовательность ДНК, способная к гибридизации в строгих условиях с клоном по подпункту (а) и
(c) генетический вариант любой из последовательностей ДНК по подпунктам (а) и (b), который кодирует полипептид, обладающий биологическим свойством природно встречающегося полипептида лиганда mpl.
17. Изолированная нуклеиновая кислота, имеющая последовательность, которая гибридизируется в умеренно строгих условиях с нуклеиновой кислотой, имеющей последовательность нуклеотидов 119-390 последовательностей 4 или 5, и кодирует лиганд mpl.
18. Геномная ДНК или кДНК, гибридизирующаяся в условиях от умеренно строгих до очень строгих с природно встречающейся молекулой нуклеиновой кислоты, имеющей последовательность, кодирующую лиганд mpl, определенный по любому из пп. 1-7.
19. Нуклеиновая кислота по любому из пп. 15-18, отличающаяся тем, что оперативно связывается с промотором.
20. Вектор экспрессии, включающий нуклеиновую кислоту по любому из пп. 15-18, оперативно связываемую с контрольной последовательностью, распознаваемой клеткой-хозяином.
21. Клетка-хозяин, трансформируемая нуклеиновой кислотой по любому из пп. 15-20, таким образом, что приобретает способность экспрессировать указанную нуклеиновую кислоту для продуцирования полипептида лиганда mpl.
22. Способ продуцирования полипептида лиганда mpl, включающий экспрессию рекомбинантной нуклеиновой кислоты по любому из пп. 15-20 в подходящей клетке-хозяине, выделение полипептида лиганда mpl из клетки-хозяина или среды культуры клетки-хозяина.
23. Способ продуцирования полипептида лиганда mpl, включающий экспрессию в клетке индигенного гена лиганда mpl в условиях регулирования промотором/усилителем, угнетающим агентом или экзогенным элементом модуляции транскрипции, который вводится в геном клетки с приближением и ориентацией, достаточными для влияния на транскрипцию ДНК, кодирующей полипептид лиганда mpl, выделение полипептида лиганда mpl.
24. Способ приготовления лекарственного средства, включающий объединение полипептида лиганда mpl по любому из пунктов 1-14, или полученного способом по п. 22, с фармацевтически приемлемым носителем.
25. Композиция, включающая полипептид лиганда mpl по любому из пп. 1-14, или полученный способом по пункту 22, и фармацевтически приемлемый носитель.
26. Композиция по п. 25, отличающаяся тем, что дополнительно включает терапевтически эффективное количество агента, выбранного из цитокинов, колониестимулирующих факторов и интерлейкинов.
27. Композиция по пункту 26, отличающаяся тем, что агент выбирается из LIF, G-CSF, GM-CSF, М-CSF, Еро, IL-1, IL-2, IL-3, IL-5, IL-6, IL-7, IL-8, IL-9 и IL-11.
28. Способ усиления испытательного образца нуклеиновой кислоты, включающий инициирование реакции полимеразы нуклеиновых кислот с нуклеиновой кислотой по п. 18.
29. Способ определения присутствия полипептида лиганда mpl, включающий гибридизацию нуклеиновой кислоты по п. 18 с испытательным образцом нуклеиновой кислоты и определение присутствия нуклеиновой кислоты полипептида лиганда mpl.
30. Способ приготовления антитела, специфичного к лиганду mpl, включающий: использование полипептида лиганда mpl по любому из пп. 1-14, или полученного способом по п. 22, для формирования иммунного ответа у субъекта и приготовление антитела, специфичного к лиганду mpl.
31. Способ по п. 30, отличающийся тем, что получают моноклональное антитело.
32. Антитело, получаемое способом по п. 30 или п. 31.
33. Антитело, способное специфически связываться с полипептидом лиганда mpl по любому из пунктов 1-14.
34. Антитело по пункту 33, отличающееся тем, что является моноклональным антителом.
35. Клеточная линия гибридомы, продуцирующая антитело по п. 32 или п. 34.
Текст
Изобретение относится к изоляции, очистке и рекомбинации или химическому синтезу белков, влияющих на жизнеспособность, размножение, видоизменение и зрелость кроветворных клеток, особенно тромбоцитообразующих клеток. Данное изобретение главным образом относится к клонированию и экспрессии нуклеиновых кислот, кодирующи х лиганды белков, способные связываться с mpl, членом сверхсемейства цитокиновых рецеаторов, и активировать его. Изобретение также относится к использованию этих белков, индивидуально или в сочетании с другими цитокинами для лечения иммунных заболеваний или нарушений кроветворения, включая тромбоцитопению. I. Кроветворная система Кроветворная система вырабатывает зрелые, имеющие высокоспециализированное назначение клетки крови, известные науке как жизненно необходимые для всех млекопитающих. Эти зрелые клетки включают: эритроциты, переносящие кислород и углекислый газ, Т- и В-лимфоциты, отвечающие за иммунитет на основе клеток или антител, тромбоциты, отвечающие за свертывание крови, и гранулоциты и макрофаги, специализирующиеся на удалении вредных веществ и шлаков и выступающие вспомогательными клетками при борьбе с инфекцией. Гранулоциты, в свою очередь, подразделяются на нейтрофилы, эозинофилы, базофилы и мачтовые клетки, "специализированные типы клеток, имеющие различные функции. Примечательно, что все эти специализированные типы зрелых клеток происходят от одного общего простейшего типа клеток, известных как плюрипотенциальные стволовые клетки, первоначально найденные в костном мозге [Dexter et. al., Ann. Re v. Cell Biol., 3:423-441 (1987)]. Зрелые высокоспециализированные клетки крови должны постоянно вырабатываться в больших количествах в течение всей жизни млекопитающего. Подавляющее большинство этих специализированных клеток крови остаются функционально активными лишь от несколько часов до нескольких недель [Cronkite et al., Blood Cells, 2:263-284 (1976)]. Таким образом, необходимо постоянное возобновление зрелых клеток крови, самого клеточного ствола, а также всех промежуточных или связующи х первичных клеточных линий, лежащих между простейшими и зрелыми клетками, чтобы поддерживать нормальное стабильное состояние клеток крови млекопитающих. Сердцем кроветворной системы является плюрипотенциальная стволовая клетка (клетки). Эти клетки сравнительно немногочисленны и подвергаются самовозобновлению путем размножения, вырабатывая дочерний ствол, или трансформируются путем нескольких этапов видоизменений, в более зрелые прародительские клетки с ограниченными линиями воспроизводства, в конечном счете образующие высокоспециализированные зрелые кровяные клетки. Например, определенные мультипотенциальные прародительские клетки, известные в литературе как CFC-Mix и происходящие от клеток ствола, подвергаются размножению (самовозобновлению) и развитию, вырабатывая колонии, содержащие различные миелоидные клетки: эритроциты, нейтрофилы, мегакариоциты (предшественники тромбоцитов), макрофаги, базофилы, эозинофилы и мачтовые клетки. Другие прародительские клетки лимфоидного происхождения подвергаются размножению и развитию в Т-клетки и Вклетки. Кроме того, между прародительскими клетками CFC-Mix и миелоидными клетками лежит другой, промежуточный, класс прародительских клеток. Эти прародительские клетки с ограниченными линиями воспроизводства классифицируются на основе потомства, которое они вырабатывают. Так, известными непосредственными предшественниками миелоидных клеток являются эритроидные колониеобразующие элементы (CFU-E) для эритроцитов, гранулоцито/макрофаго-колониеобразующие клетки (GM-CFC) для нейтрофилов и макрофагов, мегакариоцитоколониеобразующие клетки (Meg-CFC) для мегакариоцитов, эозинофил-колониеобразующие клетки (Eos-CFC) для эозинофилов и базофил-колониеобразующие клетки (Bas-CFC) для мачтовых клеток. Известны (см. ниже) или могут быть открыты также другие промежуточные прародительские клетки между плюрипотенциальными стволовыми клетками и законченными кровяными клетками, отличающиеся различной степенью ограниченности воспроизводства и возможностями самовозобновления. По всей видимости, основа нормальной системы кроветворения - снижение способности к самовозобновлению в мультипотенциальном качестве и приобретение качества ограниченности воспроизводства и законченности в развитии. Так, у одной границы спектра кроветворных клеток лежит плюрипотенциальная стволовая клетка, обладающая способностью к самовозобновлению и видоизменению во все различные видо-специфические прародительские клетки. Эта способность является основой метода трансплантации костного мозга, при котором простейшие стволовые клетки воссоздают целую систему кроветворных клеток. У другой границы спектра лежат прародительские клетки с высокой ограниченностью воспроизводства и их потомство, которое потеряло способность к самовозобновлению, но приобрело зрелую функциональную активность. Размножение и развитие стволовых клеток и прародительских клеток с ограниченными линиями воспроизводства строго контролируется различными агентами роста кроветворных клеток или цитокинами. Роль этих агентов роста в живых организмах сложна и не до конца понятна. Некоторые агенты роста, такие как интерлейкин-3 (IL-3), способны стимулировать как мультипотенциальные стволовые клетки, так и прародительские клетки нескольких ветвей потомств, включая, например, мегакариоциты. Другие агенты, такие как гранулоцито/макрофаго-колониестимулирурщий фактор (GM-CSF), первоначально был определен как действующий только на клетки GM-CFC. Позднее, однако, было обнаружено, что он также влияет на размножение и развитие также и мегакариоцитов. Так, оказалось, что IL-3 и GM-CSF имеют перекрывающиеся области биологической активности, хотя с различной мощностью. Еще позднее было открыто, что интерлейкин-6 (IL-6) и интерлейкин-11 (IL-11), не оказывая очевидного влияния на образование мег-колоний по отдельности, показывают синергический эффект с IL-3, стимулир уя развитие мегакариоцитов [Yonemura et al., Εxp. Hematol., 20:1011-1016 (1992)]. Итак, агенты роста кроветворных клеток могут влиять на рост и видоизменение одной или более линий, их активность может перекрываться с другими агентами роста во влиянии на одну клеточную линию, или может проявляться синергически с другими агентами. Оказывается также, что агенты роста могут проявлять свой эффект на различных стадиях развития клеток от плюрипотенциальных стволовы х клеток, через различные передаточные прародительские клетки с ограниченными линиями воспроизводства до зрелых кровяных клеток. Например, эритропоэтин (еро) показывает ускорение размножения только зрелых эритроидных прародительских клеток. IL-3 проявляет свой эффект раньше, действуя на простейшие стволовые клетки и промежуточные прародительские клетки с ограниченными линиями воспроизводства. Другие агенты роста, такие как агент стволовы х клеток (SCF) могут влиять даже на еще более примитивные клетки. Из вышеизложенного следует вывод, что вновь открытые агенты роста, которые влияют на жизнеспособность, размножение, видоизменение или зрелость любых кровяных клеток или их предшественников, могут быть полезны, особенно при восстановлении кроветворной системы, активность которой снижена из-за болезни или после радио- или химиотерапии. II. Мегакариоцитопоэз и вырабатывание тромбоцитов Закономерности мегакариоцитопоэза и вырабатывания тромбоцитов описан у [Ma zur, Exp. Hematol., 15:248 (1987) и Hoffman, Blood, 74:1196-1212 (1989)]. Коротко: плюрипотенциальные стволовые клетки костного мозга видоизменяются в мегакариоциты, эритроциты и клетки миелоцитных линий. Считается, что существуе т иерархия передаточных мегакариоцитных прародительских клеток между стволовыми клетками и мегакариоцитами. Было идентифицировано как минимум три класса мегакариоцитных прародительских клеток, а именно: мегакариоциты всплескоообразующего элемента (BFU-MK), мегакариоциты колониеобразующего элемента (CFU-MK) и прародительские клетки мегакариоцитов низкой плотности (LDCFU-MK). Процесс развития мегакариоцитов представляет собой последовательность из трех стадий, базирующуюся на стандартном морфологическом критерии. Первые распознаваемые члены семейства мегакариоцитов (МК или meg) - мегакариобласты. Эти клетки первоначально составляют 20-30мкм в диаметре и имеют базофильную цитоплазму и ядро невыразительной формы с сетчатым хроматином и несколькими ядрышками. Затем мегакариобласты могут содержать до 32 ядер (полиплоид), но цитоплазма остается разреженной и незрелой. По мере развития, ядро становится более выпуклым и окрашенным, количество цитоплазмы увеличивается и она становится более ацидофильной и зернистой. Более зрелые клетки этого семейства могут на периферии создавать видимость появления тромбоцитов. Обычно на стадии бластов находится менее 10% мегакариоцитов, а более 50% являются зрелыми. Произвольно выбранная морфологическая классификация, обычно применяемая к семейству мегакариоцитов, следующая: в самой ранней форме - мегакариобласты; в промежуточной форме - промегакариоциты или базофильные мегакариоциты, и в последних формах - зрелые (ацидофильные, зернистые или тромбоцит-образующие) мегакариоциты. Зрелые мегакариоциты вытягивают нити цитоплазмы в синусоидальном направлении, эти нити отсоединяются и разделяются на отдельные тромбоциты [Williams et al., Hematology, 1972]. Считается, что мегакариоцитопоэз включает несколько регулирующи х факторов [Williams et al., Br. J.Hematol., 52:173 (1982)] и [Williams et al., D. Cell Physiol., 110:101 (1982)]. Ранняя стадия мегакариоцитопоэза определена как митотическая, касающаяся, размножения клеток и инициирования колоний из CFU-MK, но на которую не влияет количество тромбоцитов [Burstein et al., J. Cell Physiol., 109:333 (1981) и Kimura et al., Exp. Hematol., 13:1048 (1985)]. Последняя стадия развития - не митотическая - включает ядерную полиплоидизацию и цитоплазматическое развитие и, вероятно, регулируется механизмом обратной связи по периферийному числу тромбоцитов [Odell et al., Blood, 48:765 (1976) и Ebbe et al., Blood, 32:787 (1968)]. Обсуждалось существование отчетливых, имеющих специфическое строение, колониестимулирующих агентов для мегакариоцитов [Mazur, Exp. Hematol., 15:340-350 (1987)]. Однако большинство авторов считают, что столь важный для жизни процесс, как вырабатывание тромбоцитов, должен регулироваться цитокином (цитокинами), ответственными за этот процесс. Гипотеза существования специфического цитокина (цитокинов) дала толчок для более чем 30-летнего поиска, но до сих пор такой цитокин не был выделен и классифицирован как единственный MK-CSF (ТРО). Хотя было описано, что MK-CSF были частично выделены из экспериментально выработанной тромбоцитопении [Hill et al., Exp. Hematol., 14:752 (1986)], из кондиционированной среды почки человеческого эмбриона [CM] (McDonald et al., J. Lab. Clin. Med., 85:59 (1975)], в человеке из пластической анемии и мочевых экстрактов идиопатической тромбоцитопенической пурпуры [Kawakita et. al., Blood, 6:556 (1983)] и плазмы [Hoffman et al., J. Clin. Invest., 75:1174 (1985)], их физиологические функции в большинстве случаев еще непонятны. Кондиционированная среда митоген-активированных селезеночных клеток фитолакки американской (PWM-SpC M) и клеточная линия мышиных миеломоноцитов WEHI-3 (WEHI-3CM) использовались в качестве среды для образования мегакариоцитов. PWM-SpCM содержит агенты, вызывающие рост CFU-MK [Metcalf et al., Pro. Natl. Acad. Sci., USA, 72:1744-1748 (1975); Quesenberry et al., Blood, 65:214 (1985); Iscove, N.N., in Hematopoietic Cell Differentiation, ICN-UCLA Symposia on Molecular and Cellular Biology, Vol.10, Golde et al., eds. (New York, Academy Press) pp.37-52 (1978)], одним из которых является интерлейкин-3 (IL-3), мультивидовой колониестимулирующий агент (multi-CSF [Burstein, Blood Cells, 11:469 (1986)]. Другие агенты в этой среде еще не идентифицированы и не выделены. WEHI-3 миелсжоноцитная клеточная линия мыши, выделяющая относительно большое количество IL-3 и меньшее количество GM-CSF. Было обнаружено, что IL-3 усиливает рост широкого спектра кроветворных клеток [Ihle et al., D.Immunol., 13:282 (1983)]. Также был обнаружен синергический эффект IL-3 со многими известными кроветворными гормонами или агентами роста [Bartelmez et al., J.Cell Physiol., 122:362-369 (1985) и Warren et al., Cell, 46:667-674 (1988)], включая эритропоэтин (ЕРО) и интерлейкин-1 (IL-1), при введении самых ранних мультипотенциальных предшественников и образовании очень больших смешанных колоний кроветворных клеток. Другие источники усилителей образования мегакариоцитов были найдены в клеточных линиях макрофагов легких и костей мыши, клеток брюшинных выделений и почечных клетках человеческого эмбриона. Несмотря на некоторые противоречивые данные [Mazur, Exp. Hematol., 15:340-350 (1987)], очевидно [Geissler et al., Br. J. Haematol., 60:233-238 (1985)], что в мегакариоцитопоэзе усиливающую роль больше играют активированные Т-лимфоциты, чем моноциты. Эти заключения наводят на мысль о том, что выделения активированных Т-лимфоцитов, такие как интерлейкины, могут быть регулирующими агентами в развитии МК [Geissler et al., Exp. Hematol., 15:845-853 (1987)]. Большое число работ по мегакариоцитопоэзу с очищенным эритропоэтином ЕРО [Vainchenker et al., Blood, 54:940 (1979); McLeod et al., Nature, 261:492-4 (1976); Williams et al., Exp. Hematol., 12:734 (1984)] показывает, что это т гормон оказывает усиливающее влияние на образование колонии МК. Это было также продемонстрировано на сывороточных и бессывороточных культура х и в отсутствие вспомогательных клеток [Williams et al., Exp. Hematol., 12:734 (1984)]. ЕРО было дано определение как участвующему скорее в одно- и двухклеточной стадии мегакариоцитопоэза, в отличие от PWM-SpCM, участвующего в четырехклеточной стадии развития мегакариоцита. Взаимодействие всех эти х агентов, как на ранних, так и на поздних стадиях мегакариоцитопоэза еще предстоит изучить. Данные, полученные в нескольких лабораториях, наводят на мысль о том, что единственными мультивидовыми агентами, которые по отдельности обладают МК-колониестимулирующей активностью, являются GM-CSF и IL-3 и в меньшей степени стимулирующий В-клетки агент IL-6 [Ikebuchi et al., Proc. Natl. Acad. Sci. USA, 84:9035 (1987)]. Позднее несколько авторов сообщили о том, что IL-11 и агент ингибирования лейкемии (LIE) взаимодействуют синергически с IL-3, увеличивая размер и плоидность мегакариоцита [Yonemura et al., British Journal of Hematology, 84:16-23 (1993); Burstein et al., J. Cell. Physiol., 153:305-312 (1992); Metcalf et al., Blood, 76:50-56 (1990); Metcalf et al., Blood, 77:2150-2153 (1991); Bruno et al., Exp. Hematol., 19:378-381 (1991); Yonemura st al., Exp Hematol., 20:1011-1016 (1992)]. Другие документы, представляющие интерес, включают: [Eppstein et al., U.S. Patent No.4,962,091; Chong, U.S. Patent No.4,879,111; Fernandas et al., U.S. Patent No.4,604,377; Wissler et al., U.S. Patent No.4,512,971; Gottlieb et al., U.S. Patent No.4,468,379; Bennett et al., U.S. Patent No.5,215,895; Kogan et al., U.S. Patent No.5,250,732; Kimura et al., Eur 3. Immunol., 20(9):1927-1931 (1990); Secor et al., J. of Immunol., 144(4):1484-1489 (1990); Warren et al., J. of Immunol., 140(1):94-99 (1988); Warren et al., Exp. Hematol., 17(11):1095-1099 (1989); Bruno et al., Exp. Hematol., 17(10):1038-1043 (1989); Tanikawa et al., Exp. Hematol., 17(8):883-888 (1989); Koike et al., Blood, 75(12):2286-2291 (1990); Lotem, Blood, 75(5):1545-1551 (1989); Rennick et al., Blood, 73(7): 1828-1835 (1989); Clutterbuck et al., Blood, 73(6):1504-1512 (1989)]. III. Тромбоцитопения Тромбоциты являются основными элементами механизма свертывания крови. Снижение уровня циркулирующих тромбоцитов, называемое тромбоцитопенией, происходит при различных клинических состояниях и заболеваниях. Диагноз тромбоцитопении в общем случае ставится при количестве тромбоцитов ниже 150´109 на литр. Главные причины тромбоцитопении могут быть разбиты на три і категории, исходя из продолжительности жизни тромбоцитов, а именно: (1) ухудшение вырабатывания тромбоцитов костным мозгом, (2) изоляция тромбоцитов в селезенке (спленомегалия) или (3) повышенная деструкция тромбоцитов в периферийной циркуляции (т.е. аутоиммунная тромбоцитопения или химио- и радиационная терапия). Кроме того, у пациентов, получающи х·большие количества быстроусвояемых, бедных на тромбоциты препаратов крови, тромбоцитопения может развиваться из-за разбавления крови. Клинические проявления тромбоцитопении зависят от степени болезни, ее причины и возможных сопутствующи х коагуляционных дефектов. В общем случае, пациенты с числом тромбоцитов между 20 и 100´109 на литр подвергаются риску чрезмерной пост-травматической потери крови, в то время, как пациенты с числом тромбоцитов ниже 20´109 на литр подвергаются риску самопроизвольной потери крови. Эти последние пациенты являются кандидатами на переливание тромбоцитов с сопутствующим риском получения иммунодефицита и вирусов. При любой степени тромбоцитопении потеря крови имеет тенденцию к ужесточению, когда причиной является не увеличение деструкции тромбоцитов, а снижение их образования. В первой ситуации ускоренное разрушение тромбоцитов приводит к циркуляции более молодых, больших по размеру и гемостатически более эффективных тромбоцитов. Тромбоцитопения может появляться от различных заболеваний, коротко описанных ниже. Более детальное описание можно найти у [Schafner, A.I., "Thrombocytopenia and Disorders of Platelet Function", Internal Medicine, 3rd Ed., John D.Hutton et al., Eds., Little Brown and Co., Boston/Toronto/London (1990)]. (а) Тромбоцитопения, обусловленная ухудшением образования тромбоцитов Причины врожденной тромбоцитопении включают органическую апластическую анемию (синдром Фанкони) и врожденную амегакариоцитную тромбоцитопению, которая может быть связана с пороком развития скелета. Развивающееся заболевание вырабатывания тромбоцитов обусловлено либо гипоплазией мегакариоцитов, либо неэффективным тромбопоэзом. Мегакариоцитная гипоплазия может быть результатом различных состояний, например, аплазии костного мозга (включая идиопатические формы или миелоугнетение при помощи химиоили радиотерапии), миелофиброза, лейкемии, инвазии костного мозга метастатической опухолью или гранулемами. В некоторых случая х токсины, инфекционные агенты или лекарства могут избирательно влиять на тромбопоэз; примеры включают временную тромбоцитопению, вызванную алкоголем и определенными вирусными инфекциями, а также легкую тромбоцитопению, связанную с приемом тиазидных мочегонных средств. Наконец, не эффективный тромбопоэз, являющийся следствием мегалобластических процессов (недостаточность фолатов или В12), также может вызвать тромбоцитопению, обычно при сопутствующей анемии и лейкопении. Текущее лечение тромбоцитопении, вызванной ухудшением образования тромбоцитов, зависит от идентификации и устранения причины расстройства костного мозга. Переливание тромбоцитов обычно применяется только для пациентов с серьезными осложнениями потери крови или для восстановления после потерь во время хирургических операций, поскольку изоиммунизация может привести к невосприимчивости к дальнейшим переливаниям. Потери из слизистых оболочек, вызванные тяжелой формой тромбоцитопении, могут быть восстановлены оральным или внутривенным введением антифибринолитических агентов. Тромбоцитная недостаточность может, однако, развиваться, если антифибринолитические агенты используются пациентом с рассеянной интраваскулярной коагуляцией (DIC). (b) Тромбоцитопения, обусловленная селезеночной изоляцией Спленомегалии, вызванной любыми причинами, может сопутствовать легкая или средняя тромбоцитопения. Это в основном пассивный процесс (гиперспленизм) селезеночной изоляции тромбоцитов, в отличие от активной деструкции тромбоцитов селезенкой в случаях иммуноопосредованной тромбоцитопении, описанйой ниже. Хотя самой распространенной причиной гиперспленизма является застойная спленомегалия из-за портальной гипертензии, обусловленной алкогольным циррозом, другие формы застойной, инфильтратной или лимфопролиферативной спленомегалии также связаны с тромбоцитопенией. Количество тромбоцитов в результате одного гиперспленизма чаще всего не падает ниже 50´10 на литр. (c) Тромбоцитопения, обусловленная неиммуно-опосредованной деструкцией тромбоцитов Тромбоцитопения может быть результатом ускоренной деструкции тромбоцитов вследствие различных неиммунологических процессов. Расстройства этого типа включают рассеянную интраваскулярную коагуляцию, протезные интраваскулярные устройства, экстракорпоральная циркуляция крови, тромботические микроангиопатии, такие как тромботическая тромбоцитная пурпура. Во всех эти х случаях циркулирующие тромбоциты, находящиеся близко к искусственным поверхностям или к аномальной васкулярной интиме, или поглощаются в этих местах, или повреждаются и затем удаляются ретикулоэндотелиальной системой. Болезненные состояния или расстройства, при которых рассеянная интраваскулярная коагуляция (DIC) может усиливаться, описаны в Braunwald et al. (eds), Harrison's Principles of Internal Medicine, 11th Ed., p.1478, McGraw Hill (1987). Интраваскулярные протезные устройства, включающие кардиальные клапаны и внутриаортные баллоны, могут вызывать от легкой до средней степени деструктивной тромбоцитопении и временной тромбоцитопении у пациентов с кардиолегочным искусственным кровообращением или гемодиализом из-за поглощения или повреждения тромбоцитов в экстракорпоральной системе. (d) Лекарственная иммунная тромбоцитопения При иммунологически опосредованной тромбоцитопении было исследовано более 100 лекарственных веществ. Однако хорошо себя зарекомендовали только хинидин, хинин, золото, сульфонамиды, цефалотин и гепарин. Лекарственная тромбоцитопения часто носит тяжелые формы и резко проявляется главным образом в течение тех дней, когда пациент принимает сенсибилизирующие медикаменты. (e) Иммунная (аутоиммунная) тромбоцитопеническая пурпура (ITP) ITP у взрослых представляет собой хроническое заболевание, характеризуемое аутоиммунной деструкцией тромбоцитов. Аутоантителами обычно являются igG, хотя другие иммуноглобулины тоже описаны. Хотя были найдены аутоантитела ITP, связанные с мембраной тромбоцитов GPII bIIIa, в большинстве случаев специфика антигенов тромбоцитов не идентифицирована. Экстраваскулярная деструкция сенсибилизированных тромбоцитов происходит в рекулоэндотелиальной системе селезенки и печени. Хотя более половины случаев ITP являются идиопатическими, многие пациенты страдают ревматическими или аутоиммунными заболеваниями (например, системная красная волчанка) или лимфопролиферативными расстройствами (хроническая лимфоцитарная лейкемия). (f) ВИЧ-обусловленная тромбоцитопения ITP является частой причиной ВИЧ-инфекции [Morris et al., Ann. Intern. Med., 96:714-717 (1982)], появляющейся на любой стадии болезни, как у пациентов с диагнозом синдрома приобретенного иммунодефицита (AIDS), с AIDS-связанным комплексом, так и у пациентов с ВИЧ-инфекцией, но без симптомов AIDS. ВИЧ-инфекция представляет собой передающуюся болезнь, характеризуемую глубоким дефицитом функции клеточного иммунитета и появлением условно-патогенных инфекций и злокачественных образований. Первичной иммунологической аномалией в результате инфекции ВИЧ является прогрессирующее истощение и функциональное ухудшение Т-лимфоцитов, выделяющих клеточный поверхностный гликопротеин CD4 [Lane et al., Ann. Rev Immunol., 3:477 (1985)]. Потеря CD4 Т-клетками, вероятно, вызывает глубокие нарушения клеточного и гуморального иммунитета, ведущие к возникновению условно-патогенных инфекций и злокачественных характеристик AIDS (Η. Lane выше). Хотя механизм ВИЧ-обусловленной ІТР не изучен, считается, что он отличен от механизма ITP, не связанной с ВИЧ-инфекцией. [Walsh et al., Ν.Eng. J.Med., 311:635-639 (1984); Ratner, Am. J. Med., 86:194-198 (1989)]. IV. Текущее лечение тромбоцитопении Тактика лечения тромбоцитопении диктуется тяжестью заболевания и срочностью клинической ситуации. Способы лечения ВИЧ-обусловленной и не-ВИЧ-обусловленной тромбоцитопении сходны, и хотя было испробовано много различных терапевтических подходов к лечению, терапия остается темой для дискуссий. Количество тромбоцитов у пациентов с диагнозом тромбоцитопения успешно увеличивается при применении глюкокортикоида (например, преднизолона), однако у большинства пациентов наблюдается неполный результат или возникновение рецидива при снижении дозы глюкокортикоида или прекращении его приема. Основываясь на исследованиях пациентов с ВИЧ-связанной тромбоцитопенией, некоторые ученые предположили, что глюкокортикоидная терапия может привести к предрасположенности к СПИДу. Глюкокортикоиды обычно назначают, когда число тромбоцитов падает ниже 20´109/литр или если происходит самопроизвольная потеря крови. Пациентам, невосприимчивым к глюкокортикоидам, может назначаться соединение формулы 4-(2-хлорфенил)-9-метил-2-[3-(4-морфолинил)-3-пропанон-1-ил]6Нтиено[3,2,f][1,2,4]триазоло[4,3,а][1,4]диазепин (WEB 2086) для успешного лечения тяжелых форм не-ВИЧ-обусловленной ITP. Пациенты с количеством тромбоцитов 37000-58000/мкл получали WEB 2086 и через 1-2 недели лечения количество тромбоцитов увеличивалось до 140000-190000/мкл. [ЕР 361,077 и Lohman et al., Lancet, 1147 (1988)]. Хотя оптимальное лечение амегакариоцитарной тромбоцитопенической пурпуры (ААТР) е ще не определено, применение антитимоцитоглобулина (ATG), лошадиной антисыворотки для ткани человеческой вилочковой железы, приводит к длительной полной ремиссии [Trimble et al., Am. J.Hematol., 37:126-127 (1991)]. Однако, последние отчеты показывают, что кроветворное действие ATG присуще тимеросалу, в котором, предположительно, белок выступает в качестве носителя серы [Panella et al., Cancer Research, 50:4429-4435 (1990)]. Хорошие результаты показывает спленэктомия. При этом удаляется основное место деструкции тромбоцитов и основной источник вырабатывания аутоантител у многих пациентов. Результатом этой процедуры является длительная ремиссия без лечения у большого количества пациентов. Однако, поскольку хирургические операции нежелательны для пациентов с иммунной недостаточностью, спленэктомия рекомендуется только в тяжелых случаях тромбоцитопении (например, при тяжелой форме ВИЧобусловленной ITP), для пациентов с неудовлетворительным результатом 2-3-недельного лечения глюкокортикоидом или рецидивом после прекращения приема глюкокортикоида. Имеющиеся исследовательские данные пока не дают представления о том, почему спленэктомия вызывает предрасположенность пациентов к СПИДу. Кроме преднизолоновой терапии и спленэктомии используются также некоторые цитотоксические агенты, например, винкристин и азидотимидин (AZT, зидовудин), которые показывают улучшение при лечении ВИЧобусловленно тромбоцитопении; однако, результаты являются еще предварительными. Из вышеизложенного следует вывод, что одним способом лечения тромбоцитопении является получение агента, способного ускорять видоизменение и развитие мегакариоцитов или их предшественников в тромбоцито-продуцирующую форму. В определении такого агента, обычно называемого "тромбопоэтин" (ТРО) была проделана значительная работа. Другие названия, часто используемые для ТРО в литературе, включают: агент стимуляции тромбоцитопоэза (TSF), мегакариоцито-колониестимулирующий агент (MK-CSF), мегакариоцитостимулирующий агент и потенцирующее средство для мегакариоцитов. Впервые действие ТРО было исследовано в 1959 году [Rak et al., Med. Exp., 1:125], и попытки выделить и очистить этот агент продолжаются до сегодняшнего дня. Существуют описания частичной очистки ТРО-активных полипептидов [см., например, Tayrien et al., J. Biol. Chem., 262:3262 [1987] and Hoffman et al., J. Clin. Invest. 75:1174 (1985)], другие утверждают, что ТРО не является отельной сущностью, а скорее представляет собой полифункциональную экспрессию известного гормона [IL-3, Sparrow et al., Prog Clin. Biol. Res., 215:123 (1986)]. Независимо от ее формы и природы, молекула, обладающая тромбопоэтической активностью, имеет огромное терапевтическое значение. Хотя ни один белок еще однозначно не идентифицирован как ТРО, несомненный интерес вызывает недавнее открытие, что mpl, предполагаемый цитокиновый рецептор, может служить передатчиком тромбопоэтического сигнала. V. Mpl - Мегакариоцитопоэтический цитокиновый рецептор Считается, что размножение и развитие кроветворных клеток строго регулируется агентами, которые положительно или отрицательно влияют на размножение и мультивидовое изменение плюрипотенциальных стволовых клеток. Это действие оказывается при посредстве связей высокого сродства внеклеточных белковых агентов со специфическими рецепторами клеточной поверхности. Эти рецепторы клеточной поверхности обладают большой степенью гомологичности и в целом классифицируются как члены сверхсемейства цитокиновых рецепторов. Члены сверхсемейства включают рецепторы для: IL-2 (с и g цепи) [Hatakeyama et al., Science, 257:379-382 (1991)], IL-3 [Itoh et al., Science, 247:324-328 (1990), Gorman et al., Proc Natl. Acad. Sci. USA, 87:5459-5463 (1990);. Kitamura et al., Cell, 66:1165-1174 (1991a); Kitamura Proc Natl. Acad. Sci. USA, 88:5082-5086 (1991b)]; IL-4 Mosleyetal., Cell, 59:335-348 (1989); IL-5 [Takaki et al., EMBO J., 9:43674374 (1990); Tavernier et al., Cell, 66:1175-1184 (1991)]; IL-6 [Yamasaki et al., Science, 241:825-828 (1988); Hibi et al., Cell, 63:1149-1157 (1990)]; IL-7 [Goodwin et al., Cell 60:941-951 (1990)], IL-9 [Renault et al., Proc Natl. Acad. Sci. USA, 89:5690-5694 (1992)]; гранулоцито-макрофаго-колониестимулирующий агент (GM-GSF) [Gearing et al., EMBO J., 8:3667-3676 (1991); Hayashida et al., Proc Natl. Acad. Sci. USA, 244:9655-9659 (1990)]; гранулоцито-колониестимулирующий агент (G-GSF) [Fukunaga et al., Cell 61:341-350 (1990a) Fukunaga et al., Proc Natl. Acad. Sci. USA, 87:8702-8706 (1990b); Larsen et al., D. Exp. Med., 172:1559-1570 (1990), ЕРО [D'Andrea et al., Cell, 57: 277-285 ([1989); Jones et al., Blood, 76:31-35 (1990)], агент ингибирования лейкемии (LIF) [Gearing et al., EMBO J., 10:2839-2848 (1991), онкостатин Μ (OSM) [Rose et al., Proc Natl. Acad. Sci. USA, 88:8641-8645 (1991), а также рецепторы для пролактина [Boutin et al., Proc Natl. Acad. Sci. USA, 88:7744-7748 (1988); Edery et al., Proc Natl. Acad. Sci. USA, 86:2112-2116 (1989)], гормон роста (GH) [Leung et al., Nature, 330:537-543 (1987)] и ресничный нейротрофический агент (CNTF) [Davis et al., Science, 253:59-63 (1991)]. Члены сверхсемейства цитокиновых рецепторов можно сгруппировать в три функциональные категории [обзор см. Nicola et al., Cell, 67:1-4 (1991)]. Первый класс включает рецепторы, состоящие из одной цепи, такие как рецептор эритропоэтина (EPO-R) или рецептор гранулоцито-колониестимулирующего агента (GCSF-R), которые связывают лиганды высокого сродства при посредстве внеклеточных доменов и генерируют внутриклеточный сигнал. Второй класс рецепторов, так называемых р-подэлементов, включает рецептор интерлейкина-6 (IL6-R), рецептор гранулоцито-макрофаго-колониестимулирующего агента (GM-GSF-R) рецептор интерлейкина-3 (IL3-Rp) и другие члены сверхсемейства, цитокиновых рецепторов. Эти рподэлементы связывают лиганды низкого сродства, но не генерируют вн утриклеточного сигнала. Рецептор с высоким сродством, обладающий способностью сигнализации, генерируется гетеродимером между рподэлементом и членом третьего класса цитокиновых рецепторов, называемых с-подэлементами, например, сс, общий с-подэлемент для трех р-подэлементов IL3-Rp и GM-CSF-R. То, что mpl является членом сверхсемейства цитокиновых рецепторов, очевидно из гомологии последовательности оснований ДНК [Gearing et al., ΕΜΒΟ J., 8:3667-3676 (1988); Bazan, Proc Natl. Acad. Sci. USA, 87:6834-6938 (1990)] [Davis et al., Science, 253:59-63 (1991), Vigon et al., Proc Natl. Acad. Sci. USA, 89:5640-5644 (1992)] и его способности переносить пролиферативные сигналы. Белковая цепь, выведенная при молекулярном клонировании c-mpl мыши, оказалась гомологичной другим цитокиновым рецепторам. Внеклеточный домен содержит 465 аминокислотных остатков и состоит из двух поддоменов, каждый с четырьмя высококонсервированными цистеинами и специфической цепью в поддомене с амино-окончанием и в поддомене с карбокси-окончанием. Лиганд-связующие внеклеточные домены предположительно имеют сходную двойную с-ствольную складчатую геометрическую структур у. Этот двойной внеклеточный домен высокогомологичен сигналопередающей цепи, общей для рецепторов IL-3, IL-5 и GM-CSF, а также связующему домену низкого сродства LIF (Vigon et al., Oncogene, 8:2607-2615 [1993]). Таким образом, mpl могут принадлежать к классу цитокиновых рецепторов, связующи х лиганды низкого сродства. Сравнение мышиного mpl и зрелого человеческого mpl P обнаружило, что последовательность оснований ДНК этих двух белков на 81% идентична. Более точный анализ показал, что последовательности внеклеточных доменов с N-окончаниями и С-окончаниями идентичны на 75% и 80% соответственно. Наиболее законсервированная область mpl представляет собой цитоплазматический домен, показывающий 91% идентичности аминокислот, с последовательностью 37 остатков около трансмембранного домена, идентичных в обоих видах. В соотве тствии с этим, mpl описывается как один из наиболее законсервированных членов свер хсемейства цитокиновых рецепторов (Vigon выше). То, что mpl является функциональным рецептором, способным передавать пролиферативный сигнал, очевидно из строения химерических рецепторов, включающих внеклеточный домен из цитокинового рецептора, имеющего высокое сродство к известному цитокину с цитоплазматическим доменом mpl. Поскольку до сих пор не описано ни одного известного лиганда для mpl, было необходимо построить химерический лиганд высокого сродства, связующий внеклеточный домен из таких цитокиновых рецепторов, как IL-4R или G-CSFR. Vigon et al., выше слил внеклеточный домен G-CSFR с трансмембранным и цитоплазматическим доменами c-mpl. IL-3-зависимую клеточную линию, BAF/B03 (Ba/F3), подвергли трансфекции химерой G-CSFR/mpl наряду с контролем полной длины G-CSFR. Клетки, зараженные химерой, росли одинаково хорошо в присутствии цитокина IL-3 и G-CSF. Подобным образом, клетки, зараженные GCSFR, также хорошо росли в присутствии IL-3 и G-CSF. Все клетки погибали в отсутствие агентов роста. Подобный опыт был проведен Skoda et al., ΕΜΒΟ J., 12(7):2645-2653 (1993), в котором внеклеточный и трансмембранный домены человеческого рецептора IL-4 (hIL-4-R) были слиты с цитоплазматическим доменом mpl мыши и этой химерой была заражена IL-3-зависимая клеточная линия Ba/F3 мыши. Клетки Ba/F3, зараженные обычным hIL-4-R, размножались нормально в присутствии как IL-4, так и IL-3. Клетки Ba/F3, зараженные hIL-4-R/mpl, размножались нормально в присутствии hIL-4 (независимо от присутствия или отсутствия IL-3), показав, что в клетках Ba/F3 цитоплазматический домен mpl содержит все элементы, необходимые для передачи пролиферативного сигнала. Эти химерические эксперименты продемонстрировали способность цитоплазматического домена mpl к пролиферативной сигнализации, но не показали, может ли внеклеточный домен mpl связывать лиганды. Эти результаты допускают как минимум две возможности, а именно, что mpl является рецептором с одной цепью (класса 1), подобно EPO-R или G-CSFR, или он является сигналопередающим с-подэлементом (класс 3), требующим р-подэлемента типа IL-3 (Skoda et al. выше). VI. Лиганд mpl - тромбопоэтин (ТРО) Как описано выше, было сделано предположение, что, сыворотка содержит единственный в своем роде агент, называемый тромбопоэтином (ТРО), который действуе т синергически с различными другими цитокинами, ускоряя рост и развитие мегакариоцитов. Ни одного такого природного агента до сих пор не было выделено из сыворотки или другого источника, хотя предпринималось большое количество попыток разными группами ученых. Даже если до сих пор неизвестно, способен ли mpl непосредственно связываться с агентом стимуляции мегакариоцитов, последние опыты показали, что mpl участвует в переносе пролиферативного сигнала от агента или агентов, найденных в сыворотке пациентов с апластическим костным мозгом (Met'hia et al., Blood, 82(5):1395-1401 [1993]). То, что единственный сывороточный колониеобразующий агент, отличный от Il-lp, IL-3, IL-4, Il-6, IL-11, SCF, ЕРО, G-CSF и GM-CSF, переносит пролиферативный сигнал через mpl, очевидно из исследования распределения c-mpl в простейших и промежуточных кроветворных клеточных линиях и из исследований обратного mpl в одной из этих клеточных линий. Используя обратную транскриптазу (RT)-PCR в иммуно-очищенных человеческих кроветворных клетках, Methia et al., выше, продемонстрировал, что сильные сигналы mpl mRNA были найдены только в очищенных клетках CD34+, мегакариоцитах и тромбоцитах. Клетки CD34+, очищенные от костного мозга (ВМ), составляют около 1% от всех клеток ВМ и обогащаются в присутствии простейших и промежуточных прародительских клеток всех видов (т.е., эритроидов, грануломакрофагов и мегакариоцитов). Обратные олигодезоксинуклеотиды mpl, как было показано, подавляют образование мегакариоцитарных колоний от плюрипотенциальных клеток CD34+, культивированных в сыворотке от пациентов с аплазией костного мозга [богатый источник мегакариоцито-колониестимулирующей активности (MK-CSA)]. Те же самые обратные олигодезоксинуклеотиды не оказывают никакого влияния на образование колоний эритроидов или грануломакрофагов. Связываются ли непосредственно mpl с лигандами, и способствует ли мегакариоцитопоэзу указанный агент сыворотки при посредстве mpl, было до сих·пор неизвестно. Однако было сделано предположение, что, если mpl действительно непосредственно связывается с лигандами, та его аминокислотная последовательность должна быть высококонсервированной и иметь перекрестную реакционноспособность с различными видами благодаря большой степени идентичности последовательностей во внеклеточных доменах mpl человека и мыши [Vigon et al., выше (1993)]. VII. Объекты изобретения В свете вышеизложенного можно сделать выводы, что в данной области существует текущая необходимость выделить и идентифицировать молекулы, способные, стимулировать размножение, видоизменение и развитие кроветворных клеток, особенно мегакариоцитов или их предшественников, для терапевтического использования при лечении тромбоцитопении. Считается, что такой молекулой является лиганд mpl, и, таким образом, существуе т следующая необходимость - выделить такой лиганд (лиганды) для оценки его роли в клеточном росте и видоизменении. В соответствии с этим, объектом данного изобретения является получение фармацевтически чистой молекулы, способной стимулировать размножение, видоизменение и/или развитие мегакариоцитов в зрелую тромбоцито-продуцирующую форму. Вторым объектом является производство молекулы в форме, пригодной для терапевтического использования при лечении расстройств кроветворения, в частности, тромбоцитопении. Следующим объектом настоящего изобретения является получение молекул нуклеиновой кислоты, кодирующей лиганды такого белка и использование этих молекул нуклеиновой кислоты для производства связующи х лигандов mpl в рекомбинантной клеточной культуре для диагностики) и терапевтического использования. Еще одним объектом является получение производных и модифицированных форм лигандов белка, включающих варьируемые последовательности аминокислот, варьируемые гликопротеиновые формы и их ковалентные производные. Дополнительным объектом является получение варьируемых полипептидных форм, сочетающи х лиганды mpl с остатками аминокислот и заместителями, из последовательности ЕРО для производства белка, способного регулировать размножение и рост прародительских клеток как тромбоцитов, так и красных кровяных телец. Еще одним дополнительным объектом является получение иммуногенов для увеличения количества антител против лигандов mpl, или их слитых форм, а также для получения антител, способных связывать такие лиганды. Эти и другие объекты изобретения будут понятны после рассмотрения детального описания в целом. Объекты изобретения достигаются путем получения изолированного белка7 способствующего размножению и развитию мегакариоцитопоэтических клеток млекопитающих, называемого "лиганд mpl" (ML) или "тромбопоэтин" (ТРО), способного стимулировать размножение, видоизменение и/или развитие мегакариоцитов в зрелую тромбоцито-продуцирующую форму. Этот в основном гомогенный белок может быть в чистом виде выделен из натурального источника способом, включающим (1) контактирование плазмы источника, содержащей молекулы лиганда mpl, которые должны быть очищены, с иммобилизованным рецепторным полипептидом, в частности, полипептидом mpl или полипептидом слитого mpl, иммобилизованным на подложке, в условиях, при которых молекулы лиганда mpl, которые должны быть очищены, селективно адсорбируются на иммобилизованном рецепторном полипептиде, (2) промывку иммобилизованного рецепторного полипептида и его подложки для удаления неадсорбированного материала и (3) элюирование молекул лиганда mpl из иммобилизованного рецепторного полипептида, на котором они адсорбированы, элюирующим буферным раствором. Предпочтительным натуральным источником является плазма или моча млекопитающего, содержащая лиганд mpl. Как вариант, млекопитающее может страдать аплазией, и иммобилизованный рецептор представляет собой слияние mplIgG. Как вариант, белок, способствующий размножению и развитию мегакариоцитопоэтических клеток, представляет собой изолированный, в основном гомогенный полипептид лиганда mpl, полученный синтетическими или рекомбинантными средствами. Полипептид "лиганда mpl" или "ТРО" данного изобретения предпочтительно имеет как минимум 70% идентичности всей последовательности ДНК с последовательностью аминокислот в высокоочищенном, в основном гомогенном полипептиде лиганда mpl свиньи и как минимум 80% идентичности последовательности ДНК с "ЕРО-доменом" полипептида лиганда mpl свиньи. Как вариант, лиганд mpl данного изобретения представляет собой зрелый человеческий лиганд mpl (hML), имеющий зрелую аминокислотную последовательность, представленную на Фиг.1 (номер последовательности: 1), или один из его вариантов или посттранскрипционных модифицированных форм, или белок, имеющий 80% идентичности с последовательностью зрелого человеческого лиганда mpl. Возможно, вариант лиганда mpl представляет собой фрагмент, в частности, амино-конечный или фрагмент "ЕРО-домена" зрелого человеческого лиганда mpl. Предпочтительно амино-конечный фрагмент сохраняет практически всю последовательность человеческого ML между первым и четвертым цистеиновыми остатками, но может содержать существенные дополнения, делеции или заместители снаружи этого участка. Согласно этому варианту осуществления изобретения, фрагментарный полипептид может быть представлен формулой X-h ML(7-151)-Y, где hML(7-151) представляет собой аминокислотную последовательность человеческого ТРО (hML) от Cys 7 до Cys 151 включительно; X представляет собой аминогруппу Cys 7 или один или более амино-конечный остаток аминокислоты зрелого hML, или добавление аминоккислотного остатка, такого как Met, Туr, или ведущие последовательности, содержащие, например, участки протеолитического расщепления (фактор Ха или тромбин); Υ представляет собой карбоксильную конечную группу C ys 151 или один или более карбоксиконечный остаток аминокислоты зрелого hML, или добавление аминокислотного остатка. Как вариант, полипептид лиганда mpl или его фрагмент может быть слитым в гетерологичный полипептид (химеру). Предпочтительный гетерологичный полипептид представляет собой цитокин, колониестимулирующий агент или интерлейкин или их фрагменты, в частности, кит-лиганд (KL), IL-1, Il-3, IL-4; Il-6, IL-11, SCF, ЕРО, GM-CSF или LIF. Как вариант, предпочтительным гетерологичным полипептидом может быть иммуноглобулиновая цепь, в частности, человеческий IG1, IG2, IG3, IG4, IGA, IGE, IGD, IGM - или их фрагменты, в частности, содержащие постоянные домены жесткой цепи IgG. Другой аспект данного изобретения представляет композицию, включающую изолированный агонист mpl, биологически активный и предпочтительно способный стимулировать включение меченых нуклеотидов (например, 3Н-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, зараженных человеческим mpl. Как вариант, агонистом mpl является биологически активный лиганд mpl, предпочтительно способный стимулировать включение 35S в циркулирующие тромбоциты в анализе мышиных ответных тромбоцитов. Подходящие агонисты mpl включают hML153, h ML(R153A, R154A), h ML2, hML3, h ML4, mML, mML2, mML3, p ML и pML2 или их фрагменты. В другом варианте изобретение предлагает изолированное антитело, способное связываться с лигандами mpl. Изолированное антитело, способное связываться с лигандами mpl, может, как вариант, быть слито со вторым полипептидом, и антитело или его слитая форма может быть использовано для выделения лиганда mpl в чистом виде из источника, как описано выше для иммобилизованного mpl. В дальнейшем развитии этого варианта изобретение предлагает способ обнаружения лиганда mpl вне организма или в организме, включающий контактирование антитела с образцом, в частности, образцом сыворотки, подозреваемой в содержании лиганда, и обнаружение его в том случае, если связывание произойдет. В следующем варианте изобретение предлагает изолированную молекулу н уклеиновой кислоты, кодирующей лиганд mpl или его фрагменты, которая может быть, как вариант, помечена обнаруживаемым элементом, и молекулу нуклеиновой кислоты, имеющую последовательность оснований, комплементарную или гибридизирующуюся в определенном диапазоне условий (от умеренно до высоко строгих) с молекулой нуклеиновой кислоты, имеющей последовательность оснований, кодирующую лиганд mpl. Предпочтительными являются молекулы нуклеиновой кислоты, кодирующие человеческие, свиные и мышиные лиганды mpl и включающие РНК и ДНК с геномами и кДНК. В дальнейшем развитии этого варианта изобретение предлагает молекулу н уклеиновой кислоты, представляющую собой ДНК, кодирующую лиганд mpl и включающую повторяемый вектор, в котором ДНК оперативно связана с контрольными последовательностями, распознаваемыми хозяином, трансформируемым вектором. Как вариант, ДНК представляет собой кДНК с последовательностью оснований, представленной на Фиг.1: 5'-3' (номер последовательности: 2), 3'-5' или ее фрагменты. Далее этот аспект включает клетки хозяина, предпочтительно клетки СНО, трансформированные вектором, и способ использования ДНК для влияния на продуцирование лиганда mpl, предпочтительно включающий экспрессию кДНК, кодирующей лиганд mpl, в культуре трансформированных клеток хозяина и извлечение лиганда mpl из клеток хозяина или среды культуры клеток хозяина. Лиганд mpl, приготовленный таким образом, предпочтительно представляет собой человеческий лиганд mpl. Далее изобретение включает способ лечения млекопитающих, страдающи х расстройством кроветворения, в частности, тромбоцитопенией, предусматривающий введение млекопитающему терапевтически эффективного количества лиганда mpl. Возможно введение лиганда mpl в сочетании с цитокином, в частности, колониестимулирующим агентом или интерлейкином. Предпочтительные колониестймулирующие агенты или интерлейкины включают кит-лиганд (KL), LIF, G-CSF, GM-CSF, M-CSF, ЕРО, IL-1, IL-3, IL-6, IL-11. Далее изобретение включает процесс выделения в чистом виде ТРО (ML) из ТРО-продуцирующего микроорганизма, включающий: (1) разрушение или лизирование-клеток, содержащих ТРО, (2) возможно, отделение растворимого материала от нерастворимого материала, содержащего ТРО, (3) солюбилизация ТРО в нерастворимом материале солюбилизирующим буфером, (4) отделение солюбилизированного ТРО от др уги х растворимых и нерастворимых материалов, (5) укладка ТРО в окислительно-восстановительном буфере и (6) отделение сложенного ТРО от развернутого ТРО. Этот процесс разработан для солюбилизации нерастворимого материала, содержащего ТРО, хаотропическим агентом, где хаотропический агент выбирается из соли гуанидина, тиоцианата натрия или мочевины. Далее процесс предусматривает отделение солюбилизированного ТРО от других растворимых и нерастворимых материалов в один или несколько этапов, выбранных из центрифугирования, гельфильтрации и хроматографии обратной фазы. Этап укладки ТРО проводится в окислительновосстановительном буфере, содержащем как окислительный, так и восстановительный агенты. Как правило, окислителем является кислород или соединение, содержащее минимум одну дисульфидную связь, а восстановителем - соединение, содержащее минимум один свободный сероводород. Предпочтительно окислитель выбирается из окисленного глютатиона (GSSG) и цистина, а восстановитель - из восстановленного глютатиона (GSH) и цистеина. Наиболее предпочтителен вариант, когда окислителем является GSSG, а восстановителем - GSH. Предпочтительно также, чтобы молярная концентрация окислителя была равна или превосходила молярную концентрацию восстановителя. Окислительновосстановительный буфер дополнительно содержит детергент, предпочтительно выбранный из CHAPS или CHAPSO, в количестве как минимум 1%. Окислительно-восстановительный буфер дополнительно содержит хлорид натрия, предпочтительно с концентрацией от 0,1 до 0,5М, и глицерин, предпочтительно с концентрацией более 15%. рН буфера предпочтительно лежит в диапазоне от 7,5 до 9,0, и этап укладки проводится при 4 градусах в течение 12-48 часов. Этот этап обеспечивает получение биологически активного ТРО, в котором между Cys, ближайшим к конечному амину домена ЕРО, и Cys, ближайшим к конечному карбоксилу, образуется дисульфидная связь. Далее изобретение включает процесс выделения в чистом виде биологически активного ТРО из микроорганизма, включающий: (1) лизирование как минимум внеклеточной мембраны микроорганизма, (2) обработка лизата, содержащего ТРО, хаотропическим агентом, (3) укладка ТРО и (4) отделение неочищенного ТРО и развернутого ТРО от сложенного ТРО. Фиг.1 показывает выведенную последовательность аминокислот (номер последовательности: 1) в кДНК человеческого лиганда mpl (hML) и кодированную последовательность нуклеотидов (номер последовательности: 2). Нуклеотиды пронумерованы в начале каждой строки. Нетранслируемые участки 5'и 3'обозначены прописными буквами. Аминокислотные остатки пронумерованы над последовательностью, начиная от последовательности Ser 1 зрелого белка лиганда mpl (ML). Границы предполагаемого экзона 3 обозначены стрелками, а потенциальные участки N-гликозилирования обведены рамками. Цистеиновые остатки обозначены точками над последовательностью. Подчеркнутые последовательности соответствуют Nконечным последовательностям, найденным в лиганде mpl, выделенном из плазмы свиньи. Фиг.2 показывает процедуру, используемую для инкорпорации в лиганд mpl 3Н-тимидина. Чтобы определить присутствие лиганда mpl из различных источников, клетки mpl Ρ Ba/F3 были лишены IL-3 в течение 24 часов в увлажненном инкубаторе при 37°С и 5% СО2 в воздухе. После IL-3-голодания клетки распределили в 96-луночных чашках с разбавленными образцами или без них, и подвергли культивированию 24 часа в инкубаторе клеточных культур. Затем добавили 20мкл среды RPMI, не содержащей сыворотки и содержащей 1мкКи 3Н-тимидина, в каждую лунку на последние 6-8 часов. Затем клетки собрали в 96луночные фильтровальные чашки и промыли водой. Фильтры сосчитали. Фиг.3 показывает действие проназы, DTT и тепла на способность АРР стимулировать размножение клеток Ba/F3-mpl. При изучении действия проназы добавляли проназу (Boehringer Mannheim) или альбумин бычьей сыворотки к Affi-gel10 (Biorad) и выдерживали отдельно с АРР в течение 18 часов при 37°C. Затем удалили смолы путем центрифугирования и исследовали надосадочную жидкость. АРР также нагревали до 80°C в течение 4 минут или обрабатывали 100мкМ DTT с последующим диализом в PBS. Фиг.4 показывает элюирование активности лиганда mpl в колонках Phenyl-Toyopearl, Blue-Sepharose и Ultralink-mpl. Фракции 4-8, элюированные из колонки mpl-сродства, были пиковыми. Фиг.5 показывает SDS-PAGE-элюирование фракций Ultralink-mpl. К 200мкл каждой фракции.2-8 добавили 1мл ацетона, содержащего 1мМ НСl при -20°С. Через 3ч. при -20°С образцы центрифугировали, а результирующие гранулы дважды промыли ацетоном при -20°С. Затем ацетоновые гранулы растворили в 30мкл SDS-солюбилизирующего буфера, обработали 100мкМ DTT и нагрели при 90°С в течение 5 минут. После этого образцы растворили в 4-20% геле SDS-полиакриламида и визуализировали белок при помощи серебряного окрашивания. Фиг.6 показывает элюирование активности лиганда mpl в SDS-PAGE. Фракцию 6 из колонки mpl-сродства растворили в 4-20% геле SDS-полиакриламида в невосстанавливающих условиях. После электрофореза гель разрезали на 12 равных частей и электроэлюировали, как описано в примерах. Электроэлюированные образцы подвергли диализу в PBS и анализу при разбавлении 1/20. Стандартом Mr при калибровке геля был стандарт Novex Mark 12. Фиг.7 показывает влияние АРР, лишенной лиганда mpl, на мегакариоцитопоэз человека. АРР, лишенная лиганда mpl, была приготовлена путем прохождения 1мл через 1-мл колонку mpl-сродства (700мкг mpl-IgG/мл NHS-superose, Pharmacia). Культуры человеческих периферических стволовы х клеток обработали 10% АРР или 10% АРР, ли шенной лиганда mpl, и культивировали 12 дней. Мегакариоцитопоэз проанализировали количественно, как описано в примерах. Фиг.8 показывает влияние mpl-IgG на стимулирование человеческого мегакариоцитопоэза при помощи АРР. Культуры человеческих периферических стволовых клеток обработали 10% АРР и культивировали 12 дней. В 0, 2 и 4-й дни добавляли mpl-IgG (0,5мкг) или ANP-R-IgG (0,5мкг). Мегакариоцитопоэз через 12 дней проанализировали количественно, как описано в примерах. Графики построены по средним значениям параллельных образцов, а в скобках приведены их действительные значения. Фиг.9 показывает обе цепи фрагмента человеческой геномной ДНК (390 bр [нуклеотидных пар]), кодирующей лиганд mpl. Показаны выведенная аминокислотная последовательность "экзона 3" (номер последовательности: 3), кодирующая последовательность (номер последовательности: 4) и ее комплемент (номер последовательности: 5). Фиг.10 показывает выведенную аминокислотную последовательность зрелого человеческого лиганда mpl (hML) (номер последовательности: 6) и зрелого человеческого эритропоэтина (номер последовательности: 7). Предсказанная аминокислотная последовательность для человеческого лиганда mpl совмещена с последовательностью человеческого эритропоэтина. Идентичные аминокислоты обведены рамками, а пустоты, введенные для оптимального совмещения, показаны пунктиром. Потенциальные участки Nгликозилирования подчеркнуты сплошной линией для hML и штриховой линией для hEPO. Два цистеина, важных для активности эритропоэтина, отмечены большими точками. Фиг.11 показывает выведенную аминокислотную последовательность изоформ зрелого человеческого лиганда mpl: hML (номер последовательности: 6), hML2 (номер последовательности: 8), hML3 (номер последовательности: 9) и hML4 (номер последовательности: 10). Идентичные аминокислоты обведены рамками, а пустоты, введенные для оптимального совмещения, показаны пунктиром. Фиг.12А, 12В и 12С показывают влияние человеческого лиганда mpl на размножение клетки Ba/F3-mpl вне организма (А). Мегакариоцитопоэз количественно оценили с использованием радиоактивного изотопа в моноклональном антителе IgG мыши, специфическом для гликопротеина GPIIbIII a (В) мегакариоцита, а тромбоцитопоэз мыши измеряли путем анализа ответных тромбоцитов. Клетки 293 были подвергнуты трансфекции методом СаРО4 [Gorman, С in DNA Cloning: A New Approach 2:143-190 (1985)] вектором pRK5 отдельно, pRK5-hML или pRK5-ML153 в течение ночи (pRK5-ML153 генерировали введением терминирующего кодона после остатка 153 в hML при помощи PCR). Затем кондиционировали среду в течение 36 часов и исследовали на стимуляцию размножения клеток Ba/F3-mpl, как описано в Примере 1 (А) или человеческий мегакариоцитопоэз вне организма (В). Мегакариоцитопоэз количественно оценили с использованием радиоактивного изотопа 125l в моноклональном антителе IgG (HP11D) мыши, специфическом для гликопротеина GPIIbIIla (В) мегакариоцита, как описано (Grant et al., Blood 69:1334-1339 [1987]). Действие частично очищенного рекомбинантного ML (rML) на вырабатывание тромбоцитов в организме (С) определяли с использованием анализа ответных тромбоцитов, описанного McDonald, Т.P. Proc. Soc. Exp. Biol, Med. 144:1006-10012 (1973). Частично очищенный rML приготовили из 200мл кондиционированной среды, содержащей рекомбинантный ML. Среду пропустили через 2-мл колонку Blue-Separose, уравновешенную в PBS, а колонку промыли PBS и элюировали PBS, содержащим по 2М мочевины и NaCl. Активную фракцию подвергли диализу в PBS и обработали 1мг/мл BSA без эндотоксина, образцы содержали менее одной единицы эндотоксина/мл. Мышам ввели 64000, 32000 или 16000 единиц rML или одного наполнителя. Каждая группа состояла из шести мышей. Полученные значения и стандартные отклонения для каждой группы показаны на рисунке. Величины ρ определяли по средним значениям двух сравнительных Т-тестов. Фиг.13 сравнивает действие изоформ и вариантов человеческого лиганда mpl на размножение клеток Ba/F3. Были исследованы hML, ложный опыт, hML2, hML3, hML(R15ЗА, R154A) при различном титре, как описано в Примере 1. Фиг.14А, 14В и 14С показывают выведенную аминокислотную последовательность (номер последовательности: 1) человеческого лиганда mpl (hML) или человеческого ТРО (hTPO) и кодирующую последовательность человеческой геномной ДНК (номер последовательности: 11). Нуклеотиды и аминокислотные остатки пронумерованы в начале каждой строки. Фиг.15 показывает SDS-PAGE очи щенного 293-rhML332 и очи щенного 293-rhML153 . Фиг.16 показывает последовательность нуклеотидов: кДНК (номер последовательности: 12) и выведенная аминокислотная последовательность (номер последовательности: 13) открытой рамки считывания изоформы ML мыши. Эта зрелая изоформа лиганда mpl мыши содержит 331 аминокислотный остаток, на 4 остатка меньше, чем в предполагаемом mML полной длины, и поэтому обозначена mML2. Нуклеотиды пронумерованы в начале каждой строки. Аминокислотные остатки пронумерованы над последовательностью, начиная с Ser 1. Потенциальные участки N-гликозилирования подчеркнуты. Цистеиновые остатки отмечены точками над последовательностью. Фиг.17 показывает последовательность кДНК (номер последовательности: 14) и предсказанную белковую последовательность (номер последовательности: 15) этой изоформы ML мыши. Нуклеотиды пронумерованы в начале каждой строки. Аминокислотные остатки пронумерованы над последовательностью, начиная с Ser 1. Эта зрелая изоформа лиганда mpl мыши содержит 335 аминокислотных остатков и считается лигандом mpl полной длины, поэтому обозначена mML. Последовательность сигналов отмечена пунктирным подчеркиванием, а возможная точка расщепления указана стрелкой. Нетранслированные участки 5' и 3' обозначены прописными буквами. Две обнаруженные делеции, являющиеся результатом попеременного сращивания (mML2 и mML3), подчеркнуты. Четыре цистеиновых остатка отмечены точками. Семь потенциальных участков N-гликозилирования обведены рамками. Фиг.18 сравнивает выведенную аминокислотную последовательность изоформы человеческого ML hML3 (номер последовательности: 9) и изоформу ML мыши, обозначенную mML3 (номер последовательности: 16). Предсказанная аминокислотная последовательность для человеческого лиганда mpl совмещена с последовательностью лиганда mpl мыши. Идентичные аминокислоты обведены рамками, а пустоты, введенные для оптимального совмещения, показаны пунктиром. Аминокислотные остатки пронумерованы в начале каждой строки. Фиг.19 сравнивает выведенные аминокислотные последовательности зрелых изоформ мышиного ML (номер последовательности: 17), свиного ML (номер последовательности: 18) и человеческого ML (номер последовательности: 6). изоформу ML мыши, обозначенную mML3 (номер последовательности: 16). Аминокислотные последовательности совмещены с наличием пустот, введенных для оптимального совмещения и обозначенных пунктиром. Аминокислоты пронумерованы в начале каждой строки, а идентичные остатки обведены рамками. Потенциальные участки N-гликозилирования обведены рамками с тенью. Цистеиновые остатки отмечены точками. Заокнсервированные диосновные аминокислотные раппорты, представляющие потенциальные участки расщепления протеазы, подчеркнуты. Четыре обнаруженные во всех трех видах последовательностей аминокислотные делеции (ML2) обведены жирными рамками. Фиг.20 показывает последовательность кДНК (номер последовательности: 19) и предсказанную последовательность зрелого белка (номер последовательности: 18) изоформы свиного ML (pML). Эта зрелая изоформа свиного лиганда mpl содержит 332 аминокислотных остатка и считается свиным лигандом mpl полной длины, обозначаемым pML. Н уклеотиды пронумерованы в начале каждой строки. Аминокислотные остатки .пронумерованы над последовательностью, начиная с Ser 1. Фиг.21 показывает последовательность кДНК (номер последовательности: 20) и предсказанную последовательность зрелого белка (номер последовательности: 21) изоформы свиного ML (pML2). Эта зрелая изоформа свиного лиганда mpl содержит 328 аминокислотных остатков и представляет собой форму свиного лиганда mpl с четырьмя делециями, обозначенную pML2. Нуклеотиды пронумерованы в начале каждой строки. Аминокислотные остатки пронумерованы над, последовательностью, начиная с Ser 1. Фиг.22 сравнивает выведенную аминокислотную последовательность изоформы свиного ML полной длины pML (номер последовательности: 18) и изоформу свиного ML, обозначенную pML2 (номер последовательности: 21). Предсказанная аминокислотная, последовательность для pML совмещена с последовательностью pML2. Идентичные аминокислоты обведены рамками, а пустоты, введенные для оптимального совмещения, показаны пунктиром. Аминокислотные остатки пронумерованы в начале каждой строки. Фиг.23 показывает особенности pSV15.ID.LL.MLORF плазмида ("полной длины", или ТРО332), используемого для трансфекции хозяйских клеток CHO-DP12 с целью получения CHO-rhTPO332. Фиг.24 показывает особенности pSV15.ID.LL.MLEPO-D плазмида ("усеченного", или ТРО153), используемого для трансфекции хозяйских клеток CHO-DP12 с целью получения СНО-гhТРО153 . Фиг.25А, 25В, 25С показывают влияние Е. coli-rhTPO (Met-1,153) на тромбоциты (А), красные кровяные тельца (В) и белые кровяные тельца (С) нормальной мыши. Двум группам по 6 самок мышей С57 В6 вводили ежедневно буфер PBS или 0,3мкг Е. coli-rhTPO(Met-1,153) (100мкл sc.). В 0 и 3-7 дни делали отбор 40мкл крови из глазничной вены. Эту кровь немедленно разбавили 10мл промышленного разбавителя и провели полный анализ на Serrono Baker Hematology Analyzer 9018. Данные представлены в виде полученных величин +/- стандартная ошибка. Фиг.26А, 26В, 26С показывают влияние Е. coli-rhTPO(Met-1,153) на тромбоциты (А), красные кровяные тельца (В) и белые кровяные тельца (С) сублетально облученной мыши. Две группы по 10 самок мышей С57 В6 сублетально облучили 750сГр гамма-излучения от источника 137Cs и вводили ежедневно буфер PBS или 3,0мкг Е. coli-rhTPO (Met-1,153) (100мкл sc). В нулевой день и в последующие промежуточные моменты времени делали отбор 40мкл крови из глазничной вены. Эту кровь немедленно разбавили 10мл промышленного разбавителя и провели полный анализ на Serrono Baker Hematology Analyzer 9018. Данные представлены в виде полученных величин +/- стандартная ошибка. Фиг.27А, 27В, 27С показывают влияние CHO-rhTPO332 на тромбоциты (А), красные кровяные тельца (эритроциты) (В) и белые кровяные тельца (лейкоциты) (С) нормальной мыши. Двум гр уппам по 6 самок мышей С57 В6 вводили ежедневно буфер PBS или 0,3мкг СНО-rhТРО332 (100мкл sc). В 0 и 3-7 дни делали отбор 40мкл крови из глазничной вены. Эту кровь немедленно разбавили 10мл промышленного разбавителя и провели полный анализ на Serrono Baker Hematology Analyzer 9018. Данные представлены в виде полученных величин +/- стандартная ошибка. Фиг.28 показывает кривые отклика на дозу для различных форм rhTPO, полученных из· различных клеточных линий. Кривые отклика на дозу были построены для rhTPO из следующи х клеточных линий: hTPO332 из CHO (полной длины, из клеток яичника китайского хомяка); hTPO(Met-1 ,153) (усеченная форма Е. соlі-производного с N-конечным метионином); hTPO332 (TPO полной длины, из человеческих клеток 293); Metless 155 E-coli (усеченная форма [rhTPO155] без конечного метионина из Е. coli). Группам из 6 самок мышей С57 В6 в течение 7 дней ежедневно вводили различные rhTPO в зависимости от группы. Каждый день делали отбор 40мкл крови из глазничной вены для полного анализа. Представленные данные показывают максимальные эффекты, наблюдаемые в различных гр уппах, и, за исключением (met 153 E-coli), эти эффекты наблюдались на 7-й день обработки. В вышеупомянутой группе (met 153 E-coli), максимальный эффект наступил на 5-й день. Данные представлены в виде полученных величин +/- стандартная ошибка. Фиг.29 показывает кривые отклика на дозу, сравнивающие активность форм полной длины и "усеченных" форм rhTPO, полученных в клетках СНО, с усеченной формой из Е. coli. Группам из·6 самок мышей С57 В6 в течение 7 дней ежедневно вводили 0,3мкг rhTPO различных типов. Во 2-7 дни сделали отбор 40мкл крови из глазничной вены для полного анализа. Группы обрабатывались усеченной формой ТРО ТР0153 из Е·coli; TPO332 (смешанная фракция) - ТРО полной длины, содержащий около 80-90% формы полной длины и 10-20% усеченных форм; ТРО332 (фракция 30K) - очищенная усеченная фракция из первоначального "смешанного" препарата; ТРО332 (Фракция 70K) - очищенная фракция полной длины из первоначального "смешанного" препарата. Данные представлены в виде полученных величин +/- стандартная ошибка. Фиг.30 наглядно представляет способ анализа KIRA ELISA для измерения ТРО. Рисунок показывает химеру MPL/Rse.gD и важные участки прародительских рецепторов, а также окончательную стр уктур у (правая часть рисунка) и блок-схему (левая часть рисунка) последовательных этапов анализа. Фиг.31 представляет собой блок-схему для анализа KIRA ELISA, показывающую каждый этап процесса. Фиг.32A-32L представляет последовательность нуклеотидов (номер последовательности: 22) вектора экспрессии pSVI17.ID.LL, используемого для экспрессии Rse.gD в Примере 17. Фиг.33 дает схематическое представление приготовления плазмида pМР1. Фиг.34 дает схематическое представление приготовления плазмида pМР21. Фиг.35 дает схематическое представление приготовления плазмйда рМР151. Фиг.36 дает схематическое представление приготовления плазмйда рМР202. Фиг.37 дает схематическое представление приготовления плазмйда рМР172. Фиг.38 дает схематическое представление приготовления плазмйда рМР210. Фиг.39 представляет собой таблицу пяти лучши х экспрессии клонов из плазмидного банка рМР210 (идентификационные номера последовательностей: 23, 24, 25, 26, 27 и 28). Фиг.40 дает схематическое представление приготовления плазмйда рМР41. Фиг.41 дает схематическое представление приготовления плазмйда рМР57. Фиг.42 дает схематическое представление приготовления плазмйда pМР251. Детальное описание изобретения Определения В целом, следующие слова или фразы при использовании в описании, примерах и формуле изобретения имеют следующие определения. "Хаотропический агент" - относится к соединениям, которые в водном растворе и при соответствующих концентрациях могут вызывать изменения в пространственной конфигурации или конформации белка путем как минимум частичного разрыва связей, ответственных за поддержание нормальной вторичной или третичной структуры белка. Такие соединения включают, например, мочевину, гуанидин-НСl и тиоцианат натрия. Для оказания влияния на конформацию белка обычно требуются высокие концентрации, обычно 49М, эти х соединений. "Цитокин" - родовой термин для белков, высвобожденных из одной клеточной популяции, которые действуют на другую клетку как внутриклеточные посредники. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. К цитокинам можно причислить гормоны роста, инсулиноподобные агенты роста, человеческий гормон роста, N-метионильный человеческий гормон роста, бычий гормон роста, гормон паращитовидной железы, тироксин, инсулин, проинсулин, релаксин, прорелаксин, гликопротеиновые гормоны, такие как гормон стимуляции фолликул (FSH), гормон стимуляции щитовидной железы (TSH) и лейтинизирующий гормон (LH), агент гематопоэтического роста, агент гепатического роста, агент роста фибробластов, пролактин, плацентальный лактоген, р-агент некроза опухолей (TNF-p и TNF-c), мышиный гонадотропин-связанный пептид, ингибин, активин, агент васкулярного эндотелиального роста, интегрин, агенты нервного роста, такие как NGF-c, агент роста тромбоцитов, трансформирующие агенты роста (TGF), такие как TGF-p и TGF-c), инсулиноподобные агенты роста I и II, эритропоэтин-стимулирующие агенты (ЕРО), остеоиндуктивные агенты, интерфероны, такие как интерферон р, -с и g, колониестимулирующие агенты (CSF), такие как макрофаг-CSF (M-CSF), гранулоцито-макрофаг-CSF (GM-CSF), гранулоцито-CSF (G-CSF), интерлейкины (IL), такие как IL-1, IL-lp, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-11, IL-12 и другие полипептидные агенты, включая LIF, SCF и кит-лиганды. Вышеупомянутые термины подразумевают белки, полученные из натуральных источников или рекомбинантных клеточных структур. Эти термины охватывают также биологически активные эквиваленты, например, различающиеся в аминокислотной последовательности одной или, несколькими аминокислотами или типом или степенью гликозилирования. Термины "лиганд mpl", "полипептид лиганда mpl", "ML", "тромбопоэтин" или "ТРО" используются здесь поочередно и включают любой полипептид, который обладает свойством связываться с mpl, членом сверхсемейства цитокиновых рецепторов, и имеет биологические свойства ML, определенные ниже. Примерным биологическим свойством является способность стимулировать инкорпорацию меченых нуклеотидов (например, Н-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, зараженных человеческим mpl P. Другой пример биологического свойства - способность стимулировать инкорпорацию 35S в циркулирующие тромбоциты в анализе мышиных ответных тромбоцитов. Это определение включает полипептид, изолированный от источника лиганда mpl, такого как апластическая свиная плазма, описанная здесь, или другого источника, например, другого вида млекопитающего, включая человека, или приготовленный рекомбинацией или синтетическим способом, и включает также различные его варианты, в том числе функциональные производные, фрагменты, аллели, изоформы и аналоги. "Фрагмент лиганда mpl" или "фрагмент ТРО" - часть натурального зрелого лиганда mpl полной длины, или последовательность с одним или более удаленным аминокислотным остатком. Делеции аминокислотных остатков могут встречаться в любом месте пептида, включая азотные или карбоксильные окончания или внутреннюю часть. Фрагмент обладает как минимум одним биологическим свойством, общим с лигандом mpl. Типичные фрагменты лигандов mpl будут иметь последовательность как минимум 10, 15, 20, 25, 30 или 40 аминокислотных остатков, идентичных последовательностям лигандов mpl, изолированных из млекопитающего, включая лиганд, изолированный из апластической свиной плазмы или лиганд человека или мыши, в частности, его ЕРО-домен. Наглядные примеры N-конечных фрагментов - hML153 или TPO(Met-11153). "Варианты лигандов mpl" или "варианты -последовательностей лигандов mpl" здесь означают биологически активные, как определено ниже, лиганды mpl, -имеющие менее 100% идентичности с лигандом mpl, изолированным из рекомбинантной клеточной культуры или апластической свиной плазмы, или человеческим лигандом, имеющим выведенную последовательность, описанную на Фиг.1 (номер последовательности: 1). В общем случае, биологически активный вариант лиганда mpl будет иметь как минимум около 70% идентичности аминокислотной последовательности с лигандом mpl, изолированным из апластической свиной плазмы или взрослой мыши или человеческим лигандом, или их фрагментами (см. Фиг.1, последовательность номер 1), предпочтительно минимум около 75%, более предпочтительно минимум около 80%, еще более предпочтительно минимум около 90% и наиболее предпочтительно минимум около 95%. "Химерический лиганд mpl" - полипептид, включающий лиганд mpl полной длины или один или более его фрагментов, слитый или химически связанный со вторым гетерологичным полипептидом или одним или более его фрагментами. Химера будет обладать как минимум одним биологическим свойством, общим с лигандом mpl. Второй полипептид обычно является цитокином, иммуноглобулином или их фрагментом. "Изолированный лиганд mpl", "высокоочищенный лиганд mpl" и "в основном·гомогенный лиганд mpl" - эти термины используются здесь поочередно и обозначают лиганд mpl, который был выделен в чистом виде из источника лиганда mpl или был приготовлен рекомбинантным или синтетическим способами и является достаточно отделенным от други х пептидов или протеинов, (1) чтобы получить как минимум 15 и предпочтительно 20 аминокислотных остатков с N-окончаниями или с внутренней аминокислотной последовательностью, используя секвенатор с вращающейся чашкой или промышленный аминокислотный секвенатор или другие опубликованные методы, или (2) чтобы подвергнуться гомогенизации в SDS-PAGE в невосстанавливающих или восстанавливающих условиях с использованием Coomassie blue или, предпочтительно, серебряного окрашивания. Гомогенность здесь означает менее 5% включений других белков из источника. Термин "биологическое свойство", используемый в сочетании с термином "лиганд mpl" или "изолированный лиганд mpl", означает обладание тромбопоэтической активностью или эффекторной или антигенной функцией или активностью в организме, которая прямо или косвенно вызывается или выполняется лигандом mpl (в его природной или преобразованной конформации) или его фрагментом. Эффекторные функции включают способность mpl связываться с любым носителем, агонизм и антагонизм mpl, особенно способность передавать пролиферативные сигналы, включая репликацию, регуляторную функцию для ДНК, модуляцию биологической активности других цитокинов, активацию рецепторов (особенно цитокиновых), деактивацию, регуляцию роста или видоизменения клеток и т.д. Антигенная функция означает обладание эпитопом, или антигенным участком, который способен к перекрестной реакции с антителами, вводимыми против природного лиганда mpl. Принципиальная антигенная функция полипептида лиганда mpl состоит в том, что он связывается со сродством как минимум около 106л/моль с антителом, вводимым против лиганда mpl, изолированного из апластической свиной плазмы. Как правило, сродство связи полипептидов составляет как минимум около 107л/моль. Наиболее предпочтительно, чтобы антигенно-активный полипептид лиганда mpl был полипептидом, который связывается с антителом, вводимым против лиганда mpl, имеющим одну из описанных выше эффекторных функций. Антитела, используемые для определения "биологической активности", представляют собой кроличьи поликлональные антитела, получаемые путем выделения лиганда mpl из рекомбинантной клеточной культуры или апластической свиной плазмы в адъюванте Freund, подкожного введения полученного вещества и усиления иммунного отклика путем внутрибрюшинного введения вещества до те х пор, пока титр антител лиганда mpl не станет постоянным. Термин "биологическая активность", используемый в сочетании с термином "лиганд mpl" или "изолированный лиганд mpl", обозначает лиганд mpl или полипептид, который показывает тромбопоэтическую активность или обладает такой же эффекторной функцией, как лиганд mpl, изолированный из апластичеркой свиной плазмы или экспрессированный в рекомбинантной клеточной культуре, описанной здесь. Принципиальная известная эффекторная функция лиганда mpl или пептида здесь - способность связываться с mpl и стимулировать инкорпорацию меченых нуклеотидов (например, 3Н-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, зараженных человеческим mpl P. Другой пример эффекторной функции -способность стимулировать инкорпорацию 35S в циркулирующие тромбоциты в анализе мышиных ответны х тромбоцитов. Еще одна известная зффекторная функция лиганда mpl - способность стимулировать вне организма человеческий мегакариоцитопоэз, который может быть проанализирован с использованием радиоактивных изотопов в моноклональных антителах, специфических для гликопротеина GPIIbIII a мегакариоцита. "Процент идентичности аминокислотной последовательности" относится к последовательности лиганда mpl, определенного здесь, как процентное содержание аминокислотных остатков в последовательности кандидата, которые идентичны остаткам лиганда mpl, изолированного из апластической свиной плазмы или лиганда мыши или человека, имеющего выведенную аминокислотную последовательность, описанную на Фиг.1 (номер последовательности: 1), после совмещения последовательностей и введения, если необходимо, пустот для достижения максимальной процентной идентичности, не рассматривая законсервированные v заместители как идентичные. Никакие N-окончания, С-окончания, внутренние добавления, делеции или вставки в последовательность лиганда mpl не должны вводиться в последовательность, как влияющие на ее идентичность или гомологичность. Так, примером биологически активных полипептидов лигандов mpl, имеющих идентичные последовательности, являются: лиганд препро-mpl, лиганд про-mpl и зрелый лиганд mpl. "Микросеквенирование лиганда mpl" может быть осуществлено любым подходящим стандартным способом, если он достаточно чувствителен. В одном из таких способов высокоочищенный полипептид, полученный из геля SDS или полученный на последнем этапе HPLC, секвенируется непосредственно при помощи автоматизированной (фенилизотиоцианатнрй) деградации Edman с использованием газофазного секвенатора АррПеД Biosystems модели 470А, оснащенного фенилтиогидантионовым (РТН) аминокислотным анализатором 120А. Кроме того, подобным образом могут быть секвенированы фрагменты лиганда mpl, полученного химическим (например, при помощи CNBr, гидроксиламина, 2-нитро-5-тиоцианобензоата) или энзиматическим (например, при помощи трипсина, клострипаина, стафилококковой протеазы) расщеплением с последующей фрагментарной очисткой (например, HPLC). РТН-аминокислоты анализируются с использованием системы данных ChromPerfect (Justice Innovations, Palo Alto, CA). Интерпретация последовательности выполняется на компьютере VAX 11/785 Digital Equipment Co., как описано в Henzel et al., J. Chromatography, 404:41-52 (1987). Как вариант, слои фракций HPLC могут быть подвергнуты электрофорезу на 5-20% SDS-PAGE, перенесены на мембрану PVDF (ProBlott, AIB, Foster City, CA) и окрашены Coomassie Brilliant Blue (Matsudiara, D. Biol. Chem., 262:10035-10038 (1987). Специфический белок, идентифицированный окрашиванием, извлекается из образца, и проводится N-конечное секвенирование в газофазном секвенаторе, описанном выше. Для выстраивания внутренних последовательностей белка фракции HPLC высушиваются в условиях вакуума (Speedvac), ресуспендируются в соответствующи х буферах и расщепляются цианогенбромидом, Lys-специфическим энзимом Lys-C (Wako Chemicals, Richmond, VA) или Asp-N (Boehringer Mannheim, Indianapolis, IN). После расщепления результирующие пептиды секвенируются в виде смеси, или после HPLC-разложения в колонке С4 развиваются при помощи пропанольного градиента в 0,1% TFA перед газофазным секвенированием. "Тромбоцитопения" определяется как количество тромбоцитов ниже 150´109 на литр крови. "Тромбопоэтическая активность" определяется как биологическая активность, которая состоит из ускорения размножения, видоизменения и/или развития мегакариоцитов или их предшественников в тромбоцито-продуцирующую форму этих клеток. Эта активность может быть измерена различными методами анализа, включая метод ответных мышиных тромбоцитов в организме, метод индуцирования антигена поверхности клетки тромбоцита, с измерением его антитромбоцитным иммуноанализом (анти-GPIIbIIIa) для мегакариобластической клеточной линии человеческой лейкемии (СМK), и метод индуцирования полиплоидизации в мегакариобластической клеточной линии (DAMI). "Тромбопоэтин" (ТРО) определяется как соединение, обладающее тромбопоэтической активностью или способностью повышать количество сывороточных тромбоцитов у млекопитающих. ТРО предпочтительно способен увеличивать количество эндогенных тромбоцитов как минимум на 10%, более предпочтительно на 50%, и наиболее предпочтительно способен поднимать количество тромбоцитов в человеке до более чем 150´10 на литр крови. "Нуклеиновая кислота изолированного лиганда mpl" - РНК или ДНК, содержащая более 16 и предпочтительно 20 и более последовательных нуклеотидных оснований, которая кодирует биологически активный лиганд mpl или его фрагмент, комплементарная к ДНК или РНК или гибридизируется с ДНК или РНК и сохраняет стабильную связь в умеренно строгих и строгих условиях. Эта РНК или ДНК свободна от, как минимум, одной нуклеиновой кислоты источника контаминации, с которой она обычно связана в природном источнике, и предпочтительно в основном свободна от любых други х РНК или ДНК млекопитающих. Фраза "свободна от, как минимум, одной нуклеиновой кислоты источника контаминации, с которой она обычно связана в природном источнике" включает случай, когда нуклеиновая кислота присутствуе т в источнике природной клетки, но находится в другом хромосомном положении или граничит с последовательностями нуклеиновых кислот, обычно не присутствующими в клетке источника. Примером нуклеиновой кислоты изолированного лиганда mpl служит ДНК или РНК, кодирующая биологически активный лиганд mpl, имеющая как минимум 75% идентичности последовательности, предпочтительно минимум 80%, более предпочтительно минимум 85%, еще более предпочтительно минимум 90% и наиболее предпочтительно минимум 95% идентичности последовательности с человеческим, мышиным или свиным лигандом mpl. "Контрольные последовательности" означают последовательности ДНК, необходимые для экспрессии оперативно связанной кодирующей последовательности в конкретном организме хозяина. Контрольные последовательности, подходящие для прокариоцитов, например, включают промотор, возможно, последовательность-оператор, участок связывания рибосом, и, возможно, другие, еще плохо исследованные, последовательности. Эукариотические клетки, как выяснено, используют промоторы, сигналы полиаденилирования и ген-усилители. "Оперативно связанные" в применении к нуклеиновым кислотам означает, что нуклеиновые кислоты находятся в функциональных взаимоотношениях с другими последовательностями нуклеиновых кислот. Например, ДНК, отвечающая за препоследовательность или секреторный лидер, оперативно связана с ДНК, отвечающей за полипептид, если она выражена как препротеин, участвующий в секреции полипетида; генусилитель или промотор оперативно связан с кодирующей последовательностью, если она влияет на транскрипцию последовательности; участок связывания в рибосоме оперативно связан с кодирующей последовательностью, если она расположена так, чтобы облегчать трансляцию. В целом, "оперативно связанные" означает, что связываемые последовательности ДНК являются смежными и, в случае секреторного лидера, смежными и находящимися в фазе считывания. Однако гены-усилители не должны быть смежными. Связывание выполняется путем лигирования в удобных ограничительных участках. Если таких участков не существует, используются синтетические олигонуклеотидные адаптеры или связующие вещества , в соответствии с обычной практикой. "Экзогенный" в применении к элементам означает последовательность нуклеиновых кислот, являющуюся чужеродной в данной клетке, или гомологичную для клетки, но находящуюся в таком положении внутри нуклеиновой кислоты хозяйской клетки, в котором обычно этот элемент не присутствует. "Клетка", "клеточная линия" и "клеточная культура" используются здесь поочередно, и такие определения включают все потомство клетки или клеточной линии. Так, например, термины "трансформанты" и "трансформированные клетки" включают первичную клетку объекта и культуры, произведенные от нее, независимо от количества пересадок. Понятно также, что все потомство может не быть полностью идентично по содержанию ДНК из-за случайных мутаций. Мутантное потомство, имеющее ту же функцию или биологическую активность, что и первоначальная клетка, также включается. Различия вышеуказанных обозначений будут видны по контексту. "Плазмиды" - автономно воспроизводимые молекулы кольцевых ДНК, имеющие независимое происхождение и обозначенные здесь прописной буквой "p", перед или после которой стоят заглавные буквы или цифры. Стартовые плазмиды здесь представляют собой или промышленно используемые, обычно доступные без ограничений, или могут быть построены из таких доступных плазмидов в соответствии с известными способами. Кроме того, другие эквивалентные плазмиды описаны в литературе и известны любому специалисту. "Ограниченное энзимное расщепление" в применении к ДНК означает каталитическое разложение внутренних фосфодиэфирных связей ДНК энзимами, которые действуют только в определенных участках последовательности ДНК. Такие энзимы называются "ограниченными эндонуклеазами". Каждая ограниченная эндонуклеаза распознает определенную последовательность ДНК, называемую "участком ограничения" и обладающую двухскладчатой симметрией. Здесь используются коммерчески доступные ограниченные энзимы, с соблюдением их реакционных условий, коэнзимов и других требований, устанавливаемых производителями энзимов. Все ограниченные энзимы обозначаются аббревиатурами, состоящими из заглавной буквы, за которой следуют др угие буквы, представляющие микроорганизм, из которого был получен данный энзим, и затем цифра, обозначающая конкретный энзим. В общем случае, используется около 1мкл плазмида или фрагмента ДНК с 1-2 единицами энзима в примерно 20мкл буферного раствора. Соответствующие буферы и количества субстрата для конкретного ограниченного энзима определяются производителем. Обычно используется инкубация в течение около 1ч. при 37°C, но условия могут варьироваться в соответствии с инструкциями поставщика. После инкубации белок или полипептид удаляется путем экстракции фенолом или хлороформом, а расщепленная нуклеиновая кислота выделяется из водной фракции осаждением этанолом. За расщеплением ограниченными энзимами может следовать бактериально-щелочной гидролиз фосфатазой фосфа тов 5'-окончаний для предохранения двух расщепленных концов фрагмента ДНК от "закольцовывания" или образования замкнутой петли, что сделает невозможной вставку другого фрагмента ДНК в участок ограничения. Если не установлено иначе, после расщепления плазмидов не производится дефосфорилирование 5'-окончаний. Методики и реагенты дефосфорилирования обычны и описаны в разделах 1.56-1.61 Sambrook et al., Volecular Cloning: A Laboratory Manual [New York: Cold Spring Harbor Laboratory Press, 1989]. "Выделение" или "изоляция" данных фрагментов ДНК из устройств для расщепления означает отделение расщепленных ДНК на полиакриламиде или геле агарозы электрофорезом, идентификацию нужного фрагмента путем сравнения его подвижности с подвижностью контрольных фрагментов ДНК известного молекулярного веса, извлечение гель-фракции, содержащей нужный фрагмент и отделение геля от ДНК. Эта методика общеизвестна. Например, см. Lawn et al., Nucleic Acids Res., 9:6103-6114 (1981), и Goeddel et al., Nucleic Acids Res., 8:4057 (1980). "Южный анализ" или "южный блоттинг" - способ, которым наличие последовательностей ДНК в композиции после расщепления ограниченной эндонуклеазой обнаруживается путем гибридизации с известным, меченым олигонуклеотидом или фрагментом ДНК. Южный анализ обычно включает электрофоретическое отделение расщепленных фрагментов ДНК на геле агарозы, денатурацию ДНК после электрофоретического отделения и пересадку ДНК на нитроцеллюлозу, нейлон или другую подходящую мембранную подложку для анализа меченным радиоактивными изотопами, биотинилированным или энзиммеченным зондом, как описано в разделах 9.37-9.52 Sambrook et.al., выше. "Северный анализ" или "Северный блоттигн" - способ, используемый для идентификации последовательностей РНК, которые гибридизируются с известным зондом, таким как олигонуклеотид, фрагмент ДНК, кДНК или ее фрагмент, или фрагмент РНК. Зонд метится радиоизотопом, таким как 32Р, или биотинилированием, или энзимом. Анализируемая РНК обычно отделяется электрофорезом на агарозе или геле полиакриламида, пересаживается на нитроцеллюлозу, нейлон или другую подходящую мембранную подложку и гибридизируется с зондом с использованием стандартной технологии, хорошо известной в данной области, например, описанной в разделах 7.39-7.52 Sambrook et al., выше. "Лигирование" - процесс образования фосфодиэфирных связей между двумя фрагментами нуклеиновой кислоты. Для лигирования двух фрагментов концы фрагментов должны быть совместимыми друг с другом. В некоторых случаях концы станут непосредственно совместимыми после расщепления эндонуклеазой. Однако может быть, необходимо сначала преобразовать, неустойчивые концы,обычно получаемые после расщепления, в прямые концы, совместимые при лигировании. Для выпрямления концов ДНК обрабатывается подходящим буфером в течение минимум 15 минут при 15°C примерно с 10 единицами фрагмента Кленова ДНК-полимеразы I или Т4 ДНК-полимеразы в присутствии четырех дезоксирибонуклеотидтрифосфатов. Затем ДНК очищается феноло-хлороформной экстракцией и этанольным осаждением. Фрагменты ДНК, которые должны быть сшиты вместе, помещаются в раствор в примерно эквимолярных количествах. Раствор также содержит АТР, буфер лигазы и лигазу типа Т4 ДНКлигазы при содержании около 10 единиц на 0,5мкг ДНК. Если ДНК должна быть лигирована с вектором, вектор сначала линеаризуется путем расщепления соответствующей ограниченной эндонуклеазой(ами). Линеаризованный фрагмент затем обрабатывается бактериально-щелочной фосфатазой или телячьей кишечной фосфатазой для предотвращения самолигирования во время этапа лигирования. "Приготовление" ДНК из клеток означает изоляцию ДНК плазмида из культуры хозяйских клеток. Обычно используемые способы приготовления ДНК - крупно- и мелкомасштабное плазмидное приготовление, описанное в разделах 1.25-1.33 Sambrook et al., выше. "Олигонуклеотиды" - короткие, состоящие из одной или двух цепей полидезоксинуклеотиды, химически синтезированные известными способами [такими как фосфотриэфирная, фосфитная или фосфорамидитная химия, использующая твердофазную те хнологию, как описано в ЕР 266,032 публ. 4 мая 1988, или через дезоксинуклеозид-Н-фосфатные промежуточные продукты, как описано у Froehler et al., Nucl. Acids Res., 14:5399-5407 (1986)]. Другие способы включают реакцию цепи полимеразы, определенную ниже, и другие автоинициирующие способы и синтезы олигонуклеотидов на твердых подложках. Все эти способы описаны в Engels et al., Agnew. Chem. Int. Ed. Engl., 28:716-734 (1989). Эти способы используются в том случае, если вся последовательность нуклеиновой кислоты гена известна или комплементарность последовательности нуклеиновой кислоты с кодирующей цепью возможна. Альтернативный способ: если целевая аминокислотная последовательность известна, можно теоретически предположить потенциальные последовательности нуклеиновых кислот, используя известные и предпочтительные кодирующие остатки для каждого аминокислотного остатка. Затем олигонуклеотиды очищаются на полиакриламидных гелях. "Реакция цепи полимеразы" или "PCR" относится к процессу или технологии, в которой малые количества, определенного участка нуклеиновой кислоты, РНК и/или ДНК, умножаются, как описано в U.S. Patent No.4,683,195, публ. 28 июля 1987. В общем случае, должна иметься информация о последовательности от концов нужного участка или за его пределами, такая, чтобы могли быть построены инициаторы олигонуклеотидов; последовательности этих инициаторов будут идентичными или подобными противоположным цепям матрицы, которая должна быть умножена. 5'-конечные нуклеотиды двух инициаторов могут соответствовать концам распространяемого материала. PCR можно использовать для распространения специфических последовательностей РНК, специфических последовательностей ДНК из общей геномной ДНК, и кДНК, транскрибированной из общей клеточной РНК, "бактериофаговых или плазмидных последовательностей, и т.д. См. Mullis et al., Cold Spring Harbor Symp. Quant. Biol., 51:263 (1987); Erlich, ed., PCR Technology, [Stockton Press, NY, 1989]. Здесь PCR рассматривается как один, но не единственный пример метода реакции нуклеиновой кислоты полимеразы для умножения нуклеиновой кислоты в испытательном образце, включающий использование известной нуклеиновой кислоты в качестве инициатора и нуклеиновой кислоты полимеразы для умножения или генерирования специфического участка нуклеиновой кислоты. "Строгие условия" - такие, которые (1) предусматривают низкую стойкость ионов и высокую температуру промывки, например 0,015Μ NaCl/0,015Μ цитрата натрия/0,1% NaDodSO4 (SDS) при 50°С, или (2) предусматривают во время гибридизации денатурирующий агент, такой как формамид, например, 50% (об./об.) формамида с 0,1% альбумина бычьей сыворотки/0,1% Ficoll/0,1% поливинилпирролидона/50мМ натриево-фосфатного буфера при рН6,5 с 750мМ цитрата натрия при 42°С. Другой пример - использование 50% формамида, 5´SSC (0,75Μ NaCl, 0,075Μ цитрата натрия), 50мМ фосфа та натрия (рН6,8), 0,1% пирофосфата натрия, 5´ раствора Denhardt, обработанная ультразвуком ДНК спермы лосося (50мкг/мл), 0,1% SDS и 10% сульфата декстрана при 42°С, с промывкой при 42°С в 0,2´SSC и 0,1% SDS. "Умеренно строгие условия" описаны в Sambrook et al., выше, и включают использование промывочного раствора и условий гибридизации (температура, стойкость ионов и % SDS), менее строгих, чем описаны выше. Пример умеренно строгих условий - инкубация в течение ночи при 37°C в растворе, включающем: 20% формамида, 5´SSC (150мМ NaCl, 15мМ тринатрийцитрата), 50мМ фосфата натрия (рН 7,6), 5´ раствора Denhardt, 10% сульфата декстрана и 20% денатурированной разрезанной ДНК спермы лосося с последующей промывкой фильтров в 1´ SSC при 37-50°С. Опытный специалист поймет, как отрегулировать температуру, стойкость ионов и т.д. для приведения в соответствие таких факторов, как длина зонда и т.п. "Анти тела" (Abs) и "иммуноглобулины" (Igs) - гликопротеины, имеющие одинаковые структурные характеристики. В то время как антитела проявляют способность связываться с конкретными антигенами, иммуноглобулины включают как антитела, так и другие антителоподобные молекулы с недостаточной специфичностью по отношению к антигенам. Полипептиды последнего вида, например, продуцируются на низких уровнях лимфатической системой, а на высоких уровнях - миеломами. "Природные антитела и иммуноглобулины" – обычно гетеротетрамерические гликопротеины величиной примерно 150 000 дальтон, состоящие из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей. Каждая легкая цепь связана с тяжелой цепью одной ковалентной дисульфидной связью, в то время как число дисульфидных связей между тяжелыми цепями варьируется в зависимости от изотипов иммуноглобулинов. Каждая тяжелая и легкая цепь через одинаковые промежутки включает внутрицепные дисульфидные мостики. Каждая тяжелая цепь имеет на одном конце переменный домен (VH), за которым следует ряд постоянных доменов. Каждая легкая цепь имеет на одном конце переменный домен (VL) и на другом конце постоянный домен; постоянный домен легкой цепи совмещается с первым постоянным доменом тяжелой цепи, а переменный домен легкой цепи совмещается с переменным доменом тяжелой цепи. В частности, считается, что взаимосвязь между переменными доменами тяжелой и легкой цепей обеспечивают аминокислотные остатки [Clothia et al., J. Моl. Biol., 186:651-663 (1985); Novotny and Haber, Proc. Natl. AcadSci. USA, 82: 4592-4596 (1985)]. Термин "переменный" относится к тому факту, что определенные участки переменных доменов значительно различаются в последовательности для различных антител и участвуют в связывании и активности каждого конкретного антитела с его конкретным антигеном. Однако переменность неравномерно распределена по всему переменному домену. Она концентрируется в тре х сегментах, называемых участками, определяющими комплементарность (CDR), или в сверхпеременных участках доменов как легких, так и тяжелых цепей. Более высококонсервированные участки переменных доменов называются рамками (FR). Переменные домены природных тяжелых и легких цепей, каждый из которых состоит из четырех участков FR, в основном принимающих с-пластинчатую конфигурацию, соединяются тремя CDR, которые образуют петли, соединяясь с с-структурой, а в некоторых случаях образуя ее часть. CDR в каждой цепи находятся в непосредственной близости друг к другу посредством участков FR, и, вместе с CDR другой цепи участвуют в образовании в антителе участка связывания с антигеном [см. Kabat et al., Sequences of Proteins of Immunological Interest National Institute of Health, Bethesda, MD (1987)]. Постоянные домены не участвуют непосредственно в связывании с антигеном, но проявляют различные эффекторные функции, такие как участие в клеточной токсичности антитела или антитело-обусловленной клеточной токсичности. Папаиновое расщепление антител продуцирует два идентичных антиген-связывающих фрагмента, называемых фрагментами "Fab", каждый из которых имеет один участок для связывания с антигеном, и остаточный фрагмент "Fc", название которого отражает его способность легко кристаллизоваться. Обработкой пепсином получают фрагмент F(ab')2, который имеет два участка связывания с антигеном и все еще способен к перекрестной связи с антигеном. "Fv" - минимальный фрагмент антитела, который содержит полный участок распознавания и связывания антигена. Этот участок состоит из димера одной тяжелой и переменного домена одной легкой цепи в жесткой, нековалентной, связи. Именно в этой конфигурации взаимодействуют три CDR каждого переменного домена, образуя участок связывания антигена на поверхности димера VH-VL. Шесть CDR вместе обусловливают свойство антитела связываться со специфическим антигеном. Однако, даже один переменный домен (или половина Fv, включающая только три CDR, специфических для антигена) обладает способностью распознавать и связывать антиген, хотя и с более низким сродством, чем целый связующий участок. Фрагмент Fab также содержит постоянный домен легкой цепи и первый постоянный домен (СН1) тяжелой цепи. Фрагменты Fab' отличаются от фрагментов Fab дополнением нескольких остатков у карбоксильного окончания СН1-домена тяжелой цепи, включая один или более цистеинов из шарнирного участка антитела. Fab'-SH - здесь обозначение для Fab', в котором остаток (остатки) цистеина в постоянных доменах содержат свободные тиоловые группы. Фрагменты антитела F(аb')2 первоначально продуцировались как пары фрагментов Fab', соединяющиеся шарнирными цистеинами. Другие химические пары фрагментов антител также известны. Термин "легкая цепь" антител (иммуноглобулинов) любого позвоночного, может быть отнесен к одному из двух ярко выраженных типов, называемых каппа и лямбда и основанных на аминокислотных последовательностях их постоянных доменов. В зависимости от аминокислотной последовательности постоянных доменов их тяжелых цепей, иммуноглобулины могут быть отнесены к различным классам. Существуе т пять основных классов иммуноглобулинов: Ig A, IgD, IgE, IgG и Ig M, и некоторые из них далее разделяются на подклассы (изотипы): IgG-1, IgG-2, IgG-3 и IgG-4; Ig A-1 и Ig A-2. Постоянные домены тяжелых цепей, которые соответствуют различным классам иммуноглобулинов, называются альфа, дельта, эпсилон, гамма и мю, соответственно. Структуры подэлементов и трехмерные конфигурации различных классов иммуноглобулинов хорошо известны. Термин "антитело", используемый в широчайшем смысле, конкретно обозначает отдельные моноклональные антитела (включая антитела-агонисты и антагонисты), композиции антител с полиэпйтопической спецификой, а также фрагменты антител (например, Fab, F(ab')2 и Fv), до тех пор, пока они проявляют желаемую биологическую активность. Термин "моноклональное антитело" здесь относится к антителу, полученному из популяции в основном гомогенных антител, т.е., отдельные антитела, составляющие популяцию, идентичны, за исключением возможных естественных мутаций, происходящих в незначительных количествах. Моноклональные антитела высокоспецифичны и направлены против отдельных участков антигенов. Далее, в отличие от препаратов обычных (поликлональных) антител, которые, как правило, включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело направлено против одной детерминанты антигена. В дополнение к их специфичности, моноклональные антитела имеют преимущество в том, что они синтезируются культурой гибридомы, не загрязненной другими иммуноглобулинами. Определение "моноклональное" показывает ту особенность антитела, что оно получено из, в основном, гомогенной популяции антител, и не означает, что это антитело получено каким-либо особым способом. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены способом гибридомы, впервые описанным Kohler & Milstein, Nature, 256:495 (1975), или способами рекомбинации ДНК [см., например, U.S. Patent No.4,816,567 (Cabilly et al.)]. Моноклональные антитела здесь особо включают "химерические" антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепи идентична или гомологична с соответствующими последовательностями в антителах, полученных от определенных видов животных или принадлежащих к определенному классу или подклассу антител, в то время как остальная часть цепи (цепей) идентична или гомологична с соответствующими последовательностями в антителах, полученных от други х видов животных или принадлежащих к другому классу или подклассу антител, а также во фрагментах этих антител, пока они проявляют желаемую биологическую активность [U.S. Patent No.4,816,567 (Cabilly et al.); Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855 (1984)]. "Гуманизированные" формы нечеловеческих (например, мышиных) антител представляют собой химерические иммуноглобулины, цепи иммуноглобулинов или их фрагменты (например, Fv, Fab, Fab', F(ab')2 или другие антиген-связующие последовательности антител), содержащие минимальные последовательности, полученные от нечеловеческого иммуноглобулина. В большей степени гуманизированные антитела являются человеческими иммуноглобулинами (антитело реципиента), в которых остатки из участков, определяющих комплементарность (CDR), реципиента заменяются остатками из CDR нечеловеческого вида (антитело донора), такого как мышь, крыса или кролик, имеющего желаемую специфичность, сродство и возможности. В некоторых случаях остатки рамки Fv человеческого иммуноглобулина заменяются соответствующими нечеловеческими остатками. Далее, гуманизированное антитело может включать остатки, которые не обнаружены ни в антителе реципиента, ни в импортируемом CDR или последовательностях рамки. Эти модификации сделаны для улучшения и оптимизации характеристик антитела. В целом, гуманизированное антитело будет содержать практически все из как минимум, одного, обычно двух, переменных доменов, в которых все или, практически все участки CDR соответствуют участкам CDR нечеловеческого иммуноглобулина и все или практически все участки FR соответствуют последовательностям участков FR человеческого иммуноглобулина. Гуманизированное антитело оптимально будет также содержать как минимум часть постоянного участка иммуноглобулина (Fc), обычно человеческого иммуноглобулина. Более детально [см. Jones et al., Nature, 321:522-525 (1986); Reichmann et al., Nature, 332:323-329 (1988); Presta, Curr. Op. Struct. Biol., 2:593-596 (1992)]. "Неиммуногенный в человеке" означает, что при контакте полипептида в фармацевтически приемлемом носителе и в терапевтически эффективном количестве с соответствующей тканью человека никакой чувствительности или сопротивления полипептиду после второго введения полипептида и после соответствующего латентного периода (т.е. от 8 до 14 дней) не было проявлено. II. Предпочтительные варианты осуществления изобретения Предпочтительными полипептидами данного изобретения являются в основном гомогенные полипептид(ы), называемые здесь лиганд(ы) mpl или тромбопоэтин (ТРО), которые обладают свойством связываться с mpl, членом сверхсемейства цитокиновых рецепторов и имеют биологическое свойство стимуляции инкорпорации меченых нуклеотидов (3Н-тимидина) в ДНК IL-3-зависимых клеток Ba/F3, подвергнутых трансфекции человеческим mpl P. Более предпочтительно, чтобы лиганд(ы) mpl представляли собой изолированный белок (белки), обладающий гематопоэтической, особенно мегакариоцитопоэтической или тромбопоэтической активностью, а именно, способностью стимулировать размножение, развитие и/или видоизменение незрелых мегакариоцитов или их предшественников в зрелую тромбоцито-продуцирующую форму. Наиболее предпочтительно, чтобы полипептиды данного изобретения представляли собой человеческие лиганд(ы) mpl, включая их фрагменты, обладающие гематопоэтйческой, мегакариоцитопоэтической или тромбопоэтической активностью. Возможно, эти человеческие лиганд(ы) mpl отличаются недостаточным гликозилированием. Другие предпочтительные человеческие лиганды mpl представляют собой "ЕРО-домены" hML, называемые здесь hML153 или hTPO153, усеченную форму hML, называемую hML245 или hTPO 245 и зрелый полипептид полной длины, имеющий аминокислотную последовательность, показанную на Фиг.1 (номер последовательности: 1), называемый hML, hML332 или hTPO332 и биологически активный замещенный вариант hML (R153A, R154A). Возможно, предпочтительными полипептидами данного изобретения являются биологически или иммунологически активные варианты лигандов mpl, выбранные из hML2, hML3, hML4, mMl, mMl2, mМІ3, pML и pML2. Возможно, предпочтительными полипептидами данного изобретения являются биологически активные варианты лигандов mpl, имеющие аминокислотные последовательности, которые как минимум на 70% идентичны человеческому лиганду mpl [см. Фиг.1 (номер последовательности: 1)], лиганду mpl мыши [см. Фиг.16 (номер последовательности: 12 и 13)], рекомбинантному свиному лиганду mpl [см. Фиг.19 (номер последовательности: 18)] или свиному лиганду mpl, изолированному из апластической свиной плазмы, предпочтительно минимум на 75%, более предпочтительно минимум на 80%, еще более предпочтительно минимум на 90% и наиболее предпочтительно минимум на 95%. Лиганды mpl, изолированные из апластической свиной плазмы, имеют следующие характеристики: (1) Частично очищенный лиганд элюируется из гель-фильтрационной колонки с PBS, PBS, содержащим 0,1% SDS или с PBS, содержащим 4М МgСl2 с Mr 60,000-70,000; (2) Активность лиганда уничтожается проназой; (3) Лиганд стабилен к низкой рН (2,5), к SDS до 0,1% и мочевине 2М; (4) Лиганд представляет собой гликопротеин, основанный на его связывании с различными лектиновыми колонками; (5) Высокоочищенный лиганд элюируется из невосстановленного SDS-PAGE с Mr 25,000-35,000. Меньшие количества активности также элюируются с Mr 18,000-22,000 и 60,000; (6) Высоко очищенный лиганд растворяется в восстановленном SDS-PAGE в виде пар с Mr 28,000 и 31,000; (7) Амино-конечные последовательности полос 18,000-22,000, 28,000 и 31,000 - одно и то же: SPAPPACDPRLLNKLLRDDHVLHGR (номер последовательности: 29); и (8) Лиганды связываются и элюируются из следующих колонок сродства: Blue-Sepharose, CM Blue-Sepharose, MONO-Q, MONO-S, Lentil lectin-Sepharose, WGA-Sepharose, Con A-Sepharose, Ether 650m Toyopearl, Butyl 650m Toyopearl, Phenyl 650m Toyopearl, и Phenyl-Sepharose. Более предпочтительными полипептидами лигандов mpl являются такие, которые кодируются человеческими геномними или кДНК, имеющими аминокислотную последовательность, описанную на Фиг.1 (номер последовательности: 1). Другие предпочтительные природные биологически активные полипептиды лигандов mpl данного изобретения включают лиганд препро-mpl, про-mpl, зрелый лиганд mpl, фрагменты лиганда mpl и их гликозилированные варианты. Другие предпочтительные полипептиды данного изобретения включают варианты и химеры последовательностей лигандов mpl. Обычно предпочтительные варианты и химеры последовательностей лигандов mpl - биологически активные варианты лигандов mpl, имеющие аминокислотные последовательности, которые как минимум на 70% идентичны человеческому лиганду mpl или свиному лиганду mpl, изолированному из апластической свиной плазмы, предпочтительно минимум на 75%, более предпочтительно минимум на 80%, еще более предпочтительно минимум на 90% и наиболее предпочтительно минимум на 95%. Примером предпочтительного варианта лиганда mpl является N-конечный домен варианта hML (называемый "ЕРО-дамен", так как его последовательность гомологична эритропоэтину). Предпочтительный ЕРО-домен hML включает около 153 первых аминокислотных остатков зрелого hML и называется hML153. Возможный предпочтительный вариант последовательности hML такой, в котором один или более основных или диосновных аминокислотных остатков в С-конечном домене замещены неосновным аминокислотным остатком (остатками) (гидрофобным, нейтральным, кислртным, ароматическим, Gly, Pro и т.д.). Предпочтительный вариант последовательности С-конечного домена hML - такой, в котором Arg-остатки 153 и 154 замещены остатками Ala. Альтернативный предпочтительный вариант последовательности hML содержит hML153 или hML332 , в котором аминоостатки 111-114 (QLPP или LPPQ) удалены или замещены различными тетрапептидными последовательностями (AGAG и т.п.). Выше указанные мутанты с делециями называются D4hML332 или D4hML153 . Предпочтительная химера - слияние между лигандом mpl или его фрагментом (определенным ниже) с гетерологическим полипептидом или его фрагментом. Например, hML153 может быть слит с фрагментом IgG для улучшения полупериода существования сыворотки или с IL-3, G-CSF или ЕРО для получения молекулы с выраженной тромбопоэтической или химерической гематопоэтической активностью. Альтернативной предпочтительной химерой человеческого лиганда mpl является "химера домена MLEPO", которая состоит из N-конечных остатков hML от 153 до 157, замещенных одним или более, но не всеми, остатками человеческого ЕРО, расположенного примерно так, как показано на Фиг.10 (номер последовательности: 7). В этом варианте осуществления изобретения химера hML имеет длину около 153166 остатков, где в последовательность hML добавляются или служат заместителями отдельные остатки или их блоки из последовательности человеческого ЕРО, в положениях, соответствующи х выравниванию, показанному на рис. 10 (номер последовательности: 6). Примерная блочная последовательность, вставленная в N-конечную часть hML, включает один или более участков N-гликозилирования в положениях (ЕРО) 24-27, 38-40 и 83-85; один или более из четырех предсказанных алифатических р-спиральных узлов в положениях (ЕРО) 9-22, 59-76, 90-107 и 132-152; другие высококонсервированные участки, включая Nконечные и С-конечные участки и остатки в положениях (ЕРО) 44-52 [см., например, Wen et al., Blood, 82:15071516 (1993) и Boissel et al., J. Biol. Chem., 268(21):15983-15993 (1993)]. Предполагается, что эта "химера доменов ML-EPO" буде т иметь смешанную тромбопоэтическую-эритропоэтическую (ТЕРО) биологическую активность. Другие предпочтительные полипептиды данного изобретения включают фрагменты лиганда mpl, имеющие последовательность из, как минимум, 10, 15, 20, 25, 30 или 40 аминокислотных остатков, которые идентичны последовательностям лиганда mpl, изолированного из апластической свиной плазмы или человеческого лиганда mpl, описанного здесь (см. Таблицу 14, Пример 24). Предпочтительным фрагментом лиганда mpl является человеческий ML[1-X], где X-153, 164, 191, 205, 207, 217, 229 или 245 [см. Фиг.1 (номер последовательности: 1), последовательность остатков 1-Х]. Другие предпочтительные фрагменты лиганда mpl включают фрагменты, полученные в результате химического или энзиматического гидролиза или расщепления очищенного лиганда. Другой предпочтительный объект изобретения - способ очистки молекул лиганда mpl, предусматривающий контактирование источника лиганда mpl, содержащего молекулы лиганда mpl, с иммобилизованным рецепторным полипептидом, особенно полипептидом mpl или слитого mpl, в условия х, при которых очищаемые молекулы лиганда mpl избирательно адсорбируются на иммобилизованном рецепторном полипептиде, промывку иммобилизованной подложки для удаления неадсорбированного материала и элюирование очищаемых молекул из иммобилизованного рецепторного полипептида элюирующим буфером. Источником, содержащим лиганд mpl, может быть плазма, в которой иммобилизованным рецептором предпочтительно является слияние mpl-IgG. Альтернативным источником, содержащим лиганд mpl, является рекомбинантная клеточная культура, в которой концентрация лиганда mpl в среде культуры или в клеточных лизатах в основном выше, чем в плазме или в других природных источниках. В этом случае описанный выше метод иммуносродства, хотя и пригоден, но обычно не используется, так как можно применить более традиционные способы очистки белков, известные в данной области. В кратком изложении, предпочтительный способ очистки для получения в основном гомогенного лиганда mpl включает: удаление посторонних частиц, а также хозяйских клеток или лизированных фрагментов, например, центрифугированием или ультрафильтрацией; возможно концентрирование белка при помощи коммерчески доступного концентрационного фильтра; отделение лиганда от други х примесей в один или несколько этапов, выбранных из способов иммуносродства, ионообмена (например, DEAE или матрицы, содержащие карбоксиметильные или сульфопропильные группы), Blue-Sepharose, CM Blue-Sepharose, MONO-Q, MONO-S, Lentil lectin-Sepharose, WGA-Sepharose, Con A-Sepharose, Ether Toyopearl, Butyl Toyopearl, Phenyl Toyopearl, протеиновый А-Sepharose, SDS-Page, обращенно-фазовая HPLC (силикагель с дополнительными алифатическими группами) или хроматография на молекулярных ситах Sephadex, и затем осаждение этанолом или сульфатом аммония. Ингибитор протеазы, такой как метилсульфонилфторид (PMSF), может быть включен в любой из вышеописанных этапов для ингибирования протеолиза. В другом предпочтительном варианте изобретение предлагает изолированное антитело, способное связываться с лигандом mpl. Предпочтительное антитело лиганда mpl [является моноклональным Kohler & Milstein, Nature, 256:495-497 (1975); Campbell, Laboratory Techniques in Biochemistry and Molecular Biology, Burdon et al., Eds, Volume 13, Elsevier Science Publishers, Amsterdam (1985); Huse et al., Science, 246:1275-1281 (1989)]. Предпочтительным антителом лиганда mpl является такое, которое связывается с лигандом mpl со сродством как минимум около 106л/моль. Более предпочтительным является антитело, которое связывается со сродством как минимум около 107л/моль. Наиболее предпочтительно, чтобы антитело, действующее против лиганда mpl, имело одну из вышеописанных эффекторных функций. Изолированным антителом, способным связываться с лигандом mpl, как вариант, может быть слито со вторым полипептидом, и полученное таким слиянием антитело может быть использовано для изолирования и очистки лиганда mpl из источника, как описано выше, для иммобилизованного полипептида mpl. В следующем предпочтительном аспекте этого варианта осуществления изобретение предлагает способ для обнаружения лиганда mpl вне организма или в организме, включающий контактирование антитела с образцом, в частности, образцом сыворотки, подозреваемой в содержании лиганда, и обнаружении, если связывание происходит. В следующем предпочтительном варианте изобретение предлагает меченую или не меченую молекулу изолированной нуклеиновой кислоты, кодирующей лиганд mpl или его фрагменты, и молекулу нуклеиновой кислоты, имеющую последовательность, комплементарную или гибридизирующую в строгих или умеренно строгих условия х молекулу н уклеиновой кислоты, имеющую последовательность, кодирующую ли ганд mpl. Предпочтительной нуклеиновой кислотой лиганда mpl является РНК или ДНК, кодирующая биологически активный лиганд mpl и имеющая как минимум 75%, более предпочтительно минимум 80%, еще более предпочтительно минимум 90% и наиболее предпочтительно минимум 95% идентичности с последовательностью человеческого лиганда mpl. Более предпочтительными молекулами изолированных нуклеиновых кислот являются последовательности ДНК, кодирующие биологически активный лиганд mpl, выбранные из (а) ДНК, основанной на кодирующем участке гена лиганда mpl млекопитающего (например, ДНК, содержащая нуклеотидную последовательность, представленную на Фиг.1 (номер последовательности: 2), или ее фрагменты); (b) ДНК, способная гибридизировать ДНК (а) в как минимум умеренно строгих условиях; (с) ДНК, являющаяся результатом вырождения ДНК (а) или (b) вследствие вырождения генетического кода. Предполагается, что новые лиганды mpl, описанные здесь, могут быть членами семейства лигандов или цитокинов, имеющих такую идентичность последовательности, чтобы их ДНК могла гибридизироваться с ДНК, представленной на Фиг.1 (номер последовательности: 2) (или ее комплементом или фрагментами) в слабо и умеренно строгих условиях. Таким образом, следующий аспект данного изобретения включает ДНК, которая гибридизируется в нестрогих и умеренно строгих условия х с ДНК, кодирующей полипептиды лигандов mpl. В следующем предпочтительном варианте осуществления данного изобретения молекулой нуклеиновой кислоты является кДНК, кодирующая лиганд mpl и, кроме того, содержащая воспроизводимый вектор, в котором кДНК оперативно связана с контрольными последовательностями, распознаваемыми хозяином, трансформированным вектором. Этот аспект включает клетки хозяина, трансформируемые вектором, и способ использования кДНК для влияния на продуцирование лиганда mpl, предусматривающий экспрессию кДНК, кодирующей лиганд mpl, в культуре трансформированных клеток хозяина и извлечение лиганда mpl из культуры клеток хозяина. Лигандом mpl, приготовленным таким образом, предпочтительно является в основном гомогенный человеческий лиганд mpl. Предпочтительной хозяйской клеткой для продуцирования лиганда mpl является клетка яичника китайского хомяка (СНО). Далее изобретение включает предпочтительный способ лечения млекопитающих, страдающих иммунологическими или гематопоэтическими расстройствами, в частности, тромбоцитопенией, включающий введение млекопитающему терапевтически эффективного количества лиганда mpl. Возможно применение лиганда mpl в сочетании с цитокином, в частности, колониестимулирующим агентом или интерлейкином. Предпочтительными колониестимулирующими, агентами или интерлейкинами являются: кит-лиганд, LIF, GCSF, GM-CSF, M-CSF, ЕРО, IL-1, IL-2, IL-3, IL-5, IL-6, IL-7, IL-8, IL-9 или IL-11. III. Способы осуществления Долгое время некоторыми авторами предполагалось, что продуцирование тромбоцитов регулируется многовидовыми специфическими гуморальными агентами. Было постулировано, что две различных цитокиновых активности, называемые мегакариоцито-колониестимулирующий агент (meg-CSF) и тромбопоэтин, регулируют мегакариоцитопоэз и тромбопоэз [Williams et al., J. Cell Physiol., 110:101-104 (1982); Wiliiams et al., Blood Cells, 15:123-133 (1989); and Gordon et al., Blood, 80:302-307 (1992)]. Согласно этой гипотезе, meg-CSF стимулирует размножение предшественников мегакариоцитов, в то время как тромбопоэтин сначала влияет на созревание более дифференцированных клеток, а, в конечном счете, высвобождает тромбоциты. Начиная с 60-х годов, возникновение и признаки активности meg-CSF и тромбопоэтина в плазме, сыворотке и моче животных и людей, изученные на случаях тромбоцитопении, были хорошо описаны [Odell et al., Proc. Soc. Exp. Biol. Med., 108:428-431 (1961); Nakeff et al., Acta Haematol., 54:340344 (1975); Specter, Proc. Soc. Exp. Biol., 108:146-25 149 (1961); Schreiner et al., J. Clin. Invest., 49:1709-1713 (1970); Ebbe, Blood, 44:605-608 (1974); Hoffman et al., N. Engl. J. Med., 305:533 (1981); Straneva et al., Exp. Hematol., 17:1122-1127 (1988); Mazur et al., Exp. Hematol., 13:1164 (1985); Mazur et al., J. Clin. Invest., 68:733741 (1981); Sheiner et al., Blood, 56:183-188 (1980); Hill et al., Exp. Hematol., 20:354-360 (1992); and Kegyi et al., Int. J. Cell Cloning, 8:236-244 (1990)]. Эти активности были описаны как видо-специфические и отличные от известных цитокинов [Hill R.J. et al., Blood 80:346 (1992); Erickson-Miller C.L. et al., Brit. D. Haematol., 84:197-203 (1993); Straneva J.E. et al., Exp. Hematol. 20:4750 (1992); and Tsukada J. et al., Blood 81:866-867 (1993)]. Поэтому, попытки выделить meg-CSF или тромбопоэтин из тромбоцитопенической плазмы или мочи были безуспешными. В соответствии с выше упомянутыми исследованиями, описывающими тромбоцитопеническую плазму, мы обнаружили, что апластическая свиная плазма (АРР), полученная от облученных свиней, стимулирует человеческий мегакариоцитопоэз вне организма. Мы обнаружили, что эта стимуляторная активность аннулируется растворимым внеклеточным доменом c-mpl, подтверждая, что АРР является потенциальным источником предполагаемого лиганда mpl (ML). Тогда мы успешно выделили лиганд mpl из АРР, и информация последовательности аминокислот была использована для изоляции кДНК мышиного, свиного и человеческого ML. Эти ML имели последовательности, гомологичные эритропоэтину и обладали meg-CSF и тромбопоэтиноподобной активностями. 1. Выделение и идентификация лиганда mpl из плазмы Как показано выше, апластичекая плазма из различных видов животных обладает активностью стимуляции гематопоэза вне организма, однако ни одного гематопоэтического стимуляторного агента, изолированного из плазмы, прежде описано не было. Один источник получения апластической плазмы - от облученных свиней. Эта апластическая свиная плазма (АРР) стимулирует человеческий гематопоэз вне организма. Чтобы определить, содержит ли АРР лиганд mpl, ее эффект был исследован измерением инкорпорации Н-тимидина в клетки Ba/F3, подвергнутые трансфекции. человеческим mpl Ρ (Ba/F3-mpl), при помощи процедуры, показанной на Фиг.2. АРР стимулир ует инкорпорацию 3Н-тимидина в клетки (Ba/F3-mpl), но не в контрольные клетки Ba/F3 (т.е., не зараженные человеческим mpl Ρ). Кроме того, в нормальной свиной плазме подобной активности не наблюдалось. Эти результаты показывают, что АРР содержит агент или агенты, которые передают пролиферативный сигнал через рецептор mpl и поэтому могут быть природными лигандами для этого рецептора. Это предположение подтвердилось, когда было обнаружено, что лечение АРР растворимым mpl-IgG прекращает стимуляторное действие АРР на клетки (Ba/F3-mpl). Активный агент АРР представляется белком, поскольку проназа, DTT или нагрев уничтожают активность АРР (Фиг.3). Этот агент также не подвергался диализу. Однако, активность была стабильна до низких рН (рН2,5 в течение 2ч.) и показывала способность связываться и элюироваться из нескольких колонок лектинсродства, доказывая, что это гликопротеин. Дальнейшее изучение структуры и идентификация этой активности проводилось путем ее выделения из АРР с использованием химеры mpl-IgG. АРР обрабатывали в соответствии с порядком, описанным в Примерах 1 и 2. В кратком изложении, лиганд mpl выделяли с использованием гидрофобной интерактивной хроматографии (НІС), иммобилизовали с использованием окрашивающей хроматографии и хроматографии по mpl-сродству. Выделение активного агента на каждом этапе показано на рис. 4 , а выделение складок - в таблице 1. Общее количество активного агента, выделенного через колонку mpl-сродства, составило около 10%. Фракция пиковой активности (F6) из колонки mpl-сродства имеет расчетную величину 9,8´106 единиц/мг. Общее выделение из 5 литров АРР составило около 4´106 складок (0,8 единиц/мг к 3,3´106 единиц/мг) с восстановлением 83´106 складок в белок (250г на 3мкг). Мы оценили специфическую активность лиганда, элюированного из колонки mpl-сродства, в 3´10ед./мг. Таблица 1 Выделение лиганда mpl Образец АРР Фенил Blue-Sep. mpl (місі) (Fxns 5-7) Объем мл 5000 4700 640 12 Белок мг/мл 50 0.8 0.93 5´10-4 Ед/мл Единиц 40 40 400 1666 200 000 200 000 256 000 20 000 Специфическая активность, ед/мг 0.8 50 430 3 300 000 Выход % 94 128 10 Выделение 1 62 538 4 100 000 Белок определяли анализом Bradford. Концентрацию белка фракций 5-7 mpl-элюирования оценивали на основе интенсивности серебряного окрашивания SDS-геля. Одна единица определяется как вызывающая 50% максимального стимулирования размножения клеток Ba/F3-mpl. Анализ фракций, элюированных из колонки mpl-сродства (SDS-PAGE, 4-20%, гель Novex) в условиях восстановителя, обнаружил присутствие нескольких белков (Фиг.5). Белки с интенсивным серебряным окрашиванием показали средние величины Mr 66000, 55000, 30000, 28000 и 18000-22000. Чтобы определить, какой из этих белков стимулирует размножение клеточной культуры Ba/F3-mpl, белки элюировали из геля, как описано в Примере 2. Результаты опыта показали, что основная часть активного агента была элюирована из слоя геля, содержавшего белки с Mr 28000-32000, меньшая активность - из слоя с 18000-22000 (Фиг.6). Единственные белки, видимые в этих участках, имели Mr 30000, 28000 и 18000-22000. Чтобы идентифицировать и получить белковую последовательность для белков, найденных в этом участке геля (т.е., полосы при 30, 28 и 1822кДа), эти три белка подвергли электроблоттингу в PVD F и определили последовательность, как описано в Примере 3. Полученные амино-конечные последовательности приведены в таблице 2. Таблица 2 Амино-конечные последовательности лиганда mpl послед.№30 послед.№31 послед.№32 Компьютерный анализ показал, что эти аминокислотные последовательности найдены впервые. Поскольку все три последовательности одинаковы, было сделано предположение, что белки 30, 28 и 1822кДа родственные и могут являться различными формами одного вновь открытого белка. Более того, этот белок (белки) и представлялся природным лигандом mpl из-за его активности, определенной на SDS-PAGE в том же участке (28000-32000) 4-20% геля. Кроме того, при гель-фильтрационной хроматографии с использованием колонки Superose 12 (Pharmacia) был получен частично очищенный лиганд с Mr 17 000-30 000. Предполагается, что различные Mr-формы лиганда являются результатом протеолиза или гликозилирования или других пост- или предтрансляционных модификаций. Как описано ранее, РНК обратного человеческого mpl аннулирует мегакариоцитопоэз в культурах человеческого костного мозга, обогащенных родительскими клетками CD 34+, не влияя на видоизменение други х видов кроветворных клеток (Methla et al., выше). Этот результат приводит к предположению, что рецептор mpl может играть некоторую роль в видоизменении и размножении мегакариоцитов вне организма. Чтобы полнее изучить роль лиганда mpl в мегакариоцитопоэзе, сравнили влияние АРР и АРР, лишенной лиганда mpl, на человеческий мегакариоцитопоэз вне организма. Влияние АРР на человеческий мегакариоцитопоэз определяли с использованием модификации анализа жидкостно-суспензионного мегакариоцитопоэза, описанной в Примере 4. При этом анализе клетки человеческого периферического ствола (PSC) обрабатывались АРР до и после хроматографии по mpl-IgG-сродству. Стимуляция GPIIbllia мегакариоцитопоэза подсчитывали при посредстве антитела 125I-anti-IIb lllа (Фиг.7). Как показано на Фиг.7, 10% АРР вызывает приблизительно 3-кратное усиление стимуляции, в то время как АРР, ли шенная лиганда mpl, не оказывает никакого влияния. Примечателен также тот факт, что АРР, лишенная лиганда mpl, не вызывает размножения клеток Ba/F3-mpl. В другом опыте растворимый человеческий mpl-IgG, добавляемый на 0, 2 и 4 день к культурам, содержащим 10% АРР, нейтрализовал стимуляторное действие АРР на человеческий мегакариоцитопоэз (Фиг.8). Эти результаты показывают, что лиганд mpl играет определенную роль в регулировании человеческого мегакариоцитопоэза и поэтому может быть использован при лечении тромбоцитопении. 2. Молекулярное клонирование лиганда mpl На основе амино-конечной аминокислотной последовательности, полученной из белков 30, 28 и 18-22кДа (см. Таблицу 2), были построены два вырожденных олигонуклеотидных первичных фонда, используемые для усиления свиной геномной ДНК при помощи PCR. Разумно было предположить, что если амино-конечная аминокислотная последовательность кодируется отдельным экзоном, то длина точного продукта PCR должна составлять 69 оснований. Был найден фрагмент ДНК такого размера и субклонирован в pGEMT. Последовательности олигонуклеотидных инициаторов PCR и три полученных клона показаны в Примере 5. Аминокислотная последовательность (PRLLNKLLR [номер последовательности: 33]) пептида, закодированного между инициаторами PCR, оказалась идентичной последовательности, полученной путем амино-конечного белкового секвенирования свиного лиганда (см. остатки 9-17 в последовательностях свиных белков 28 и 30кДа). Синтетический олигонуклеотид, основанный на последовательности фрагмента PCR, был использован для просеивания библиотеки человеческих геномних ДНК. Был сконструирован 45-мерный олигонуклеотид, обозначенный pR45, и синтезирован на базе последовательности фрагмента, PCR. Этот олигонуклеотид имел следующую последовательность: 5'GCC-GTG-AAG-GAC-GTG-GTC-GTC-ACG-AAG-C AG-TTT-ATT-ТAG-GAG-TCG 3' послед.№34 Этот дезоксиолигонуклеотид был использован для отбора библиотеки человеческих геномних ДНК в ggem12 в нестрогих условиях гибридизации и промывки в соответствии с Примером 6. Позитивные клоны были собраны, очищены и проанализированы методами ограниченного картирования и южного блоттинга. Фрагмент EcoRl-Xbal (390 оснований), гибридизированный с 45-мером, был субклонирован в pBluescript SK-. Последовательность ДНК этого клона подтвердила, что ДНК, кодирующая человеческий гомолог свиного лиганда mpl, изолирована. Последовательность человеческой ДНК и выведенная аминокислотная последовательность показаны на рис. 9 (номера последовательностей: 3 и 4). Предсказанные положения интронов в геномной последовательности также указаны стрелками и определяют предполагаемый экзон ("экзон 3"). На основе последовательности человеческого "экзона 3" (Пример 6) были синтезированы олигонуклеотиды, соответствующие концам 3' и 5' последовательности экзона. Эти 2 инициатора были использованы в реакциях PCR в качестве шаблона кДНК, приготовленного из различных тканей человека. Ожидаемый размер точного продукта PCR составлял 140 оснований. После анализа продуктов PCR на 12% полиакриламидном геле фрагмент ДНК ожидаемого размера был обнаружен в библиотеках кДНК, полученных из взрослой человеческой почки, клетках 293 зародышевой почки и кДНК, полученной из человеческой зародышевой печени. Библиотека кДНК зародышевой печени (7´106 клонов) в лямбдa-DR2 была, в свою очередь, просеяна при помощи того же 45-мерного олигонуклеотида, используемого для просеивания человеческой геномной библиотеки и библиотеки кДНК зародышевой печени в нестрогих условиях гибридизации. Позитивные клоны были собраны, очищены, и размер вставки был определен по PCR. Один клон со вставкой 1,8kb был выбран для дальнейшего анализа. Используя процедуры,·описанные в Примере 7, были получены нуклеотид и выведенная аминокислотная последовательность человеческого лиганда mpl (hML). Эти последовательности представлены на Фиг.1 (номера последовательностей: 1 и 2). 3. Структура человеческого лиганда mpl (hML) Человеческий лиганд mpl (hML) последовательности кДНК, Фиг.1 (номер последовательности: 2), включает 1774 нуклеотида, за которыми следует поли (А) хвост. Он содержит 215 нуклеотидов нетранслируемой последовательности 5' и 498 нуклеотидов нетранслируемой последовательности 3'. Предполагаемый инициирующий кодон в нуклеотидной позиции (216-218) находится внутри общей типичной последовательности, предпочтительной для инициирования эукариотической трансляции. Открытая рамка считывания состоит из 1059 нуклеотидов и кодирует 353 аминокислотных полипептида, начиная с позиции нуклеотида 220. N-окончание предсказанной аминокислотной последовательности высокогидрофобно и, вероятно, соответствуе т сигнальному пептиду. Компьютерный анализ предсказанной аминокислотной последовательности [von Hei jne et al., Eur. J. Biochem., 133:17-21 (1983)] показывает потенциальный участок расщепления для сигнальной пептидазы между остатками 21 и 22. Расщепление в этой позиции будет генерировать зрелый полипептид из 332 аминокислотных остатков, начиная с амино-конечной последовательности, полученной из лиганда mpl, очищенного из свиной плазмы. Предсказанный негликозилированный молекулярный вес лиганда из 332 аминокислотных остатков - около 38кДа. Существует 6 потенциальных участков N-гликозилирования и 4 цистеиновых остатка. Сравнение последовательности лиганда mpl с базой данных последовательностей Genbank обнаруживает 23% идентичности между амино-конечными 153 остатками зрелого человеческого лиганда mpl - и человеческим эритропоэтином, Фиг.10 (номера последовательности: 6 и 7). Если принять в расчет консервативные заместители, этот участок hML показывает 50% подобия человеческому эритропоэтину (hEPO). ЕРО и h ML содержат 4 цистеина. Три из четырех цистеинов в hML законсервированы, включая первый и последний цистеин. Целенаправленные мутагенные эксперименты показали, что первый и последний цистеины эритропоэтина образуют дисульфидную связь, необходимую для функции [Wang, F.F. et al., Endocrinology 116:2286-2292 (1983)]. По аналогии, первый и последний цистеины hML тоже могут образовывать критическую дисульфидную связь. Ни один из участков гликозилирования в hML не законсервирован. Все потенциальные N-связанные участки гликозилирования hML расположены в карбоксиконечной половине полипептида hML. Подобно hEPO, мРНК hML не содержит ни общей типичной полиаденилированной последовательности AAU AAA, ни регуляторного элемента AUUUA, который присутствует в нетранслируемых участках 3' многих цитокинов и, как предполагается, оказывает влияние на стабильность мРНК [Shaw et al., Cell, 46:659-667 (1986)]. Анализ северного блоттинга показал низкие уровни транскрипции РНК hML (1,8kb) как зародышевой, так и во взрослой почках. После длительной выдержки во взрослой почке могла быть найдена более слабая полоса того же размера. При сравнении, человеческий эритропоэтин выражается в зародышевой печени и, как отклик на гипоксию, во взрослой почке и печени [Jacobs et al., Nature, 313:804-809 (198); Bondurant et al., Molec. Cell. Biol., 6:2731-2733 (1986)]. Важность С-конечной части hML е ще предстоит изучить. Судя по наличию шести потенциальных участков для N-гликозилирования и способности лиганда связываться с колонками лектин-сродства, эта часть hML, вероятно, гликозилирована. В некоторых опытах с гелевым элюированием мы наблюдали активность с Mr около 60 000, которая может представлять гликозилированную молекулу полной длины. Следовательно, Сконечная часть может служить для стабилизации и увеличения полупериода существования циркулирующего hML. В случае эритропоэтина негликозилированная форма обладает вне организма полной биологической активностью, но имеет значительно сниженный полупериод существования по сравнению с гликозилированным эритропоэтином [Takeuchi et al., J. Biol. Chem., 265:12127-12130 (1990); Narhi et al., J. Biol. Chem., 266:23022-23026 (1991); Spivack et al., Blood, 7:90-99 (1989)]. С-конечный домен hML содержит 2 диосновных аминокислотных последовательности (раппорт Arg-Arg в позициях 153-154 и 245-246), которые могут служить в качестве потенциальных рабочих участков. Расщепление на этих участках может отвечать за генерирование форм 30, 28 и 18-22кДа лиганда mpl, изолированного из АРР. Примечательно, что последовательность Аrg153-Arg154 располагается сразу после эритропоэтин-подобного домена ML. Эти наблюдения показывают, что ML полной длины может представлять собой белок-предшественник, который подвергается ограниченному протеолизу для генерирования зрелого лиганда. 4. Изоформы и варианты человеческого лиганда mpl Изоформы, или альтернативно сращенные формы человеческого лиганда mpl были обнаружены при помощи PCR в человеческой взрослой печени. В кратком изложении, инициаторы были синтезированы в соответствии с каждым концом кодовой последовательности hML, так же как были выбраны внутренние участки последовательности. Эти инициаторы использовали в RT-PCR для усиления РНК взрослой человеческой печени, как описано в Примере 10. Кроме формы полной длины, обозначенной hML, исследовали или выводили три других формы, обозначенные hML2, hML3, h ML4. Зрелые выведенные аминокислотные последовательности всех четырех изоформ представлены на Фиг.11 (номера последовательностей: 6, 8, 9 и 10). hML3 имеет делецию 116 нуклеотидов в позиции 700, что приводит к аминокислотной-делеции и сдвигу рамки. Теперь кДНК кодирует зрелый полипептид длиной в 265 аминокислот, являющийся производным от последовательности hML в позиции аминокислотного остатка 139. Наконец, hML4 имеет делецию 12 нуклеотидов после нуклеотидной позиции 618 (также обнаруженную в последовательностях мыши и свиньи (см. ниже) и делецию 116 оснований, обнаруженную в hML3. Хотя ни один клон с единственной делецией 12 оснований (после нуклеотида 619) не был выделен в человеке (обозначение hML2), эта форма, вероятно, существует, поскольку такая изоформа была идентифицирована у мыши и свиньи (см. ниже), и поскольку она была идентифицирована в сочетании с делецией 116 в hML4. Были сконструированы замещенный вариант hML, в котором диосновная последовательность Arg153"Arg154 заменена двумя аланиновыми остатками, и усеченная форма "ЕРО-домена" hML, чтобы определить, необходима ли полная длина ML для биологической активности. Замещенный вариант диосновной последовательности Arg153"Arg 154 , называемый hML(R153A, R154A) был сконструирован с использованием PCR, как описано в Примере 10. Усеченная форма "ЕРО-домена", hML153 , также была создана с использованием PCR при введении стопорного кодона после Arg153 . 5. Экспрессия рекомбинантного человеческого лиганда mpl (rhML) во временно трансфецированных клетках (293) человеческой эмбриональной почки Чтобы убедиться, что клонированная человеческая кДНК кодирует лиганд для mpl, лиганд был экспрессирован в клетках 293 млекопитающего под контролем непосредственного цитомегаловирусного промотора ранней стадии, с использованием векторов экспрессии pRK5-hML или pRK5-hML153. Было обнаружено, что супернатанты из временно трансфецированных человеческих эмбриональных клеток 293 почки стимулируют инкорпорацию 3Н-тимидина в клетки Ba/F3-mpl, но не в родительские клетки Ba/F3 (Фиг.12А). Среда из клеток 293, трансфецированных одним вектором pRK, не содержала этой активности. Добавление к среде mpl-IgG аннулировало стимуляцию (данные не показаны). Эти результаты показывают, что клонированная кДНК кодирует функциональный человеческий ML (hML). Чтобы определить, может ли "ЕРО-домен" самостоятельно связывать и активировать mpl, в клетках 293 была экспрессирована усеченная форма hML, rhML153. Было обнаружено, что супернатанты из трансфецированных клеток обладают активностью, подобной активности, присутствующей в супернатантах из клеток, выражающих h ML полной длины (Фиг.12А), что показывает, что С-конечный домен ML не обязателен для связывания и активации c-mpl. 6. Лиганд mpl стимулирует мегакариоцитопоэз и тромбопоэз Как rhML, так и усеченные формы rhML153 рекомбинантного hML стимулировали человеческий мегакариоццтопоэз вне организма (Фиг.12В). Этот эффект наблюдался в отсутствие други х экзогенно добавляемых агентов гематопоэтического роста. За исключением IL-3, mpl был единственным испытанным агентом гематопоэтического роста, проявляющим такую активность. IL-11, IL-6, Il-1, эритропоэтин, G-CSF, Il-9, LIF, кит-лиганд (KL), M-CSF, OSM и GM-CSF не оказывали влияния на мегакариоцитопоэз при отдельных испытаниях в нашем исследовании (данные не показаны). Этот результат показывает, что ML обладает мегакариоцитостимулирующей активностью и определяет для ML роль в регулировании мегакариоцитопоэза. Тромбопоэтическая активность, присутствующая в плазме тромбоцитопенических животных, проявила способность стимуляции продуцирования тромбоцитов при анализе по методу мышиного образца ответных тромбоцитов [McDonald, Proc. Soc. Exp. Biol. Med., 14:1006-1001 (1973) и McDonald et al., Scand. 3. Haematol., 16:326-334 (1976)]. В этой модели мышей подвергли острой тромбоцитопении, используя особую антитромбоцитную сыворотку, что приводит к предсказуемому ответному продуцированию тромбоцитов. Такие иммуно-тромбоцитопенические мыши более чувствительны к экзогенным тромбопоэтиноподобным активностям, чем нормальные мыши [McDonald, Proc. Soc. Exp. Biol. Med., 14:1006-1001 (1973)], как эксгипоксические мыши более чувствительны к эритропоэтину, чем нормальные мыши [McDonald, et al., J. Lab. Clin. Med., 77:134-143 (1971)]. Чтобы определить, стимулирует ли rML продуцирование тромбоцитов в организме, мышам с ответным тромбоцитозом вводили частично очищенный rhML. Затем посчитали количество тромбоцитов и инкорпорацию 35S в тромбоциты. Введение мышам 64000 или 32000 единиц rML значительно увеличило продуцирование тромбоцитов, что очевидно из 20%-го увеличения количества тромбоцитов (р=0,0005 и 0,0001 соответственно) и 40%-го увеличения инкорпорации 35S в тромбоциты (р=0,003) у обработанных мышей по сравнению с контрольными мышами, .которым вводили только наполнитель (Фиг.12С). Этот уровень стимуляции сравним с уровнем, наблюдаемым в этой модели при применении IL-6 (данные не показаны). Введение 16000 единиц заметно не стимулировало продуцирование тромбоцитов. Эти результаты показывают, что ML стимулируе т продуцирование тромбоцитов в зависимости от дозы и поэтому обладает тромбопоэтиноподобной активностью. Клетки 293 подвергли также трансфекции другими изоформами hML, описанными выше, и исследовали супернатанты путем анализа размножения клеток Ba/F3-mpl (Фиг.13). Показанные на рисунке hML2 и hML3 не проявили ощутимой активности при этом анализе, однако активность hML(R153A, R154A) была подобна hML и hML153 , показывая, что процессы на диосновном участке Arg153-Arg154 ни необходимы, ни вредны для активности. 7. Мегакариоцитопоэз и лиганд mpl Было сделано предположение, что мегакариоцитопоэз регулируется на многих клеточных уровнях [Williams et al., Bood Cells, 15:123-133 (1989)]. В основном это предположение базируется на наблюдении, что определенные агенты гематопоэтического роста стимулируют размножение предшественников мегакариоцитов, в то время как другие главным образом влияют на развитие клеток в зрелую форму. Результаты, представленные здесь, приводят к выводу, что ML действует как агент размножения, так и развития. То, что ML стимулир ует размножение, доказывает несколько фактов. Во-первых, АРР стимулир ует и размножение, и развитие человеческих мегакариоцитов вне организма, и эта стимуляция полностью ингибируется mpl-IgG (Фиг.7 и 8). Кроме того, ингибирование образования колонии мегакариоцитов обратными олигонуклеотидами c-mpl [Methia et al., Blood, 82:1395-1401 (1993)] и открытие факта, что c-mpl может переносить пролиферативный сигнал в клетки, в которые он вносится [Scoda et al., EMBO, 12:26452653 (1993) и Vigon et al., Oncogene, 8:2607-2615 (1993)] также означает, что ML стимулирует размножение. Очевидная экспрессия c-mpl на всех стадиях видоизменения мегакариоцитов [Methia et al., Blood, 82:13951401 (1993)] и способность рекомбинантного ML ускоренно стимулировать продуцирование тромбоцитов в организме означает, что ML также влияет и на развитие. Доступность рекомбинантного ML делает возможным тща тельное изучение его роли в регулировании мегакариоцитопоэза и тромбоцитопоэза, а также его потенциальной способности влиять на другие виды кроветворных клеток. 8. Изоляция гена человеческого лиганда mpl (TPO). Клоны человеческой геномной ДНК гена ТРО были изолированы путем просеивания человеческой геномной библиотеки в lgem12 с pR45, в нестрогих или в строгих условия х с фрагментом, соответствующим половине 3' последовательности человеческой кДНК для лиганда mpl. Были изолированы два перекрывающихся l-клона размером в 35kb. Два перекрывающихся фрагмента (BamH1 и EcoRI), содержащие полный ген ТРО, были субклонированы и секвенированы (см. Фиг.14А, 14В и 14С). Структура человеческого гена включает 6 экзонов в пределах 7kb геномной ДНК. Границы всех экзон/интрон-переходов содержат общий для генов млекопитающих раппорт [Shapiro, Μ.В., et al., Nucl. Acids Res. 15:7155 (1987)]. Экзон 1 и экзон 2 содержат нетранслируемую последовательность 5' и четыре начальных аминокислоты сигнального пептида. Остальная часть секреторного сигнала и первые 26 аминокислот зрелого белка кодируются в пределах экзона 3. Целый карбоксильный домен и нетранслируемая последовательность 3', а также около 50 аминокислот эритропоэтин-подобного домена кодируются в пределах экзона 6. Четыре аминокислоты, участвующие в делеции, наблюдаемой в пределах hML-2 (hTPO-2), расположены в конце 5' экзона 6. Анализ человеческой геномной ДНК методом южного блоттинга показал, что ген для ТРО присутствует в единственной копии. Хромосомное расположение гена было определено флуоресцентной гибридизацией на месте, (FISH) и отнесено к хромосоме 3q27-28. 9. Экспрессия и очистка ТРО из клеток 293 Приготовление и очистка ML ТРО из клеток 293 детально описано в Примере 19. В кратком изложении, кДНК, соответствующая целой открытой считывающей рамки ТРО, была получена при помощи PCR с использованием pRK5-hml I. Продукт PCR был очищен и клонирован между участками ограничения Clal и Xbal плазмида pRK5tkneo (pRKS-производный вектор, модифицированный для выражения неомицинрезистентного гена в усло виях активации тимидинкиназой) для получения вектора pRKStkneo.ORF (векторная последовательность для целой открытой считывающей рамки). Вторая векторная последовательность для гомологичного домена ERO была генерирована тем же путем, но с использованием других инициаторов PCR, для получения окончательной конструкции, называемой pRK5tkneoEPO-D. Этими двумя конструкциями были заражены клетки человеческой эмбриональной почки методом СаРО4, после чего отобрали неомицин-резистентные клоны и допустили их рост до слияния. С использованием анализа размножения Ba/F3-mpl оценили экспрессию МL153 или ML154 из этих клонов в кондиционированной среде. Очистка rhML332 проводилась, как описано в Примере 19. В кратком изложении, кондиционированную среду 293-rhML332 ввели в колонку Blue-Sepharose (Pharmacia) и затем промывали буфером, содержащим 2М мочевины и 1М NaCl. Затем элюат из Blue-Sepharose непосредственно перенесли в колонку WGA-Sepharose, промыли 10 колоночными объемами буфера, содержащего 0,5М N-ацетил-D-глюкозамина. Элюат из WGASepharose перенесли в колонку C4-HPLC (Synchrom, Inc.) и элюировали с непрерывным пропанольным градиентом. Посредством SDS-PAGE очи щенный 293-rhML332 мигрирует в виде широкой полосы в участок геля 68-80кДа (см. Фиг.15). Очистка rhML153 также проводилась, как в Примере 19. В кратком изложении, кондиционированную среду 293-rhML153 элюировали в колонке Blue-Sepharose, как описано для rhML332. Затем элюат из Blue-Sepharose непосредственно перенесли в колонку mpl-сродства, как описано выше. rhML153, элюированный из колонки mpl-сродства, подвергли очистке до гомогенности, используя колонку C4-HPLC в тех же условиях, что и для rhML332. Посредством SDS-PAGE очищенный 293-rhML153 мигрирует в две широких и две узких полосы с Mr около 18 000-22 000 (см. Фиг.15). 10. Лиганд mpl мыши Фрагмент ДНК, соответствующий кодированному участку лиганда mpl, была получена при помощи PCR, очищен и помечен в присутствии 32P-dATP и 32P-dCTP. Этот образец был использован для просеивания 106 клонов библиотеки кДНК печени мыши в lGT10. Клон мыши, Фиг.16 (номера последовательностей: 12 и 13), содержащий 1443 вставленных пар оснований, был изолирован и секвенирован. Предполагаемый инициирующий кодон в нуклеотидной позиции 138-141 находился в пределах общей типичной последовательности, подходящей для эукариотического трансляционного инициирования [Kozak, Μ. J. Cell Biol., 108:229-241 (1989)]. Эта последовательность определяет открытую рамку считывания 1056 нуклеотидов, которая предопределяет первичный трансляционный продукт 352 аминокислот. Края этой рамки образуют 137 нуклеотидов нетранслируемой последовательности 5' и 247 нуклеотидов последовательности 3'. За нетранслируемым участком 3' ненаблюдается поли(А) хвоста, показывая, что клон, вероятно, неполный. N-окончание предсказанной аминокислотной последовательности высокогидрофобно и, вероятно, представляет сигнальный пептид. Компьютерный анализ [von Hei jne, G. Eur. J. Biochem. 133:17-21 (1983)] показывает потенциальный участок расщепления для сигнальной пептидазы между остатками 21 и 22. Расщепление в этой позиции будет генерировать зрелый полипептид из 331 аминокислоты (35кДа), идентифицированный как mML331 (или mML2 по причинам, изложенным ниже). Последовательность содержит 4 цистеина, все законсервированы в человеческой последовательности, и семь потенциальных Nгликозилированных участков, 5 из которых законсервированы в человеческой последовательности. И снова, как в случае hML, все семь потенциальных N-гликозилированных участков расположены в С-конечной половине белка. При сравнении с человеческим ML была обнаружена значительная идентичность для нуклеотидной и выведенной аминокислотной последовательностей в "ЕРО-доменах" этих ML. Однако, при совмещении выведенных аминокислотных последовательностей человеческого и мышиного ML мышиная последовательность показала наличие тетрапептидной делеции между остатками 111-114, соответствующими 12 нуклеотидным делециям после позиции 618 в человеческой (см. выше) и свиной (см. ниже) кДНК. В соответствии с этим были исследованы дополнительные клоны для обнаружения возможных изоформ ML мыши. Один клон кодировал полипептид из 335 аминокислотных выведенных последовательностей, содержащий "отсутствующий" тетрапептид LPLQ. Эта форма предположительно является мышиным ML полной длины и названа mML или mML335. Нуклеотидная и выведенная аминокислотная последовательность для mML представлены на Фиг.17 (номера последовательностей: 14 и 15). Этот клон кДНК состоит из 1443 пар оснований и оканчивается поли(А) хвостом. Он содержит открытую рамку считывания (1068 оснований). Края этой рамки образуют 134 основания нетранслируемой последовательности 5' и 241 основание последовательности 3'. Предполагаемый инициирующий кодон находится в нуклеотидной позиции 138-140. Открытая рамка считывания кодирует предсказанный белок из 356 аминокислот, первые 21 из которых высокогидрофобны и функционируют, скорее всего, как секреторный сигнал. Наконец, был изолирован и секвенирован третий клон мыши, содержащий делецию 116 нуклеотидов, соответствующую h ML3. Поэтому данная мышиная изоформа была обозначена mML3. Сравнение выведенных аминокислотных последовательностей этих двух изоформ показано на Фиг.18 (номера последовательностей: 9 и 16). Общая идентичность аминокислотных последовательностей между ML человека и мыши, Фиг.19 (номера последовательностей 6 и 17), составляет 72%, но эта гомология неравномерно распределена. Участок, определенный как "ЕРО-домен" (аминокислоты 1-153) для человеческой последовательности и 1-149 для мышиной) лучше законсервирован (86% гомологии), чем карбокси-конечный участок белка (62% гомологии). Это служит еще одним доказательством того, что только "ЕРО-домен" важен для биологической активности белка. Интересно, что из двух раппортов диосновных кислот, найденных в h ML, только диосновный раппорт, следующий сразу за "ЕРО-доменом" (позиции остатков 153-154) в человеческой последовательности, присутствует в мышиной последовательности. Это согласуется с предположением, что ML·- полной длины может представлять белок-предшественник, который подвергается ограниченному протеолизу, генерируя зрелый лиганд. Как альтернатива, протеолиз между Arg 153 и Arg154 может образовать зазор hML. Вектор экспрессии, содержащий целую кодирующую последовательность mML, временно ввели в клетки 293, как описано в Примере 1. Кондиционированная среда из этих клеток стимулировала инкорпорацию 3Нтимидина в клетки Ba/F3, выражающие mpl мыши или человека, но не влияла на родительскую (не содержащую mpl) клеточную линию. Это показывает, что клонированная кДНК mpl мыши кодирует функциональный лиганд, способный активировать рецептор ML (mpl) как мыши, так и человека. 11. Свиной лиганд mpl Свиную кДНК ML (pML) изолировали при помощи RACE PCR, как описано в Примере 13. кДНК-продукт размером 1342 оснований нашли в почке и субклонировали. Несколько клонов секвенировали и обнаружили свиной лиганд mpl из 332 аминокислотных остатков, обозначив его pML (или pML332), имеющий нуклеотидную и выведенную аминокислотную последовательность, показанную на Фиг.20 (номера последовательностей: 18 и 19). И снова была идентифицирована вторая форма, обозначенная pML2, кодирующая белок с делецией 4 аминокислот (228 аминокислотных остатков) (см. Фиг.21 (номер последовательности: 21)). Сравнение аминокислотных последовательностей pML и pML2 показывает, что последняя форма идентична, за исключением того, что тетрапептид QLPP, соответствующий остаткам 111-114, отсутствует (см. Фиг.22 (номера последовательностей: 18 и 21)). Четыре аминокислотных делеции, наблюдаемые в кДНК ML мыши и свиньи, находятся в одной и той же позиции в пределах предсказанных белков. Сравнение предсказанных аминокислотных последовательностей зрелого ML человека, мыши и свиньи, Фиг.19 (номера последовательностей: 6, 17 и 18), показывает, что общая идентичность последовательностей составляет 72% между мышью и человеком, 68% между мышью и свиньей и 73% между свиньей и человеком. Значительно большая гомология наблюдается в амино-конечной половине ML (ЕРО-гомологичный домен). Этот домен идентичен на 80-84% между любыми двумя видами, в то время как карбокси-конечная половина (углеводородный домен) показывает лишь 57-67% идентичности. Диосновный аминокислотный раппорт, который может представлять участок протеазного расщепления, находится на карбоксильном конце эритропоэтин-гомологичного домена. Этот раппорт законсервирован во всех трех видах в этой позиции [Фиг.19 (номера последовательностей: 6, 17 и 18)]. Второй диосновный участок, имеющийся в позициях 245 и 246 в человеческой последовательности, не содержится в последовательностях мыши и свиньи. Последовательность ML мыши и свиньи содержит 4 цистеина, законсервированные в человеческой последовательности. Имеется 7 участков N-гликозилирования в мышином лиганде и 6 - в свином. Все потенциальные участки N-гликозилирования снова находятся в С-конечной половине белка. 12. Экспрессия и очистка ТРО из клеток яичника китайского хомяка (СНО) Векторы экспрессии, используемые для трансфекции клеток СНО, обозначаются pSV15.ID.LL.MLORF (полная длина или ТРО332) и pSV15.ID.LL.MLEPO-D (усеченная форма или ТРО153). Представляющие интерес свойства эти х плазмидов показаны на Фиг.23 и 24. Процедуры трансфекции описаны в Примере 20. В кратком изложении, кДНК, соответствующую целой открытой рамке считывания ТРО, получили при помощи PCR. Продукт PCR очистили и клонировали между двумя участками ограничения (Clal и Sall) плазмида pSV15.lD.LL для получения вектора pSV15.ID.LL.MLORF. Вторую конструкцию, соответствующую ЕРО-гомологичному домену, генерировали тем же способом, но с использованием обратного инициатора (EPOD.Sal). Окончательная конструкция последовательности вектора для ЕРО-гомологичного домена ТОР названа pSV15.ID.LL.MLEPO-D. Эти две конструкции линеаризовали при помощи Notl и ввели в клетки яичника китайского хомяка [клетки CHO-DP12, ЕР 307,247 публ. 15 марта 1989] путем электропорации. 107 клеток электропорировали в устройстве электропорации BRL (350В, 330мФ, низкая емкость) в присутствии 10, 25 и 50мг ДНК, как описано в [Andreason, G.L. J. Tissue Cult. Meth. 15,56 (1993)]. На следующий день после трансфекции произвели расщепление клеток в избирательной среде DHFR (высокоглюкозный DMEM-F12 50:50 без глицина, 2мМ глютамина, 2-5% диализированной зародышевой бычьей сыворотки). Через 10-15 дней отдельные колонии перенесли в 96-луночные чашки и оставили расти до слияния. Экспрессию МL153 и ML 332 в кондиционированной среде из этих клонов оценили с использованием анализа размножения Ba/F3-mpl (описанного в Примере 1). Процесс очистки и изоляции ТРО из собранной жидкости клеточной культуры СНО описан в Примере 20. В кратком изложении, собранную жидкость клеточной структуры (HCCF) вводят в колонку Blue-Sepharose (Pharmacia) при соотношении около 100л HCCF на литр смолы. После этого колонку промывают 3-5 колоночными объемами буфера, затем 3-5 объемами буфера, содержащего 2,0Μ мочевины и 1,0М NaCl. Элюат из Blue-Sepharose, содержащий ТРО, переносят в колонку Wheat Germ Lectin Sepharose (Pharmacia), уравновешенную в элюирующем буфере Blue-Sepharose при соотношении 8-16мл элюата BlueSepharose на мл смолы. После этого колонку промывают 2-3 колоночными объемами балансировочного буфера. Затем ТРО элюируют 2-5 колоночными объемами буфера, содержащего 2,0М мочевины и 0,5М Nацетил-D-глюкозамина. Элюат Wheat Germ Lectin, содержащий ТРО, подкисляют и добавляют С 12Е8 к конечной концентрации 0,04%. Результирующий пул вводят в колонку обращенной фазы С4, уравновешенную в 0,1% TFA, 0,04% С12Е8 при нагрузке около 0,2-0,5мг белка на мл смолы. Белок элюируют в ацетонитриле с двухфазным линейным градиентом, содержащем 0,1% TFA и 0,04% C12Е8, и образуют п ул на основе SDS-PAGE. Затем полученный пул С4 разбавляют и диафильтруют примерно 6 объемами буфера на Amicon YM или подобной ультрафильтрационной мембране с отсечкой молекул весом 10000-30000 Дальтон. Результирующий диафильтрат можно затем непосредственно обрабатывать или далее концентрировать ультрафильтрацией. Диафильтрат/концентрат обычно регулируется до конечной концентрации 0,01% Tween80. Весь диафильтрат/концентрат или его часть, эквивалентную 2-5% рассчитанного объема колонки переносят в колонку Sephacryl S-300 HR (Pharmacia), уравновешенную в буфере, содержащем 0,01%, Tween80, и хроматографируют. ТРО-содержащие фракции, очищенные от скоплений и продуктов протеолитической деградации, объединяют в пул на основе SDS-PAGE. Результирующий пул фильтр уют и хранят при 2-8°С. 13. Способы трансформирования и инициирования синтеза ТРО в микроорганизме, а также изоляции, очистки и укладки ТРО, образованного в нем. Конструирование векторов экспрессии Е. соli ТРО детально описано в Примере 21. В кратком изложении, для выражения первых 155 аминокислот ТРО, начиная с низкомолекулярного лидера, разного для разных структур, были построены плазмиды рМР21, рМР151, рМР41, рМР57 и рМР202. Лидеры предназначены в первую очередь для высокоуровневого трансляционного инициирования и ускоренной очистки. Плазмиды рМР210-1, -Т8, -21, -22, -24, -25 сконструированы для выражения первых 153 аминокислот ТРО начиная с инициирующего метионина, и отличаются только частотой повторения кодона первых 6 аминокислот ТРО, в то время как плазмид рМР251 является производным от рМР21-1, в котором карбокси-терминированный конец ТРО увеличен на две аминокислоты. Все указанные плазмиды продуцируют высокие уровни внутриклеточной экспрессии ТРО в E. coli при инициировании триптофан-промотором [Yansura, D.G. et al. Methods in Enzymology (Goeddel, D.V., Ed.) 185:54-60, Academic Press, San Diego (1990)]. Плазмиды pMP1 и рМР172 являются промежуточными при конструировании вышеупомянутых плазмидов внутриклеточной экспрессии ТРО. Упомянутые плазмиды внутриклеточной экспрессии ТРО использовались для трансформирования Е. соli с использованием метода теплового удара СаСl2 [Mandel, Μ. et al. J. Моl. Biol., 53:159-162, (1970)] и других процедур, описанных в Примере 21. В кратком изложении, трансформированные клетки сначала выращивали при 37°C до тех пор, пока оптическая плотность (600нм) культуры не достигла примерно 2-3. Затем культуру разбавили и, после выращивания с аэрацией, добавили кислоту. Затем культуру оставили на 15 часов для дальнейшего роста с аэрацией, после чего клетки собрали путем центрифугирования. Процессы изоляции, очистки и конформационной укладки, приведенные ниже для получения биологически активного, соответствующим образом уложенного человеческого ТРО или его фрагментов описаны в Примерах 22 и 23 и могут быть использованы для извлечения любого варианта ТРО, включая Ν и С-конечные расширенные формы. Другие процессы, подходящие для укладки рекомбинантного или синтетического ТРО, см. в следующих патентах: Builder et al., U.S. Patent 4,511,502; Jones et al., U.S. Patent 4,512,922; Olson et al., U.S. Patent 4,518,526; Builder et al., U.S. Patent 4,620,948; общее описание процесса извлечения и конформационной укладки для различных рекомбинантных белков, выраженных в нерастворимой форме в Е. coli. А. Извлечение нерастворимого ТРО Микроорганизм, например, Е. coli, выражающий ТРО., закодированный любым подходящим плазмидом, подвергают ферментации в условиях, в которых ТРО осаждается в виде нерастворимых "тел отражения". Как вариант, клетки сначала промываются в деструктирующем клеточном буфере. Обычно около 100г клеток ресуспендируют в примерно 10 объемах клеточного деструктирующего буфера (например, 10мМ Tris, 5мМ EDTA, рН8) с гомогенизатором, например, Polytron, и центрифугируют при 5000´г в течение 30 минут. Затем клетки лизируют с использованием любой обычной технологии, такой как тонизирующий удар, обработка ультразвуком, циклическим давлением, химические или энзиматические методы. Например, вышеупомянутые промываемые клеточные гранулы можно суспендировать в следующи х 10 объемах клеточного деструктирующего буфера с гомогенизатором и пропустить суспензию через устройство деструктирования клеток LH Cell Disruoter (LH Inceltech, Inc>) или через Microfluidizer (Microfluidics International) в соответствии с инструкциями производителя. Затем особое вещество, содержащее ТРО, отделяется от жидкой фазы и, возможно, промывается любой подходящей жидкостью. Например, суспензия клеточного лизата центрифугир уется при 5000´г в течение 30мин., ресуспендируется и, возможно, центрифугир уется второй раз для образования промытых гранул "тел отражения". Промытый гранулят может быть использован сразу или замораживается для хранения (например, при -70°С). В. Солюбилизация и очистка мономерного ТРО Нерастворимый ТРО в грануляте "тел отражения" затем солюбилизируется солюбилизирующим буфером, который содержит хаотропический агент и обычно действует при щелочной рН и содержит восстановительный агент для повышения выхода мономерного ТРО. Характерными хаотропическими агентами являются мочевина, гуанидин НСІ и тиоцианат натрия. Предпочтительный хаотропический агент гуанидин НСl. Концентрация хаотропического агента составляет обычно 4-9М, предпочтительно 6-8М. рН солюбилизирующего буфера поддерживается любым подходящим буфером в диапазоне 7,5-9,5, предпочтительно 8,0-9,0 и наиболее предпочтительно - 8,0. Предпочтительно солюбилизирующий буфер также содержит восстановительный агент, способствующий образованию мономерной формы ТРО. Подходящие восстановительные агенты включают органические соединения, содержащие свободный тиол (RSH). Характерными восстановительными агентами являются дитиотреитол (DTT), дитиоэритритол (DTE), меркаптоэтанол, глютатион (GSH), цистеамин и цистеин. Предпочтительным восстановительным агентом является дитиотреитол (DTT). Как вариант, солюбилизирующий буфер может содержать мягкий окислительный агент (например, молекулярный кислород) и сульфитную соль для образования мономерного ТРО п утем сульфитолиза. В этом варианте результирующий TPO-S-сульфонат позднее подвергается укладке в присутствии окислительно-восстановительного буфера (например, GSH/GSSG) для формирования соответствующим образом сложенного ТРО. Белок ТРО обычно подвергается дальнейшей очистке с использованием, например, центрифугирования, гель-фильтрационной хроматографии и обращенно-фазовой колоночной хроматографии. Приведенная в качестве иллюстрации, следующая методика обеспечивает достаточный выход мономерного ТРО. Гранулят "тел отражения" ресуспендируются примерно в 5 объемах (вес.) солюбилизирующего буфера (20мМ Tris, pH8, с 6-8М гуанидина и 25мМ DTT) и перемешивается 1-3 часа, или в течение ночи, при 4°C для солюбилизации белка ТРО. При этом также полезны высокие концентрации мочевины (6-8М), но в результате это приводит к некоторому снижению выхода по сравнению с гуанидином. После солюбилизации раствор центрифугир уют при 30 000´г в течение 30мин. для получения прозрачного супернатанта, содержащего денатурированный мономерный белок ТРО. Затем супернатант хроматографируют в гель-фильтрационной колонке Superdex 200 (Pharmacia, 2,6´60см) при скорости течения 2мл/мин и элюируют 20мМ фосфа та натрия при рН6,0 с 10мМ DTT. Собирают фракции, содержащие мономерный денатурированный белок ТРО (элюированные между 1.60 и 200мл). Белок ТРО подвергают дальнейшей очистке в полу-подготавливаемой колонке обращенной фазы С4 (VYD AC 2´20см). При скорости 5мл/мин, образец обрабатывается в колонке, уравновешенной в 0,1% TFA (три фтор уксусной кислоты) с 30% ацетонитрила. Белок элюируют с линейным градиентом ацетонитрила (30-60% за 60мин.). Очищенный восстановленный белок элюируют с примерным содержанием ацетонитрила 50%. Этот материал используется для укладки и получения биологически активного варианта ТРО. С. Укладка ТРО для генерирования биологически активной формы После солюбилизации и дальнейшей очистки ТРО получают биологически активную форму п утем укладки денатурированного мономерного ТРО в окислительно-восстановительном буфере. Благодаря высокой активности ТРО (половина максимальной стимуляции в клетках Ba/F3 достигается приблизительно при 3пг/мл), возможно получение биологически активного материала при использовании многих различных буферов, детергентов и окислительно-восстановительных условий. Однако в большинстве таких сред получаются лишь небольшие количества соответствующим образом сложенного материала (50%. Была обнаружена пригодность многих различных детергентов для получения хотя бы некоторого количества правильно сложенного материала, в том числе Triton Х-100, додецил-бета-мальтозид, CHAPS, CHAPSO, SDS, саркосил, Tween 20 и Tween 80, Zwittergent 3-14 и другие. Самыми предпочтительными из них оказались детергенты семейства CHAPS (CHAPS и CH APSO), которые наилучшим образом участвовали в реакции укладки и ограничивали агрегацию белка и образование неправильных дисульфидных связей. Наиболее предпочтительными были уровни содержания CHAPS выше 1%. Для повышения выхода использовался хлорид натрия, при оптимальном содержании от 0,1М до 0,5М. Для ограничения металл-катализируемого окисления (и агрегации), которое наблюдалось в некоторых препаратах, было предпочтительно присутствие EDTA (1-5мМ) в окислительно-восстановительном буфере. Оптимальные условия укладки обеспечивали концентрации глицерина выше 15%. Для максимального выхода было важно, чтобы окислительно-восстановительная пара в буфере состояла из окисленного и восстановленного органического тиола (RSH). Подходящие пары включают меркаптоэтанол, тлютатион (GSH), цистеамин, цистеин и их соответствующие окисленные формы. Предпочтительными окислительновосстановительными парами были глютатион (GSH):окисленный глютатион (GSSG), цистеин:цистин. Наиболее предпочтительная пара - глютатион (GSH): окисленный глютатион (GSSG). Повышенный выход наблюдался в том случае, когда мольное число окисленного члена пары было равно или превышало мольное число восстановленного члена. Значения рН между 7,5 и 9 были оптимальными для укладки этих вариантов ТРО. Органические растворители (этанол, ацетонитрил, метанол) применялись при концентрациях 10-15% или ниже. Более высокие уровни органических растворителей увеличивали количество неправильно сложенных форм. Наиболее подходящими были Tris и фосфатный буфер. Инкубация при 4°C также повышала выход правильно сложенного ТРО. Выход сложенного материала 40-60% (от количества восстановленного и денатурированного ТРО, используемого для реакции укладки) типичен для препаратов ТРО, которые очищали на . первом этапе при помощи колонки С4. Активный материал может быть получен при использовании менее чистых препаратов (т.е. сразу после колонки Superdex или после первоначальной экстракции "тел отражения"), хотя вы ход в этом случае меньше из-за сильного осаждения и помех, создаваемых другими (не-ТРО) белками во время процесса укладки. Поскольку ТРО содержит 4 цистеиновых остатка, возможно генерирование трех различных дисульфидных версий этого белка: дисульфидные связи между цистеиноверсия 1 выми остатками 1-4 и 2-3 дисульфидные связи между цистеиноверсия 2 выми остатками 1-2 и 3-4 дисульфидные связи между цистеиноверсия 3 выми остатками 1-3 и 2-4 Во время первых исследований при определении условий укладки было выделено несколько различных пиков, содержащих белок ТРО, в обращенно-фазной хроматографической колонке С4. Только один из этих пиков обладал значительной биологической активностью, определенно при помощи анализа клеток Ba/F3. В соответствии с этим условия укладки были оптимизированы в направлении увеличения выхода этой версии. В таких условиях вы ход неуложенных версий составлял менее 10-20% от общего количества мономерного ТРО, полученного на этапе солюбилизации. Масс-спектрометрией и секвенированием белка была определена дисульфидная модель биологически активного белка - 1-4 и 2-3, где цистеины нумеруются последовательно от амино-окончания. Эта цистеиновая модель с поперечными связями согласуется с известной дисульфидной моделью родственной молекулы эритропоэтина. D. Биологическая активность рекомбинантного сложенного ТРО Сложенный и очищенный ТРО обладает активностью при исследовании вне организма и в организме. Например, при анализе Ba/F3 половинно-максимальное стимулирование инкорпорации тимидина в клетки Ba/F3 для ТРО (Met-1 1-153) было достигнуто при 3,3пг/мл (0,3пМ). В ELISA на базе mpl-рецептора половинномаксимальная активность наблюдалась при 1,9нг/мл (120пМ). В нормальных и миелоугнетенных животных, облученных почти смертельной дозой рентгеновского излучения, сложенный ТРО (Met-1 1-153) имел высокий потенциал (активность была видна уже при дозах 30нг/мышь) стимуляции продуцирования новых тромбоцитов. Подобная биологическая активность наблюдалась для других форм ТРО, уложенных в соответствии с вышеописанной методикой (см. Фиг.25, 26 и 28). 14. Способы измерения тромбопоэтической активности Тромбопоэтическая активность может быть измерена различными методами анализа, включая Ва/F3анализ лиганда mpl, описанный в Примере 1, анализ ответного синтеза мышиных тромбоцитов, метод инициирования антигенов клеточной поверхности тромбоцитов с измерением антитромбоцитов (anti-GPIIblllа) для мегакариобластической клеточной линии человеческой лейкемии (СМК) [см. Sato et al., Brit. J. Haematol., 72:184-190 (1989)] (см. также жидкостно-суспензионный анализ мегакариоцитопоэза, описанный в Примере 4) и метод инициирования полиплоидизации в мегакариобластической клеточной линии (DAMI)·- [см. Ogura et al., Blood, 72(1):49-60 (1988)]. Развитие мегакариоцитов из незрелых, в основном не синтезирующи х ДНК клеток, в морфологически идентифицируемые мегакариоциты представляет собой процесс, включающий появление цитоплазматических органелл, вырабатывание мембранных антигенов (GPIIbIII a) эндорепликацию и высвобождение тромбоцитов, как описано в "Предпосылках изобретения". Предположительно, должен существова ть видо-специфический промотор (т.е. лиганд mpl) развития мегакариоцитов, инициирующий хотя бы некоторые из этих изменений в незрелых мегакариоцитах, ведущих к высвобождению и ослаблению тромбоцитопении. Таким образом, были разработаны различные виды анализа для измерения этих параметров в незрелых клеточных линиях мегакариоцитов, т.е. в клетках СМK и DAMI. Анализ СМK (Пример 4) исследует появление специфического маркера тромбоцитов, GPII bIIIa, и распространение тромбоцитов. Анализ DAMI (Пример 15) измеряет эндорепликацию с момента увеличения плоидности и пробы на зрелые мегакариоциты. Распознаваемые мегакариоциты имеют величины плоидности 2N, 4N, 8N, 16N, 32N и т.д. Наконец, анализ мышиных ответных тромбоцитов, проводимый в организме (Пример 16) полезен для проверки действия вводимых исследуемых соединений (здесь лиганда mpl) на рост количества тромбоцитов. Для измерения активности ТРО были разработаны два дополнительных метода анализа вне организма. Первый - активация рецептора киназы (KIRA) ELISA, при котором клетки СНО заражаются химерой mpl-Rse, и тирозин-фосфорилированный Rse измеряется методом ELISA после действия на mpl-участок химеры лиганда mpl (см. Пример 17). Второй метод - ELISA на основе рецептора, в котором чашка ELISA, покрытая кроличьим античеловеческим IgG захватывает mpl-IgG человеческого химерического рецептора, который связывается с исследуемым лигандом mpl. Биотинилированное поликлональное антитело кролика к лиганду mpl (TPO15s) используется для обнаружения связанного лиганда mpl, который измеряется с использованием стрептавидинпероксидазы, как описано в примере 18. 15. Биологический отклик в организме нормальных и сублетально облученных мышей, обработанных ТРО Нормальные и сублетально облученные мыши обрабатывались усеченной формой и полным ТРО, изолированным из клеток яичника китайского хомяка (СНО), Е. соli и человеческих эмбриональных почечных клеток (293). Обе формы ТРО, полученного из клеток трех хозяев, стимулировали продуцирование тромбоцитов у мышей, однако, ТРО полной длины, изолированный из СНО, по всей видимости, продуцировал наибольший отклик в организме. Эти результаты показывают, что для оптимальной активности в организме может быть необходима соответствующее гликозилирование карбокси-конечного домена. (a) Е. coli-rhTPO(Met-1, 153) "Met"-форму домена ЕРО (Met в позиции -1 плюс первые 153 остатка человеческого ТРО), продуцированную в Е. coli (см. Пример 23) вводили ежедневно нормальной самке мыши С57 В6, как описано в пояснениях к Фиг.25А, 25В и 25С. Эти рисунки показывают, что негликозилированная усеченная форма ТРО, продуцированного в Е. coli и сложенного, как описано выше, способна стимулировать примерно двукратное увеличение продуцирования тромбоцитов у нормальной мыши без' влияния на популяцию красных или белых кровяных телец. Та же молекула, вводимая ежедневно сублетально облученной (137Cs) самке мыши С57 В6, как описано в пояснениях к Фиг.26А, 26В и 26С, стимулировала восстановление тромобоцитов и устранила их полный упадок, но не оказала никакого эффекта на эритроциты или лейкоциты. (b) CHO-rhTPO332 ТРО полной длины, полученный из клеток СНО и вводимый ежедневно нормальной" самке мыши С57 В6, как описано в пояснениях к Фиг.27А, 27В и 27С, стимулировал примерно пятикратное увеличение продуцирования тромбоцитов у нормальной мыши без влияния на популяцию красных или белых кровяных телец. (c) CHO-rhTPO332, Ε. Coli-rhTPO(Met-1,153), 293-rhTPO332 и Ε.·coli-rhTPO155 Были построены кривые отклика на дозу при обработке нормальной мыши rhTPO, полученного из различных клеточных линий (CHO-rhTPO332, Ε·coli-rhTPO(Met-1, 153), 293-rhTPO332 и Ε. coli-rhTPO155), как описано в пояснении к Фиг.28. Этот рисунок показывает, что все испытанные формы молекулы стимулируют продуцирование тромбоцитов, однако форма полной длины, полученная из СНО, имеет наивысшую активность в организме. (d) CHO-rhTPО153 , C YO-rhTPO - ”усеченная”и CHO-rhTPО332 Были также построены кривые отклика на дозу при обработке · нормальной мыши различными формами rhTPO, полученного из клеток СНО (CHO-rhTPO153, C YO-rhTPO "усеченная" и CHO-rhTPO332), как описано в пояснении к Фиг.29. Эта фигура показывает, что все испытанные формы молекулы стимулируют продуцирование тромбоцитов, однако форма полной длины (70кДа) имеет наивысшую активность в организме. 16. Общий способ приготовления лиганда mpl и его вариантов Предпочтительно лиганд mpl получают по стандартной методике рекомбинации, которая включает продуцирование полипептида лиганда mpl путем культивирования клеток, подвергнутых трансфекции, для экспрессии нуклеиновой кислоты лиганда mpl (обычно путем трансформации клеток вектором экспрессии) и извлечения полипептида из клеток. Однако, было сделано предположение, что лиганд mpl может быть получен путем гомологичной рекомбинации, или при помощи методики рекомбинантного продуцирования, которая использует управляющие элементы, вводимые в клетки, уже содержащие ДНК, кодирующую лиганд mpl. Например, мощный элемент-промотор/усилитель, угнетающий элемент или экзогенный элемент модулирования транскрипции могут быть введены в геном клетки хозяина в приближении и с ориентацией, достаточными для оказания влияния на транскрипцию ДНК, кодирующей нужный лиганд mpl. He управляющий элемент кодирует лиганд mpl, а сама ДНК соответствует геному хозяйской клетки. Затем можно осуществить просеивание полученных культур для получения клеток, вырабатывающих полипептид рецептора данного изобретения, или для увеличения или уменьшения уровней экспрессии, по желанию. Таким образом, изобретение предлагает способ получения лиганда mpl, предусматривающий вставку в геном клетки, содержащей молекулу н уклеиновой кислоты лиганда mpl, элемента модулирования транскрипции, в приближении и с ориентацией, достаточными для оказания влияния на транскрипцию ДНК, кодирующей нужный лиганд mpl, с возможным последующим, этапом культивирования клетки, содержащей элемент модулирования транскрипции и молекулы нуклеиновой кислоты. Изобретение также решает, предлагает хозяйскую клетку, содержащую врожденную нуклеиновую кислоту лиганда mpl, оперативно связываемую с экзогенными управляющими последовательностями, распознаваемыми хозяйской клеткой. А. Изоляция ДНК, кодирующей полипептид лиганда mpl ДНК, кодирующая полипептид лиганда mpl, может быть получена из любой библиотеки кДНК, приготовленной из ткани, которая предположительно содержит мРНК лиганда mpl и выражает ее на обнаруживаемом уровне. Ген лиганда mpl может быть также получен из библиотеки геномних ДНК или путем олигонуклеотидного синтеза вне организма из полного нуклеотида или аминокислотной последовательности. Библиотеки просеиваются в соответствии с зондами, сконструированными для идентификации гена, представляющего интерес, или закодированного им белка. Для библиотек экспрессии кДНК подходящими зондами являются моноклональные или поликлональные антитела, которые распознают лиганды mpl и особым образом с ними связываются. Для библиотек кДНК подходящими зондами являются олигонуклеотиды, состоящие в длину примерно из 20-80 оснований, которые кодируют известные или предполагаемые участки кДНК лиганда mpl из одного и того же или разных видов животных, а также комплементарные или гомологичные кДНК или их фрагменты, которые кодируют тот же или подобный ген. Соответствующие зонды для просеивания библиотек геномних ДНК включают (но не ограничиваются ими) олигонуклеотиды, кДНК или их фрагменты, которые кодируют то т же или подобный ген, и/или гомологичные геномние ДНК или их фрагменты. Просеивание кДНК или геномной библиотеки при помощи выбранного зонда может проводиться с использованием стандартной методики, как описано в главах 10-12 Sambrook et al., выше. Альтернативой является изолирование гена, кодирующего лиганд mpl, чтобы использовать методику PCR, как описано в разделе 14 Sambrook et al., выше. Эта методика требует использования олигонуклеотидных зондов, гибридизирующих ДНК, кодирующую лиганд mpl. Способы выбора олигонуклеотидов описаны ниже. Предпочтительный способ практического осуществления данного изобретения - использование тща тельно отобранных олигонуклеотидных последовательностей для просеивания библиотек кДНК из различных тканей, предпочтительно человеческой или свиной почки (взрослой или зародышевой), или клеточных линий печени. Например, библиотеки кДНК клеточных линий человеческой зародышевой печени просеиваются при помощи олигонуклеотидных зондов. Иногда и человеческие геномные библиотеки могут просеиваться при помощи олигонуклеотидных зондов. Олигонуклеотидные последовательности, выбранные в качестве зондов, должны иметь достаточную длину и быть достаточно недвусмысленными, чтобы, избежать ложных результатов. Н ужная нуклеотидная последовательность (последовательности) обычно конструируются на основе участков лиганда mpl, имеющих минимальную избыточность кодонов. Олигонуклеотиды могут быть дегенерированы одной или более позициях. Использование дегенерированных олигонуклеотидов особенно важно, если просеивается библиотека, полученная от вида, наличие в котором предпочтительного кодона неизвестно. Олигонуклеотид должен быть помечен, так чтобы он мог быть обнаружен после гибридизации с ДНК в просеиваемой библиотеке. Предпочтительный способ введения метки - использование АТР (например, g32Р) и полинуклеотидной киназы для мечения радиоактивным изотопом конца 5' олигонуклеотида. Однако могут быть использованы и другие способы мечения олигонуклеотида, включая, например, биотинилирование или энзимную метку. Особый интерес представляет нуклеиновая кислота лиганда mpl, кодирующая полипептид лиганда mpl полной длины. В некоторых предпочтительных вариантах осуществлейия изобретения последовательность нуклеиновой кислоты включает в себя природную сигнальную последовательность лиганда mpl. Нуклеиновая кислота, содержащая всю последовательность, кодирующую белок, получают просеиванием выбранных библиотек кДНК или геномних библиотек с использованием выведенной аминокислотной последовательности. В. Варианты аминокислотной последовательности природного лиганда mpl Варианты аминокислотной последовательности природного лиганда mpl получают путем введения соответствующи х нуклеотидных изменений в ДНК лиганда mpl или путем синтеза вне организма желаемого полипептида лиганда mpl. Такие варианты включают, например, делеции, вставки или замещения в остатках аминокислотной последовательности свиного лиганда mpl. Например, карбокси-конечная часть зрелого лиганда mpl полной длины может быть удалена путем протеолитического расщепления (в организме или вне организма) или путем клонирования и экспрессии фрагмента ДНК, кодирующего лиганд mpl полной длины, для продуцирования биологически активного варианта. Любая комбинация проводимых делеций, вставок и замещений должна быть достигнута в конечной конструкции, с учетом того, чтобы эта конечная конструкция обладала желаемой биологической активностью. Аминокислотные изменения могут также изменять посттрансляционные процессы лиганда mpl, такие как изменение количества или положения участков гликозилирования. При конструировании вариантов аминокислотных последовательностей лиганда mpl размещение участков мутаций и природа мутаций будут зависеть от характеристик модифицируемого лиганда mpl. Участки мутаций могут модифицироваться отдельно или группами, например, (1) замещением сначала консервативными аминокислотами, а затем более радикальными,в зависимости от достигнуты х результатов, (2) делецией целевого остатка, или (3) вставкой остатков того же или другого класса рядом с выбранным участком, или комбинацией вариантов 1-3. Удобная методика идентификации определенных остатков или участков полипептида лиганда mpl, выбранных для мутагенеза, называется "мутагенез с аланиновым сканированием", описанный Cunningham and Wells, Science, 244:1081-1085 [1989]. Остаток или группа целевых остатков идентифицируется (например, заряженные остатки, такие как arg, asp, his, lys и glu) и заменяются любой, но предпочтительно нейтральной или положительно заряженной аминокислотой (наиболее предпочтительно аланином или полиаланином) для оказания влияния на взаимодействие аминокислот с водной средой снаружи клетки. Затем домены, показывающие функциональную чувствительность к замещениям, подвергаются усовершенствованию путем введения тех же или других вариантов рядом или непосредственно в места замещения. Таким образом, в то время как участок введения вариаций аминокислотных последовательностей определяется предварительно, природа мутации не нуждается в предопределении. Например, чтобы оптимизировать действие мутации на данном участке, проводится мутагенез с аланиновым сканированием (или случайный мутагенез) в целевом кодоне или, участке, и выраженные варианты лиганда mpl просеиваются для получения оптимальной комбинации желаемой активности. Существуют две основные переменные при конструировании вариантов аминокислотных последовательностей: расположение участка мутации и природа мутации. Например, варианты полипептида лиганда mpl включают варианты последовательности лиганда mpl и могут представлять встречающиеся в природе аллели (которые не требуют манипуляций с ДНК лиганда mpl) или предварительно определенные мутантные формы, полученные мутацией ДНК, приводящей к аллели или к варианту, не встречающемуся в природе. В основном, расположение и природа выбранных мутаций будут зависеть от характеристик модифицируемого лиганда mpl. Делеции аминокислотной последовательности обычно варьируются в диапазоне от 1 до 30 остатков, более предпочтительно от 1 до 10 остатков, и обычно являются смежными. Как альтернатива, делеции аминокислотной последовательности лиганда mpl могут включать часть или целый карбокси-конечный
ДивитисяДодаткова інформація
Назва патенту англійськоюPolypeptide of mpl ligand (variants), isolated molecul of nucleic acid encoding polypeptide of mpl ligand (variants), expressing vector, host cell, method for manufacturing polypeptide of mpl ligand (variants), composition, method for drug manufacturing, method for detecting polypeptide of mpl ligand, antibody, method for producing antibody, hybridoma cell line
Назва патенту російськоюПолипептид лиганда mpl (варианты), изолированная молекула нуклеиновой кислоты, кодирующей полипептид лиганда mpl, вектор экспрессии, клетка-хозяин, способ продуцирования полипептида лиганда mpl (варианты), композиция, способ изготовления лекарственного средства, способ определения присутствия лиганда полипептида mpl, антитело, способ изготовления антитела, клеточная линия гибридомы
МПК / Мітки
МПК: A61K 38/00, C12N 15/19, C07K 19/00, C07K 14/52, A61K 38/22, A61P 7/00, C12N 15/16, C12Q 1/68, C12N 15/09, C12N 15/12, C07H 21/04, C12N 5/10, A61P 7/02, C12P 21/02, C12P 21/08
Мітки: композиція, молекула, кислоти, нуклеїнової, вектор, кодує, експресії, поліпептид, ліганду, ізольована, клітина-хазяїн, продукування, спосіб, варіанти, поліпептиду
Код посилання
<a href="https://ua.patents.su/161-72869-polipeptid-ligandu-mpl-varianti-izolovana-molekula-nuklenovo-kisloti-shho-kodueh-polipeptid-ligandu-mpl-vektor-ekspresi-klitina-khazyan-sposib-produkuvannya-polipeptidu-ligandu-mpl.html" target="_blank" rel="follow" title="База патентів України">Поліпептид ліганду mpl (варіанти), ізольована молекула нуклеїнової кислоти, що кодує поліпептид ліганду mpl, вектор експресії, клітина-хазяїн, спосіб продукування поліпептиду ліганду mpl (варіанти), композиція,</a>
Послідовність днк, що кодує поліпептид, який зв’язується з mort-1 (варіанти), вектор експресії, що включає послідовність днк, клітина- хазяїн, виділений поліпептид та фармацевтична композиція, що містить поліп

Номер патенту: 72423
Опубліковано: 15.03.2005
Автори: Болдін Марк, Гольцев Юрій, Воллах Девід, Гончаров Таня
МПК: C07K 14/715, A61K 48/00, A61P 31/12, A61K 45/00, C12N 5/10, A61P 35/00, C12N 9/64, C07K 16/28, C12N 15/09, C07K 14/47, C07K 14/705
Мітки: поліпептид, експресії, клітина, фармацевтична, днк, хазяїн, композиція, включає, вектор, варіанти, виділений, кодує, mort-1, містить, послідовність, зв'язується, поліп
Формула / Реферат:
1. Послідовність ДНК, що кодує поліпептид, який зв'язується з MORT-1, де MORT-1 є білком, який зв'язується з внутрішньоклітинним доменом FAS-R, і який зв'язується з білком TRADD, який зв'язується з внутрішньоклітинним доменом p55-TNF-R, де зазначений поліпептид має:a) послідовність, що містить залишки 1-182 послідовності SEQ ID №5;b) послідовність, що містить фрагмент а), де фрагмент зв'язується з MORT-1;c) послідовність,...
Гібридні білки, фармацевтична композиція, що містить гібридний білок, ізольована молекула днк, що містить гібридний білок, вектор експресії, клітина-хазяїн, спосіб продукування гібридного білка, спосіб індукува

Номер патенту: 52646
Опубліковано: 15.01.2003
Автори: Джеймсон Брендфорд А., Чаппел Скотт С., Кампбелл Роберт К.
МПК: C07K 14/715, C07K 14/59
Мітки: експресії, композиція, білок, білки, містить, ізольована, вектор, молекула, фармацевтична, білка, індукува, продукування, клітина-хазяїн, гібридні, днк, гібридного, гібридний, спосіб
Формула / Реферат:
1. Гібридний білок, який містить дві різні коекспресовані амінокислотні послідовності, що утворюють гетеродимер, кожна з яких містить:(a) щонайменше одну амінокислотну послідовність, вибрану з групи, що складається з ланцюга гомомерного рецептора, ланцюга гетеромерного рецептора, ліганду, відмінного від гонадотропіну, фрагмента згаданого ланцюга згаданого гомогенного рецептора, згаданого ланцюга згаданого гетеромерного рецептора або...
Молекула нуклеїнової кислоти, що кодує поліпептид, який являє собою варіант ліпази, яка стимулюється солями жовчі (bssl) і має її активність (варіанти), поліпептид (варіанти), фармацевтична композиція

Номер патенту: 48109
Опубліковано: 15.08.2002
Автори: Лундберг Леннарт, Ханссон Леннарт, Тьорнелль Ян, Блекберг Ларс, Хернелл Олле, Стрьомквіст Матс, Едлунд Мікаель
МПК: C12N 5/10, A23C 9/20, A01K 67/027, C12N 15/85, A61K 38/46, A61P 3/06, C07H 21/04, A23L 1/305, C12N 9/18, C12N 15/86, C12N 15/09
Мітки: кислоти, кодує, молекула, стимулюється, варіанти, яка, композиція, ліпази, фармацевтична, має, являє, солями, активність, bssl, собою, нуклеїнової, жовчі, варіант, поліпептид
Формула / Реферат:
1. Молекула нуклеиновой кислоты, кодирующая полипептид, который представляет собой вариант стимулируемой солями желчи липазы (BSSL) и имеет ее активность, включающий менее чем 722 аминокислот, у которого одна или более, но не все из аминокислот участка, соответствующего аминокислотам 536-722 представленной последовательности, обозначаемой как SEQ ID NO 3 в списке последовательностей, делетированы.2. Молекула нуклеиновой...
Виділена молекула нуклеїнової кислоти (варіанти), химерний рослинний ген (варіанти), трансформуючий рослину вектор (варіанти), рослинна клітина (варіанти), спосіб одержання рослини, що проявляє стійкість до гер

Номер патенту: 44217
Опубліковано: 15.02.2002
Автори: Бог Моллі, Лебрюн Мішель, Фрейссіне Жорж, Томас Террі
МПК: C12N 15/09, C12N 9/10, A01H 1/00, C07K 14/415, C12N 15/82
Мітки: виділена, проявляє, трансформуючий, рослину, рослинній, варіанти, кислоти, ген, химерний, нуклеїнової, стійкість, рослини, вектор, клітина, одержання, рослинна, молекула, гер, спосіб
Формула / Реферат:
1. Выделенная молекула нуклеиновой кислоты, содержащая регуляторный элемент гена гелиантинина, где указанный регуляторный элемент управляет семя-специфичной генной экспрессией в растении и включает нукдеотиды 2304-2401 SEQ ID NO: 1: 2. Выделенная молекула нуклеиновой кислоты, содержащая регуляторный элемент гена гелиантинина, где указанный регуляторный элемент управляет корень-специфичной генной экспрессией в...
Saf-поліпептид, фрагмент днк, що кодує saf-поліпептид, рекомбінантна плазмідна днк для експресії safполіпептиду (варіанти), штам грибів streptomyces lividans, спосіб експресії saf-поліпептиду

Номер патенту: 27696
Опубліковано: 16.10.2000
Автори: Гарсіа Томас Вігаль, Жіль Хосе Антоніо, Ортега Антоніо Даза, Мартін Хуан Франціско
МПК: C12N 15/09, C07K 14/37, C12N 1/21, C12P 21/02, C12N 15/31, C07K 14/36, C07K 14/41, C07K 14/00
Мітки: варіанти, експресії, saf-поліпептиду, спосіб, плазмідна, streptomyces, lividans, днк, фрагмент, рекомбінантна, штам, грибів, safполіпептиду, saf-поліпептид, кодує
Текст:
...от штамма АТСС 10137 S. griseus. 7.2 кб Bgl II фрагмент от pULADI и 1 кб Bgl II фрагмент от pULAD3 субклониовали раздельно в обеих ориентациях в ВамНI-переваренной pUC 19. Все эти плазмидные конструкции проверялись в клетках Е. coli M103 и затем использовались для трансформирования Е. coli E15 (Sartiy и др. J. Bacteriol, 145:288-292,1981) и Е. coli A 1046, в В-ХР не было обнаружено никаких синих колоний, позволяющих предположить, что...
Попередній патент: Спосіб лазерного гравірування форм флексодруку
Наступний патент: Спосіб лікування раку та спосіб пригнічення росту пухлинних клітин з застосуванням інгібіторів нааладази
Випадковий патент: Спосіб установлення зв'язку в комп'ютерних мережах