Малярійні вакцини
Номер патенту: 95293
Опубліковано: 25.07.2011
Автори: Ядава Анджалі, Маршан Мартін, Оукенгаус Крістіан Ф., Коен Джозеф Д.







Формула / Реферат
1. Імуногенна білкова частинка, яка включає наступні мономери:
a. злитий білок, що включає послідовності, які походять від CS білка P. vivax та S антигену гепатиту B (CSV-S), та
b. злитий білок, що включає послідовності, які походять від CS білка P. falciparum та S антигену вірусу гепатиту B (RTS), та
c. необов‘язково, S антиген, що походить від гепатиту B.
2. Частинка згідно з будь-яким з попередніх пунктів, де антиген вірусу гепатиту В є таким, що походить від adw серотипу.
3. Частинка згідно з будь-яким з попередніх пунктів, яка додатково включає один або більше додаткових антигенів, що походять від P. falciparum та/або P. vivax.
4. Частинка згідно з будь-яким з пунктів 1-4, де компонент Р. vivax являє собою гібридний поліпептид, вибраний з DBP, PvTRAP, PvMSP2, PvMSP4, PvMSP5, PvMSP6, PvMSP7, PvMSP8, PvMSP9, PvAMA1 та RBP.
5. Частинка згідно з будь-яким з пунктів 1-4, яка включає послідовність гібридного білка, що оточує спорозоїт, як показано у SEQ ID No 13.
6. Композиція, що включає частинку згідно з будь-яким з попередніх пунктів та ад‘ювант.
7. Композиція згідно з пунктом 6, де ад‘ювант є вибраним з групи, що включає:
• солі металу, такі як гідроксид алюмінію або фосфат алюмінію,
• емульсії масло-у-воді,
• агоністи Тoll-подібних рецепторів (такі, як агоніст Toll-подібного рецептора 2, агоніст Toll-подібного рецептора 3, агоніст Toll-подібного рецептора 4, агоніст Toll-подібного рецептора 7, агоніст Toll-подібного рецептора 8 та агоніст Toll-подібного рецептора 9),
• сапоніни, наприклад, Quil A та його похідні, такі, як QS7 та/або QS21,
• CpG-вмісні олігонуклеотиди,
• 3D-MPL,
•(2-дезокси-6-o-[2-дезокси-2-[(R)-3-додеканоїлокситетрадеканоїламіно]-4-o-фосфоно-β-D-глюкопіранозил]-2-[(R)-3-гідрокситетрадеканоїламіно]-α-D-глюкопіранозилдигідрофосфат),
• DP (3S,9 R)-3-[(R)-додеканоїлокситетрадеканоїламіно]-4-oксo-5-aзa-9(R)-[(R)-3-гідрокситетрадеканоїламіно]декан-1,10-діол,1,10-біс(дигідрофосфат), та
• MP-Ac DP (3S-, 9R)-3-[(R)-додеканоїлокситетрадеканоїламіно]-4-oксo-5-aзa-9-[(R)-3-гідрокситетрадеканоїламіно]декан-1,10-діол,1-дигідрофосфат 10-(6-аміногексаноат),
або їх комбінації.
8. Композиція згідно з пунктом 6 або 7, де ад‘ювант є вибраним з групи, що включає:
• сапонін, асоційований із сіллю металу, такою, як гідроксид алюмінію або фосфат алюмінію,
• 3D MPL, QS21 та CpG олігонуклеотид, наприклад, у вигляді композиції масло-у-воді,
• сапонін у формі ліпосоми, наприклад, що додатково включає QS21 та стерин, та
• ISCOM.
9. Композиція згідно з будь-яким з пунктів 6-8, яка додатково включає один або більше додаткових антигенів, що походять від P. falciparum та/або P. vivax, у суміші.
10. Композиція згідно з будь-яким з пунктів 6-9, де композиція є вакциною для парентерального застосування.
11. Частинка, як визначено у будь-якому з пунктів 1-5, або композиція, як визначено у будь-якому з пунктів 6-10, для застосування у лікуванні.
12. Застосування білка, як визначено у будь-якому з пунктів 1-5, або композиції згідно з будь-яким з пунктів 6-10 для виробництва лікарського засобу для лікування/запобігання малярії.
13. Спосіб лікування пацієнта, чутливого до інфекції Plasmodium, що включає введення ефективної кількості білка, як визначено у будь-якому з пунктів 1-5, або композиції, як визначено у будь-якому з пунктів 6-11, зокрема, у вигляді вакцини.
14. Хазяїн (вектор/плазміда), що кодує послідовності ДНК, які є компонентами частинки, як визначено у будь-якому з пунктів 1-5, де вказаний вектор/плазміда є прийнятною(им) для вбудовування ДНК у хазяїна, для виробництва релевантної частинки, або вказана плазміда є здатною за прийнятних умов до виробництва компонентів, які поєднуються у релевантну частинку.
15. Спосіб одержання частинки, як визначено у будь-якому з пунктів 1-5, де спосіб включає експресію нуклеотидної послідовності, що кодує вказані білки, у прийнятному хазяїні та відновлення продукту.
16. Спосіб згідно з пунктом 15, де хазяїн являє собою дріжджі.
17. Спосіб згідно з пунктом 16, де дріжджі є вибраними з групи, яка включає:
Saccharomyces, Schizosaccharomycees, Kluveromyces, Pichia (наприклад Pichia pastoria), Hansenula, (наприклад Hansenula polymorpha), Yarowia, Schwaniomyces, Schizosaccharomyces, Zygoaccharomyces, такі як Saccharomyces cerevisiae, S. carlsberoensis, K. lactis, Y1834 та DC5.
18. Спосіб згідно з будь-яким з пунктів 16-17, де продукт відновлюють шляхом лізису хазяйської клітини при обробці прийнятною композицією, що включає поверхнево-активну сполуку.
19. Спосіб згідно з пунктом 18, де поверхнево-активна сполука є вибраною з групи, що включає: Tвін (такий, як Tвін 20), поліоксіетиленові етери, поліетиленгліколь.
20. Продукт, який одержаний за способом, описаним у будь-якому з пунктів 15-19.
Текст
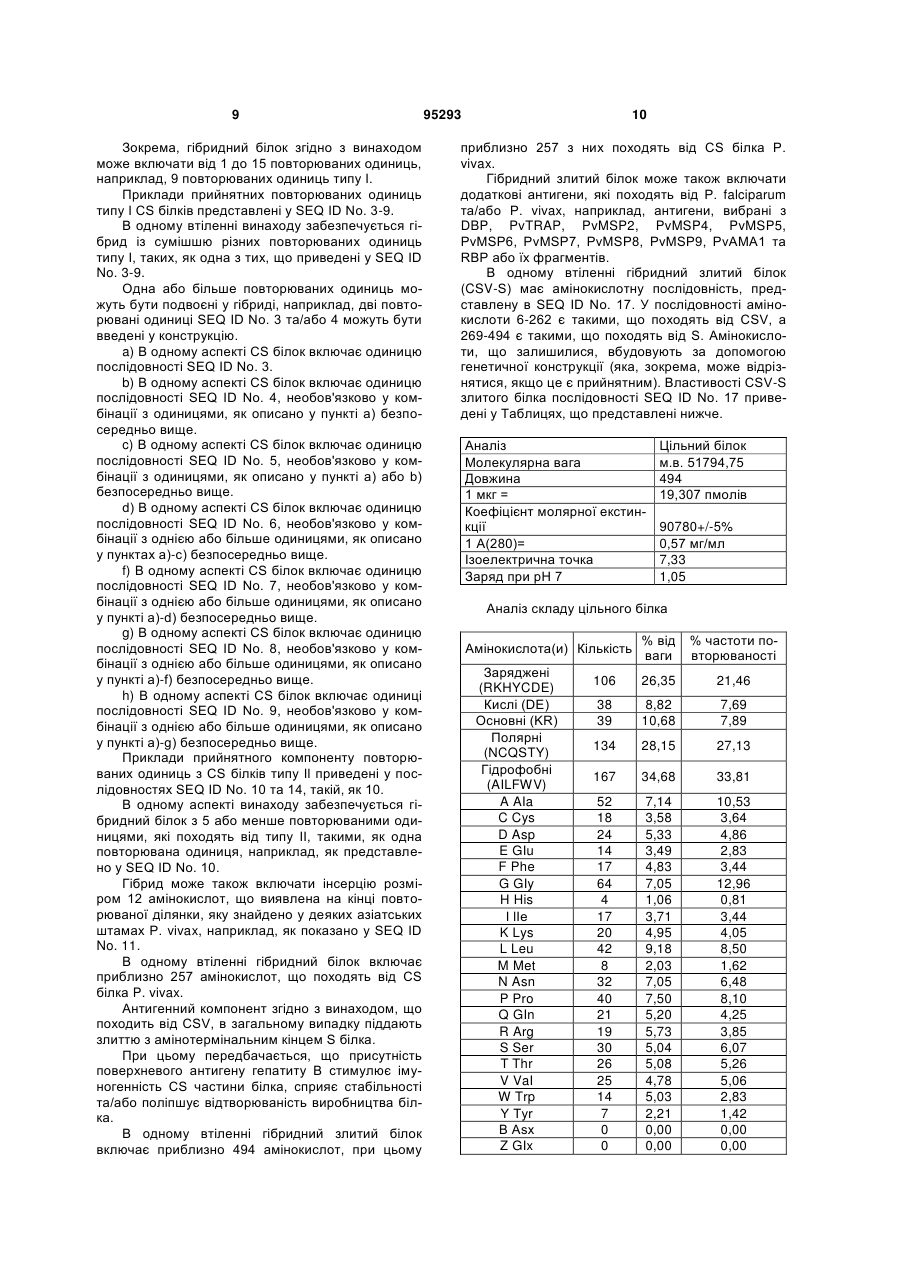
1. Імуногенна білкова частинка, яка включає наступні мономери: a. злитий білок, що включає послідовності, які походять від CS білка P. vivax та S антигену гепатиту B (CSV-S), та b. злитий білок, що включає послідовності, які походять від CS білка P. falciparum та S антигену вірусу гепатиту B (RTS), та c. необов„язково, S антиген, що походить від гепатиту B. 2. Частинка згідно з будь-яким з попередніх пунктів, де антиген вірусу гепатиту В є таким, що походить від adw серотипу. 3. Частинка згідно з будь-яким з попередніх пунктів, яка додатково включає один або більше додаткових антигенів, що походять від P. falciparum та/або P. vivax. 4. Частинка згідно з будь-яким з пунктів 1-4, де компонент Р. vivax являє собою гібридний поліпептид, вибраний з DBP, PvTRAP, PvMSP2, PvMSP4, 2 (19) 1 3 95293 4 9. Композиція згідно з будь-яким з пунктів 6-8, яка додатково включає один або більше додаткових антигенів, що походять від P. falciparum та/або P. vivax, у суміші. 10. Композиція згідно з будь-яким з пунктів 6-9, де композиція є вакциною для парентерального застосування. 11. Частинка, як визначено у будь-якому з пунктів 1-5, або композиція, як визначено у будь-якому з пунктів 6-10, для застосування у лікуванні. 12. Застосування білка, як визначено у будь-якому з пунктів 1-5, або композиції згідно з будь-яким з пунктів 6-10 для виробництва лікарського засобу для лікування/запобігання малярії. 13. Спосіб лікування пацієнта, чутливого до інфекції Plasmodium, що включає введення ефективної кількості білка, як визначено у будь-якому з пунктів 1-5, або композиції, як визначено у будь-якому з пунктів 6-11, зокрема, у вигляді вакцини. 14. Хазяїн (вектор/плазміда), що кодує послідовності ДНК, які є компонентами частинки, як визначено у будь-якому з пунктів 1-5, де вказаний вектор/плазміда є прийнятною(им) для вбудовування ДНК у хазяїна, для виробництва релевантної частинки, або вказана плазміда є здатною за прийнятних умов до виробництва компонентів, які поєднуються у релевантну частинку. 15. Спосіб одержання частинки, як визначено у будь-якому з пунктів 1-5, де спосіб включає експресію нуклеотидної послідовності, що кодує вказані білки, у прийнятному хазяїні та відновлення продукту. 16. Спосіб згідно з пунктом 15, де хазяїн являє собою дріжджі. 17. Спосіб згідно з пунктом 16, де дріжджі є вибраними з групи, яка включає: Saccharomyces, Schizosaccharomycees, Kluveromyces, Pichia (наприклад Pichia pastoria), Hansenula, (наприклад Hansenula polymorpha), Yarowia, Schwaniomyces, Schizosaccharomyces, Zygoaccharomyces, такі як Saccharomyces cerevisiae, S. carlsberoensis, K. lactis, Y1834 та DC5. 18. Спосіб згідно з будь-яким з пунктів 16-17, де продукт відновлюють шляхом лізису хазяйської клітини при обробці прийнятною композицією, що включає поверхнево-активну сполуку. 19. Спосіб згідно з пунктом 18, де поверхневоактивна сполука є вибраною з групи, що включає: Tвін (такий, як Tвін 20), поліоксіетиленові етери, поліетиленгліколь. 20. Продукт, який одержаний за способом, описаним у будь-якому з пунктів 15-19. Даний винахід відноситься до нових ліпопротеїнових частинок, способів їх одержання та очистки, їх застосування у медицині, зокрема, у запобіганні малярійних інфекцій, до композицій/вакцин, що містять білок або антитіла проти білкових частинок, таких, як моноклональні або поліклональні антитіла, та до їх застосування, зокрема, у терапії. Малярія представляє собою одну з основних проблем охорони здоров'я у світі з більше, ніж 2-4 мільйонами людей, які вмирають від цього захворювання кожного року. Одна з найбільш переважаючих форм захворювання спричинюється протозойним паразитом Р. vivax, який виявлений у тропічних та субтропічних районах. Цікавим є те, що паразит може повністю завершувати свій москітний цикл при температурах, нижчих 15°C, що дозволяє йому розповсюджуватися у районах помірного клімату. Одна з найбільш гострих форм цього захворювання спричинюється протозойним паразитом Plasmodium falciparum, що є відповідальним за найбільшу смертність, притаманну малярії. Життєвий цикл Plasmodium є складним та потребує двох хазяїв, людини та москіта, для свого завершення. Інфекція у людини ініціюється інокуляцією спорозоїтів у слину інфікованого москіта. Спорозоїти переміщуються до печінки та там інфікують гепатоцити, де вони піддаються диференціації через екзоеритроцитарну внутрішньоклітинну стадію у стадію мерозоїту, який інфікує еритроцити крові (RBC) для ініціації циклічної реплікації на безстатевій стадії у крові. RBC цикл завершується диференціацією ряду мерозоїтів у статеву стадію гаметоцитів, які споживаються москітом, там вони розвиваються через серію стадій у середній кишці з виникненням спорозоїтів, які переміщуються у слинну залозу. Завдяки тому факту, що захворювання, яке спричинюється P. vivax, лише в деяких випадках буває летальним, зусилля щодо запобігання та лікування малярії були сфокусовані на більш смертельній формі захворювання, що спричинюється Plasmodium falciparum (P. falciparum). Незважаючи на те, що захворювання, спричинене P. vivax, звичайно не приводить до смерті пацієнта, завдяки кількості випадків, яка зростає, значному впливу на якість життя пацієнта, зростанню кількості повідомлень про тяжкі випадки захворювання, що приводять до анемії та смерті, економічним втратам, залишається необхідність в ефективній вакцинації проти цього захворювання. Крім того, одна вакцина, що є здатною забезпечити захист проти обох причин цього захворювання, буде бажаною. Одна з характеристик P. vivax полягає у тому, що деякі штами є здатними спричинювати затриману інфекцію, залишаючись у латентному стані у печінці перед появою у периферичній кровоносній системі до прояву клінічних симптомів. Таким чином, індивідууми, наприклад, коли подорожують через інфіковані райони, можуть бути інфіковані та можуть не проявляти симптомів протягом кількох місяців. Це спричинює існування можливості поширення захворювання, з цієї причини особам, які подорожують до інфікованих районів, не дозволяється здавати кров для переливання протягом 5 визначеного періоду часу після подорожі до інфікованого району. Малярійна інфекція, спричинена P. vivax, залишається латентною у печінці, хоча паразит піддається пре-еритроцитарній шизогонії. Якщо паразит контролюється на цій стадії, перед тим, як він зникає з печінки, у пацієнта не спостерігається ніяких клінічних симптомів захворювання. Стадія спорозоїту Plasmodium була ідентифікована як потенційна мішень малярійної вакцини. Вакцинація при використанні дезактивованого (опроміненого) спорозоїту була показана як така, що індукує захист проти експериментальної малярії людини (Am. J, Trop. Med. Hyg 24: 297-402, 1975). Проте не існувало можливості практично та технічно виробляти малярійну вакцину для загальної популяції на основі цієї методики, використовуючи опромінені спорозоїти. Основний поверхневий білок спорозоїту є відомим як білок, що оточує спорозоїт (CS білок). Як передбачається він є втягненим у рухливість та інвазію спорозоїту під час його проходження від початкового сайту інокуляції москітом у систему кровообігу, коли він мігрує до печінки. CS білок видів Plasmodia характеризується центральним повторюваним доменом (повторювана ділянка), що є фланкованим неповторюваними аміно (N-термінальними) та карбокси (Стермінальними) фрагментами. Центральний домен Р. vivax складається з кількох блоків повторюваної одиниці, що звичайно містить тандем з 9 амінокислот. У деяких азіатських штамах після центральної повторюваної ділянки є присутньою додаткова послідовність, що складається приблизно з 12 амінокислот (див. SEQ ID No. 11). Функція останньої є невідомою. Проте деякі вважають, що вказані амінокислоти можуть бути пов'язаними із затриманим початком виявлення клінічних симптомів захворювання, проте ця гіпотеза не була досліджена. Передбачається, що N-термінальна ділянка характеризується послідовністю з 5 амінокислот, що є відомою як ділянка І (див. SEQ ID No. 1). Також передбачається, що С-термінальна ділянка характеризується тим, що вона включає послідовність з 18 амінокислот, що є відомою як ділянка II. Остання містить фрагмент адгезії до клітини, який є високо консервативним серед усіх малярійних CS білків (див. SEQ ID No. 2). Декілька груп запропонували субодиничні вакцини на основі білка, що оточує спорозоїт. Дві з цих двох вакцин піддавалися клінічному випробуванню; одна являє собою синтетичний пептид, інша є рекомбінантним білком (Ballou та ін., Lancet: і 1277 (1987) та Herrington та ін. Nature 328:257 (1987)). Ці вакцини були успішними у стимуляції анти-спорозоїтної відповіді. Проте величина відповіді була невтішною, при цьому деякі вакцини взагалі не давали відповіді. Крім того, відсутність «бустингу» рівнів антитіла після подальших ін'єкцій та результати in vitro аналізів проліферації лімфоцитів дають змогу запропонувати, що Т-клітини більшості цих волонтерів не впізнавали імунодомінантного повтору. Проте один волонтер у цьому дослідженні не розвивав паразитемію. 95293 6 WO 93/10152 та WO 98/05355 описують вакцини, що походять від CS білка P falciparum та демонструють деякий прогрес у вакцинації проти P. falciparum при використанні описаного підходу, див. також Неррnеr та ін. 2005, Vaccine 23, 224350. CS білок у P. falciparum має центральну повторювану ділянку, що є консервативною. На противагу до цього є відомими принаймні дві форми (позначаються як VK210 або тип І та VK247 або тип II) CS білка для P. vivax. Це робить більш складною ідентифікацію конструкції CS білка з усіма бажаними властивостями, такими, як імуногенність, що забезпечує загальний захист проти P. vivax, незважаючи на специфічний тип CS білка, оскільки антитіла, спрямовані проти центральної повторюваної ділянки типу І, з необхідністю не впізнають епітопи на відповідній ділянці типу II та навпаки. Рекомбінантний CS білок P. vivax було експресовано та проаналізовано як вакцину у 1980-1990 роках з обмеженим успіхом (Collins та ін., 1989. Am. J. Тrор. Med. Hyg. 40, 455-64). Була зроблена деяка робота з розробки вакцини на основі поліантигенних пептидів (МАР) при використанні одного або більше епітопів, які є такими, що перехресно зв'язуються (Nardelli та Tarn, 1995, Pharm. Biotechnol. 6, 803-19). Даний винахід забезпечує антигенні частинки для застосування в малярійних вакцинах, які, як передбачається, генерують гуморальну відповідь, а також клітинну імунну відповідь. Антигенні частинки, як передбачається, індукують утворення антитіл проти CS білка P falciparum та P. vivax типу І та типу II. Антиген може також індукувати Тхелперні клітини, наприклад, Th1 та/або Тh2 клітини. Згідно з цим даний винахід забезпечує імуногенні білкові частинки, що включають наступні мономери: a. злитий білок, що включає послідовності, які походять від CS білка P. vivax та S антигену гепатиту В (CSV-S), та b. злитий білок, що включає послідовності, які походять від CS білка Р. falciparum та S антигену гепатиту В (RTS), та необов'язково c. S антиген, який походить від вірусу гепатиту В. Список послідовностей SEQ. ID. No. 1 Ділянка І N-термінального кінця SEQ. ID. No. 2 Ділянка Il С-термінального кінця SEQ. ID. No. 3-9 Різноманітні повторювані одиниці типу I CS білка SEQ. ID. No. 10 Основна повторювана одиниця типу Il CS білка SEQ. ID. No. 11 Додаткові амінокислоти, виявлені в азіатських штамах SEQ. ID. No. 12 Нуклеотидна послідовність гібридного білка CSV (оптимізована для експресії в Е. соlі) SEQ. ID. No. 13 Амінокислотна послідовність гібридного білка CSV SEQ. ID. No. 14 Мінорна повторювана одиниця типу Il CS білка SEQ. ID. No. 15 Нуклеотидна послідовність 7 для гібридного білка CSV (оптимізована для експресії у дріжджах) SEQ. ID. No. 16 Нуклеотидна послідовність для гібридного злитого білка CSV-S SEQ. ID. No. 17 Амінокислотна послідовність для гібридного злитого білка CSV-S SEQ. ID No. 18 Нуклеотидна послідовність для RTS експресійної касети та спрогнозований RTS1S білок Фігури: Фіг.1 Плазмідна карта для pRIT15546 являє собою дріжджовий епісомальний вектор Фіг.2 Плазмідна карта плазміди pGF1-S2, одержаної GSK1 що використовується при «злитті» бажаного антигену з S антигеном гепатиту В. Клонування гетерологічних послідовностей ДНК між Smal сайтами (після вирізання Smal фрагменту ДНК розміром 12 п.о.) утворює злиття у рамці з геном S Фіг.3 Плазмідна карта pRIT15582. Перетравлювання за допомогою Xhol вивільняє фрагмент лінійної ДНК розміром 8,5 кб, що несе CSV-S експресійну касету плюс LEU2 селективний маркер, що використовується для вбудовування у дріжджову хромосому Фіг.4 Рестрикційна карта лінійного Xhol фрагменту, що використовується для інтеграції CSV-S касети Фіг.5 Вестерн-блотінг аналіз рекомбінантних білків, що експресуються у штамі Y1835. Секція А: ВБ, виявлений за допомогою анти-S антитіла Завантажені зразки (100 мкг загального білка/комірка): 1: Y1631 (штам, що продукує RTS,S, для порівняння) 2: Y1835 3: Y1835 4: Y1834 Секція В: ВБ, виявлений за допомогою антиSV антитіла Завантажені зразки (100 мкг загального білка/комірка): 1: Y1631 (штам, що продукує RTS,S, для порівняння) 2: Y1295 3: Y1835 4: Y1834 5: nr (інша конструкція CSVS) 6: nr (інша конструкція - тільки S-антиген) Фіг. 6 Електронна мікрографія CSV-S,S змішаних частинок, що продукуються штамом Y1835 CSV-S,S частинки очищали із розчинних екстрактів клітин (на основі процесу очистки RTS,S) та піддавали електронномікроскопічному аналізу. Частинки піддавали візуалізації після негативного забарвлювання за допомогою вольфрамової кислоти. Шкала є еквівалентною 100 нм. Фіг. 7 Вестерн-блотінг для рекомбінантних білків, що експресуються у штамі Y1845. ВБ виявлений за допомогою анти-S антитіла Кількість завантаженого загального білка приведена у дужках 1: Y1835 (100 мкг) 2: Y1631 (100 мкг - штам, що продукує RTS,S, 95293 8 для порівняння) 3: Y1845 (100 мкг) 4: Y1845 (50 мкг) 5: Y1845 (25 мкг) Фіг.8 CsCI аналіз густини вільного від клітин екстракту, одержаного зі штаму Y1845 Таким чином, злитий білок CSV-S, що використовується у винаході, включає частину, що має походження від CS білка P. vivax (CSV). Цей CSV антиген може бути нативним білком, таким, як виявлено у типі I CS білків P. vivax та/або таким, як виявлено у білках типу Il P. vivax. Альтернативно, CSV білок може являти собою гібридний білок або химерний білок, що включає елементи із вказаних CS білків типу І та II. Коли останній є злитим із S антигеном, такий буде називатися в даній заявці гібридним злитим білком. CSV-S використовується в даній заявці як генеричний термін для охоплення злитих білків, що включають послідовність/фрагмент з CS білка P. vivax та послідовність/фрагмент з S-антигену гепатиту В. RTS використовується в даній заявці як генеричний термін для охоплення злитих білків, що включають послідовність/фрагмент з CS білка P. falciparum та послідовність/фрагмент з S-антигену гепатиту В. Гібридні/химерні білки будуть у загальному випадку включати: принаймні одну повторювану одиницю, що походить від центрального повторюваного сегменту білка типу I1 що оточує спорозоїт P. vivax, та принаймні одну повторювану одиницю, що походить від центрального повторюваного сегменту білка типу II, що оточує спорозоїт P. vivax. В загальному випадку гібридний білок буде також включати N-термінальний фрагмент з CS білка Plasmodium, такого, як P. vivax, наприклад, фрагмент, що включає ділянку І, таку, як амінокислотна послідовність, представлена в SEQ ID No. 1. Звичайно гібридний білок буде містити Стермінальний фрагмент з CS білка Plasmodium, такого, як P. vivax, наприклад, фрагмент, що включає ділянку II, таку, як амінокислотна послідовність, представлена в SEQ ID No. 2. При відсутності бажання бути зв'язаним з теорією, передбачається, що N- та C-термінальні фрагменти включають декілька епітопів T- та Вклітин. Будь-який прийнятний штам P. vivax може використовуватися у винаході, включаючи: штами Latina, America (тобто SaI 1, Belem), Korean, China, Thailand, Indonesia, India та Vietnam. Конструкція, що представлена у SEQ ID No 13, базується на корейському штамі (зокрема, на південнокорейському штамі). P. vivax з CS білками типу І є більш переважним, ніж P. vivax з CS білками типу II. Таким чином, в одному аспекті винаходу використовується CS білок з типу І. В альтернативному аспекті винаходу забезпечується гібридний білок, що включає повторювану одиницю з типу І та повторювану одиницю з типу II, наприклад, де у гібрид включається більше повторюваних одиниць типу І, ніж повторюваних одиниць типу II. 9 Зокрема, гібридний білок згідно з винаходом може включати від 1 до 15 повторюваних одиниць, наприклад, 9 повторюваних одиниць типу І. Приклади прийнятних повторюваних одиниць типу I CS білків представлені у SEQ ID No. 3-9. В одному втіленні винаходу забезпечується гібрид із сумішшю різних повторюваних одиниць типу І, таких, як одна з тих, що приведені у SEQ ID No. 3-9. Одна або більше повторюваних одиниць можуть бути подвоєні у гібриді, наприклад, дві повторювані одиниці SEQ ID No. 3 та/або 4 можуть бути введені у конструкцію. a) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 3. b) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 4, необов'язково у комбінації з одиницями, як описано у пункті а) безпосередньо вище. c) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 5, необов'язково у комбінації з одиницями, як описано у пункті а) або b) безпосередньо вище. d) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 6, необов'язково у комбінації з однією або більше одиницями, як описано у пунктах а)-с) безпосередньо вище. f) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 7, необов'язково у комбінації з однією або більше одиницями, як описано у пункті а)-d) безпосередньо вище. g) В одному аспекті CS білок включає одиницю послідовності SEQ ID No. 8, необов'язково у комбінації з однією або більше одиницями, як описано у пункті а)-f) безпосередньо вище. h) В одному аспекті CS білок включає одиниці послідовності SEQ ID No. 9, необов'язково у комбінації з однією або більше одиницями, як описано у пункті а)-g) безпосередньо вище. Приклади прийнятного компоненту повторюваних одиниць з CS білків типу Il приведені у послідовностях SEQ ID No. 10 та 14, такій, як 10. В одному аспекті винаходу забезпечується гібридний білок з 5 або менше повторюваними одиницями, які походять від типу II, такими, як одна повторювана одиниця, наприклад, як представлено у SEQ ID No. 10. Гібрид може також включати інсерцію розміром 12 амінокислот, що виявлена на кінці повторюваної ділянки, яку знайдено у деяких азіатських штамах P. vivax, наприклад, як показано у SEQ ID No. 11. В одному втіленні гібридний білок включає приблизно 257 амінокислот, що походять від CS білка P. vivax. Антигенний компонент згідно з винаходом, що походить від CSV, в загальному випадку піддають злиттю з амінотермінальним кінцем S білка. При цьому передбачається, що присутність поверхневого антигену гепатиту В стимулює імуногенність CS частини білка, сприяє стабільності та/або поліпшує відтворюваність виробництва білка. В одному втіленні гібридний злитий білок включає приблизно 494 амінокислот, при цьому 95293 10 приблизно 257 з них походять від CS білка P. vivax. Гібридний злитий білок може також включати додаткові антигени, які походять від P. falciparum та/або P. vivax, наприклад, антигени, вибрані з DBP, PvTRAP, PvMSP2, PvMSP4, PvMSP5, PvMSP6, PvMSP7, PvMSP8, PvMSP9, PvAMA1 та RBP або їх фрагментів. В одному втіленні гібридний злитий білок (CSV-S) має амінокислотну послідовність, представлену в SEQ ID No. 17. У послідовності амінокислоти 6-262 є такими, що походять від CSV, а 269-494 є такими, що походять від S. Амінокислоти, що залишилися, вбудовують за допомогою генетичної конструкції (яка, зокрема, може відрізнятися, якщо це є прийнятним). Властивості CSV-S злитого білка послідовності SEQ ID No. 17 приведені у Таблицях, що представлені нижче. Аналіз Молекулярна вага Довжина 1 мкг = Коефіцієнт молярної екстинкції 1 А(280)= Ізоелектрична точка Заряд при рН 7 Цільний білок м.в. 51794,75 494 19,307 пмолів 90780+/-5% 0,57 мг/мл 7,33 1,05 Аналіз складу цільного білка % від ваги % частоти повторюваності 106 26,35 21,46 38 39 8,82 10,68 7,69 7,89 134 28,15 27,13 167 34,68 33,81 52 18 24 14 17 64 4 17 20 42 8 32 40 21 19 30 26 25 14 7 0 0 7,14 3,58 5,33 3,49 4,83 7,05 1,06 3,71 4,95 9,18 2,03 7,05 7,50 5,20 5,73 5,04 5,08 4,78 5,03 2,21 0,00 0,00 10,53 3,64 4,86 2,83 3,44 12,96 0,81 3,44 4,05 8,50 1,62 6,48 8,10 4,25 3,85 6,07 5,26 5,06 2,83 1,42 0,00 0,00 Амінокислота(и) Кількість Заряджені (RKHYCDE) Кислі (DE) Основні (KR) Полярні (NCQSTY) Гідрофобні (AILFWV) A AIa C Cys D Asp E GIu F Phe G GIy H His I lІe K Lys L Leu M Met N Asn P Pro Q GIn R Arg S Ser T Thr V VaI W Trp Y Туr В Asx Z GIx 11 Х Ххх Теr 0 1 95293 0,00 0,00 0,00 0,20 Нуклеотидна послідовність для білка послідовності SEQ ID No. 17 приведена у SEQ ID No. 16. Компонент білкових частинок згідно з винаходом, що має назву RTS (тобто таких, що походять від P. falciparum), може бути одержаний так, як описано у WO 93/10152, що включає опис RTS* (зі штаму NF54/3D7 P. falciparum). В одному або більше втіленнях винаходу антиген, що походить від P. falciparum та використовується у злитому білку, може суттєво являти собою цілий білок CS. В одному втіленні винаходу використовується S-антиген повної довжини. В іншому втіленні використовується фрагмент вказаного S-антигену. Злитий білок CSV-S може, наприклад, бути одержаним при використанні плазміди pGF1-S2 (див. Фіг.2 та приклади для додаткових деталей), який у разі вбудовування прийнятної послідовності, яка відповідає CSV, у сайт клонування Saml може за прийнятних умов продукувати злитий білок CSV-S. Послідовності ДНК, що кодують білки згідно з даним винаходом, є в одному втіленні фланкованими елементами контролю транскрипції, бажано такими, що походять від генів дріжджів, та є вбудованими в експресійний вектор. Сконструйована експресійна касета для гібридних білків згідно з винаходом може, наприклад, характеризуватися наступними ознаками: • Промоторна послідовність, яка походить, наприклад, від гену TDH3 S.cerevisiae. • Послідовності, що кодують прийнятний злитий білок. • Послідовність термінації транскрипції, що міститься у межах послідовності, яка походить, наприклад, від гену ARG3 S. cerevisiae. Прикладом специфічного промотора є промотор з гену TDH3 S.cerevisiae, Musti та ін. Винахід також охоплює вектори, які використовуються при одержанні гібридного злитого білка. Для вбудовування послідовності, яка кодує гібридний злитий білок у прийнятного хазяїна, потім може використовуватися прийнятна плазміда. Приклад прийнятної плазміди являє собою pRIT15546, 2 мкм вектор для розміщення прийнятної експресійної касети, див. Фіг.1 та Приклади для додаткових деталей. Плазміда буде у загальному випадку містити вбудований маркер для сприяння селекції, наприклад, ген, що кодує стійкість до антибіотиків, або LEU2, або HIS ауксотрофність. В загальному випадку хазяїн буде мати експресійну касету для кожного злитого білка у частинці та може також мати одну або більше експресійних касет для S антигену, інтегрованого у геном. Винахід також відноситься до хазяйської клітини, трансформованої вектором згідно з винаходом. Хазяйські клітини можуть бути прокаріотичними або еукаріотичними клітинами, але переважно, являють собою дріжджі, наприклад, Saccharomyces (наприклад, Saccharomyces 12 cerevisiae, такий, як DC5 у ATCC базі даних (депозитний номер 20820), під назвою RIT DC5 сіr(о). Депозитор: Smith Kline-RIT) та дріжджі, відмінні від Saccharomyces. Такі включають Schizosaccharomyces (наприклад, Schizosaccharomyces pombe) Kluyveromyces (наприклад, Kluyveromyces lactis), Pichia (наприклад, Pichia pastoris), Hansenula (наприклад, Hansenula polymorpha), Yarrowia (наприклад, Yarrowia lipolytica) та Schwanniomyces (наприклад, Schwanniomyces occidentalis). В одному аспекті винахід відноситься до рекомбінантного дріжджового штаму Y1834 (та до його застосування), що використовується для експресії злитого білка, див. Приклади для його одержання. В іншому втіленні винаходу забезпечується рекомбінантний дріжджовий штам Y1835 або Y1845 (та до їх застосування) для експресії злитого білка згідно з винаходом, див. Приклади для додаткових деталей. Нуклеотидні послідовності або їх частина (така, як частина, що кодує CS/гібридний білок, але необов'язково не та частина, що кодує білок S)1 використовувані в даній заявці, можуть бути оптимізовані за кодонами для експресії у хазяїні, такому, як дріжджі. Винахід також охоплює хазяїна, що включає полінуклеотид, такий, як ДНК, що кодує два або більше компонентів частинки, яка використовується у даному винаході. В одному втіленні хазяйська клітина включає експресійну касету для злитого білка, що походить від P. vivax, та експресійну касету для злитого білка, що походить від P. falciparum. В деяких хазяйських організмах, таких, як клітини дріжджів, експресований злитий білок (що включає S антиген) піддається спонтанній зборці у білкову структуру/частинку, яка складається з багаточисленних мономерів вказаних злитих білків. Коли дріжджі експресують два різні злиті білки, то такі, як передбачається, сумісно піддаються зборці з утворенням частинок. Коли вибраний реципієнтний дріжджовий штам вже несе у своєму геномі декілька інтегрованих копій експресійних касет S антигену гепатиту В, то частинки, які піддаються зборці, включають мономери незлитого S антигену. Такі частинки можуть також називатися вірусоподібними частинками (VLP). Ці частинки можуть також бути описані як мультимерні ліпопротеїнові частинки. Альтернативно, ці частинки можуть бути одержані при використанні ряду способів, наприклад, шляхом злиття кожного антигену, що походить від Plasmodium, з іншим партнером злиття (наприклад, антигенами вірусу гепатиту В або вірусним структурним білком) та експресії конструкції у прийнятному хазяїні, такому, як дріжджі або бактерії. Таким чином, забезпечується імуногенна білкова частинка, що включає наступні мономери: a. злитий білок, що включає послідовності, які походять від CS білка P.vivax, та b. злитий білок, що включає послідовності, які походять від CS білка Р. falciparum. У додатковому аспекті винахід забезпечує 13 злитий білок, що включає: a) послідовність, яка походить від CS білка P. vivax (така, як послідовність з повторюваної ділянки типу І та/або типу II) b) послідовність, яка походить від CS білка P. falciparum (така, як послідовність його повторюваної ділянки), та c) послідовність S-антигену гепатиту В. Таким чином, винахід охоплює білкову частинку, що включає злитий білок, який походить від CS білка P.vivax, та злитий білок, що походить від CS білка P. falciparum, де антиген, злитий з антигеном Plasomdium, є вибраним для індукції утворення/зборки ліпопротеїнових частинок, коли вказані злиті білки експресуються у прийнятному хазяїні. Таким чином, винахід забезпечує VLP, що включає CSV-S та RTS одиниці. В одному аспекті винахід забезпечує частинку, що суттєво складається з CSV-S та RTS одиниць. В альтернативному аспекті одержані частинки включають або складаються суттєво з CSV-S, RTS та S одиниць. Можуть використовуватися різні підходи до створення дріжджів для одержання вказаних частинок, наприклад, експресійна касета для злитого білка може бути вбудованою у геном дріжджів, що вже містить експресійну касету для будь-яких необхідних злитих білків та/або S антигену. При цьому спеціаліст в даній області техніки, що працює в цій галузі, є здатним одержати прийнятного хазяїна для приготування частинок згідно з винаходом. Таким чином, в одному аспекті винахід забезпечує прийнятного хазяїна, такого, як дріжджі, що включає ДНК, яка кодує CSV-S, RTS, та необов'язково S. В альтернативному аспекті прийнятний хазяїн являє собою одну або більше плазміду, здатну до експресії білка у поєднанні з, наприклад, дріжджами. Не маючи бажання пов'язуватися з будь-якою теорією, передбачається, що поверхнево-активні сполуки, які використовуються для вивільнення білка з клітини, можуть також сприяти стабілізації ліпопротеїнових частинок. Існує гіпотеза, що ліпопротеїнові частинки винаходу можуть робити свій внесок у додаткову стимуляцію in vivo імунної відповіді на антигенний(і) білок(білки). В одному аспекті винахід забезпечує дефектний за реплікацією вірусний вектор, що кодує один або більше CS білків, наприклад, такого(их), що кодує(кодують) CS білок(білки), які є включеними у частинки згідно з винаходом. Прийнятні вірусні вектори можуть являти собою такі, що походять аденовірусних векторів, аденоасоційованих вірусних векторів (AAV), векторів на основі вірусу кору, лентівірусів, альфавірусів, бакуловірусів, вірусу простого герпесу, та поксвірусів, таких, як вірус коров'ячої віспи, вірус віспи птахів, вірус віспи голубів, вірус віспи канарок, віспи свиней та віспи овецьх/кіз. Методика одержання аденовірусних векторів, що кодують малярійний антиген, є, наприклад, описаною у WO 2004/055187. Білок, що кодується вектором, може, наприклад, бути модифікованим для запобігання глікозилювання білка під час експресії, наприклад, де 95293 14 які серинові залишки можуть бути замінені на аланінові залишки для зниження глікозилювання. Вірусні вектори, які використовуються у винаході, можуть бути рекомбінантними. Аденовірус Аденовірусні вектори даного винаходу включають один або більше гетерологічних полінуклеотидів (ДНК), які кодують один або більше імуногенних поліпептидів. Аденовірусні вектори для застосування у даному винаході можуть бути такими, що походять від спектру хазяїв-ссавців. Аденовіруси (які в даній заявці називаються "Ad" або "Adv") мають характерну морфологію з ікосаедричним капсидом, який складається з трьох основних білків, гексону (II), пентонової основи (III) та шишкоподібних волокон (IV), разом з рядом інших мінорних білків, VI, VIII, IX, IІІa та IVa2 (Russell W.C. 2000, Gen Viriol, 81:2573-2604). Вірусний геном являє собою лінійну, дволанцюгову ДНК з термінальним білком, який є приєднаним ковалентно до 5'-термінального кінця, що має інвертовані кінцеві повтори (ITR). Вірусна ДНК є безпосередньо асоційованою з високо основним білком VII та маленьким пептидом, що називається мю. Інший білок, V, є спакованим з цим комплексом ДНК-білок та забезпечує структурний зв'язок з капсидом через білок VI. Вірус також містить протеазу, яка кодується вірусом, що є необхідною для процесингу деяких структурних білків з одержанням зрілого інфекційного вірусу. Було ізольовано більше 100 різних серотипів аденовірусу, які інфікують різні види ссавців, 51 з них мають людське походження. Таким чином, один або більше аденовірусних векторів можуть походити від людського аденовірусу. Приклади таких, аденовірусів, що походять від людських, являють собою Ad 1, Ad2, Ad4, Ad5, Ad6, Ad11, Ad24, Ad34, Ad35, Ad50/51, зокрема, Ad5, Ad11 та Ad35. Людські серотипи були класифіковані на шість підвидів (A-F) на основі ряду біологічних, хімічних, імунологічних та структурних критеріїв. Незважаючи на те, що вектори на основі Ad5 екстенсивно використовуються у ряді експериментів з генної терапії, існують обмеження для застосування аденовірусних векторів Ad5 та іншої групи C завдяки попередньо існуючому імунітету у загальній популяції з причини природної інфекції. Члени Ad5 та іншої групи C мають тенденцію бути серед найбільш домінуючих серотипів. Імунітет до існуючих векторів може розвиватися як результат піддання впливу вектору під час лікування. Ці типи попередньо існуючого імунітету або імунітету, що розвинувся, до векторів на основі домінуючих серотипів можуть обмежувати ефективність генної терапії або зусилля по вакцинації. Альтернативні аденовірусні серотипи, таким чином, являють собою дуже важливі цілі, які переслідуються при розробці систем для доставки генів, що є здатними, уникати імунної відповіді хазяїна. Одна така область альтернативних серотипів являє собою такі, що походять від приматів, відмінних від людини, зокрема, від аденовірусів шимпанзе. Див. патент США №6,083,716, який описує геном двох аденовірусів шимпанзе. 15 Було показано, що аденовірусні вектори шимпанзе ("Pan" або "С") індукують сильні імунні відповіді на трансгенні продукти так ефективно, як й людські аденовірусні вектори (Fitzgerald та ін. J. Immunol. 170:1416). Аденовіруси приматів, відмінних від людини, можуть бути ізольовані з мезентеричних лімфатичних вузлів шимпанзе. Аденовіруси шимпанзе є в достатній мірі подібними до людського аденовірусу підтипу С, що дозволяє проводити реплікацію Е1 делетованого вірусу у клітинах HEK 293. Незважаючи на це, аденовіруси шимпанзе є філогенетично відмінними від більш загальних людських серотипів (Ad2 та Ad5). Pan6 є найменш близькоспорідненим та серологічно відрізняється від Pan5, 7 та 9. Таким чином, один або більше аденовірусних векторів можуть походити від аденовірусів приматів, відмінних від людини, наприклад, від аденовірусу шимпанзе, такого, як один із серотипів Pan5, Pan6, Pan7 та Pan9. Аденовірусні вектори можуть також походити більше, ніж від одного вірусного серотипу, та кожний серотип може походити з того самого або іншого джерела. Наприклад, вони можуть походити від більше, ніж з одного, серотипу людини та/або більше, ніж з одного, серотипу примату, відмінного від людини. Способи конструювання химерних аденовірусних векторів є розкритими у WO2005/001103. Існують деякі обмеження за розмірами, асоційовані з інсерціею гетерологічної ДНК в аденовірус. Людські аденовіруси мають здатність упаковувати до 105% довжини геному дикого типу (Bert та ін. 1993, J Virol 67 (10), 5911-21). Більш низька границя пакування для людських аденовірусів була показана як така, що становить 75% довжину геному дикого типу (Parks та ін. 1995, J Virol 71(4), 3293-8). Один приклад аденовірусів, корисних згідно з даним винаходом, являє собою аденовіруси, які відрізняються від домінуючих природних серотипів у людській популяції, такі, як Ad2 та Ad5. Це надає можливості уникати індукції потужних імунних відповідей проти вектора, які обмежує ефективність подальших введень того самого серотипу шляхом блокування поглинання вектора за допомогою нейтралізуючого антитіла та токсичності, що виявляє свій вплив. Таким чином, аденовірус може являти собою аденовірус, який не є домінуючим існуючим у природі серотипом людського вірусу. Аденовіруси, ізольовані з тварин, мають імунологічно відмінний капсид, гексон, пентон, волокнисті компоненти, але філогенетично є близькоспорідненими. Зокрема, вірус може являти собою не-людський аденовірус, такий, як мавпячий аденовірус та, зокрема, аденовірус шимпанзе, такий, як Pan5, 6, 7 або 9. Приклади таких штамів є описаними у WO 03/000283 та є доступними з Американської колекції типових культур, 10801 University Boulevard, Manassas, Virginia 20110-2209, та інших джерел. Бажані штами аденовірусів шимпанзе являють собою Pan5 [ATCC VR-591], Pan6 [ATCC VR-592] та Pan7 [ATCC VR-593]. 95293 16 Застосування аденовірусів шимпанзе, як передбачається, є бажаними у порівнянні із застосуванням серотипів людських аденовірусів, оскільки є відсутнім попередньо існуючий імунітет, зокрема, є відсутніми попередньо існуючі антитіла, до аденовірусів у цільовій популяції. Перехресна реакція аденовірусів шимпанзе з попередньо існуючими антитілами є присутньою тільки у 2% цільової популяції у порівнянні з 35% у випадку деяких кандидатних людських аденовірусів. Аденовіруси шимпанзе відрізняються від більш загальних людських підтипів Ad2 та Ad5, але є більш спорідненими з людським Ad4 підгрупи Е, який не є домінуючим підтипом. Pan 6 є менш близькоспорідненим з Pan5, 7 та 9. Аденовірус згідно з винаходом може бути дефектним за реплікацією. Це означає, що він має знижену здатність до реплікації в некомплементарних клітинах у порівнянні з вірусом дикого типу. Це може бути викликане мутацією вірусу, наприклад, делецією гену, що є втягнений у реплікацію, наприклад, делецією Е1а, E1b, Е3 або Е4 гену. Аденовірусні вектори згідно з даним винаходом можуть походити від аденовірусу, що є дефектним за реплікацією, включаючи функціональну Е1 делецію. Таким чином, аденовірусні вектори згідно з винаходом можуть бути дефектними за реплікацією завдяки відсутності здатності експресувати аденовірусні Е1 та Е1b, тобто є функціонально делегованими у Е1а та Е1b. Рекомбінантні аденовіруси можуть також нести функціональні делеції в інших генах [див. WO 03/000283], наприклад, делеції у генах Е3 або Е4 (або в їх частинах, таких, як частина Е3). Аденовірусний затриманий ранній ген Е3 може бути елімінований з аденовірусної послідовності, яка утворює частину рекомбінантного аденовірусу. Функція Е3 не є необхідною для утворення рекомбінантної аденовірусної частинки. Таким чином, не є необхідним замінювати функцію цього генного продукту для того, щоб спакувати рекомбінантний аденовірус, корисний у даному винаході. В одному окремому втіленні рекомбінантні аденовіруси мають функціонально делеговані гени Е1 та Е3. Конструювання таких векторів є описаним у Roy та ін., Human Gene Therapy 15:519-530, 2004. В одному аспекті аденовірус має делетовані Е1 та Е4 та делетовану частину Е3. Рекомбінантні аденовіруси можуть також бути сконструйовані як такі, що мають функціональну делецію гену Е4, незважаючи на це, може бути бажаним зберігти ORF6 функцію Е4. Аденовірусні вектори згідно з винаходом можуть також містити делецію затриманого раннього гену Е2а. Можуть також бути зроблені делеції у будь-якому з пізніх генів L1-L5 аденовірусного генома. Подібно до цього, делеції у проміжних генах IX та IV також можуть бути корисними. Можуть бути зроблені інші делеції в інших структурних або неструктурних генах аденовірусів. Зазначені вище делеції можуть використовуватися індивідуально, тобто послідовність аденовірусу для застосування у даному винаході може містити тільки делеції Е1. Альтернативно, делеції цільних генів або їх частини, що ефективно руйнують їх 17 біологічну активність, можуть також використовуватися у будь-якій комбінації. Наприклад, в одному з векторів аденовірусні послідовності можуть мати делеції гену Е1 та гену Е4, або генів Е1, Е2а та Е3, або генів Е1 та Е3 (такі, як функціональні делеції Е1а та Е1b, та делецію принаймні частини Е3), або генів Е1, Е2а та Е4, з делецією або без делеції Е3, тощо. Такі делеції можуть бути повними або частковими делеціями цих генів та можуть використовуватися у комбінації з іншими мутаціями, такі, як чутливі до температури мутації, для досягнення бажаного результату. Аденовірусні вектори можуть бути одержані на будь-якій прийнятній клітинній лінії, в якій вірус є здатним до реплікації. Зокрема, можуть використовуватися комплементарні клітинні лінії, які забезпечують фактори, відсутні у вірусному векторі, що приводить до їх порушених характеристик реплікації (таких, як Е1 та/або Е4). Без обмеження такі клітинні лінії можуть являти собою HeLa [ATCC депозитний номер CCL 2], А549 [ATCC депозитний номер CCL 185], HEK 293, KB [CCL 17], Detroit [наприклад, Detroit 510, CCL 72] та WI-38 [CCL 75] клітини, серед інших. Усі ці клітинні лінії є доступними з Американської колекції типових культур, 10801 University Boulevard, Manassas, Virginia 20110-2209. Інші прийнятні вихідні клітинні лінії можуть бути одержані з інших джерел, таких, як PER.C6© клітини, як представлено клітинами, задепонованими під ECACC номером 96022940 в Європейській колекції культур клітин тваринних (ECACC) при Центрі прикладної мікробіології та дослідження (CAMR, UK) або Her 96 клітини (Crucell). Винахід охоплює застосування відомих клітинних ліній для одержання вірусного вектора, що кодує білок згідно з даним винаходом. Полінуклеотидні послідовності, які кодують імуногенні CS поліпептиди, можуть бути оптимізовані за кодонами для клітин ссавців. Така оптимізація за кодонами є детально описаною у WO 05/025614. Винахід також охоплює композиції, що включають частинки згідно з винаходом та вірусний вектор, що містить малярійний антиген, зокрема, малярійний антиген, що має спільне походження із вказаними частинками, та необов'язково ад'ювант. У контексті цього опису «наповнювач» відноситься до компоненту у фармацевтичній композиції, що не виявляє терапевтичного ефекту сам по собі. Розріджувач потрапляє у визначення наповнювача. Термін «імуногенний» у контексті даного винаходу призначений для віднесення до здатності викликати імунну відповідь. Ця відповідь може, наприклад, бути тоді, коли ліпопротеїнові частинки вводяться у складі прийнятної композиції, яка може включати/вимагати присутності прийнятного ад'юванта. Бустер-ін'єкція, що включає дозу, яка є такою самою або меншою за вихідну дозу, може бути необхідною для одержання бажаної імуногенної відповіді. Композиція/фармацевтичний склад згідно з винаходом може також включати суміш одного або більше додаткових антигенів, таких, як ті, що походять від P. falciparum та/або P. 95293 18 vivax, наприклад, де антигени є вибраними з DBP, PvTRAP, PvMSP2, PvMSP4, PvMSP5, PvMSP6, PvMSP7, PvMSP8, PvMSP9, PvAMA1 та RBP або їх фрагментів. Інший приклад антигенів, що походять від P. falciparum, включає PfEMP-1, Pfs 16 антиген, MSP1, MSP-3, LSA-1, LSA-3, АМА-1 та TRAP. Інші антигени Plasmodium включають EBA, GLURP, RAPI, RAP2, Sequestrin, Pf332, STARP, SALSA, PfEXPI, Pfs25, Pfs28, PFS27/25, Pfs48/45, Pfs230 P. falciparum та їх аналогів з інших Plasmodium spp. Композиція/фармацевтичний склад згідно з винаходом може також включати частинки RTS,S (як описано у WO 93/10152) у суміші із частинками згідно з винаходом. В одному втіленні конструкція вірусного вектора є такою, як описано у WO 2004/055187. У вакцині згідно з винаходом водний розчин частинок може використовуватися безпосередньо. Альтернативно, білок, який піддавався або не піддавався попередній ліофілізації, може змішуватися або піддаватися абсорбції з будь-яким з відомих ад'ювантів, які включають, але без обмеження, алюм, мураміловий дипептид, сапоніни, такі, як QuiІ А. Особливими ад'ювантами є ті, що вибрані з групи солей металів, емульсій масло-у-воді, агоністів Toll-подібних рецепторів, (зокрема, агоніст Toll-подібного рецептора 2, агоніст Toll-подібного рецептора 3, агоніст Toll-подібного рецептора 4, агоніст Toll-подібного рецептора 7, агоніст Tollподібного рецептора 8 та агоніст ToII-подібного рецептора 9), сапонінів та їх комбінацій, за умови, що солі металів використовуються тільки у комбінації з іншим ад'ювантом, але не окремо, якщо вони не рецептовані таким чином, що не більше, ніж приблизно 60% антигенів піддається адсорбції на солі металу. Зокрема, не більше, ніж приблизно 50%, наприклад, 40% антигенів піддається адсорбції на солі металу, та в одному втіленні не більше, ніж приблизно 30% антигенів піддається адсорбції на солі металу. Рівень антитіла, адсорбованого на солі металу, може визначатися за допомогою методик, що є добре відомими з рівня техніки. Рівень вільного антигену може бути підвищений, наприклад, шляхом рецептування композиції у присутності іонів фосфату, наприклад, забуференого фосфатом фізіологічного розчину, або шляхом підвищення співвідношення антигену до солі металу. В одному втіленні ад'ювант не включає солі металу як єдиного ад'юванту. В одному втіленні ад'ювант не включає солі металу. В одному втіленні ад'ювант являє собою ліганд Toll-подібного рецептора (TLR) 4, бажано агоніст, такий, як похідна ліпіду А, зокрема, монофосфорил ліпід А або більш бажано 3деацилований монофосфорил ліпід А (3D-MPL). 3D-MPL є доступним від GlaxoSmithKline Biologicals North America та, в першу чергу, поліпшує CD4+ Т-клітинні відповіді з IFN-g (TM) фенотипом. Він може бути одержаний згідно зі способами, розкритими в GB 2 220 211 А. Хімічно він являє собою суміш 3-деацилованого монофосфорил ліпіду А з 3, 4, 5 або 6 ацилованими ланцюгами. Бажано використовувати у композиціях згідно з 19 даним винаходом маленькі частинки 3D-MPL. Маленькі частинки 3D-MPL мають такий розмір частинок, що вони можуть стерильно фільтруватися через фільтр 0,22 мкм. Такі препарати є описаними у міжнародній патентній заявці WO 94/21292. Синтетичні похідні ліпіду А є відомими та, як передбачається, є TLR 4 агоністами. Такі включають, але без обмеження: OM 174 (2-дезокси-6-О-[2-дезокси-2-[(R)-3дедоканоїлокситетра-деканоїламіно]-4-oфocфoнo-β-D-глюкoпipaнoзил]-2-[(R)-3гiдpoкcитeтpaдeкaнoїлaмiнo]--Dглюкопіранозилдигідрофосфат), (WO 95/14026) OM 294 DP (3S,9R)-3-[(R)додеканоїлокситетра-деканоїламіно]-4-оксо-5-аза9(R)-[(R)-3-гідрокситетрадеканоїламіно]декан-1,10діол,1,10-біс(дигідроофосфат) (WO 99/64301 та WO 00/0462) OM 197 MP-Ac DP (3S,9R)-3-[(R)додеканоїлокситетра-деканоїламіно]-4-оксо-5-аза9-[(R)-3-гідрокситетрадеканоїламіно]декан-1,10діол,1-дигідроофосфат 10-(6-аміногексаноат) (WO 01/46127) Типово, коли використовується 3D-MPL, антиген та 3D-MPL доставляються з алюмом або представлені в емульсії масло-у-воді, або у вигляді емульсії декількох масел-у-воді. Введення 3D-MPL є бажаним, оскільки він являє собою стимулятор відповідей ефекторних Т-клітин. Інші TLR4 ліганди, які можуть використовуватися, являють собою алкілглюкозамінідфосфати (AGP), такі, як ті, що розкриті в WO 9850399 або US 6303347 (способи для одержання AGP є також розкритими) або фармацевтично прийнятні солі AGP, як розкрито в US 6764840. Деякі AGP є TLR4 агоністами, а деякі є TLR4 антагоністами. Обидва, як передбачається, є корисними як ад'юванти. Інший бажаний імуностимулятор для застосування в даному винаході являє собою QuiІ А та його похідні. QuiІ А є препаратом сапоніну, ізольованим з південноамериканського дерева Quilaja Saponaria Molina, що був вперше описаний як такий, що володіє ад'ювантною активністю Dalsgaard та ін. у 1974 ("Сапонінові ад'юванти", Archiv. fur die gesamte Virusforschung, том. 44, Springer Verlag, Berlin, стор. 243-254). Очищені фрагменти QuiІ А були ізольовані шляхом ВЕРХ, що зберігає ад'ювантну активність без токсичності, асоційованої з Quil A (EP 0 362 278), наприклад, QS7 та QS21 (що є також відомими як QA7 та QA21). QS-21 є природним сапоніном, що одержаний з кори Quillaja saponaria Molina, він індукує CD8+ цитотоксичність T клітин (CTL), ThI клітин та переважну відповідь lgG2a антитіла, а також є переважним сапоніном у контексті даного винаходу. Були описані окремі композиції QS21, які, зокрема, додатково включають стерин (WО 96/33739). Співвідношення QS21:стерин буде типово мати порядок від 1:100 до 1:1 ваг./ваг. В загальному випадку є присутнім надлишок стерину, а співвідношення QS21:стерин є принаймні 1:2 ваг./ваг. Є типовим, коли для введення людині QS21 та стерин будуть присутніми у вакцині в інтервалі від приблизно 1 мкг до приблизно 100 мкг, наприклад, від приблизно 10 мкг до приблизно 50 95293 20 мкг на дозу. Ліпосоми у загальному випадку містять нейтральний ліпід, наприклад, фосфатидилхолін, який звичайно є некристалічним при кімнатній температурі, наприклад, фосфатидилхолін яєчного жовтка, діолеїл фосфатидилхолін або дилаурил фосфатидилхолін. Ліпосоми можуть також містити заряджений ліпід, який підвищує стабільність структури ліпосома-0821 для ліпосом, що складаються із насичених ліпідів. У цих випадках кількість зарядженого ліпіду часто становить 1-20% ваг./ваг., наприклад, 5-10%. Співвідношення стерину до фосфоліпіду складає 1-50% (мол./мол.), наприклад, 20-25%. Такі композиції можуть містити MPL (3деацилований монофосфорил ліпід А, що також є відомим як 3D-MPL). 3D-MPL є відомим з GB 2220211 (Ribi) як суміш 3 типів 3-деацилованого монофосфорил ліпіду А з 4, 5 або 6 ацилованими ланцюгами, що виробляється Ribi lmmunochem, Montana. Сапоніни, які утворюють частину згідно з даним винаходом, можуть бути відокремлені у формі міцел, змішаних міцел, (переважно, але не виключно з солями жовчних кислот) або можуть бути у формі ISCOM матриць (EP O 109 942 В1), ліпосоми або споріднені колоїдні структури, такі, як червоподібні або кільцеподібні мультимерні комплекси, або ліпідні/шаруваті структури та ламели, коли рецептуються з холестерином та ліпідом, або у формі емульсії масло-у-воді (наприклад, як у WO 95/17210). Сапоніни бажано можуть бути асоційовані із солями металів, такими, як гідроокис алюмінію або фосфат алюмінію (WO 98/15287). Звичайно сапоніни є представленими у формі ліпосоми, ISCOM або у формі емульсії масло-уводі. Імуностимуляторні олігонуклеотиди або будьякі інші агоністи Toll-подібного рецептора (TLR) 9 можуть також використовуватися. Бажані олігонуклеотиди для застосування в ад'ювантах або вакцинах згідно з даним винаходом являють собою CpG-вмісні олігонуклеотиди, бажано такі, що містять два або більше динуклеотидних CpG фрагментів, відокремлених принаймні трьома, більш бажано принаймні шістьома або більше нуклеотидами. CpG фрагмент являє собою цитозиновий нуклеотид, після якого йде гуаніновий нуклеотид. CpG олігонуклеотиди згідно з даним винаходом являють собою типово дезоксинуклеотиди. У бажаному втіленні інтернуклеотид в олігонуклеотиді являє собою фосфородитіоат, або більше бажано фосфородитіоатний зв'язок, хоча фосфодіестер та інші інтернуклеотидні зв'язки також передбачаються в об'ємі даного винаходу. Також в об'єм даного винаходу включаються олігонуклеотиди зі змішаними інтернуклеотидними зв'язками. Способи одержання фосфородитіоатних олігонуклеотидів або фосфородитіоату є описаними у US 5,666,153, US 5,278,302 та WO 95/26204. Приклади бажаних олігонуклеотидів мають наступні послідовності. Послідовністі бажано містять фосфородитіоатний модифікований інтернуклеотидний зв'язок. 21 Альтернативні CpG олігонуклеотиди можуть включати бажані послідовності, приведені вище, в яких вони мають несуттєві делеції або доповнення. CpG олігонуклеотиди, що використовуються в даному винаході, можуть бути синтезовані у відповідності із будь-якими способами, відомими в області техніки (наприклад, див. EP 468520). Так, ці олігонуклеотиди можуть бути синтезовані при використанні автоматичного синтезатора. Приклади TLR 2 агоніста включають пептидоглікан або ліпопротеїн. Імідазохіноліни, такі, як іміквімод та резиквімод є відомими TLR7 агоністами. Одноланцюгова PHK є також відомим TLR агоністом (TLR8 у людей та TLR7 мишей), де одноланцюгова PHK та полі IC (поліінозиноваполіцитидилова кислота - комерційний синтетичний міметик вірусної PHK) є прикладами TLR 3 агоністів. 3D-MPL є прикладом TLR4 агоніста, у той час, як CPG являє собою приклад TLR9 агоніста. Може бути включеним імуностимулятор, альтернативно або на доповнення. В одному втіленні цей імуностимулятор буде являти собою 3 деацилований монофосфорил ліпід A (3D-MPL). В одному аспекті ад'ювант включає 3D-MPL. В одному аспекті ад'ювант включає QS21. В одному аспекті ад'ювант включає CpG. В одному аспекті ад'ювант є рецептованим у вигляді емульсії масло-у-воді. В одному аспекті ад'ювант є рецептованим у вигляді ліпосоми. Ад'ювантні комбінації включають 3D-MPL та QS21 (EP 0 671 948 BІ), емульсії масло-у-воді, що включають 3D-MPL та QS21 (WO 95/17210, WO 98/56414), або 3D-MPL, рецептований з іншими носіями (EP 0 689 454 BІ). Інші бажані ад'ювантні системи включають комбінацію 3D-MPL, QS21 та a CpG олігонуклеотиду, як описано в US 6558670 та US 6544518. В одному втіленні даного винаходу забезпечується вакцина, що включає частинки, описані в даній заявці, у комбінації з 3D-MPL та носієм. Типово носій буде представляти собою емульсію масло-у-воді або алюм. Білкові частинки даного винаходу можуть також бути інкапсульовані у мікрочастинки, такі, як ліпосоми. Вакцинні препарати є в загальному випадку є описаними у джерелі «Нові тенденції та удосконалення у галузі вакцин», під ред. Voller та ін., University Park Press, Baltimore, Maryland, U.S.A., 1978. Інкапсуляція у ліпосомах є описаною, наприклад, у Fullerton, патент США №4,235,877. Кількість білкових частинок згідно з даним винаходом, що є присутньою у кожній вакцинній дозі, 95293 22 вибирають як кількість, яка індукує імунопротективну відповідь без значних, шкідливих побічних ефектів у типових вакцинах. Така кількість буде варіювати в залежності від специфічного імуногену, що використовується та від того, чи містить ця вакцина ад'ювант. В загальному випадку, передбачається, що кожна доза буде включати 1-1000 мкг білка, бажано 1-200 мкг, найбільш бажано 10100 мкг. Оптимальна кількість для конкретної вакцини може бути встановлена за допомогою стандартних досліджень, що втягують спостереження за титрами антитіла та інші відповіді у суб'єктів. Після цього суб'єкти будуть переважно одержувати бустерну імунізацію у період приблизно 4 тижні, а після цього повторюють бустерні імунізації настільки довго, скільки зберігається ризик інфекції. Імунна відповідь на білок згідно з цим винаходом поліпшується шляхом застосування ад'юванту та/або імуностимулятора. Використовувана кількість 3D MPL є в загальному випадку низькою, але в залежності від вакцинної композиції може складати 1-1000 мкг на дозу, наприклад, 1-500 мкг на дозу, та таку від 1 до 100 мкг на дозу. Кількість CpG або імуностимуляторних олігонуклеотидів в ад'ювантах або вакцинах згідно з даним винаходом є в загальному випадку низькою, але в залежності від вакцинної композиції може складати - 1000 мкг на дозу, наприклад, 1-500 мкг на дозу, та більш бажано від 1 до 100 мкг на дозу. Кількість сапоніну для застосування в ад'ювантах згідно з даним винаходом може складати таку в інтервалі 1-1000 мкг на дозу, бажано 1-500 мкг на дозу, більш бажано 1-250 мкг на дозу, та найбільш бажано від 1 до 100 мкг на дозу. Композиції згідно з даним винаходом можуть використовуватися як з профілактичною, так і терапевтичною метою. Терапія включає профілактичне лікування. Згідно з цим винахід забезпечує вакцинну композицію, як описано в даній заявці, для застосування у медицині, наприклад, для лікування (включаючи профілактику) малярії. Додатковий аспект даного винаходу полягає у забезпеченні способу одержання гібридного білка згідно з винаходом, при цьому вказаний спосіб включає експресію послідовності ДНК, що кодує білок, у прийнятному хазяїні, бажано, дріжджах, та виділення продукту. Додатковий аспект винаходу полягає у забезпеченні способу лікування пацієнта, чутливого до інфекції Plasmodium, шляхом введення ефективної кількості вакцини, як описано в даній заявці вище. У додатковому аспекті забезпечується комбінація для лікування, що включає: імуногенну частинку згідно з винаходом, вірусний вектор, який кодує малярійний антиген, такий, як вказаний гібридний антиген. Наприклад, якщо вказана частинка та вказаний вірусний вектор вводяться сумісно, то вони, наприклад, можуть бути змішані для одночасного введення або, альтернативно, можуть бути рецептовані для послідовного введення. Як використовується в даній заявці термін «сумісно» означає, що вказана комбінація компо 23 нентів вводиться у межах періоду часу не більше 12 годин, наприклад, протягом періоду часу більше 1 години, типово за один прийом, наприклад, у ході того самого візиту до лікаря, наприклад, коли вони вводяться послідовно або одночасно. Винахід також включає режими примуваннябустингу різними компонентами, описаними у даній заявці, наприклад, примування проводять вірусним вектором, а бустинг-ін'єкції проводять вказаними частинками, або навпаки. У контексті даного опису термін «такий, що включає» інтерпретується як такий, що містить. Аспекти винаходу, що включають певний елемент, також призначені для охоплення вказаних аспектів або суттєво складаються з релевантних елементів. Приклади, приведені нижче, надані для ілюстрації методології, що може використовуватися для одержання частинок згідно з винаходом. Приклади можуть або не можуть утворювати аспект винаходу. ПРИКЛАДИ Приклад 1 Опис штаму Y1834 Дріжджовий рекомбінантний штам Y1834 можу використовуватися для експресії злитого білка. Він складається із хазяйського штаму Saccharomyces cerevisiae DC5, трансформованого рекомбінантним експресійним вектором pRIT15546. DC5 являє собою лабораторний штам дріжджів (ATCC No: 20820) з наступним генотипом: leu2-3, Ieu2-112, his3, саn1-11. Подвійна leu-2 мутація дозволяє проводити селекцію на введення та підтримання вектора pRIT15546, що несе функціональну копію гена LEU-2. Тільки ті клітини, що несуть вектор з LEU-2 геном, можуть рости при відсутності лейцину у ростовому середовищі. Вектор pRIT15546 є дріжджовим епісомальним експресійним вектором (вектор на основі 2), що несе експресійну касету. Рекомбінантна експресія направляється промотором, що походить від дріжджового гена TDH3 (конститутивна експресія). Конструювання вектора pRIT15546 детально приведене нижче. Конструювання вектора pRIT15546. Синтетичний ген із прийнятною переважністю кодонів для дріжджової експресії конструювали та субклонували у вектор pUC57. Одержану плазміду pUC57/CSV та дріжджовий експресійний вектор pGf1-S2 піддавали рестрикції при використанні прийнятних ферментів. Вектор pGf1-S2 конструювали (у GSK) при використанні багатоетапної процедури клонування. Цей вектор, який вже несе S експресійну касету, дозволяє проводити конструювання злитих генів, таке, як N-термінальне злиття у рамці з геном S вірусу гепатиту В. Заключний експресійний вектор після перевірки послідовності мав назву pRIT15546 (Фіг.3) Трансформація штаму DC5. Leu- та his-ауксотрофний DC5 штам піддавали трансформації рекомбінантною плазмідою pRIT15546 при використанні стандартного пропису для трансформації дріжджів. Трансформовані клітини висаджували на агарові селективні планшети. Відбирали один трансформант, який одержу 95293 24 вав офіційну назву Y1834. Експресія рекомбінантного білка: Y1834 вирощували при 30°C у середовищі YNB (дріжджове з доданням азотистої основи, яке одержують від Kracker Scientific lnc) мінімальному середовищі, доповненому 8 мкг/мл гістидину до оптичної густини (620 нм) 0,5. Потім клітини збирали та одержували клітинні екстракти. Приготування екстракту: Клітини ресуспендували у буфері для диспергування та піддавали механічному руйнуванню (скляні кульки). Екстракт центрифугували протягом 15 хвилин при 5000 об./хв. Супернатантну фракцію розганяли на 4-20% SDS - поліакриламідному гелі. Буфер для диспе- буфер на основі 50 мМ фосргування: фату Na (рН: 7,5) 4 мМ EDTA Твін-20 0,5% + суміш інгібіторів протеаз (повна/ ROCHE) Концентрація клі- 100 мл культури (оптична густин: тина (OD): 0,5) у 5 мл диспергувального буфера = концентрація 10 OD одиниць/мл. Освітлення сиро- екстракт центрифугували прового екстракту: тягом 15 хвилин при 5000 об./хв. Визначення рекомбінантного білка Освітлені екстракти розганяли на 4-20% SDSполіакриламідному гелі, білки переносили на нітроцелюлозну мембрану та піддавали імунозабарвлюванню. Вестерн-блотінг аналіз: Реагент = Мишаче моноклональне антитіло анти-S (одержане GSK Biologicals) - (розведення: 1/500) Анти-S антитіла, які є комерційно доступними, можуть замінювати ті, що використовуються у цьому способі. Альтернативно, можуть використовуватися анти-CSV антитіла, наприклад, ті, що є відомими як MR4, доступні від NIH. Приклад 2 Опис штаму Y1835 Дріжджовий рекомбінантний штам Y1835 одночасно експресував CSV-S злитий білок та S антиген. Для одержання штаму, що сумісно експресує CSV-S та S білки, штам Y1295, Saccharomyces cerevisiae, який вже несе п'ять інтегрованих копій S експресійних касет, піддавали трансформації за допомогою рекомбінантного інтегративного експресійного вектора pRIT15582. Штам Y1295 конструювали на GSK шляхом багатоетапної процедури трансформації. Конструювання штаму Y1295 є описаним у WO 93/10152. Штам Y1295 має наступний генотип: leu2-3, Ieu2-112, gall Leu-2 мутація дозволяє проводити селекцію на поглинання лінійного фрагменту ДНК, що походить від pRIT15582, що несе CSV-S касету та функціональний ген LEU2. Вектор pRIT15582 являє собою інтегративний дріжджовий вектор (Ту-вектор), що несе CSV-S експресійну касету. Рекомбінантна експресія направляється промотором, що походить від дріж 25 джового гена TDH3 (конститутивна експресія). Конструювання вектора pRIT15582 у деталях приведене нижче. Конструювання pRIT15582 інтегративного вектора. Вихідний матеріал, який використовувася для конструювання вектора pRIT15582, являв собою експресійну плазміду pRIT15546 (Фіг.1). Конструювання цієї плазміди є описаним у Прикладі 1. Перетравлювання pRIT15546 за допомогою Hindlll ендонуклеази вивільняє ДНК фрагмент довжиною 3706 п.о., що відповідає повній експресійній касеті CSV-S (pTDH3+CSV-S+tARG3). Цей Hindlll ДНК фрагмент (після заповнення при використанні Т4 ДНК полімерази) вбудовували у Ту-інтегративний вектор pRIT13144 в унікальному Sall клонуючому сайті (такий, що піддавали рестрикції за допомогою Sall/оброблений Т4). Одержана плазміда pRIT15582 містила на доповнення до експресійної касети дріжджовий ген LEU2 як селективний маркер (Фіг.3). Перетравлювання pRIT15582 за допомогою Xhol ендонуклеази вивільняло лінійний фрагмент довжиною 8500 п.о., показаний на Фіг.4, що може бути інтегрований у дріжджовий геном шляхом гомологічної рекомбінації вільних кінців за допомогою резидентних Ту-елементів. Трансформація штаму Y1295. Для одержання штаму, що експресує як S, так CSV-S білки, штам Y1295 піддавали трансформації за допомогою лінійного Xhol фрагменту довжиною 8500 п.о. (Фіг.4) із селекцією на Leu+ колонії. Одержували декілька інтегрантів, що містять набори обох експресійних касет, що є присутніми у геномі при різних співвідношеннях. Один трансформант, який несе чотири копії CSV-S касет відбирали та давали офіційне позначення Y1835. Експресія рекомбінантного білка: Y1835 вирощували при 30°C у середовищі YNB (дріжджове з азотистою основою, яке одержують від Kracker Scientific Inc.) мінімальному середовищі до оптичної густини (620 нм) приблизно 0,5 (0,8). Потім клітини збирали та одержували клітинні екстракти. Аналіз експресії ПРОДУКТІВ за допомогою імуноблотінгу Приготування екстракту: Клітини ресуспендували у буфері для диспергування та піддавали механічному руйнуванню (скляні кульки). Екстракт центрифугували протягом 15 хвилин при 5000 об./хв. Супернатантну фракцію розганяли на 12,5%. SDS - поліакриламідному гелі Буфер для диспер- 50 мМ фосфату Na (рН: 7,5) гування: 4 мМ EDTA Твін-20 0,5% + суміш інгібіторів протеаз (Повна/ ROCHE) Концентрація клі- 100 мл культури (оптична густин: тина (OD): 0,5) у 2,5 мл диспергувального буфера = концентрація 20 OD одиниць/мл. Освітлення сиро- екстракт центрифугували вого екстракту: протягом 5-10 хвилин при 5000 об./хв. 95293 26 Визначення рекомбінантного білка Освітлені екстракти розганяли на 12,5% SDSполіакриламідному гелі, білки переносили на нітроцелюлозну мембрану та піддавали імунозабарвлюванню. Вестерн-блотінг аналіз (Фіг.5) Реагенти 1/ Мишаче моноклональне антитіло анти-S (одержане GSK Biologicals)- (розведення: 1/250) 2/ Кроляче поліклональне антитіло анти-CSV (люб'язно надане WRAIR) - розведення 1/20000 Анти-S антитіла, а також анти-Р. vivax/CSP антитіла, які є комерційно доступними, можуть замінювати ті, що використовуються у цьому способі. Приклад 3: Опис штаму Y1845 Дріжджовий рекомбінантний штам Y1845 одночасно експресує CSV-S злитий білок, RTS злитий білок та S антиген. Для одержання штаму, який сумісно експресує CSV-S, RTS та S білки, штам Y1295 Saccharomyces cerevisiae, який вже несе 5 інтегрованих копій S експресійних касет, піддавали трансформації за допомогою еквімолярного розчину RTS та CSV-S лінійних фрагментів ДНК, що походять від pRIT13540 та pRIT15582 інтегративних векторів, відповідно. Штам Y1295 конструювали на GSK шляхом багатоетапної процедури трансформації. Конструювання Y1295 штаму є описаним у WO 93/10152. Штам Y1295 має наступний генотип: leu2-3, Ieu2-112, gall Leu-2 мутація дозволяє проводити селекцію на поглинання лінійного фрагменту ДНК, який походить від pRIT15582, що несе CSV-S касету та функціональний ген LEU2. Вектори pRIT13540 та pRIT15582 являють собою інтегративні дріжджові вектори (вектори на основі Ту), що несуть RTS та CSV-S експресійні касети, відповідно. Для обох векторів рекомбінантна експресія направляється промотором, що походить від дріжджового гена TDH3 (конститутивна експресія). Конструювання вектора pRIT15582 у деталях приведене у Прикладі 2. Конструювання вектора pRIT13540 є описаним у WO 93/10152. Одержання лінійних інтегративних фрагментів ДНК. Інтегративні вектори pRIT13540 та pRIT15582 обидва піддавали рестрикції Xhol ендонуклеазою, вивільняючи ДНК фрагменти довжиною 8200 п.о. та 8500 п.о., відповідно. Ці фрагменти можуть бути інтегровані у дріжджовий геном шляхом гомологічної рекомбінації вільних кінців з резидентними Туелементами. Трансформація штаму Y1295. Для одержання штаму, що експресує три білки RTS, CSV-S та S, штам Y1295 піддавали трансформації за допомогою еквімолярного розчину лінійних Xhol фрагментів довжиною 8200 п.о. та 8500 п.о. (Фіг.4) із селекцією на Leu+ колонії. Одержували декілька інтегрантів, які містять набори двох експресійних касет, що є присутніми у геномі при різних співвідношеннях. Один трансформант, що несе чотири копії CSV-S та дві копії RTS (на доповнення до п'яти копій S касет), відбирали та давали офіційне позначення Y1845. Експресія рекомбінантного білка: Y1845 вирощували при 30°C у середовищі 27 YNB (дріжджове з азотистою основою, яке одержують від Kracker Scientific lnc) мінімальному середовищі до оптичної густини (620 нм) приблизно 0,5. Потім клітини збирали та одержували клітинні екстракти. Аналіз експресії продуктів за допомогою імуноблотінгу Приготування екстракту: Клітини ресуспендували у буфері для диспергування та піддавали механічному руйнуванню (скляні кульки). Екстракт центрифугували протягом 15 хвилин при 5000 об./хв. Супернатантну фракцію розганяли на 12,5% SDS - поліакриламідному гелі Буфер для диспер- 50 мМ фосфату Na (рН: 7,5) гування: 4 мМ EDTA Твін-20 0,5% + суміш інгібіторів протеаз (Повна/ ROCHE) Концентрація клі- 100 мл культури (оптична густин: тина (OD): 0,5) у 5 мл диспергувального буфера = концентрація 10 OD одиниць/мл. Освітлення сиро- екстракт центрифугували вого екстракту: протягом 5-10 хвилин при 5000 об./хв. Визначення рекомбінантного білка Освітлені екстракти розганяли на 12,5% SDSполіакриламідному гелі, білки переносили на нітроцелюлозну мембрану та піддавали імунозабарвлюванню. Вестерн-блотінг аналіз (Фіг.5) Реагент Мишаче моноклональне антитіло анти-S (одержане GSK Biologicals) - (розведення: 1/500) Центрифугування у градієнті густини CsCI: Утворення частинок у штамі Y1845 піддавали аналізу за допомогою центрифугування у градієнті густини CsCI. Сирові екстракти (~20 мг загального білка) аналізували у градієнті 12 мл 1,5 M CsCI (88 годин при 40000 об./хв., +8°C на Beckman 70.1 роторі Ті). Збирали фракції (~0,6 мл) та аналізували шляхом імуноблотінгу при використанні анти-S антитіла. Як показано на Фіг.8, піки вестернблотінгу виявлялися для тих самих фракцій (№№10 та 11) градієнта, що відповідало густині 3 спливання 1,21 та 1,20 г/см . Утворення триплет 95293 28 них частинок підтримувалося градієнтним аналізом. Посилання 1. Harford N, Cabezon T, Colau В, та ін., "Construction and Characterization of a Saccharomyces cerevisiae Strain (RIT4376) Expressing Hepatitis B Surface Antigen", Postgrad Med J 63, Supp. 2: 65-70, 1987. 2. Jacobs E, Rutgers T, Voet P, та ін., "Simultaneous Synthesis and Assembly of Various Hepatitis B Surface Proteins in Saccharomyces cerevisiae", Gene 80: 279-291, 1989. 3. Vieira J and Messing J, "The pUC plasmids, an M13mp7-Derived System for Insertion Mutagenesis and Sequencing with Synthetic Universal Primers", *_Gene 19: 259-268, 1982. 4. Hinnen A, Hicks JB1 and Fink GR1 "Transformation of Yeast", Proc Natl Acad Sci USA 75: 1929-1933, 1980. 5. Broach JR, Strathern JN, and Hicks JB, "Transformation in Yeast Development of a Hybrid Cloning Vector and Isolation of the CAN 1 Gene", Gene 8:121-133,1979. 6. Zhang H, та ін., "Double Stranded SDNA Sequencing as a Choice for DNA Sequencing", Nucleic Acids Research 16:1220, 1988. 7. Dame JB, Williams JL. Mc Cutchan TF, та ін., "Structure of the Gene Encoding the Immunodominant Surface Antigen on the Sporozoites of the Human Malaria Parasite Plasmodium falciparum", Science 225: 593-599, 1984. 8. Valenzuela P, Gray P, Quiroga M, та ін., "Nucleotide Sequences of the Gene Coding for the Major Protein of Hepatitis B Virus Surface Antigen", Nature 280: 815-819, 1979. 9. In SS, Kee-Hoyung L, Young RK, та ін., "Comparison of Immunological Responses to Various Types of Circumsporozoite Proteins of Plasmodium vivax in Malaria Patients of Korea", Microbiol. Immunol. 48(2): 119-123, 2004;Microbiol. Immunol. 2004; 48(2): 119-123. 10. Rathore D, Sacci JB1 de Ia Vega P, та ін., "Binding and Invasion of Liver Cells by Plasmodium falciparum Sporozoites", J. Biol. Chem. 277(9): 70927098, 2002. Rathore та ін., 2002, J. Biol. Chem. 277, 7092-8. 29 95293 30 31 95293 32 33 95293 34 35 95293 36 37 95293 38 39 95293 40 41 95293 42 43 Комп‟ютерна верстка А. Рябко 95293 Підписне 44 Тираж 23 прим. Міністерство освіти і науки України Державний департамент інтелектуальної власності, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601
ДивитисяДодаткова інформація
Назва патенту англійськоюVaccines for malaria
Автори англійськоюCohen Joseph D, Marchand Martine, Ockenhouse Christian F, Yadava Anjali
Назва патенту російськоюМалярийные вакцины
Автори російськоюКоэн Джозеф Д., Маршан Мартин, Оукенгаус Кристиан Ф., Ядава Анджали
МПК / Мітки
МПК: A61P 31/00, C07K 14/445, A61K 39/015
Код посилання
<a href="https://ua.patents.su/22-95293-malyarijjni-vakcini.html" target="_blank" rel="follow" title="База патентів України">Малярійні вакцини</a>
Авірулентний ізолят lawsonia intracellularis європейського походження, вакцини для імунізації тварин, застосування заявленого авірулентного ізоляту lawsonia intracellularis як вакцини, спосіб одержання вакцини,

Номер патенту: 87665
Опубліковано: 10.08.2009
Автори: Кроулл Джеремі Дж., Ніттел Джеффрі П., Руфф Майкл Б.
МПК: C12R 1/01, C12Q 1/02, A61K 35/16, A61K 39/02, G01N 33/554, A61B 10/00, C12N 1/20
Мітки: імунізації, ізолят, застосування, intracellularis, ізоляту, європейського, тварин, авірулентний, спосіб, походження, одержання, вакцини, заявленого, авірулентного, lawsonia
Формула / Реферат:
1. Авірулентний ізолят Lawsonia intracellularis, де авірулентний ізолят являє собою ізолят Lawsonia intracellularis, депонований під реєстраційним номером ATCC PTA-4926, або будь-який авірулентний ізолят Lawsonia intracellularis європейського походження, який відрізняється тим, що зазначений авірулентний Lawsonia intracellularis не виділяється з фекаліями на 14 день після...
Композиція вакцини (варіанти), спосіб стабілізації qs21 відносно гідролізу (варіанти), спосіб приготування композиції вакцини

Номер патенту: 56132
Опубліковано: 15.05.2003
Автори: Гарсон Наталі Марі Жозеф Клод, Фріде Мартін
МПК: A61K 39/02, A61K 39/39, A61K 39/12
Мітки: вакцини, відносної, спосіб, варіанти, стабілізації, гідролізу, приготування, композиція, композиції
Формула / Реферат:
1. Композиция вакцины, содержащая антиген, QS21 и стерин, отличающаяся тем, что весовое соотношение QS21 и стерина составляет от 1:1 до 1:100.2. Композиция по п. 1, отличающаяся тем, что весовое соотношение QS21 и стерина составляет от 1:1 до 1:5.3. Композиция по п. 1, отличающаяся тем, что весовоесоотношение QS21 и стерина составляет 1:2.4. Композиция по любому из пп. 1-3, отличающаяся тем, что...
Композиція вакцини проти малярії та спосіб лікування або попередження захворювання малярією з використанням цієї композиції

Номер патенту: 68336
Опубліковано: 16.08.2004
Автор: Кохен Джозеф
МПК: A61K 39/002, A61K 39/29, A61K 39/015, A61K 39/39
Мітки: малярії, малярією, попередження, лікування, композиція, використанням, спосіб, захворювання, вакцини, композиції, цієї
Формула / Реферат:
1. Композиція протималярійної вакцини, що містить принаймні два специфічні для малярії антигени, в комбінації з ад¢ювантом, що є вибірковим стимулятором відповіді ТН1 клітин, і де принаймні два малярійних антигени вибрані з групи, яка включає:а) гібридний протеїн, що включає всю С-термінальну ділянку циркумспорозоїтного (CS) протеїну, чотири або більше подвійних повторів імунодомінантної ділянки та поверхневий антиген вірусу...
Мутант коров’ячого вірусу герпесу типу 1 (bhv-1), композиція вакцини для вакцинації тварин для захисту їх від bhv-1, композиція вакцини для вакцинації тварин для захисту їх від патогена

Номер патенту: 40571
Опубліковано: 15.08.2001
Автори: Рейсевейк Франціскус Антоніус Марія, Оірсот Йоханнес Теодорус ван, Має Ооже Кам'єль
МПК: C12N 7/01, A61P 31/12, C12N 15/09, G01N 33/569, G01N 33/53, C12N 15/00, A61K 39/395, C07K 14/06, A61K 39/265, A61K 39/245, C12N 7/00, C12P 21/08, C12N 15/83, C12Q 1/70, C12N 15/869
Мітки: герпесу, мутант, захисту, патогена, типу, тварин, вірусу, вакцинації, вакцини, коров'ячого, композиція, bhv-1
Формула / Реферат:
1.Мутант коровьего вируса герпеса типа 1 (BHV-1), имеющий делецию в gЕ-гене гликопротеина, причем gЕ-ген гликопротеина расположен в US-регионе между сайтами рестрикции Pst 1 и Sma 1, причем указанная делеция позволяет мутанту отличаться серологически от BHV-1 дикого типа.2.Мутант BHV-1 по п.1, отличающийся тем, что указанная делеция в gЕ-гене гликопротеина вызвана аттенуацией.3.Мутант BHV-1 по п. 2, отличающийся тем, что...
Спосіб отримання інактивованої вакцини проти лейкозу великої рогатої худоби

Номер патенту: 33513
Опубліковано: 25.06.2008
Автори: Нагаєві Галина Григорівна, Нагаєв Евген Миколайович
МПК: A61K 39/002, C12N 5/00
Мітки: спосіб, вакцини, худоби, інактивованої, лейкозу, отримання, рогатої, великої
Формула / Реферат:
Спосіб отримання інактивованої вакцини проти лейкозу великої рогатої худоби, що включає культивування вірусвмісних лейкоцитів периферійної крові великої рогатої худоби, хворої на лейкоз, їх виділення з культуральної речовини з наступною гомогенізацією, обробку отриманого лізату 10 % 3М розчином хлористого калію та тритоном х - 100, його інкубування при 4±1 °С, розведення обробленого лізату забуференим фізіологічним розчином до кінцевого...
Попередній патент: Спосіб запобігання утворенню пилу із зернистих матеріалів
Наступний патент: Спосіб автоматичного відводу повітря з унітаза і пристрій для його здійснення
Випадковий патент: Пристрій для розвитку руки та навчання письму з використанням персонального комп'ютера