Промотор з органо-специфічною активністю у рослинах цукрового буряку, що індукується при зберіганні
Номер патенту: 97622
Опубліковано: 12.03.2012
Автори: Роттуес Александер, Хель Райнхард, Шталь Дітмар Юрген







Формула / Реферат
1. Промотор з органо-специфічною активністю у рослинах цукрового буряку, який у запасаючих органах рослин при зберіганні при 17 ºС або при 26-28 ºС виявляє вищу активність, ніж в інших частинах цих рослин, та тим, що його активність після збирання запасаючих органів змінюється та стає вищою, ніж перед їх збиранням.
2. Промотор за п. 1, який відрізняється тим, що його активність у запасаючих органах піддається вимірюванню методом візуалізації плям РНК, і активність перед збиранням запасаючих органів за порівнюваних умов дослідження виявляється меншою ніж 20 %, відповідно до варіанта, якому віддається перевага, меншою ніж 10 %, та, зокрема, менше ніж 5 %.
3. Промотор, який містить:
a) послідовність нуклеотидів за SEQ ID NO: 1 або
b) послідовність нуклеотидів за SEQ ID NO: 2 або,
c) послідовність нуклеотидів за SEQ ID NO: 3 або,
d) послідовність нуклеотидів за SEQ ID NO: 4 або,
е) послідовність нуклеотидів за SEQ ID NO: 5 або,
f) послідовність нуклеотидів, комплементарну до послідовностей а)-е), або
g) послідовність нуклеотидів, гідбридизовану з однією з послідовностей нуклеотидів a)-f).
4. Похідні промотору за п. 3, що мають функціональну активність промотору за п. 1.
5. Вектор для трансформації Agrobacterium tumefaciens, який включає промотор за будь-яким з пп. 1-3 або похідну за п. 4.
6. Еукаріотна або прокаріотна клітина-хазяїн, яка включає промотор за будь-яким з пп. 1-3 або похідну за п. 4.
7. Трансгенна рослина цукрового буряку або її частини, які включають промотор або похідні за одним із пп. 1-4.
8. Трансгенна рослина або її частини за п. 7, трансформовані промотором за будь-яким з пп. 1-3 або похідним за п. 4 для впливу на метаболізм у запасаючих органах.
9. Трансгенна рослина або її частини за п. 7, яка відрізняється тим, що рослиною є Beta vulgaris.
10. Трансгенне насіння рослини за п. 7 або 8 для продукування трансгенної рослини або її частин за п. 7 або 8.
11. Застосування промотору за одним із пп. 1-3 або похідних за п. 4 для одержання трансгенної рослини цукрового буряку, що має одну або кілька з наведених нижче властивостей:
a) покращений метаболізм вуглеводів у запасаючих органах після збирання,
b) покращений метаболізм азоту у запасаючих органах після збирання,
c) покращену стійкість проти висихання та покращений водний статус у запасаючих органах після збирання,
d) покращену холодостійкість та морозостійкість запасаючих органів після збирання,
e) підвищену резистентність або стійкість запасаючих органів проти патогенних організмів після збирання,
f) покращений вторинний обмін речовин у запасаючих органах після збирання.
Текст
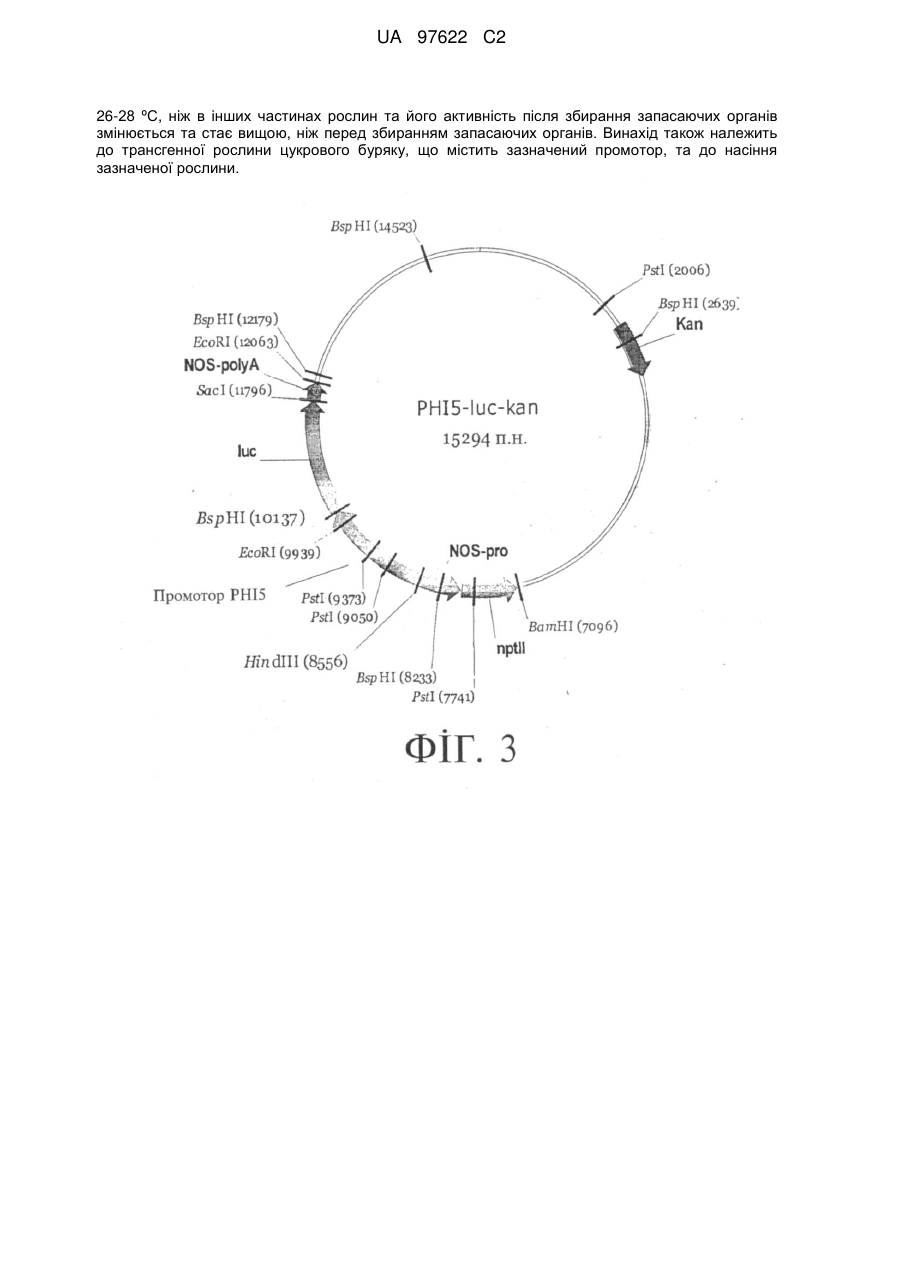
Реферат: Винахід належить до промотору з органо-специфічною активністю у рослинах цукрового буряку, який виявляє у запасаючих органах рослин вищу активність при зберіганні при 17 ºС або при UA 97622 C2 (12) UA 97622 C2 26-28 ºС, ніж в інших частинах рослин та його активність після збирання запасаючих органів змінюється та стає вищою, ніж перед збиранням запасаючих органів. Винахід також належить до трансгенної рослини цукрового буряку, що містить зазначений промотор, та до насіння зазначеної рослини. UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 Цей винахід стосується промотору з органо-специфічною активністю у рослинах, його застосування, а також трансгенних рослин. Згідно з Нільсоном (Nilsson, 2000), корисні рослини з точки зору придатності для зберігання можна розділити на три групи. Збираний матеріал першої групи, до якої належать такі рослини, як капуста, броколі, цвітна капуста, спаржа та шпінат, охоплює листя, пагони, квітки та бруньки. Органи цих рослин мають низьку здатність до нагромадження води і після збирання швидко старіють. До другої групи належать рослини з м'ясистими плодами, наприклад, томати, гарбузи та груші. Плоди цих рослин визрівають та старіють під час зберігання. До третьої групи належать дволітні рослини. Рослини з дворічним життєвим циклом, наприклад, цукровий буряк, цикорій, морква, цибуля або артишок, на протязі першого року утворюють та розвивають запасаючий орган, який наступного року забезпечує цвітіння та утворення насіння. Запасаючі органи після збирання врожаю зазнають численних фізіологічних змін, які впливають на якість запасаючих органів та кількість речовини в них. Ці фізіологічні зміни, з одного боку, виникають внаслідок механічних пошкоджень під час збирання, наприклад, порізів та ударів, а з іншого боку обумовлені впливом зберігання та пов'язаної з ним втрати води, є наслідками вимушеного або природного стану спокою або результатами пристосування до холоду. Отже, для уможливлення впливу на інтенсивність обміну речовин у запасаючих органах після збирання врожаю необхідні цілеспрямовані процеси, які переважно регулюються специфічними промоторами. Безумовно відомі гени, які активуються у пагонах спаржі або у квітках броколі після збирання врожаю. Навпаки, у запасаючих органах рослин такі гени невідомі, тому до цього часу не існує посилань на відповідні промотори. Таким чином, метою цього винаходу є запропонувати промотор, за допомогою якого можна впливати на фізіологічні зміни у запасаючих органах рослин після їх збирання, пов'язані з обміном речовин. Згідно з цим винаходом, засобом досягнення цієї мети є промотор, що має ознаки, вказані у п. 1 формули винаходу. Нижче подано більш детальні визначення термінів та понять, вжитих у цій заявці. У значенні, пов'язаному з цим винаходом, запасаючими органами рослини є частини рослини, призначені для нагромадження вуглеводів, наприклад, сахарози, крохмалю або інуліну, та/або азотвмісних сполук, наприклад, протеїнів або амінокислот. Типовим запасаючим органом є, наприклад, корінь або гіпокотиль. Запасаючими органами паросткового типу можуть бути бульби картоплі, топінамбуру та міжвузля стеблини цукрової тростини. Подальшими прикладами запасаючих органів є кореневі бульби, наприклад, ямсу, маніоки та батату. Дволітні рослини потребують для життєвого циклу дворічного періоду. У перший рік закладаються запасаючі органи рослин. У другий рік відбувається цвітіння та утворення насіння з використанням резервів поживних речовин, нагромаджених у запасаючому органі. Термін «промотор» означає послідовність нуклеотидних основ у нуклеїновій кислоті, яка керує експресією гена, у певних випадках залежно від екзогенних та ендогенних факторів. До цих факторів належать, наприклад, індуктори, репресори та аналогічні протеїни, що зв'язують ДНК, але також і фактори оточуючого середовища. Промотор може складатися з кількох елементів. Однак у будь-якому разі він включає регуляторний елемент, який відповідає за транскрипцію гена, що знаходиться під його контролем. Промотор, який є більш активним у запасаючому органі рослини після збирання, ніж до збирання, і активність якого індукується таким чином, виявляє у зібраних коренях певну активність (вимірювану методом візуалізації плям РНК); у незібраних коренях при порівнянних умовах досліду така активність становить, наприклад, менше ніж 20%, відповідно до варіанта, якому віддається перевага, менше ніж 10% та, зокрема, менше ніж 5%. Ця специфічність може виникнути лише з деяким запізненням після збирання та на протязі зберігання. «Похідні» промотору являють собою скорочені або подовжені, або частково ідентичні версії або гомологи цього промотору з однаковими або в основному однаковими властивостями. Термін «пряма протигрибкова (або антибактеріальна) дія» означає, що генні продукти чинять безпосередній протигрибковий вплив, наприклад, розчиняють клітинні оболонки або кодують фітоалексинсинтази або метаболіти, які пригнічують обмін речовин у грибках або мікроорганізмах. Термін «непряма протигрибкова (або антибактеріальна) дія» означає, що генні продукти активують рослинний генний захист. До таких генів належать, наприклад, гени резистентності, компоненти шляхів передачі сигналів (наприклад, кінази, фосфатази), фактори транскрипції або ферменти, які продукують сигнальні речовини (наприклад, ферменти, що утворюють етилен, саліцилову кислоту, жасмонат, реакційноздатні кисневмісні сполуки, монооксид азоту). Термін «інфекція» означає найбільш ранній момент часу, у який обмін речовин грибка (або росту грибка) стає готовим до проникнення у тканину організму-хазяїна. До таких явищ належать, наприклад, виростання гіфів або утворення специфічних інфекційних структур, наприклад, пенетраційних гіфів або апресоріїв. 1 UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 Термін «гомологія» означає в даному описі гомологію щонайменше 70% площини ДНК, яку можна визначити відомими способами, наприклад, зіставленням послідовностей за допомогою комп'ютера (Альтшуль та ін. - S.F. Altschul et аl., 1990, J. Моl. Biol. 215:403-410). Термін «комплементарна послідовність нуклеотидів» стосовно двонитчастої ДНК означає, що друга нитка ДНК, яка доповнює першу нитку ДНК, у відповідності з правилами парування основ містить нуклеотидні основи, які відповідають основам першої нитки. Термін «гібридизація» у значенні, вживаному в цьому описі, означає гібридизацію у звичайних умовах, наприклад, описаних Сембруком та ін. (Sambrook et аl., 1989), а у варіантах, яким віддається перевага - у жорстких умовах. Жорсткими умовами гібридизації є, наприклад, гібридизація у 4xSSC при 65°C з подальшим кількаразовим промиванням у 0,1xSSC при 65°C при загальній тривалості приблизно 1 год. Менш жорсткими умовами гібридизації є, наприклад, гібридизація у 4xSSC при 37°С з подальшим кількаразовим промиванням у 1xSSC при кімнатній температурі. Вислів «жорсткі умови гібридизації» може означати також гібридизащю при 68°С у 0,25 M фосфаті натрію, рН 7,2, 7% SDS, 1 мМ EDTA та 1% BSA протягом 16 год із подальшим дворазовим промиванням 2xSSC та 0,1% SDS при 68°С. Нижче подано більш детальний опис винаходу з посиланнями на фігури та приклади. Промотор відповідно до цього винаходу є активним після збирання у запасаючих органах рослин, наприклад, у коренях цукрового буряка, моркви та цикорію або у бульбах картоплі. Навпаки, у запасаючих органах або інших частинах рослин до збирання його активність є низькою або взагалі не виявляється. Цю властивість можна застосувати для покращення обміну речовин у запасаючих органах рослин після збирання. Застосування промотору уможливлює також одержання трансгенних рослин та частин цих рослин, наприклад, насіння. Відповідно до варіантів, яким віддається перевага, у запасаючих органах рослин методом візуалізації плям PHK може вимірюватися активність промотору відповідно до цього винаходу, яка при порівнянних умовах досліду на запасаючих органах перед збиранням становить менше ніж 20%, відповідно до варіанта, якому віддається перевага, менше ніж 10% та, зокрема, менше ніж 5%. Згідно з подальшим варіантом здійснення винаходу, промотор містить: a) послідовність нуклеотидів за SEQ ID NO 1 або b) послідовність нуклеотидів за SEQ ID NO 2 або c) послідовність нуклеотидів за SEQ ID NO 3 або d) послідовність нуклеотидів за SEQ ID NO 4 або e) послідовність нуклеотидів за SEQ ID NO 5 або f) послідовність нуклеотидів, комплементарну до послідовностей а)-е), або g) послідовність нуклеотидів, гібридизовану з однією з послідовностей нуклеотидів a)-f). Винахід передбачає також похідні такого промотору. Такі похідні більш детально визначені вище та включають також фрагменти ДНК промотору, представлені у картах рестриктів, а також фрагменти ДНК, які можна одержати із застосуванням не названих явно наявних на ринку рестрикційних ендонуклеаз. Винахід стосується, крім того, трансгенних рослин, які можна трансформувати із застосуванням промотору відповідно до цього винаходу. Винахід стосується також застосування промотору відповідно до цього винаходу або його похідних для одержання трансгенних рослин, які мають одну або кілька нижчевказаних властивостей: a) покращений метаболізм вуглеводів у запасаючих органах після збирання, b) покращений метаболізм азоту у запасаючих органах після збирання, c) покращену стійкість проти висихання та покращений стан води у запасаючих органах після збирання, d) покращену холодостійкість та морозостійкість запасаючих органів після збирання, e) підвищену резистентність або стійкість запасаючих органів проти патогенних організмів після збирання, f) покращений вторинний обмін речовин у запасаючих органах після збирання. Тому промотор відповідно до цього винаходу можна застосовувати для послаблення розщеплення цукру у зібраних цукрових буряках та нагромадження інвертованого цукру або для запобігання цим явищам. Для цієї мети слід, наприклад, експресувати у зібраних коренях ген інгібітора інвертази та пригнітити активність клітинних інвертаз. Промотор відповідно до цього винаходу можна застосовувати також для досягнення підвищеного виходу інуліну та продукування молекул інуліну з подовженими ланцюгами у коренях цикорію. Для цього слід послабити розщеплення інуліну у зібраному цикорії або запобігти такому розщепленню, для чого слід послабити активність фруктозил-екзогідролази у зібраних коренях за допомогою антисенсибілізатора або інформаційної РНК. Промотор відповідно до цього винаходу можна застосовувати також для послаблення «підсолоджування під впливом холоду» зібраних та закладених на зберігання бульб картоплі. Для цього слід експресувати у зібраних бульбах ген інгібітора інвертази та пригнітити активність клітинних інвертаз. 2 UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 Промотор відповідно до цього винаходу можна, крім того, застосовувати для зниження у запасаючих органах рослин після збирання вмісту «шкідливого азоту», що піддається екстракції, наприклад, амінокислот. Підвищені концентрації азотвмісних сполук у запасаючих органах рослин часто знижують поживно-фізіологічну цінність продукції або утруднюють добування нагромаджених речовин, наприклад, сахарози з коренів цукрових буряків. Зниження вмісту «шкідливого азоту», що піддається екстракції, у коренях можна досягти шляхом забезпечення посиленого синтезу протеїнів з амінокислот у запасаючих органах за допомогою промоторів. На відміну від амінокислот, протеїни при добуванні цукру з цукрових буряків випадають в осад і тому не екстрагуються. Промотор відповідно до цього винаходу можна застосовувати також для підвищення холодо- та морозостійкості зібраних запасаючих органів. Для цього слід, наприклад, експресувати за допомогою промотору фактори транскрипції для холодо- та морозостійкості або протеїнів для захисту від холоду або морозу. За допомогою промотору відповідно до цього винаходу можна також покращити стійкість зібраних запасаючих органів проти захворювань. Зібрані запасаючі органи можуть бути інфіковані численними грибками, що розвиваються в грунті, наприклад, представниками виду Fusarium, або бактеріями, наприклад, Erwinia carotovora - збудниками мокрої гнилі бульб картоплі. Застосування комбінації індукованого промотору з геном, продукт якого виявляє пряму або непряму протигрибкову або антибактеріальну дію, уможливлює забезпечення резистентності до грибків або бактерій та покращення придатності запасаючих органів рослин для зберігання. Нарешті, промотор відповідно до цього винаходу можна застосовувати для покращення вторинного обміну речовин у запасаючих органах рослин. Бульби картоплі є найважливішим джерелом вітаміну C у Середній Європі. Однак вміст вітаміну C у бульбах під час зберігання знижується. За допомогою промотору це розщеплення вітаміну можна послабити або запобігти йому. ФІГУРИ На Фіг. 1 показано індукцію індукованих після збирання (PHI) генів 7, 20, 153 та 227 цукрового буряка після збирання коренів буряка, визначену методом візуалізації плям РНК. Аналізувалася експресія генів у листі та коренях цукрового буряка безпосередньо перед збиранням (0d), а потім у різні моменти часу після збирання. Для цього корені зберігали протягом 1 доби, 4 діб, 7 діб, 14 діб, 21 діб, 28 діб та 35 діб при 17°С або протягом 1 доби, 4 діб та 14 діб при 26-28°C. Для аналізу у кожний відповідний момент часу виділяли у денатурувальному формальдегід-агарозному гелі 10 мкг загальної клітинної PHK та гібридизували із фрагментами кодувальної ДНК генів РНІ7, РНІ20, РНІ153 та РНІ227. На Фіг. 2 показано індукцію експресії індукованого після збирання (PHI) гена 5 у коренях цукрового буряка після збирання, визначену методом RT-PCR. Безпосередньо перед збиранням (0d) транскрипт РНІ5 спостерігається у коренях у дуже незначній кількості, а у листі - у невеликій кількості. У сукупностях кДНК, одержаних з РНК, виділеної з коренів після 1 доби, 4 діб, 7 діб, 14 діб, 21 доби, 28 діб, 35 діб та 46 діб зберігання при 17°С та після 1 доби, 3 діб, 4 діб, 7 діб та 14 діб при 26-28°С, спостерігається значна кількість транскрипту РНІ5. Контроль експресії гена глщеринальдегід-3-фосфат-дегідрогенази (GAPDH) свідчить, що для реакцій RT-PCR застосовувалися однакові кількості кДНК. На Фіг. 3 показано вектор гена-репортера PHI5-luc-kan з трансляційним злиттям між промотором РНІ5 та геном люциферази з Photinus pyralis. Промотор РНІ5 у векторі PHI5-luc-kan містить нуклеотиди положень 1-1587 нуоеотидної послідовності SEQ ID NO 1 та може бути виділений з вектора за допомогою HindIII та BspHI та комбінований з іншими генами. На Фіг. 4 показано вектор гена-репортера PHI7-luc-kan з трансляційним злиттям між промотором РНІ7 та геном люциферази з Photinus pyralis. Промотор РНІ7 у векторі PHI7-luc-kan містить нуклеотиди положень 1-2695 нуклеотидної послідовності SEQ ID NO 2 та може бути виділений з вектора за допомогою HindIII та Ncol та комбінований з іншими генами. На Фіг. 5 показано вектор гена-репортера PHI20-luc-kan з трансляційним злиттям між промотором РНІ20 та геном люциферази з Photinus pyralis. Промотор РНІ20 у векторі PHI20-luc-kan містить нуклеотиди положень 1-2102 нуклеотидної послідовності SEQ ID NO 3 та може бути виділений з вектора за допомогою HindIII та Ncol та комбінований з іншими генами. На Фіг. 6 показано вектор гена-репортера PHI153-luc-kan з трансляційним злиттям між промотором РНІ153 та геном люциферази з Photinus pyralis. Промотор РНІ153 у векторі PHI153-luc-kan містить нуклеотиди положень 1-5829 нуклеотидної послідовності SEQ ID NO 4 та може бути виділений з вектора за допомогою HindIII та Ncol та комбінований з іншими генами. На Фіг. 7 показано вектор гена-репортера PHI227-luc-kan з трансляційним злиттям між промотором РНІ227 та геном люциферази з Photinus pyralis. Промотор РНІ227 у векторі PHI227-luc-kan містить нуклеотиди положень 1-1117 нуклеотидної послідовності SEQ ID NO 5 та може бути виділений з вектора за допомогою HindIII та Ncol та комбінований з іншими генами. Виявлення ІНДУКЦІЇ експресії генів РНІ7, РНІ20, РНІ153 та РНІ227 у зібраних коренях цукрових буряків Посівний матеріал цукрового буряка висіювали у поле навесні, і буряк вирощували згідно зі звичайною 3 UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 сільськогосподарською практикою. Корені рослин у віці 24 тижнів восени збирали та обробляли протягом 30 с у наявній на ринку бетономішалці (фірми Attika) для створення поверхневих пошкоджень та ударів, типових для механічного збирання. Негайно після цього запасаючі органи закладали на зберігання при 17°С та при 26-28°C. З буряків, що зберігалися при 17°С, відбирали проби (по 5 коренів) через 1 добу, 3 доби, 4 доби, 7 діб, 14 діб, 21 добу, 28 діб, 35 діб та 46 діб після збирання, а з буряків, що зберігалися при 26-28°С - через 1 добу, 3 доби, 4 доби, 7 діб та 14 діб після збирання, і виділяли з проб загальну клітинну PHK за Логеманом та ін (Logemann et аl., 1987). Дослідження експресії генів, індукованої після збирання виконували методом візуалізації плям PHK за Сембруком та ін. (Sambrook et аl., 1989). Для цього 10 мкг загальної клітинної PHK із проб листя та коренів, відібраних на полі безпосередньо перед збиранням (0d), а також із коренів, що зберігалися протягом 1-35 діб, розділяли на денатурувальному формальдегід-агарозному гелі. Відділену способом електрофорезу PHK способом нанесення плям за допомогою капілярів переносили на нейлонову мембрану Hybond N (виробник фірма Amersham Pharmacia Biotech, Freiburg). Радіоактивне мічення 20 нг кДНК-клону 32 індукованих після збирання (PHI) генів 7, 20, 153 та 227 50 мкКі P-dATP (6000 Кі/ммоль, Amersham Pharmacia Biotech, Freiburg) здійснювали за допомогою набору праймерів Prime-It II Random Primer Kit (фірма Stratagene GmbH, Heidelberg) згідно з інструкціями виробника. Після цього фільтр із PHK гібридизували з міченими зондами, промивали та експонували на рентгеноплівці. Ген РНІ7 перед збиранням не експресувався у листі та дуже слабко експресувався у коренях цукрового буряка. Після збирання та під час зберігання мала місце сильна експресія гена РНІ7 у коренях. При 28°С індукція була сильнішою, ніж при 17°С (Фіг. 1). Ген РНІ20 перед збиранням слабко експресувався у листі та у коренях цукрового буряка. Після збирання та під час зберігання мала місце сильна довготривала індукція гена РНІ20 у запасаючих органах. При 17°С та 28°С інтенсивність індукції була порівнянною (Фіг. 1). Ген РНІ153 перед збиранням не експресувався ні у листі, ні у коренях цукрового буряка. Після збирання та під час зберігання мала місце сильна індукція гена РНІ153 у коренях. При 17°C та 28°С інтенсивність індукції була порівнянною (Фіг. 1). Ген РНІ227 перед збиранням слабко експресувався у листі та у коренях цукрового буряка. Після збирання та під час зберігання мала місце сильна індукція та сильна довготривала експресія гена РНІ227 у коренях. При 17°C та 28°С інтенсивність індукції була порівнянною (Фіг. 1). Виявлення індукції експресії гена РНІ5 у зібраних коренях цукрових буряків Для виявлення індукції експресії РНІ5 після збирання коренів цукрового буряка був проведений експеримент методом RT-PCR. З 5 мкг загальної клітинної PHK із листя та коренів, відібраних на полі безпосередньо перед збиранням (0d), виготовляли сукупність кДНК за допомогою набору для синтезу RevertAid H Minus cDNA Synthese Kit (продукт фірми MBI Fermentas) згідно з інструкціями виробника. Подальші сукупності кДНК виготовляли з РНК, виділених із коренів буряка, які зберігалися протягом 1 доби, 4 діб, 7 діб, 14 діб, 21 доби, 28 діб, 35 діб та 46 діб зберігання при 17°C та протягом 1 доби, 3 діб, 4 діб, 7 діб та 14 діб при 26-28°С. Експресію гена РНІ5 виявляли з використанням праймерів РНІ5-1 (GTG CAA GGA TTC TGG CAC CCG TCG GTG G) та РНІ5-2 (GTA TGG GCC GCG GCA GAT CCA GGT AGC G) за допомогою Taq-полімерази (продукт фірми Q-Biogene) згідно з інструкціями виробника. Для контролю детектували експресію гена гліцеринальдегід-3-фосфат-дегідрогенази (GAPDH) з використанням праймерів GAPDH-1 (ATG TTT AAG TAC GAC AGT GTT CAC G) та GAPDH-2 (ATG TGA AGG TCT GAC TTG TAT TCG T) з метою посвідчення, що для реакцій RT-PCR застосовувалися однакові кількості кДНК. Дослідження методом RT-PCR показало, що перед збиранням (0d) ген РНІ5 слабко експресувався у листі та дуже слабко у коренях. У буряках, що зберігалися при 17°C та 28°С, виявлено сильну експресію і, отже, індукцію гена РНІ5 у запасаючих органах після збирання та на протязі зберігання (Фіг. 2). У буряках, що зберігалися при 17°С, у період з 1-го по 7-й день після збирання виявлено явно послаблену експресію гена РНІ5 у порівнянні з періодом з 14-го по 46-й день. У буряках, що зберігалися при 26-28°С, у 1-й та 3й дні після збирання виявлено явно послаблену експресію гена РНІ5 у порівнянні з 4-м, 7-м та 14-м днями. Експресія гена GAPDH була однаковою у всіх пробах (Фіг. 2). Ці результати показали, що експресія гена РНІ5 індукувалася після збирання. При цьому індукція на протязі перших діб зберігання, яка фізіологічно визначається наслідками механічних пошкоджень, явно послаблена у порівнянні із фазою подальшого зберігання та пов'язаних із ним фізіологічних змін. Класифікація РНІ-ПРОМОТОРІВ Усі промотори РНІ5, РНІ7, РНІ20, РНІ153 та РНІ227 індукуються після збирання у коренях цукрового буряка в ході процесу збирання та подальшого зберігання (Фіг. 1 та Фіг. 2). Поряд з цією загальною властивістю індукції після збирання згадані п'ять промоторів відрізняються один від одного з точки зору активності у грунті перед збиранням, впливу механічних пошкоджень та зберігання на кінетику індукції та впливу температури зберігання на інтенсивність індукції промоторів. Ці відмінності дозволяють виділити чотири підкласи промоторів, що індукуються після збирання (Таблиця 1). Промотор PHI 153 належить до першого підкласу РНІ-промоторів, для яких транскрипт гена у 4 UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 запасаючому органі перед збиранням не виявляється. Промотори РНІ20 та РНІ227 належать до другого підкласу РНІ-промоторів, які перед збиранням виявляють слабку активність у запасаючому органі та індукція яких після збирання як при відносно низьких (17°C), так і при підвищених (28°C) температурах має однакову інтенсивність і, отже, не залежить від температури. Промотор РНІ7 належить до третього підкласу РНІ-промоторів, які перед збиранням виявляють слабку активність у запасаючому органі та індукція яких при підвищених (28°C) температурах є явно інтенсивнішою, ніж при відносно низьких (17°C) температурах. Промотор РНІ5 належить до четвертого підкласу РНІ-промоторів, які перед збиранням виявляють слабку активність у запасаючому органі та активність яких помірно зростає внаслідок механічних пошкоджень при збиранні та сильно зростає під час зберігання. Злиття ПРОМОТОРІВ РНІ5, РНІ7, РНІ20, РНІ153 та РНІ227 з геном люциферази з Photinus pyralis Для виявлення активності промоторів РНІ5, РНІ7, РНІ20, РНІ153 та РНІ227 у цукровому буряку ці промотори були трансляційно злиті з геном люциферази з Photinus pyralis та трансформовані у цукровому буряку. Для цього промотор РНІ5 був клонований у лінеаризованому Hindlll-BspHI бінарному векторі pGPTV-kan у формі фрагмента Hindlll-BspHI, а промотори РНІ7, РНІ20, РНІ153 та РНІ227 - як фрагмент Hindlll-Ncol у тому самому векторі, лінеаризованому Hindlll-Ncol (Беккер та ін. - Becker et al., 1992). Одержані вектори мають позначення PHI5-luc-kan (Фіг. 3), PHI7-luc-kan (Фіг. 4), PHI20-luc-kan (Фіг. 5), PHI153-luc-kan (Фіг. 6) та PHI227-luc-kan (Фіг. 7). Ці бінарні вектори були трансформовані у бактерії Agrobacterium tumefaciens, штам С58С1, з резидентною плазмідою pGV2260 методом прямої трансформації ДНК (Ан - An, 1987). Селекцію рекомбінантних клонів A. tumefaciens виконували із застосуванням антибіотика канаміцину (50 мг/л). Трансформацію цукрових буряків виконували за Ліндсеєм та ін. (Lindsey et аl., 1991) із застосуванням антибіотика канаміцину. Трансгенність рослин перевіряли методом PCR. Застосування праймерів GTGGAGAGGCTATTCGGTA та CCACCATGATATTCGGCAAG викликало збільшення фрагмента ДНК розміром 553 пари основ з гена nptII. Реакцію PCR виконували із застосуванням 10 нг геномної ДНК, концентрації праймера 0,2 мкМ при температурі замикання циклу 55°С у середовищі Multicycler РТС-200 (фірма MJ Research, Watertown, USA). Ген люциферази вектора PHI5-luc-kan (Фіг. 3) можна вивільнити у формі фрагмента BspHI-SacI, а ген люциферази векторів PHI7-luc-kan (Фіг. 4), PHI20-luc-kan (Фіг. 5), PHI153-luc-kan (Фіг. 6) та PHI227-luc-kan (Фіг. 7) - у формі фрагмента NcoI-SacI, і замінити іншим геном, який має бути експресований, наприклад, геном інгібітора інвертази. Для цього ген, експресія якого необхідна, має також бути у формі фрагмента NcoI-SacI. За альтернативним варіантом, промотор РНІ5 можна виділити у формі фрагмента Hindlll-BspHI, а промотори РНІ7, РНІ20, РНІ153 та РНІ227 - у формі фрагмента Hindlll-Ncol, та ввести їх у відповідні вектори експресії. Виявлення активності промотору РНІ5 v коренях цукрового буряка під час зберігання Трансгенні цукрові буряки, трансформовані конструкцією гена-репортера PHI5-luc-kan, вирощували в умовах теплиці. У віці 20 тижнів рослини збирали, і корені зберігали протягом 6 тижнів. Активність промотору визначали у листі та коренях перед збиранням та щотижня у коренях протягом зберігання шляхом вимірювання активності гена-репортера. Активність люциферази Photinus pyralis визначали за допомогою системи Luciferase Assay System (фірма Promega, Mannheim, Німеччина) у люмінометрі Sirius (Berthold Detection System GmbH, Pforzheim, Німеччина) згідно з інструкціями виробника. Для одержання придатного для аналізу екстракту ферменту спочатку визначали масу проби тканини. Проби листя гомогенізували з додаванням морського піску у 10кратному об'ємі (на одиницю маси) буфера пасивного лізису (PLB) у ступці, а проби коренів - у наявному на ринку кухонному подрібнювачі (Warring Blender). Надосадову рідину переносили у пробірку Еппендорфа місткістю 1,5 мл та центрифугували при 4°C протягом 5 хв при 20000 g. Відбирали прозору надосадову рідину та додавали до неї 10 мкл неочищеного екстракту для вимірювання активності люциферази Photinus. Активність промотору РНІ5 у листі та коренях перед збиранням була низькою. Навпаки, у зібраних та зберіганих коренях восьми незалежних трансформантів активність цього промотору зростала сильно або дуже сильно. Згідно з результатами дослідження гена-репортера, які узгоджуються з результатами RTPCR, активність промотору РНІ5 сильно індукується у коренях цукрового буряка після збирання. Трансгенні рослини з особливими властивостями Із застосуванням промотору відповідно до цього винаходу можна одержати трансгенні рослини з особливими властивостями: a) покращеним метаболізмом вуглеводів у запасаючих органах після збирання, b) покращеним метаболізмом азоту у запасаючих органах після збирання, c) покращеною стійкістю проти висихання та покращеним водним статусом у запасаючих органах після збирання, 5 UA 97622 C2 5 10 15 20 25 30 35 40 45 50 55 60 d) покращеною холодостійкістю та морозостійкістю запасаючих органів після збирання, e) підвищеною резистентністю або стійкістю запасаючих органів проти патогенних організмів після збирання, f) покращеним вторинним обміном речовин у запасаючих органах після збирання. Покращений метаболізм вуглеводів у запасаючих органах після збирання Метаболізм вуглеводів у запасаючих органах рослин можна у різних відношеннях покращити за допомогою промотора відповідно до цього винаходу. Механічні впливи під час збирання та фізіологічні зміни під час зберігання цукрових буряків та моркви можуть спричинити посилений гідроліз та зниження вмісту сахарози та нагромадження інвертованого цукру (Бурба - Burba, 1973; Смед та ін. - Smed et аl., 1996; Галіндо та ін. - Galindo et аl., 2004). Пов'язані із цим втрати цукру, що мають порядок від 0,01% до 0,025% на добу, знижують кількість цукру, що може бути добутий із цукрових буряків. Утворення інвертованого цукру спричиняє технологічні утруднення при промисловому добуванні цукру (Бурба, 1976). Промотори відповідно до цього винаходу можна використати для послаблення гідролізу сахарози та утворення інвертованого цукру. Для цієї мети слід забезпечити посилення експресії гена інгібітора інвертази у зібраних коренях або пригнітити активність гена або генів сахарозосинтетази або гена інвертази шляхом застосування антисенсибілізатора або інформаційної РНК. Промотори відповідно до цього винаходу можна застосовувати також для забезпечення підвищеного виходу поліфруктанів, наприклад, інуліну, та продукування молекул інуліну з подовженими ланцюгами у коренях цикорію. Для цього слід послабити розщеплення інуліну у зібраному цикорії або запобігти такому розщепленню, для чого послабити активність фруктозил-екзогідролази у зібраних коренях шляхом застосування антисенсибілізатора або інформаційної РНК. Покращений метаболізм азоту у запасаючих органах після збирання Промотор відповідно до цього винаходу можна застосовувати для зниження у запасаючих органах рослин після збирання вмісту «шкідливого азоту», що піддається екстракції, наприклад, амінокислот. Підвищені концентрації азотвмісних сполук у запасаючих органах рослин часто знижують поживнофізіологічну цінність продукції або утруднюють добування нагромаджених речовин, наприклад, сахарози з коренів цукрових буряків. Шляхом посилення експресії відповідних ферментів, транскрипційних факторів, нагромаджувальних протеїнів тощо можна досягти посилення синтезу протеїнів з амінокислот у запасаючих органах. На відміну від амінокислот, протеїни при добуванні цукру з цукрових буряків випадають в осад і тому не екстрагуються. Покращення стійкості запасаючих органів проти фітопатогенних ґрунтових грибків та бактерій Промотор відповідно до цього винаходу у поєднанні з геном або комбінацією генів можна застосувати також для забезпечення прямої або непрямої протигрибкової дії у запасаючих органах рослин. Протигрибкова дія забезпечує підвищену резистентність або стійкість проти грибків після збирання та під час зберігання. Для цієї мети промотор трансляційно або транскрипційно зливають із генами захисту проти патогенних факторів, генні продукти яких виявляють пряму або непряму протигрибкову або антибактеріальну дію. Комбінації промотору з геном клонують у бінарний трансформаційний вектор pGPTV та трансформують трансформат, одержаний за допомогою A. tumefaciens, у цукрові буряки, моркву або картоплю. Трансгенність рослин перевіряють, як описано вище, методом PCR, а експресію генів у коренях або бульбах контролюють методом візуалізації плям РНК. Підвищену резистентність запасаючих органів проти грибків або бактерій спостерігають при випробуваннях на резистентність. Несподівано виявлено, що індукована після збирання експресія генів захисту від патогенних факторів не спричиняла малорослості або зниження врожайності, які часто спостерігалися при конститутивній експресії під час вегетативного розвитку рослин (Гейль та Болдуїн - Heil and Baldwin, 2002). Посилання Altschul S.F. et al. (1990). Basic Local Alignment search tool, J. Моl. Biol. 215: 403-410. An G. (1987). Binary Ti vectors for plant transformation and promoter analysis. Methods Enzymol. 153, 292305. Becker D., Kemper E., Schell J., and Masterson R. (1992). New plant binary vectors with selectable markers located proximal to the left T-DNA border. Plant Моl. Biol. 20(6): 1195-1197. Burba M. (1976). Atmung und Saccharosestoffwechsel lagernder Zuckerrüben. Zeitschrift für die Zuckerindustrie 26: 647-658. Galindo F.G., Herppich W., Gekas V., and Sjöholm I. (2004). Factors affecting quality and postharvest properties of vegetables: Integration of water relations and metabolism. Critical Reviews in Food Science and Nutrition 44: 139-154. Heil M. and Baldwin I.T. (2002). Fitness costs of induced resistance: emerging experimental support for a slippery concept. Trends Plant Sci. 2002 Feb.; 7(2): 61-67. Lindsey K,. Gallois P., and Eady C (1991) Regeneration and transformation of sugar beet by Agrobacterium tumefaciens. Plant Tissue Culture Manual B7: 1-13; Kluwer Academic Publishers. 6 UA 97622 C2 5 Logemann E., Parniske M., Hahlbrock K. (1995). Modes of expression and common structural features of the complete phenylalanine ammonia-lyase gene family in parsley. Proc. Natl. Acad. Sci. USA. 1995 92(13): 59055909. Nilsson T. (2000). Postharvest storage and handling of vegetables., In Fruit and Vegetable Quality (an integrated view), Seite 96-121. Herausgeber Shewfew R.L. and Bruckner B., Technomic Publishing Inc. Sambrook J., Fritsch E.F., and Maniatis T. (1989). In Molecular Cloning, A Laboratrory Manual (Cold Spring Harbor Laboratory Press, New York). Smed E., Augustinussen E., and Stensen J.K. (1996). Loss of sugar in injured sugar beet losses from lifting, th storing and washing. Proceedings of the 59 IIRB Congress, 533-545. 10 7 UA 97622 C2 8 UA 97622 C2 9 UA 97622 C2 10 UA 97622 C2 11 UA 97622 C2 ФОРМУЛА ВИНАХОДУ 5 10 15 20 1. Промотор з органо-специфічною активністю у рослинах цукрового буряку, який у запасаючих органах рослин при зберіганні при 17 ºС або при 26-28 ºС виявляє вищу активність, ніж в інших частинах цих рослин, та тим, що його активність після збирання запасаючих органів змінюється та стає вищою, ніж перед їх збиранням. 2. Промотор за п. 1, який відрізняється тим, що його активність у запасаючих органах піддається вимірюванню методом візуалізації плям РНК, і активність перед збиранням запасаючих органів за порівнюваних умов дослідження виявляється меншою ніж 20 %, відповідно до варіанта, якому віддається перевага, меншою ніж 10 %, та, зокрема, менше ніж 5 %. 3. Промотор, який містить: a) послідовність нуклеотидів за SEQ ID NO: 1 або b) послідовність нуклеотидів за SEQ ID NO: 2 або, c) послідовність нуклеотидів за SEQ ID NO: 3 або, d) послідовність нуклеотидів за SEQ ID NO: 4 або, е) послідовність нуклеотидів за SEQ ID NO: 5 або, f) послідовність нуклеотидів, комплементарну до послідовностей а)-е), або g) послідовність нуклеотидів, гібридизовану з однією з послідовностей нуклеотидів a)-f). 4. Похідні промотору за п. 3, що мають функціональну активність промотору за п. 1. 12 UA 97622 C2 5 10 15 20 5. Вектор для трансформації Agrobacterium tumefaciens, який включає промотор за будь-яким з пп. 1-3 або похідну за п. 4. 6. Еукаріотна або прокаріотна клітина-хазяїн, яка включає промотор за будь-яким з пп. 1-3 або похідну за п. 4. 7. Трансгенна рослина цукрового буряку або її частини, які включають промотор або похідні за одним із пп. 1-4. 8. Трансгенна рослина або її частини за п. 7, трансформовані промотором за будь-яким з пп. 13 або похідним за п. 4 для впливу на метаболізм у запасаючих органах. 9. Трансгенна рослина або її частини за п. 7, яка відрізняється тим, що рослиною є Beta vulgaris. 10. Трансгенне насіння рослини за п. 7 або 8 для продукування трансгенної рослини або її частин за п. 7 або 8. 11. Застосування промотору за одним із пп. 1-3 або похідних за п. 4 для одержання трансгенної рослини цукрового буряку, що має одну або кілька з наведених нижче властивостей: a) покращений метаболізм вуглеводів у запасаючих органах після збирання, b) покращений метаболізм азоту у запасаючих органах після збирання, c) покращену стійкість проти висихання та покращений водний статус у запасаючих органах після збирання, d) покращену холодостійкість та морозостійкість запасаючих органів після збирання, e) підвищену резистентність або стійкість запасаючих органів проти патогенних організмів після збирання, f) покращений вторинний обмін речовин у запасаючих органах після збирання. 13 UA 97622 C2 14 UA 97622 C2 15 UA 97622 C2 16 UA 97622 C2 17 UA 97622 C2 Комп’ютерна верстка А. Крулевський Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 18
ДивитисяДодаткова інформація
Назва патенту англійськоюPromoter with organo-specific activity in sugar beet plants, induced at storing
Автори англійськоюHell Reinhardt, Rottues Alexander, STAHL DIETMAR JUERGEN
Назва патенту російськоюПромотор с органоспецифической активностью в растениях сахарной свеклы, который индуцируется при хранении
Автори російськоюХель Райнхард, Роттуес Александер, Шталь Дитмар Юрген
МПК / Мітки
МПК: C12N 15/82, A01H 5/00
Мітки: промотор, органо-специфічною, буряку, рослинах, індукується, зберіганні, цукрового, активністю
Код посилання
<a href="https://ua.patents.su/20-97622-promotor-z-organo-specifichnoyu-aktivnistyu-u-roslinakh-cukrovogo-buryaku-shho-indukuehtsya-pri-zberiganni.html" target="_blank" rel="follow" title="База патентів України">Промотор з органо-специфічною активністю у рослинах цукрового буряку, що індукується при зберіганні</a>
Промотор для епідерміс-специфічної трансгенної експресії в рослинах

Номер патенту: 86781
Опубліковано: 25.05.2009
Автори: Дудлер Роберт, Швайцер Патрік, Панструга Ральф, Шульце-Леферт Пауль
МПК: C12N 15/82
Мітки: трансгенної, промотор, експресії, рослинах, епідерміс-специфічної
Формула / Реферат:
1. Промоторний регіон зі специфічністю до рослинного епідермісу, що включає першу послідовність, що походить із промотору гену GSTA1 та являє собою SEQ ID No 1, та другу послідовність, що походить із інтрону гену WIR1a та являє собою SEQ ID No 2, причому перша послідовність та/або друга послідовність промоторного регіона має ідентичність послідовності від щонайменше 90 % з промоторними послідовностями, наведеними в SEQ ID No 1 та/або SEQ ID...
Рослинний матеріал цукрового буряку, стійкий до імідазолінонового гербіциду, спосіб надання рослинам буряку стійкості до гербіцидів та спосіб боротьби з бур’янами

Номер патенту: 72429
Опубліковано: 15.03.2005
Автори: Пеннер Дональд, Райт Террі Р.
МПК: C12N 15/09, A01H 5/00, C12N 9/88, A01H 1/04, C12N 5/10
Мітки: цукрового, буряку, стійкості, надання, гербіцидів, імідазолінонового, рослинній, боротьби, гербіциду, бур'янами, стійкий, спосіб, матеріал, рослинам
Формула / Реферат:
1. Рослинний матеріал цукрового буряку, який складається з мутантних клітин з мутантним геном ацетолактатсинтази, що кодує цю синтазу, в якому нуклеотид змінено з гуаніну на аденін у позиції 337, мутантні клітини мають стійкість до імідазолінонового гербіциду, і ця стійкість здатна передаватися звичайним кросбридингом рослин, виведених з цих клітин, і клітини здатні регенерувати у рослину.2. Матеріал за п. 1, який відрізняється тим, що...
Рослинний матеріал цукрового буряку, стійкий до імідазолінонового та сульфонілсечовинного гербіциду, спосіб надання рослинам буряку стійкості до гербіцидів та спосіб боротьби з бур’янами

Номер патенту: 72428
Опубліковано: 15.03.2005
Автори: Пеннер Дональд, Райт Террі Р.
МПК: C12N 5/10, C12N 9/88, A01H 1/04, A01H 5/00, C12N 15/09, C12N 5/00
Мітки: гербіциду, рослинній, імідазолінонового, матеріал, цукрового, стійкий, буряку, гербіцидів, боротьби, рослинам, стійкості, бур'янами, спосіб, сульфонілсечовинного, надання
Формула / Реферат:
1. Рослинний матеріал цукрового буряку, який складається з мутантних клітин з мутантним геном ацетолактатсинтази, що кодує цю синтазу, в якому нуклеотиди змінено з цитозину на тимін у позиції 562 та з гуаніну на аденін у позиції 337, мутантні клітини мають стійкість як до імідазолінонових, так і до сульфонілсечовинних гербіцидів, і ця стійкість передається звичайним кросбридингом рослин, виведених з цих клітин, і клітини здатні регенерувати у...
Спосіб вирощування цукрового буряку

Номер патенту: 34684
Опубліковано: 15.03.2001
Автори: Герасименко Станіслав Михайлович, Пономаренко Сергій Платонович, Сакало Валентина Дмитрівна
МПК: A01C 1/00, A01N 43/46, A01P 21/00
Мітки: цукрового, буряку, спосіб, вирощування
Формула / Реферат:
Спосіб вирощування цукрового буряку, що включає передпосівну обробку насіння, висівання його в грунт, обробку посівів у період вегетації, вирощування в природних умовах до збору коренеплодів, який відрізняється тим, що передпосівну обробку насіння проводять регулятором росту бетастимуліном з розрахунку 15 мл на 1 т насіння, в період вегетації обробку посівів у фазі від 8-10 листків до змикання рядків проводять емістимом С з розрахунку 10 мл у...
Регулятор росту цукрового буряку

Номер патенту: 30087
Опубліковано: 17.12.2001
Автори: Пономаренко Сергій Платонович, Боровикова Галина Семенівна, Герасименко Станіслав Михайлович
МПК: C07D 307/62, A01N 43/40, A01P 21/00
Мітки: росту, регулятор, буряку, цукрового
Формула / Реферат:
Регулятор росту цукрового буряку, що містить в основі емістим С, який відрізняється тим, що додатково містить комплекс N-оксиду 2,6-диметилпіридину з щавлевою кислотою при такому співвідношенні компонентів, мас., %: емістим С 80-90 комплекс N-оксиду 2,6-диметилпіридину 4-9 вода решта.