Виділений поліпептид, який зв’язується з рецептором інсуліноподібного фактора росту 1







Формула / Реферат
1. Виділений поліпептид, який зв′язується з рецептором інсуліноподібного фактора росту 1, що містить поліпептид з послідовністю, представленою в SEQ ID NO:1-13.
2. Виділений полінуклеотид, що містить полінуклеотид, який кодує поліпептид, що містить амінокислотну послідовність, представлену в SEQ ID NO:1-13.
3. Виділений полінуклеотид, що містить полінуклеотид з послідовністю, представленою в SEQ ID NO:14-26, або його комплементарною послідовністю.
4. Виділений вектор, що містить полінуклеотид з послідовністю, представленою в SEQ ID NO:14-26.
5. Вектор за п. 4, що являє собою вектор експресії.
6. Виділена клітина-хазяїн, що містить вектор за п. 4.
7. Виділений злитий білок, що містить поліпептид з послідовністю, приведеною в SEQ ID NO:1-13, злитий з другим поліпептидом.
8. Злитий білок за п. 7, де другий поліпептид кодує імуноглобулін або його фрагмент.
9. Спосіб експресії поліпептиду, що включає стадії:
a) забезпечення клітини-хазяїна за п. 6; і
b) культивування клітини-хазяїна в умовах, достатніх для експресії поліпептиду з послідовністю, приведеною в SEQ ID NO:1-13.
10. Спосіб доставки терапевтичного агента через клітини ендотелію, що включає:
a) кон'югацію терапевтичного агента з поліпептидом, що містить поліпептид з послідовністю, приведеною в SEQ ID NO:1, 2, 4, 8 або 12, з утворенням кон'югата;
b) приведення кон'югата в контакт з клітинами ендотелію; і
с) вимірювання кількості кон'югата, доставленої через клітини ендотелію.
11. Спосіб за п. 10, де клітини ендотелію утворюють гематоенцефалічний бар'єр.
Текст
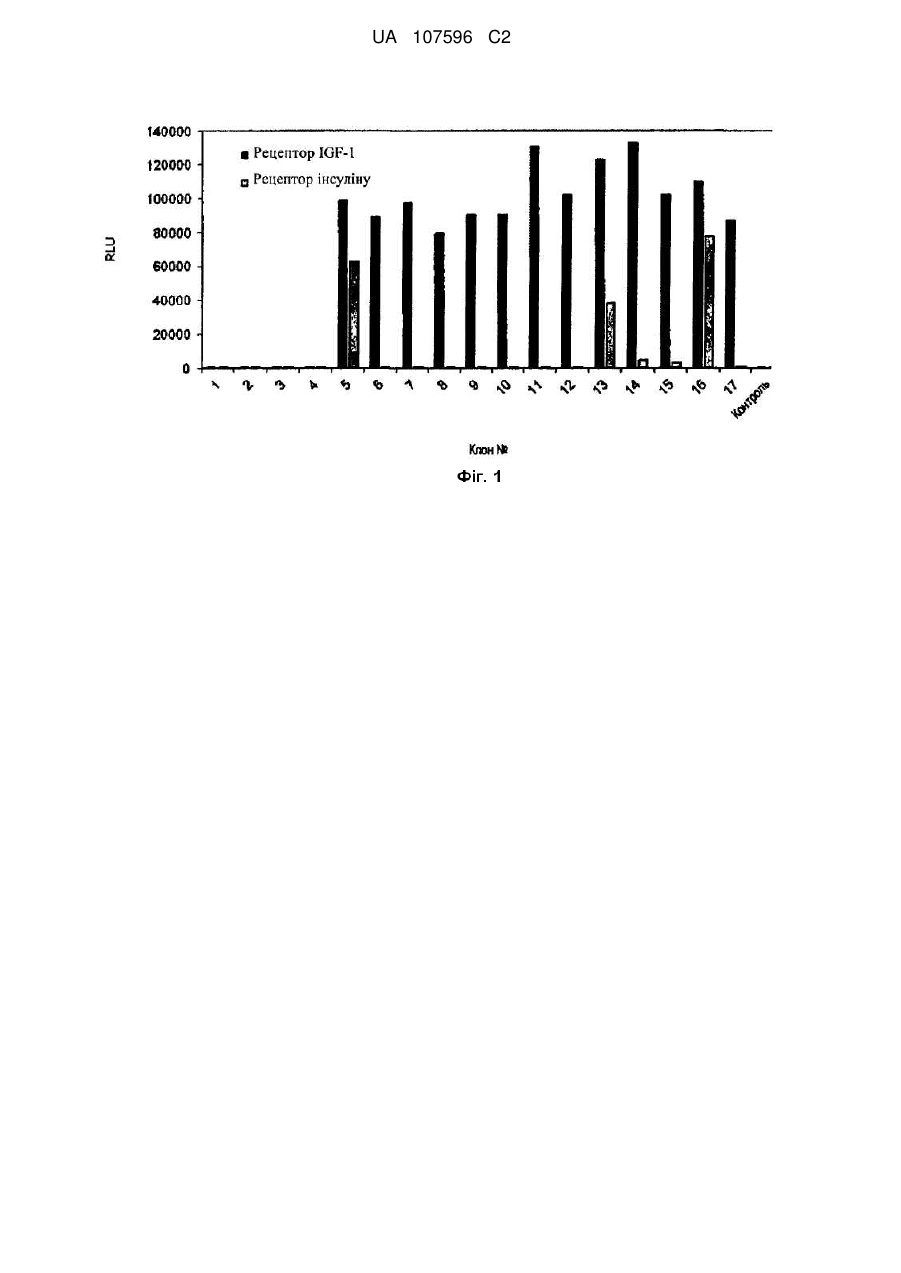
Реферат: Винахід належить до виділеного поліпептиду, який зв′язується з рецептором інсуліноподібного фактора росту 1. Також винахід належить до способу експресії вказаного поліпептиду. UA 107596 C2 (12) UA 107596 C2 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 ГАЛУЗЬ ТЕХНІКИ, ДО ЯКОЇ НАЛЕЖИТЬ ВИНАХІД Даний винахід стосується пептидів, що зв'язуються з рецептором інсуліноподібного фактора росту 1, полінуклеотидів, що їх кодують, а також способів їх отримання і застосування. ПЕРЕДУМОВИ СТВОРЕННЯ ВИНАХОДУ Останні досягнення в галузі біомедичних досліджень і високопродуктивному скринінгу лікарських препаратів дозволили запропонувати множину потенційних терапевтичних агентів для лікування захворювань, пов'язаних з центральною нервовою системою. Однак багато з потенційних терапевтичних агентів виявляється незадовільними при тестуванні in vivo через недостатньо ефективний транспорт через гематоенцефалічний бар'єр (ГЕБ). Терапевтичні агенти можуть транспортуватися через ГЕБ декількома способами, включаючи насичувані транспортні системи, адсорбційний трансцитоз, коли терапевтичний агент, що транспортується, інтерналізується клітиною в ГЕБ і доставляється на аблюмінальну поверхню для вивільнення у внутрішньоклітинний рідинний компартмент мозку, трансмембранну дифузію, причому терапевтичний агент розчиняється в ліпідному бішарі, з якого складаються мембрани клітин, які складають ГЕБ, а також позаклітинні шляхи, коли терапевтичний агент використовує залишкову проникність ГЕБ. Випробувано декілька підходів до модифікації терапевтичних агентів для зміни їх проникаючої здатності через ГЕБ, включаючи кон'югацію з білками, природно проникаючими через ГЕБ, такими як інсулін, інсуліноподібний фактор росту 1 і 2 (IGF-1, IGF-2), лептин і трансферин (заявка на патент США № US2007/0081992), зшивання поліпептидів з катіонізованими антитілами, що зв'язуються з певними клітинними рецепторами, такими як інсуліновий рецептор (патент США № 7388079) або трансфериновий рецептор (патент США № 6329508; Zhang and Pardridge, Brain Res. 889:49-56, 2001); зв'язування терапевтичних агентів з синтетичними полімерами, такими як полі(бутилціаноакрилат) або поліакриламід, покритий полісорбатом 80 (заявка на патент США № 2002/0009491, заявка на патент США № 2002/0013266; заявка на патент США № 2006/0051317), і використання ліпосом або імуноліпосом. Для всіх підходів, що використовуються сьогодні, до підвищення ефективності транспорту терапевтичних агентів через ГЕБ характерна невисока ефективність через конкуренцію з ендогенними лігандами, недостатній транспорт терапевтичного агента в паренхіму мозку і деградацію терапевтичних агентів внаслідок доставки його в лізосоми. Таким чином, існує потреба в розробці способів транспорту терапевтичних агентів через ГЕБ. КОРОТКИЙ ОПИС ФІГУР На фіг. 1 показано зв'язування окремих лізатів фагів з IGF1R і IR. На фіг. 2 показано зв'язування злитого з AP білка з IGF1R. КОРОТКИЙ ОПИС ВИНАХОДУ Один аспект даного винаходу являє собою виділений поліпептид, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1-13. Інший аспект даного винаходу являє собою виділений полінуклеотид, що містить полінуклеотид, який кодує поліпептид, що містить амінокислотну послідовність, представлену в SEQ ID NO: 1-13. Інший аспект даного винаходу являє собою виділений полінуклеотид, що містить полінуклеотид з послідовністю, приведеною в SEQ ID NO: 14-26, або його комплементарною послідовністю. Інший аспект даного винаходу являє собою виділений вектор, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 14-26. Інший аспект даного винаходу являє собою виділену клітину-хазяїна, що містить вектор, який складає предмет даного винаходу. Інший аспект даного винаходу являє собою виділений злитий білок, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1-13, злитий з другим поліпептидом. Інший аспект даного винаходу являє собою спосіб експресії поліпептиду, що включає стадії: a. забезпечення клітини-хазяїна, що складає предмет даного винаходу; і b. культивування клітини-хазяїна в умовах, достатніх для експресії поліпептиду з послідовністю, приведеною в SEQ ID NO: 1-13. Інший аспект даного винаходу являє собою спосіб доставки терапевтичного агента через клітини ендотелію, що включає a. кон'югацію терапевтичного агента з поліпептидом, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1, 2, 4, 8 або 12, з утворенням кон'югата; b. приведення кон'югата в контакт з клітинами ендотелію; і 1 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 с. вимірювання кількості кон'югата, доставленої через клітини ендотелію. ДОКЛАДНИЙ ОПИС ВИНАХОДУ Всі публікації, що згадуються в даному описі, включаючи, без обмежень, патенти і заявки, включені в цей документ шляхом посилання, як якби вони були повністю викладені безпосередньо в цьому документі. Використовувані в даному описі і в формулі винаходу форми однини включають посилання на множинні об'єкти, якщо інше не випливає явно з контексту. Так, наприклад, посилання на "поліпептид" має на увазі посилання на один або більше поліпептидів і включає їх еквіваленти, відомі фахівцям в даній галузі. Всі технічні і наукові терміни, що використовуються в цьому документі, якщо тільки не дане інше їх визначення, мають загальноприйняте значення, зрозуміле будь-якому фахівцеві в галузі даного винаходу. У цьому документі описані приклади композицій і способів, хоча для перевірки або реалізації на практики даного винаходу можуть бути використані будь-які композиції і способи, подібні або еквівалентні описаним в цьому документі. Термін "поліпептид" означає молекулу, що містить щонайменше два амінокислотні залишки, зв'язані пептидним зв'язком з утворенням поліпептиду. Малі поліпептиди, що містять менше ніж 50 амінокислотних залишків, можуть називатися "пептидами". Поліпептиди можуть також називатися "білками". Термін "полінуклеотид" означає молекулу, що містить ланцюжок нуклеотидів, ковалентно пов'язаних через цукрофосфатний кістяк або через іншу еквівалентну ковалентну хімічну структуру. Дволанцюжкові і одноланцюжкові молекули ДНК і РНК являють собою типові приклади полінуклеотидів. Термін "комплементарна послідовність" означає другу виділену полінуклеотидну послідовність, що антипаралельна першій виділеній полінуклеотидній послідовності і містить нуклеотиди, комплементарні нуклеотидам першої полінуклеотидної послідовності. Як правило, такі "комплементарні послідовності" при контакті з першою виділеною полінуклеотидною послідовністю у відповідних умовах здатні до утворення дволанцюжкових полінуклеотидних молекул, таких як дволанцюжкові молекули ДНК або дволанцюжкові молекули РНК. Термін "вектор" означає полінуклеотид, який може подвоюватися в біологічній системі або переміщуватися між такими системами. Полінуклеотиди-вектори, як правило, містять елементи, такі як точки початку реплікації, сигнал поліаденілування або маркери вибору, що забезпечують дуплікацію або збереження таких полінуклеотидів в біологічній системі. Прикладами згаданих біологічних систем можуть служити клітини, віруси, тваринні, рослини і реконструйовані біологічні системи, що використовують біологічні компоненти, здатні до подвоєння вектора. Полінуклеотидами, що містять вектор, можуть бути молекули ДНК або РНК, або їх гібриди. Термін "вектор експресії" означає вектор, який можна використовувати в біологічній системі або реконструйованій біологічній системі для прямої трансляції поліпептиду, закодованого полінуклеотидною послідовністю, присутньою у векторі експресії. Використовуваний в цьому документі термін "гематоенцефалічний бар'єр" або "ГЕБ" означає межу розділу між кров'ю периферичної системи кровообігу і головним і спинним мозком, яка утворена щільними з'єднаннями в ендотеліальних плазматичних мембранах капілярів мозку і створює виключно щільний бар'єр, що обмежує транспорт молекул в мозок, навіть таких малих, як молекула сечовини з молекулярною масою 60 Да. Гематоенцефалічний бар'єр в головному мозку, гематоспинномозковий бар'єр в спинному мозку і гематоретинальний бар'єр в сітківці являють собою безперервні капілярні бар'єри в центральній нервовій системі (ЦНС) і збірно називаються гематоенцефалічним бар'єром. Термін "антитіло" означає молекулу, яка специфічно зв'язується з антигеном, і включає димерні, тримерні і мультимерні антитіла, а також химерні, гуманізовані і повністю людські антитіла. Крім того, антитіло може являти собою ціле антитіло або функціональний фрагмент молекули антитіла, такий як фрагмент, що зберігає щонайменше її функцію зв'язування з антигеном, що включають Fab, F(ab´), F(ab´)2, scFv, dsFv і діатіла. Наприклад, фрагменти антитіла можуть бути отримані з використанням протеолітичних ферментів (наприклад, ціле антитіло розщеплюється папаїном для отримання фрагментів Fab, а обробка пепсином дає фрагменти F(ab´)2). Технології отримання і застосування різних антитіл добре відомі в даній галузі (під ред. Ausubel, et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., м. Нью-Йорк, 1987-2001; Sambrook, et al., Molecular Cloning: А Laboratory Manual, 2-е видання, Колд-Спринг-Харбор, штат Нью-Йорк, 1989; Harlow and Lane, Antibodies, а Laboratory Manual, Колд-Спринг-Харбор, штат Нью-Йорк, 1989; під ред. Colligan, et al., Current Protocols in Immunology, John Wiley & Sons, Inc., м. Нью-Йорк, 1994-2001; Colligan et al., Current Protocols in Protein Science, John Wiley & Sons, м. Нью-Йорк, штат Нью-Йорк, 1997-2001; Kohler et al., Nature 2 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 256:495-497, 1975; патент США № 4816567, Queen et al., Proc. Natl. Acad. Sci. 86:10029-10033, 1989). Наприклад, повністю людські моноклональні антитіла, що не включають ніяких послідовностей, що не стосуються людини, можуть бути отримані з допомогою трансгенних по людському імуноглобуліну мишей або за допомогою бібліотек фагових дисплеїв (Lonberg et al., Nature 368:856-859, 1994; Fishwild et al., Nature Biotech, 14:845-851, 1996; Mendez et al., Nature Genetics 15:146-156, 1997; Knappik et al., J. Mol. Biol. 296:57-86, 2000; Krebs et al., J. Immunol. Meth. 265:67-84, 2001). Молекула або препарат антитіла "специфічно зв'язується" із заданим антигеном, якщо вони зв'язуються з даним антигеном з вищою афінністю і специфічним, на противагу неспецифічному, чином в порівнянні з іншим, відмінним від першого, антигеном. Іншими словами, "специфічне зв'язування" молекули або препарату антитіла можна використовувати для розрізнення двох різних поліпептидів. Використовуваний в цьому документі термін "рецептор інсуліноподібного фактора росту 1", або "IGF1R", означає IGF1R людини (GenBank: NP_000866) з амінокислотною послідовністю, приведеною в SEQ ID NO: 27. Для отримання зрілого білка про-поліпептид IGF1R розщеплюється на альфа- і бета-ланцюги. Альфа-ланцюг включає амінокислотні залишки 31740 послідовності SEQ ID NO: 27, а бета-ланцюг включає амінокислотні залишки 741-1367 послідовності SEQ ID NO: 27. Використовуваний в цьому документі термін "розчинний IGF1R", або "sIGF1R", означає позаклітинний домен IGF1R (амінокислотні залишки 31-932 послідовності SEQ ID NO: 27). Розчинний IGF1R може являти собою позаклітинний домен нерозщепленого про-поліпептиду або позаклітинну домен зрілого IGF1R (амінокислотні залишки 31-740, які утворюють альфа-ланцюг, і амінокислотні залишки 741-932, які утворюють позаклітинну частину бета-ланцюга). Використовуваний в цьому документі термін "кон'югат" означає химерну молекулу, що містить пептид, який складає предмет даного винаходу, з амінокислотною послідовністю, приведеною в SEQ ID NO: 1-13, і терапевтичний агент. Термін "кон'югований" або "кон'югування" означає, що терапевтичний(-і) агент(- и) і пептиди, що складає предмет даного винаходу, фізично зв'язані один з одним, наприклад, ковалентними хімічними зв'язками, фізичними силами, такими як сили ван-дер-Ваальса або гідрофобні взаємодії, шляхом інкапсулювання, вбудовування або комбінацій цих способів. Терапевтичний(- і) агент(- и) і пептиди, що складає предмет даного винаходу, можуть бути пов'язані один з одним хімічними зв'язками через спиртову, кислотну, карбонільну тіолову або аміногрупу з використанням добре відомих способів хімічного синтезу (див., наприклад, заявку на патент США № US2010/0028370). Терапевтичний агент може бути пов'язаний з пептидами, що складають предмет даного винаходу, через групу-лінкер. Прикладами лінкерів є гліцин-збагачені лінкери, такі як Gly3SerGly3Ser (SEQ ID NO: 28) або Gly4SerGly4SerGly4Ser (SEQ ID NO: 29). Якщо терапевтичний агент і пептиди, що складають предмет даного винаходу, кон'юговані один з одним через ковалентний зв'язок або пептид і терапевтичного агента являє собою поліпептид, то такий кон'югат як ціле називається "злитий білок". Таким чином, термін "злитий білок" означає поліпептид, утворений з двох (або) більше гетерологічних поліпептидів, які звичайно не зливаються разом в одній амінокислотній послідовності. Злиті білки можуть загалом бути отримані з використанням способів на основі рекомбінантних нуклеїнових кислот, тобто шляхом транскрипції і трансляції продукту злиття рекомбінантних генів, де злиття включає сегмент, що кодує поліпептид, який складає предмет даного винаходу, і сегмент, що кодує гетерологічний поліпептид. Використовуваний в цьому документі термін "терапевтичний агент" означає молекулу, що вводиться суб'єкту для отримання бажаного терапевтичного ефекту. Суб'єктів є людина або тварина, включаючи ссавців і приматів. Приклади терапевтичних агентів включають білки, антитіла, пептиди, малі молекули або полінуклеотиди. Терапевтичні агенти також можуть являти собою токсини або радіоізотопи, коли передбачуваний терапевтичний ефект полягає, наприклад, в знищенні ракових клітин. У даному винаході представлені виділені поліпептиди, які зв'язуються з IGF1R, полінуклеотиди, що кодують поліпептиди, вектори, що містять полінуклеотиди, виділені клітинихазяїни, поліпептиди, які можуть бути отримані при експресії полінуклеотидів, способи експресії поліпептидів, що складають предмет даного винаходу, і способи використання полінуклеотидів і поліпептидів, які складають предмет даного винаходу. Поліпептиди, що складають предмет даного винаходу, зв'язуються з IGF1R і шляхом трансцитозу переносяться через клітини ендотелію. Оскільки IFG1R експресується на клітинах ендотелію в гематоенцефалічному бар'єрі (ГЕБ), поліпептиди, що складають предмет даного винаходу, можуть забезпечувати можливість доставки терапевтичних агентів через ГЕБ. 3 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 Один аспект даного винаходу являє собою виділений поліпептид, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1-13. Поліпептиди, що складають предмет даного винаходу, можуть бути отримані шляхом хімічного синтезу, такого як твердофазний синтез пептидів, в автоматичному синтезаторі пептидів. У альтернативному варіанті здійснення поліпептиди, що складають предмет даного винаходу, можуть бути отримані на основі полінуклеотидів, що кодують ці поліпептиди з використанням безклітинних систем експресії, таких як системи експресії на основі лізату ретикулоцитів, системи експресії на основі екстракту з пшеничних зародків і системи експресії на основі екстракту Escherichia coli. Поліпептиди, що складають предмет даного винаходу, також можуть бути отримані шляхом експресії і виділення з клітин-носіїв нуклеотидної послідовності, що складає предмет даного винаходу, з використанням добре відомих фахівцям способів, таких як рекомбінантна експресія поліпептидів, що легко виділяються з афінними мітками. Фахівцеві в даній галузі також відомі інші способи отримання поліпептидів, що складають предмет даного винаходу. Інший аспект даного винаходу являє собою виділений злитий білок, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1-13, злитий з другим поліпептидом. Вказаний другий поліпептид може являти собою лідерну послідовність або секреторну послідовність. Вказаний другий поліпептид може являти собою терапевтичний агент, злитий з пептидами, що складають предмет даного винаходу. Терапевтичний агент і пептид, що складає предмет даного винаходу, можуть бути злиті один з одним різними способами. С-Кінець або N-кінець пептиду, що складає предмет даного винаходу, може бути безпосередньо пов'язаний з N-кінцем або С-кінцем, відповідно, терапевтичного агента через амідний зв'язок або пептидний лінкер. Терапевтичні агенти можуть бути пов'язані з пептидом, що складає предмет даного винаходу, з використанням добре відомого в даній галузі хімічного крос-поєднання. Інший аспект даного винаходу являє собою виділений полінуклеотид, що містить полінуклеотид, який кодує поліпептид, що становить предмет даного винаходу. Полінуклеотиди, які складають предмет даного винаходу, можуть бути отримані шляхом хімічного синтезу, такого як твердофазний синтез полінуклеотидів, в автоматичному синтезаторі полінуклеотидів. У альтернативному варіанті здійснення полінуклеотиди, що складають предмет даного винаходу, можуть бути отримані з використанням інших способів, таких як ПЛРдуплікація, векторна дуплікація або способи маніпуляції ДНК з використанням рестрикційних ферментів. Способи виробництва або отримання полінуклеотидів з відомою заданою послідовністю добре відомі фахівцям в даній галузі. Полінуклеотиди, що складають предмет даного винаходу, також можуть містити щонайменше одну некодуючу послідовність, таку як транскрибовані, але послідовності, що не транслюються, сигнали термінації, сайти зв'язування рибосоми, послідовності, що стабілізують мРНК, інтрони і сигнали поліаденілування. Полінуклеотидні послідовності також можуть включати в себе додаткові послідовності, що кодують додаткові амінокислоти. Такі додаткові полінуклеотидні послідовності можуть, наприклад, кодувати маркер або мітку, наприклад, гексагістидиновий пептид (Gentz et al., Proc. Natl. Acad. Sci. (США) 86:821-284, 1989) або гемаглютинінову пептидну мітку (Wilson et al., Cell 37:767-778, 1984), що полегшують очищення злитих білків. Прикладами полінуклеотидів є полінуклеотиди з послідовністю, приведеною в SEQ ID NO: 14-26. Інший варіант здійснення даного винаходу являє собою вектор, що містить виділений полінуклеотид з послідовністю, приведеною в SEQ ID NO: 14-26. Вектори, що складають предмет даного винаходу, використовують для збереження полінуклеотидів, подвоєння полінуклеотидів або індукцій експресії поліпептидів, кодованих вектором, що складає предмет даного винаходу, в біологічних системах, включаючи реконструйовані біологічні системи. Вектори можуть мати хромосомальне, епісомальне і вірусне походження. До них можуть належати вектори, отримані з бактеріальних плазмід, бактеріофагів, транспозонів, епісом дріжджів, вставних елементів, хромосомальних елементів дріжджів, бакуловірусів, паповавірусів, таких як SV40, вірусів коров'ячої віспи, аденовірусів, вірусів пташиної віспи, вірусів псевдосказ, пікорнавірусів і ретровірусів, а також вектори, отримані з комбінацій вказаних вище елементів, такі як косміди і фагміди. Вектори, що складають предмет даного винаходу, можуть бути приготовані у вигляді мікрочастинок з допоміжними речовинами, ліпідами, буферними речовинами або іншими ексципієнтами, залежно від конкретного застосування. У одному варіанті здійснення даного винаходу вектор являє собою вектор експресії. Вектори експресії, як правило, включають в себе елементи послідовності нуклеїнових кислот, які дозволяють контролювати, регулювати, активувати або допускати експресію поліпептидів, що 4 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 кодуються даним вектором. Вказані елементи можуть містити сайти зв'язування енхансера транскрипції, сайти ініціація РНК-полімерази, сайти зв'язування рибосом і інші сайти, сприяючі експресії закодованих поліпептидів в даній системі експресії. Вказані системи експресії можуть являти собою добре відомі фахівцям в даній галузі клітинні або безклітинні системи. Також добре відомі елементи послідовності нуклеїнових кислот і послідовності вихідного вектора, придатні для використання в процесі експресії закодованих поліпептидів. Приклад отриманого на основі плазміди вектора експресії, який можна використовувати для експресії поліпептидів, що складають предмет даного винаходу, містить точку початку реплікації Е. coli, ген хлорамфенікол-ацетилтрансферази (CAT), промотор T7 бактеріофага, сигнальну послідовність pelB і послідовність термінатора T7. Інший варіант здійснення даного винаходу являє собою виділену клітину-хазяїна, що містить вектор, який складає предмет даного винаходу. Типові приклади клітин-хазяїнів включають клітини архей; бактеріальні клітини, такі як Streptococci, Staphylococci, Enterococci, Е. coli, Streptomyces, ціанобактерії, В. subtilis і S. aureus; клітини грибів, такі як Kluveromyces, Saccharomyces, Basidomycete, Candida albicans або Aspergillus; клітини комах, такі як Drosophila S2 і Spodoptera Sf9; клітини тварин, такі як CHO, COS, HeLa, C127, 3T3, BHK, 293, CV-1, клітини меланоми Боуеса і мієломи; і клітини рослин, такі як клітини голонасінних або покритонасінних рослин. Клітини-хазяїни для реалізації способів, що складають предмет даного винаходу, можуть бути представлені у вигляді окремих клітин або популяцій клітин. Популяції клітин можуть являти собою популяцію виділених або культивованих клітин або популяцію клітин, присутніх в матриксі, такому як тканина. Введення полінуклеотиду, такого як вектор, в клітину-хазяїна можна здійснити способами, добре відомими фахівцям в даній галузі (Davis et al., Basic Methods in Molecular Biology, 2-е вид., Appleton & Lange, м. Норуолк, штат Коннектикут, 1994; Sambrook et al., Molecular Cloning: А Laboratory Manual, 3-вид., Cold Spring Harbor Laboratory Press, Колд-Спринг-Харбор, штат НьюЙорк, 2001). До цих способів належать трансфекція з використанням фосфату кальцію, трансфекція з обробкою DEAE-декстраном, мікроін'єкція, трансфекція через катіонні ліпіди, електропорація, трансдукція, введення при зішкрібанні, балістичне введення і інфікування. Структура поліпептидів або фрагментів, що складають предмет даного винаходу, може бути модифікована для таких цілей, як підвищення специфічності, стабільності, розчинності субстрату і т. п. Наприклад, можна отримати модифікований поліпептид, в якому послідовність амінокислот змінена, наприклад, шляхом амінокислотної заміни, делеції або вставки. Очікується, що виділена заміна лейцину на ізолейцин або валін, аспартату на глутамат, треоніну на серин, або інша аналогічна заміна амінокислоти на близьку по структурі амінокислоту (тобто консервативні мутації) в більшості, але не у всіх, випадках не буде чинити великого впливу на біологічну активність молекули, що отримується. Консервативними називають заміни, що проводяться всередині сімейства амінокислот з близькою структурою бічних ланцюгів. Генетично кодовані амінокислоти можна розділити на чотири сімейства: (1) кислотні (аспартат, глутамат); (2) основні (лізин, аргінін, гістидин); (3) неполярні (аланін, валін, лейцин, ізолейцин, пролін, фенілаланін, метіонін, триптофан); і (4) незаряджені полярні (гліцин, аспарагін, глутамін, цистеїн, серин, треонін, тирозин). Фенілаланін, триптофан і тирозин іноді об'єднують в одну групу ароматичних амінокислот. Альтернативно, набір амінокислот може бути розділений на наступні групи: (1) кислі (аспартат, глутамат); (2) основні (лізин, аргінін, гістидин), (3) аліфатичні (гліцин, аланін, валін, лейцин, ізолейцин, серин, треонін), причому серин і треонін можна необов'язково згрупувати в окрему групу аліфатичних гідроксильних амінокислот; (4) ароматичні (фенілаланін, тирозин, триптофан); (5) амідні (аспарагін, глутамін); і (6) сірковмісні (цистеїн і метіонін) (Stryer (ред.), Biochemistry, 2-е видання, WH Freeman and Co., 1981). Щоб швидко визначити можливість отримання функціонального гомолога в результаті зміни амінокислотної послідовності поліпептиду або його фрагмента, можна оцінити здатність модифікованого поліпептиду або фрагмента викликати відгук аналогічно немодифікованому поліпептиду або фрагменту з використанням описаних в цьому документі тестів. Пептиди, поліпептиди або білки з більше ніж однією заміною також можуть бути легко перевірені аналогічним чином. Поліпептиди, що складають предмет даного винаходу, можна також приготувати у вигляді суміші з фармацевтично прийнятним носієм або розріджувачем. Для цих цілей можна використовувати різні водні носії, наприклад, 0,4 %-ий сольовий розчин, 0,3 %-ий гліцин і т. п. Вказані розчини стерильні і звичайно не містять твердих домішок. Стерилізацію вказаних розчинів можна проводити з використанням загальноприйнятих, добре відомих способів стерилізації (наприклад, фільтруванням). Композиції можуть містити фармацевтично прийнятні допоміжні речовини, необхідні для імітування фізіологічних умов, наприклад, агенти для 5 UA 107596 C2 5 10 15 20 25 30 35 40 45 50 55 60 коректування і підтримки pH. Концентрація поліпептидів, що складають предмет даного винаходу, в такій фармацевтичній композиції може значно варіюватися, наприклад, від менше ніж приблизно 0,5 %, звичайно щонайменше приблизно 1 %, до 15 або 20 % ваг., і визначається, насамперед, на основі об'єму рідини, її в'язкості і інших факторів відповідно до конкретного вибраного режиму введення. Фахівці в даній галузі легко визначать відповідне терапевтично ефективне дозування. Певне дозування можна, при необхідності, вводити повторно через відповідні проміжки часу, вибрані лікуючим лікарем або іншим фахівцем у відповідній галузі (наприклад, медичною сестрою, ветеринаром або ветеринарним фельдшером) в період лікування. Поліпептиди, що складають предмет даного винаходу, можуть бути ліофілізовані для зберігання і згодом перед використанням відновлені у відповідному носії. Цей спосіб зарекомендував себе як ефективний для стандартних білкових препаратів. Способи ліофілізації і відновлення добре відомі фахівцям. Інший варіант здійснення даного винаходу являє собою спосіб експресії поліпептиду, що включає стадії забезпечення клітини-хазяїна, що складає предмет даного винаходу; і культивування клітини-хазяїна в умовах, достатніх для експресії щонайменше одного поліпептиду, що складає предмет даного винаходу. Клітини-хазяїни можуть культивуватися в будь-яких умовах, що забезпечують підтримування або збільшення чисельності даного типу клітин і достатніх для експресії поліпептиду. Умови культивування, середовища і відповідні способи обробки, достатні для експресії поліпептидів, добре відомі фахівцям в даній галузі. Наприклад, багато типів клітин ссавців можна культивувати в аеробний умовах при 37 °C з використанням відповідним чином буферизованого середовища DMEM, тоді як клітини бактерій, дріжджів і інші види клітин можна культивувати при 37 °C в умовах відповідної атмосфери в середовищі LB. Експресія поліпептиду в рамках способів, що складають предмет даного винаходу, може бути підтверджена з використанням різних добре відомих способів. Наприклад, експресія поліпептиду може бути підтверджена з використанням виявляючих реагентів, таких як антитіла, з використанням, наприклад, FACS-аналізу або імунофлуоресцентних способів детектування або електрофорезом в ПААГ з додецилсульфатом натрію, або ВЕРХ. Інший аспект даного винаходу являє собою спосіб доставки терапевтичного агента через клітини ендотелію, що включає: a. кон'югацію терапевтичного агента з поліпептидом, що містить поліпептид з послідовністю, приведеною в SEQ ID NO: 1, 2, 4, 8 або 12, з утворенням кон'югата; b. приведення кон'югата в контакт з клітинами ендотелію; і с. вимірювання кількості кон'югата, доставленої через клітини ендотелію. Поліпептиди, що складають предмет даного винаходу, полегшують доставку терапевтичного агента через клітини ендотелію шляхом зв'язування поліпептидів, що складають предмет даного винаходу, з IGF1R. Пептиди вибирають таким чином, щоб кон'югований з ними терапевтичний агент не перешкоджав зв'язуванню поліпептидів, що складають предмет даного винаходу, з IGF1R. У даному винаході описана кон'югація білка з молекулярною масою приблизно 100 кДа (921 амінокислота) з поліпептидами, що складають предмет даного винаходу, без втрати трансцитозної активності. Інші поліпептиди порівнянних розмірів, ймовірно, також можуть бути успішно кон'юговані з поліпептидами, що складають предмет даного винаходу, і доставлятися через клітини ендотелію в мозок. Ефективність доставки кон'югата через клітини ендотелію може бути виміряна in vitro або in vivo за допомогою добре відомих способів. Типові вимірювання in vitro можна проводити з використанням поляризованих моношарів клітин ендотелію, вимірюючи трансцитоз кон'югата із застосуванням, наприклад, антитіл до кон'югата. Вимірювання in vivo можна провести у пацієнта з використанням, наприклад, радіоактивно мічених кон'югатів, вимірюючи їх розподіл в мозку після введення. Між результатами вимірювання in vitro і in vivo спостерігали хорошу кореляцію. Наприклад, в роботі Perrier et al. продемонстрована хороша кореляція (R=0,94) між коефіцієнтами проникності ряду сполук і відповідними коефіцієнтами перенесення через гематоенцефалічний бар'єр in vivo у гризунів при використанні спільних культур астроцитів і клітин ендотелію мозку щурів (Perrier et al., Brain Res. 1150:1-13, 2007). Даний винахід далі буде описаний з посиланням на конкретні необмежуючі приклади. Приклад 1 Виявлення пептидів, що зв'язуються з IGF1R Фаговий пенінг Бібліотеки фагів pIX з випадковими пептидами генерували слідуючи способам, описаним в заявці на патент США № US2010/0021477, і використовували їх як джерело пептидів, що 6 UA 107596 C2 5 10 15 20 25 зв'язуються з IGF1R людини. На даній бібліотеці провели пенінг в розчині за біотинілованою формою очищеного розчинного IGF1R (sIGF1R) з гексагстидиновою міткою на С-кінці (R&D Systems, м. Міннеаполіс, штат Міннесота) протягом трьох циклів. Біотинілування sIGF1R проводили за допомогою реагенту EZ-Link Sulfo-NHS-LC-Biotin формату No-Weigh в мікротрубках (Pierce, м. Рокфорд, штат Іллінойс). Через великий розмір sIGF1R (~330 кДа) для 1 циклу очищення використали гранули Tetralink Avidin завдяки їх приблизно 10-кратній зв'язувальній здатності, в порівнянні з магнітними гранулами Dynal. Кожний з 384 отриманих в результаті пенінгу окремих фаголізатів перевірили на специфічність зв'язування з sIGF1R, використовуючи фаговий твердофазний імуноферментний аналіз (ІФА). Коротко, на стінки ямки імунологічний планшетів Black Maxisorp (Nunc, м. Рочестер, штат Нью-Йорк) сорбували по 100 мкл/ямку 5 мкг/мл розчину sIGF1R (R&D Systems, м. Міннеаполіс, штат Міннесота), потім додавали в ямку по 25 мкл фаголізату і визначали проходження реакції за допомогою антитіла anti-M13-HRP (EMD Biosciences, м. Гібстаун, штат Нью-Джерсі) і субстрат пероксидази (Roche, м. Індіанаполіс, штат Індіана), фіксуючи сигнал спектрофотометром для зчитування планшетів TEKAN. Як негативний контроль використали неспоріднений білок, що достовірно не проходить через ГЕБ. Позитивні лизати визначали як клони, для яких специфічний сигнал sIGF1R утроє перевищував фоновий сигнал негативного контролю. Усього було отримано 13 клонів з унікальними пептидними послідовностями, що достовірно зв'язуються з sIGF1R (таблиця 1). 3 з цих 13 клонів (клони 5, 13 і 16) перехресно взаємодіяли з рецепторами інсуліну. Пептиди з клонів 5-14, 16 і 17 клонували всередині рамки зчитування як злиті білки пептиду і лужної фосфатази-His6 (peptide-AP) в модифікований вектор pET20b+, що містить клонований ген хлорамфенікол-ацетилтрансферази (CAT). Отримані злиті білки типу peptide-AP експресували в бактеріях і очищали за допомогою системи очищення Ni-NTA (EMD Biosciences, м. Гібстаун, штат Нью-Джерсі) згідно з інструкціями виробника. Амінокислотна послідовність використаної лужний фосфатази приведена в SEQ ID NO: 30. Таблиця 1a Поліпептидні послідовності виявлених пептидів, що зв'язуються з IGF1R Пептидна послідовність TGCDFPELCRGCHP AECEWPWLTLELCQS PFCYSGGPLPYPCTY PVCPSFCYDQYVCPT FTCAVYSLSELDCRD LSCYDPTLRTLYCHV HTCFYPTLMPPELCFD SNCPPLDMRLTELCVM WHCTPLTQIADPGSIIHILECTV VECDTPSITFSPGLEALFWNTCSP MTCAWHTLHTDPGLTPQLTLPCIY AGCPSPMPPVDPGFYSAIVQLCRE DDIDEFLHQLHNLVNNVH 30 35 Номер клону 5 6 7 8 9 10 11 12 13 14 15 16 17 SEQ ID NO пептиду 1 2 3 4 5 6 7 8 9 10 11 12 13 Ефективність зв'язування очищених злитих білків peptide-AP з sIGF1R оцінювали методом імуноферментного аналізу (ІФА) по зв'язуванню з іммобілізованим sIGF1R. Коротко, бактерії, трансформовані кожним з векторів експресії злитих білків peptide-AP, вирощували протягом ночі, і на наступний день культури центрифугували при швидкості 4 500 об/хв і температурі 4 °C. Супернатант збирали, і 75 мкл кожного супернатанту тестували на зв'язування з 2 мкг/мл sIGF1R, іммобілізованого на планшетах для імуноферментного аналізу. Пов'язані пептиди визначали за допомогою субстрату Attophos (Roche, м. Індіанаполіс, штат Індіана) згідно з рекомендаціями виробника з використанням спектрофотометра для зчитування планшетів Molecular Devices M5. Злиті білки пептидних клонів 7 і 10 експресувались погано. Всі інші злиті білки peptide-AP зв'язувалися з sIGF1R (фіг. 2). 7 UA 107596 C2 Таблиця 1b Полінуклеотидні послідовності пептидів, що зв'язуються з IGF1R Номер SEQ Послідовність клону ID NO: 5 14 ACGGGTTGTGATTTTCCGGAGTTGTGTCGTGGTTGTCATCCG 6 15 GCTGAGTGTGAGTGGCCGTGGCTTACGCTGGAGCTTTGTCAGTCT 7 16 CCTTTTTGTTATTCTGGTGGGCCGCTGCCGTATCCTTGTACGTAT 8 17 CCTGTGTGTCCGTCGTTTTGTTATGATCAGTATGTGTGTCCGACT 9 18 TTTACGTGTGCTGTTTATTCGTTGTCTGAGCTGGATTGTAGGGAT 10 19 TTGAGTTGTTATGATCCGACGCTGCGTACGTTGTATTGTCATGTT 11 20 CATACGTGTTTTTATCCTACGCTGATGCCTCCTGAGCTGTGTTTCGAT 12 21 AGTAATTGTCCTCCGTTGGATATGCGGCTGACTGAGCTTTGTGTTATG TGGCATTGTACTCCGCTGACGCAGATTGCTGATCCAGGCCAGAGTATTATTCATA 13 22 TTTTGGAGTGTACTGTT GTTGAGTGTGATACGCCGTCTATTACTTTTTCGCCAGGCCTGGAGGCGCTGTTTT 14 23 GGAATACGTGTTCTCCT ATGACGTGTGCGTGGCATACTTTGCATACGGATCCAGGCCTTACTCCGCAGCTGA 15 24 GGCTGCCTTGTATTTAT GCGGGGTGTCCGAGTCCGATGCCGCCGGTGGATCCAGGCTTTTATAGTGCTATT 16 25 GTGCAGCTGTGTAGGGAG 17 26 GATGACATAGACGAATTTCTTCATCAACTCCACAACCTAGTAAACAATGTTCAC 5 10 15 20 25 30 35 Приклад 2 Характеристики пептидів, що зв'язуються з IGF1R Виявлені пептиди, що зв'язуються з IGF1R, клонували всередині рамки зчитування до Скінця домену IgG білка G (PG) в модифікований вектор pET17b (EMD Chemicals, м. Гібстаун, штат Нью-Джерсі), що має незалежний від лігування сайт клонування (LIC), отримавши злиті білки PG-peptide. Домен білка G, що зв'язується з IgG, стабільний, що полегшило виділення злитого білка з бактеріальних лізатів. Амінокислотна послідовність використаного домену IgG білка G приведена в SEQ ID NO: 31. Злиті білки PG-peptide експресували в індукованих 1 мМ IPTG бактеріях і очищали на гранулах IgG Sepharose (GE Healthcare Life Sciences, м. Піскатауей, штат Нью-Джерсі) з бактеріальних лізатів, очищених центрифугуванням в режимі 16000g при 4 °C протягом 20 хв. Відносну афінність зв'язування злитих білків PG-peptide з sIGF1R вимірювали методом імуноферментного аналізу. Визначення величин Kd проводили з використанням програмного забезпечення GraphPad Prism 4 по рівнянню для одного сайту зв'язування. По 100 мкл розчину 2 мкг/мл sIGF1R (R&D Systems, м. Міннеаполіс, штат Міннесота) в сольово-фосфатному буфері Дульбекко (DPBS (-/-)) протягом ночі сорбували на стінки 96-ямкових планшетів Black Maxisorp (Nunc, м. Рочестер, штат Нью-Йорк) при 4 °C. Потім планшети промивали буфером TBST і блокували ямку 200 мкл/ямку буфером Starting Block T20 (TBS) (Pierce, м. Рокфорд, штат Іллінойс) протягом 1 години при кімнатній температурі при струшуванні зі швидкістю 300 об/хв. У ямку додали по 75 мкл/ямку очищеного розчину злитих білків PG-peptide в послідовному розведенні, починаючи з концентрації 40 мкМ, і інкубували при кімнатній температурі протягом 1 години при перемішуванні зі швидкістю 300 об/хв; потім об'єм розчину в ямку доводили до 100 мкл буфером Starting Block T20. Планшети тричі промивали буфером TBST, в кожну ямку вносили по 100 мкл/ямку мічених пероксидазою антитіл кролика (1:5000) (Rockland, м. Гілбертсвілль, штат Пенсільванія), планшети знову промивали і детектували сигнал, використовуючи по 100 мкл/ямку субстрату пероксидази (Roche, м. Індіанаполіс, штат Індіана). Інтенсивність хемілюмінесценції визначали спектрофотометром для зчитування планшетів Molecular Devices M5. Для проведення конкурентного аналізу до буфера Starting Block T20, що використовується для доведення об'єму рідини в ямку до 100 мкл, додавали інсуліноподібний фактор росту 1 (IGF-1) (R&D Systems, м. Міннеаполіс, штат Міннесота) до кінцевої концентрації 400 нM. Характеристики злитих білків PG-peptide приведені в таблиці 2. Більшість пептидів зв'язувалися з відносною афінністю в області низької концентрації в мкM (0,75-8 мкМ). Злитий білок з пептидом клону 17 відрізнявся тим, що крива профілю зв'язування для нього мала 8 UA 107596 C2 дзвоноподібну форму. Така поведінка аналогічно зв'язуванню інсуліну з інсуліновим рецептором і IGF-1 до IFG1R, лігандів, які при зв'язуванні індукують конформаційні зсуви в рецепторах і мають негативні профілі кооперативності при вищих концентраціях лігандів. Результати конкурентного аналізу з використанням IGF-1 представлені в таблиці 2. 5 Таблиця 2 Номер клону 5 6 7 8 9 10 11 12 13 14 15 16 17 10 15 20 25 30 35 40 SEQ ID NO пептиду 1 2 3 4 5 6 7 8 9 10 11 12 13 EC50(мкM) 0,75 1 1,8 1,6 3,5 4,3 1,2 1,8 >50 0,85 2,6 8,2 не проводили Конкурує з IGF-1 так так так так так Трансцитоз так так не проводили так ні не проводили ні так ні ні не проводили так ні Приклад 3 Модель ГЕБ in vitro Вибрані пептиди, що зв'язуються з sIGF1R, додатково вивчили на моделі гематоенцефалічного бар'єра in vitro і моделі клітин ендотелію мікросудин мозку щура. Клітини ендотелію капілярів мозку щура підготували, слідуючи роботі Perriere et al., J. Neurochem. 93:279-289, 2005. Коротко, мозок 6-8-тижневих самців щурів лінії Sprague Dawley розвертали на хроматографічному папері 3MM для видалення оболонок головного мозку, розрізали сагітально, відсікали білу речовину і залишали тільки кору, яку потім ретельно подрібнювали. Подрібнену кору головного мозку переносили в поліпропіленову конічну пробірку об'ємом 50 мл, що містить 20 мл модифікованого за способом Дульбекко середовища Ігла (DMEM) з додаванням 39 од./мл ДНКази I (Worthington, м. Лейквуд, штат Нью-Джерсі) і 0,7 мг/мл колагенази типу 2 (Worthington, м. Лейквуд, штат Нью-Джерсі) в кінцевій концентрації, і інкубували при 37 °C, обережно перемішуючи, протягом 1,25 години. Після швидкого центрифугування отриману пелету повторно розчиняли в 20 мл 20 % розчину BSA (Sigma, м. Сент-Луїс, штат Міссурі) в DMEM, центрифугували, і виділену збагачену мікросудинами пелету розщеплювали другий раз в 20 мл середовища DMEM з додаванням 39 од./мл ДНКази I і 1 мг/мл колагенази/диспази (Roche, м. Індіанаполіс, штат Індіана) при 37 °C протягом 1 години. Продукт розщеплення швидко центрифугували, отриману в результаті пелету повторно розчиняли в 2 мл середовища DMEM і потім наносили понад 33 % безперервного градієнту Percoll, центрифугували, і збагачену фракцію мікросудин витягували і ще раз швидко центрифугували. Пелету з клітинами повторно розчиняли в 10 мл повного середовища для вирощування клітин ендотелію мікросудин мозку щурів (DMEM, 20 % отриманих з плазми сироватки (PDS), 100 мкг/мл гепарину, 2 мМ L-глутаміну, 100 од./мл пеніциліну, 100 мкг/мл стрептоміцину, 0,25 мкг/мл амфотерицину і висівали на 10 см чашки Петрі на 4 години при 37 °C в атмосфері 5 % CO2. Через 4 години решту клітин, що залишилися незакріпленими, відсмоктували піпеткою і підраховували в гемацитометрі з трипановим синім. Клітини висівали у верхній камері мембран Transwell (розмір пір 0,4 мкм, діаметр 1,12 см, Corning, м. Актон, штат Массачусетс), оброблених 400 мкг/мл колагену типу IV (Sigma, м. Сент-Луїс, штат Міссурі) і 100 5 мкг/мл фібронектину (Sigma, м. Сент-Луїс, штат Міссурі) зі щільністю 6×10 клітин/мл, 500 мкл на ямку. У нижню камеру вміщували 1 мл поживного середовища. У обидві камери додавали 4 мкг/мл пуроміцину (Clontech, м. Маунтин-Вию, штат Каліфорнія) і інкубували планшети при 37 °C в атмосфері 5 % CO2 протягом ночі. На наступний день поживне середовище замінювали на свіже повне культуральне середовище з додаванням 4 мкг/мл пуроміцину, і клітини знову вміщували в інкубатор на ніч. На наступний день поживне середовище замінювали на свіже повне культуральне середовище, заміну проводили ще раз через два дні. Культури клітин оцінювали візуально до досягнення ступеня злиття колоній 100 %, приблизно через 6-7 днів після висівання. Розроблена in vitro модель ГЕБ мала високий трансендотеліальний 9 UA 107596 C2 2 5 10 15 електричний опір (>100 Ом-см ), виміряний за допомогою системи Millicell-ERS (Millipore, м. Біллеріка, штат Массачусетс), і дуже низьку проникність для Na-флуоресцеїну (приблизно 1-6 5×10 см/с). У верхню камеру з моделлю ГЕБ in vitro додавали 25 мкг очищених злитих білків peptide-AP, і трансцитозовані злиті білки peptide-AP визначали через 15 і 30 хвилин в нижній камері за допомогою імуноферментного аналізу. Коротко, 75 мкл кожного зразка переносили на планшети, покриті моноклональними антитілами миші до антибактеріальної лужної фосфатази в концентрації 5 мкг/мл (Sigma, м. Сент-Луїс, штат Міссурі). Планшет інкубували протягом 1 години, промивали, виявляли сигнал за допомогою субстрату Attophos (Roche, м. Індіанаполіс, штат Індіана) згідно з рекомендаціями виробника і вимірювали результат з використанням спектрофотометра для зчитування планшетів Molecular Devices M5 з довжиною хвилі збудження 440 нм і довжиною хвилі випромінювання 550 нм. Отримані результати приведені в таблиці 2. Після приведеного повного опису даного винаходу фахівцеві в даній галузі буде зрозуміло, що можлива множина змін і модифікацій, що не виходять за межі суті або об'єму прикладеної формули винаходу. 10 UA 107596 C2 11 UA 107596 C2 12 UA 107596 C2 13 UA 107596 C2 14 UA 107596 C2 15 UA 107596 C2 16 UA 107596 C2 17 UA 107596 C2 18 UA 107596 C2 19 UA 107596 C2 20 UA 107596 C2 21 UA 107596 C2 22 UA 107596 C2 ФОРМУЛА ВИНАХОДУ 5 10 15 20 25 1. Виділений поліпептид, який зв'язується з рецептором інсуліноподібного фактора росту 1, що містить поліпептид з послідовністю, представленою в SEQ ID NO:1-13. 2. Виділений полінуклеотид, що містить полінуклеотид, який кодує поліпептид, що містить амінокислотну послідовність, представлену в SEQ ID NO:1-13. 3. Виділений полінуклеотид, що містить полінуклеотид з послідовністю, представленою в SEQ ID NO:14-26, або його комплементарною послідовністю. 4. Виділений вектор, що містить полінуклеотид з послідовністю, представленою в SEQ ID NO:14-26. 5. Вектор за п. 4, що являє собою вектор експресії. 6. Виділена клітина-хазяїн, що містить вектор за п. 4. 7. Виділений злитий білок, що містить поліпептид з послідовністю, приведеною в SEQ ID NO:113, злитий з другим поліпептидом. 8. Злитий білок за п. 7, де другий поліпептид кодує імуноглобулін або його фрагмент. 9. Спосіб експресії поліпептиду, що включає стадії: a) забезпечення клітини-хазяїна за п. 6; і b) культивування клітини-хазяїна в умовах, достатніх для експресії поліпептиду з послідовністю, приведеною в SEQ ID NO:1-13. 10. Спосіб доставки терапевтичного агента через клітини ендотелію, що включає: a) кон'югацію терапевтичного агента з поліпептидом, що містить поліпептид з послідовністю, приведеною в SEQ ID NO:1, 2, 4, 8 або 12, з утворенням кон'югата; b) приведення кон'югата в контакт з клітинами ендотелію; і с) вимірювання кількості кон'югата, доставленої через клітини ендотелію. 11. Спосіб за п. 10, де клітини ендотелію утворюють гематоенцефалічний бар'єр. 23 UA 107596 C2 Комп’ютерна верстка Л. Ціхановська Державна служба інтелектуальної власності України, вул. Урицького, 45, м. Київ, МСП, 03680, Україна ДП “Український інститут промислової власності”, вул. Глазунова, 1, м. Київ – 42, 01601 24
ДивитисяДодаткова інформація
Назва патенту англійськоюInsulin-like growth factor 1 receptor binding peptides
Автори англійськоюDiem, Michael, O'Neil, Karen
Автори російськоюДим Майкл, О'Нил Карин
МПК / Мітки
МПК: C12P 21/06, A61K 39/00, A61K 38/00, C12P 21/04
Мітки: виділений, рецептором, зв'язується, поліпептид, інсуліноподібного, фактора, росту
Код посилання
<a href="https://ua.patents.su/26-107596-vidilenijj-polipeptid-yakijj-zvyazuehtsya-z-receptorom-insulinopodibnogo-faktora-rostu-1.html" target="_blank" rel="follow" title="База патентів України">Виділений поліпептид, який зв’язується з рецептором інсуліноподібного фактора росту 1</a>
Моноклональне антитіло людини, яке специфічно зв’язується з рецептором інсуліноподібного фактора росту і (igf-ir)

Номер патенту: 87804
Опубліковано: 25.08.2009
Автори: Міллер Пенелопа Е., Кохен Брюс Д., Біб Жан, Мойєр Джеймс Д., Корвалан Хосе Р., Галло Майкл
МПК: A61K 39/395, C12N 5/20, C12N 15/13, C12N 15/64, G01N 33/574, C12P 21/08, A61K 49/00, C07K 16/18, A61P 35/00
Мітки: яке, росту, специфічно, igf-ir, людини, зв'язується, моноклональне, фактора, антитіло, інсуліноподібного, рецептором
Формула / Реферат:
1. Людське моноклональне антитіло або його антигензв'язувальна частина, яка специфічно зв'язується з рецептором інсуліноподібного фактора росту I (IGF-IR), де вказане антитіло містить важкий ланцюг і легкий ланцюг, де вказаний легкий ланцюг кодується геном Vκ А30 зародкової лінії людини, і де амінокислотні послідовності CDR1, CDR2 і CDR3 важкого ланцюга та амінокислотні послідовності CDR1, CDR2 і CDR3 легкого ланцюга вибрані з групи, що...
Стабілізовані поліпептиди інсуліноподібного фактора росту

Номер патенту: 97953
Опубліковано: 10.04.2012
Автори: Гласс Девід Джонатан, Форнаро Мара
МПК: A61P 3/10, C07K 14/65, C12N 15/27, A61K 38/18
Мітки: стабілізовані, поліпептиди, росту, фактора, інсуліноподібного
Формула / Реферат:
1. Поліпептид, що включає модифікований людський білок-попередник IGF-1, який містить IGF-1 та пептид Е, у якому залишки G1, P2 та E3 видалені, та амінокислота в положенні 37 має заміну R37А, а також видалені амінокислоти у положеннях 71 і 72.2. Поліпептид за п. 1, у якому білок-попередник містить Ea-пептид. 3. Поліпептид за п. 1, у якому білок-попередник містить Eb-пептид. 4. Поліпептид за п. 3, у якому останні сім...
Виділений поліпептид baff-r (рецептор фактора активації в-клітин сімейства tnf)

Номер патенту: 83458
Опубліковано: 25.07.2008
Автори: Амброуз Крістін М., Томпсон Джеффрі С.
МПК: G01N 33/68, C07K 16/28, A61K 48/00, C07K 14/715, C12N 15/12, C12N 15/62, A61K 38/17, C12Q 1/68, G01N 33/53, C12N 15/10, C07H 21/04
Мітки: рецептор, поліпептид, виділений, активації, фактора, в-клітин, baff-r, сімейства
Формула / Реферат:
1. Виділений поліпептид BAFF-R (рецептор фактора активації В-клітин сімейства TNF), що містить амінокислотну послідовність, вибрану з групи, яка складається з:(a) SEQ ID NО:5;(b) фрагмента SEQ ID NО:5, що зв’язується з BAFF (фактор активації В-клітин сімейства TNF);(с) амінокислотної послідовності, яка зв'язується з BAFF і щонайменше на 70 % ідентична SEQ ID NО:5, або фрагмента SEQ ID NО:5;(d) амінокислотної...
Послідовність днк, що кодує поліпептид, який зв’язується з mort-1 (варіанти), вектор експресії, що включає послідовність днк, клітина- хазяїн, виділений поліпептид та фармацевтична композиція, що містить поліп

Номер патенту: 72423
Опубліковано: 15.03.2005
Автори: Гольцев Юрій, Гончаров Таня, Воллах Девід, Болдін Марк
МПК: C07K 14/715, A61P 35/00, C12N 9/64, A61K 48/00, C07K 16/28, A61P 31/12, A61K 45/00, C12N 15/09, C07K 14/47, C07K 14/705, C12N 5/10
Мітки: послідовність, днк, експресії, поліпептид, виділений, варіанти, mort-1, поліп, фармацевтична, хазяїн, зв'язується, клітина, вектор, включає, кодує, композиція, містить
Формула / Реферат:
1. Послідовність ДНК, що кодує поліпептид, який зв'язується з MORT-1, де MORT-1 є білком, який зв'язується з внутрішньоклітинним доменом FAS-R, і який зв'язується з білком TRADD, який зв'язується з внутрішньоклітинним доменом p55-TNF-R, де зазначений поліпептид має:a) послідовність, що містить залишки 1-182 послідовності SEQ ID №5;b) послідовність, що містить фрагмент а), де фрагмент зв'язується з MORT-1;c) послідовність,...
Кон’югати інсуліноподібного фактора росту-1 (іфр-1) і поліетиленгліколю

Номер патенту: 93666
Опубліковано: 10.03.2011
Автори: Хессе Фрідеріке, Метцгер Фрідріх, Шаубмар Андреас, Ланг Курт, Фозер Штефан, Амреін Беат, Ланцендьорфер Мартін, Кюнкеле Клаус-Петер, Регула Йорг
МПК: A61P 25/28, A61K 38/30, A61K 47/48
Мітки: фактора, кон'югати, інсуліноподібного, поліетиленгліколю, іфр-1, росту-1
Формула / Реферат:
1. Кон'югат, що включає варіант інсуліноподібного фактора росту І (ІФР-І) і поліетиленгліколь (ПЕГ), який відрізняється тим, що зазначений варіант ІФР-І містить заміни (заміну) амінокислот у положеннях 27, 37, 65, 68 у послідовності вихідного ІФР-I таким чином, що одна або декілька амінокислот у положеннях 37, 65, 68 є лізином (К), а амінокислота в положенні 27 є полярною амінокислотою, але не лізином, причому зазначений ПЕГ є кон'югованим...
Попередній патент: Освітлювальний пристрій білого світла з управлінням кольоровою температурою світлового потоку
Наступний патент: Блок для охолодження і очищення повітря, передбачений для установки для кріогенної дистиляції повітря
Випадковий патент: Стикове з'єднання стінових панелей з плитами перекриття